feronia receptor-like kinase regulates rho gtpase ... · feronia receptor-like kinase regulates rho...
TRANSCRIPT

FERONIA receptor-like kinase regulates RHO GTPasesignaling of root hair developmentQiaohong Duana, Daniel Kitaa,b, Chao Lia, Alice Y. Cheunga,b,c,1, and Hen-Ming Wua,b
aDepartment of Biochemistry and Molecular Biology, bMolecular Cell Biology Program, and cPlant Biology Graduate Program, University of Massachusetts,Amherst, MA 01003
Edited by June B. Nasrallah, Cornell University, Ithaca, NY, and approved August 27, 2010 (received for review April 20, 2010)
Plant RHO GTPases (RAC/ROPs) mediate multiple extracellularsignals ranging from hormone to stress and regulate diversecellular processes important for polarized cell growth, differentia-tion, development, reproduction, and responses to the environ-ment. They shuttle between the GDP-bound inactive state and theGTP-bound activated state and their activation is predominantlymediated by a family of guanine nucleotide exchange factors(GEFs) referred to as ROPGEFs. Using the Arabidopsis ROPGEF1 asbait, we identified members of a receptor-like kinase (RLK) familyas potential upstream regulators for RAC/ROP signaling. NADPHoxidase-derived reactive oxygen species (ROS) are emerging as im-portant regulators for growth and development and play a crucialrole in mediating RAC/ROP-regulated root hair development, a po-larized cell growth process. We therefore screened T-DNA insertionmutants in these RLKs for root hair defects and found that muta-tions in one of them, At3g51550 encoding the FERONIA (FER) re-ceptor-like kinase, induced severe root hair defects. We show thatthe fer phenotypes correlated with reduced levels of active RAC/ROPs and NADPH oxidase-dependent, auxin-regulated ROS accu-mulation in roots and root hairs and that up-regulating RAC/ROPsignaling in fer countered the mutant phenotypes. Taken together,these observations strongly support FER as an upstream regulatorfor the RAC/ROP-signaled pathway that controls ROS-mediatedroot hair development. Moreover, FER was pulled down by ROP2GTPase in a guanine nucleotide-regulated manner implying a dy-namic signaling complex involving FER, a ROPGEF, and a RAC/ROP.
RAC/ROP | reactive oxygen species | ROPGEF | signal transduction |surface regulator
RHO GTPases are a large family of related monomeric GTP-binding proteins that serve diverse signaling functions (1).
Plant RAC/ROPs form a unique clade of RHO-like GTPases andplay important roles in regulating cell growth and polarity estab-lishment, hormone- and stress-induced responses that underliegrowth, development, reproduction, and interactions with the en-vironment (2–4). Several RAC/ROP effectors have been identified(see Fig. S1A). These include a family of CRIB-domain proteins(RICs) and a coiled-coil domain protein (ICR1). RICs are knownto mediate intracellular [Ca2+] and actin dynamics in polarizedpollen tubes and coordinate actin and microtubule organization inleaf epidermal cells (2), whereas ICR1 regulates secretion andauxin distribution during early embryo development in Arabidopsis(5, 6). Interaction between RAC/ROPs and NADPH oxidaseunderlies activation of pathogen-elicited reactive oxygen species(ROS)-mediated defense response in rice (7). Upstream of RAC/ROPs, the hormones auxin and abscisic acid can activate and in-activate, respectively, these small GTPases, which in turn signalhormone-regulated responses (8–10). Given their involvement indiverse biological processes, RAC/ROPs may act as integrationpoints for crosstalk between multiple signaling pathways (2).Besides being mediators of stress-induced responses, increasing
evidence shows that ROS also serve important regulatory func-tions in growth and development (11–15). In plants, multipletransmembrane spanning NADPH oxidases, homologous to thecatalytic gp91phox subunit of the mammalian enzyme, have been
found to underlie several RAC/ROP-mediated, ROS-regulatedgrowth and stress-induced responses. In studies involving theArabidopsis ROOT HAIR DEFECTIVE2 (RHD2/RBOHC) geneencoding a NADPH oxidase and SUPERCENTIPEDE, whichencodes a RAC/ROP negative regulator, guanine nucleotide dis-sociation inhibitor (GDI) (see Fig. S1A), Dolan and colleaguesprovided compelling evidence for aRAC/ROP-mediatedNADPHoxidase-dependent pathway for ROS-regulated root hair de-velopment (16, 17). Specifically, they showed that NADPH oxi-dase-derived ROS is required for polarized root hair growth andregulated RAC/ROP activity controls production and spatiallyregulated accumulation of ROS at the tip of emerging and elon-gating root hairs. Similarly, up-regulating RAC/ROP activity byoverexpressing a constitutively active RAC/ROP in transgenicArabidopsis seedlings also induced ectopic accumulation ofNADPH oxidase-derived ROS and defective root hairs (18).Although relying on guanine nucleotide exchange for activation
is conserved for RAC/ROPs (Fig. S1A), their activation is largelydependent on a unique guanine exchange factor (GEF) familyreferred to as ROPGEFs (19–21). ROPGEFs have variableamino- and carboxyl-terminal domains and a conserved centrallylocated nucleotide exchange activity domain, which exclusivelyactivatesRAC/ROPs but notRHOGTPases fromother organisms.Little is known about signaling components upstream of ROP-GEFs. To date, two pollen-specific leucine-rich repeat receptor-like kinases (LRR RLKs) (22), the tomato LePRK2, and Arabi-dopsis AtPRK2a are the only surface receptors known to interactwithROPGEFs. These PRKs regulate RAC/ROP-mediated pollentube growth, another polarized cell type in plant (23–25).Of the hundreds of RLKs in Arabidopsis (26), several of the
17-membered Catharanthus roseus receptor-like kinase (RLK)-related family have been identified as regulators for variousgrowth-related processes (27). Loss of function in one of these,THESEUS1 (THE1), suppressed the short hypocotyl phenotypein a cellulose synthase mutant, leading to the suggestion that itacts as a sensor for cell wall defects to signal growth inhibition(28). Although single mutations in THE1 and the relatedHERCULES1 (HERK1) did not induce noticeable phenotype,the1 herk1 double mutant showed inhibited growth (29). Re-ducing the level of another related RLK, FERONIA (FER), alsoresulted in growth inhibition and induced altered hormoneresponses (29, 30). FER was identified as a regulator for femalefertility, mediating pollen tube rupture in the female gameto-phyte to discharge sperm for fertilization and preventing multi-ple tube entrance into an already fertilized ovule (31–33). The
