evolution and the aquatic ecosystem: defining unique units …€¦ · · 2009-10-06evolution and...
TRANSCRIPT

EVOLUTION AND THE AQUATIC ECOSYSTEM: DEFINING UNIQUE UNITS IN POPULATION CONSERVATION
JENNIFER L. NIELSEN Editor
DENNIS A. POWERS Consulring Editor
American Fisheries Society Symposium 17
Proceeditzgs of rlte Symposium on Evolution and rltc Aquatic Ecosystem: Defining Unique Utzits in Populatiorl Conscn~ariotl
Held at Monterey, California. USA 23-25 May 1994
American Fisheries Society Bethesda, Maryland
1995 - .r

The American Fisheries Society Symposium series is a registered serial. Suggested citation formats follow.
Entire book
Nielsen, J. L.. editor. 1995. Evolution and the aquatic ecosystem: defining unique units in population conservation. American Fisheries Society Symposium 17, Bethesda, Maryland.
Article within the book
Healey, M. C., and A. Prince. 1995. Scales of variation in life history tactics of Pacific salmon and the conservation of phenotype and genotype. Pages 176-184 in J. L. Nielsen, editor. Evolution and the aquatic ecosystem: defining unique units in population conservation. American Fisheries Society Symposium 17, Bethesda, Maryland.
O Copyright by the American Fisheries Society, 1995
All rights reserved. Photocopying for internal or personal use. or for the internal or personal use of specific clients, is permitted by AFS provided that the appropriate fee is paid directly to Copyright Clearance, Center (CCC). 773 Rosewood Drive. Danvers. Massachusetts 01923, USA; phone 508-750-84!HI. ,Request authorization to make multiple copies for classroom use from CCC. These permissions do not -extend to electronic distribution or long-term storage of articles or to copying for resale, promotion. advertising, general distribution. or creation of new collective works. For such uses, permission or license must be
.. . obtained from AFS.
Library of Congress Catalog Card Number: 95-083247
ISBN 0-913235-94-6 ISSN 0892-2284
Printed in the United States of America on recycled, acid-free paper.
American Fisheries Society . 5410 Grosvenor Lane, Suite 110
Bethesda, Maryland 20814-2199. USA
Preface . . . Introduction
Evolutiona Endangere R S. Waplt
Selection o P. R Muna'
Session Ow Morpholog R I. Behnk
Processes a G. Smitlz, J.
Systematics 'and Conser R L Mayd,
A Role for L Bematch
Evolutiona~ Coregonid R B. Phil14
Faunas of 1 M. Mina an
Genetic Pol F. M. Utter.
Sessiotr Ove The Releva G. W. Barlc

Amencan Fiheries Sociery SflpOsium 17:334-249. 1995 C Copyngn~ by the American Firherim Sociery 1995
*
A Disturbance-Based Ecosystem Approach to Maintaining and Restoring Freshwater Habitats of
Evolutionarily Significant Units of Anadromous' Salmonids in the Pacific Northwest
G. H. REEVES U.S. Fowsr Service, pacific Nonhwesf Rerea~h Sfation, 3200 SW Jeffenon Way, Cowallis, Oregon 97331, USA
L E. BENDA Depanmenf of Geological Sciences, Unive&y of WashLrgron. Sea& Washingron 98195. USA
U.S. Fans Service Pacific N o n h w t Research Station
Technology Cenvr, Wcyerhaeuser Company, T a w m Washingron 98477, US4
F o m Sewice. Pacific Nonhwcn RCKMh Station
Absnacf.-To preserve and recover evolutionarily significant units (ESUs) of anadromous salmo- nids 0ncoh.vnchus spp. in the Pacific Northwest. long-term and short-term ecological processes that create and matntain freshwater habitats must be restored and protected. Aquatic ecosystems throu* out the region are dynamic in space and time. and lack of consideration of their dynamic aspects has limited the effectiveness of habitat restoration program. Riverine-riparian ecosystems used by anadromous salmonids were naturally subjected to periodic catastrophic disturbances. after which they moved through a series of recovery states over periods of decades to centuries. Consequently the landscape was a mosaic of varying habitat conditions, some that were suitable for anadromous salmonids and some that were not. Life history adaptations of salmon, such as straying of adults. mo\.ement of juveniles. and high fecundity r a t e allowed populations of anadromous salmonids to persist in this dynamic environment. Perspectives gained from natural cycles of disturbance and recovery of the aquatic envifonment must be incorporated into recovery plans for freshwater habitats. In general. we do npt advocate renuning to the natural disturbance regime, which may include large-scale catastrophic processes such as stand-replacing wildtirtr This may be an impossibility given patterns of humin aevelopment in the region. We believe that it is more prudent to modify human- imposed disturbance regimes to create and maintain the nmssary range of habitat conditions in space (I@' h) and time (10'-102 years) within and among watersheds across the distributional range of an ESU. An additional component of any recovery plan. which is imperative in the short-term. is the establishment of watershed reserves that contain the best a t i n g habitats and indude the most ecologically intact watersheds.
Biodi\.eni? is not a 'set-aside' issue that' can be physi- and degradation. overexploitation in spon and com- al ly isolated in a few. or even many. resewes.. . . We mercial fisheries, variable ocean conditions. and ef- must see lhe larger t'k--stewardshi~ of all the species fects of hatchery practices, are responsible for the on all of the landscape with every activity we undertake as human beings-a task without spatial and temporal depressed status of these fish (Nehlsen et al. 1991). boundaries. (J. F. Franklin 1993) The relative importance of each in contributing to
the decline of an ESU undoubtedly varies across the Agencies responsible for the development of re- region. Any recovery program must address and
covery plans for evolutionarily significant units incorporate consideration of all responsible factors (ESUs: Waples 1991) of anadromous salmohids to be successful. Oncori~ytrchw spp. in the Pacific Northwest (PNW) The most common factor associated with declines of the United States face difficult tasks. First is the of anadromous salmonids is habitat degradation. identification of ESUs. Second is the identification which includes destruction and modification of of factors that contribute to the decline of a panic- freshwater and estuarine habitats (Nehlsen et a!. ular ESU. X suite of factors. including habitat loss 1991; Frissell 1993). Stream and river Systems
334 .

DISTURBANCE-BASED H.4BITAT RECOVERY P U W 335
throughout the PNW have been extensively altered to accomplish this. Williams et al. (1989) also noted by human activities such as agriculture, urbaniza- that the failure to address this concern may be a tion. and timber harvest (Bisson et al. 1992). Fea- major reason no fish species has ever been recov- tures of altered ecosystems include changes (gener- ered after listing under the U.S. Endangered Spe- ally reductions) in species diversity, changes in cies Act (ESA: 16 U.S.C. $5 1531 to 1544). species distributions. and losses of habitat types or The purpose of this paper is to examine compo- ecosystem states (Holling 1973; Rappon et al. 1985: nents of strategies necessary to provide habitat for Steedman and Regier 1987). Li et a[. (1987). Bisson ESUs of anadromous salmonids in the PNW. Spe- et (1992)v and Reeves et (1993) noted that cifically. we will consider the role of natural distur- native salmonid assemblages are simplified in wa- bances in creating and maintaining habitats tersheds that have been impacted by various human how an understanding of [his role be incor- activities. Native nonsalmonids or introduced spe- porated into long-tem recovey planning. cies often dominate fish communities in altered ecosystems (Li et al. 1987; Bisson et al. 1992). Hab- itat degradation is widespread across the region as a Ecosystem and Spatiotemporal result of past and present activities (Bisson et al. Considerations 1992; McIntosh et al. 1994). Degradation of terres- trial ecosystems in the PNW (Thomas et al. 1993) May (1994) noted that the most pressing chal- .
and elsewhere (e.g., Wilcove ei al. 1986; Rolstad lenge to conservation biology is the need to under- 1991) has resulted in similar changes in terrestrial stand the responses of organismsover large tempo- species assemblages. . . ral and spatial scales. Some relationships between
~ & t and many present approaches to manage- habitat condition and individual salmonid response rilent of freshwater habitats of anadromous salmo- have been well established at the scales of habitat nick have focused on mitigating losses rather than unit'(e.g.. Bisson et al. 1982; Nickelson et al. 1992), preventing them. This strategy has generally not stream reach (e.g., Murphy et al. 1989). and ( t o a been successful.(Biison et al. 1992) and habitat loss lesser extent) watershed (Schlosser 1991). But there and degradation continue. Williams et al. (1989) is little understanding about how biological entities also found that such a strategy failed to halt the such as ESUs may respond to habitat patterns at decline of habitat quantity and quality 'for other large spatial scales.-'dn initial hurdle in recovery freshwater fishes. Naturally variable ocean condi- planning for ESUs is identifying appropriate spatial tions increase the importance of freshwater habitats and temporal scales on which to focus.
to anadromous salmonids (Thomas et al. 1993). As The ESA requires that ecosystems be considered a iesblt of this dependence on'freshwater habitats in the development of recovery plans. The ESUs of and the extensive, amount of habitat degradation anadromous salmonids generally encompass large that has occurred, protection and restoration of geographic areas (e.g., Snake River basin in Idaho, upslope and fluvial processes that create and main- upper Sacramento River and its tributaries in north- tain habitats must be an integral component of any em California). It is difficult to delineate the fresh- recovery program. water ecosystem of an ESU over such large areas.
Habitat losses may result from human activities We believe that it is reasonable- to consider the that directly destroy habitats or change the long- composite of individual watersheds within the geo- term dynamics of ecosystems (Rapport et al. 1985; graphic range of an ESU to be the "ecosystem" and Webb and Thomas 1994). Recent proposals for to direct conservation and recovery efforts for fresh- restoring and protecting habitats of at-risk fishes water habitats toward the populations that make up (e-g., Reeves and Sedell 1992; Thomas et al. 1993; an ESU. Currens et al. (in press) suggest that ap- Moyle and Yoshiyama 1994) addressed habitat de- propriate temporal scales for populations are sev- struction. primarily through the establishment of eral de~ades to centuries and that spatial Scales watershed-level reserves in which human impacts should begin at the watershed level (Figure 1). M- would be minimized, as advocated by. Sheldon though temporal considerations have not been ad- (1988) and Williams et al. (1989). We are not aware dressed explicitly, recent proposals for restoring of anyone who has explicitly addressed long-term and conserving freshwater habitats of anadromous ecosystem dynamics in the context of fish conserva- salmonids have emphasized watersheds (e.g.. tion. Williams et al. (1989) called for recovery ef- Reeves and Sedell 1992; Thomas et al. 1993; Moyle forts to restore and conserve ecosystems rather than and Yoshiyama 1994). We concur with this d i m -

