enzymes: the catalysts of life - warner pacific collegeclasspages.warnerpacific.edu/bdupriest/bio...
TRANSCRIPT

© 2012 Pearson Education, Inc.
Lectures by
Kathleen Fitzpatrick Simon Fraser University
Enzymes:
The Catalysts
of Life
Chapter 6

© 2012 Pearson Education, Inc.
Activation Energy and the
Metastable State
• Many thermodynamically feasible reactions in a cell
that could occur do not proceed at any appreciable
rate
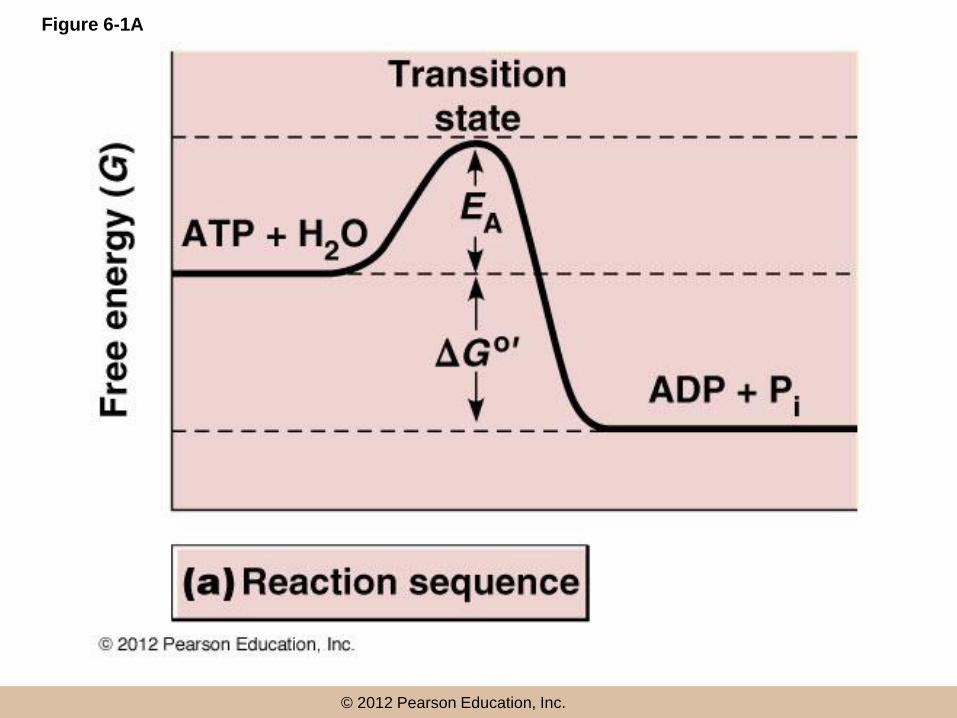
• For example, the hydrolysis of ATP has G = –7.3
kcal/mol
• ATP + H2O ADP + Pi
• However, ATP dissolved in water remains stable for several days

© 2012 Pearson Education, Inc.
Before a Chemical Reaction Can
Occur, the Activation Energy Barrier
Must Be Overcome
• Molecules that could react with one another often
do not because they lack sufficient energy
• Each reaction has a specific activation energy, EA
• EA: the minimum amount of energy required before
collisions between the reactants will give rise to
products

© 2012 Pearson Education, Inc.
Transition state
• Reactants need to reach an intermediate
chemical stage called the transition state
• The transition state has a higher free energy
than that of the initial reactants
© 2012 Pearson Education, Inc.
Figure 6-1A

© 2012 Pearson Education, Inc.
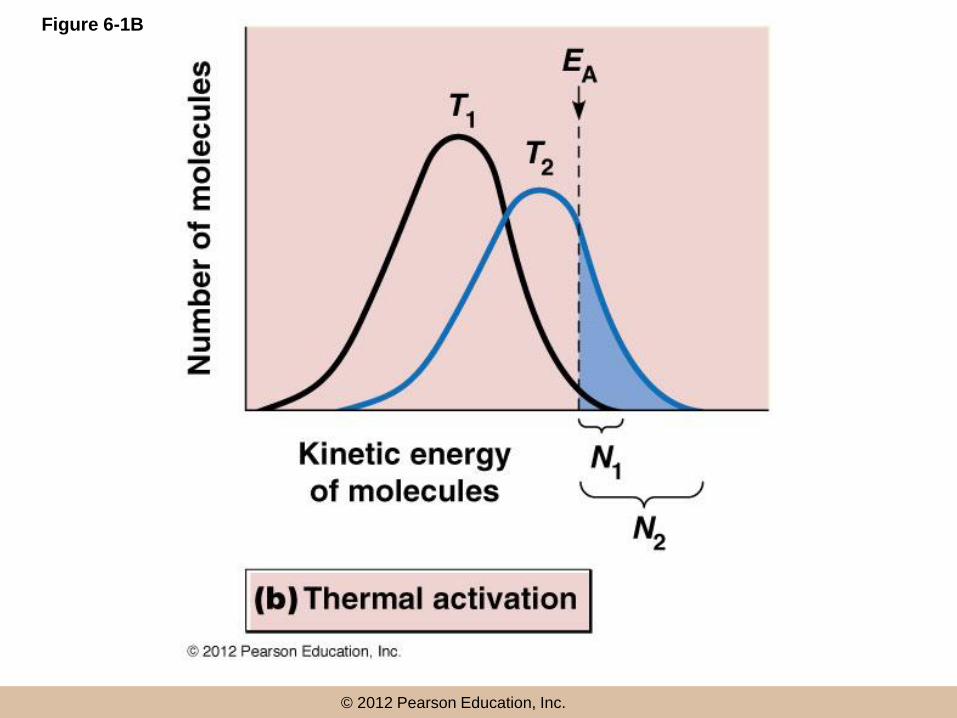
Activation energy barrier
• The rate of a reaction is always proportional
to the fraction of molecules with an energy
equal to or greater than EA
• The only molecules that are able to react at a
given time are those with enough energy to
exceed the activation energy barrier, EA
© 2012 Pearson Education, Inc.
Figure 6-1B

© 2012 Pearson Education, Inc.
The Metastable State Is a Result of
the Activation Barrier
• For most reactions at normal cell temperature, the
activation energy is so high that few molecules
can exceed the EA barrier
• Reactants that are thermodynamically unstable,
but lack sufficient EA, are said to be in a
metastable state
• Life depends on high EAs that prevent most
reactions in the absence of catalysts

© 2012 Pearson Education, Inc.
Catalysts Overcome the Activation
Energy Barrier
• The EA barrier must be overcome in order for
needed reactions to occur
• This can be achieved by either increasing the
energy content of molecules or by lowering
the EA requirement

© 2012 Pearson Education, Inc.
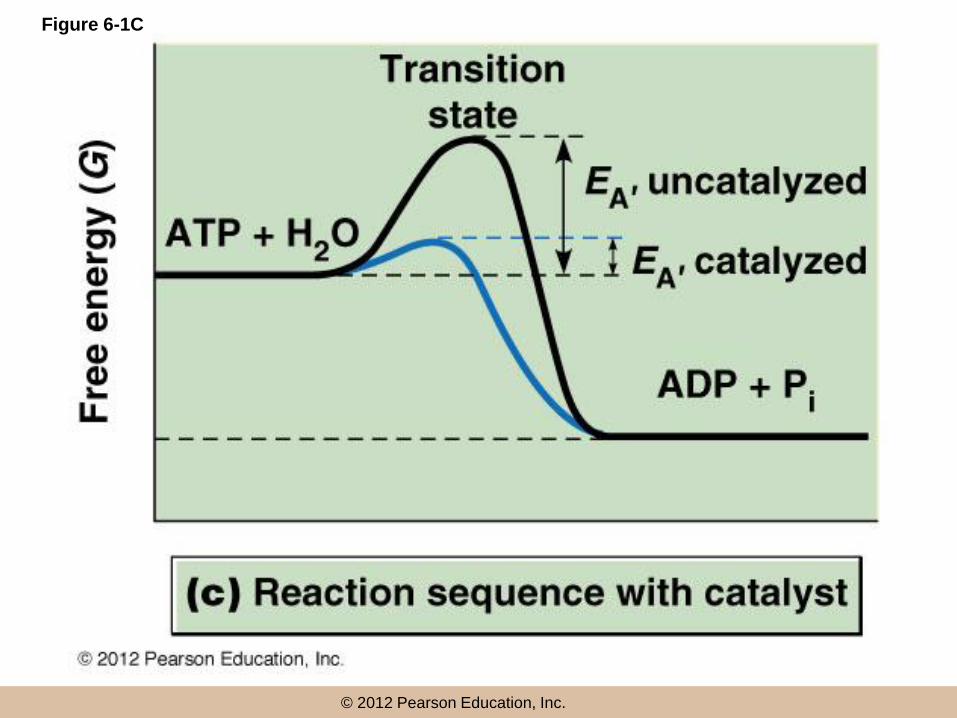
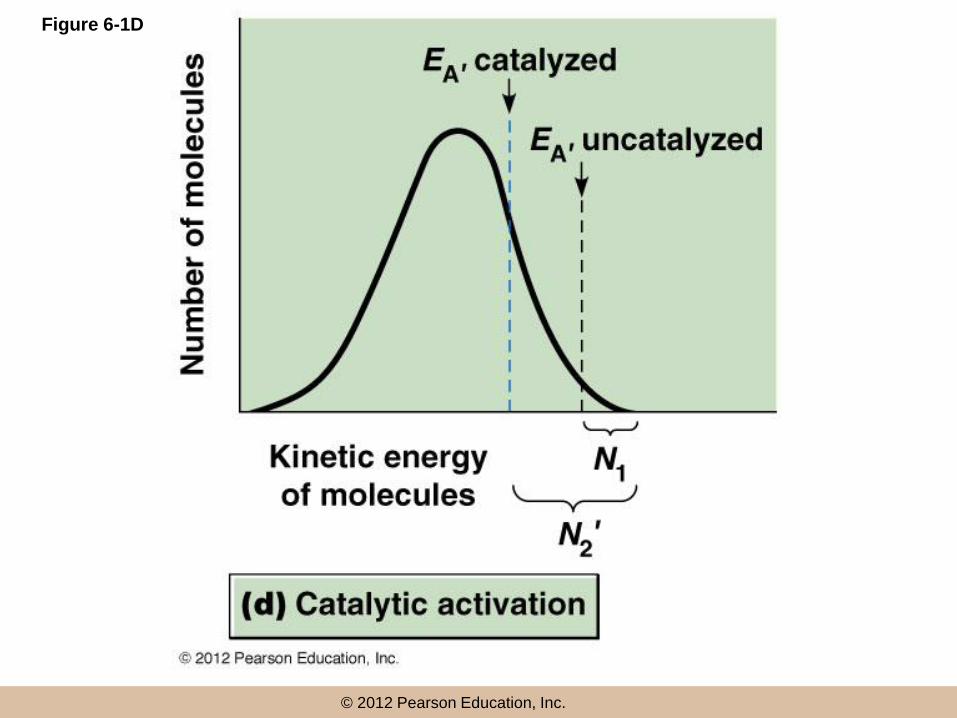
Lowering activation energy
• If reactants can be bound on a surface and
brought close together, their interaction will be
favored and the required EA will be reduced
• A catalyst enhances the rate of a reaction by
providing such a surface and effectively lowering
EA
• Catalysts themselves proceed through the
reaction unaltered
© 2012 Pearson Education, Inc.
Figure 6-1C
© 2012 Pearson Education, Inc.
Figure 6-1D

