epigenetic research in cancer epidemiology: trends...
TRANSCRIPT

Minireview
Epigenetic Research in Cancer Epidemiology: Trends,Opportunities, and Challenges
Mukesh Verma1, Scott Rogers1, Rao L. Divi1, Sheri D. Schully1, Stefanie Nelson1, L. Joseph Su1,Sharon A. Ross2, Susan Pilch3, Deborah M. Winn1, and Muin J. Khoury1,4
AbstractEpigenetics is emerging as an important field in cancer epidemiology that promises to provide insights into
gene regulation and facilitate cancer control throughout the cancer care continuum. Increasingly, investigators
are incorporating epigenetic analysis into the studies of etiology andoutcomes. Tounderstand current progress
and trends in the inclusion of epigenetics in cancer epidemiology,we evaluated thepublished literature and the
National Cancer Institute (NCI)–supported research grant awards in this field to identify trends in epigenetics
research. We present a summary of the epidemiologic studies in NCI’s grant portfolio (from January 2005
through December 2012) and in the scientific literature published during the same period, irrespective of
support from the NCI. Blood cells and tumor tissue were the most commonly used biospecimens in these
studies, although buccal cells, cervical cells, sputum, and stool samples were also used. DNA methylation
profiling was the focus of the majority of studies, but several studies also measured microRNA profiles. We
illustrate here the current status of epidemiologic studies that are evaluating epigenetic changes in large
populations. The incorporation of epigenomic assessments in cancer epidemiology studies has and is likely to
continue to provide important insights into the field of cancer research. Cancer Epidemiol Biomarkers Prev; 23(2);
223–33. �2013 AACR.
IntroductionCancer is both a genetic and epigenetic disease (1).
Epigenetics is defined as heritable changes in geneexpression that are not the result of changes in the DNAsequence. Four major epigenetic mechanisms are DNAmethylation, histone modification, nucleosome remo-deling, and microRNAs (miRNAs), which have beenshown to play a role in a number of complex diseases,including cancer (2, 3). DNA methylation in particularhas been extensively assessed in breast, colon, esoph-ageal, lung, pancreas, ovary, prostate, and other cancers(4–12). Through their effects on genomic stability andgene expression, epigenetic changes influence carcino-genesis from initiation through progression, throughouta person’s lifespan, and, in some cases, across genera-tions (13). Epigenetic events that are relevant to cancerrisk are believed to occur early in cancer development,
thus, may serve as potential "first hits" for tumorigen-esis. Epigenetic marks reflect both an individual’s genet-ic background and exposure to different environmentalfactors, and thus may be useful for understanding theimpact of environmental exposures in carcinogenesis(14). Because epigenetic changes occur before or duringearly tumor development, they can be modulated bydiet, drugs, and other external factors such as infectiousagents; epigenetic profiling may provide clues to miti-gate an individual’s risk of cancer (15–17). Mill andHeijmans recently proposed that improved understand-ing of the mechanism of cancer progression can beunderstood by studying epigenetics in populations asa part of an integrated functional genomic study (18).Epigenetic changes in comparison with genetic ones arereversible and are acquired in a gradual manner and thisfeature provides a huge potential for cancer preventionstrategies. In addition, therapies targeting epigeneticmechanisms have been shown to modify or inhibit geneexpression and some have shown modest effects inclinical research settings.
To understand the current state of the field of epige-netics in cancer epidemiology, we evaluated the researchproject grant (RPG) awards funded by the National Can-cer Institute (NCI) and the published literature in PubMedfor trends in epigenetic research in cancer epidemiologyacross the cancer control continuum in studies conductedin human populations. This report presents a summary ofour findings, particularly in the context of studying riskand cancer-relevant exposures, including nutrition and
Authors' Affiliations: 1Epidemiology and Genomics Research Program,Division of Cancer Control and Population Sciences; 2Division of CancerPrevention, National Cancer Institute; 3Office of the Director, InformationResources and Services Branch, NIH, Bethesda, Maryland; and 4Office ofPublic Health Genomics, Centers for Disease Control and Prevention,Atlanta, Georgia
Corresponding Author: Mukesh Verma, Epidemiology and GenomicsResearch Program, Division of Cancer Control and Population Sciences,National Cancer Institute, NIH, 6130 Executive Boulevard, Rockville, MD20852. Phone: 301-594-7344; Fax: 301-435-6609; E-mail:[email protected]
doi: 10.1158/1055-9965.EPI-13-0573
�2013 American Association for Cancer Research.
CancerEpidemiology,
Biomarkers& Prevention
www.aacrjournals.org 223
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573

infectious agents, as well as practical matters such as thetype of cancers being studied, and the methods andtechniques that are both emerging and commonly used.Overall, we sought to present an overview of the progressin the inclusion of epigenetics in cancer epidemiologystudies and to identify scientific questions related toepigenetics that cancer epidemiology can address.
Materials and MethodsCriteria and terms used for identifying cancerepigenetics and epidemiology grants andpublications (search strategy and analysis)
NCI-supported RPGs related to epigenetic epidemiol-ogy funded from January 1, 2005, to December 31, 2012,were included in the portfolio analysis, and the scientificterms used in analyzing grants in different categories areshown in Table 1. The portfolio was analyzed usingNCI’sPortfolio Management Application software version 13.4.Search and selection criteria used for the grant proposal tobe classified as "epigenetic epidemiology" study were asfollows: "One OR more terms from column 1 of Table 1"AND "one ORmore terms from column 2 of Table 1 AND"Human." In addition, the criteria for inclusion of a projectin the analysiswere as follows: (i) the focus of the project iscancer; (ii) study involves human subjects; (iii) focus of atleast one of the specific aims in the project is cancerepigenetics; and (iv) had at least 100 cases. We excludedstudies that focused solely on polymorphisms in genesencoding DNA methyltransferases or miRNAs. Afterapplying these criteria and exclusions, 79 RPGs wereidentified for further analysis. The authors of this report
coded the grant abstracts for and analyzed the data bystudydesign, organ site, biospecimen typeused, exposureevaluated (if applicable), and method/technology usedfor epigenetic analysis.
In addition, we searched the published literature onepigenetic epidemiology in PubMed from January 1,2005, to December 31, 2012, using the following searchcriteria: (epigenesis, genetic[mh] OR epigenomics[mh]OR "DNA methylation" OR methylation[ti] OR "histonemodification" OR CIMP[tiab] OR microRNAs[mh]OR "CpG Islands/genetics"[mh] OR methylation[ti])AND neoplasms[mh] AND (epidemiologic studies[mh]OR risk[mh] OR "population-based"[tiab] OR "oddsratio"[tiab] OR hazard[tiab] OR cohort�[tiab]) ANDHumans[Mesh]. The following elimination criteria wereapplied: (i) studies with less than 100 cancer cases; (ii)studies published before 2005; (iii) experimental studiesin animals; (iv) review articles and articles reportingmeta-analyses; (v) letters, commentaries, editorials, andnews articles; and (vi) studies that are not clearly epi-demiologic. This search yielded 486 publications thatare relevant for analysis. As with the grant analysisoutlined above, authors listed above coded the publica-tions on and analyzed the data by study design, organsite, biospecimen type used, exposure evaluated (ifapplicable), and method/technology used for epigenet-ic analysis. To check all manual coding, each publicationand grant was coded by two different authors and anydiscrepancies were resolved.