REEVES ET AL
FIGURE I.-A general hierarchial model of biological levels of organization for anadromous salmonids and the spatial and temporal scales that influence them (from Currcns et al.. in press).
mentation purposes, the individual watershed is the appropriate focus for recovery plans.
Within watersheds, recovery programs for ESUs must address not only root causes directly respon- sible for the immediate loss of habitat quanti iand quality &@o ecosystem processes that create and maintain habitats through time. In developing an ecosystem approach to the conservation and resto- ration of endangered organisms, it must be recog- nized that ecoGtems are generally dynamic in space and time because of natural .disturbances, particularly at large spaciotemporal scales (Botkin 1990). , .
A mosaic of conditions occurs within an ecosys- tem at any time as a consequence of disturbances (White and Pickett 1985). Any disturbed patch de- velops different habitat conditions or states over time. The assemblage of organisms in a particular patch changes with changing habitat conditions (Ta- ble 1; Huff and Raley 1991; Raphael 1991). Points along the trajectory of disturbance and recovery represent various states in the potential range of states that an ecosystem may exhibit. The locations of patches in particular states shift across the land- scape due to the stochastic nature of most natural disturbances. In the PNW, terrestrial ecosystems are very dynamic in space and time as a result of natural disturbances such as fire and wind (Agee 1991, 1993). Holling (1973) noted that if resources are to be sustained, the dynamic nature of ecosys- tems and the need to maintain the diversity of ecosystem states must be recognized. Attempts to view and manage systems and resources in a static contexr may increase the rate of extinction of some organisms (Holling 1973).
Persistence in Dynamic Environments
It is unlikely that individual populations persist over long tenns at the local scale in a d ~ a m i c environment (Hanski and Gilpin 1991; McCauley 1991; Mange1 and Tier 1994). In dynamic environ- ments, " . . . some patches are empty (but liable for colonization), while others are occupied (but liable to extinction). Ln such circumstances, the lights of individual patches wink on and off unpredictably, but the overall average level of illumination-the overall density of the metapopulation-may remain
TAB= 1.-Bird species found in d i r c n t sera1 stages of Douglas-fir fomts of Oregon and Washington (from Huff and Raley 1991).
Seral rraee S~cc ies Earh. Mid kte
Chestnut-backed chickadee Panu nrfacau
Hermit warbler Dendmica occidnYalic
Western flycatcher ,, . Lnp- d i m , . ,, ! '. . :;
Winter wren .T*aooglodyru'
Red-breasted nuthatch Sin4 caM&Nic
Swainson's thrush cuthanu LLrlUhlus
American mbm Twkrr migrmorLcr
Northern spotted owl SIlir occlduuolil CaUriM
Pileated woodpecker Drpzopw pileanu
Varied thrush LxOmunaNLrr

DISTGRBXNCE-BASED HABITAT RECOi'ERY PLLY 337
relatively steady" (May 1994). Metapopulations siry and effect of disturbance vay with channel sire persist in dynamic environments through a suite of and location within the watershed (Benda 199.1). adaptations. Response to change varies with the level of biological organization (Karr and Freemark 1985: White and Pickett 1985).Physiological. mor- An Oregot1 Evan~ple
phological. and behavioral aaaptations &at the The natural disturbance regime in the central individual level. Life history patterns (Stearns Oregon Coast Range includes infrequent stand- 1977). reproductive rates,' and modes of dispersal resetting wildfires and frequent intense winter rain- (Vrijenhoek 1985) are adaptations at the popula- storms. Wildfires reduce the soil-binding capacity of tion level. roots.' When intense rainstorms saturate soils dur-
Several studies have documented the response of . ing periods of low root strength. concentrated land- terrestrial populations to periodic catastrophic dis- sliding into channels and debris flows may result. turbances. Christensen ( 1985) cited examples of de- Such naturally occurring disturbances in stream clines in small-mammal populations after fires in channels can have both immediate impacts on and shrublands. Populations recovered after the vegeti- long-term implications for anadromous salmonids. tion did, and immigration from surrounding areas Immediate impacts include direct mortality. habitat was a primary factor in the mammal recoveries. destruction. elimination of access to spawning and Colonizers of perturbed areas may be genetically rearing sites. and temporary reduction or elimina- predisposed to disperse (Sjorgen 1991) surplus to other populations (Hanski 1985; Pulliam 1988) or chance &rivals (Goodman 1987). Such adaptations increase the probability that metapopulations will persist through time'.
The ~ 6 a r n i c Aquatic Environment
Aquatic ecologists and managers often do not have the long-term dynamic view of ecosystems held by terrestrial ecologists (White and Pickett 1985) and advocated b i Holling (1973). Streams in the PNW (Resh et al. 1988) and elsewhere (Pringle et al. 1988; Reice 1994) are dynamic within rela- tively short $-me Fames; typically a year to a decade,
at the watershed scale, in response to floodsor mass wasting (Swanston 1991). It is generalty held that biological populations (some of them but not the
tion of food resources: 'Longer-term kffects may be positive. however. landslidesand debris flows'intro- duce essential habitat elements; sucii~as large'wood and sediment. into 'channels:and 'affect.:s'torage. of these- materials. The configuratiofibof-channel net- works:..the delivet?.. :storage; and .tianspon'of:iedi- ment .and wood. and the decomposit ion~'of!wd~~ debris interact to rreare, maintain;:'and 'distribute fish ,habitat over the long term. , -..: ' " :' ! i i, ..?, ::
Three streams in the central . Oregon* 'Coast Range were examined'to~~explore-"some "of .the.re- sponses of salmonids and their habitats to',the mat- ural disturbance regime (G. H. Reeves, U.S. Forest Senice. Pacific Northwest Research Station. un- published data). The :streams have giadients be-
w e e n 1 and 25% and drainage'areas.betwee'n 14 and 18 km'. Benda (1994) examined these and other streams as part of a study to model watershed - -
entire assemblage)' and physical features of these erosion and sedimentation. Summer habitats and systems recover relatively quickly after such distur- bances (e.g., Bisson et al. 1988; Lamberti et al. 1991; Pearson et al. 1992). Similar short-term responses of lotic fishes to disturbances have been noted in other areas (e.g., Hanson and Waters 1974; Mat- thews 1986). Over extended periods, habitat condi- tions in streams of similar size within a geomorphic region should be relatively uniform within and among watersheds (Vannote et al. 1980).
In contrast to terrestrial ecology, no theory pre- dicts the mosaic of aquatic conditions or ecological states caused by disturbances and the correspond- ing responses of fish populations over extended periods. Minshall et al. (1989), Naiman et al. (1992). and Benda (1994) have proposed that aquatic ecosystems are dynamic in space and time - 9 t h ~ . . ~ ~ ~ P I C ~ P A r ~ l l ~ Th- nma :--:-
assemblages of juvenile anadromous salmonids were inventoried in 1988 and 1989. The time since catastrophic wildfire and hillslope failure differed among streams.
The watershed of Harvey Creek was burned by an intense wildfire in the late 18005, and the forest was principally 90-100-year-old Douglas fir Pseudotsuga rnedesii at the time of the study. The .channel contained.,a large volume of sediment in storage throughout the lower portion of the drainage net- work and thus was considered to be in an aggrada- tional state (mean depth of deposits, 1.8 m). Evi- dence of burned wood in the channel indicated widespread landsliding followed the fire. Gravel was the dominant substrate (Figure 2). Larger substrate panicles and large woody debris were buried in the ---..-I A ---- :.- ---- - - - t - , ---- A..-.L n n - \