Author contributions: Q.D., D.K., A.Y.C., and H.-M.W. designed research; Q.D., D.K., C.L.,and H.-M.W. performed research; Q.D., D.K., and H.-M.W. contributed new reagents/analytic tools; Q.D., D.K., C.L., A.Y.C., and H.-M.W. analyzed data; and Q.D., D.K.,A.Y.C., and H.-M.W. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
See Commentary on page 17461.1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1005366107/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1005366107 PNAS | October 12, 2010 | vol. 107 | no. 41 | 17821–17826
PLANTBIOLO
GY
SEECO
MMEN
TARY

pollen-specific ANXUR1 and ANXUR2 are needed for pollentube growth as double anx1 anx2 mutant pollen tubes ruptureprecociously and fail to reach the ovules for fertilization (34, 35).Together, these findings suggest that these RLKs play importantroles in controlling cell growth and integrity, mediating con-sequences that could be as diagonally opposed as growth andnongrowth. Here, we show that FER is a ROPGEF-interactingRLK and regulates RAC/ROP-signaled and ROS-mediated roothair development, revealing a surface regulator for a signalingpathway that regulates ROS-mediated root hair growth and alsobroadly impacts growth and development.
ResultsFER RLK Is a Broadly Expressed ROPGEF-Interacting RLK. UsingROPGEF1 as bait in a yeast two-hybrid screen, we identifiedmembers of theArabidopsis CrRLK family as potential interactingproteins for ROPGEFs (Fig. 1A and Fig. S1B andC). Focusing onone of these, At3g51550, we used bimolecular fluorescence com-plementation (BiFC) assays (36) in protoplasts to confirm that thisRLKalso interacted withROPGEF1 in plant cells (Fig. 1B). Giventhe importance of RAC/ROP signaling to polarized cell growth(15–18) and that root hairs are more accessible than pollen tubesfor observation, we initially screened T-DNA insertion mutants inthese RLKs for root hair defects (see SI Materials and Methods)and found that mutations in At3g51550, which turned out to beidentical to FER when its sequence was elucidated (33), uniquelyinduced severe root hair defect (see below). Hence we adopt theFER nomenclature and the family as FER related. We also in-cluded in our studies siréne (srn), whose mutation is allelic with fer,the other originally described mutations, and inducing similar fe-male gametophytic defects as fer (31–33).FER promoter (pFER)::GUS analysis in transgenic Arabidopsis
indicates that in addition to expression in the ovule (33), FER isbroadly expressed (Fig. 1C) as previously shown by transcriptanalyses (33, 37). pFER expressed FER–GFP was localized to
the plasma membrane of diverse cell types, including emergingand growing root hairs (Fig. 1D).
FER Is Essential for Root Hair Development. We identified two T-DNA–induced alleles, fer-4 and fer-5 (Fig. 2A and Fig. S2A). Theoriginally described fer and srn are both in the vicinity of fer-4(33) (Fig. 2A). RT-PCR analysis showed that fer-4, with its T-DNA insertion in the extracellular domain-coding region, isa null mutant, whereas fer-5 harbors truncated transcripts (Fig.2B), consistent with its T-DNA insert being closed to the end ofits kinase domain-coding region of the gene. fer-4 and srn showed
Fig. 1. FER is a broadly expressed ROPGEF-interacting RLK. (A) The kinasedomain of FER [FER(K)] and ROPGEF1 (GEF1) interact in yeast two-hybridassays, complementing histidine deficiency (First and Third rows). FER(K)interacts with several other GEFs and ROPGEF1 interacts with RAC/ROP (Fig.S1 B and C) (19, 20). (B) FER and ROPGEF1 interact in BiFC assays in proto-plasts. YFP image for the interacting YN–EE–GEF1 and FER–HA–YC was ac-quired by autoexposure (Top); the same exposure condition was used forcontrol protoplasts expressing one of the partners (Middle and Bottom).(Scale bar, 10 μm.) (C) pFER::GUS expression pattern in 10-d-old seedlingsand roots and leaf from 10-d-old plants. Arrowheads, root hairs. [Scale bars,1 mm (seedling and inflorescence), 50 μm (root and stigma), and 500 μm(leaf).] (D) FER–GFP localization in pFER::FER-GFP transformed Arabidopsis.[Scale bars, 100 μm (root), 50 μm (trichome), and 10 μm (root, leaf epidermis,and root hairs).]
Fig. 2. Mutations in FER induce root hair and other vegetative phenotypes.(A) Domain map for FER. Locations of the T-DNA insertion alleles fer-4(GABI_GK106A06) and fer-5 (Salk_ 029056c), fer, and srn (31–33) and primersused in PCR analyses are indicated. SP, signal peptide; TM, transmembranedomain; UTR, untranslated region. Mature FER is ∼807 aa long. Genomic DNAanalysis for fer-4 and fer-5 is shown in Fig. S2A. (B) RT-PCR analysis for FERexpression. FER(K), PCRused primers 3 and4; FER(Ex), PCRused primers 1 and 2.(C) Root hairs from 4-d-old wild-type (WT), fer-4, fer-5 and srn, and pFER::FER-GFP complemented fer-4, -5 seedlings. (Scale bar, 100 μm.) See Fig. S2B for FER–GFP expression in the complemented fer seedlings. (D) fer mutations inducea broad spectrum of root hair defects. Top shows normal root hairs (Left) andfermutation-induced collapsed (the most severe), burst [seen with dischargedcytoplasm (arrowhead)], arrested, truncated, split, and short (the least severe)root hairs. (Scale bar, 100 μm.) The histogram shows distribution of the dif-ferent classes of root hairs in WT and mutant seedlings. Each data bar repre-sents the mean ± SD where n = 1,200 root hairs from 15–24 four-day-oldseedlings. Δs highlight normal root hairs from each sample. (E) fer-inducedgrowth suppression. (Scale bars, 10 mm (25 d) and 50 mm (45 d.) (F) fer-4induces trichome defects. Arrows, collapsed; arrowheads, more than threebranches. Approximately 30% of trichomes in fer-4 seedlings were normalcompared with >90% in WT (Fig. S2C). (G) fer-4 and srn show similar re-productive phenotype. (Scale bar, 2 mm.)
