endogenous activation of nicotinic receptors mediates...
TRANSCRIPT

Respiratory Sinus ArrhythmiaEndogenous Activation of Nicotinic Receptors Mediates RespiratoryModulation of Brainstem Cardioinhibitory Parasympathetic Neurons
Robert A. Neff, Jijiang Wang, Sunit Baxi, Cory Evans, David Mendelowitz
Abstract—The heart rate increases during inspiration and decreases during expiration. This respiratory sinus arrhythmia(RSA) occurs by modulation of premotor cardioinhibitory parasympathetic neuron (CPN) activity. However, RSA hasnot been fully characterized in rats, and despite the critical role of CPNs in the generation of RSA, little is known aboutthe mechanisms that mediate this cardiorespiratory interaction. This study demonstrates that RSA in conscious rats issimilar to that in other species. The mechanism of RSA was then examined in vitro. Rhythmic inspiratory-relatedactivity was recorded from the hypoglossal rootlet of 700- to 800-mm medullary sections. CPNs were identified byretrograde fluorescent labeling, and neurotransmission to CPNs was examined using patch-clamp electrophysiologicaltechniques. During inspiratory bursts, the frequency of both spontaneousg-aminobutyric acidergic (GABAergic) andspontaneous glycinergic synaptic events in CPNs was significantly increased. Focal application of the nicotinicantagonist dihydro-b-erythroidine in ana4b2-selective concentration (3mmol/L) abolished the respiratory-evokedincrease in GABAergic frequency. In contrast, the increase in glycinergic frequency during inspiration was not alteredby nicotinic antagonists. Prenatal nicotine exposure exaggerated the increase in GABAergic frequency duringinspiration and enhanced GABAergic synaptic amplitude both between and during inspiratory events. Glycinergicsynaptic frequency and amplitude were unchanged by prenatal nicotine exposure. This study establishes a neurochem-ical link between neurons essential for respiration and CPNs, reveals a functional role for endogenous acetylcholinerelease and the activation of nicotinic receptors in the generation of RSA, and demonstrates that this cardiorespiratoryinteraction is exaggerated in rats prenatally exposed to nicotine.(Circ Res. 2003;93:●●●-●●●.)
Key Words: nucleus ambiguusn vagal activity n respiratory sinus arrhythmian prenatal nicotinen sudden infant death syndrome
The heart rate increases during inspiration and decreasesduring the postinspiration/postexpiration period. This
respiratory-related change in heart rate, respiratory sinusarrhythmia (RSA), helps to match pulmonary blood flow tolung inflation and to maintain an appropriate diffusion gra-dient for oxygen in the lungs.1–3 RSA has been observed inneonatal4 and adult5,6 humans, baboons,7 dogs,1 rabbits,8 andseals9 but has not been well characterized in rats.
Heart rate is controlled by the activity of premotor car-dioinhibitory parasympathetic neurons (CPNs) in the brain-stem, and RSA is mediated in part by central respiratorymodulation of CPN activity. CPNs are primarily located inthe nucleus ambiguus (NA), in proximity to neurons thoughtto be essential for respiratory rhythmogenesis.2,3,10–13CPNsin the NA are intrinsically silent and therefore rely onsynaptic inputs to dictate their activity.14
Although the pathways and transmitters responsible forrespiratory modulation of CPNs are unknown,g-aminobutyric acid (GABA), glycine, and acetylcholine
(ACh) are all neurotransmitters that have been implicated inthe central generation of RSA. CPNs are inhibited duringinspiration, and this inhibition has been reversed by theintracellular injection of Cl2 (Gilbey et al15). This suggeststhat GABA- and/or glycine-mediated chloride channels maybe involved in respiratory modulation of CPNs. Paradoxi-cally, however, one author of the Gilbey et al study describedin a later review that the inhibition of CPNs during inspirationcould not be inhibited by the GABAA antagonist bicucullineor the glycine antagonist strychnine.2
ACh has been shown to inhibit CPN activity,15 and recentwork has shown that endogenous ACh activates presynapticnicotinic receptors, which enhance both GABAergic andglycinergic inputs to CPNs.16 The possible involvement ofnicotinic receptors in mediating RSA is interesting becauseprenatal nicotine augments parasympathetic and reducessympathetic control of the heart rate17 and is among thehighest risk factors for sudden infant death syndrome(SIDS).18,19 Infants that succumb to SIDS often experience a
Original received April 1, 2003; revision received July 29, 2003; accepted July 30, 2003.From the Department of Pharmacology, The George Washington University, Washington, DC.Correspondence to Robert A. Neff, Department of Pharmacology, The George Washington University, 2300 Eye St NW, Washington, DC 20037.
E-mail [email protected]© 2003 American Heart Association, Inc.
Circulation Researchis available at http://www.circresaha.org DOI: 10.1161/01.RES.0000090361.45027.5B
1
by guest on June 30, 2018http://circres.ahajournals.org/
Dow
nloaded from

sustained bradycardia, which is preceded or accompanied bya life-threatening apnea.19,20 These life-threatening events inSIDS victims are thought to be caused by exaggerated centralcardiorespiratory interactions.17,19
The aim of the present study was to test whether rats havean RSA pattern similar to that of other species and toelucidate the cellular mechanisms responsible for the respi-ratory modulation of CPNs. Specifically, we tested thehypothesis that heart rate increases during inspiration in rats.In addition, we tested whether CPNs receive increasedGABAergic and glycinergic synaptic inputs duringinspiratory-related activity. Furthermore, we examinedwhether the respiratory modulation of CPNs is dependent onendogenous activation of nicotinic receptors. Because ourwork demonstrates that endogenous activation of nicotinicreceptors is responsible for RSA, we tested whether prenatalnicotine exposure alters these cardiorespiratory interactions.