REEVES ET AL
Bedrock Boulder Cobble Gravel Fines
FIGURE I.-Substrate composition in three streams of the central Oregon Coast Range that had differing histories of major natural disturbance. The time since the last major natural disturbance was -100 yean for Hamy Creek, 160-180 years for Franklin Creek. and more than 330 years for Skate Creek. Cr = Creck (G. H. Retvcs unpublished data collected in July 1988 and 1989).
- r
. I
. - usually formed by scour around large wood. were the most common habitat units but were not hy- draulically complex. Fewer pieces of large wood were observed in Harvey Creek than in the other study streams (Table 2). because wood deposited in the channel by the hillslope failure had been buried beneath sediment and %little ,wood was being re- cruited from the relatively young surrounding forest The juvenile salmonid assemblage was numerically dominated by age-0 coho salmon Oncorfiyndruc kisutch, but age-1 steelhead 0. mykk (about 1%) and cutthroat trout 0. clarki (about 1%) were also present (Table 3).
The Skate Creek watershed was forested by trees more than 330 years of age, suggesting that the stream had not been subjected to a fire or hillslope failure for a long time. Habitat conditions in the stream were very simple. The substrate was pre- dominantly bedrock and boulders with small, local-
ized patches of stored sediment (Figure 2). Ritaes were thin sheets of water flowing over bedrock Although large wood was more abundant than in the other streams examined (Table 2). the lack of a deformable gravel bed greatly limited the wood's effectiveness in forming pools. Therefore, pools were shallow (mean depth, 0.1 m) and often in bedrock depressions. Juvenile coho salmon were the only salmonids found in Skate Creek (Table 3).
Franklin Creek was intermediate in time since disturbance. Based on the present vegetation, we estimated that catastrophic wildfire and landsliding occurred 160-180 years ago in this watershed. Mean depth of sediment in the channel was 0.7 m, and there was a greater array and more even d i u - tion of substrate types.than in the other streams (Figure 2). Mean pool depth was 0.35 m, less than half the mean depth of pools in Harvey Creek. As a result of sediment transport from the channel that
TABLE 2.-Mean number Of pieces Of large wood TAB= 3.40mposition of the assemblage of juvenile ('OJ in diameter and '3 long) per loo anadromous salmonids in three streams of the ecntral three streams of the central Oregon Coast Range h a t oregon aast R~~~~ that had diElcring ,.,istoria of major had dsehg hhtories of dkm*ance natural &tubana (G. He Reeves UnPUbwed dam al- (G. H. Reeves, unpublished data collected in July 1988
in July 1988 and 1989). and 1989).
Mean percent of estimated total ombus Ycan since last major Mean piem Age-1 Age-l
Stream disturbana of wood/100 m Stream coho *on steelhead cutthroat uout
Harvey Creek 90-100 7.9 . Harvey Creek 98.0 1 .O 1.0 Franklii Creek 160-180 1U Franklin Creek 85.0 123 2 3 Skate Creek >3M 23 5 Skate Creek . 100.0 0.0 0.0
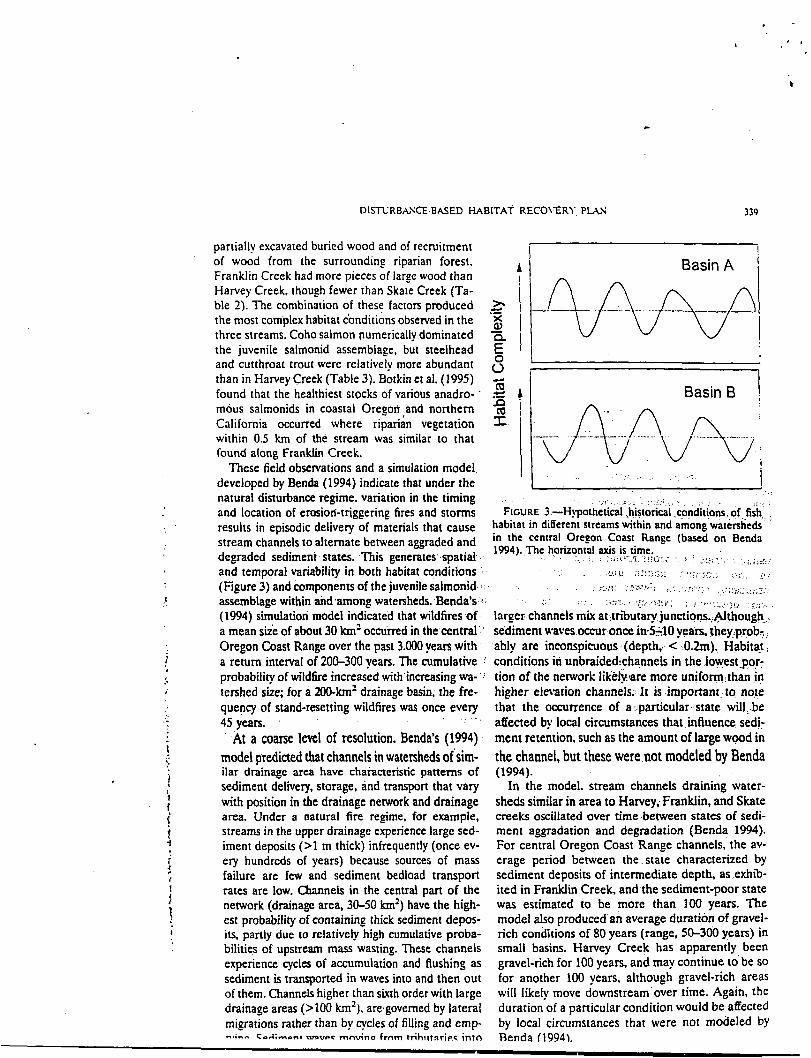
partially excavated buried wood and of recruitment I ! of wood from the surrounding riparian forest. A Basin A i Franklin Creek had more pieces of large wood than Harvey Creek, though fewer than Skate Creek (Ta- ble 2). The combination of these facton produced .- 3 the most complex habitat cbnditions observed in the 1 three streams. Coho salmon numerically dominated - the juvenile salmonid assemblage. but steelhead 1 1 and cutthroat trout were relatively more abundant 6 than in Harvey Creek (Table 3). Botkin et al. (1995) found that the healthiest stocks of various anadro- ' Basin 6 mous salmonids in coastal Oregori ,and northern California occurred where riparian vegetation within 0.5 krn of the stream was similar to that found along Franklin Creek.
These field observations and a simulation model. .. . , . - ... -. ..
developed by Benda (1994) indicate that under the natural disturbance regime. variation in the timing . .,. ..,. .I. .. . . . , . , . . ._ ., . ._ . ": ! . i i .. . . ,
and location of erosiori-triggering fires and storms FIGURE 3.-~!~otheti.al ,historical ,copditipns.:~f,fishd ': results in episodic delivery of materials that cause habitat in ditTerent streams within and among watersheds stream channels to alternate beween a w d e d and in 'he central Oregon Coast Range (based on Benda
1994). The horizontal a.+s ,is ti?: - degraded sedimeni states. This generates spatial .. : , : <!,:,.!::Y ,::,I,.i, , e : . .. %..:-,. ..' . . . . ,:;;;z4: and temporal variability in both habitat conditions . . : - >-',.i.;:.z . . . . .. , .! ,. .,!%..,; .. .- . ,,:-:, c (Figure 3) and components of the juvenile salmonid :: .- . . . -..L:j:li ~.-..;. , . . . . , ,. ... , .jc-x:, .. ... ,.::. + ... .
.. . assemblage \Githin ahd ,among watersheds. -Bendalsi: ... ;.! , :,z:. , -7z:,,,:.;:;r: : :'.'. ... . :.:'r .;7-,.
(1994) simulation model indicated that wildfires of . larger channels mix at,tributary junctiqns.,@though; a mean sizk of about 30 Ian' occu'md in the central:' sediment waves occur once in-540 years, they?prob-: Oregon Coast Range over the past 3.000 years with ' ably are inconspicuous .(depth,,. C ;O.Zm). Habitat, a return interval of 200-300 yean. The cumulative i conditions in unbraidedrchannels in the loyest,pqr- probability of wildfire increased with,increasing wa-': tion of the network likelylare more u n i f o ~ ~ t h a n in tershed size; for a 200-km' drainage basin, the fre- higher elevation channels.-.It is .important , t o note quency of stand-resetting wildfires was once every that the occurrence of a..;panicular..state will:..be
. . . . 45 years. : affected by local circumstances that. influence sedk At a coarse level of resolution. Benda's (1994) ment retention. such as the amount of large wood in -
model predicted that channels in watersheds of'sim- the channel. but these were .not modeled by Benda ilar drainage area have chaiacteristic panerns of (1994). sediment delivery, storage, and transport that vary In the model, stream channels draining water- with position in the drainage network and drainage sheds similar in area to Harvey, Franklin, and Skate area. Under a n a ~ r a i fire regime, for example, creeks oscillated over time .between states of sedi- streams in the upper drainage experience large sed- ment aggradation and degradation (Benda 1994). iment deposits (>1 m thick) infrequently (once ev- For central Oregon Coast Range channels, the av- ery hundreds of years) because sources of mass erage period between the. state characterized by failure are few and sediment bedload transport sediment deposits of intermediate depth, as .exhib- rates are low. Channels in the central part of the ited in Franklin Creek, and the sediment-poor state network (drainage area, 30-50 krn2) have the high- was estimated to be more than 100 years. The est probability of containing thick sediment depos- model also ~roduced'an average duration of gravel- its, partly due to relatively high cumulative proba- rich conditions of 80 yean (range, 50-300 yean) in bilities of upstream mass wasting. These channels small basins. Harvey Creek has apparently been experience cycles of accumulation and flushing as gravel-rich for 100 years, and may continue to'be SO
sediment is transported in waves into and then out for another 100 years, although gravel-rich areas of them. Channels higher than sixth order with large will likely move downstream'over time. Again, the drainage areas (>I00 krn2). aregoverned by lateral duration of a particular condition would be affected migrations rather than by cycles of filling and emp- by local circumstances that were not modeled by h e ; - - C -A ;mont ~ V P C mnvino frnm t r ih~rrnr ie~ into Renda (1994).