© 2012 Pearson Education, Inc. © 2012 Pearson Education, Inc.
An increase in temperature increases the rate
at which a spontaneous reaction occurs in a
test tube because _____.
a. an increase in temperature lowers the energy of activation (EA)
b. an increase in temperature makes all molecules more reactive
c. an increase in temperature increases the proportion of molecules that have sufficient kinetic energy to react
d. an increase in temperature acts like a catalyst

© 2012 Pearson Education, Inc. © 2012 Pearson Education, Inc.
A catalyst increases the rate of a reaction
by _____.
a. lowering EA and thus making G more negative
b. lowering EA without having any effect on G
c. lowering EA and thus shifting the equilibrium in favor of a negative G
d. lowering EA and thus increasing the chance that reactants will collide

© 2012 Pearson Education, Inc.
Enzymes as Biological Catalysts
• All catalysts share three basic properties
– They increase reaction rates by lowering the
EA required
– They form transient, reversible complexes with
substrate molecules
– They change the rate at which equilibrium is
achieved, not the position of the equilibrium
• Organic catalysts are enzymes

© 2012 Pearson Education, Inc.
The Active Site
• Every enzyme contains a characteristic cluster of amino acids that forms the active site
• This results from the three dimensional folding of the protein, and is where substrates bind and catalysis takes place
• The active site is usually a groove or pocket that accommodates the intended substrate(s) with high affinity
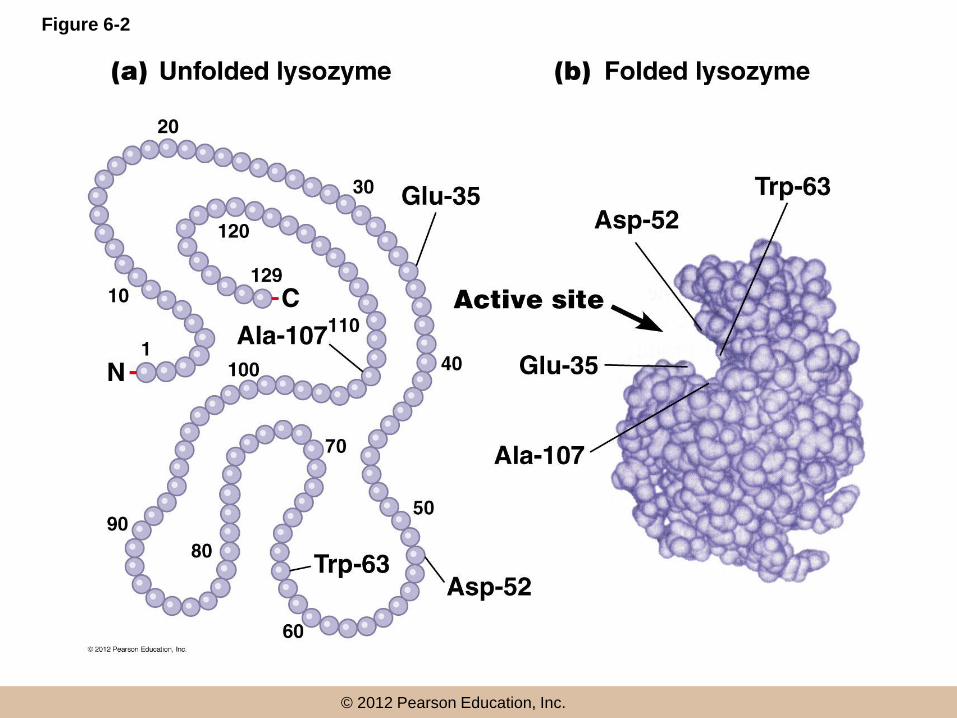
© 2012 Pearson Education, Inc.
Figure 6-2

© 2012 Pearson Education, Inc.
Cofactors
• Some enzymes contain nonprotein cofactors needed for catalytic activity, often because they function as electron acceptors
• These are called prosthetic groups and are usually metal ions or small organic molecules called coenzymes
• Coenzymes are derivatives of vitamins

© 2012 Pearson Education, Inc.
Enzyme Specificity
• Due to the shape and chemistry of the active
site, enzymes have a very high substrate
specificity
• Inorganic catalysts are very nonspecific whereas
similar reactions in biological systems generally
have a much higher level of specificity
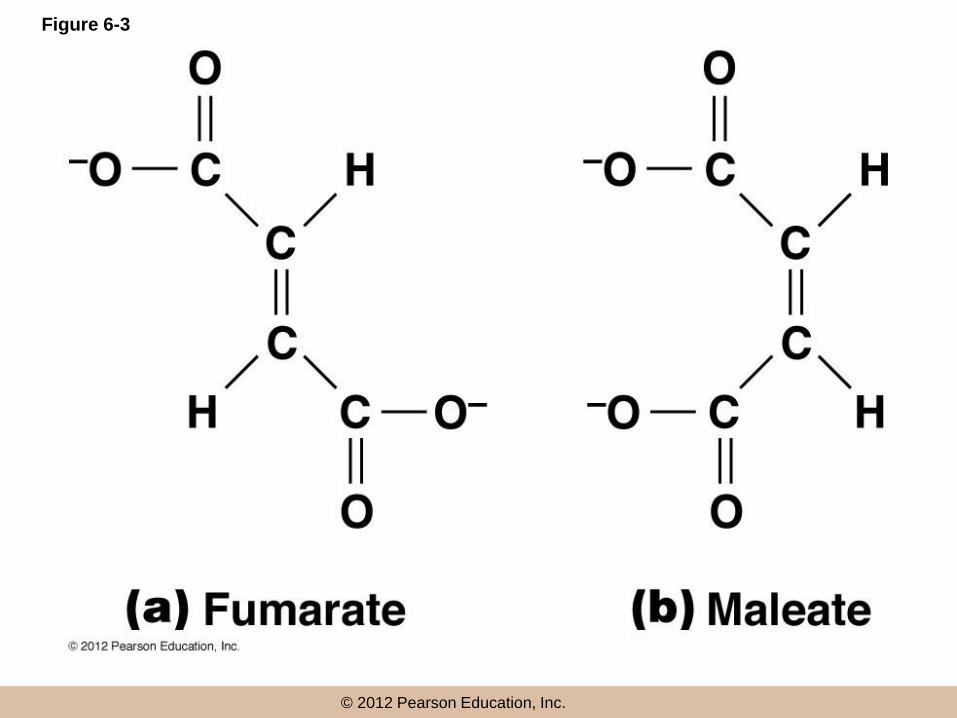
© 2012 Pearson Education, Inc.
Figure 6-3

© 2012 Pearson Education, Inc.
Group specificity
• Some enzymes will accept a number of closely
related substrates
• Others accept any of an entire group of
substrates sharing a common feature
• This group specificity is most often seen in
enzymes involved in degradation of polymers

© 2012 Pearson Education, Inc. © 2012 Pearson Education, Inc.
The active site of an enzyme is important
because it _____.
a. provides a small compartment with a higher temperature, allowing reactants to have enough energy to react
b. is altered by each reaction, explaining why cells must continuously take in energy and food to synthesize new proteins
c. provides a reactive surface to which products bind tightly
d. provides a reactive surface that lowers EA

© 2012 Pearson Education, Inc. © 2012 Pearson Education, Inc.
Enzymes exhibit more specificity than
inorganic catalysts because _____.
a. enzymes are genetically determined
b. enzymes operate over a narrower
range of temperatures than inorganic
catalysts
c. inorganic catalysts are able to catalyze
a much wider range of redox reactions
d. the shape and chemistry of the active
site of an enzyme restricts the
molecules that can bind to it