Materials and MethodsPlethysmographic/Blood Pressure RecordingsAdult female Sprague-Dawley rats were anesthetized with a combi-nation of ketamine (40 mg/kg) and xylazine (5 mg/kg IP, PhoenixPharmaceuticals). The femoral artery was exposed and catheterizedwith Micro-Renthane tubing (Braintree Scientific) that had beensoaked overnight in heparinized bacteriostatic saline. The animalswere placed in a Covance infusion harness (Instech Labs, Inc) andallowed to recover for 24 to 48 hours. After recovery, the unanes-thetized, freely moving rats were placed in a whole-body plethys-mographic chamber, which allowed simultaneous measurement ofblood pressure, heart rate, and respiratory airflow using BiosystemXA software (Buxco Electronics, Inc). Only measurements recordedduring periods in which the animals were awake and sedentary wereanalyzed.
Fluorescent Labeling of CPNs and MedullarySlice PreparationsNeonatal Sprague-Dawley rats (P1-P5, Hilltop, Scottdale, Pa) wereinitially exposed to isoflurane (Abbott Laboratories) until anesthe-tized and cooled to'4°C. A right thoracotomy was performed, andthe retrograde fluorescent tracer X-rhodamine-5- (and -6)-isothiocyanate (Molecular Probes) was injected into the fat pads atthe base of the heart. After 24 hours of recovery, each animal wasanesthetized with isoflurane and decapitated, and the head wasplaced in a 4°C physiological saline solution (mmol/L: NaCl 140,KCl 5, CaCl2 2, glucose 5, and HEPES 10) bubbled with 100% O2,pH 7.4. All animal procedures were performed with the approval ofthe Animal Care and Use Committee of The George WashingtonUniversity in accordance with the recommendations of the panel oneuthanasia of the American Veterinary Medical Association and theNational Institutes of Health publicationGuide for the Care and Useof Laboratory Animals. The medulla was removed with care topreserve the hypoglossal cranial nerve rootlet. The medulla wasmounted on a cutting block and placed into a vibrating blademicrotome (Leica). Serial transverse sections were sliced in arostrocaudal progression until the inferior olives and the NA could bevisualized on the rostral surface of the tissue. A single thick (700- to800-mm) section that included CPNs, the hypoglossal nerve rootlet,the pre-Botzinger complex, and the rostral portion of the hypoglossalnucleus was cut and transferred to a recording chamber and perfused(4 mL/min) with room temperature aCSF (mmol/L: NaCl 125, KCl3, CaCl2 2, NaHCO3 26, glucose 5, and HEPES 5) equilibrated with95% O2/5% CO2, pH 7.4.
Recording Respiratory Network ActivityThe thick medullary slice preparation contains the pre-Botzingercomplex, local circuits for motor output generation, and respiratory
hypoglossal motor neurons, which generate inspiratory-phase motordischarge in hypoglossal cranial nerves.13 As seen in other similarmedullary respiratory slice preparations, the frequency ofrespiratory-related hypoglossal discharge is significantly lower thanthat in in vivo preparations; this is likely due to the reducedtemperature of the preparation and the absence of sensory input tothe medulla.21 Spontaneous respiratory-related activity was recordedby monitoring the motor neuron population activity from hypoglos-sal nerve rootlets using a suction electrode. Hypoglossal rootletactivity was amplified (50 000 times), filtered (10- to 300-Hzbandpass, CWE Inc), and adjacent-averaged (50-ms windows).Respiratory activity was also electronically integrated (t550 ms,CWE Inc) during experiments examining glycinergic synaptic inputsto CPNs.
Patch-Clamp TechniquesCPNs in the NA were identified by the presence of the fluorescenttracer.22 Patch pipettes (2.5 to 3.5 MV) were visually guided to thesurface of individual CPNs using differential interference optics andinfrared illumination (Zeiss). Patch pipettes contained (mmol/L) KCl150, MgCl2 4, EGTA 2, Na-ATP 2, and HEPES 10, pH 7.4. Thispipette solution causes the Cl2 current induced by the activation ofGABA or glycine receptors to be recorded as an inward current(calculated reversal potential of Cl2 4 mV). Voltage-clamp record-ings were made with an Axopatch 200B and pClamp 8 software(Axon Instruments). All synaptic activity in CPNs was recorded at280 mV. Only preparations in which synaptic activity increased inCPNs during inspiration (in 95 of 117 [81%] of the preparations)were used for further experimentation and analysis. Only one cellwas recorded per nucleus for an experiment. In 12 slices, anadditional cell was recorded from the same slice in the contralateralNA.
Focal Drug ApplicationFocal drug application was performed using a pneumatic Picopumppressure delivery system (WPI). Drugs were ejected from a patchpipette positioned within 30mm from the patched CPN. Themaximum range of drug application has been previously determinedto 100 to 120mm downstream from the drug pipette and consider-ably less behind the drug pipette.23 GABAergic neurotransmissionwas isolated by focal application ofD-2-amino-5-phosphonovalerate(AP-5, 50 mmol/L), 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX,50mmol/L), and strychnine (1 mmol/L) to blockN-methyl-D-aspartate (NMDA), non-NMDA, and glycinergic receptors, respec-tively. Glycinergic neurotransmission was isolated by focal applica-tion of AP-5, CNQX, and gabazine (25mmol/L) to block NMDA,non-NMDA, and GABAA receptors, respectively. Nicotinic recep-tors were blocked with dihydro-b-erythroidine (DHbE, 100mmol/L)or curare (10mmol/L). The role of different receptor subtypes wastested by applying DHbE at concentration selective for thea4b2
nicotinic receptor (3mmol/L),24 and a7 nicotinic receptor subtypeswere tested with thea7 nicotinic receptor antagonista-bungarotoxin(aBTX, 100 nmol/L). All drugs were obtained from Sigma.