Juvenile salmonid assemblages are likely associ- Adaprarions of Anadromorcs Salmonids ated with each state predicted by the model. Ben- &adromous salmonid populations in Padc da's (1994) simulation indicated that sediment sup- are well adapted to dynamic ply would be limited at any given location in these Adaptations include straying by adults. high small streams a majority of the time. Based on field and mobilinl of jUveda. Straying by observations, coho salmon would have dominated is genetically conmlle& directly or indirectly such simplified habitats. When a channel segment (Quinn 1984). and aids the reatablisbent of pop was not in this degraded state. it would shift be- ulations in disturbed - on large (Neave 1958) tween states of aggradation and intermediate sedi- and local scales (Rickcr 1989). Strays would be ment supply. Two additional salmonid species. reproductively most successful where local popula- steelhead and cutthroat trout, are expected to occur tions have been reducedor extirpated (Tallman and in aggraded channels. The intermediate state is Healey 1994). provided there are suitable spawning characterized by intermediate sediment depths and and rearing ccmditions. Individuals from more than more complex habitat. which should support a ju- one population may recolonize depopulated areas, venile salmonid assemblage containing greater pro- increasing the genetic diversity of the new popula- portions of trout. Benda (1994) has developed long- tion. term average probabilities for the time a channel hhvements juveniles from natal to segment would have spent in each state. Applied to areas the estabrkhment of new
a population of channels (those with similar gradi- P ~ P ~ ~ ~ ~ ~ ~ ~ . may be genetically
ent, drainage area, etc.) for a particular time, these pmmmmed to move; may be -laced
probabilities can be used to estimate the landscape- from high-density populations (Northcote 1992).
scale mosaic of habitat conditions or biodiversity. Chapman (1962) suggested that juvenile salmonids that were unable to obtain territories and migrated
For example, in watersheds of an area similar to downstream were less fit individuals. However, at those in the field example (approximately 25 km2), some may vohrntsdly il eMdcm im- the frequency distribution developed by Benda prwes swivaL Trhapw and Hanma. (1983) (1994) indicates that a majority of channel segments found that jwenile coho salmon moving -- in the central Oregon Coast Range should have stream in a small ~ r i ~ i ~ h columbia stream took up limited sediment supplies at any parti~ular time and residence in unoccupied habit- and grew rapidly. thus should contain relatively siyple habitats. High fecundity contributes to the establishment
A natural mosaic of habitat conditions for and growth of a local population if conditic- are anadromous salmonids has likely existed elsewhere favorable. Pacific salmon are relatively fecund for in the PNW; the features and relative proportion of benthic-spawning fishes with large eggs Pink each channel state should vary with climate, vege- salmon Oncorhynchus ~ u s c h u , the smallest spe- tation. drainage pattern, and spatial scale. Meyer et cies, typically possess 1,200-1,900 eggs per female al. (1992) found cycles of aggradation and degrada- (Heard 1991). Adult female chinook salmon 0. tion associated with wildfires and hillslope failures t s h n ~ c h a , the largest species, may contain more in a Wyoming stream like those just described for than 17,000 eggs (HealeY 1991). Both high fecundity the central Oregon Coast Range. It seems reason- and large eggs contribute to the reproduck sue- able to assume that channel conditions over time CeSS of species whose Young have extended ~ e ~ o d s were similar to those observed in the Oregon of hmfFvel residence These traits also streams we examined. growth when conditions are suitable. 1
In summary, the natural disturbance regime of the central Oregon Coasr Range is described by the Human of Regimes frequency, size, and spatial distribution of wildfires Natural ecomems generally have a large capac- and landslides, and this regime has been responsible ity to absorb change without being dramatically for developing a range of channel conditions within altered. Resilience of an easystem is the degree to and among watersheds. The structure and compo- which the system can be disturbed and still return to sition of the juvenile anadromous salmonid assem- a domain of behavior m which processes and inter- .blage varies with channel conditions. A disturbance actions function as before (Holling 1973). If a dis- regime that resembles this natural regime must be turbance exceeds the resilience of the system, the incorporated into any recovery plan for freshwater domain may shift and the system will develop new habitats of ESUs of anadromous salmonids. conditions or states that had not previously been

DISTURBANCE-BASED HABITAT RECOVERY PL\h' 34 1
exhibited. Yount and Niemi (1990). modifying the disturbance definition of Bender et al. (1984). dis- tinguished "pulse" disturbances from "ptess" dis- turbances. A pulse disturbance allows an ecosystem to remain within its normal bounds or domain and to recover the conditions that were present prior to disturbance. A press disturbance forces an ecosys- tem to a different domain or set of conditions. Yount and Niemi (1991) considered many anthro- pogenic disruptions, such as timber harvesting and urbanization, to be press disturbances. Gum and Wallace (1984) hypothesized that stkam biota may not be able to recover from the effects of anthro- pogenic disturbances because these have no ana- logues in the natural disturbance regime. and or- ganisms may not have evolved the appropriate breadth of habitat or reproductive requirements.
Modifications in the type of disturbance or in the frequency and magnitude of natural disturbances can alter the species composition, habitat features. and resilience of an emsystem (White and Picken 1985; Hobbs and Huenneke 1992). Alteration or loss of habitats as a result of changes in the disturbance regime can bring on extirpation of some species. in- creases in species favored by available habitats. and invasions of exotic organisms (Levin 1974; Harrison and Quinn 1989; Hansen and Urban 1992). We also believe that changes in the legacy of disturbance (the conditions that exist immediately following a distur- bance) may be another important component of dis- turbance regimes that can be altered. Changes in the legacy can infiuence a system's resiliency by altering habitat conditions created immediately following a disturbance illad by altering future conditions.
ESUs of anadromous salmonids must accommo- date the d ~ ~ a m i c nature of the PNW landscape. Given the dynamic nature of terrestrial ecosystems (Agee 1991, 1993). the links between terrestrial processes and aquatic ecosystems. the apparent ad- aptations of anadromous fish for persisting in a dynamic environment. and the limited available ev- idence (based on central Oregon Coast Range streams) of non-steady-state behavior of sedimen- tation and habitats. we believe a dynamic.approach is advisable in any recoven prosram. In the.follow- ing sections. we describe the components ..that should be included in this approach.'
We do not mean to imply that every human action or activity causes a press disturbance: the impact of anthropogenic disruptions is minimal in some ecosystems (e.g., Attiwill 1994a, 1994b). How- ever, we believe human activities that affect anadro- mous salmonids and their habitats, such as timber harvesting, urbanization, and agriculture, do gener- ate press disturbances. These disturbances can re- '
sult in the loss of habitats or ecosystem states nec- essary for various anadromous salmonids (Hicks et al. 199L; Bisson et al. 1992). Human activities in the PNW have altered the recovery potential of ecosys- tems, which may be as responsible for the decline of habitat as the direct impact of the activity itself.
Waters/~ed Scale Resenmes: Sllon- a~ld - Long-term Considerations" ... .
. . . , . . , ? . , ,.. "' ~ ... .. , ..;:. . . . Anadromous , : ,..., salmonids'exhibit .. ,. t yp ica l f e i i t~~~s of
"patchy.populations": .. , I :; .-.,. !<., thei eGst in a'd$i@ic , ,_ * . . . - en"- . ronment and..have good.~iipersal~abilities'(Hamdin ,... -.:- - :.:; 1991,. i994j, . .:!< ' ~ohe&at i .on r . . . :. ,,c,:....b,..-j 'of patchy'.populat~ons $., . ,r.-.. .>?...,.....-
requires the Lconse@ation,~of :%merous' paiches..of .--: ,. -c .-<. - :- ,. ...... . suitable hatiitai'and .i..,-.r -,., .. . ".the" pote.ntial.. for. . d i sped l am&_e t h e m ' ( ~ e ~ ~ ~ i f9gl::'igg4)::~~&&j&-j spice , . . ,: , . . ." . . ing of rejewCi:bhou,d .de-pen"d',o." , ,j$6.&~$+&,~
. ...... / . dispersal '~ha*de.*;iid, of -thy"ip'e/iii'.!6f c."&~
(Simberloff .1988).'[.Rieman' and : McInt$k?~(1995) used logistic ft$~essioii~fo' invebiigate ihe:influence of patch size;-as well'as'stream'width andpadient, on populations of .bull trout Salr*elinur~confienncs at the reach. stream:-and watershed scales;: This approach could:.be ..helpful in identifying critical features of reserves for anadromous:salmonids,,~In our current' thinking, on reserve planning for ESUs
A Disturbance-Based Ecosystem Approach to Freshwater Habitat Recovery ,
We believe that any long-term propam for re-
of anadromous ra&onids, we ron~ide~patches to be watersheds. the size of which should depend on the species and geographic location. i t is difficult to predict' the exact number of patches required to sustain an organism (Lawton et al. 1994). Lande (1988) could do this for the northern spotted owl because data were available on essential life history variables. It is unlikely that predictions could be obtained for many other species, including ESUs of anadromous salmonids, because necessary life his- tory data are often lacking (Lawton et al. 1994).
In the Short term, reserves should be established in watersheds with good habitat conditions and functionally intact ecosystems to provide protection for these remaining areas. Reserves of this type are likely to be found in wildernesses and roadless areas on federal lands. Examples of watersheds that fulfill this requirement include some of the key water- sheds identified by Reeves and Sedell (1992). the