17822 | www.pnas.org/cgi/doi/10.1073/pnas.1005366107 Duan et al.

indistinguishable and multiple phenotypes, whereas fer-5’sdefects were milder and more restricted (see below).Consistent with the high expression level of FER in root hairs
(37) (Fig. 1C), fer-4, fer-5, and srn showed readily observable roothair defects that ranged from collapsed, burst, to short (Fig. 2 Cand D). Although the level of normal root hairs was similarly low,the majority of fer-4 and srn root hairs collapsed shortly afteremergence, whereas many of the fer-5 root hairs were shorter thanwild type (WT) or burst after some growth was achieved (Fig. 2 Cand D). fer-5 was negligibly affected in growth and development,whereas fer-4 showed significantly reduced stature, developed se-vere trichome defects (Fig. 2 E and F and Fig. S2C), and re-productively mimicked fer and srn (Fig.2G). The root hair defectsin fer-4, -5 were complemented by pFER::FER-GFP (Fig. 2C andFig. S2B), as were other growth and reproduction phenotypes infer-4 (e.g., Fig. 2G). Together these observations imply that FER’srole in growth and development is considerably broader thanregulating female gametophytic functions (31–33). Although ki-nase-deleted and extracellular domain-located mutations in someRLKs act in a dominant negative manner (38, 39), the fact that theT-DNA insert is at the end of the kinase domain and that het-erozygous fer-5 plants were nondistinguishable from wild-typeplants suggest fer-5 is more likely to have lost some aspects ofFER’s multiple functions that rely on an intact C-terminal region,whereas the null mutants fer-4, srn, and fer have lost all ofFER’s functions.
FER Functions in a RAC/ROP Signaling Pathway and Mediates Auxin-Regulated Root Hair Development. It is well established that auxinplays an important role in regulating root hair development (e.g.,refs. 40, 41) and we showed earlier that RAC/ROPs mediateauxin-signaled gene derepression via 26S proteasome-mediatedrepressor proteolysis (9, 10). We therefore asked whether FERfunctions in a RAC/ROP signaling pathway and also acts asa regulator for auxin-regulated root hair development. We ob-served that when grown in the presence of auxin, differencesbetween WT and fer root hairs were further amplified becauseroot hair elongation in WT seedlings was stimulated by auxin,whereas fer-4, fer-5, and srn root hairs failed to respond andremained short (Fig. 3 A–C and Fig. S2D). pFER::FER-GFPcomplemented this phenotype, restoring auxin-stimulated roothair growth in fer seedlings (Fig. 3 A and B).To establish that FER indeed acted upstream of RAC/ROPs,
we examined whether increasing RAC/ROP signaling capacity byoverexpressing GFP–ROP2 may counteract the root hair defectsin fer-4 and fer-5. Indeed fer-5 root hairs were restored to normal,including ability to respond to auxin stimulation (Fig. 3 D and Eand see also Fig. S3), consistent with reduced RAC/ROP signalingas underlying the fer-induced root hair defects under normalgrowth conditions and decreased sensitivity to auxin stimulation ofroot hair growth. However, a rescued fer-4 was not recovered.Possibly, the higher level of RAC/ROP overexpression required tocounteract fer-4’s deficiency could have prevented transgenic plantregeneration. Alternatively, and more pertinent to FER’s multiplefunctionality as suggested by the multiple phenotypes induced byfer-4 and other null or knockdown mutations (29–33), it is possiblethat fer-5 is defective only in FER’s RAC/ROP-interacting ca-pacity, whereas fer-4 lacks all of FER’s functions, including thosepotentially requiring an intact extracellular domain.To substantiate the FER and RAC/ROP signaling linkage on
a biochemical level, we used a functional pulldown assay thatspecifically targets activated RAC/ROPs (5, 8, 9). Indeed, thelevel of activated RAC/ROPs was reduced in fer-4, -5 seedlingsrelative to WT (Fig. 3F and Fig. S2E) consistent with FER beingnecessary for RAC/ROP activation thus working as an upstreamregulator for these small GTPases. Moreover, ROP2 maltose-binding protein (MBP) efficiently pulled down pFER-expressedFER–GFP from seedling microsomal proteins or when FER–
HA was overexpressed in mesophyll or root protoplasts (Fig. 3G–I), suggesting complex formation via their mutual interactionwith ROPGEFs, several of which are known to interact withFER (Fig. S1B).It is known that most ROPGEFs preferentially bind to GDP-
bound RAC/ROPs versus the GTP-bound activated form (19, 20),presumably upon nucleotide exchange stimulated by upstreamsignaling, the activatedGTPases would be released to interact withdownstream effectors. Therefore the presence of a signalingcomplex involving an upstream regulator, a ROPGEF and a RAC/ROP should exist but would likely be transient in vivo (see Fig.3K). To substantiate that the observed pulldown of FER by RAC/ROPs (Fig. 3G andH Left) was functionally relevant, we includedeither GDP or GTP in the pulldown reaction to determinewhether these guanine nucleotides differentially affect the pull-down efficiency. GDP and GTP would shift RAC/ROPs to theinactive or activated state, respectively, mimicking the action ofthese guanine nucleotides under physiological conditions. Indeed,pulldown of FER–HA was substantially enhanced by GDP butreduced by GTP (Fig. 3 H Right and I), consistent with the ob-servation that most ROPGEFs (e.g., ROPGEF4, Fig. 3J) arepreferentially pulled down by GDP-bound RAC/ROPs but not asefficiently by the GTP-bound form (19, 20). Together withobservations that ROPGEFs interact with FER (Fig. 1 A and Band Fig. S1B) and with RAC/ROPs (19, 20; Fig. S1C), results fromthese pulldown assays are consistent with a dynamic signalingcomplex involving interactions between FER, a ROPGEF, anda RAC/ROP, mediating nucleotide exchange and activating thesmall GTPases (Fig. 3J).