Prenatal Nicotine ExposureAdult female rats were anesthetized with ketamine (40 mg/kg)/xylazine (5 mg/kg IP, Phoenix Pharmaceuticals) on the third day ofgestation and implanted with Alzet osmotic minipumps (Durect)containing (2)-nicotine (56.1 mg/mL bacteriostatic saline, Sigma).Osmotic minipumps were chosen to avoid the high plasma nicotineconcentrations and subsequent episodic fetal hypoxia/ischemia thatcan be produced by nicotine injections.25 Pumps delivered 2.1 mgnicotine per day, a level approximately equivalent to levels thatoccur in moderate to heavy smokers, for 28 days.17
Data Analysis
Plethysmographic/Blood Pressure ExperimentsHeart rate intervals were measured during inspiratory and subsequentexpiratory periods in 6 animals using Acqknowledge (version 3.7.3,
2 Circulation Research September 19, 2003
by guest on June 30, 2018http://circres.ahajournals.org/
Dow
nloaded from

Biopac Systems). Heart rate was recorded from at least 20 respira-tory cycles while the animal was awake and sedentary. Data arepresented as mean6SEM. Statistical comparisons were made usingpaired Studentt tests. A value ofP,0.05 indicates significantdifferences.
ElectrophysiologySynaptic events were detected using MiniAnalysis (version 5.6.12,Synaptosoft). The frequency and amplitude of inhibitory postsynap-tic currents (IPSCs) that occurred in CPNs during inspiratory-relatedhypoglossal activity were averaged to determine inspiratory values.Control values were determined by averaging IPSCs that occurred ina 5- to 10-second window ending 1 second before inspiratory burstonset and beginning at least 3 seconds after the end of any previousinspiratory activity. All data are presented as mean6SEM. Statisticalcomparisons were made using paired or unpaired Studentt tests, asappropriate. A value ofP,0.05 indicates significant differences.
ResultsSprague-Dawley Rats Exhibit RSA Pattern Similarto That of Other SpeciesRespiratory airflow (Figure 1, top) and blood pressure (Figure1, bottom) were simultaneously recorded in 6 conscious,freely moving rats. During inspiration, the average heartbeatinterval decreased significantly from 17164 ms during expi-ration to 16864 ms during inspiration (P,0.05). This changein heart period corresponds to an average inspiratory-relatedincrease in heart rate of 6.961.9 bpm (P,0.05).
CPNs Are Inhibited During Inspiration byEndogenous Nicotinic ACh Receptor–MediatedIncreases in GABAergic ActivityTo determine the cellular basis of RSA, the synaptic activityof CPNs was measured in vitro. GABAergic neurotransmis-sion was isolated by focal application of the glutamatergicand glycinergic antagonists AP-5, CNQX, and strychnine in22 cells (from 17 preparations). The focal application of theseantagonists did not significantly alter the frequency (P.0.05)or duration (P.0.05) of the respiratory activity. Duringinspiration, the frequency of GABAergic synaptic inputs to
CPNs was significantly increased (basal 5.260.7 Hz, inspi-ration 10.561.3 Hz,P,0.01, n522 cells; Figure 2a). AllIPSCs under these recording conditions were blocked byfocal application of the GABAA antagonist gabazine (Figure2b). Focal application of the nicotinic antagonist curaresignificantly reduced the inspiratory-related increase inGABAergic synaptic frequency (control basal 3.566.0 Hz,control inspiration 7.661.3 Hz, curare basal 2.660.4 Hz,curare inspiration 4.460.6 Hz,P,0.05, n512 cells from 10preparations; Figure 2c) but did not significantly affect thebasal frequency of IPSCs between bursts (P,0.05, n512cells). GABAergic synaptic amplitude was not significantlyaltered by inspiratory activity or by the application of curare(P.0.05, n512 cells).
Determination of the nAChR Subtype MediatingInspiratory-Related Increase in GABAergic Inputto CPNsTo determine the specific nicotinic ACh receptor (nAChR)subtype mediating the respiratory-related increase inGABAergic frequency, subtype-selective nicotinic antago-nists were used. Focal application ofaBTX (a7-selectivenicotinic antagonist) had no significant effect on the fre-quency of GABAergic synaptic inputs to CPNs betweeninspiratory bursts (control 4.961.2 Hz, aBTX 5.561.2 Hz,P.0.05, n54 cells) and did not alter the inspiratory-relatedincrease in GABAergic synaptic frequency (control basal5.061.2 Hz, control inspiration 15.262.7 Hz, aBTX basal5.761.1 Hz,aBTX inspiration 11.462.4,P.0.05, n54 cellsfrom 4 preparations). Focal application of the nicotinicantagonist DHbE, in a concentration selective for thea4b2
receptor subtype (3mmol/L), did not affect GABAergicsynaptic frequency between bursts (control 8.561.7 Hz,DHbE 6.461.2 Hz,P.0.05, n56 cells from 3 preparations)but did abolish the increase in GABAergic frequency duringinspiration (control basal 8.561.7 Hz, control inspiration13.962.7 Hz, DHbE control 6.461.2 Hz, DHbE inspiration
Figure 1. Heart rate increases during inspiration (Insp) and decreases during expiration (Exp) in the rat. Blood pressure (BP) and respi-ratory airflow were recorded simultaneously in unanesthetized, freely moving rats (n56 animals). The heartbeat-to-heartbeat intervalwas significantly decreased from 17164 to 16764 ms during Insp (P,0.05).
Neff et al Mechanism of Respiratory Sinus Arrhythmia 3
by guest on June 30, 2018http://circres.ahajournals.org/
Dow
nloaded from

8.261.8 Hz, P,0.05, n56 cells; Figure 3). GABAergicsynaptic amplitude was not significantly altered byaBTX(control 49.768.2 pA,aBTX 40.568.9 pA,P.0.05, n54cells) or DHbE (control 57.2611.4 pA, DHbE 52.969.2 pA,P.0.05, n56 cells).
