expression and regulation of functional oxytocin receptors
TRANSCRIPT

Expression and Regulation of Functional Oxytocin Receptors in Bovine T 1
Lymphocytes1 2
3
Short Title: Oxytocin receptors in bovine immune cells 4
5
Kalidou Ndiaye, Daniel H. Poole and Joy L. Pate 6
Department of Animal Sciences, 7
The Ohio State University/Ohio Agricultural Research and Development Center, 8
1680 Madison Ave, Wooster, Ohio 44691 9
10
1 This project was supported by National Research Initiative Competitive Grant no. 2004-11
35203-14789 from the USDA Cooperative State Research, Education, and Extension 12
Service Animal Reproduction Program. Salaries and research support also provided by State 13
and Federal funds appropriated. 14
15
Address correspondence to: 16
Dr. Joy L. Pate 17
Department of Animal Sciences 18
The Ohio State University/Ohio Agricultural Research and Development Center 19
1680 Madison Avenue, Wooster, OH 44691 20
Fax: 330-263-3949 21
Email: [email protected] 22
23
BOR Papers in Press. Published on December 19, 2007 as DOI:10.1095/biolreprod.107.065938
Copyright 2007 by The Society for the Study of Reproduction.

2
ABSTRACT 1
The corpus luteum (CL) produces oxytocin (OXT) which has been proposed to 2
regulate the pulsatile release of prostaglandin F2 alpha during luteolysis in ruminants. 3
This action of oxytocin is mediated via oxytocin receptors (OXTR) present on uterine 4
epithelial cells. It is hypothesized that luteal oxytocin acts as a paracrine regulator of 5
resident immune cells. In the present study, OXTR mRNA expression in bovine 6
lymphocytes was analyzed as well as its regulation during the estrous cycle. OXTR 7
transcripts were observed in freshly purified bovine peripheral blood mononuclear cells 8
(PBMC) and T lymphocytes. OXTR mRNA in bovine lymphocytes was numerically 9
greater but not significantly different on day 3 than on day 19 of the estrous cycle (P = 10
0.091). In cultured T cells, estradiol (E2) treatment significantly increased the steady-state 11
concentrations of OXTR mRNA, but the stimulatory effect of E2 was inhibited by the 12
addition of progesterone (P4). Each of the major T cell subsets (CD4+, CD8+, and gamma 13
delta+) expressed OXTR mRNA with no significant difference in expression among them. 14
Western blot analyses demonstrated the presence of the bovine OXTR protein at about 45 15
kDa in lymphocytes, as well as expression of the 14 kDa precursor of oxytocin. When 16
lymphocytes were treated with OXT, intracellular concentrations of calcium ([Ca2+]i) 17
were rapidly and dramatically increased. This study demonstrated that bovine 18
lymphocytes express OXT receptors and that this expression can be regulated in a 19
steroid-dependent manner. Furthermore, OXT elicited a functional [Ca2+]i response in T 20
lymphocytes supporting the possibility that OXT within the CL could act as a paracrine 21
or autocrine regulator of resident T lymphocytes. 22
Keywords: Bovine; oxytocin receptor; lymphocytes; oxytocin; calcium.23

3
INTRODUCTION 1
The nonapeptide oxytocin (OXT) is mainly produced by the hypothalamus and 2
transported in neurosecretory vesicles to the posterior pituitary where it is stored or 3
released into the bloodstream after specific stimulation. The major endocrine functions of 4
OXT include uterine contraction during parturition and the contraction of the mammary 5
gland for milk ejection [1]. In addition, there is evidence that the corpus luteum (CL) of 6
ruminants also synthesizes and secretes oxytocin [2, 3]. This discovery led to the 7
proposal that oxytocin stimulates the pulsatile release of prostaglandin F2α (PGF2α) from 8
the uterus, resulting in regression of the CL [4-7]. It has also been shown that oxytocin is 9
involved in the regulation of the proliferation of various cell types including mammary 10
gland epithelial cells and endometrial epithelial cells under both physiological and 11
neoplastic conditions [8-10]. 12
The biological actions of OXT are exerted through specific, membrane-bound 13
OXT receptors (OXTR) [11-13]. The structure of human and bovine OXTR proteins has 14
been elucidated [14, 15] and the predicted amino acid sequences indicate that the OXTR 15
belongs to the large family of G-protein coupled receptors with a typical seven 16
transmembrane domain structure [15]. Upon stimulation by OXT, OXTR initiates a 17
signal cascade resulting in the hydrolysis of phosphatidylinositol 4, 5-bisphosphate by 18
phospholipase C to diacylglycerol, which stimulates a protein kinase C, and to inositol 19
triphosphate [16]. The inositol triphosphate pathway leads to intracellular calcium (Ca2+) 20
release from intracellular stores and the diffusion of Ca2+ into the cytosol for induction of 21
specific biological responses. The traditional role of OXT has been extended by several 22
reports about its biological activities and by the presence of OXTR mRNA in tissues that 23

4
have not previously been considered as conventional targets for OXT [10, 17, 18]. For 1
instance, receptors for oxytocin have been recently identified in the thymus of embryonic 2
mice [19] and in freshly isolated lymphocytes from human peripheral blood [20]. 3
In the reproductive system, immune cells are involved in the progression of 4
luteolysis once it has been initiated by PGF2α and likely contribute to both continued 5
decline in progesterone production as well as structural demise of the CL [21-23]. The 6
CL is composed of a heterogeneous mixture of cell types that consist of steroidogenic 7
cells and nonsteroidogenic cells, including vascular endothelial cells, fibroblasts and 8
immune cells. In the cow, lymphocytes are present in the fully functional CL and increase 9
at the time of luteal regression [24, 25]. In addition, cultured bovine luteal cells were 10
shown to be potent stimulators of T lymphocyte proliferation [26]. It is proposed that 11
communication between steroidogenic or endothelial cells and immune cells occurs 12
throughout the lifespan of the CL, not only during luteolysis. The demonstration of OXTR 13
mRNA expression in the CL of sheep [27] and pigs [28], and in cultured bovine luteal 14
cells [29] suggested an autocrine or paracrine role for luteal OXT. Acting as a paracrine 15
factor, it is plausible that OXT is involved in the regulation of the function of immune 16
cells that reside in the CL. 17
The long term goal of these studies is to identify factors involved in luteal cell 18
regulation of T lymphocytes. Functional status of immune cells within the CL are likely 19
regulated by a balance of paracrine factors and alterations of this balance may influence 20
whether immune cells regulate normal homeostasis of luteal tissue or exert 21
proinflammatory function during luteolysis. Luteal oxytocin is proposed to be one of 22
these factors. Therefore, the specific objectives of this study were to determine if bovine 23

5
immune cells express OXTR mRNA, if temporal changes in expression occur during the 1
estrous cycle, how such expression is regulated, and if OXTR proteins in lymphocytes 2
are functional. 3
4

6
MATERIALS AND METHODS 1
1. Isolation of PBMC and T Lymphocytes 2
Peripheral blood mononuclear cells (PBMC) were isolated from whole blood 3
collected via jugular venipuncture on days 3, 11, and 19 (D3, D11, and D19) of the 4
estrous cycle (day of estrus = day 0; n = 4 cows on each day). The lymphocyte-rich white 5
blood cell layer was obtained following centrifugation of whole blood and PBMC were 6
isolated by centrifugation over Ficoll-Paque Plus (Amersham Pharmacia Biotech). An 7
aliquot of fresh PBMC was taken for total RNA extraction. The T lymphocytes were 8
separated from the PBMC immunomagnetically by positive selection with anti-CD2 and 9
anti-gamma-delta (γδ) antibodies using the AutoMACS Separator (Miltenyi Biotech, 10
Germany) as recommended by the manufacturer. Briefly, PBMC (1x108 cells) were 11
incubated with mouse anti-CD2 (MUC2A) and anti-γδ (α-TcR1-N24 GB21A; VMRD, 12
Inc., Pullman, WA) antibodies (5µg/ml for each antibody), washed as previously 13
described [30] and labeled with rat anti-mouse IgG2a+b microbeads (Miltenyi Biotech). 14
Anti-CD2 antibodies were used to selectively target the CD2+ T lymphocytes including 15
natural killer (NK) T cells, while anti-γδ antibodies target the gamma-delta T 16
lymphocytes. This separation procedure yielded a population of cells that was 17
approximately 95% pure T lymphocytes as determined by fluorescent labeling with anti-18
CD3 (MM1A; T cell receptor) and anti-CD335 (MCA2365; NK cells) antibodies 19
(VMRD, Inc.). The negative fraction that contains the macrophages and B cells was 20
collected to analyze OXTR mRNA expression for comparison with purified T cells. The 21
number of viable T cells was determined using the Guava ViaCount Flex Reagent in the 22
Guava EasyCyte system (Guava Technologies, Inc.). The T lymphocytes (TC) were 23