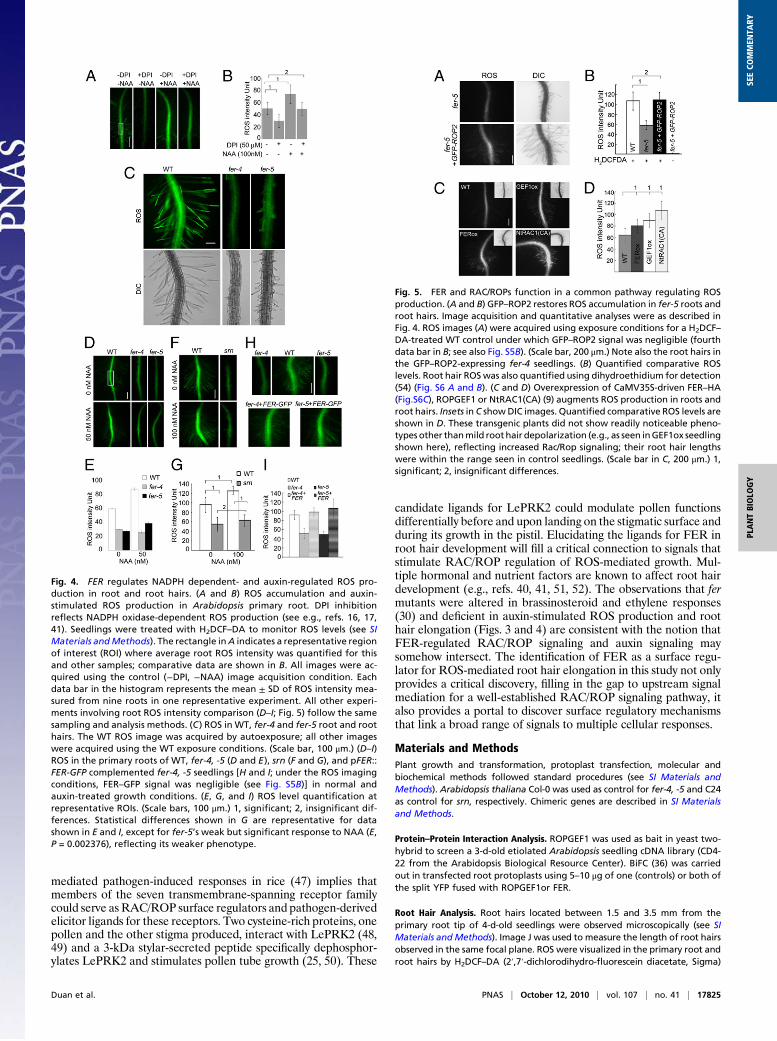
FER Regulates RAC/ROP-Mediated NADPH Oxidase-Dependent ROSAccumulation in Root and Root Hairs. The root hair defects in fermutants closely resemble those seen in rhd2 (Fig. S4). Togetherwith the FER, RAC/ROP signaling linkage (Fig. 3) and the RAC/ROP linkage to NADPH oxidase-dependent ROS-mediated roothair development (15–18), it seemed highly plausible that FERacted upstream of the RAC/ROP-regulated pathway. We there-fore examined whether fer mutants were compromised in RAC/ROP-regulated ROS accumulation.Roots and root hairs are known to show auxin-regulated diphe-
nylene iodonium (DPI)-sensitive NADPH oxidase-dependentROS accumulation and growth (e.g., refs. 13, 16, 17, 41, 42; Fig. 4Aand B; Fig. S5A). When treated with H2DCF–DA, a commonlyused ROS-detection stain, fer root and root hairs showed that theyaccumulated significantly lower levels ofROSrelative toWT(Fig. 4C andD–G, 0 nMNAA)and theywere alsononresponsive toauxin-stimulatedROSaccumulation (Fig. 4D–G,NAA-treated samples).Like the root hair morphological defects, the reduced ROS phe-notype in ferwas complemented by pFER::FER-GFP (Fig. 4H andI) and counteredbyoverexpressingROP2–GFP(Fig. 5A andB andsee also Fig. S5B and Fig. S6A andB). The corollary experiment ofoverexpressing RAC/ROPs, ROPGEF1, or FER also resulted inincreased ROS accumulation in transgenic seedling roots (Fig. 5Cand D and Fig. S6 C and D), further supporting that FER, ROP-GEFs, and RAC/ROPs act in a common pathway that controlsNADPH oxidase-dependent ROS production.