© 2012 Pearson Education, Inc.
Enzyme Diversity and Nomenclature
• Thousands of different enzymes have been
identified, with enormous diversity
• Names have been given to enzymes based on
substrate (protease, ribonuclease, amylase), or
function (trypsin, catalase)
• Under the Enzyme Commission (EC), enzymes are
divided into six major classes based on general
function
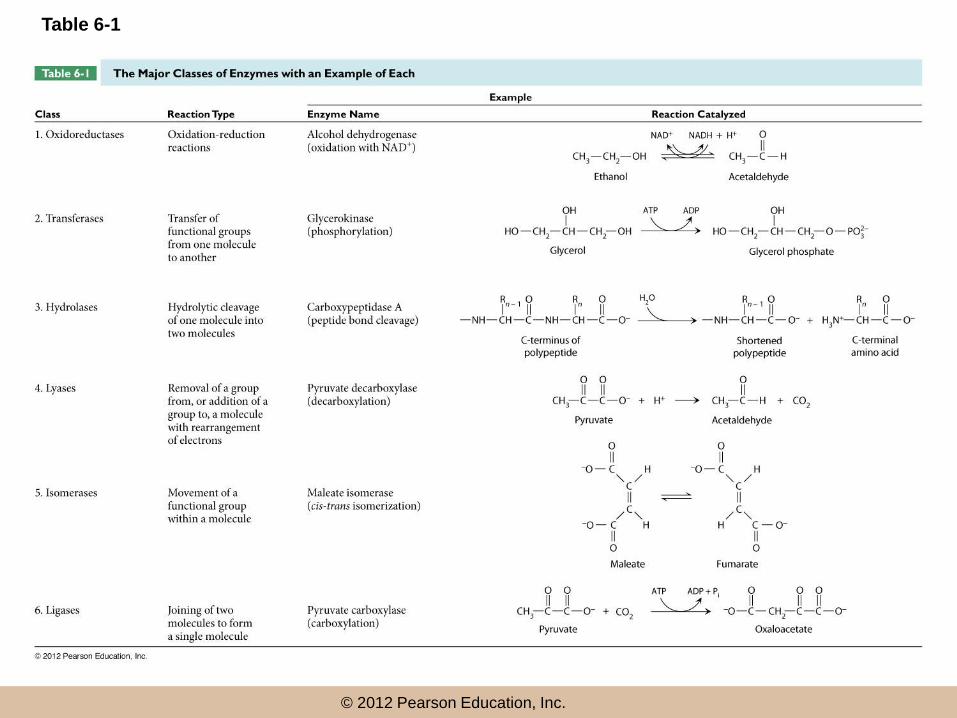
© 2012 Pearson Education, Inc.
Table 6-1
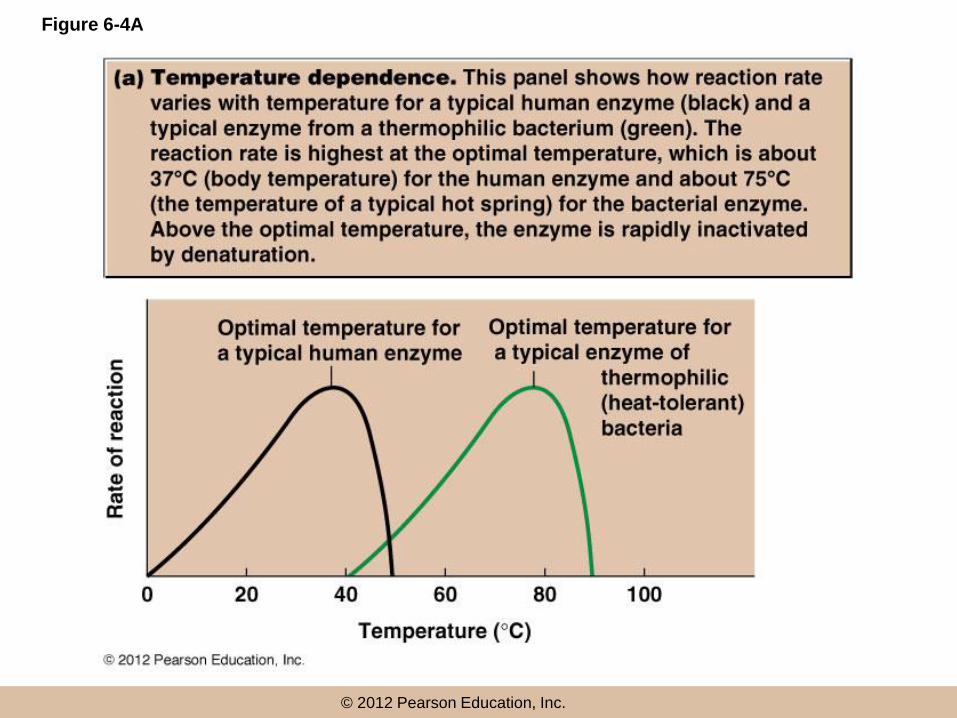
© 2012 Pearson Education, Inc.
Figure 6-4A

© 2012 Pearson Education, Inc.
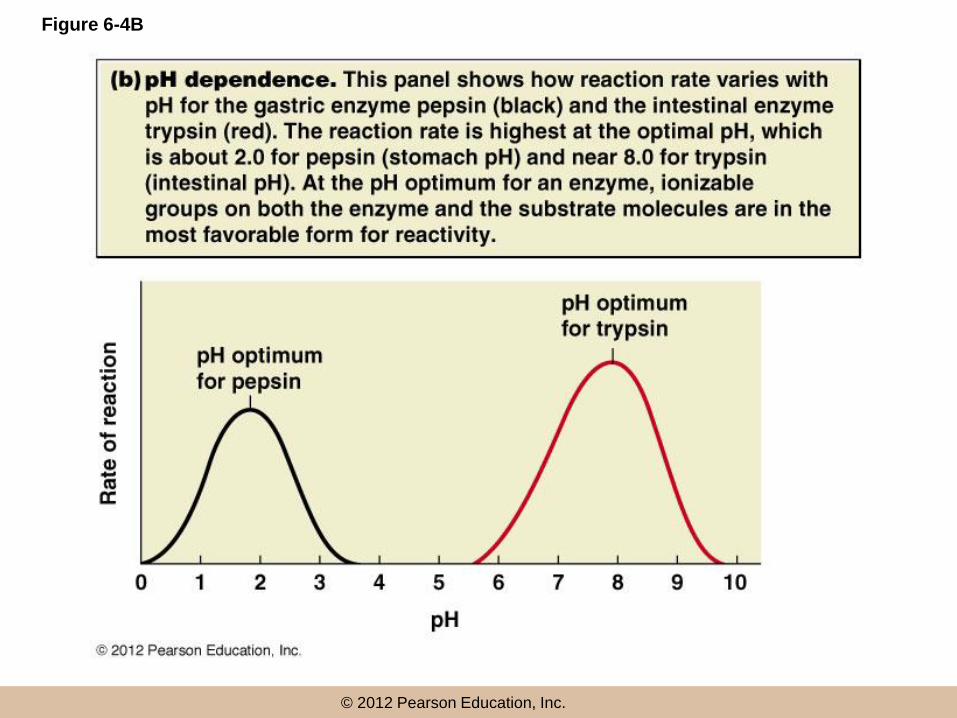
Sensitivity to pH
• Most enzymes are active within a pH range of
about 3–4 units
• pH dependence is usually due to the presence of
charged amino acids at the active site or on the
substrate
• pH changes affect the charge of such residues,
and can disrupt ionic and hydrogen bonds
© 2012 Pearson Education, Inc.
Figure 6-4B

© 2012 Pearson Education, Inc.
Sensitivity to Other Factors
• Enzymes are sensitive to factors such as molecules
and ions that act as inhibitors or activators
• Most enzymes are also sensitive to ionic strength of
the environment
– affects hydrogen bonding and ionic interactions
needed to maintain tertiary conformation

© 2012 Pearson Education, Inc.
Substrate Binding, Activation, and
Catalysis Occur at the Active Site
• Because of the precise chemical fit between
the active site of the enzyme and its
substrates, enzymes are highly specific

© 2012 Pearson Education, Inc.
Substrate Binding
• Once at the active site, the substrate molecules
are bound to the enzyme surface in the right
orientation to facilitate the reaction
• Substrate binding usually involves hydrogen
bonds, ionic bonds, or both
• Substrate binding is readily reversible

© 2012 Pearson Education, Inc.
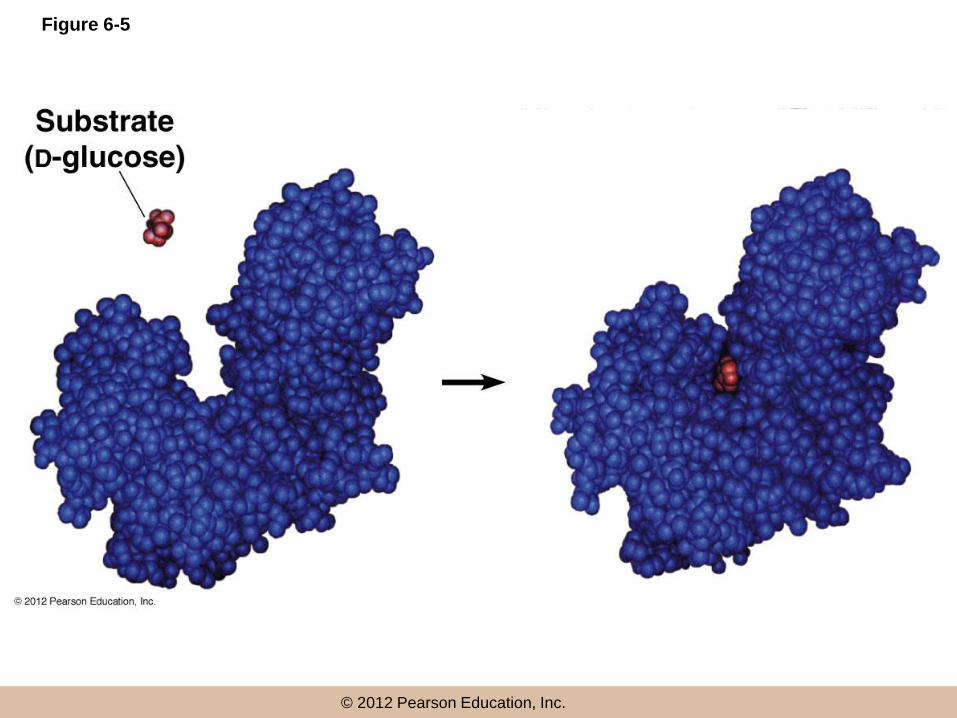
The induced-fit model
• In the past, the enzyme was seen as rigid, with
the substrate fitting into the active site like a key
in a lock (lock-and-key model)
• A more accurate view is the induced-fit model,
in which substrate binding at the active site
induces a conformational change in the shape of
the enzyme
© 2012 Pearson Education, Inc.
Figure 6-5


© 2012 Pearson Education, Inc.
Substrate Activation
• The role of the active site is to recognize and
bind the appropriate substrate and also to
activate it by providing the right environment
for catalysis
• This is called substrate activation, which
proceeds via several possible mechanisms