7
subsequently divided for culture or for total RNA extraction. T cells were cultured in the 1
presence or absence of Concanavalin A (125µg/ml; Calbiochem), 10ng/ml of 2
progesterone (P4; Sigma-Aldrich), and/or 15pg/ml of 17β-estradiol (E2; Sigma-Aldrich). 3
Cultures were carried out for 72 hours at 37°C in RPMI-1640 medium containing 10% 4
fetal bovine serum (Gibco, Invitrogen Corporation). At the end of the culture, T cells 5
were collected for total RNA extraction and for OXTR mRNA quantification by reverse 6
transcription followed by quantitative polymerase chain reaction (RT-qPCR). A portion 7
of the freshly collected PBMC were further separated into different subpopulations using 8
anti-CD4, anti-CD8, and anti-γδ antibodies (VMRD, Inc.), separately, in order to analyze 9
OXTR mRNA expression in these different T cell subsets. 10
11
2. Collection and Dissociation of Corpora Lutea and Coculture Experiments 12
Corpora Lutea (CL) were collected transvaginally from cyclic cows (n = 4) during 13
the midluteal phase (day 11; day 0 = day of estrus) of the estrous cycle. Immediately 14
following CL removal, 500 ml of jugular venous blood were collected as described 15
above. Handling of animals and surgical procedures were conducted according to 16
protocols approved by the Institutional Laboratory Animal Care and Use Committee of 17
the Ohio State University. Dissociation of luteal tissues was performed according to the 18
procedure described previously [31]. Luteal tissues were minced and placed in 24mM 19
Hepes-buffered Ham’s F-12 culture medium (Gibco, Invitrogen Corporation) containing 20
0.5% bovine serum albumin (Sigma Aldrich, Inc.), 20µg/ml gentamycin (Gentamycin 21
Reagent Solution; Invitrogen Corporation), and 2000U/g tissue collagenase type I 22
(Worthington Biochemical Corporation). Dissociation, dispersion, as well as 23

8
centrifugations and washes were performed as previously described [31]. Following 1
dissociation, luteal cells were resuspended in Ham’s F-12 culture medium and the 2
number of viable cells was counted using the Guava ViaCount program in the Guava 3
EasyCyte system (Guava Technologies, Inc.) The luteal cells (LC; 5x105 cells) were 4
cocultured with TC (2x106 cells) in RPMI-1640 medium containing 10% heat-inactivated 5
fetal bovine serum and treated with progesterone (0.5µM), oxytocin (OXT, 100ng/ml; 6
GenScript Corporation), and anti-oxytocin polyclonal antibody (Chemicon International; 7
50ng/ml). The concentration of anti-OXT that would effectively bind all of the OXT in 8
the culture medium was calculated based on a preliminary experiment performed to 9
determine OXT concentrations secreted by luteal cells over the 72-hour culture. The 10
assay for OXT was kindly performed in Dr. Joanne Fortune’s laboratory, and the average 11
concentration of OXT in luteal cell-conditioned medium was 1.75ng/ml. Cocultures of 12
LC and TC were also carried out in the presence or absence of 50µg/ml of 13
aminoglutethimide (AG; Sigma-Aldrich), an inhibitor of the cytochrome P450 side-chain 14
cleavage enzyme. This concentration of AG has been previously shown to inhibit 15
endogenous P4 synthesis by cultured bovine luteal cells [32]. Additionally, exogenous P4 16
was added to the coculture in the presence of AG. All TC-LC cocultures were performed 17
in a humidified atmosphere of 5% CO2 in air at 37°C. Total RNA was extracted from the 18
TC after 72 hours of coculture to quantify OXTR mRNA by RT-qPCR. In a separate 19
experiment, T cells were isolated from luteal tissues following the same procedure 20
described above. These T cells referred to as resident T cells were collected for RNA 21
extraction and for detection of OXTR mRNA. 22
23

9
3. Total RNA Extraction and RT-qPCR 1
Total RNA was extracted from freshly purified PBMC and TC as well as from 2
cultured cells using TRIzol reagent (Invitrogen, Life Technologies). The concentrations 3
of total RNA were determined by measurement of optical density at 260nm in the 4
UV/Vis spectrophotometer (Beckmann). Total RNA was treated with RNase-free DNase 5
I (Roche Molecular Biochemicals) to eliminate genomic DNA contamination. RT-qPCR 6
was used to detect OXTR mRNA in total RNA from fresh PBMC, fresh TC, cultured TC 7
treated with steroids (P4, E2), and TC cocultured with LC followed by treatment with P4, 8
oxytocin, and anti-oxytocin. Oligonucleotide primers specific for OXTR transcript 9
(forward: 5’-ATCCGCACGGTCAAGATG-3’; reverse: 5’-10
AGAGGAAGCGCTGCACAA-3’) were designed to amplify a 226 base pair (bp) OXTR 11
cDNA fragment. These primers were designed from the bovine OXTR mRNA sequence 12
[14] (GenBank accession number: AF101724). Total RNA from bovine uterus was used 13
as positive control in the present study. Glyceraldehyde-3-phosphate dehydrogenase 14
(GAPDH) cDNA fragment was amplified as a constitutively expressed gene with the 15
following primers (forward: 5’-AAGATTGTCAGCAATGCC-3’; reverse: 5’-16
ACAGACACGTTGGGAG-3’). Total RNA (2µg) was reverse transcribed using the 17
iScript cDNA Synthesis Kit (Bio-Rad Laboratories) according to the protocol of the 18
manufacturer. Following the reverse transcription reaction, quantitative PCR was 19
performed on the MJ Research Opticon 2 (Bio-Rad Laboratories) using the iQSYBR 20
Green Supermix (Bio-Rad Laboratories) for a total of 35 cycles under the following 21
conditions: denaturing, 94°C for 30 seconds; annealing, 56°C for 45 seconds; and 22
extension, 72°C for 60 seconds, followed by an extra elongation of 5 minutes at 72°C. 23

10
Homologous standard curve prepared from purified OXTR cDNA PCR product was used 1
to calculate the steady-state concentration of OXTR mRNA in triplicate wells for each 2
sample. The PCR amplification products were electrophoretically separated on 1.5% 3
agarose gels and visualized with ethidium bromide. For initial validation, the specific 4
band corresponding to the size of the expected OXTR cDNA fragment was cut and 5
purified using the QIAquick Gel Extraction Kit (Qiagen Sciences) for sequence 6
confirmation. A control sample that was not reverse transcribed was used to confirm that 7
the product obtained was not amplified from genomic DNA. 8
9
4. Protein Extraction and Immunoblotting 10
Fresh PBMC were isolated on D3, D11, and D19 of the estrous cycle from four 11
different cows at each time. The proteins were extracted from lymphocytes using the 12
CelLytic MT Cell Lysis Reagent (Sigma-Aldrich Biotechnology) in the presence of the 13
protease inhibitor cocktail (Sigma-Aldrich Biotechnology) following the manufacturer’s 14
protocol and quantified according to the method of Bradford [33] (Bio-Rad Protein 15
Assay, Bio-Rad Laboratories). Protein samples (100µg) were subjected to electrophoresis 16
on a 12% SDS-polyacrylamide gel and the separated proteins were blotted onto 17
polyvinylidene difluoride membranes (PVDF; Hybond-P, Amersham Pharmacia 18
Biotech). Western analyses were performed as previously described [34] using a 19
polyclonal rabbit anti-OXTR antibody (Sigma-Aldrich; catalog #: O4389), raised against 20
a synthetic peptide corresponding to the N-terminal extracellular domain of the oxytocin 21
receptor. First, membranes were incubated with blocking buffer (20mM Tris pH 7.4, 22
150mM NaCl, 5% non-fat dry milk, and 0.05% Tween-20) for 2 hours at room 23