REEVES ET AL
Systems should qualify based on the extent of hab- itat degradation and the degree to which their nat- ural diversity and ecological processes are retained. Examples of such watenheds are some of the key watenheds identified by Reeves and Sedell (1992). some tier 1 key watenheds identified by Thomas et al. (1993). the class I11 waters of Moyle and Sato (1991). and the class I11 waters of Moyle and Yo- shiyama's (1994) aquatic diversity management ar- eas. Restoration programs implemented in' these watersheds should be holistic in their approach. They should address instream habitat concerns. prevent, further degradation. and restore ecological processes that create and maintain instream habi- tats.
It is imperative to recognize and acknowledge that identified reserves will experience natural and, often. anthropogenic disturbances. Thus, simply putting aside a fuced set of watenheds as reserves may not provide habitats of sufficient quantity and quality to ensure long-~enn persistence of ESUs. Conservation reserves have generally been estab-
i lished and managed without consideration of long- term disturbance dynamics and the biological and evolutionary processes that influence organisms contained within them (Western 1989). Conse- quently. their populations may have higher pmba- bilities of extirpation in the long term than ex- pected. Reasons for this include isolation of reserves from surrounding areas of suitable habitat resulting from habitat fragmentation (MacArthur and Wilson 1967; Diamond and May 1976); resuic- tion or elimination of migration and dispersal (Elsenberg and Hanis 1989; Harris and Elsenberg 1989); and boundary effects associated with sur- rounding areas, such as invasion of native and ex- otic competitots, disease, and pollution (Shone- wald-Cox 1983; Wilcox 1990). Hales (1989) and
FIGURE 4.-Distribution of tier 1 key watersheds iden- White and Branon (1980) noted that in dynamic tified by Thomas et al. (1993). landscapes, reserves may act as holding islands that
persist only for relatively short ecological periods (100-200 years). Reserves should be large enough
the tier 1 key watersheds of Thomas et al. (1993) to allow operation of the natural disturbance re- (Figure 4). Ideally these reserves should be distrib- gime and to support a mosaic of patches with dif- uted across the range of an ESU and should contain ferent biological and physical attributes (Pickett subpopulations of it. Because of the critical impor- and Thompson 1978). ' tance of these watersheds in the short term, activi- Gotelli (1991) noted that reserve strategies such ties within them should be minimized or modified as those proposed by Hamson (1991,1994) do not to protect the integrity of existing physical and eco- address the longevity of patches. This is a major logical conditions. concern in dynamic environments like those of the
Identification of watersheds that have the best PNW. Modification of the strategy proposed by potential for betng restored should also be a short- Hamson (1991, 1994) to accommodate a dynarnlc ;em prioricy of any recovery stratep. There water environment is a prudent approach in the d&lop sheds could serve as the next generation of reserves. ment of a recovery strategy for anadrornous salrno-

Natural
DISTCRBANCE-BASED H.-\BITAT RECO\'ERY PL4S .33
present fying practices to create a human-influenced distur- Future
FIGCRE 5.--Conceptual representation of the range of conditions experienced by aquatic ecoqstems historically. currentl!.. and under. a new disturbance regime (modified from H. Rcgier. University of Toronto. personal commu- nication).
nid habitats. Specifically, there is need for a shifting mosaic of reserves that change location in response to the ability of specific watersheds to provide suit- able habitat conditions.
A New Human-influenced Disturbance Regirne
Under natural wildfire regimes of the PNW. the condition of freshwater habitats for anadromous salmonids was likely regulated by episodic delivery of sediment and wood to the channel. Given that human demands on ecosystems will only increase. we believe that returning the entire landscape to the natural wildfire regime will not be possible. There- fore. human activities will have to be molded into an analogous disturbance regime if habitats are to re- cover and persist. First must come an understanding of how the natural disturbance regime created and
maintained habitats for anadromous salmonids through time and how i t has been modified by human activity. Then it will be necessary to identify those human activities that can be altered to main- tain desired ecological processes and leave the leg- acy that allows recovery and persistence of required freshwater habitats. In other words. the character of anthropogenic disruption must be shifted from a press to a pulse disturbance (Yount and Niemi 1990) (Figure 5). The following is an example of how we believe timber harvest and associated activ- ities, as currently practiced on federal lands in the central Oregon Coast Range, have affected habitat and biodiversity of anadromous salmonids and how these could be adjusted to help create suitable con- ditions in space and time. We believe that timber management may offer more immediate opponuni- ties than agricultural or urban processes for modi-
bance regime that maintains components of the natural regime.
Disturbance caused by timber harvest differs from.stand-resetting wildfires in the central Oregon Coast Range in several respects. One difference is the legacy of the disturbance. Wildfires. left large amounts of standing and douned wood (Agee 1991). which was often delivered to channels along with sediment in storm-generated landslides (Benda 1991). This promoted development of high- quality habitats as sediment wa; transported from the system. leaving the wood behind (Benda'l99.1). Timber harvest. as it is generally practiced. reduces the amount of large wood available to streams (Hicks et al. 1991: .Reeves el al. ,1993: Ralph et al. 1994). so when hanlest-related hillsiope failures-oc- cur: sediment is the prima? materialdeli;;e&d to the channel (Hicks et al. 1991). Because large.\vood is an integral component of aquatic @bitats ...l̂ ii:;.= a n d a major influence on sediment transponand stciEge. the potential for developing complex habitats,,, a much lower when small rather- thanL-lar@:iimTunts of wood are in the channel. ~onse~uentl$;;clfaiinels ma!. be simpler following timber harvestiti<" . .:!,=*.,. \hey are after wildfires. r+jp;& .;i.g<.; ;
The interval between events also affects the con- ditions that develop after a disturba?ce.,(~gbbs and Huenneke 1992). Under the natural'.$i~turb&ce regime. variation in the timing and loqiion of ero- sion-triggering fires and storms probably caused stream channels to alternate benveen aggraded and degraded sediment states, generating temporal vari- ability in both fish habitats and assemblages of ju- venile salmonids. Wildfires occurred on average about once every 300 years in'the central Oregon
Coast Range (Benda 1994). In watersheds smaller than 30 km', postfire development of the most di- verse physical and biological stream conditions may have taken 150 yean or more (see earlier discus- sion). Timber harvest generally occurs at intervals of 60-80 years on public lands and 40-50 years on private timberlands. This may not allow sufficient time for the development of conditions necessary to support the array of fishes found under natural disturbance regimes.
A third difference between timber harvest and a disturbance regime dominated by wildfire is the spatial distribution of each. Based on a fire fre- quency of once every 300 years. Benda (1994) esti- mated that on average, 15-25% of the forest in the central Oregon Coast Range would have been in early successional stages because of recent wildfires, In contrast, the area affected by timber harvest 1s

3 4 REEVES ET L
much greater. For example. in the Mapleton Dis- relatively lowquality habitats across the landscape trict of the Siuslaw National Forest. which contains and eliminate the potential for attaining the most the watersheds studied by Benda (1993). approxi- complex habitat states. mately 35% of the forest is in early succession In summary. the differences between present tim- (J. Martin. Siuslaw National Forest, personal com- ber harvest disturbance regime and the natural dis- munication). If private lands were included. the turbance regime have important implications for percentage would be greater. The present forested stream ecosystems and anadromous salmonids. landscape is more homogeneous with respect to Stream habitat at a point in the channel. is less seral stage than it was historically. Just as the dis- complex under the timber harvest regime (Hicks et tribution of terrestrial habitat has been altered by al. 1991; Bisson et al. 1992) than under the natural switching from a wildfire-driven to a harvest-driven regime. and the potential for achieving greater com- disturbance regime. it is also possible that the dis- plexity is also reduced. This is primarily a result of tribution of aquatic habitats is different today than the reduced legacy and shorter interval between ~t was under the natural disturbance regime and disturbance events under the timber harvest regime. thus less capable of supporting a diverse juvenile In addition. landscape-level habitat heterogeneity is salmonid assemblage. reduced under the harvest regime because the dis-
A fourth difference between the natural wildfire- turbance is more dispersed and widespread. driven and the current harvest-driven regime is the The new disturbance regime created by timber size of disturbance and the landscape pattern gen- harvest should address the concerns just listed The erated by the disturbance. Timber on federal lands l e g a j of hillslope failures associated with timber has typically been managed by widely dispersed management activities needs to include more large activities: approximately 174,000 krn of roads exist wood. Ben& (1990) identified the attributesof first- across public lands in the range of the northern and second-order streams that favor the delivery of spotted owl (Thomas et al. 1993). and many millions desirable material to fib-bearing channels. Increas- of hectares'have been affected by small harvests of ing the extent of riparian protection along these approximately 16 ha. Wildfires. on the other hand, streams. as proposed by Thomas et al. (1993). ob- often generate a larger but more concentrated dis- viously increases the potential delivery of wood. turbance. When wildfires occurred in the central Such a strategy may not result in wood loadings as Oregon Coast Range, they tended to 'be large large as occurred naturally because trees away from (mean, 3,000 ha), stand-resetting fires (Benda the riparian zone will have been removed. However, 1994). Conkquently, the spatial pattern and this strategy should increase wood loadings beyond amount of sediment delivered to channels would what is currently possible and should allow channels likely be different under these two disturbance re- to develop more complex habitats. gimes. In naturally burned areas. storms occurring Longer intervals between harvest rotations could during periods of low root strensth would generate be another component of this new disturbance re- large volumes of sediment from nearly synchronous gime. In single basins in the central Oregon Coast hillslope failures and channels would become ag- Range, the desirable interval may be 150-200 years, graded. Subsequently, delivery would be reduced although this is a fust approximation. The exact while source areas recharged. This, coupled with interval would depend on the magnitude and areal downstream flushing of stored sediments, would extent of the natural disturbance regime and the bring the channel to an intermediate level of sedi- time it takes for favorable habitat conditions to ment storage and a corresponding period of high- develop if adequate large wood and sediment are quality habitat. In unburned watersheds, sediment available. It will be Werent in other regions. Ex- delivery rates would remain low. In contrast. timber tending rotation time would also provide benefits to harvest activities are dispersed; thus, we presume many terrestrial organisms. that mass wasting would be more widely distributed Concentrating rather than dispersing manage- and would deliver sediment at elevated rates in ment activities could be another element of the new most managed watersheds. Storm-generated land- disturbance regime. 'lbis would more closely resem- slides would be asynchronous, being governed ble the pattern generated by natural disturbances through time by harvest schedules. Cycles of chan- than does the current practice of dispersing activlty nel aggradation and degradation probably would in small areas. For example, if a basin has four not be apparent and sediment delivery, at a land- subwatersheds it may be better to concentrate ac- scape scale, would likely be chronic rather than tivities In one for an extended per~od (50-75 years) episodic. These factors would conspire to produce than to operate in 25% of each one at any time