DiscussionIn showing that FER regulates RAC/ROP-signaled NADPHoxidase-dependent ROS-mediated root hair growth, our resultsreveal a surface regulator for a well-established signaling path-way that is not only important for the growth of these polarizedcells but also broadly implicated in growth, development, andstress management (11–15). That FER is broadly expressed (Fig.1) and loss-of-function fer mutants are pleiotropic and severelygrowth suppressed (Figs. 2–4) suggest FER plays an importantand general role in growth promotion throughout development.The presence of ROS in the growing regions of vegetative tissues
Duan et al. PNAS | October 12, 2010 | vol. 107 | no. 41 | 17823
PLANTBIOLO
GY
SEECO
MMEN
TARY

and that inhibiting NADPH oxidase-dependent ROS productioncorrelates with impaired growth (e.g., refs. 14, 16, 42) are con-sistent with the suggestion that FER targets the NADPH oxidase-regulated pathway to promote growth in a broad range of celltypes. The fact that ROS both promote and repress growth sug-gests the functional involvement of ROS in cell growth is complex.Using the diversely functioning RAC/ROPs as regulators forNADPH oxidase would be a versatile strategy that could maximizeinput regulation of ROS production and output modulation ofROS-mediated responses. Discovery of FER as an upstreamregulator for ROS-regulated growth, potentially in a large numberof cell types, opens the way to dissect how ROS-mediated sig-naling may be regulated to meet their roles under diverse con-ditions. On the other hand, given the multiple downstreampathways that RAC/ROPs are known to regulate (2–6), it is alsopossible that FER activates other RAC/ROP effectors to controladditional target systems, in different or even the same cell types.The fact that FER is required for pollen tube rupture inside thefemale gametophyte (31–33) suggests cell death, albeit noncell au-tonomously, as theultimate target.Therefore, a complete dissectionof FER functions throughout growth and development will beneeded to fully establish the potentially multiple signal-responsepathways that the broadly expressed FER is likely to mediate.Establishing the FER to RAC/ROP signaling linkage suggests
members of the broader FER-like RLK family (27) may also act assurface regulators for signal relay to regulate these molecularswitches. Thus far, four other Arabidopsis FER-like RLKs havebeen implicated in regulating growth, reproduction, and stress-related processes (28, 29, 34, 35). These seemingly diverse path-ways may nonetheless share certain functional themes. For in-stance, with the well-established role of apoplastic ROS incontrolling wall stiffness and extensibility (14, 43–45), it is con-ceivable that FER would mediate a ROS environment thatweakens the pollen tube wall, whereas ANX1 and ANX2 would doso to maintain its integrity. Given the role of NADPH oxidase-derived ROS in secondary cell wall synthesis and that RAC/ROPshave been implicated as regulating this process (46), it is con-ceivable for ROS to be involved in the THE1-mediated cell wallintegrity-dependent growth regulatory process. Taken together,available information on these FER-related RLKs suggests thatthe functional roles for at least a subset of these receptors may beinduced by or impact cell wall integrity and that ROS, with theirability to support cell expansion and induce wall stiffness, could becentral to the processes that these RLKs mediate.Defined signal-receptor linkages for RAC/ROP-regulated
pathways remain to be established. The observation of an α-sub-unit of a heterotrimeric G protein acting upstream of RAC/ROP-
Fig. 3. fermutants show reduced auxin and RAC/ROP signaling capacity. (A)Four-day-old WT, mutant, and pFER::FER-GFP complemented mutant seed-lings grown under standard (0 nM NAA) or auxin-supplemented conditions.Auxin-treated WT and fer-4, -5 mutants are shown magnified on the Right.(Scale bar, 2 mm.) (B) Root hair length comparison between WT, fer-5, andcomplemented fer-5 under standard and auxin-supplement growth con-ditions. Data bars represent average of the mean root hair lengths fromtriplicate samples ± SE. For each replicate, 300 root hairs from at least nineplants were measured. (C) Root hair length comparison between WT andfer-4 (Fig. S2D shows data for srn). (D and E) Overexpression of GFP–ROP2rescued fer-5–induced root hair morphological defects (D), restored root hairelongation (E, compare white and black bars), and auxin-stimulated roothair elongation (E, compare 0, 50, and 100 nM NAA data bars). See Fig. S3 forgenotyping of these rescued plants. (Scale bar, 100 μm.) (F) Pulldown ofactive RAC/ROPs. ICR1–MBP was used for fer-4; PBD–GST (8, 9) was used for
fer-5. Anti-NtRac1 antibody (9) was used for detection. The fer-4 blot wasvisualized by chemiluminescence, the fer-5 blot by alkaline phosphatasereaction. Quantification of data is shown in Fig. S2E. (G–J) ROP2–MBP pull-down of pFER-expressed FER–GFP from microsomal proteins from trans-formed seedlings (G), of FER–HA expressed in transiently transfectedmesophyll (H) and root (I) protoplasts, and of GEF4–HA expressed in meso-phyll protoplasts (J). Lower panels show input bait proteins on Ponçeau-stained blots. Anti-GFP (G) and anti-HA (H–J) were used for detection. *bandin G was unrelated to the experiment. (K) Sketch showing guanine nucle-otide-regulated FER–RAC/ROP interaction as suggested by results shown inG–J. Presumably, in vivo, signal activation-induced changes in the interactingROPGEF and RAC/ROP would weaken the GEF–RAC/ROP interaction, recy-cling the ROPGEF, and releasing activated RAC/ROP to interact with effectorsfor downstream signaling. Data for WT, fer-5, fer5 + FER–GFP and fer5 +GFP–ROP2 shown in B and Ewere collected in the same experiments thus thedata for WT and fer-5 are used in both panels. Mutant root hair data areoverestimations because only hairs with measurable lengths were includedin the data set. Brackets and the numbers 1 and 2 in C denote statisticalcomparisons (1, significant; 2, insignificant) as described in Materials andMethods; these differences are representative for data shown in B and E.