11
temperature followed by an overnight (12-16 hours) incubation at 4°C with anti-OXTR 1
antibody at a final concentration of 0.5µg/ml. Membranes were washed twice with TBS-2
Tween (20mM Tris pH 7.4, 150mM NaCl, 0.05% Tween-20), and incubated with the 3
horseradish peroxidase-labeled anti-rabbit secondary antibody (Amersham Biosciences) 4
at a dilution of 1:20,000. The antigen-antibody complex was visualized using the 5
enhanced chemiluminescence system (ECL Western Blotting Analysis System; 6
Amersham Biosciences) following the manufacturer’s protocol. Membranes were 7
exposed to Kodak Biomax light films (Kodak) and the films were developed in the SRX-8
101A Konica film processor (Konica Corporation, Japan). Membranes were also 9
incubated with anti-oxytocin to determine if bovine lymphocytes express oxytocin along 10
with its receptor. Beta actin was used as an internal control to verify the integrity of 11
proteins in the samples. 12
13
5. Intracellular Calcium Measurements 14
To test OXT actions on intracellular free Ca2+ concentrations ([Ca2+]i), T 15
lymphocytes were isolated as described above, washed with serum-free RPMI-1640 and 16
loaded with 1µM of the fluorescent Ca2+ indicator, Fura-2 AM (Molecular Probes, 17
Eugene, OR) essentially according to the manufacturer’s protocol and as described by 18
Braileanu et al. [35]. Briefly, cells were incubated with Fura-2 at 37°C for 40 minutes in 19
serum-free culture medium. The cells were washed for 30 minutes in calcium-free buffer 20
(140mM NaCl, 5mM KCl, 2mM MgCl2, 10mM Hepes, and 6mM glucose, pH 7.4) to 21
remove unincorporated Fura-2 prior to [Ca2+]i measurements. Fura-2-loaded cells were 22
treated with OXT (100ng/ml), and the measurements were performed using the BioTek 23

12
FL600F Fluorescence Plate Reader (Bio-Tek Instruments, Inc.). [Ca2+]i were determined 1
by the fluorescence measurements at 340 and 360 nm. The specificity of the response 2
was evaluated by the treatment of cells with arginine vasopressin (AVP; Sigma-Aldrich). 3
Results were obtained from separate experiments using T cells from four different 4
animals. Each experiment was performed in triplicate, and the concentration in each 5
replicate represents the average of 10 individual readings. Values were corrected for 6
background fluorescence. 7
8
6. Statistical Analysis 9
Gene expression data were log-transformed and analyzed using covariate analysis within 10
the mixed model of SAS (SAS Inst. Inc., Cary, NC) with GAPDH as the covariate. Data 11
were presented as least-square means ± SEM and differences were considered significant 12
at p < 0.05. For calcium measurements, three-way analysis of variance was performed to 13
determine whether differences existed among the different treatments. The Student-14
Newman-Keuls procedure was used to determine differences among specific means. 15
Differences were considered significant at p < 0.05. 16
17
18

13
RESULTS 1
1. OXTR mRNA expression in bovine lymphocytes 2
Real-time PCR was initially conducted to determine if OXTR mRNA are expressed 3
in bovine lymphocytes. Experiments were performed on freshly collected PBMC, freshly 4
purified T and B cells, as well as cultured T cells. After a maximal number of 5
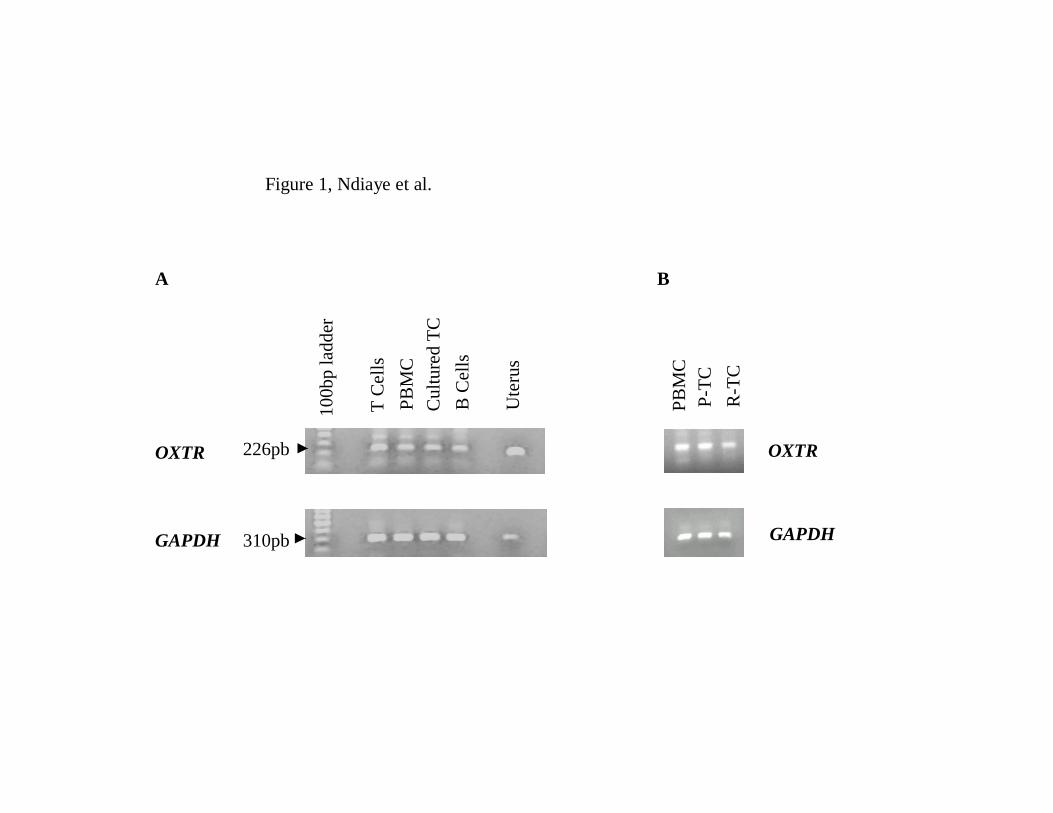
amplification cycles OXTR cDNA was detected in all samples analyzed with a specific 6
226 base pair amplicon (Figure 1A). The same amplicon was observed in the uterine 7
sample used as a positive control. Sequencing results confirmed that the observed 8
amplicon was indeed OXTR transcript. This transcript was also detected in the resident T 9
cells isolated from luteal tissues (Figure 1B). Quantitative analyses showed that OXTR 10
mRNA was greater in T cells than in PBMC (p < 0.05; figure 2A) or purified B 11
cells/macrophages (p < 0.05; Figure 2B) indicating that T cells are the main lymphocyte 12
population that expresses OXTR mRNA. In addition, OXTR mRNA concentrations were 13
greater in freshly isolated TC compared with cultured TC (p < 0.05; Figure 2C). The 14
expression of OXTR mRNA was also observed in peripheral blood T cells collected at 15
three different stages of the estrous cycle. There was considerable variation among 16
animals in expression of OXTR mRNA, such that there was no significant difference in 17
OXTR mRNA concentrations among the early (D3), mid (D11), and late (D19) stages of 18
the estrous cycle (Figure 3). However, OXTR mRNA expression appeared numerically 19
lower in cells collected on D19 of the estrous cycle compared to D3 (p = 0.091). 20
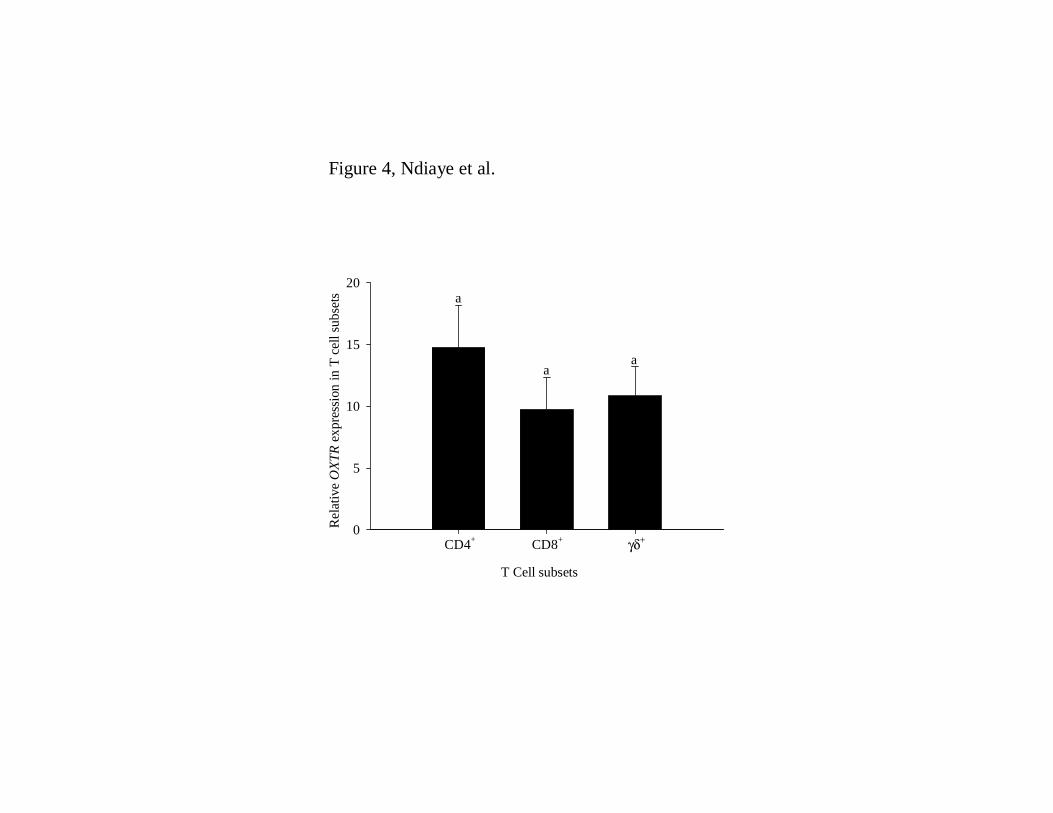
Additionally, OXTR transcripts were found in each of the major T cell subsets including 21
CD4+, CD8+, and γδ+ T cells with no significant differences in the steady-state 22
concentrations of OXTR mRNA among T cell subsets (Figure 4). 23