© 2012 Pearson Education, Inc.
Three common mechanisms of
substrate activation
• Bond distortion, making it more susceptible to
catalytic attack
• Proton transfer, which increases reactivity of
substrate
• Electron transfer, resulting in temporary covalent
bonds between enzyme, substrate

© 2012 Pearson Education, Inc.
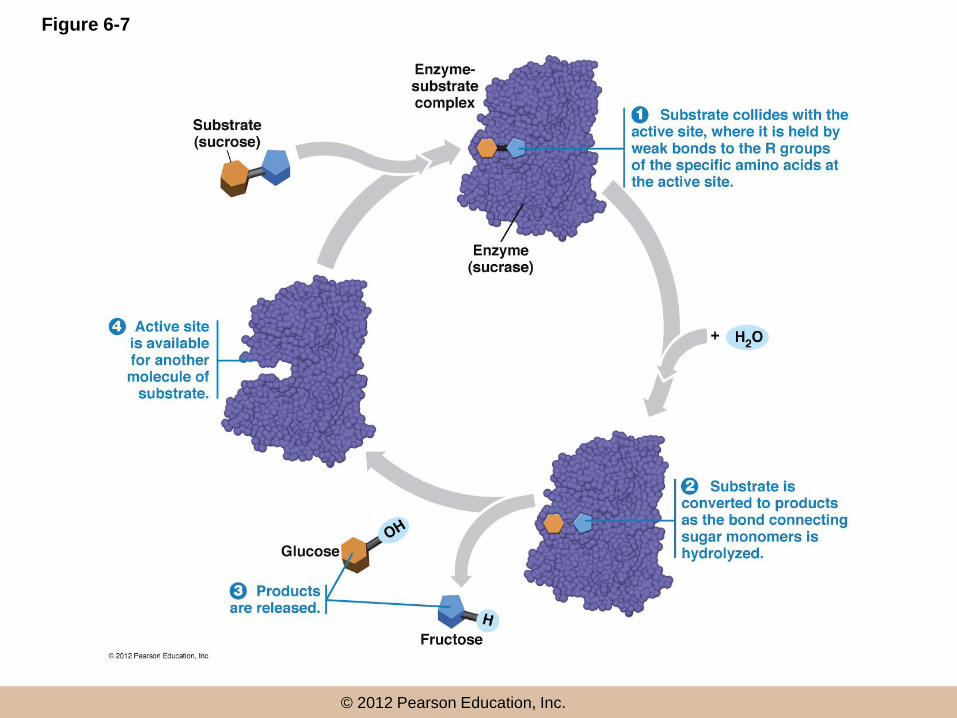
The Catalytic Event
• The sequence of events
– 1. The random collision of a substrate
molecule with the active site results in it
binding there
– 2. Substrate binding induces a conformational
change that tightens the fit, facilitating the
conversion of substrate into products

© 2012 Pearson Education, Inc.
The Catalytic Event (continued)
• The sequence of events
– 3. The products are then released from the
active site
– 4. The enzyme molecule returns to the original
conformation with the active site available for
another molecule of substrate
© 2012 Pearson Education, Inc.
Figure 6-6
© 2012 Pearson Education, Inc.
Figure 6-7

© 2012 Pearson Education, Inc. © 2012 Pearson Education, Inc.
When an enzyme binds to a substrate and
“activates” it, activation means that _____.
a. the enzyme has increased the kinetic energy in the substrate, making it more likely to react
b. the enzyme has lowered G for the reaction
c. the induced fit has subjected the substrate to a chemical environment that lowers EA
d. the enzyme has undergone a conformational change, consistent with the induced fit model

© 2012 Pearson Education, Inc.
Enzyme Kinetics
• Enzyme kinetics describes the quantitative
aspects of enzyme catalysis and the rate of
substrate conversion into products
• Reaction rates are influenced by factors such
as the concentrations of substrates, products,
and inhibitors

© 2012 Pearson Education, Inc.
Initial reaction rates
• Initial reaction rates are measured over a brief
time, during which the substrate concentration
has not yet decreased enough to affect the rate
of reaction

© 2012 Pearson Education, Inc.
Most Enzymes Display Michaelis–
Menten Kinetics
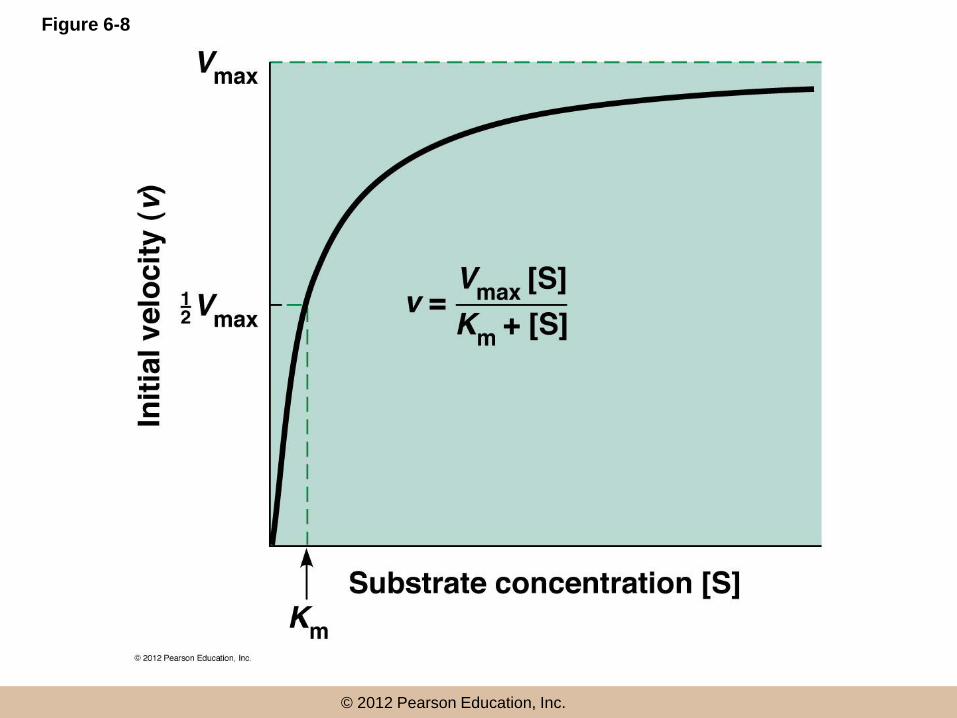
• Initial reaction velocity (v), the rate of change in product concentration per unit time, depends on the substrate concentration [S]
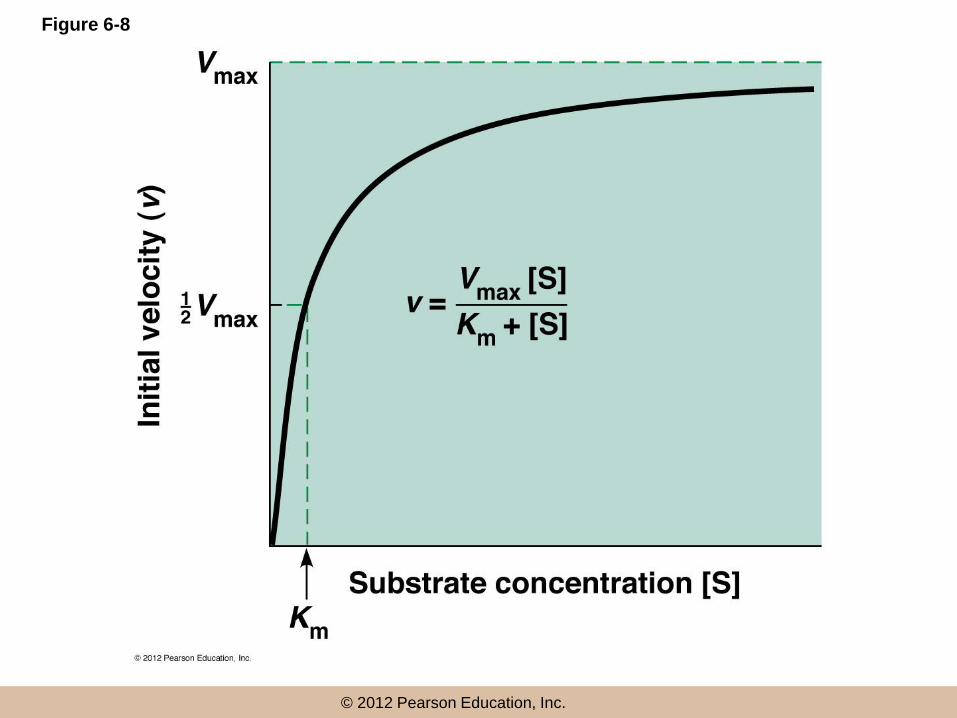
© 2012 Pearson Education, Inc.
Figure 6-8

© 2012 Pearson Education, Inc. © 2012 Pearson Education, Inc.
To find Vmax for an enzyme, it’s important to
measure the initial velocity because _____.
a. enzymes can catalyze both the forward and backward reaction as the ratio of products:reactants increases
b. the initial velocity is the only one that comes from the induced fit caused by the enzyme
c. the initial velocity is the only one directly related to G for the reaction
d. degradation of the enzyme as the reaction proceeds makes the reaction progressively slower, artificially lowering Vmax

© 2012 Pearson Education, Inc.
The Michaelis–Menten Equation
• Michaelis and Menten postulated a theory of
enzyme action
• Enzyme E first reacts with the substrate, to
form a transient complex, ES
• ES then undergoes the catalytic reaction to
generate E and P

© 2012 Pearson Education, Inc.
The Michaelis–Menten Equation
(continued)
•
• The above model, under steady state conditions
gives the Michaelis–Menten equation
•
• Km (the Michaelis constant) = the concentration of
substrate that gives half maximum velocity
© 2012 Pearson Education, Inc.
Figure 6-8

© 2012 Pearson Education, Inc.
What Is the Meaning of Vmax and
Km?
• We can understand the relationship between
v and [S], and the meaning of Vmax and Km by
considering three cases regarding [S]