ResultsThe type of exposures/modifiers proposed to be exam-
ined in NCI-supported grants and those examined inpublications is shown in Fig. 1. The grant proposalssupported by the NCI most often studied the influenceof nutrition, drugs/treatment, or infectious agents. Othergrants focused on energy balance, exogenous hormones,chemical exposures (e.g., pesticides), and physical expo-sures (e.g., radiation). In the literature, the majority ofpublications explored the effect of nutrition, smoking,drugs and treatments, and infectious agents on epigeneticprocesses.
The results from the literature analysis revealed that thenumber of publications addressing epigenetic changes incancers of the colon were the largest, followed by breastand lung (Fig. 2). The NCI-supported grant portfolioanalysis indicated that in the field of epigenetics, breastcancer was the most frequently studied cancer type (Fig.2). The next two most frequently investigated cancerswere colorectal and lung cancer, in that order.Other organsites examined for epigenetic changes included pancreas,ovary, liver, gastric, and head and neck cancers.
The portfolio analysis found that 67 of 79 grantsplanned to analyze methylation levels, mostly at selectedindividual loci (data not shown). Some of these grantsalso proposed to assessCpG islandmethylator phenotype(CIMP) status, promoter methylation, and microsatelliteinstability. Investigators proposed to assess histone
Table 1. Grant portfolio search and codingcategories
"HUMAN" AND "NEOPLASM" AND one or moreepigenetic terms from the first column and one ormore epidemiology terms from the second column
Epigenetics terms Epidemiology terms
Epigenetics EpidemiologyMethylation Case–controlEpigenome CohortHistone Epidemiology methodsAcetylation Observational studiesImprinting Risk assessmentMicroRNA Causal associationChromatin compaction Association studiesCIMP Nested case–control studies
Clinical trialEtiologyRisk factorsSusceptibilityPrognosisPreventionPopulation-based
Verma et al.
Cancer Epidemiol Biomarkers Prev; 23(2) February 2014 Cancer Epidemiology, Biomarkers & Prevention224
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573

modifications along with methylation levels in cancers ofthe bladder, cervix, and prostate, and also in myelodys-plastic syndrome. Noncoding RNAs, particularly miR-NAs, were the focus of seven studies, with 5 studiesplanned to assess methylation at specific loci along withmiRNAs.Concerning biospecimen types (Fig. 3) used for epige-
netic analysis, tumor tissues (40 grants) and blood (35grants) were the most predominant specimen types col-
lected, although a few grants proposed using buccal cells,paraffin-embedded tissues, plasma, sputum,mouthwash,urine, DNA from blood spot, stool, toenail, and saliva forepigenetic analysis. Thirteen grant proposals used bothblood and tumor tissue specimens for comparison anal-ysis,whereas 22 grant proposals usednormal tissue speci-mens for comparison with tumor specific epigenetic mar-kers. The portfolio analysis also found that 53 of 79 studiesused tumor tissue for epigenetic analysis, whereas 13
0
5
10
15
20
25
30
% o
f To
tal
Exposure/modifier
Grants Publications
Alcoh
ol
Arsen
ic
Che
micals
Dru
g/tre
atm
ent
Endog
enou
s ho
rmon
es
Energ
y ba
lanc
e
Exoge
nous
hor
mon
es
Imm
unolog
ic
Infe
ctious
age
nts
Nut
rition
Physica
l exp
osur
e
Smok
ing/
toba
cco
Oth
ers
Figure 1. Number of NCI grants andPubMed-indexed publications incancer epigenetic epidemiology byexposure evaluated in the study.Different types of exposures areshown in the figure and discussedin the text.
0
5
10
15
20
25
30
35
40
45
% o
f To
tal
Cancer site
Grants Publications
ALL
AM
LBla
dder
Bra
inBre
ast
Cer
vica
lCol
orec
tal
End
omet
rial
Eso
phag
usG
astri
c
Hea
d an
d ne
ckLe
ukem
iaLi
ver
Lung
Ora
lO
varia
n
Ped
iatri
c ge
rm c
ell
Pro
stat
ePan
crea
ticRen
alTh
yroi
d
Figure 2. Number of NCI grants andPubMed-indexed publications incancer epigenetic epidemiology bycancer site. Breast cancer site isthe most studied cancer site in ourgrant portfolio, whereas colorectalcancer site is in publicationanalysis.
Epigenetics in Cancer Epidemiology
www.aacrjournals.org Cancer Epidemiol Biomarkers Prev; 23(2) February 2014 225
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573

studies planned to analyze epigenetic changes simulta-neously in both leukocyte and tumor DNA. Table 2shows the types of biospecimens used for epigeneticanalysis in published cancer epidemiology studies. Sim-ilar to the trend in grant proposals, both tissue samplesand blood were the frequently used biospecimens forepigenetic analysis in published studies (19). The pub-
lished literature also suggested that investigators usedbuccal cells, sputum, urine, cervical swab, and pancre-atic fluid for epigenetic analysis in epidemiologic stud-ies (Fig. 3), albeit less frequently. Several investigatorssought to determine whether blood cells could be usedas a surrogate tissue for examining epigenetic profiles intumor tissue (19, 20).
0
10
20
30
40
50%
of
To
tal
Biospecimen type
Grants Publications
Tissu
e an
d bloo
d
Bucca
l cells
Bronc
heal la
vage
Bone
mar
row
Cer
vica
l swab
Exfoliate
d ce
lls
Nor
mal tiss
ue
Pancr
eatic
fluid
Pleur
al e
ffusion
Plasm
a
PBMC
Saliva
Sputu
mSto
ol
Toe n
ail
Tumor
tiss
ue
Urin
e
Figure 3. Number of NCI grants andPubMed-indexed publications incancer epigenetic epidemiology bybiospecimen used in the study. Forepidemiologic studies, the mostfrequently collected sample istumor tissue followed by blood.
Table 2. Examples of the types of biospecimens examined in published cancer epidemiology studies ofepigenetics, 2005–2012
BiospecimensExamples of types of cancer studied (all involved methylation marksunless noted in parentheses)
Blood Bladder cancer (43), breast cancer (44, 45), cervical cancer (46), colon cancer(47), esophageal cancer (miRNA profile; ref. 48), gastric cancer (49), headand neck cancer (50), leukemia (51), liver cancer (52), liver cancer (miRNAprofile; ref. 53), lung cancer (54), prostate cancer (55), and renal cancer (56)
Duodenal secretion Pancreatic cancer (57)Exfoliated cells from oral rinse Head and neck cancer (58)Formalin-fixed, paraffin-embedded tissues Lung cancer (histone modifications; ref. 59), prostate cancer (59, 60), rectal
cancer (61), and renal cancer (histone modifications; ref. 59)Pancreatic secretion Pancreatic cancer (62)Salivary rinse Lung cancer (63)Sputum Lung cancer (64, 65)Tumor tissue Bladder cancer (66), brain cancer (miRNAprofiling; ref. 67), breast cancer (68),
colon cancer (69, 70), esophageal cancer (71), gastric cancer (72),glioblastoma (73), head and neck cancer (74), laryngeal and hypolaryngealcancer (75), liver cancer (76), lung cancer (59, 77, 78), neuroblastoma (79),oral cancer (80), ovarian cancer (81, 82), pancreatic cancer (57, 83),prostate cancer (59, 84), rectal cancer (85), and renal cancer (59, 86)
Urine Bladder cancer (87), prostate cancer (88, 89)Uterine/cervix swab Cervical cancer (46)
Verma et al.