14
1
2. Western blot analyses 2
To investigate whether OXTR protein expression paralleled that of the mRNA, 3
western blot analyses were performed on lymphocyte protein samples from D3, D11, and 4
D19 of the estrous cycle. A specific band of the expected molecular weight of about 45 5
kDa corresponding to the bovine OXTR protein was observed in all protein extracts from 6
lymphocytes collected at different times of the estrous cycle (Figure 5; panel A). Proteins 7
extracted from uterus were used as positive control and a single protein was recognized 8
by the anti-OXTR antibody. Interestingly, one more band at about 31 kDa appeared in all 9
lymphocyte samples (Figure 5; panel A). Both the 45 kDa and 31 kDa proteins appeared 10
similar for the cells collected at each sampling time. Western blot analyses performed on 11
the same samples also showed presence of oxytocin in the lymphocytes at a molecular 12
weight of about 14 kDa, consistent with the size of OXT-neurophysin 1, the precursor 13
protein to OXT (Figure 5; panel B). The presence of this precursor indicates production 14
of oxytocin by bovine lymphocytes. The same size protein was observed in a pituitary 15
sample used as a positive control for OXT. Beta actin protein used as an internal control 16
was recognized in all samples by the monoclonal anti-beta actin antibody and no changes 17
were observed throughout the estrous cycle. 18
19
3. OXTR mRNA regulation by steroids 20
In order to better understand the regulation of OXTR mRNA expression in T cells, 21
the effects of progesterone and estradiol were analyzed. T cells were cultured in the 22
presence or absence of Con A and treated with P4 and E2, separately, and in combination. 23

15
E2 significantly increased OXTR mRNA expression (p < 0.05; Figure 6, left panel). The 1
effect of P4 only on the steady-state concentrations of OXTR mRNA was not significant 2
compared with the control cells (p = 0.092; Figure 6, left panel), but progesterone 3
completely inhibited E2-stimulated OXTR mRNA expression. Surprisingly, the 4
stimulatory effect of E2 on OXTR mRNA expression by T cells was completely abrogated 5
when the cells were cultured in the presence of Con A (Figure 6, right panel). In fact, 6
expression of OXTR mRNA in Con A-stimulated T cells treated with E2 was lower than 7
in control cells (p < 0.05; figure 6, right panel), and the steady-state concentrations of 8
OXTR mRNA in P4-treated cells was intermediate between controls and E2-treated cells 9
(p = 0.077). In Con A-treated cells, OXTR mRNA expression was not different in P4 + 10
E2-treated cells compared to either steroid administered alone (Figure 6, right panel). 11
12
4. TC-LC coculture 13
To further analyze P4 effects on OXTR mRNA expression by T cells, coculture 14
experiments were conducted using T cells and luteal cells from D11 CL. T cells that were 15
cultured in the presence of luteal cells tended (p = 0.0516) to have lower amounts of 16
OXTR mRNA compared to TC cultured alone, presumably due to the progesterone 17
produced by the cultured luteal cells (Figure 7). Treatment of the TC-LC coculture with 18
aminogluthetimide (AG), which inhibits the production of endogenous P4, caused a 19
significant increase in the steady-state concentrations of OXTR mRNA compared to the 20
TC-LC with no AG (p < 0.05; Figure 7). Addition of exogenous P4 to the coculture 21
reversed the effect causing a decrease in OXTR mRNA expression by T cells as compared 22