© 2012 Pearson Education, Inc.
Case 1: Very Low Substrate
Concentration ([S] << Km)
• If [S] << Km
• Then, Km + [S] = [Km]
•
• So at very low [S], the initial velocity of the
reaction is roughly proportional to [S]

© 2012 Pearson Education, Inc.
Case 2: Very High Substrate
Concentration ([S] >> Km)
• If [S] >> Km
• Then, Km + [S] = [S]
•
• So at very high [S], the initial velocity of the reaction
is independent of variation in [S] and Vmax is the
velocity at saturating substrate concentrations

© 2012 Pearson Education, Inc.
Vmax
• Vmax is an upper limit determined by
– The time required for the actual catalytic reaction
– How many enzyme molecules are present
• The only way to increase Vmax is to increase
enzyme concentration
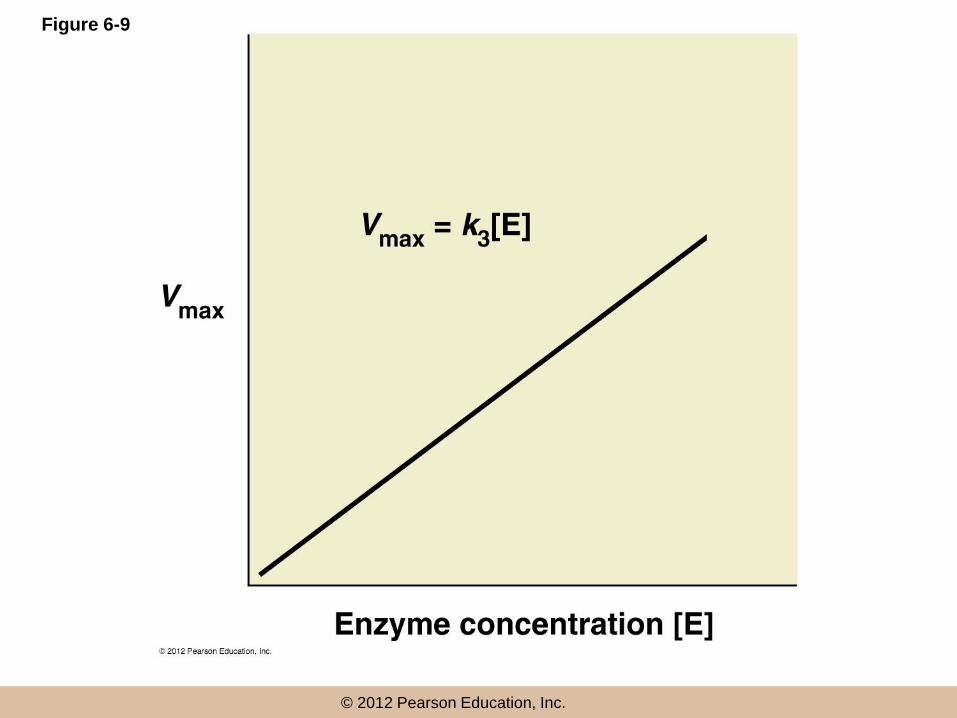
© 2012 Pearson Education, Inc.
Figure 6-9

© 2012 Pearson Education, Inc.
Case 3: ([S] = Km)
• If [S] is equal to Km
• [
• This shows that Km is the specific substrate
concentration at which the reaction proceeds
at one half its maximum velocity

© 2012 Pearson Education, Inc.
Why Are Km and Vmax Important to
Cell Biologists?
• The lower the Km value for a given enzyme and
substrate, the lower the [S] range in which the
enzyme is effective
• Vmax is important, as a measure of the potential
maximum rate of the reaction
• By knowing Vmax, Km, and the in vivo substrate
concentration, we can estimate the likely rate of
the reaction under cellular conditions
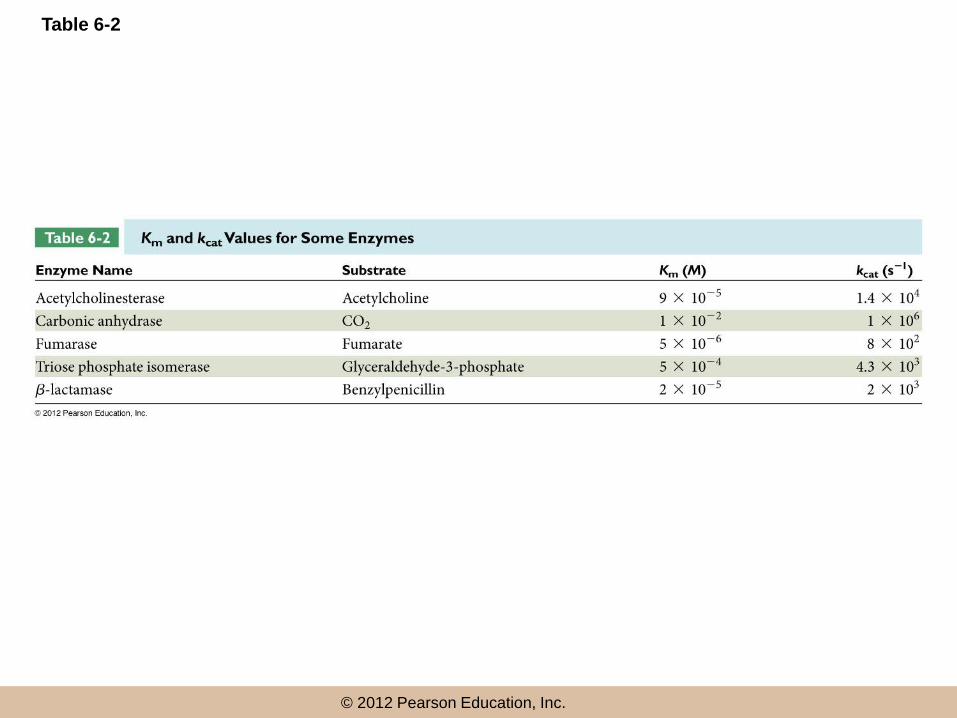
© 2012 Pearson Education, Inc.
Table 6-2

© 2012 Pearson Education, Inc.
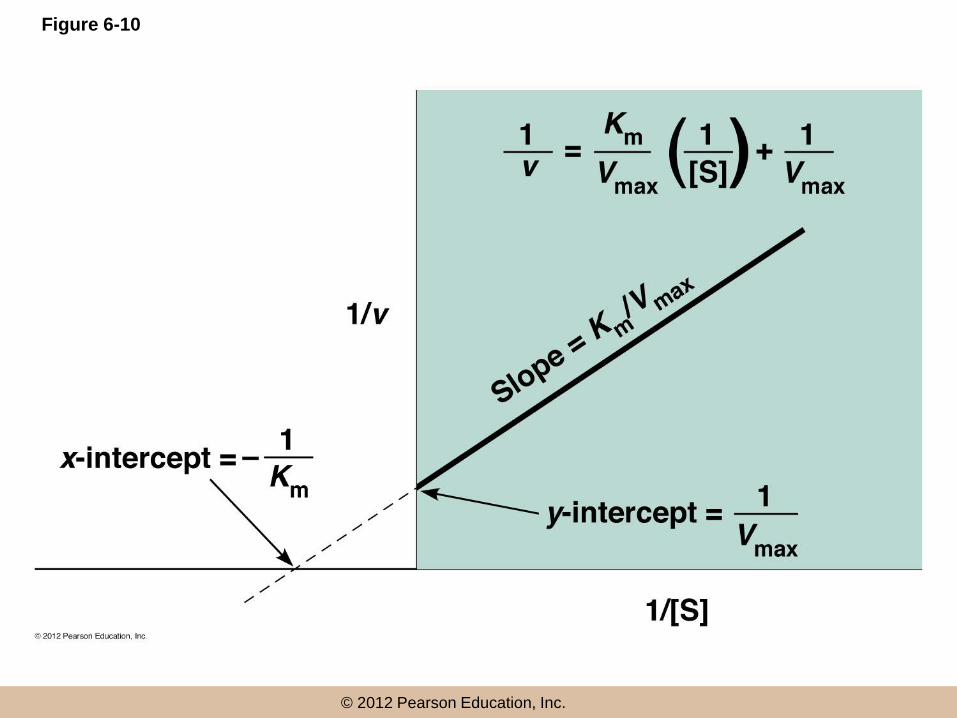
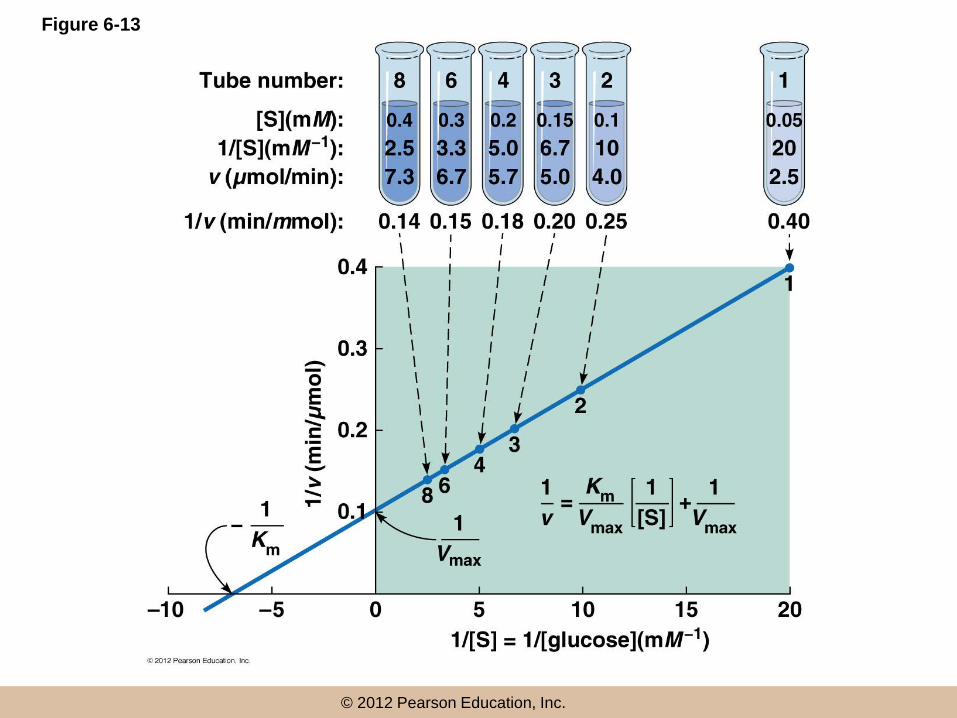
The Double-Reciprocal Plot Is a Useful
Means of Linearizing Kinetic Data
• Lineweaver and Burk inverted both sides of
equation 6-7 to give
•
• This is known as the Lineweaver–Burk equation