Cancer Epidemiol Biomarkers Prev; 23(2) February 2014 Cancer Epidemiology, Biomarkers & Prevention226
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573

Case–control studydesignwas themost predominantlyused study design type both in RPGs and in publications(Fig. 4). Investigators also used case-only cohort andnested case–control studies for epigenetic studies, but toa lesser extent. Eight grant proposals used either amixed-methods approach or different approaches to addresscancer etiology questions.Several methodologies are currently used to generate a
global viewofDNAmethylation,miRNAexpression, andhistone modifications in various cell types. As notedearlier, most of the projects examined both global and
tumor-specific DNA methylation changes (11, 20, 21).Investigators used several different types of technologiesfor methylation analyses (Fig. 5), and the most commonlyused were methylation-specific PCR, Methyl Light, andpyrosequencing-based technologies (22). Ulrich and col-leagues described a method that screens a broad scale offactors in relation to DNA methylation (23). In grantssupported by the NCI, Methyl Light and pyrosequencingwere most commonly used technologies for methylationprofiling; whereas methylation-specific PCR, MethylLight, was the routinely used technology in the published
0
10
20
30
40
50
60
Nested
Case–control
CohortCases onlyCase–control Others
% o
f T
ota
l
Study design
Grants Publications
Figure 4. Number of NCI grants andPubMed-indexed publications incancer epigenetic epidemiologyby study design. The mostcommon study design was case–control study in publications andgrants.
0
20
40
60
80
100
120
140
160
% o
f To
tal
Method/technology
Grants Publications
Affy
mat
rix ti
ling
arra
y
Bisul
fite
sequ
encing
Chi
p-on
-Chi
pCO
BRA
DNM
T as
say
EZ
DNA m
ethy
latio
n
Gol
den
gate
bea
d ar
ray
HPLC
Illum
ina
bead
array
Liqu
id c
hrom
atog
raph
y
LUM
A a
ssay
Mas
s ar
ray
with
epi
type
r
Met
hyl lig
ht
Met
hyla
tion-
spec
ific
PCR
Micro
arra
y
miR
NA p
rofilin
gM
SI
Pyr
oseq
uenc
ing
qPCR
qRT-
PCR
TaqM
anO
ther
Figure 5. Number of NCI grants andPubMed-indexed publications incancer epigenetic epidemiology bymethod or technology used in thestudy. AML, acute myelogenousleukemia; ALL, acutelymphoblastic leukemia; COBRA,combined bisulfate restrictionanalysis; DNMTassay, DNAmethyltransferase assay; HPLC, highpressure liquid chromatography;LUMA assay, LUminometricmethylation assay; MSI,microsatellite instability; qRT-PCR,quantitative reverse transcriptasePCR.
Epigenetics in Cancer Epidemiology
www.aacrjournals.org Cancer Epidemiol Biomarkers Prev; 23(2) February 2014 227
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573

studies (Fig. 5). Both grant awards and publications usedmostly PCR-based miRNA analysis technologies. How-ever, a recent trend is to use microarray or sequencing-based technologies because of their high-throughput andbroad dynamic range. Pyrosequencing of methylationregions was the preferred method in publications inwhich global methylation was studied, see for exampleref. 21. Chromatin immunoprecipitation methods usingantibodies for specific histone modifications, such asChIP-PCR, ChIP-on-ChIP (Chip onmicroarray Chip), andChIP-SAGE (ChIP combined with serial analysis of geneexpression), were routinely used for understanding his-tonemodifications and associatedDNAsequences in bothgrant awards and published articles, whereasmore recentpublications used ChIP-Seq.
Overall, the major hypotheses being addressed in boththe grant portfolio and the publications include (i) wheth-er diet or environmental exposure is associated withspecific epigenetic marks, patterns, and/or miRNAexpression, and whether epigenetic factors were relatedto cancer risk or survival from cancer; (ii) whether anepigenetic profile in blood or other accessible biospeci-mens is related to the epigenetic profile observed in thetumor; and (iii) whether an epigenetic pattern detected intumor tissue differs from that of the normal tissue sur-rounding it.
In the publications studies, there are numerous exam-ples of epidemiologic studies that reported the associationof methylation and risk of different cancers. Severalexamples will illustrate this. In a case–control studynested in the prospective Shanghai Women’s HealthStudy, 192 gastric cancer cases and 384 matched controlswere used to analyze methylation of Alu and long inter-spersed nucleotide elements (LINE), and results indicatedthat hypomethylation of these repeat sequences was asso-ciated with increased risk of cancer (24). However, breastcancerwas not associatedwithAlu andLINEhypomethy-lation, as reported in another group of participants from acohort in New York (25). A case–control study designdemonstrated an association of HPV16 DNA hyper-methylation with high-grade intraepithelial cervical can-cer (26). Characterization of methylated promoters ofother oncogenic papilloma viruses, HPV18, HPV31, andHPV41, indicated that methylation of these viruses was ageneral phenomenon and diagnostic assays, based onthese results, could be developed (27). A case–controlstudy showed a correlation between promoter methyla-tion and testicular tumors (28). Constitutional BRCA1promoter methylation determined in peripheral bloodwas shown to strongly predispose toward the develop-ment of tumors with features that resemble BRCA1-mut-ated tumors (29). Thus, screening of peripheral blood forBRCA1 methylation may predict risk for breast cancer.In another study, analysis of tumor tissues indicated thatinsulin growth factor II (IGF-II) promoter methylationwas associated with ovarian cancer progression (30). Inone nested case–control study, hypermethylation of runt-related transcription factor 3 (RUNX3) was associated
with advanced gastric lesions, which were susceptibleto Helicobacter pylori infection, a risk factor for gastriccancer (31). Inactivation of PTEN due to hypermethyla-tion was associated with cervical cancer (32). Studies inColon Cancer Family Registry indicated that relatives ofcolorectal cancer patients with hypermethylation ofMLH1 gene might be at increased risk of colorectal andgastric cancer and possibly ovarian and liver cancer (33).
Less commonly observed in the publications analysisare the effects of epigenetic factors on cancer survival,with very few examples illustrating this. Formalin-fixedtumor tissues from patients with non–small cell lungcancer were analyzed for hypermethylation of selectedgenes (p16, MGM2, DAPK, RASSF1, CDH1, LET7-3-a, andPTEN), and results indicated that p16 hypermethylationwas associated with a worse outcome in patients with age60 years or younger but not in older than 60 years (12).Serum DNA shed from tumors was used to evaluatemethylation of specific genes associated with breast can-cer in a case–control study (34). Delgado-Cruzata andcolleagues demonstrated that global DNA methylationlevels measured in white blood cells may be a potentialbiomarker of breast cancer risk even within families athigher risk of cancer (21).