16
to AG-treated cells (p < 0.05). Neither OXT nor anti-OXT affected OXTR mRNA 1
expression compared to the TC-LC control. 2
3
5. Calcium measurements 4
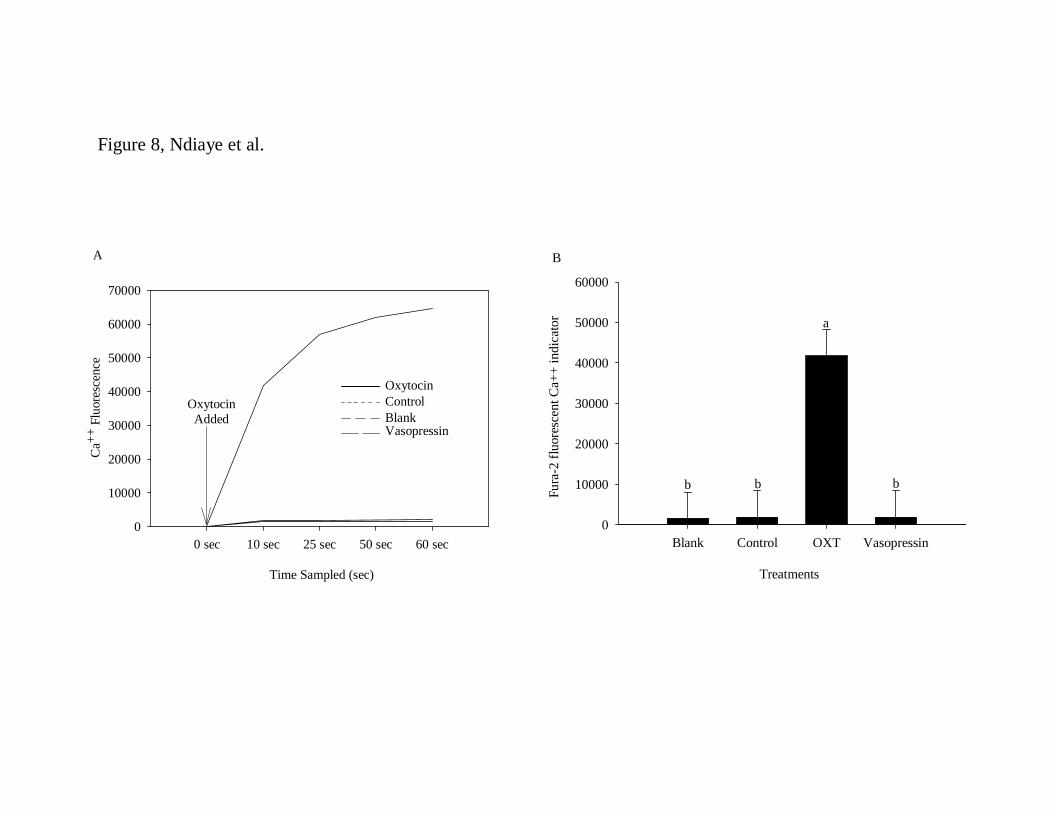
To determine if the OXTR protein found in the T cells is biologically active, the 5
intracellular concentrations of calcium were measured using the fura-2 calcium indicator. 6
When T cells were treated with OXT, intracellular concentrations of calcium ([Ca2+]i) 7
were rapidly and dramatically increased as compared with non-treated cells (p < 0.05; 8
Figure 8, panels A and B). The specificity of the Ca2+ response to OXT was demonstrated 9
by lack of increase in ([Ca2+]i) after treatment with vasopressin (Figure 8). 10
11

17
DISCUSSION 1
In the present study, OXT receptor mRNA and protein expression was demonstrated 2
in bovine peripheral blood mononuclear cells and T lymphocytes. The expression of 3
OXTR mRNA in PBMC was lower than in purified T cells indicating that T cells are the 4
main white blood cell components that express OXTR mRNA, thus purification of T cells 5
from macrophages and B lymphocytes enriched the OXTR message relative to GAPDH. 6
This was confirmed by the observation that the negative fraction of PBMC, containing 7
macrophages and B lymphocytes, had very little expression of OXTR mRNA compared 8
with specifically purified T lymphocytes. Cultured T cells exhibited lower expression of 9
OXTR mRNA compared with freshly purified T cells, perhaps because the culture 10
medium did not contain factors that stimulate or sustain OXTR mRNA expression in T 11
lymphocytes. It is also possible that OXTR mRNA is downregulated during the isolation 12
procedure, and more than 72 hours in culture is required for T cells to resume expression 13
of OXTR mRNA in amounts comparable to the fresh cells. Alternatively, autocrine 14
factors that downregulate OXTR mRNA may be produced during the culture. All the 15
subsets of T cells (CD4+, CD8+, and γδ+) expressed OXTR mRNA suggesting an 16
important role for the OXT/OXTR system in the response of these immune cells. 17
OXTR transcripts were quantified by RT-qPCR in bovine lymphocytes and appeared 18
differentially regulated by estradiol and progesterone in this in vitro model. Several 19
studies have shown that OXTR mRNA from uterus and mammary gland tissues is 20
upregulated by E2 and downregulated by P4 in vivo and in vitro [36-40]. In the rat uterus, 21
OXTR mRNA was shown to be upregulated when concentrations of circulating estrogen 22
are high and P4 concentrations are low [40, 41]. Similarly, in ewes, E2 treatment induced 23

18
an increase in endometrial OXTR mRNA expression [42-44], but treatment with E2 after 1
several days of continuous P4 treatment decreased OXTR mRNA expression [45, 46]. 2
This observation may suggest that OXTR mRNA stimulation by E2 requires the absence 3
or the decline of P4 in the cellular environment and/or an adequate balance of E2/P4 rather 4
than only an increase in E2 concentrations. In the present experiment, E2 significantly 5
increased OXTR mRNA expression in T cells cultured for 72 hours. This stimulatory 6
effect of E2 on OXTR mRNA lymphocytes agrees with previous studies examining OXTR 7
mRNA in reproductive tissues. This result suggests that E2 may have an effect on 8
immune cell functions by regulating OXTR mRNA expression from the T cells. However, 9
the effect of E2 was completely abrogated when the T cells were stimulated with 10
concanavalin A (Con A). It is not clear if this surprising observation is the result of a 11
direct inhibition of E2 action or its receptor by Con A, or if stimulated T cells produce 12
autocrine/paracrine factors that antagonize the response to E2. 13
The steroid hormone P4 was also shown to regulate OXTR mRNA expression. One of 14
the functions of P4 is the maintenance of uterine quiescence by decreasing uterine 15
sensitivity to oxytocin. Previous studies showed that P4 had a negative effect on OXTR 16
mRNA expression in endometrial epithelial cells [47-49]. In the endometrial cells, the 17
OXTR gene was spontaneously upregulated when the cells were explanted away from the 18
influence of circulating P4 [49]. Another in vitro study showed a decrease in OXTR 19
mRNA numbers when bovine endometrial epithelial cells were exposed to P4 and treated 20
with E2, suggesting that the combination of E2 with P4 leads to a low responsiveness of 21
the endometrium to OXT by decreasing OXTR mRNA [48]. In bovine peripheral 22
lymphocytes, OXTR mRNA expression was highest on day 3 of the estrous cycle, began 23

19
to decline by day 11, and was further decreased by day 19. The decrease in lymphocyte 1
OXTR mRNA as the estrous cycle progressed is likely due to elevated P4 at midcycle 2
compared to day 3. Estradiol concentrations would be expected to increase around day 19 3
of the estrous cycle, but progesterone concentrations are still elevated at that time. 4
Although E2 stimulated OXTR mRNA expression in T cells in vitro, its action was 5
completely inhibited by simultaneous treatment with progesterone. Therefore, the 6
temporal pattern of expression of OXTR mRNA in circulating lymphocytes is consistent 7
with the observed steroid hormone effects on cultured lymphocyte OXTR mRNA 8
expression. The decrease of the steady-state concentration of OXTR mRNA by the 9
combination of P4 and E2 might suggest regulation of T cell responsiveness to OXT by 10
these two steroids. This implies that resident T cells might be relatively unresponsive to 11
OXT when P4 synthesis is maximal, although the effect of P4 was primarily to antagonize 12
E2 stimulation of OXTR mRNA. Since the bovine CL does not contain appreciable 13
amounts of E2, the local effect of progesterone on lymphocyte OXTR mRNA may be less 14
pronounced. Further, during times of tissue remodeling, such as during luteinization and 15
luteolysis, P4 concentrations are lower, perhaps allowing for increased expression of 16
OXTR mRNA. The importance of hormonal regulation of OXTR mRNA to the function 17
of peripheral blood and resident T cells remains to be determined. 18
To determine if P4 synthesis by luteal cells could affect OXTR mRNA expression by 19
T lymphocytes, T cells were cocultured with luteal cells. In the presence of AG, an 20
inhibitor of endogenous P4 production by luteal cells, OXTR mRNA expression was 21
greater compared to its expression from cells cultured in the absence of AG. When 22
exogenous P4 was added to the cocultures in the presence of AG, a decrease in OXTR 23