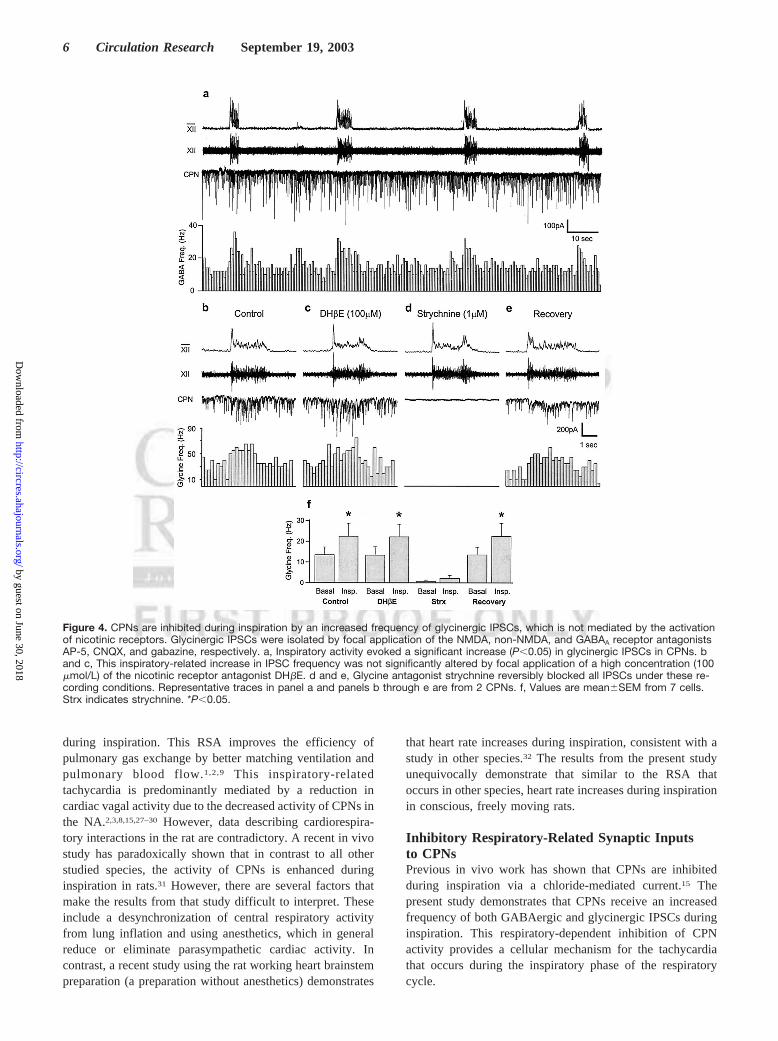
Glycinergic Respiratory Inputs to CPNsIn an additional 13 cells (from 10 preparations), glycinergicactivity was isolated by focal application of the glutamatergicand GABAergic antagonists AP-5, CNQX, and gabazine. Theapplication of these antagonists did not significantly alter thefrequency (P.0.05) or duration (P.0.05) of the respiratoryactivity. During inspiration, glycinergic synaptic frequencywas also significantly increased (basal 11.062.2 Hz, inspira-tion 19.063.5 Hz,P,0.01, n513 cells; Figure 4a). Focalapplication of the nicotinic antagonist DHbE in a highconcentration that blocks all nicotinic receptors (100mmol/L,Figures 4b, 4c, and 4f) did not significantly alter theinspiratory-related increase in glycinergic frequency (controlbasal 13.463.8 Hz, control inspiration 22.366.3 Hz, DHbE
13.464.0 to 22.066.1 Hz,P.0.05, n57 cells from 5preparations). DHbE did not significantly alter the frequencyof glycinergic synaptic events between inspiratory events(DHbE, n57 cells,P.0.05). All IPSCs under these record-ing conditions were reversibly blocked by focal application ofstrychnine (Figures 4d through 4f). Glycinergic amplitudewas not altered by respiratory activity (basal 52.065.1 pA,inspiration 59.669.1 pA,P.0.05, n513 cells) or by appli-cation of DHbE (control 52.065.1 pA, DHbE 44.967.7 pA,P.0.05, n57 cells). Application of 100mmol/L DHbE didnot significantly alter the frequency (P.0.05) or duration(P.0.05) of the respiratory activity.
Effect of Prenatal Nicotine Exposure onGABAergic and Glycinergic Synaptic Inputsto CPNs
GABAThe frequency of GABAergic synaptic events increased460690% in animals prenatally exposed to nicotine (n518
Figure 2. Endogenous activation of nicotinic receptors mediates GABAergic inhibition of premotor CPNs during inspiration. Inspiratory-related bursting activity was recorded from the hypoglossal rootlet (XII), rectified, and adjacent-averaged (XII with bar above). Fluores-cently identified CPNs were patch-clamped in the whole-cell configuration, and GABAergic IPSCs were isolated by focal application ofthe NMDA, non-NMDA, and glycine receptor antagonists AP-5 (50 mmol/L), CNQX (50 mmol/L), and strychnine (1 mmol/L), respectively.a, During inspiratory activity, the frequency of GABAergic IPSCs in CPNs was significantly increased (P,0.05). b, GABAA antagonistgabazine blocked all IPSCs. c, Nicotinic receptor antagonist curare (10 mmol/L) significantly inhibited (P,0.05) the inspiratory-relatedincrease in GABAergic synaptic frequency in CPNs. Representative traces in panels a, b, and c are from 3 CPNs.