DiscussionThrough our analyses of theNCI grant portfolio and the
published literature on the use of epigenetic markers inepidemiologic research, we identified a number of chal-lenges and opportunities for the field (Table 3). There istremendous potential to integrate the knowledge of epi-genetics research with genetic characteristics, environ-mental predisposition, and lifestyle factors in cancer epi-demiologic studies. The field of cancer epidemiologyresearch should leverage the use of epigenetic informa-tion when used appropriately to advance translationalresearch and to enhance practice in cancer prevention,detection, diagnosis, and treatment.
Breast, lung, colon, and prostate cancers were the moststudied organ sites in grants and publications, whereasa few studies investigated other cancer sites. This resultwas anticipated because these are among the most com-mon cancers and epidemiologists would likely haveaccess to higher numbers of samples from these cancertypes. The cancer site in which NCI-supported research isyielding many promising epidemiologic insights is coloncancer, particularly the identification of CIMP, a molec-ular subtype of colon cancer. Traditional histopathologicexamination cannot distinguish CIMPþwith CIMP� indi-viduals (35, 36). A prospective study of duration ofsmoking cessation and colorectal cancer risk was basedon CIMP analysis and results indicated a protective effectof smoking cessation on a DNA methylation–related car-cinogenesis pathway leading toCIMPþphenotype (37). Inthe Netherlands Cohort Study, body size and physicalactivity were associated with risk of colorectal cancer inCIMPþ and CIMP� individuals (38). Central adiposityincreased and high physical activity decreased the risk of
Verma et al.
Cancer Epidemiol Biomarkers Prev; 23(2) February 2014 Cancer Epidemiology, Biomarkers & Prevention228
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573
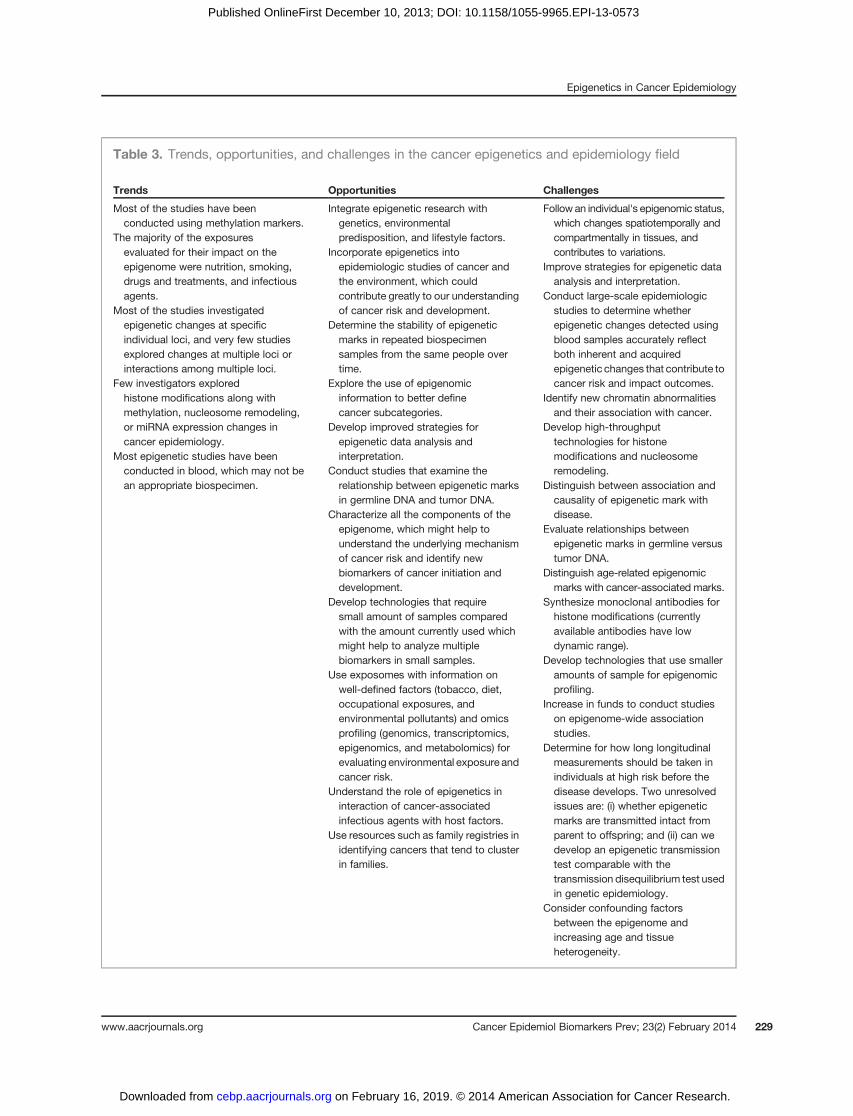
Table 3. Trends, opportunities, and challenges in the cancer epigenetics and epidemiology field
Trends Opportunities Challenges
Most of the studies have beenconducted using methylation markers.
Integrate epigenetic research withgenetics, environmentalpredisposition, and lifestyle factors.
Follow an individual's epigenomic status,which changes spatiotemporally andcompartmentally in tissues, andcontributes to variations.
The majority of the exposuresevaluated for their impact on theepigenome were nutrition, smoking,drugs and treatments, and infectiousagents.
Incorporate epigenetics intoepidemiologic studies of cancer andthe environment, which couldcontribute greatly to our understandingof cancer risk and development.
Improve strategies for epigenetic dataanalysis and interpretation.
Most of the studies investigatedepigenetic changes at specificindividual loci, and very few studiesexplored changes at multiple loci orinteractions among multiple loci.
Determine the stability of epigeneticmarks in repeated biospecimensamples from the same people overtime.
Conduct large-scale epidemiologicstudies to determine whetherepigenetic changes detected usingblood samples accurately reflectboth inherent and acquiredepigenetic changes that contribute tocancer risk and impact outcomes.Few investigators explored
histone modifications along withmethylation, nucleosome remodeling,or miRNA expression changes incancer epidemiology.
Explore the use of epigenomicinformation to better definecancer subcategories.
Identify new chromatin abnormalitiesand their association with cancer.
Most epigenetic studies have beenconducted in blood, which may not bean appropriate biospecimen.
Develop improved strategies forepigenetic data analysis andinterpretation.
Develop high-throughputtechnologies for histonemodifications and nucleosomeremodeling.Conduct studies that examine the
relationship between epigenetic marksin germline DNA and tumor DNA.
Distinguish between association andcausality of epigenetic mark withdisease.Characterize all the components of the
epigenome, which might help tounderstand the underlying mechanismof cancer risk and identify newbiomarkers of cancer initiation anddevelopment.
Evaluate relationships betweenepigenetic marks in germline versustumor DNA.
Develop technologies that requiresmall amount of samples comparedwith the amount currently used whichmight help to analyze multiplebiomarkers in small samples.
Distinguish age-related epigenomicmarks with cancer-associated marks.
Use exposomes with information onwell-defined factors (tobacco, diet,occupational exposures, andenvironmental pollutants) and omicsprofiling (genomics, transcriptomics,epigenomics, and metabolomics) forevaluating environmental exposure andcancer risk.