DISTURBAYCE-BASED HABITAT RECOVERY PUS 345
A. Dispersion of Activity
Time 1 1 Time 2
Time 3
, . , . .
. . . .
6. concentration of ~ c t i v i t ~
- Time 1 . I Time 2
Time 3 . 1 Time4
(Figure 6). Grant ( 1990) modeled such a scenario to determine its effects on patterns of peak flow and found that there was little difference benveen the nvo approaches. Franklin and Forman (1967) be- lieved that dispersing activity (Figure 6A) increases habitat and landscape fragmentation and is more detrimental than concentrating activities (Figure 68) to terrestrial organisms that require late-suc- cessional forests. We believe that concentrating ac- tivity would have similar benefits for the aquatic biota if the elements discussed previousl!. are':in- cluded. This approach could also be linked toplan- ning future resenses and reducing risks in resenpes. so it merits consideration in the de\*elopment of habitat recoven' efforts.
All of the elements discus&.ed above must be in- cluded in the development of a new disturbance regime if the re$me is~&;be successful,at creating and "maintaining bb i t a t i ' i ..;. for r ..,. anadronibus , . . - saimo- nids. .Exclusion of ,.any ,,element greatly reduces the potential for slt%eis:"Oui L . . _ _.. cifncept of designing a disturba.nce"regime arourid hiiman ,ac'iivitiescouid complement , . .. . p&n; $(oth&'iTrategies piop&&d for mariagement o f ;the' :ce,"'$tral 0r 'ego";~oasi ' ;~ange (Noss 1993)l:and. othei ..,-?,.. .$& '... .. of 'the- PNW- (e.g., .. ,.;-,'.
' ~ T h o t k et a!. 1993): ~ h t 5 ~ e ' ~ g ~ for reserves in jkhich ". '....,!.,:<
human activitj':is c"&jled'oi,,elimigaied. . ...., .. :. Thepro- posed new disturbance. 'iegime could.be apelied to
' . . ) . . ' . , .
'areas outside 'hi SUCK iese+e. e t e m . particularly . . . . .:.. . . . ., 2 :...:::- . in ttif short t~rm.,'It~couldz:~lso guide manigement siiategies in res'e(ves"whkre:Iimited hunian activity is allowed. The long-term goal'of this effofort. would be to create rehgia to replace and'cornplement refugia in permanently'desi~ated~reservk. such as wilderness areas and other withdrawn lands.'
Conclusions
Plans directed at the freshwater habitat for ESUs of anadromous salmonids in the Pacific Northwest must be focused on restoring and maintaining eco- system processes that create and maintain habitats through time. It is important to insure that as good habitats '.wink out," either through anthropogenic or natural disturbances or through development into new ecological states, others "wink on." Des- ignating 'the most intact remaining aquatic ecosys- tems as reserves is essential for meeting near-term requirements. In the long term. a static reserve system alone is unlikely to meet the requirements of
FIGURE 6.-Examples of patterns resulting from (A) these fish. Management must also be directed at
dispersing and (I) concentrating land manapmen[ ar- developing the next generation of reserves. Strate- tivities in a watershed over time (modified from Grant gies should be designed and implemented that trea! 149111 land manapemen1 a c t i v i r i e c e c r l i c t r r r h a n r ~ +=vrnrc

346 R E E E S ET AL.
be man~pulated so as to retain the ecological pro- while the senior author was a visitor at Waikato cesses necessary t o create and malntaln freshwater University. Hamilton. hew Zealand. We give spe- habitat through time. Although necessary for cial thanks to Jennifer Nielsen. who organized this anadromous salmonids, the approach of moving symposium and was very patient with us as a e wrote resenes and managing periodic disturbances may this paper. not be suitable for locally endemic or immobile biota. It is imperative to consider the needs of other organisms in the development of any habitat recov- References
en. program for ESUs of anadromous salmonids. Agee. J. K. 1991. Fire histop of Douglas-fir foresrs in the Xlany hurdles must be overcome to make our Pacific Northwest. Pages 25-34 i r ~ L F. Rugg~ero.
approach effective. First. biologists. managers. and K.B. Aubry. A. B. Care!*. and M- H- HUB. technical
planners need to think in longer time frames than coordinators. Wildlife and vegetation of unmanaged Douglmfir forests. U.S. Forest Service General they are generally accustomed to using. They need ,-,,chnifal~epon PNW-GTR-a5.
to acknowledge that ecosystems are dynamic in Agee. J. K. 1993. Ere ccolog of Pacific ~ o n h ; for- space and time over these longer periods. Simply ests. Island Press. Washington. DC. designating reserves and expecting these to function Attiwill. P. M. 1994a. The disturbance of fomt ecosys- as such for extended periods may be unrealistic: tems: the basis for conxcative management. Forest some benefits may accrue in the short tenn. but in Ecology and Management 63:247-M0.
Attiwill. P. M. 1994b. Ecolqical disturbance and the the long run it is unlikely that habitats of sufficient conxrvath memcnt of in Aus- quality and quantity will be available to sustain ,&A F~~~~~ Ecology and Management 0.1-346.
ESUS of anadromous salmonids. Expectations d Bend& L E. 1990. m e influence of debris flowson than- about habitat conditions in streams must change: a . nels and valley floors in the Orepn Coast Range, stream \\.ill not always have suitable habitats for USA. Earth Surface: Processes and Landforms 15: anadromous salmonids, and all streams should not 457-466. C h i c h e r , UK. be expected to have suitable habitats a t the same '- lgg4. Stochastic geomorpholOP in a humid
mountain laadscape. Doctoral disscnauon. Liniver- tlme. A consequence of a dynamic view is that. si, of Wathington, perspectives must be regional (Holling 1973). I h e Bender. E. & T. J. c=. and M. E. Gilpin. 1984. Per- percentage of the landscape that should contain turbation utpcrimcnts in community ecology: theory suitable habitats must be identified and the tempo- and practice. Ecology 65:l-13. ral and spatial distributions of these habitats deter- Bi*on- P. A. J- Nielsen. R. A. Palinason. and L E. mined. Grove. 1982 A system of naming habitat types In
small streams, with examples of habitat u t i l i i n by Finally. disturbance must be recognized as an salmonids during low stnamflow. Pages 62-73 in integral component of any long-term strategy. This N. B. mm ~ ~ ~ i s i " ~ ~ and will be a difficult hurdle to overcome. It requires of aquatic habitat in~ntory infonnahon. American educating resource managers, scientists. administra- Fisheries Society, Western Division, Beth& Mary- tors. politicians, and the public so they realize that land. periodic disturbance is not necessarily negative. T o P- '- and J- W. 1988.
Summer production of coho salmon stacked in the contrary, disturbance may be necessary in order ML SL nMms aher the 19w erup. to have productive habitats for ESUs of anadro- tion. Transadoas of the Amencan Fisheries Society mous salmonids in the PNW over long periods. 1 17322-335.
Acknowledgments
The development and refinement of the ideas within and context of this paper benefited from discussions and interactions with several people in- cluding Bernard Bormann. Ken Currens, Tom Dunne, Gordon Grant, Robert Gresswell, Brendan Hicks. Jim Hall, Hiram Li, Bruce Marcot, Tom Northcote, Scott Overton, Henry Regier, Bruce Rieman. Tom Spies, Fred Swanson, and Tommy Williams. Robert Gresswell. and Reed Noss pro- vided helpful reviews of early versions of this manu- script. Much of the research for this paper was done
Bisson. P. A, T. P. Quinn, G. H. Reeves. and S. V. Gregory. ,1992 Best management practices, cumula- tive effects, and long-term trends in fish abundance in Pacific Nonhwest river systems. Pages 189-232 in R. J. Naiman. editor. Watershed management bal- ancing sustainability and environmental change. Springer-Verlag. New York.
Botkin. D. B. 1990. Discordant harmonies: a new ecology for the twenty-first century. Oxford University Press, New York.
Botkin. D. B.. K. Cummins. T. Dunne. H. Regiu. M. So- bel. and L Talbo~ 1995. Status and future of salmon of western Oregon and northern California: findings and options. Center for the Study of the Environ- ment. Report #8. Santa Barbara. California
Chapman. D. W. 1962. Aggressive behavior in juvenile