4 Circulation Research September 19, 2003
by guest on June 30, 2018http://circres.ahajournals.org/
Dow
nloaded from

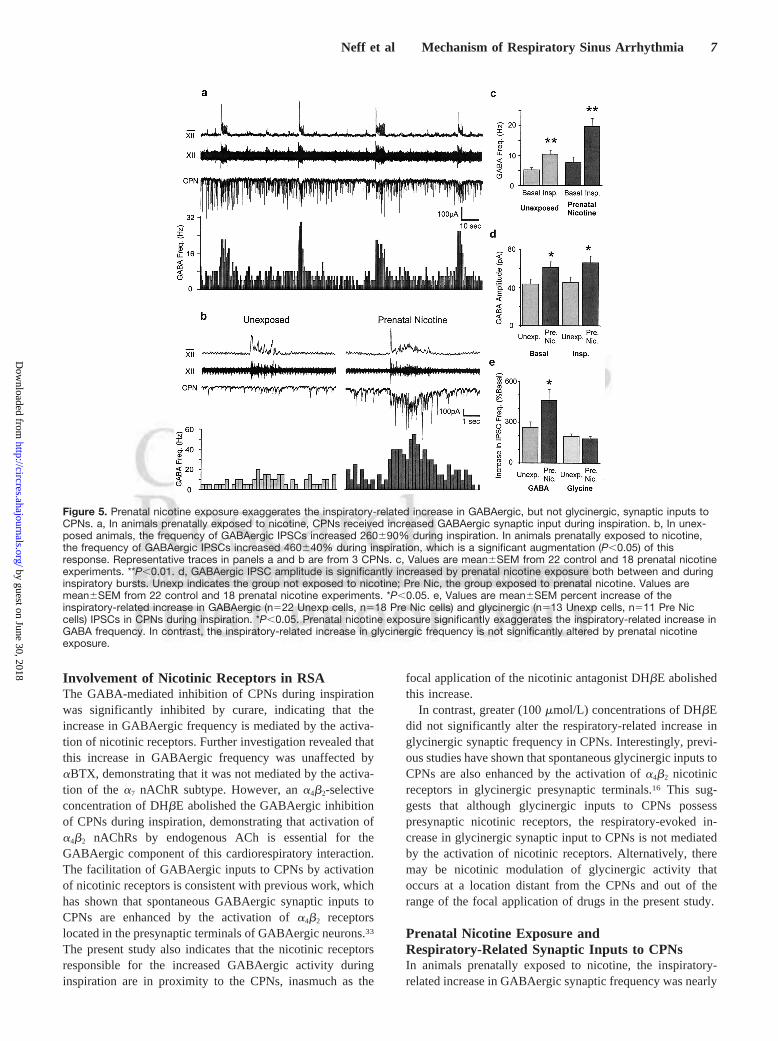
cells from 17 preparations), a significant exaggeration of the260640% increase observed in unexposed animals (n522cells, P,0.01; Figures 5a through 5c and 5e). Prenatalnicotine did not significantly alter the frequency of GABAer-gic synaptic events between inspiratory bursts (unexposed5.260.7 Hz [n522 cells], prenatal nicotine 7.760.7 [n518cells], P.0.05). Focal application of DHbE at a concentra-tion (3 mmol/L) selective fora4b2 nicotinic receptors signif-icantly inhibited the inspiratory-related increase in GABAer-gic frequency in animals prenatally exposed to nicotine(control basal 6.261.2 Hz, control inspiration 14.862.2 Hz,DHbE basal 2.860.8 Hz, DHbE inspiration 4.661.3 Hz,P,0.05, n56 cells from 5 preparations). In addition, DHbEsignificantly reduced basal GABAergic synaptic frequency(P.0.05, n56 cells).
Prenatal nicotine also significantly increased GABAergicsynaptic amplitude relative to unexposed animals both be-tween (unexposed 43.764.5 pA [n522 cells], prenatal nico-tine 61.065.5 pA [n518 cells],P,0.05) and during (unex-posed 45.365.3 pA [n522 cells], prenatal nicotine 66.166.2pA [n518 cells],P,0.05) inspiratory bursts (Figures 5d and5e). All GABAergic synaptic events were reversibly blockedby focal application of gabazine.
GlycineThe inspiratory-related increase in glycinergic frequency wasnot significantly altered in animals prenatally exposed to
nicotine (179617%, n511 cells from 8 animals) comparedwith unexposed animals (191615%, n513 cells,P.0.05;Figure 5e). Glycinergic synaptic amplitude was not signifi-cantly altered by prenatal nicotine exposure between (unex-posed 52.065.0 pA [n513 cells], prenatal nicotine 43.365.5pA [n511 cells],P.0.05) or during inspiratory bursts (un-exposed 59.669.1 pA [n513 cells], prenatal nicotine45.267.9 pA [n511 cells],P,0.05). All glycinergic IPSCswere reversibly blocked by focal application of strychnine.
DiscussionThere are four major findings from the present study: (1)Heart rate increases during inspiration and decreases duringexpiration in conscious, unrestrained rats. (2) CPNs in thebrainstem are inhibited during inspiration by an increase inboth GABAergic and glycinergic synaptic inputs. (3) Therespiratory-related increase in GABAergic activity, but notglycinergic activity, is mediated by the endogenous activationof a4b2 nicotinic ACh receptors. (4) Prenatal nicotine expo-sure significantly exaggerates the GABA-mediated, but notglycine-mediated, inhibition of CPNs during inspiration.
RSA Pattern in RatsIt is well established in many species (including neonatal4 andadult5,6 humans, baboons,7 dogs,26 seals,9 and rabbits8) thatthe heart rate increases during inspiration and decreases
Figure 3. a4b2 nicotinic receptors mediate the inspiratory-related GABAergic inhibition of CPNs. GABAergic IPSCs were isolated byfocal application of the NMDA, non-NMDA, and glycine receptor antagonists AP-5, CNQX, and strychnine, respectively. a, Inspiratoryactivity evoked a significant increase (P,0.01) in the frequency of GABAergic IPSCs in CPNs. b, This inspiratory-related increase inGABA synaptic frequency was abolished by focal application of the nicotinic receptor antagonist DHbE at a concentration selective forthe a4b2 receptor subtype (3 mmol/L). DHbE did not significantly change GABA synaptic frequency between bursts. c and d, Gabazine(25 mmol/L) reversibly blocked all IPSCs under these recording conditions. Representative traces in panels a through d are from thesame CPN. e, Values are mean6SEM from 6 cells. *P,0.05 and **P,0.01.