© 2012 Pearson Education, Inc.
The double-reciprocal plot
• A plot of 1/v vs 1/[S] is called the double-reciprocal
plot
• This linear plot takes the general form of y = mx + b,
where m is the slope and b the y-intercept
• The slope is Km/Vmax, the y-intercept is 1/Vmax, and
the x-intercept is –1/Km
© 2012 Pearson Education, Inc.
Figure 6-10

© 2012 Pearson Education, Inc.
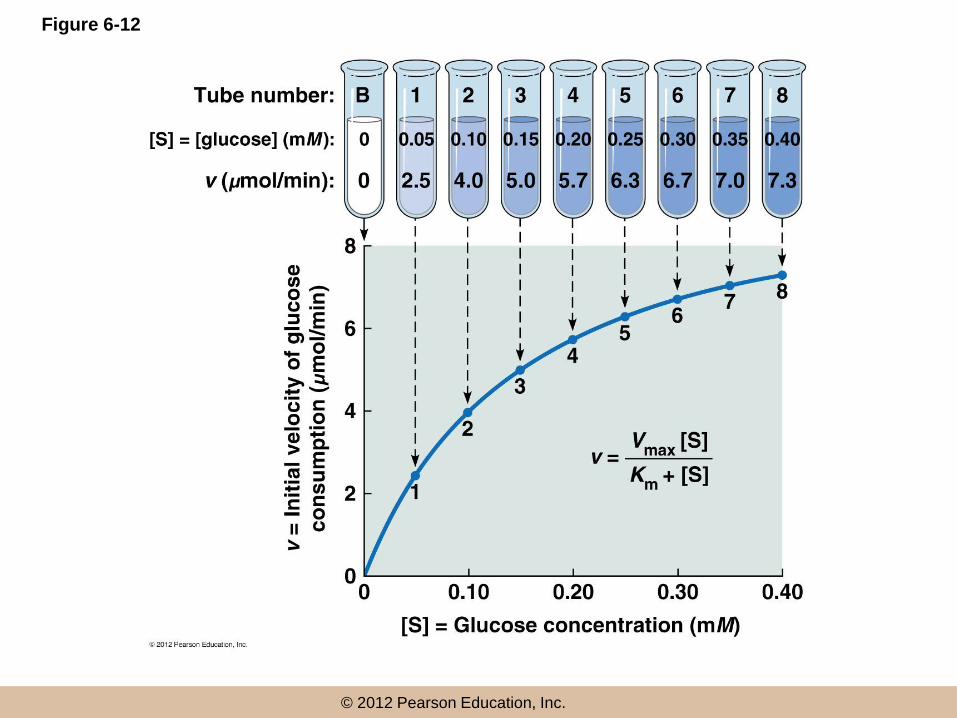
Determining Km and Vmax: An Example
• Consider the following reaction, important in energy
metabolism:
– glucose + ATP → glucose-6-phosphate + ADP
• To analyze this reaction, begin by determining initial
velocity at several substrate concentrations
• For two substrates, they must be varied one at a time,
with saturating levels of the other
hexokinase
© 2012 Pearson Education, Inc.
Figure 6-12
© 2012 Pearson Education, Inc.
Figure 6-13

© 2012 Pearson Education, Inc.
Enzyme Inhibitors Act Either
Irreversibly or Reversibly
• Enzymes are influenced (mostly inhibited) by
products, alternative substrates, substrate
analogs, drugs, toxins, and allosteric effectors
• The inhibition of enzyme activity plays a vital
role as a control mechanism in cells
• Drugs and poisons frequently exert their effects
by inhibition of specific enzymes

© 2012 Pearson Education, Inc.
Inhibitors important to enzymologists
• Inhibitors of greatest use to enzymologists are
substrate analogs and transition state analogs

© 2012 Pearson Education, Inc.
Reversible and irreversible inhibition
• Irreversible inhibitors, which bind the enzyme
covalently, cause permanent loss of catalytic
activity and are generally toxic to cells
– For example, heavy metal ions, nerve gas poisons,
some insecticides
• Reversible inhibitors bind enzymes
noncovalently and can dissociate from the
enzyme

© 2012 Pearson Education, Inc.
Reversible inhibition (continued)
• The fraction of enzyme available for use in a cell
depends on the concentration of the inhibitor
and how easily the enzyme and inhibitor can
dissociate
• The two forms of reversible inhibitors are
competitive inhibitors and noncompetitive
inhibitors

© 2012 Pearson Education, Inc.
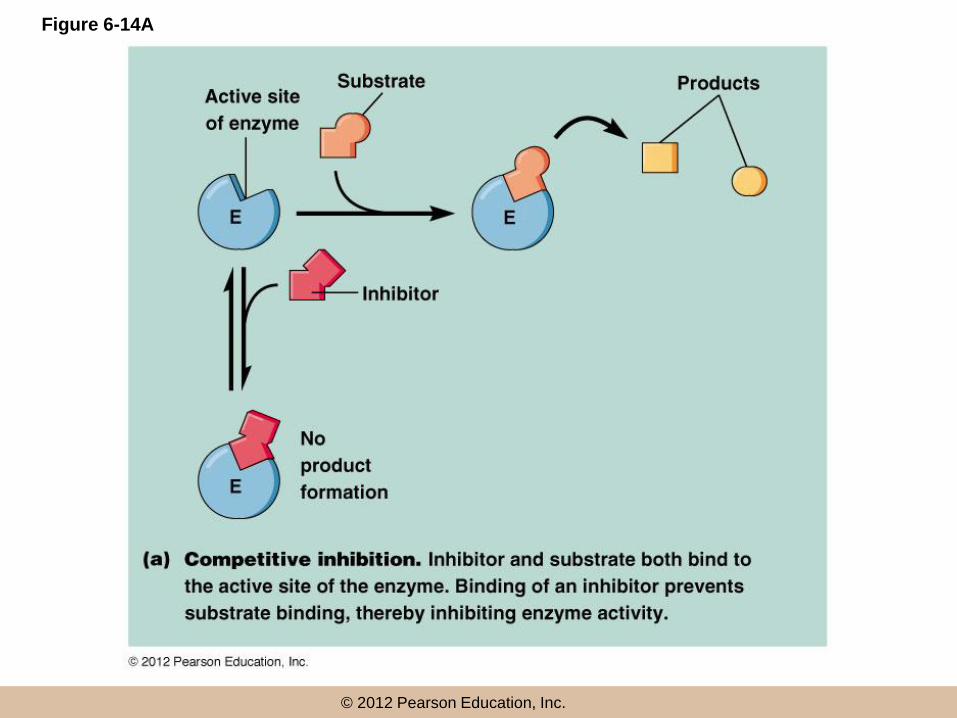
Competitive inhibition
• Competitive inhibitors bind the active site of an
enzyme and so compete with substrate for the
active site
• Enzyme activity is inhibited directly because
active sites are bound to inhibitors, preventing
the substrate from binding
© 2012 Pearson Education, Inc.
Figure 6-14A

© 2012 Pearson Education, Inc.
Noncompetitive inhibition
• Noncompetitive inhibitors bind the enzyme
molecule outside of the active site
• They inhibit activity indirectly by causing a
conformation change in the enzyme that
– Inhibits substrate binding at the active site, or
– Reduces catalytic activity at the active site
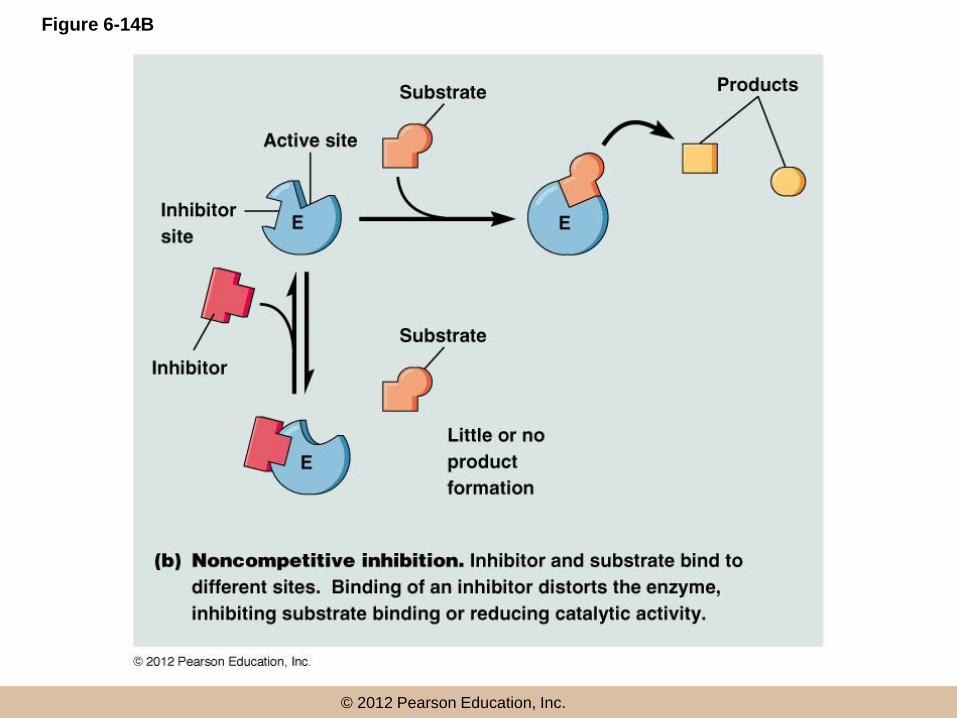
© 2012 Pearson Education, Inc.
Figure 6-14B

© 2012 Pearson Education, Inc. © 2012 Pearson Education, Inc.
How would you expect a competitive inhibitor to
affect enzyme function?
a. By raising the Km without affecting Vmax, because infinite amounts of substrate would wash out the inhibitor.
b. By lowering the Vmax without affecting Km, because the enzyme still binds well to its natural substrate.
c. By lowering the Vmax without affecting Km, because in the presence of the inhibitor, there is essentially less enzyme, and Vmax
is directly proportional to the enzyme concentration.