Synthesize monoclonal antibodies forhistone modifications (currentlyavailable antibodies have lowdynamic range).
Understand the role of epigenetics ininteraction of cancer-associatedinfectious agents with host factors.
Develop technologies that use smalleramounts of sample for epigenomicprofiling.
Use resources such as family registries inidentifying cancers that tend to clusterin families.
Increase in funds to conduct studieson epigenome-wide associationstudies.
Determine for how long longitudinalmeasurements should be taken inindividuals at high risk before thedisease develops. Two unresolvedissues are: (i) whether epigeneticmarks are transmitted intact fromparent to offspring; and (ii) can wedevelop an epigenetic transmissiontest comparable with thetransmission disequilibrium test usedin genetic epidemiology.
Consider confounding factorsbetween the epigenome andincreasing age and tissueheterogeneity.
Epigenetics in Cancer Epidemiology
www.aacrjournals.org Cancer Epidemiol Biomarkers Prev; 23(2) February 2014 229
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573

colorectal cancer in this population. In a large prospec-tive cohort of women (the Nurses’ Health Study), dif-ferential effects of B vitamins, alcohol, and methionineintake was observed in CIMPþ and CIMP� individuals(39). In another independent prospective study of olderwomen, no link with alcohol intake was observed irre-spective of the CIMP and BRAF mutation status (40).Examples discussed above also indicate that differentrisk factors have different effects on colorectal cancer inCIMPþ and CIMP� individuals. A recently completedprospective study involving 134,204 individuals indi-cated a protective effect of smoking cessation on a DNAmethylation–related pathway leading to high CIMPcolorectal cancer (37). Similar series of studies shouldbe considered for other cancer types in which CIMPphenotype has been reported such as bladder, ovarian,and prostate cancer.
These epidemiologic findings complement other linesof evidence that epigenetic events can be driver events inthe pathogenesis of colorectal cancer. Also, epigeneticevents cooperate with genetic events in the progressionof colonic mucosa to colorectal cancer. The number ofgenes affected by epigenetics is more than the number ofgenes mutated in colorectal cancer (41).
Our analysis indicated that most of the grants andpublications that we examined evaluated either global orspecific DNA methylation status and/or microRNA pro-files. This is likely because the methodologies usedto assess methylation and detect miRNAs are moreamenable for the high-throughput, large-scale studiesperformed in epidemiologic settings. In contrast, fewinvestigators explored histone modifications along withmethylation, nucleosome remodeling, or miRNA expres-sion changes in cancer epidemiology. Histone modifica-tions have been reported to play critical roles in cancerdevelopment, and aberrant patterns of histone modifica-tions are linked toDNAmethylation changes. In addition,the studies that we examined did not use genome-wideagnostic approaches or integrated approaches to assessepigenetic changes that influence cancers, although weanticipate an increase in these types of studies. Currently,ChIP-based methods are being used for assessing chro-matin structures on a genome-scale, but the limited avail-ability of specific antibodies for histonemodifications thatefficiently precipitate chromatin, low dynamic range, andthe labor-intensive, time-consuming, and costly nature ofthese studies preclude their use in epidemiology studies.In addition, technologies that use smaller amounts ofbiospecimens are needed for it to be feasible for epide-miologists to incorporate such experiments into theirstudies.
Our analysis showed that tumor tissues and bloodwerethe most predominant specimen type used for epigeneticresearch in epidemiology. An individual’s epigenomicstatus changes spatiotemporally and compartmentally intissues (42). Blood samples contain different types of cellswithhalf-livesvarying froma fewhours to severalyears. Inaddition, diverse cell types may have different inherent
epigenetic characteristics and may harbor or sustain dif-ferent levels of epigenetic changes (19). Large-scale epide-miologic studies are needed to determine whether epige-netic changes detected using blood samples accuratelyreflect both inherent and acquired epigenetic changes thatcontribute to cancer risk and impact outcomes (19, 42). Inaddition, such studies could examinewhether these inher-ent or acquired epigenetic changes reflect the pattern ofepigenetic changes in the tumor tissues.
Future research in this area includes developing imp-roved strategies for epigenetic data analysis and interpre-tation, determining the stability of epigenetic marks inrepeated biospecimen samples from the same peopleover time, and studies that examine the relationshipbetween epigenetic marks in germline DNA and tumorDNA. Resources that are particularly valuable includestudies that have prospectively collected and stored bios-pecimens, and thus allow for the analysis of epigeneticprofiles well before cancer starts developing. The serialsampling of biospecimens at multiple time points acrossthe life course may provide additional value allowinginsights into the temporal variation in epigenetic marksover time. Some important research resources that couldbe explored for such studies include some of the largecancer prevention trials (such as NCI’s Prostate, Lung,Colorectal, and Ovarian Cancer Screening Trial thatobtained repeated biosamples during the trials). Someconfounding factors need to be considered in studies ofepigenomics and cancer risk, particularly age. Moreover,becausemultiple types of exposures (e.g., alcohol, tobaccosmoking) may modify one’s epigenomic profile, theseexposures need to be controlled for when examining anyone exposure. Studies that correlate epigenetic character-istics in different tissues and body fluids in the samepersons would be very useful to validate the use of easilyaccessible biospecimens such as white blood cells instudies of cancer risk.
In infectious-agent–associated cancers (cervical, liver,gastric cancer, nasopharyngeal carcinoma, and Kaposisarcoma), epigenetic epidemiology has emerged as anoth-er promising area for future research. However, thesestudies face temporal causality problem. In addition, thereversible and context-dependent nature of epigeneticchanges poses challenges to epidemiologic studies. Toovercome these challenges, pathogen-associated epige-netic studies should be accompanied by comprehensivelongitudinal (multistage andmulti-individual) and trans-generational data.
Investment of resources is needed in this area of cancerepigenetics and epidemiology. Epigenetics hold substan-tial potential for furthering our understanding of themolecular mechanisms of health-related risks due toenvironmental exposure and individual susceptibility.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Received June 3, 2013; revised November 18, 2013; accepted November23, 2013; published OnlineFirst December 10, 2013.
Verma et al.
Cancer Epidemiol Biomarkers Prev; 23(2) February 2014 Cancer Epidemiology, Biomarkers & Prevention230
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573

References1. Baylin SB, Jones PA. A decade of exploring the cancer epigenome -
biological and translational implications. Nat Rev Cancer 2011;11:726–34.
2. Jones P. Out of Africa and into epigenetics: discovering reprogram-ming drugs. Nat Cell Biol 2011;13:2.
3. Feinberg AP. Genome-scale approaches to the epigenetics of com-mon human disease. Virchows Archiv 2010;456:13–21.
4. Khare S, Verma M. Epigenetics of colon cancer. Methods Mol Biol2012;863:177–85.
5. Kumar D, VermaM.Methods in cancer epigenetics and epidemiology.Methods Mol Biol (Clifton, NJ) 2009;471:273–88.
6. Verma M, Maruvada P, Srivastava S. Epigenetics and cancer. Crit RevClin Lab Sci 2004;41:585–607.
7. Verma M, Dunn BK, Ross S, Jain P, Wang W, Hayes R, et al. Earlydetection and risk assessment: proceedings and recommendationsfrom theWorkshoponEpigenetics inCancerPrevention.AnnNYAcadSci 2003;983:298–319.