Neff et al Mechanism of Respiratory Sinus Arrhythmia 5
by guest on June 30, 2018http://circres.ahajournals.org/
Dow
nloaded from
during inspiration. This RSA improves the efficiency ofpulmonary gas exchange by better matching ventilation andpulmonary blood flow.1,2,9 This inspiratory-relatedtachycardia is predominantly mediated by a reduction incardiac vagal activity due to the decreased activity of CPNs inthe NA.2,3,8,15,27–30However, data describing cardiorespira-tory interactions in the rat are contradictory. A recent in vivostudy has paradoxically shown that in contrast to all otherstudied species, the activity of CPNs is enhanced duringinspiration in rats.31 However, there are several factors thatmake the results from that study difficult to interpret. Theseinclude a desynchronization of central respiratory activityfrom lung inflation and using anesthetics, which in generalreduce or eliminate parasympathetic cardiac activity. Incontrast, a recent study using the rat working heart brainstempreparation (a preparation without anesthetics) demonstrates
that heart rate increases during inspiration, consistent with astudy in other species.32 The results from the present studyunequivocally demonstrate that similar to the RSA thatoccurs in other species, heart rate increases during inspirationin conscious, freely moving rats.
Inhibitory Respiratory-Related Synaptic Inputsto CPNsPrevious in vivo work has shown that CPNs are inhibitedduring inspiration via a chloride-mediated current.15 Thepresent study demonstrates that CPNs receive an increasedfrequency of both GABAergic and glycinergic IPSCs duringinspiration. This respiratory-dependent inhibition of CPNactivity provides a cellular mechanism for the tachycardiathat occurs during the inspiratory phase of the respiratorycycle.
Figure 4. CPNs are inhibited during inspiration by an increased frequency of glycinergic IPSCs, which is not mediated by the activationof nicotinic receptors. Glycinergic IPSCs were isolated by focal application of the NMDA, non-NMDA, and GABAA receptor antagonistsAP-5, CNQX, and gabazine, respectively. a, Inspiratory activity evoked a significant increase (P,0.05) in glycinergic IPSCs in CPNs. band c, This inspiratory-related increase in IPSC frequency was not significantly altered by focal application of a high concentration (100mmol/L) of the nicotinic receptor antagonist DHbE. d and e, Glycine antagonist strychnine reversibly blocked all IPSCs under these re-cording conditions. Representative traces in panel a and panels b through e are from 2 CPNs. f, Values are mean6SEM from 7 cells.Strx indicates strychnine. *P,0.05.
6 Circulation Research September 19, 2003
by guest on June 30, 2018http://circres.ahajournals.org/
Dow
nloaded from
Involvement of Nicotinic Receptors in RSAThe GABA-mediated inhibition of CPNs during inspirationwas significantly inhibited by curare, indicating that theincrease in GABAergic frequency is mediated by the activa-tion of nicotinic receptors. Further investigation revealed thatthis increase in GABAergic frequency was unaffected byaBTX, demonstrating that it was not mediated by the activa-tion of the a7 nAChR subtype. However, ana4b2-selectiveconcentration of DHbE abolished the GABAergic inhibitionof CPNs during inspiration, demonstrating that activation ofa4b2 nAChRs by endogenous ACh is essential for theGABAergic component of this cardiorespiratory interaction.The facilitation of GABAergic inputs to CPNs by activationof nicotinic receptors is consistent with previous work, whichhas shown that spontaneous GABAergic synaptic inputs toCPNs are enhanced by the activation ofa4b2 receptorslocated in the presynaptic terminals of GABAergic neurons.33
The present study also indicates that the nicotinic receptorsresponsible for the increased GABAergic activity duringinspiration are in proximity to the CPNs, inasmuch as the
focal application of the nicotinic antagonist DHbE abolishedthis increase.
In contrast, greater (100mmol/L) concentrations of DHbEdid not significantly alter the respiratory-related increase inglycinergic synaptic frequency in CPNs. Interestingly, previ-ous studies have shown that spontaneous glycinergic inputs toCPNs are also enhanced by the activation ofa4b2 nicotinicreceptors in glycinergic presynaptic terminals.16 This sug-gests that although glycinergic inputs to CPNs possesspresynaptic nicotinic receptors, the respiratory-evoked in-crease in glycinergic synaptic input to CPNs is not mediatedby the activation of nicotinic receptors. Alternatively, theremay be nicotinic modulation of glycinergic activity thatoccurs at a location distant from the CPNs and out of therange of the focal application of drugs in the present study.
Prenatal Nicotine Exposure andRespiratory-Related Synaptic Inputs to CPNsIn animals prenatally exposed to nicotine, the inspiratory-related increase in GABAergic synaptic frequency was nearly
Figure 5. Prenatal nicotine exposure exaggerates the inspiratory-related increase in GABAergic, but not glycinergic, synaptic inputs toCPNs. a, In animals prenatally exposed to nicotine, CPNs received increased GABAergic synaptic input during inspiration. b, In unex-posed animals, the frequency of GABAergic IPSCs increased 260690% during inspiration. In animals prenatally exposed to nicotine,the frequency of GABAergic IPSCs increased 460640% during inspiration, which is a significant augmentation (P,0.05) of thisresponse. Representative traces in panels a and b are from 3 CPNs. c, Values are mean6SEM from 22 control and 18 prenatal nicotineexperiments. **P,0.01. d, GABAergic IPSC amplitude is significantly increased by prenatal nicotine exposure both between and duringinspiratory bursts. Unexp indicates the group not exposed to nicotine; Pre Nic, the group exposed to prenatal nicotine. Values aremean6SEM from 22 control and 18 prenatal nicotine experiments. *P,0.05. e, Values are mean6SEM percent increase of theinspiratory-related increase in GABAergic (n522 Unexp cells, n518 Pre Nic cells) and glycinergic (n513 Unexp cells, n511 Pre Niccells) IPSCs in CPNs during inspiration. *P,0.05. Prenatal nicotine exposure significantly exaggerates the inspiratory-related increase inGABA frequency. In contrast, the inspiratory-related increase in glycinergic frequency is not significantly altered by prenatal nicotineexposure.
Neff et al Mechanism of Respiratory Sinus Arrhythmia 7
by guest on June 30, 2018http://circres.ahajournals.org/
Dow
nloaded from

twice that of unexposed animals, whereas the inspiratory-dependent increase in glycinergic frequency was not signifi-cantly different in control and nicotine-exposed animals. Thisis consistent with the results that nicotinic antagonists abol-ished the inspiratory-related increase in GABAergic fre-quency to CPNs but did not alter the respiratory modulationof glycinergic IPSCs to CPNs. In addition to enhancing theincrease in frequency of GABAergic synaptic inputs to CPNsduring inspiration, prenatal nicotine exposure also caused asignificant increase in the amplitude of both spontaneous andinspiratory-evoked GABAergic synaptic inputs to CPNs.