17824 | www.pnas.org/cgi/doi/10.1073/pnas.1005366107 Duan et al.
mediated pathogen-induced responses in rice (47) implies thatmembers of the seven transmembrane-spanning receptor familycould serve as RAC/ROP surface regulators and pathogen-derivedelicitor ligands for these receptors. Two cysteine-rich proteins, onepollen and the other stigma produced, interact with LePRK2 (48,49) and a 3-kDa stylar-secreted peptide specifically dephosphor-ylates LePRK2 and stimulates pollen tube growth (25, 50). These
candidate ligands for LePRK2 could modulate pollen functionsdifferentially before and upon landing on the stigmatic surface andduring its growth in the pistil. Elucidating the ligands for FER inroot hair development will fill a critical connection to signals thatstimulate RAC/ROP regulation of ROS-mediated growth. Mul-tiple hormonal and nutrient factors are known to affect root hairdevelopment (e.g., refs. 40, 41, 51, 52). The observations that fermutants were altered in brassinosteroid and ethylene responses(30) and deficient in auxin-stimulated ROS production and roothair elongation (Figs. 3 and 4) are consistent with the notion thatFER-regulated RAC/ROP signaling and auxin signaling maysomehow intersect. The identification of FER as a surface regu-lator for ROS-mediated root hair elongation in this study not onlyprovides a critical discovery, filling in the gap to upstream signalmediation for a well-established RAC/ROP signaling pathway, italso provides a portal to discover surface regulatory mechanismsthat link a broad range of signals to multiple cellular responses.
Materials and MethodsPlant growth and transformation, protoplast transfection, molecular andbiochemical methods followed standard procedures (see SI Materials andMethods). Arabidopsis thaliana Col-0 was used as control for fer-4, -5 and C24as control for srn, respectively. Chimeric genes are described in SI Materialsand Methods.
Protein–Protein Interaction Analysis. ROPGEF1 was used as bait in yeast two-hybrid to screen a 3-d-old etiolated Arabidopsis seedling cDNA library (CD4-22 from the Arabidopsis Biological Resource Center). BiFC (36) was carriedout in transfected root protoplasts using 5–10 μg of one (controls) or both ofthe split YFP fused with ROPGEF1or FER.
Root Hair Analysis. Root hairs located between 1.5 and 3.5 mm from theprimary root tip of 4-d-old seedlings were observed microscopically (see SIMaterials andMethods). Image J was used to measure the length of root hairsobserved in the same focal plane. ROS were visualized in the primary root androot hairs by H2DCF–DA (2′,7′-dichlorodihydro-fluorescein diacetate, Sigma)
Fig. 4. FER regulates NADPH dependent- and auxin-regulated ROS pro-duction in root and root hairs. (A and B) ROS accumulation and auxin-stimulated ROS production in Arabidopsis primary root. DPI inhibitionreflects NADPH oxidase-dependent ROS production (see e.g., refs. 16, 17,41). Seedlings were treated with H2DCF–DA to monitor ROS levels (see SIMaterials andMethods). The rectangle inA indicates a representative regionof interest (ROI) where average root ROS intensity was quantified for thisand other samples; comparative data are shown in B. All images were ac-quired using the control (−DPI, −NAA) image acquisition condition. Eachdata bar in the histogram represents the mean ± SD of ROS intensity mea-sured from nine roots in one representative experiment. All other experi-ments involving root ROS intensity comparison (D–I; Fig. 5) follow the samesampling and analysis methods. (C) ROS in WT, fer-4 and fer-5 root and roothairs. The WT ROS image was acquired by autoexposure; all other imageswere acquired using the WT exposure conditions. (Scale bar, 100 μm.) (D–I)ROS in the primary roots of WT, fer-4, -5 (D and E), srn (F and G), and pFER::FER-GFP complemented fer-4, -5 seedlings [H and I; under the ROS imagingconditions, FER–GFP signal was negligible (see Fig. S5B)] in normal andauxin-treated growth conditions. (E, G, and I) ROS level quantification atrepresentative ROIs. (Scale bars, 100 μm.) 1, significant; 2, insignificant dif-ferences. Statistical differences shown in G are representative for datashown in E and I, except for fer-5’s weak but significant response to NAA (E,P = 0.002376), reflecting its weaker phenotype.
Fig. 5. FER and RAC/ROPs function in a common pathway regulating ROSproduction. (A and B) GFP–ROP2 restores ROS accumulation in fer-5 roots androot hairs. Image acquisition and quantitative analyses were as described inFig. 4. ROS images (A) were acquired using exposure conditions for a H2DCF–DA-treated WT control under which GFP–ROP2 signal was negligible (fourthdata bar in B; see also Fig. S5B). (Scale bar, 200 μm.) Note also the root hairs inthe GFP–ROP2-expressing fer-4 seedlings. (B) Quantified comparative ROSlevels. Root hair ROSwas also quantified using dihydroethidium for detection(54) (Fig. S6 A and B). (C and D) Overexpression of CaMV35S-driven FER–HA(Fig.S6C), ROPGEF1 or NtRAC1(CA) (9) augments ROS production in roots androot hairs. Insets in C show DIC images. Quantified comparative ROS levels areshown in D. These transgenic plants did not show readily noticeable pheno-types other thanmild roothair depolarization (e.g., as seen inGEF1ox seedlingshown here), reflecting increased Rac/Rop signaling; their root hair lengthswere within the range seen in control seedlings. (Scale bar in C, 200 μm.) 1,significant; 2, insignificant differences.
Duan et al. PNAS | October 12, 2010 | vol. 107 | no. 41 | 17825
PLANTBIOLO
GY
SEECO
MMEN
TARY

(16, 17, 42, 53) and the superoxide-detecting dihydroethidium (DHE) as analternate dye (see SI Materials and Methods) (54). DPI, a commonly used in-hibitor of membrane-associated NADPH oxidase and other flavin-containingenzymes was used (50 μM) to differentiate NADPH oxidase produced ROSfrom those generated from other sources (e.g., refs. 16, 17, 42, 53). ROS wereimaged using a FITC filter (Ex460-500, DM505, Em510-560) for H2DCF–DAsignal or a rhodamine (Ex546/10, DM565LP, Em 590LP filter for dihydroethi-dium signal). ROS fluorescence intensity within a fixed region of interest (ROI)was quantified using Adobe Photoshop v.7.0. Experiments were repeated atleast three times. Student’s t-tests were used for statistical analyses. Pairwisedata comparisons in Figs. 3–5 are indicated by brackets above the data bars; 1denotes significant (P < 0.05), 2 denotes insignificant (P > 0.05) difference.Statistics details are shown in SI Materials and Methods.