8. Dunn BK, Verma M, Umar A. Epigenetics in cancer prevention: earlydetection and risk assessment: introduction. Ann N Y Acad Sci 2003;983:1–4.
9. Verma M, Srivastava S. Epigenetics in cancer: implications for earlydetection and prevention. Lancet Oncol 2002;3:755–63.
10. Feinberg AP. Epigenetics at the epicenter of modern medicine. JAMA2008;299:1345–50.
11. IwagamiS,BabaY,WatanabeM,ShigakiH,MiyakeK, IshimotoT, et al.LINE-1 hypomethylation is associated with a poor prognosis amongpatients with curatively resected esophageal squamous cell carcino-ma. Ann Surg 2013;257:449–55.
12. Bradly DP, Gattuso P, Pool M, Basu S, Liptay M, Bonomi P, et al.CDKN2A (p16) promoter hypermethylation influences the out-come in young lung cancer patients. Diagn Mol Pathol 2012;21:207–13.
13. Brasset E, Chambeyron S. Epigenetics and transgenerational inher-itance. Genome Biol 2013;14:306.
14. Wild CP, Scalbert A, Herceg Z. Measuring the exposome: a powerfulbasis for evaluating environmental exposures and cancer risk. EnvironMol Mutagen 2013;54:480–99.
15. Hou L, Zhang X, Wang D, Baccarelli A. Environmental chemicalexposures and human epigenetics. Int J Epidemiol 2012;41:79–105.
16. Costa FF. Epigenomics in cancer management. Cancer Manag Res2010;2:255–65.
17. Verma M. Cancer control and prevention: nutrition and epigenetics.Curr Opin Clin Nutr Metab Care 2013;16:376–84.
18. Mill J, HeijmansBT. Frompromises to practical strategies in epigeneticepidemiology. Nat Rev Genet 2013;14:585–94.
19. Terry MB, Delgado-Cruzata L, Vin-Raviv N, Wu HC, Santella RM. DNAmethylation in white blood cells: association with risk factors inepidemiologic studies. Epigenetics 2011;6:828–37.
20. Huang WY, Su LJ, Hayes RB, Moore LE, Katki HA, Berndt SI, et al.Prospective study of genomic hypomethylation of leukocyte DNA andcolorectal cancer risk. Cancer Epidemiol Biomarkers Prev 2012;21:2014–21.
21. Delgado-Cruzata L, Wu HC, Perrin M, Liao Y, Kappil MA, Ferris JS,et al. Global DNA methylation levels in white blood cell DNA fromsisters discordant for breast cancer from the New York site of theBreast Cancer Family Registry. Epigenetics 2012;7:868–74.
22. Eads CA, Danenberg KD, Kawakami K, Saltz LB, Blake C, Shibata D,et al. Methy Light: a high-throughput assay to measure DNA methyl-ation. Nucleic Acids Res 2000;28:E32.
23. UlrichCM,ToriolaAT, Koepl LM,Sandifer T, PooleEM,DugganC, et al.Metabolic, hormonal and immunological associationswith global DNAmethylation among postmenopausal women. Epigenetics 2012;7:1020–8.
24. Gao Y, Baccarelli A, Shu XO, Ji BT, Yu K, Tarantini L, et al. Bloodleukocyte Alu and LINE-1 methylation and gastric cancer risk inthe Shanghai Women's Health Study. Br J Cancer 2012;106:585–91.
25. Wu HC, Delgado-Cruzata L, Flom JD, Perrin M, Liao Y, Ferris JS,et al. Repetitive element DNA methylation levels in white blood cell
DNA from sisters discordant for breast cancer from the New Yorksite of the Breast Cancer Family Registry. Carcinogenesis 2012;33:1946–52.
26. Mirabello L, Schiffman M, Ghosh A, Rodriguez AC, Vasiljevic N,WentzensenN, et al. ElevatedmethylationofHPV16DNA is associatedwith the development of high grade cervical intraepithelial neoplasia.Int J Cancer 2013;132:1412–22.
27. Wentzensen N, Sun C, Ghosh A, Kinney W, Mirabello L, Wacholder S,et al. Methylation of HPV18, HPV31, andHPV45 genomes and cervicalintraepithelial neoplasia grade 3. JNatl Cancer Inst 2012;104:1738–49.
28. Mirabello L, Kratz CP, Savage SA, Greene MH. Promoter methylationof candidate genes associated with familial testicular cancer. Int J MolEpidemiol Genet 2012;3:213–27.
29. Wong EM, Southey MC, Fox SB, Brown MA, Dowty JG, Jenkins MA,et al. Constitutional methylation of the BRCA1 promoter is specificallyassociated with BRCA1mutation-associated pathology in early-onsetbreast cancer. Cancer Prev Res (Phila) 2011;4:23–33.
30. Qian B, Katsaros D, Lu L, Canuto EM, Benedetto C, Beeghly-Fadiel A,et al. IGF-II promoter specific methylation and expression in epithelialovarian cancer and their associations with disease characteristics.Oncol Rep 2011;25:203–13.
31. Li WQ, Pan KF, Zhang Y, Dong CX, Zhang L, Ma JL, et al. RUNX3methylation and expression associated with advanced precancerousgastric lesions in a Chinese population. Carcinogenesis 2011;32:406–10.
32. Rizvi MM, Alam MS, Ali A, Mehdi SJ, Batra S, Mandal AK. Aberrantpromoter methylation and inactivation of PTEN gene in cervical car-cinoma from Indian population. J Cancer Res Clin Oncol 2011;137:1255–62.
33. Levine AJ, Win AK, Buchanan DD, Jenkins MA, Baron JA, Young JP,et al. Cancer risks for the relatives of colorectal cancer cases with amethylated MLH1 promoter region: data from the Colorectal CancerFamily Registry. Cancer Prev Res (Phila) 2012;5:328–35.
34. Sturgeon SR, Balasubramanian R, Schairer C, Muss HB, Ziegler RG,Arcaro KF. Detection of promoter methylation of tumor suppressorgenes in serumDNAof breast cancer cases and benign breast diseasecontrols. Epigenetics 2012;7:1258–67.
35. Budinska E, Popovici V, Tejpar S, D'Ario G, LapiqueN, Sikora KO, et al.Gene expression patterns unveil a new level of molecular heteroge-neity in colorectal cancer. J Pathol 2013; 231:63–76.
36. Hughes LA, Melotte V, de Schrijver J, de Maat M, Smit VT, Bovee JV,et al. The CpG islandmethylator phenotype: what's in a name?CancerRes 2013;73:5858–68.
37. NishiharaR,Morikawa T, KuchibaA, Lochhead P, YamauchiM, Liao X,et al. A prospective study of duration of smoking cessation andcolorectal cancer risk by epigenetics-related tumor classification. AmJ Epidemiol 2013;178:84–100.