Other studies have shown that thea4b2 nAChR subtype issignificantly upregulated in rat brains chronically exposed tonicotine34,35 and thata4b2 receptors chronically exposed tonicotine exhibit enhanced responses to ACh and are lesssensitive to desensitization.36 A greater number of, enhancedresponses to, and reduced desensitization ofa4b2 nicotinicreceptors may be responsible for the prenatal nicotine-induced exaggeration of thea4b2-mediated increase inGABAergic synaptic frequency in CPNs during inspiration.
The a4b2 receptor antagonist DHbE (3 mmol/L) signifi-cantly inhibited the inspiratory-related increase in GABAer-gic frequency in animals prenatally exposed to nicotine,indicating that activation ofa4b2 nicotinic receptors byendogenous ACh also mediates this cardiorespiratory inter-action in animals prenatally exposed to nicotine. In addition,a4b2 block significantly reduced the frequency of spontane-ous, non–inspiratory-related GABAergic synaptic inputs toCPNs in animals prenatally exposed to nicotine but not inunexposed animals. This suggests that prenatal nicotineaugments endogenous cholinergic control of GABAergicinputs to CPNs in animals prenatally exposed to nicotine.
These alterations in cardiorespiratory control with prenatalnicotine exposure may be clinically important. Maternalcigarette smoking is highly correlated with SIDS, and it hasbeen suggested that SIDS is caused by an alteration ofbrainstem sites responsible for cardiorespiratory con-trol.17,19,37,38Infants that subsequently succumb to SIDS haveheart rates higher than those in other infants.39 The exagger-ated amplitude of all GABAergic inputs to CPNs and en-hanced increase in GABAergic synaptic frequency duringinspiration observed the present study would be expected toevoke an elevated basal heart rate and a greater than normaltachycardia during inspiration. This nicotine-mediated facil-itation of GABAergic neurotransmission to CPNs providespossible mechanisms for the elevated heart rate in SIDSvictims and the elevated heart rate prevalent in smokers.
In summary, rats possess normal RSA, and CPNs areinhibited during inspiration by an increase in the frequency ofboth GABAergic and glycinergic synaptic inputs. Theinspiratory-related increase in GABAergic synaptic fre-quency is mediated by the activation ofa4b2 nAChRs and issignificantly exaggerated by prenatal exposure to nicotine.These data show a neurochemical link between the neuronsessential for respiration and neurons that control heart rateand reveal a physiological role of endogenous ACh releaseand the activation of nicotinic receptors in the generation ofRSA.
AcknowledgmentsThis study was supported by NIH National Heart, Lung, and BloodInstitute grants HL-72006 and HL-59895 to Dr Mendelowitz. DrNeff was supported by the Jocelyn Beard Moran Memorial Fellow-ship from the American Heart Association, Mid-Atlantic affiliate.
References1. Hayano J, Yasuma F, Okada A, Mukai S, Fujinami T. Respiratory sinus
arrhythmia: a phenomenon improving pulmonary gas exchange and cir-culatory efficiency.Circulation. 1996;94:842–847.
2. Loewy AD, Spyer KM, eds.Central Regulation of Autonomic Functions.New York, NY: Oxford University Press; 1990.
3. Taylor EW, Jordan D, Coote JH. Central control of the cardiovascular andrespiratory systems and their interactions in vertebrates.Physiol Rev.1999;79:855–916.
4. Hathorn MK. Respiratory sinus arrhythmia in new-born infants.J Physiol. 1987;385:1–12.
5. Eckberg DL. Human sinus arrhythmia as an index of vagal cardiacoutflow. J Appl Physiol. 1983;54:961–966.
6. Hirsch JA, Bishop B. Respiratory sinus arrhythmia in humans: howbreathing pattern modulates heart rate.Am J Physiol. 1981;241:H620–H629.
7. Myers MM, Fifer W, Haiken J, Stark RI. Relationships between breathingactivity and heart rate in fetal baboons.Am J Physiol. 1990;258:R1479–R1485.
8. Jordan D, Khalid ME, Schneiderman N, Spyer KM. The location andproperties of preganglionic vagal cardiomotor neurones in the rabbit.Pflugers Arch. 1982;395:244–250.
9. Castellini MA, Rea LD, Sanders JL, Castellini JM, Zenteno-Savin T.Developmental changes in cardiorespiratory patterns of sleep-associatedapnea in northern elephant seals.Am J Physiol. 1994;267:R1294–R1301.
10. Machado BH, Brody MJ. Role of the nucleus ambiguus in the regulationof heart rate and arterial pressure.Hypertension. 1988;11:602–607.
11. McAllen RM, Spyer KM. The location of cardiac vagal preganglionicmotoneurones in the medulla of the cat.J Physiol. 1976;258:187–204.
12. Mendelowitz D. Advances in parasympathetic control of heart rate andcardiac function.News Physiol Sci. 1999;14:155–161.
13. Smith JC, Ellenberger HH, Ballanyi K, Richter DW, Feldman JL. Pre-Botzinger complex: a brainstem region that may generate respiratoryrhythm in mammals.Science. 1991;254:726–729.
14. Mendelowitz D. Firing properties of identified parasympathetic cardiacneurons in nucleus ambiguus.Am J Physiol. 1996;271:H2609–H2614.
15. Gilbey MP, Jordan D, Richter DW, Spyer KM. Synaptic mechanismsinvolved in the inspiratory modulation of vagal cardio-inhibitoryneurones in the cat.J Physiol. 1984;356:65–78.
16. Wang J, Wang X, Irnaten M, Venkatesan P, Evans C, Baxi S, Mende-lowitz D. Endogenous acetylcholine and nicotine activation enhancesGABAergic and glycinergic inputs to cardiac vagal neurons.J Neuro-physiol. 2003;89:2473–2481.