Pulldown Assays. Pulldown assays for activated RAC/ROPs in 7-d-old WT andfer seedling roots used ICR1 (5)–MBP or PBD–GST (refs. 8, 9 and SI Materials
and Methods) as bait, based on the rationale that direct effectors of RHOGTPases specifically bind to their activated form. ROP2–MBP was used to pulldown pFER-expressed FER–GFP from seedlings, 35S-expressed FER–HA andROPGEF4–HA from mesophyll and root protoplasts. When guanine nucleo-tides were included in these assays, ROP2–MBP resin was pretreated with10 mM GTP or 10 mM GDP for 2 h and the pulldown assays were carried outwith 1 mM GTP or 10 mM GDP in the buffer. Details are described in SIMaterials and Methods.
ACKNOWLEDGMENTS. We thank Dr. F. Berger (National University ofSingapore) for srn seeds and Dr. J.-S. Lee (Ewha Womans University, Korea)for ROS detection protocol. We thank Dr. L. Tao for generating the Arabi-dopsis 35S-NtRac1(CA) transgenic plants. We thank a reviewer for suggest-ing DHE as an alternate dye for ROS detection. This work was supported byNational Science Foundation Grant IOB0544222 and US Department of Ag-riculture Grant CSREES 2004-35304-14837.
1. Etienne-Manneville S, Hall A (2002) Rho GTPases in cell biology. Nature 420:629–635.2. Nibau C, Wu H-M, Cheung AY (2006) RAC/ROP GTPases: ‘Hubs’ for signal integration
and diversification in plants. Trends Plant Sci 11:309–315.3. Yang Z, Fu Y (2007) ROP/RAC GTPase signaling. Curr Opin Plant Biol 10:490–494.4. Kost B (2008) Spatial control of Rho (Rac-Rop) signaling in tip-growing plant cells.
Trends Cell Biol 18:119–127.5. Lavy M, et al. (2007) A Novel ROP/RAC effector links cell polarity, root-meristem
maintenance, and vesicle trafficking. Curr Biol 17:947–952.6. Hazek O, et al. (2010) A rho scaffold integrates the secretory system with feedback
mechanisms in regulation of auxin distribution. PLoS Biol 8:e1000281.7. Wong HL, et al. (2007) Regulation of rice NADPH oxidase by binding of Rac GTPase to
its N-terminal extension. Plant Cell 19:4022–4034.8. Lemichez E, et al. (2001) Inactivation of AtRac1 by abscisic acid is essential for stomatal
closure. Genes Dev 15:1808–1816.9. Tao LZ, Cheung AY, Wu HM (2002) Plant Rac-like GTPases are activated by auxin and
mediate auxin-responsive gene expression. Plant Cell 14:2745–2760.10. Tao LZ, Cheung AY, Nibau C, Wu HM (2005) RAC GTPases in tobacco and Arabidopsis
mediate auxin-induced formation of proteolytically active nuclear protein bodies thatcontain AUX/IAA proteins. Plant Cell 17:2369–2383.
11. Werner E (2004) GTPases and reactive oxygen species: Switches for killing andsignaling. J Cell Sci 117:143–153.
12. Torres MA (2010) ROS in biotic interactions. Physiol Plant 138:414–429.13. Jaspers P, Kangasjärvi J (2010) Reactive oxygen species in abiotic stress signaling.
Physiol Plant 138:405–413.14. Swanson S, Gilroy S (2010) ROS in plant development. Physiol Plant 138:384–392.15. Carol RJ, Dolan L (2006) The role of reactive oxygen species in cell growth: Lessons
from root hairs. J Exp Bot 57:1829–1834.16. Foreman J, et al. (2003) Reactive oxygen species produced by NADPH oxidase regulate
plant cell growth. Nature 422:442–446.17. Carol RJ, et al. (2005) A RhoGDP dissociation inhibitor spatially regulates growth in
root hair cells. Nature 438:1013–1016.18. Jones MA, Raymond MJ, Yang Z, Smirnoff N (2007) NADPH oxidase-dependent
reactive oxygen species formation required for root hair growth depends on ROPGTPase. J Exp Bot 58:1261–1270.
19. Berken A, Thomas C, Wittinghofer A (2005) A new family of RhoGEFs activates theRop molecular switch in plants. Nature 436:1176–1180.
20. Gu Y, Li S, Lord EM, Yang Z (2006) Members of a novel class of Arabidopsis Rhoguanine nucleotide exchange factors control Rho GTPase-dependent polar growth.Plant Cell 18:366–381.
21. Basu D, Le J, Zakharova T, Mallery EL, Szymanski DB (2008) A SPIKE1 signalingcomplex controls actin-dependent cell morphogenesis through the heteromericWAVE and ARP2/3 complexes. Proc Natl Acad Sci USA 105:4044–4049.
22. Torii KU (2004) Leucine-rich repeat receptor kinases in plants: Structure, function, andsignal transduction pathways. Int Rev Cytol 234:1–46.
23. Kaothien P, et al. (2005) Kinase partner protein interacts with the LePRK1 and LePRK2receptor kinases and plays a role in polarized pollen tube growth. Plant J 42:492–503.
24. Zhang Y, McCormick S (2007) A distinct mechanism regulating a pollen-specificguanine nucleotide exchange factor for the small GTPase Rop in Arabidopsis thaliana.Proc Natl Acad Sci USA 104:18830–18835.
25. Zhang D, et al. (2008) The pollen receptor kinase LePRK2 mediates growth-promotingsignals and positively regulates pollen germination and tube growth. Plant Physiol148:1368–1379.
26. Shiu S-H, Bleecker AB (2003) Expansion of the receptor-like kinase/Pelle gene familyand receptor-like proteins in Arabidopsis. Plant Physiol 132:530–543.