38. Hughes LA, Simons CC, van den Brandt PA, Goldbohm RA, de GoeijAF, de Bruine AP, et al. Body size, physical activity and risk ofcolorectal cancerwith orwithout theCpG islandmethylator phenotype(CIMP). PloS ONE 2011;6:e18571.
39. Schernhammer ES, Giovannucci E, Baba Y, Fuchs CS, Ogino S. Bvitamins, methionine and alcohol intake and risk of colon cancer inrelation to BRAF mutation and CpG island methylator phenotype(CIMP). PloS ONE 2011;6:e21102.
40. Razzak AA, Oxentenko AS, Vierkant RA, Tillmans LS, Wang AH,Weisenberger DJ, et al. Alcohol intake and colorectal cancer risk bymolecularly defined subtypes in a prospective study of older women.Cancer Prev Res (Phila) 2011;4:2035–43.
41. Lao VV, Grady WM. Epigenetics and colorectal cancer. Nat RevGastroenterol Hepatol 2011;8:686–700.
42. Relton CL, DaveySmith G. Is epidemiology ready for epigenetics? Int JEpidemiol 2012;41:5–9.
43. Cash HL, Tao L, Yuan JM, Marsit CJ, Houseman EA, Xiang YB, et al.LINE-1 hypomethylation is associated with bladder cancer risk amongnonsmoking Chinese. Int J Cancer 2012;130:1151–9.
44. Alshatwi AA. Breast cancer risk, dietary intake, and methylenetetrahy-drofolate reductase (MTHFR)single nucleotide polymorphisms. FoodChem Toxicol 2010;48:1881–5.
Epigenetics in Cancer Epidemiology
www.aacrjournals.org Cancer Epidemiol Biomarkers Prev; 23(2) February 2014 231
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573

45. Cebrian A, PharoahPD, AhmedS, Ropero S, FragaMF, Smith PL, et al.Genetic variants in epigenetic genes and breast cancer risk. Carcino-genesis 2006;27:1661–9.
46. Chung MT, Sytwu HK, Yan MD, Shih YL, Chang CC, Yu MH, et al.Promoter methylation of SFRPs gene family in cervical cancer. Gyne-col Oncol 2009;112:301–6.
47. Kaaks R, Stattin P, Villar S, Poetsch AR, Dossus L, Nieters A, et al.Insulin-like growth factor-II methylation status in lymphocyte DNA andcolon cancer risk in the Northern Sweden Health and Disease cohort.Cancer Res 2009;69:5400–5.
48. Zhang C, Wang C, Chen X, Yang C, Li K, Wang J, et al. Expressionprofile of microRNAs in serum: a fingerprint for esophageal squamouscell carcinoma. Clin Chem 2010;56:1871–9.
49. Liu R, Zhang C, Hu Z, Li G, Wang C, Yang C, et al. A five-microRNAsignature identified from genome-wide serum microRNA expressionprofiling serves as a fingerprint for gastric cancer diagnosis. Eur JCancer (Oxford, England : 1990) 2011;47:784–91.
50. Hsiung DT, Marsit CJ, Houseman EA, Eddy K, Furniss CS, McCleanMD, et al. Global DNA methylation level in whole blood as a biomarkerin head and neck squamous cell carcinoma. Cancer Epidemiol Bio-markers Prev 2007;16:108–14.
51. Claus R, Hackanson B, Poetsch AR, Zucknick M, Sonnet M, Blagitko-Dorfs N, et al. Quantitative analyses of DAPK1methylation in AML andMDS. Int J Cancer 2012;131:E138–42.
52. Akkiz H, Bayram S, Bekar A, Akgollu E, Ulger Y. A functional poly-morphism in pre-microRNA-196a-2 contributes to the susceptibility ofhepatocellular carcinoma in a Turkish population: a case-controlstudy. J Viral Hepat 2011;18:e399–407.
53. Zhou J, Yu L,GaoX, Hu J,Wang J, Dai Z, et al. PlasmamicroRNApanelto diagnose hepatitis B virus-related hepatocellular carcinoma. J ClinOncol 2011;29:4781–8.
54. Cao Y, Miao XP, Huang MY, Deng L, Liang XM, Lin DX, et al. Poly-morphisms of methylenetetrahydrofolate reductase are associatedwith a high risk of nasopharyngeal carcinoma in a smoking populationfrom Southern China. Mol Carcinog 2010;49:928–34.
55. Bastian PJ, Palapattu GS, Yegnasubramanian S, Rogers CG, Lin X,Mangold LA, et al. CpG island hypermethylation profile in the serum ofmen with clinically localized and hormone refractory metastatic pros-tate cancer. J Urol 2008;179:529–34.
56. Liao LM, Brennan P, van Bemmel DM, Zaridze D, Matveev V, Janout V,et al. LINE-1 methylation levels in leukocyte DNA and risk of renal cellcancer. PloS ONE 2011;6:e27361.
57. Matsubayashi H, Sato N, Brune K, Blackford AL, Hruban RH, CantoM,et al. Age- and disease-related methylation of multiple genes in non-neoplastic duodenum and in duodenal juice. Clin Cancer Res2005;11:573–83.
58. Franzmann EJ, Reategui EP, Pedroso F, Pernas FG, Karakullukcu BM,Carraway KL, et al. Soluble CD44 is a potential marker for the earlydetection of head andneck cancer. Cancer Epidemiol Biomarkers Prev2007;16:1348–55.
59. Seligson DB, Horvath S, McBrianMA, Mah V, Yu H, Tze S, et al. Globallevels of histone modifications predict prognosis in different cancers.Am J Pathol 2009;174:1619–28.
60. Weiss G, Cottrell S, Distler J, Schatz P, Kristiansen G, Ittmann M, et al.DNA methylation of the PITX2 gene promoter region is a strongindependent prognostic marker of biochemical recurrence in patientswith prostate cancer after radical prostatectomy. J Urol 2009;181:1678–85.
61. CurtinK,SamowitzWS,UlrichCM,Wolff RK,Herrick JS,CaanBJ, et al.Nutrients in folate-mediated, one-carbon metabolism and the risk ofrectal tumors in men and women. Nutr Cancer 2011;63:357–66.
62. Matsubayashi H, Canto M, Sato N, Klein A, Abe T, Yamashita K, et al.DNA methylation alterations in the pancreatic juice of patients withsuspected pancreatic disease. Cancer Res 2006;66:1208–17.
63. SunW, Zaboli D,WangH, Liu Y, Arnaoutakis D, Khan T, et al. Detectionof TIMP3 promoter hypermethylation in salivary rinse as an indepen-dent predictor of local recurrence-free survival in head and neckcancer. Clin Cancer Res 2012;18:1082–91.
64. TessemaM, Belinsky SA. Mining the epigenome for methylated genesin lung cancer. Proc Am Thorac Soc 2008;5:806–10.
65. BaryshnikovaE,DestroA, InfanteMV,CavutoS,Cariboni U, AlloisioM,et al. Molecular alterations in spontaneous sputum of cancer-freeheavy smokers: results from a large screening program. Clin CancerRes 2008;14:1913–9.
66. Dudziec E,Miah S, Choudhry HM, OwenHC, Blizard S, GloverM, et al.Hypermethylation of CpG islands and shores around specific micro-RNAs and mirtrons is associated with the phenotype and presence ofbladder cancer. Clin Cancer Res 2011;17:1287–96.