17. Slotkin TA, Saleh JL, McCook EC, Seidler FJ. Impaired cardiac functionduring postnatal hypoxia in rats exposed to nicotine prenatally: impli-cations for perinatal morbidity and mortality, and for sudden infant deathsyndrome.Teratology. 1997;55:177–184.
18. Taylor JA, Sanderson M. A reexamination of the risk factors for thesudden infant death syndrome.J Pediatr. 1995;126:887–891.
19. Meny RG, Carroll JL, Carbone MT, Kelly DH. Cardiorespiratory record-ings from infants dying suddenly and unexpectedly at home.Pediatrics.1994;93:44–49.
20. Cote A, Hum C, Brouillette RT, Themens M. Frequency and timing ofrecurrent events in infants using home cardiorespiratory monitors.J Pediatr. 1998;132:783–789.
21. Rekling JC, Feldman JL. PreBotzinger complex and pacemaker neurons:hypothesized site and kernel for respiratory rhythm generation.Annu RevPhysiol. 1998;60:385–405.
22. Mendelowitz D, Kunze DL. Identification and dissociation of cardiovas-cular neurons from the medulla for patch clamp analysis.Neurosci Lett.1991;132:217–221.
23. Wang J, Irnaten M, Venkatesan P, Evans C, Baxi S, Mendelowitz D.Synaptic activation of hypoglossal respiratory motorneurons during inspi-ration in rats.Neurosci Lett. 2002;332:195–199.
24. Alkondon M, Albuquerque EX. Diversity of nicotinic acetylcholinereceptors in rat hippocampal neurons, I: pharmacological and functionalevidence for distinct structural subtypes.J Pharmacol Exp Ther. 1993;265:1455–1473.
8 Circulation Research September 19, 2003
by guest on June 30, 2018http://circres.ahajournals.org/
Dow
nloaded from

25. Slotkin TA. Fetal nicotine or cocaine exposure: which one is worse?J Pharmacol Exp Ther. 1998;285:931–945.
26. Warner MR, deTarnowsky JM, Whitson CC, Loeb JM. Beat-by-beatmodulation of AV conduction, II: autonomic neural mechanisms.Am JPhysiol. 1986;251:H1134–H1142.
27. Kollai M, Koizumi K. Reciprocal and non-reciprocal action of the vagaland sympathetic nerves innervating the heart.J Auton Nerv Syst. 1979;1:33–52.
28. Kunze DL. Reflex discharge patterns of cardiac vagal efferent fibres.J Physiol. 1972;222:1–15.
29. Anrep G, Pascual F, Rossler R. Respiratory variations of the heart rate, I:the reflex mechanism of the respiratory sinus arrhythmia.Proc R SocLond B Biol Sci. 1936;119:191–217.
30. Anrep G, Pascual F, Rossler R. Respiratory variations of the heart rate, II:the central mechanism of the respiratory sinus arrhythmia and the inter-relations between the central and the reflex mechanisms.Proc R Soc LondB Biol Sci. 1936;119:218–232.
31. Rentero N, Cividjian A, Trevaks D, Pequignot JM, Quintin L, McAllenRM. Activity patterns of cardiac vagal motoneurons in rat nucleusambiguus.Am J Physiol. 2002;283:R1327–R1334.
32. Pickering AE, Waki H, Headley PM, Paton JF. Investigation of systemicbupivacaine toxicity using the in situ perfused working heart-brainstempreparation of the rat.Anesthesiology. 2002;97:1550–1556.
33. Wang J, Irnaten M, Mendelowitz D. Characteristics of spontaneous andevoked GABAergic synaptic currents in cardiac vagal neurons in rats.Brain Res. 2001;889:78–83.
34. Peng X, Gerzanich V, Anand R, Whiting PJ, Lindstrom J. Nicotine-induced increase in neuronal nicotinic receptors results from a decrease inthe rate of receptor turnover.Mol Pharmacol. 1994;46:523–530.
35. Flores CM, Rogers SW, Pabreza LA, Wolfe BB, Kellar KJ. A subtype ofnicotinic cholinergic receptor in rat brain is composed ofa4 and b2subunits and is up-regulated by chronic nicotine treatment.MolPharmacol. 1992;41:31–37.
36. Buisson B, Bertrand D. Chronic exposure to nicotine upregulates thehumana4b2 nicotinic acetylcholine receptor function.J Neurosci. 2001;21:1819–1829.
37. Nachmanoff DB, Panigrahy A, Filiano JJ, Mandell F, Sleeper LA,Valdes-Dapena M, Krous HF, White WF, Kinney HC. Brainstem3H-nicotine receptor binding in the sudden infant death syndrome.J Neu-ropathol Exp Neurol. 1998;57:1018–1025.
38. St-John WM, Leiter JC. Maternal nicotine depresses eupneic ventilationof neonatal rats.Neurosci Lett. 1999;267:206–208.
39. Schechtman VL, Raetz SL, Harper RK, Garfinkel A, Wilson AJ, SouthallDP, Harper RM. Dynamic analysis of cardiac R-R intervals in normalinfants and in infants who subsequently succumbed to the sudden infantdeath syndrome.Pediatr Res. 1992;31:606–612.
Neff et al Mechanism of Respiratory Sinus Arrhythmia 9
by guest on June 30, 2018http://circres.ahajournals.org/
Dow
nloaded from

Robert A. Neff, Jijiang Wang, Sunit Baxi, Cory Evans and David MendelowitzRespiratory Modulation of Brainstem Cardioinhibitory Parasympathetic Neurons
Respiratory Sinus Arrhythmia. Endogenous Activation of Nicotinic Receptors Mediates
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2003 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
published online August 7, 2003;Circ Res.
http://circres.ahajournals.org/content/early/2003/08/07/01.RES.0000090361.45027.5B.citationWorld Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on June 30, 2018http://circres.ahajournals.org/
Dow
nloaded from