© 2012 Pearson Education, Inc.
Enzyme Regulation
• Enzyme rates must be continuously adjusted to keep them tuned to the needs of the cell
• Regulation that depends on interactions of substrates and products with an enzyme is called substrate-level regulation
• Increases in substrate levels result in increased reaction rates, whereas increased product levels lead to lower rates

© 2012 Pearson Education, Inc.
Allosteric regulation and covalent
modification
• Cells can turn enzymes on and off as needed by
two mechanisms: allosteric regulation and
covalent modification
• Usually enzymes regulated this way catalyze the
first step of a multi-step sequence
• By regulating the first step of a process, cells are
able to regulate the entire process

© 2012 Pearson Education, Inc.
Allosteric Enzymes Are Regulated
by Molecules Other than Reactants
and Products
• Allosteric regulation is the single most important
control mechanism whereby the rates of
enzymatic reactions are adjusted to meet the cell’s
needs

© 2012 Pearson Education, Inc.
Feedback Inhibition
• It is not in the best interests of a cell for
enzymatic reactions to proceed at the maximum
rate
• In feedback (or end-product) inhibition, the
final product of an enzyme pathway negatively
regulates an earlier step in the pathway
•
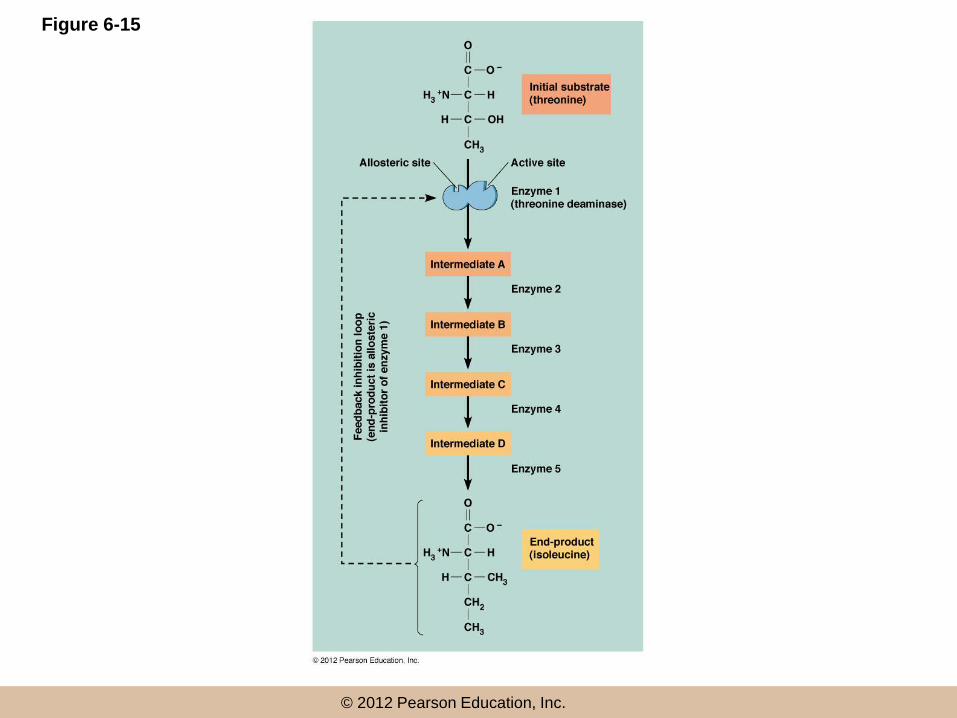
© 2012 Pearson Education, Inc.
Figure 6-15

© 2012 Pearson Education, Inc.
Allosteric Regulation
• Allosteric enzymes have two conformations, one in which it has affinity for the substrate(s) and one in which it does not
• Allosteric regulation makes use of this property by regulating the conformation of the enzyme
• An allosteric effector regulates enzyme activity by binding and stabilizing one of the conformations

© 2012 Pearson Education, Inc.
Allosteric regulation (continued)
• An allosteric effector binds a site called an allosteric (or regulatory) site, distinct from the active site
• The allosteric effector may be an activator or inhibitor, depending on its effect on the enzyme
• Inhibitors shift the equilibrium between the two enzyme states to the low affinity form; activators favor the high affinity form
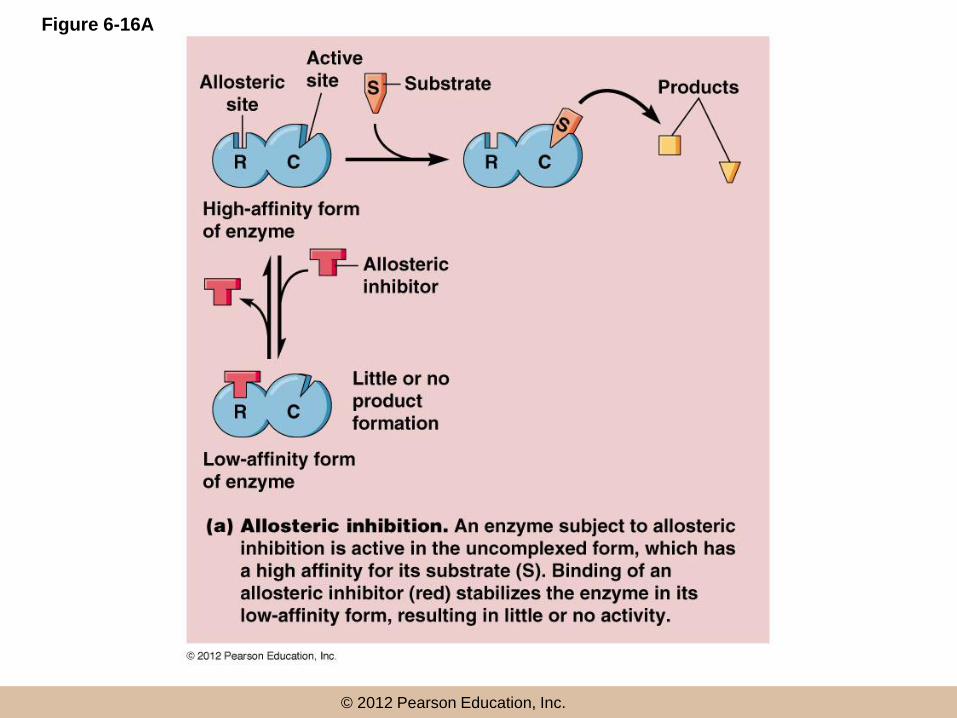
© 2012 Pearson Education, Inc.
Figure 6-16A
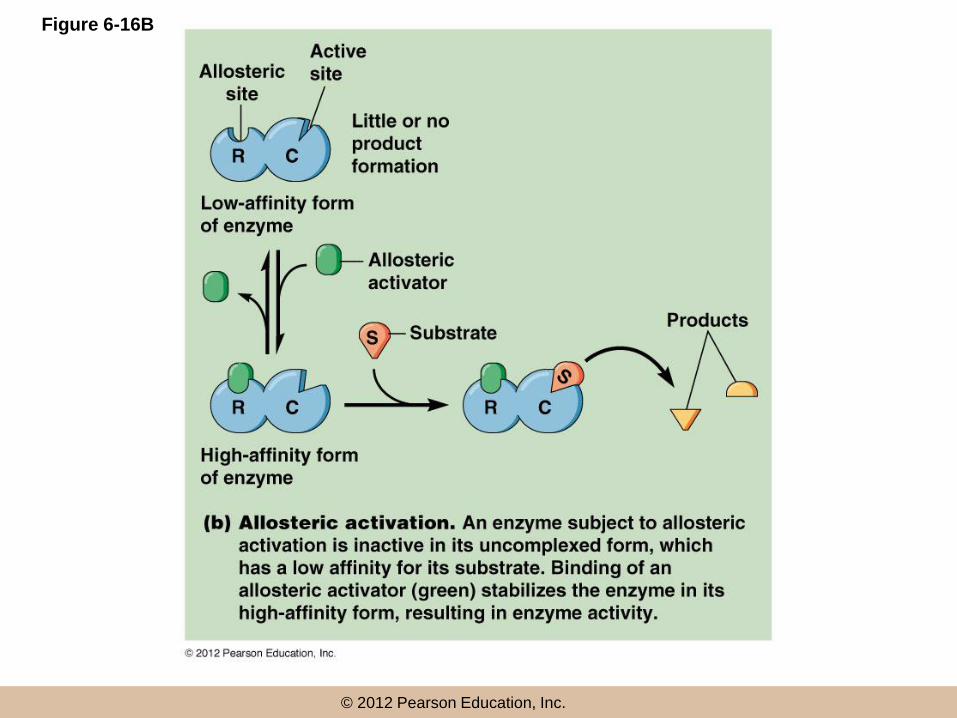
© 2012 Pearson Education, Inc.
Figure 6-16B

© 2012 Pearson Education, Inc.
Allosteric enzymes
• Most allosteric enzymes are large, multisubunit proteins with an active or allosteric site on each subunit
• Active and allosteric sites are on different subunits, the catalytic and regulatory subunits, respectively
• Binding of allosteric effectors alters the shape of both catalytic and regulatory subunits

© 2012 Pearson Education, Inc.
Allosteric Enzymes Exhibit Cooperative
Interactions Between Subunits
• Many allosteric enzymes exhibit cooperativity
• As multiple catalytic sites bind substrate
molecules, the enzyme changes conformation,
which alters affinity for the substrate
• In positive cooperativity the conformation change
increases affinity for substrate; in negative
cooperativity, affinity for substrate is decreased

© 2012 Pearson Education, Inc. © 2012 Pearson Education, Inc.
Which of the following must be true of enzymes
that are regulated allosterically?
a. The enzyme must have at least one domain or subunit that binds to the regulatory compound, and at least one catalytic domain or subunit.
b. The enzyme must never catalyze the reverse reaction.
c. The allosteric regulator may bind to the active site.
d. The enzyme must be part of a biosynthetic pathway (such as one that synthesizes the amino acid tryptophan).