67. Mueller WC, Spector Y, Edmonston TB, St Cyr B, Jaeger D, Lass U,et al. Accurate classification of metastatic brain tumors using a novelmicroRNA-based test. Oncologist 2011;16:165–74.
68. Dietrich D, Krispin M, Dietrich J, Fassbender A, Lewin J, Harbeck N,et al. CDO1 promoter methylation is a biomarker for outcome predic-tion of anthracycline treated, estrogen receptor-positive, lymph node-positive breast cancer patients. BMC Cancer 2010;10:247.
69. Rawson JB, Mrkonjic M, Daftary D, Dicks E, Buchanan DD, Young-husband HB, et al. Promoter methylation of Wnt5a is associated withmicrosatellite instability and BRAF V600E mutation in two large popu-lations of colorectal cancer patients. Br J Cancer 2011;104:1906–12.
70. Worthley DL, Whitehall VL, Buttenshaw RL, Irahara N, Greco SA,Ramsnes I, et al. DNAmethylationwithin the normal colorectal mucosais associated with pathway-specific predisposition to cancer. Onco-gene 2010;29:1653–62.
71. Jin Z, Cheng Y, Gu W, Zheng Y, Sato F, Mori Y, et al. A multicenter,double-blinded validation study of methylation biomarkers for pro-gression prediction in Barrett's esophagus. Cancer Res 2009;69:4112–5.
72. Chang MS, Uozaki H, Chong JM, Ushiku T, Sakuma K, Ishikawa S,et al. CpG island methylation status in gastric carcinoma with andwithout infection of Epstein-Barr virus. Clin Cancer Res 2006;12:2995–3002.
73. Delfino KR, Serao NV, Southey BR, Rodriguez-Zas SL. Therapy-,gender- and race-specific microRNA markers, target genes and net-works related to glioblastoma recurrence and survival. Cancer Geno-mics Proteomics 2011;8:173–83.
74. Avissar M, McClean MD, Kelsey KT, Marsit CJ. MicroRNA expressionin head and neck cancer associates with alcohol consumption andsurvival. Carcinogenesis 2009;30:2059–63.
75. Dikshit RP, Gillio-Tos A, Brennan P, De Marco L, Fiano V, Martinez-Penuela JM, et al. Hypermethylation, risk factors, clinical character-istics, and survival in 235 patients with laryngeal and hypopharyngealcancers. Cancer 2007;110:1745–51.
76. Ji J, Shi J, Budhu A, Yu Z, Forgues M, Roessler S, et al. MicroRNAexpression, survival, and response to interferon in liver cancer. NEngl JMed 2009;361:1437–47.
77. Limsui D, Vierkant RA, Tillmans LS, Wang AH, Weisenberger DJ, LairdPW, et al. Cigarette smoking and colorectal cancer risk by molecularlydefined subtypes. J Natl Cancer Inst 2010;102:1012–22.
78. Brock MV, Hooker CM, Ota-Machida E, Han Y, Guo M, Ames S, et al.DNAmethylationmarkers andearly recurrence in stage I lungcancer.NEngl J Med 2008;358:1118–28.
79. Bray I, Bryan K, Prenter S, Buckley PG, Foley NH, Murphy DM, et al.Widespread dysregulation of MiRNAs by MYCN amplification andchromosomal imbalances in neuroblastoma: association of miRNAexpression with survival. PloS ONE 2009;4:e7850.
80. Cao J, Zhou J, Gao Y, Gu L, Meng H, Liu H, et al. Methylation of p16CpG island associated with malignant progression of oral epithelialdysplasia: a prospective cohort study. Clin Cancer Res 2009;15:5178–83.
81. An J, Wei Q, Liu Z, Lu KH, Cheng X, Mills GB, et al. Messenger RNAexpression andmethylation of candidate tumor-suppressor genes andrisk of ovarian cancer-a case-control analysis. Int J Mol EpidemiolGenet 2010;1:1–10.
82. DaiW, Teodoridis JM, Zeller C,GrahamJ,Hersey J, Flanagan JM, et al.Systematic CpG islands methylation profiling of genes in the wntpathway in epithelial ovarian cancer identifies biomarkers of progres-sion-free survival. Clin Cancer Res 2011;17:4052–62.
83. Donahue TR, Tran LM, Hill R, Li Y, Kovochich A, Calvopina JH, et al.Integrative survival-based molecular profiling of human pancreaticcancer. Clin Cancer Res 2012;18:1352–63.
Verma et al.
Cancer Epidemiol Biomarkers Prev; 23(2) February 2014 Cancer Epidemiology, Biomarkers & Prevention232
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573

84. Kim SJ, Kelly WK, Fu A, Haines K, Hoffman A, Zheng T, et al. Genome-widemethylation analysis identifies involvementofTNF-alphamediatedcancer pathways in prostate cancer. Cancer Lett 2011;302:47–53.
85. Slattery ML, Curtin K, Wolff RK, Herrick JS, Caan BJ, Samowitz W.Diet, physical activity, and body size associations with rectal tumormutations and epigenetic changes. Cancer Causes Control 2010;21:1237–45.
86. Nickerson ML, Jaeger E, Shi Y, Durocher JA, Mahurkar S, Zaridze D,et al. Improved identification of von Hippel-Lindau gene alterations inclear cell renal tumors. Clin Cancer Res 2008;14:4726–34.
87. Costa VL, Henrique R, Danielsen SA, Duarte-Pereira S, Eknaes M,Skotheim RI, et al. Three epigenetic biomarkers, GDF15, TMEFF2, andVIM, accurately predict bladder cancer from DNA-based analyses ofurine samples. Clin Cancer Res 2010;16:5842–51.
88. Baden J, Green G, Painter J, Curtin K, Markiewicz J, Jones J, et al.Multicenter evaluation of an investigational prostate cancer methyla-tion assay. J Urol 2009;182:1186–93.
89. Vener T, Derecho C, Baden J, Wang H, Rajpurohit Y, Skelton J, et al.Development of a multiplexed urine assay for prostate cancer diag-nosis. Clin Chem 2008;54:874–82.
Epigenetics in Cancer Epidemiology
www.aacrjournals.org Cancer Epidemiol Biomarkers Prev; 23(2) February 2014 233
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573

2014;23:223-233. Published OnlineFirst December 10, 2013.Cancer Epidemiol Biomarkers Prev Mukesh Verma, Scott Rogers, Rao L. Divi, et al. Opportunities, and ChallengesEpigenetic Research in Cancer Epidemiology: Trends,
Updated version
10.1158/1055-9965.EPI-13-0573doi:
Access the most recent version of this article at:
Cited articles
http://cebp.aacrjournals.org/content/23/2/223.full#ref-list-1
This article cites 89 articles, 25 of which you can access for free at:
Citing articles
http://cebp.aacrjournals.org/content/23/2/223.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cebp.aacrjournals.org/content/23/2/223To request permission to re-use all or part of this article, use this link
on February 16, 2019. © 2014 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Published OnlineFirst December 10, 2013; DOI: 10.1158/1055-9965.EPI-13-0573