20
mRNA expression was observed. These data suggest that P4 produced by luteal cells may 1
downregulate T lymphocyte expression of OXTR mRNA in this coculture system. In 2
addition to P4, it is also possible that other factors, alone or in synergy with P4, may affect 3
TC expression of OXTR mRNA. No effect of OXT or anti-OXT was observed on OXTR 4
mRNA expression. This may indicate that OXT does not regulate its own receptor, 5
although suppression of OXTR mRNA expression by luteal cell-produced progesterone in 6
those cultures may have masked any effects of OXT. 7
It is generally held that steroid hormones E2 and P4 act at a genomic level by binding 8
to nuclear receptors and modulating the expression of specific target genes. While there is 9
no E2 response element on the bovine or ovine OXTR gene promoter regions [50], it has 10
been reported that estrogen receptor can act through other sites on gene promoters [51]. 11
Despite the well-described effects of P4, the molecular mechanisms that mediate its 12
actions on lymphocytes are still mostly unknown. Although P4 appears to be responsible 13
for the downregulation of OXTR mRNA expression, this effect is most likely an indirect 14
nongenomic effect since there is no P4 response element reported on the promoter region 15
of the OXTR gene [50, 52], and bovine T lymphocytes do not express nuclear P4 16
receptors [30]. Several reports suggest that P4 may act at the cell membrane through 17
specific receptors [53-55] supporting the notion that P4 may act through membrane 18
receptors to regulate OXTR mRNA expression by lymphocytes. 19
The OXTR protein was observed in lymphocyte samples at all stages of the estrous 20
cycle. A specific band was recognized both in the lymphocytes and the uterus samples. 21
The deduced amino acids from bovine OXTR protein [14] predicted a putative molecular 22
weight of 43.3 kDa which is similar to the size observed in the present study at about 45 23

21
kDa. An additional protein reacting with the anti-OXTR antibody was observed at about 1
31 kDa in lymphocytes but not in the uterus. It is not clear if the two bands observed in 2
the lymphocytes represent two different, unrelated receptors from different splicing 3
events of the OXTR gene, or if different post-translational modifications occur as 4
suggested by Kojro et al. [56]. Interestingly, in the pregnant rat uterus, a single OXTR 5
band at 66 kDa was observed while in the non-pregnant rat a second band of 30 kDa was 6
also described [39]. The nature of the 30 kDa form was not identified but could indicate 7
possible modifications of the OXTR protein dependent on the physiological status of the 8
tissue or the animal. The larger molecular weight form of OXTR protein is likely the 9
functional protein in correlation with what has been previously reported in the rabbit 10
myometrium and amnion [57]. 11
Oxytocin treatment induced an increase in [(Ca2+)]i in T cells suggesting a functional 12
response mediated by OXTR protein. Functional OXTR protein in T lymphocytes makes 13
these immune cells potential targets for OXT. In addition, the present study also showed 14
that bovine lymphocytes express OXT as demonstrated by western blot analysis. The 15
presence of OXT and its receptor in the lymphocytes implies that OXT can have not only 16
paracrine actions but also autocrine actions that could regulate the function of the 17
lymphocytes. In summary, the present data provide evidence that OXTR mRNA 18
expression in bovine lymphocytes is regulated in a steroid-dependent manner. Moreover, 19
OXT elicited a functional [Ca2+]i response in T lymphocytes supporting the hypothesis 20
that OXT may act as a paracrine and/or autocrine regulator of resident immune cell 21
functions within the CL throughout the estrous cycle. It is possible that, in addition to its 22
well-known functions on reproductive tissues, OXT may exert an alternate role by 23

22
regulating and maintaining a balance of anti-inflammatory and pro-inflammatory 1
cytokines within the CL. Further studies will determine the nature of the T cell response 2
to oxytocin. 3
4

23
ACKNOWLEDGMENTS 1
The authors wish to thank Mr. Justin Fear for assistance with sample collection and T cell 2
isolation, Dr Joanne Fortune for the assay of oxytocin and Dr William Weiss for 3
assistance with the statistical analyses. 4
5
6
7
8
9
10
11
12
13
14
15
16

24
REFERENCES 1
2
1. Fuchs AR, Fuchs F, Husslein P, Soloff MS. Oxytocin receptors in the human uterus 3 during pregnancy and parturition. Am J Obstet Gynecol 1984;150(6):734-741. 4
2. Schams D. Regulation of bovine intra-luteal function by peptide hormones. J Physiol 5 Pharmacol 1992; 43(4 Suppl 1):117-129. 6
3. Wathes DC. Possible actions of gonadal oxytocin and vasopressin. J Reprod Fertil 7 1984; 71(1):315-345. 8
4. Flint AP, Lamming GE, Stewart HJ, Abayasekara DR. The role of the endometrial 9 oxytocin receptor in determining the length of the sterile oestrous cycle and ensuring 10 maintenance of luteal function in early pregnancy in ruminants. Philos Trans R Soc 11 Lond B Biol Sci 1994; 344(1309):291-304. 12
5. McCracken JA, Custer EE, Lamsa JC. Luteolysis: A neuroendocrine-mediated event. 13 Physiol Rev 1999; 79(2):263-323. 14
6. Skarzynski DJ, Bogacki M, Kotwica J. Changes in ovarian oxytocin secretion as an 15 indicator of corpus luteum response to prostaglandin F(2alpha) treatment in cattle. 16 Theriogenology 1997; 48(5):733-742. 17
7. Wathes DC, Lamming GE. The oxytocin receptor, luteolysis and the maintenance of 18 pregnancy. J Reprod Fertil Suppl; 49:53-67. 19
8. Taylor AH, Ang VT, Jenkins JS, Silverlight JJ, Coombes RC, Luqmani YA. Interaction 20 of vasopressin and oxytocin with human breast carcinoma cells. Cancer Res 1990; 21 50(24):7882-7886. 22
9. Cassoni P, Fulcheri E, Carcangiu ML, Stella A, Deaglio S, Bussolati G. Oxytocin 23 receptors in human adenocarcinomas of the endometrium: Presence and biological 24 significance. J Pathol 2000; 190(4):470-477. 25
10. Thibonnier M, Berti-Mattera LN, Dulin N, Conarty DM, Mattera R. Signal transduction 26 pathways of the human V1-vascular, V2-renal, V3-pituitary vasopressin and oxytocin 27 receptors. Prog Brain Res 1998; 119:147-161. 28
11. Kendrick KM. Oxytocin, motherhood and bonding. Exp Physiol 2000; 85 Spec 29 No:111S-124S. 30
12. Kimura T, Takemura M, Nomura S, Nobunaga T, Kubota Y, Inoue T, Hashimoto K, 31 Kumazawa I, Ito Y, Ohashi K, Koyama M, Azuma C, Kitamura Y, Saji F. Expression 32 of oxytocin receptor in human pregnant myometrium. Endocrinology 1996; 137(2):780-33 785. 34