coho salmon as a cause of emigration. Journal of the Fishcries Research Board of Canada lY:1047-1080.
Christensen, N. L 1985. Shrubland hre regimes and their evolutionar)' consequences. Pages 86-100 k S. T. A. Pickett and P. S. White. editors. The ecology of nat- ural disturbance and patch dynamics. Academic Press. Orlando. Florida. .
Currens. K. P.. and six coauthors. In press. A hierarchial approach to conservation genetics and production of anadromous salmonids in the Columbia River basin. US. National hlarine Fisheries Service Fishery Bul- letin.
Diamond. J. M.. and R. hJ. Ma\: 1976. Island biqeogra- phy and the design of nature reserves. Pages 16.3-186 in R. M. May. editor. Theoretical &ology. Saunders. Philadelphia.
Elsenberg. J. F.. and 1. D. Harris. 1989. Conservation: a consideration of evolution. population. and life- history. Pages 99-108 in D. Western and M. Pearl. editors. Consewation for the twenty-first century. Oxford University Press, ,New York.
Franklin, J. F. 1993. Preserving biodiversity: species. eco- systems, or landscapes?. Ecological Applications
. 3:202-205. Franklin. J. F- and R. T. T. Fonnan. 1987. Creating land-
scape patterns fomt cutting:. ecological conse- quences and principles. Lankape Ecology 1:5-18.
Frisxll, C. A. ,1993. Top010gy of extinction and endan- germent of native fishes in the 'Pacific Northwest and California (U.S.A.). Conservation Biology 7:342-354.
Goodman. D. 1987. Consideration of stochastic demog- raphy in the design and management of reserves. Natural Resources Modeling 1:205-234. Tempe. ~ r -
... ,
izona Gotelli. N. J. 1991. Metapopulation 'models: the rescue
effect. the propaple rain. and the core-satellite hy- pothesis. American Naturalist 13876&776.
Grant. G. E 1990. Hydrologic, geamorphic and aquatic habitat implications of old and new forestry. Pages 35-53 in k F. Pearson and D. A. Challenger, editors. Foress-tnanaged and &Id: differences and conse- quences. University of British Columbia. Vancouver.
Gum. M. E, and J. B. Wallace. 1984. Substrate-medi- atcd mponse of invertebrates to disturbance. Ecol- ogy 65:1556-1569.
Hales. D. 1989. Changing concepts of nalional parks. Pages 139-149 in .D. Western and M. Pearl. editors. Conservation for the twenty-first century. Oxford University Press, New York
Hansen. k 1, and D. L. Urban. 1992. Avian response to landscape patterns: the role of species life histories. Landscape Ecology 7:16%180.
Hanski. I. 1985. Single-species metapopulation dynam- ics: conapts, models. and observations. Biological Journal of the Linnean Society 4217-38.
Hanski. I.. and M. Gilpin. 1991. Metapopulation dynam- in: brief history and conceptual domain. Biological Journal of the Linnean Society 423-16.
Hanson. D. L and T. F. Waters. 1974. Recovery of standing crop and production rate of a brook trout population in a Rood-damaged stream. Transactions of the American Fisheries Societ). 103:431439.
Harris. L. D.. and J. F. Elsenberg. 1989. Enhanced link- ages: neccssan. steps for success in consemation of faunal diveniry. Pages 166.181 in D. Western and hl. Psarl. cditiors. Conservation for the. nventy-first centun.. Osford Universi~ Press. New York.
Harrison. S. 1991. Local extirpation in a metapopulation contest: an empirical evaluation. Biological Journal of he Linnean Society 4273-88.
Harrison. S. 1994. hletapopula~ions and consemation. Pages 11 1-128 ill R. I. Edwards. R. M. May. and N. R. \\'ebb. editors. Large-scale e m l o p and conser- vation b i o l o ~ . Blackwell Scientific Publications. Lon- don.
Harrison. S.. and J . F. Ouinn. 1989. Correlated environ- ments andthe persistence of metapopulations. Oikos 56:293-298. .
Heale?. hl. C. 1991. Lifc histoy of chinook salmon (011- corlt~r~cl~~is rsha\cyscl~a ). P a ~ s 31 1-394 i t1 C. Gioot and L hlargolis. editors. Pacific salmon life histories. University of British Columbia Press. Vancouver.
Heard. H'. R. 1991. Life history of pink salmon (011;
c o r l ~ ~ ~ ~ c l ~ i u gorbilsclra). Pages 121-230 br C. Groot and L. hiargolis. editors. Pacific salmon life histories. University of British Columbia'Press. Vancouver.
Hicks. B. J.. J. D. Hall. P. A:' Bisson. and'J, R. .Sedell. , - 1991.. Responses of salm6iidsc to habitat changes.
American Fisheries Socieiy Special Publication 19: 483518. . . ; .- .,, .. . . . . . . . . . - . ..:. 7: ? ;
Hobbs. R J.. and L. F. ~uenneke.:.1992~~isturbance. diversity. and invasion: implications for conse~ation. Conseneation Biology 6:324-337.. . .: ., . .
Holling. C. S. 1973. Resilience and stability of ecoloiical systems. Annual Review of Ecologv and Systematics 4:1-23.
Huff. M. H.. and C. M. ~ a l e y . 1991. ~ e ~ i o n a l . ~ a t t e m s of diurnal breeding bird communities .in Oregon and Washinnton. Pages 177-206 in L. F. Ruggiero. K. B. Aubry. A. B. Carey, and M. H. Huff. technical coor- dinators. Wildlife and vegetation of u v g e d Doug- las-fir forests. U.S. Forest Service General Technical Report PNW-GTR-285.
Kan, J. R. and K. E. Fnemark. 1985. Disturbance and vertebrates: an integrative perspective. Pages 153- 168 in S. T. A. Pickett and P. S. White, editors. The ecologv of natural disturbance and patch dynamics. Academic Press. Orlando. Florida.
Larnbeni. G. A.. S. V. Gregory, L. R. Ashkenas. R. C. Wildman. and K M. S. Moore. 1991. Stream ecosys- tem recover). following a catastrophic debris flow. Canadian Journal of Fisheries and Aquatic Sciences 48:196-208.
Lande. R 1988. Genetics and demography in biological consen.ation. Science 241: 1455-1459.
Lawton. J:H.. S. Nee. A. J. Letcher. and P. H. Harvey. 1994. Animal distributions: patterns and processes. Pages 41-58 in R. J. Edwards. R. M. May, and N. R. Webb. editors. Large-scale ecology and conservation biology. Blackwell Scientific Publications. London.
Levjn, S. 1974. Dispersion and population interactions. American Naturalist 108:207-228.
Li; H. W.. C. B. Schreck. C. E. Bond. and E. Rexstad. 1987. Factors influencine changes in fish assemblages