© 2012 Pearson Education, Inc.
Enzymes Can Also Be Regulated by
the Addition or Removal of Chemical
Groups
• Many enzymes are subject to covalent
modification
• Activity is regulated by addition or removal of
groups, such as phosphate, methyl, acetyl
groups, etc.

© 2012 Pearson Education, Inc.
Phosphorylation and
Dephosphorylation
• The reversible addition of phosphate groups is a common covalent modification
• Phosphorylation occurs most commonly by transfer of a phosphate group from ATP to the hydroxyl group of Ser, Thr, or Tyr residues in a protein
• Protein kinases catalyze the phosphorylation of other proteins

© 2012 Pearson Education, Inc.
Dephosphorylation
• Dephosphorylation, the removal of phosphate groups from proteins, is catalyzed by protein phosphatases
• Depending on the enzyme, phosphorylation may be associated with activation or inhibition of the enzyme
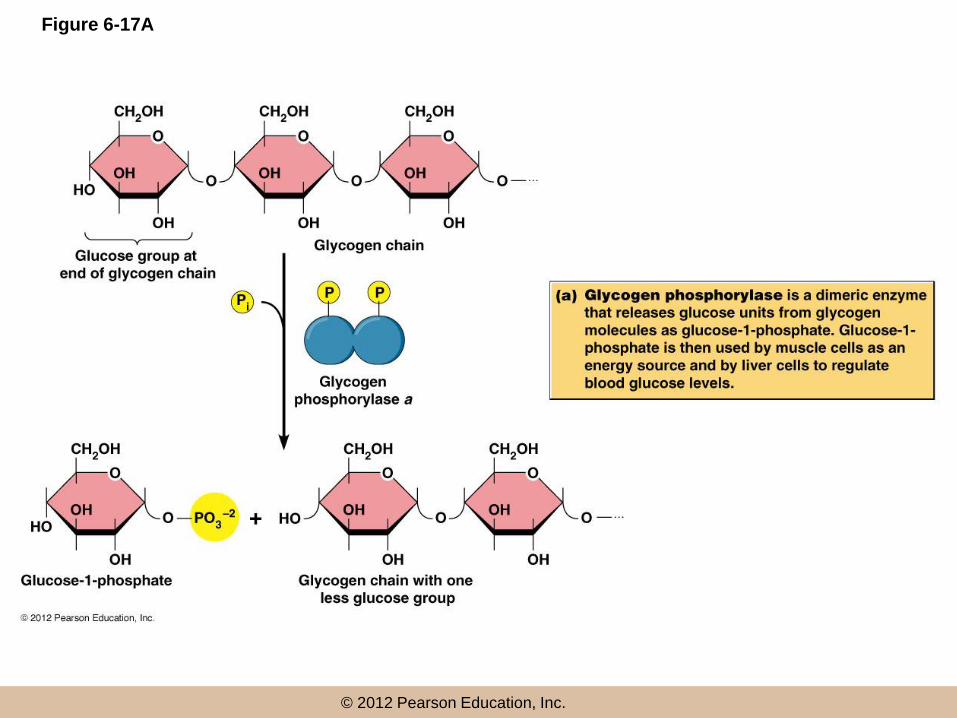
• Fisher and Krebs won the Nobel prize for their work on glycogen phosphorylase
© 2012 Pearson Education, Inc.
Figure 6-17A

© 2012 Pearson Education, Inc.
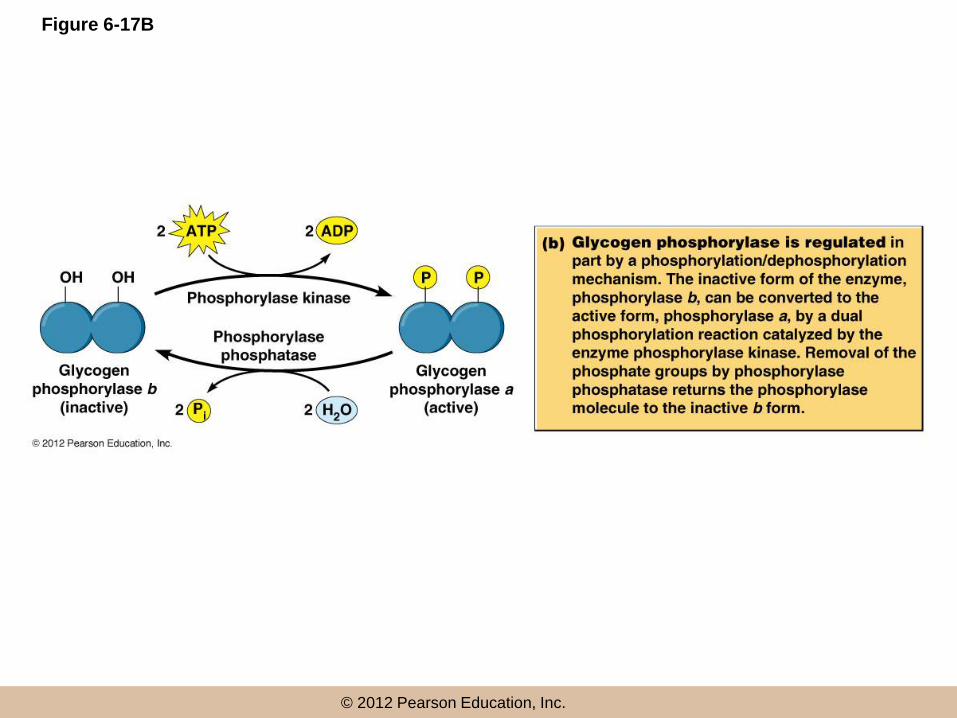
Regulation of glycogen phosphorylase
• Glycogen phosphorylase exists as two inter-convertible forms
– An active, phosphorylated form (glycogen phosphorylase-a)
– An inactive, non-phosphorylated form (glycogen phosphorylase-b)
• The enzymes responsible
– Phosphorylase kinase phosphorylates the enzyme
– Phosphorylase phosphatase removes the phosphate
© 2012 Pearson Education, Inc.
Figure 6-17B

© 2012 Pearson Education, Inc.
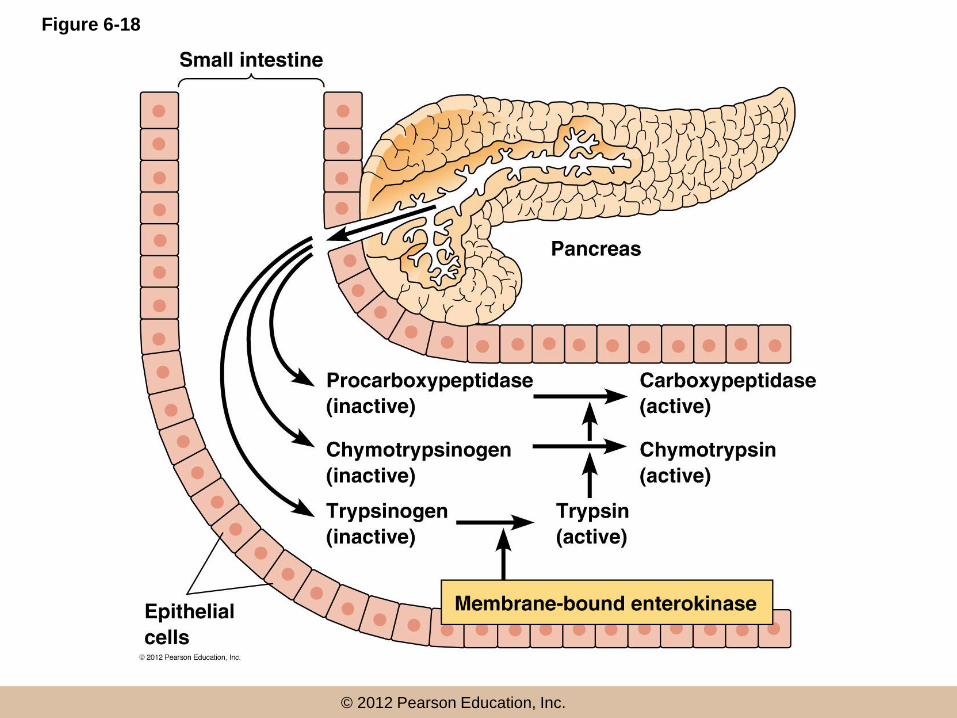
Proteolytic Cleavage
• The activation of a protein by a one-time, irreversible removal of part of the polypeptide chain is called proteolytic cleavage
• Proteolytic enzymes of the pancreas, trypsin, chymotrypsin, and carboxypeptidase, are examples of enzymes synthesized in inactive form (as zymogens) and activated by cleavage as needed
© 2012 Pearson Education, Inc.
Figure 6-18

© 2012 Pearson Education, Inc.
RNA Molecules as Enzymes:
Ribozymes
• Some RNA molecules have been found to have
catalytic activity; these are called ribozymes
• Self-splicing rRNA from Tetrahymena thermophila
and ribonuclease P are examples
• It is thought by some that RNA catalysts predate
protein catalysts, and even DNA

© 2012 Pearson Education, Inc. © 2012 Pearson Education, Inc.
Which of the following steps in the regulation of glycogen
phosphorylase is incorrect?
a. First, glycogen phosphorylase b and ATP bind to the active site of phosphorylase kinase.
b. Next, glycogen phosphorylase catalyzes the transfer of phosphate molecules from ATP onto glycogen phosphorylase, resulting in active glycogen phosphorylase a.
c. Active glycogen phosporylase a catalyzes the breaking down of glycogen.
d. Glycogen phosphorylase a and water bind to the active site of phosphorylase phosphatase, which catalyzes the removal of the phosphate groups from glycogen phosphorylase a.
e. The result is inactive glycogen phosphorylase b.