25
13. Wathes DC, Borwick SC, Timmons PM, Leung ST, Thornton S. Oxytocin receptor 1 expression in human term and preterm gestational tissues prior to and following the 2 onset of labour. J Endocrinol 1999; 161(1):143-151. 3
14. Bathgate R, Rust W, Balvers M, Hartung S, Morley S, Ivell R. Structure and expression 4 of the bovine oxytocin receptor gene. DNA Cell Biol 1995; 14(12):1037-1048. 5
15. Kimura T, Tanizawa O, Mori K, Brownstein MJ, Okayama H. Structure and expression 6 of a human oxytocin receptor. Nature 1992; 356(6369):526-529. 7
16. Gimpl G, Fahrenholz F. The oxytocin receptor system: Structure, function, and 8 regulation. Physiol Rev 2001; 81(2):629-683. 9
17. Copland JA, Jeng YJ, Strakova Z, Ives KL, Hellmich MR, Soloff MS. Demonstration of 10 functional oxytocin receptors in human breast Hs578T cells and their up-regulation 11 through a protein kinase C-dependent pathway. Endocrinology 1999; 140(5):2258-12 2267. 13
18. Jankowski M, Wang D, Hajjar F, Mukaddam-Daher S, McCann SM, Gutkowska J. 14 Oxytocin and its receptors are synthesized in the rat vasculature. Proc Natl Acad Sci U 15 S A 2000; 97(11):6207-6211. 16
19. Hansenne I, Rasier G, Pequeux C, Brilot F, Renard C, Breton C, Greimers R, Legros JJ, 17 Geenen V, Martens HJ. Ontogenesis and functional aspects of oxytocin and vasopressin 18 gene expression in the thymus network. J Neuroimmunol 2005; 158(1-2):67-75. 19
20. Yamaguchi Y, Yamada K, Suzuki T, Wu YP, Kita K, Takahashi S, Ichinose M, Suzuki 20 N. Induction of uPA release in human peripheral blood lymphocytes by [deamino-21 cysl,D-Arg8]-vasopressin (dDAVP). Am J Physiol Endocrinol Metab 2004; 287(5): 22 E970-976. 23
21. Pate JL. Involvement of immune cells in regulation of ovarian function. J Reprod Fertil 24 Suppl 1995; 49:365-377. 25
22. Pate JL, Landis Keyes P. Immune cells in the corpus luteum: Friends or foes? 26 Reproduction 2001; 122(5):665-676. 27
23. Townson DH, Pate JL. Regulation of prostaglandin synthesis by interleukin-1 beta in 28 cultured bovine luteal cells. Biol Reprod 1994; 51(3):480-485. 29
24. Penny LA, Armstrong D, Bramley TA, Webb R, Collins RA, Watson ED. Immune cells 30 and cytokine production in the bovine corpus luteum throughout the oestrous cycle and 31 after induced luteolysis. J Reprod Fertil 1999; 115(1):87-96. 32
25. Townson DH, Tsang PC, Butler WR, Frajblat M, Griel LC,Jr, Johnson CJ, Milvae RA, 33 Niksic GM, Pate JL. Relationship of fertility to ovarian follicular waves before breeding 34 in dairy cows. J Anim Sci 2002; 80(4):1053-1058. 35 36

26
26. Petroff M, Coggeshall KM, Jones LS, Pate JL. Bovine luteal cells elicit major 1 histocompatibility complex class II-dependent T-cell proliferation. Biol Reprod 1997; 2 57(4):887-893. 3
27. Sernia C, Gemmell RT, Thomas WG. Oxytocin receptors in the ovine corpus luteum. J 4 Endocrinol 1989; 121(1):117-123. 5
28. Pitzel L, Jarry H, Wuttke W. Demonstration of oxytocin receptors in porcine corpora 6 lutea: Effects of the cycle stage and the distribution on small and large luteal cells. Biol 7 Reprod 1993; 48(3):640-646. 8
29. Okuda K, Miyamoto A, Sauerwein H, Schweigert FJ, Schams D. Evidence for oxytocin 9 receptors in cultured bovine luteal cells. Biol Reprod 1992; 46(6):1001-1006. 10
30. Cannon MJ, Petroff MG, Pate JL. Effects of prostaglandin F2alpha and progesterone on 11 the ability of bovine luteal cells to stimulate T lymphocyte proliferation. Biol Reprod 12 2003; 69(2):695-700. 13
31. Pate JL. Isolation and culture of fully differenciated bovine luteal cells. Methods in 14 Toxicology 1993; 3B:358-370. 15
32. Grusenmeyer DP, Pate JL. Localization of prostaglandin F2 alpha inhibition of 16 lipoprotein use by bovine luteal cells. J Reprod Fertil 1992; 94(2):311-318. 17
33. Bradford MM. A rapid and sensitive method for the quantitation of microgram 18 quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976; 19 72:248-254. 20
34. Bedard J, Brule S, Price CA, Silversides DW, Lussier JG. Serine protease inhibitor-E2 21 (SERPINE2) is differentially expressed in granulosa cells of dominant follicle in cattle. 22 Mol Reprod Dev 2003; 64(2):152-165. 23
35. Braileanu GT, Simasko SM, Uzumcu M, Mirando MA. Intracellular free calcium in 24 response to oxytocin in pig endometrial cells. Mol Cell Endocrinol 1999; 155(1-2):77-25 83. 26
36. Amico JA, Rauk PN, Cai HM. Estradiol and progesterone regulate oxytocin receptor 27 binding and expression in human breast cancer cell lines. Endocrine 2002; 18(1):79-84. 28
37. Maggi M, Magini A, Fiscella A, Giannini S, Fantoni G, Toffoletti F, Massi G, Serio M. 29 Sex steroid modulation of neurohypophysial hormone receptors in human nonpregnant 30 myometrium. J Clin Endocrinol Metab 1992; 74(2):385-392. 31
38. Soloff MS, Fernstrom MA, Periyasamy S, Soloff S, Baldwin S, Wieder M. Regulation 32 of oxytocin receptor concentration in rat uterine explants by estrogen and progesterone. 33 Can J Biochem Cell Biol 1983; 61(7):625-630. 34 35

27
39. Umscheid CA, Wu WX, Gordan P, Nathanielsz PW. Up-regulation of oxytocin receptor 1 messenger ribonucleic acid and protein by estradiol in the cervix of ovariectomized rat. 2 Biol Reprod 1998; 59(5):1131-1138. 3
40. Zingg HH, Rozen F, Breton C, Larcher A, Neculcea J, Chu K, Russo C, Arslan A. 4 Gonadal steroid regulation of oxytocin and oxytocin receptor gene expression. Adv Exp 5 Med Biol; 395:395-404. 6
41. Larcher A, Neculcea J, Breton C, Arslan A, Rozen F, Russo C, Zingg HH. Oxytocin 7 receptor gene expression in the rat uterus during pregnancy and the estrous cycle and in 8 response to gonadal steroid treatment. Endocrinology 1995; 136(12):5350-5356. 9
42. Beard AP, Lamming GE. Oestradiol concentration and the development of the uterine 10 oxytocin receptor and oxytocin-induced PGF2 alpha release in ewes. J Reprod Fertil 11 1994; 100(2):469-475. 12
43. Spencer TE, Ing NH, Ott TL, Mayes JS, Becker WC, Watson GH, Mirando MA, Brazer 13 FW. Intrauterine injection of ovine interferon-tau alters oestrogen receptor and oxytocin 14 receptor expression in the endometrium of cyclic ewes. J Mol Endocrinol 1995; 15 15(2):203-220. 16
44. Vallet JL, Lamming GE, Batten M. Control of endometrial oxytocin receptor and 17 uterine response to oxytocin by progesterone and oestradiol in the ewe. J Reprod Fertil 18 1990; 90(2):625-634. 19
45. Wathes DC, Mann GE, Payne JH, Riley PR, Stevenson KR, Lamming GE. Regulation 20 of oxytocin, oestradiol and progesterone receptor concentrations in different uterine 21 regions by oestradiol, progesterone and oxytocin in ovariectomized ewes. J Endocrinol 22 1996; 151(3):375-393. 23
46. Zhang J, Weston PG, Hixon JE. Role of progesterone and oestradiol in the regulation of 24 uterine oxytocin receptors in ewes. J Reprod Fertil 1992; 94(2):395-404. 25
47. Horn S, Bathgate R, Lioutas C, Bracken K, Ivell R. Bovine endometrial epithelial cells 26 as a model system to study oxytocin receptor regulation. Hum Reprod Update 1998; 27 4(5):605-614. 28
48. Kombe A, Sirois J, Goff AK. Prolonged progesterone treatment of endometrial 29 epithelial cells modifies the effect of estradiol on their sensitivity to oxytocin. Steroids 30 2003; 68(7-8):651-658. 31
49. Sheldrick EL, Flick-Smith HC, Dos Santos Cruz GJ. Oxytocin receptor binding activity 32 in cultured ovine endometrium. J Reprod Fertil 1993; 98(2):521-528. 33
50. Ivell R, Walther N. The role of sex steroids in the oxytocin hormone system. Mol Cell 34 Endocrinol 1999; 151(1-2):95-101. 35 36