REEVES ET A L
of Pacific Konh\vest streams. Pages 193-202 itt W. J. gon coastal streams. Canadian Journal of Fisheries htatthews and D. C. Heins. editors. Community and . and Aquatic Sciences 49:783-789. evolutionary ecology of North American stream Northcotc. T. G. 1992. Migration and residency in stream fishes. University of Oklahoma Press. Norman. salmonids--some ecological considerations and evo-
MacArthur. R. H.. and E. 0. Wilson. 1967. The theoy of lutionary consequences. Nordic Journal of Freshwa- island biogeography. Princeton University Press. ter Research 6675-17. Princeton. New Jersey. Noss. R. F, 1993. A conservation plan for the Oregon
Mangel. M.. and C. Tier. 1994. Four facts evely conser- coast range: some preliminary suggestions. Natural vation biologist should know about persistence. Ecol- Areas Journal 13276-290. Rockford, Illinois. ogy 753607-614. Pearson. T. N, H. W. Li. and G. A. Lambcrti. 1992.
Matthews. W. J. 1986. Fish community structure in a lnfluencc of habitat complexity on resistance to temperate stream: stability, persistence. and a cata- flooding and resilience of stream fish assemblages. strophic Rood. Copcia 1986:38&397. Transactions of the American Fisheries Society 121:
Ma!.. R. M. 1994. The effects of spatial scale on ecologi- 327436. cal questions and answers. Pages 1-17 in R. J. Ed- Pickett. S. T. X.and J. N. Thompson. 1978. Patch dy. wards. R. ht. May. and N. R. Webb. editors, h g e - namia and the design of nature reserves. Biological scale ecoloe\. and conservation biology. BlacLwell Conservation 1327-37. Scientific Publications. London. Pringle. C. M.. and seven coauthors. 1988. Patch dynam-
%fcCauley. D. E. 1991. Genetic consequences of local ia in lotic ?%ems: the stream as a mosaic. Journal of population eninaion and recolonization. Trends in the North American Benthological Society 7(4):503- Ecology & Evolution 6(1):5-8. 324.
hfclnrosh. B. A.. and six coauthors., 1994. Historical Pulliam. H. R 1988: Soums. sinks. and population reg- changes in fish habitat for select river basins of east- ulation. American Naturalist 132652461. em Oregon and Washington. Northwest ,Science Quinn. T. F. 1984. Homing and straying in Pacific 68(Special lssue):36-53. .salmon. Pa-= 357-362 in J. D. McCleave. G. P.
Meyer. G. A.. S. G. Wells. R. C. Balling. ~ r , k d A-J. T. Arnold. J. J. Dodsoa and W. H. NeiL editors Mech- Jull. 1991. Rrsponsc of alluvial systems to fire'and a n h s of migration in fish. Plenum New York. climate change in Yellowstone N a t i o d Park. . , Nature ~ a l p h . S. C- G. C. Poole. L ~ " ~ o n ~ u e s t . and R. J. 357: 147-150. Naiman. 1994. Stream channel morphology and
Minshall. G.' W.. J. T. Brock, 'arid J. D. v;&. -'1989.: '.woody debris in logged and unlogged basins of west- Wildfires and Yellowstone's stream ewsystexps-Bio- ein Washington. Canadian Journal of Fisheries and' Science 39:707-715. Aquatic Sciences 51:37-51. ,
Moyle. P. B.. and G. M. Sato., 1!%1. ~n"th~' , 'd&gn'of , ~ a i h a e ~ M. G. !9?L Vertebrate spehes richncss within preserves to protect native fishes. Pag,? 155-169 in and among xral stages of Douglas-fir~hardwood for- W. L. Minckley and J. E. ' ~ e k o i ; tditon.;Battle ,, ests of nonhwestern California. Pages 415424 in against exlinction: native fish management in the L F. Ruggiem. K Br Aubry. A. B. Carey. and M. H. American west. University of Arizona-Press, Tuscon.
"
Huff. technical coordinators. Wildlife and vegetation Moyle. P. BT. and R M. Yoshiyama. 1994. Protection of of unmanaged Douglas-fir forests. US. Fomt Ser-
aquatic biodiversity in California: a five-tiered ap- ' via General.Technieal Report PNW-GTR-285. proach. Fisheries 19(2):&19. Rapport. D. J, K A. Regicr. and T. C. Hutchinson. 1985.
Murphy. M. L. J. Heifetz J. F. Thedinga, S. W. Johnson, Ecosystem behavior under strm. American Natural- and K V. Koski. 1989. Habitat utilization by juvc- ist 125:617-640. nile Pacilic salmon (Oncorhynchus) in the glacial Reeves. G. H.. and J. R Sedell. 1992. An ecosystem ap- Taku River, southeast Alaska. Canadian Journal of proach to the conservation and management of fresh- Fisheries and Aquatic Sciences 46:1677-1685. water habitat for anadromous salmonids in the Pacific
Naiman. R. J.. and eight coauthors. 1992 Fundamen@ Northwest. Transactions of the North American elements of ecologically healthy watersheds in the Widlife and Natural Resources Conference 57:408- Pacific Northwest coastal ecoregion. Pages 127-188 in , 415. R. J. Naiman. editor. Watershed management: bal- Reeves, G. H- F. H. Everen and J. R. Sedcll. 1993. ancing sustainability and environmental change. Diversity of juvenile anadromous salmonid assem- Springer-Verlag. New York. blages in coastal Oregon basins with different levels
Neave. F. 1958. The origin and speciation of Oncow- of timber harves~ Transactions of the American Fish- chus. Transactions of the Royal Society of Canada eries Society 122309-317. 5225-39. Reice. S. R. 1994. Nonequilibrium determinancf of bio-
Nehlsen. W.. J. E. Williams, and J. A. Lichatowich. 1991. logical community structure. American Scientist 82: Pacific salmon at the crossroads: stocks at risk from 424-435. California. Oregon, Idaho, and Washington. Fisher- Resh, V. H.. and nine coauthors. 1988. The role of dis- ies 16(2):4-21. turbance in stream ecology. Journal of the Nonh
Nickelson. T. E.. J. D. Rodgen. S. L. Johnson, and M. F. American Benthological Society 7:433-455. Solazzi. 1992. Seasonal changes in habitat use by ju- Ricker. W. E. 1989. History and present state of odd-vear
;othilk Coho sdlffmn:(Ou&rhvnchus kisutch) in Ore- pink salmon runs of the Frasier River region. Cana- ??.,: 2.:: 1 r . i .,:.:...>2

DISTURBANCE-BASED HA BITAT RECOVERY PLM 349
dian Technical Report of Fisheries and Aquatic Sci- ences 17021-37.
Rieman. 9 . E.. and J. D. Xfclnryre. 1995. Occurrence of bull trout in naturally fragmented habitat patches of varied size. Transactions of the American Fisheries Sociey 1.?4:385-296.
Rolstad. J. 1991. Consequences of forest fragmentation for the dynamics of bird populations: conceptual is- sues and evidence. Biological Journal of the Linnean Sociep 41:349-163.
Schlosser. 1. J. 1991. Stream fish ecology: a landscape perspective. BioScience 4 1:701-712.
Shonrwald-Cos. C. hi. 1983. Guidelines to management: a beginning attcmpt. Pages 414415 in C. M. Shon- ewald-Cos. 5. ht. Chambers. B. MacBryde. and L. Thomas. editors. Genetics and conservation: a ref- rrencc for managing wild animal and plant popula- tions. Benjamin Gummings Co.. Menlo Park. Califor- nia.
Sheldon. A. 1. 1988. Conservation of stream fishes: pat- tcms of diversity. rariry. and risk. Conxrvation Biol- ogy 2: 149-156.
Simbcrloff. D. 1988. The contribution of population and community bioloa to conservation science. Annual Review of Ecology and Systematics 19:473-511.
Sjorgen. P. 1991. Extinction and iyladon ,gradients in metapopulations: the case of the pool frog (Rana lessotiae). Biological Journal of the Linnean Society 42:135-147.
Stearns. S. C. 1977. The evolution of life history train:.a critique of the theoff and a reiilew-of the data. An- nual Review of Ecolog and Systematia 8:145-171.
Stcedman. R. J.. and H. J. Regier. 1987. Ecosystem sci- ence for the Great Lakes: perspectives on depdative and rehabilitati\.e transformation.i Canadian Journal of Fisheries and Aquatic Sciences 44(Supplement): 95-103.
Swanston. D. N. 1991. Natural processes. American Fish- eries Societ): Special Publication 19:139-379.
Tallman. R. F.. and M. C. Healey. 1994. Homing, stray- ing. and gene now among seasonally separated pop- ulations of chum salmon (Oncorhynchur keta). Cana- dian Journal of Fisheries and Aquatic Sciences 51: 577-588.
Thomas. J. W.. and the Forest Ecosystem Management ksessment Team. 1993. Forest ecosystem manage- ment: an ecological. economic. and social assessment. Repon of the Foren Ecosystem Management Assess- ment Team. ljnited States Department of Agricul- ture. Forest Service. Portland. Oregon.
Tschaplinski, P. J.. and G. F. Hartman. 1983. Winter dis- tribution of jwenile coho salmon (Oncorfiynchus kicuiclt) before and after logging in Carnation Creek,
British Columbia. and some implications for overwin- ter survival. Canadian Journal of Fisheries and Aquatic Sciences 10:152461.
Vannote. R. L. G. W. Minshall. K. W. Cummins. J. R. Sedell. and C. E. Cushing. 1980. The river contin- uum concept. Canadian Journal of Fisheries and Aquatic Sciences 37:130-137.
Vrijenhoek. R. C. 1985. Animal population genetics and disturbances: the effects of local esrinction and re-
. colonization on heterogeneiry and fitness. Pages 365- 285 br S. T. A. Pickeu and P. S. White: editors. The ecology of natural disturbance and patch djnamics: Academic Press. Orlando. Florida.
Waples. R. S. 1991. Pacific salmon. Ot rcod~~~~ch i r s spp.. and the definition of "species" under the Endangered Spccies Act. U.S. National Marine Fisheries Scvice Marine Fisheries Review 53(3):11-22.
Webb. N. R.. and J. A. Thomas. 1991. Consenring insect habitats in heathland biotypes: a question of scale. Pages 129-151 irr R. J. Edwards. R. hl. May. and N. R. Webb. editon. Large-scale ecology and conser- vation bioloe. Blackueell Scientific Publications. Lon- don.
Western. D: 1989. Conxrvation without parks: svildlife in rural landscapes. Pages 156-165 in D. Western and M. C. Pearl. editors. Conservation for'the twentyfirst century. Oxford University Press. ~ e y , Y o r k .
White. P. S.. and S. P. Bratton. 1980. After preservation: the philosophical and practical problems of change. Biolagidl Conservation 18:241-35,
White. P. S.:and S. T. A. Pickett. 1985:gatural distur- . .
bance and patch dynamics: an introdunion. Pages 3-13 in S. T. A. Pickett and P. S. White. editors. The ecology of natural disturbance and patch dynamics. Academic Press. Orlando. Florida.
Wilcove. D. S.; C. H. h-lclellan. and A. P. ~ o b s o n . 1986. Habitat fragmentation in the tempeiate zone. Pages 237-256in M. E Soule. editor. Consen-ation biology the science of scarcity and diversity. Si'nauer. Sunder- land. Massachusetts:
Wilcor'B. A. 1990. In siru conservation of genetic re- sources. Pages 45-77 in G. H. Orions. G. M. Brown. Jr., W. E. Kunin, and J. E. Swierzbinski. editors. The preservation and valuation of biological resources. University of Washington Press. Seattle.
Williams, J. E.. and seven coauthors. 1989. Fishes of North America endangered. threatened, or of special concern. Fisheries 14(6):2-20.
Yount. J. D.. and G. J. Niemi. 1990. Recovery of lotic communities and ecosystems from disturbance-a narrative review of case studies. Environmental Man- agement 14547-570.
Rmkrard by USDA Fond knb lor ollklrl uu