28
51. Safe S. Transcriptional activation of genes by 17 beta-estradiol through estrogen 1 receptor-Sp1 interactions. Vitam Horm 2001; 62:231-252. 2
52. Rozen F, Russo C, Banville D, Zingg HH. Structure, characterization, and expression of 3 the rat oxytocin receptor gene. Proc Natl Acad Sci U S A 1995; 92(1):200-204. 4
53. Cai Z, Stocco C. Expression and regulation of progestin membrane receptors in the rat 5 corpus luteum. Endocrinology 2005; 146(12):5522-5532. 6
54. Thomas P, Pang Y, Zhu Y, Detweiler C, Doughty K. Multiple rapid progestin actions 7 and progestin membrane receptor subtypes in fish. Steroids 2004; 69(8-9):567-573. 8
55. Zhu Y, Bond J, Thomas P. Identification, classification, and partial characterization of 9 genes in humans and other vertebrates homologous to a fish membrane progestin 10 receptor. Proc Natl Acad Sci U S A 2003; 100(5):2237-2242. 11
56. Kojro E, Hackenberg M, Zsigo J, Fahrenholz F. Identification and enzymatic 12 deglycosylation of the myometrial oxytocin receptor using a radioiodinated 13 photoreactive antagonist. J Biol Chem 1991; 266(32):21416-21421. 14 15
57. Hinko A, Soloff MS, Potier M. Molecular size characterization of oxytocin receptors 16 in rabbit amnion. Endocrinology 1992; 130(6):3554-3559. 17
18
19
20
21
22
23
24
25

29
Figure legends 1
2
Figure 1. Detection of OXTR cDNA in bovine lymphocytes by PCR. A) Total RNA was 3
extracted from fresh peripheral blood mononuclear cells (PBMC), fresh T and B 4
lymphocytes, and cultured T cells (TC). B) Total RNA was extracted from PBMC, 5
peripheral blood T cells (P-TC) and resident T cells (R-TC; T cells isolated from within 6
luteal tissues). Uterus was used as a positive control sample. Specific amplification of the 7
OXTR cDNA was observed in all samples. The bands represent PCR product obtained 8
after 35 cycles. 9
10
Figure 2. Analysis of OXTR mRNA expression in bovine peripheral white blood cells. 11
OXTR mRNA expression in fresh T cells was compared to fresh PBMC (A; n = 6 cows), 12
B cells/macrophages (B; n = 4), and cultured T cells (C; n = 4). Data are presented as 13
least-square means ± SEM and different letters denote means that differed significantly (p 14
< 0.05). 15
16
Figure 3. OXTR mRNA expression throughout the estrous cycle. Expression of OXTR 17
mRNA was analyzed in freshly purified T cells from day 3, day 11, and day 19 of the 18
estrous cycle. Data are presented as least-square means ± SEM (n = 4 cows). 19
20
Figure 4. OXTR mRNA expression in different T cell subsets. Different T cell subsets 21
were isolated using specific antibodies and analyzed for OXTR mRNA expression by RT-22
qPCR. Data are presented as least-square means ± SEM (n = 6 cows). 23

30
1
Figure 5. Representative western blot analyses using antibodies against OXTR and OXT 2
for protein expression in bovine lymphocytes. OXTR (A) and OXT-associated 3
neurophysin 1 (B) proteins were observed in bovine lymphocytes throughout the estrous 4
cycle, with corresponding beta actin blots. The experiment was repeated using cells from 5
4 animals with similar results. 6
7
Figure 6: Regulation of OXTR mRNA expression in bovine T lymphocytes by 8
progesterone (P4; 10ng/ml), and estradiol (E2; 15pg/ml). Left panel: Cells cultured 9
without Con A. Right panel: Cells cultured with Con A. Bars represent relative 10
concentrations of OXTR mRNA (mean ± SEM). Different letters denote significant 11
differences within Con A-treated groups (p < 0.05; n = 4 cows). 12
13
Figure 7: Effects of P4, OXT, and anti-OXT on OXTR mRNA expression in TC-LC 14
cocultures. LC = luteal cells; AG = aminogluthetimide. Bars represent relative 15
concentrations of OXTR mRNA (mean ± SEM) and different letters denote significant 16
differences (p < 0.05; n = 3 cows). 17
18
Figure 8: Oxytocin-stimulated [Ca2+]i mobilization in Fura-2-loaded T lymphocytes. 19
Lymphocytes were stimulated with oxytocin (100ng/ml) or with vasopressin (100ng/ml). 20
A) Time course of Ca2+ mobilization after OXT treatment in a representative experiment. 21
B) Peak Ca2+ concentrations in response to OXT (mean ± SEM; n = 4 separate 22
experiments). Blank = Fura 2-nonloaded T cells; Control = Fura 2-loaded T cells without 23
treatments. 24
T C
ells
PB
MC
Cul
ture
d T
C
B C
ells
Ute
rus
100b
p la
dder
OXTR
GAPDH
226pb
310pb
Figure 1, Ndiaye et al.
A B
PB
MC
P-T
C
R-T
C
OXTR
GAPDH

A
T cells PBMC0
1
2
3
4
5
Cell types
Rel
ativ
e O
XT
R e
xpre
ssio
n in
lym
phoc
ytes
b
a
B
T cells B cells/Macro0
1
2
3
4
5a
b
Rel
ativ
e O
XT
R e
xpre
ssio
n in
lym
phoc
ytes
Cell types
C
Fresh T cells Cultured TC0
2
4
6
8
10
12
14
16
18
T Lymphocytes
Rel
ativ
e O
XTR
exp
ress
ion
in T
cel
ls
a
b
Figure 2, Ndiaye et al.

0
2
4
6
8
10
12
14
16
Estrous cycle days
Rel
ativ
e O
XTR
exp
ress
ion
in T
cel
ls a
a
a
3 11 19
Figure 3, Ndiaye et al.
0
5
10
15
20
T Cell subsets
Rel
ativ
e O
XT
R e
xpre
ssio
n in
T c
ell s
ubse
ts a
aa
CD4+ CD8+ γδ+
Figure 4, Ndiaye et al.

A) OXTR
D3 D11 D19 Uterus
45
kDa
31
Lymphocytes
B) OXT
D3 D11 D19 Pit
14
kDa
Lymphocytes
Figure 5, Ndiaye et al.
β actin β actin

Treatments
Rel
ativ
e O
XTR
exp
ress
ion
in T
cel
ls
0
2
4
6
8
10
12
b
a
bb z
y
y,z
y,z
p = 0.077p = 0.092
P4+E2P4 E2Control P4+E2P4 E2Control
- Con A + Con A
Figure 6, Ndiaye et al.

p=0.0516p=0.0516
0
1
2
3
4
a
b
b bb b
Control (TC)
TC-LC AG AG+P4 OXT a-OXT
Rel
ativ
e O
XT
R e
xpre
ssio
n in
T c
ells
TC+LC
Treatments
p = 0.0516
Figure 7, Ndiaye et al.
A
Time Sampled (sec)
0 sec 10 sec 25 sec 50 sec 60 sec0
10000
20000
30000
40000
50000
60000
70000
OxytocinAdded
Ca+
+ Fl
uore
scen
ce
Oxytocin
BlankVasopressin
Control
B
Blank Control OXT Vasopressin0
10000
20000
30000
40000
50000
60000
a
bb
Fura
-2 f
luor
esce
nt C
a++
indi
cato
r
Treatments
b
Figure 8, Ndiaye et al.