27. Hématy K, Höfte H (2008) Novel receptor kinases involved in growth regulation. CurrOpin Plant Biol 11:321–328.
28. Hématy K, et al. (2007) A receptor-like kinase mediates the response of Arabidopsiscells to the inhibition of cellulose synthesis. Curr Biol 17:922–931.
29. Guo H, et al. (2009) Three related receptor-like kinases are required for optimal cellelongation in Arabidopsis thaliana. Proc Natl Acad Sci USA 106:7648–7653.
30. Deslauriers SD, Larsen PB (2010) FERONIA is a key modulator of brassinosteroid andethylene responsiveness in Arabidopsis hypocotyls. Mol Plant 3:626–640.
31. Rotman N, et al. (2003) Female control of male gamete delivery during fertilization inArabidopsis thaliana. Curr Biol 13:432–436.
32. Huck N, Moore JM, Federer M, Grossniklaus U (2003) The Arabidopsis mutant feroniadisrupts the female gametophytic control of pollen tube reception. Development130:2149–2159.
33. Escobar-Restrepo JM, et al. (2007) The FERONIA receptor-like kinase mediates male-female interactions during pollen tube reception. Science 317:656–660.
34. Miyazaki S, et al. (2009) ANXUR1 and 2, sister genes to FERONIA/SIRENE, are malefactors for coordinated fertilization. Curr Biol 19:1327–1331.
35. Boisson-Dernier A, et al. (2009) Disruption of the pollen-expressed FERONIAhomologs ANXUR1 and ANXUR2 triggers pollen tube discharge. Development 136:3279–3288.
36. Kerppola TK (2008) Bimolecular fluorescence complementation: Visualization ofmolecular interactions in living cells. Methods Cell Biol 85:431–470.
37. Zimmermann P, Hirsch-Hoffmann M, Hennig L, GruissemW (2004) GENEVESTIGATOR.Arabidopsis microarray database and analysis toolbox. Plant Physiol 136:2621–2632.
38. Diévart A, et al. (2003) CLAVATA1 dominant-negative alleles reveal functionaloverlap between multiple receptor kinases that regulate meristem and organdevelopment. Plant Cell 15:1198–1211.
39. Shpak ED, Lakeman MB, Torii KU (2003) Dominant-negative receptor uncoversredundancy in the Arabidopsis ERECTA leucine-rich repeat receptor-like kinasesignaling pathway that regulates organ shape. Plant Cell 15:1095–1110.
40. Jones AR, et al. (2009) Auxin transport through non-hair cells sustains root-hairdevelopment. Nat Cell Biol 11:78–84.
41. Pitts RJ, Cernac A, Estelle M (1998) Auxin and ethylene promote root hair elongationin Arabidopsis. Plant J 16:553–560.
42. Joo JH, Bae YS, Lee JS (2001) Role of auxin-induced reactive oxygen species in rootgravitropism. Plant Physiol 126:1055–1060.
43. Knight MR (2007) New ideas on root hair growth appear from the flanks. Proc NatlAcad Sci USA 104:20649–20650.
44. Monshausen GB, Bibikova TN, Messerli MA, Shi C, Gilroy S (2007) Oscillations inextracellular pH and reactive oxygen species modulate tip growth of Arabidopsis roothairs. Proc Natl Acad Sci USA 104:20996–21001.
45. Macpherson N, et al. (2008) NADPH oxidase involvement in cellular integrity. Planta227:1415–1418.
46. Potikha TS, Collins CC, Johnson DI, Delmer DP, Levine A (1999) The involvement ofhydrogen peroxide in the differentiation of secondary walls in cotton fibers. PlantPhysiol 119:849–858.
47. Suharsono U, et al. (2002) The heterotrimeric G protein alpha subunit acts upstreamof the small GTPase Rac in disease resistance of rice. Proc Natl Acad Sci USA 99:13307–13312.
48. Tang W, Kelley D, Ezcurra I, Cotter R, McCormick S (2004) LeSTIG1, an extracellularbinding partner for the pollen receptor kinases LePRK1 and LePRK2, promotes pollentube growth in vitro. Plant J 39:343–353.
49. Tang W, Ezcurra I, Muschietti J, McCormick S (2002) A cysteine-rich extracellularprotein, LAT52, interacts with the extracellular domain of the pollen receptor kinaseLePRK2. Plant Cell 14:2277–2287.
50. Wengier DL, Mazzella MA, Salem TM, McCormick S, Muschietti JP (2010) STIL,a peculiar molecule from styles, specifically dephosphorylates the pollen receptorkinase LePRK2 and stimulates pollen tube growth in vitro. BMC Plant Biol 10:33.
51. Zhang YJ, Lynch JP, Brown KM (2003) Ethylene and phosphorus availability haveinteracting yet distinct effects on root hair development. J Exp Bot 54:2351–2361.
52. Perry P, Linke B, Schmidt W (2007) Reprogramming of root epidermal cells in responseto nutrient deficiency. Biochem Soc Trans 35:161–163.
53. Allan AC, Fluhr R (1997) Two distinct sources of elicited reactive oxygen species intobacco epidermal cells. Plant Cell 9:1559–1572.
54. Bucana C, Saiki I, Nayar R (1986) Uptake and accumulation of the vital dye hydroethidinein neoplastic cells. J Histochem Cytochem 34:1109–1115.
17826 | www.pnas.org/cgi/doi/10.1073/pnas.1005366107 Duan et al.

Supporting InformationDuan et al. 10.1073/pnas.1005366107SI Materials and MethodsPlant Growth and Plant and Protoplast Transformation. Arabidopsisseeds were surface sterilized and germinated on B5 mediumsupplemented with 1% sucrose and solidified by 0.7% agar.Seeds were cold treated at 4 °C for 2 d for synchronization beforetransferring to 22 °C for germination. When supplemented withauxin, 1-naphthaleneacetic acid (NAA) was added to concen-trations indicated in the figures. For growth to maturity, 10-d-oldseedlings were transferred to soil and maintained in a growthchamber at 22 °C under 16/8 h light/dark cycles. Arabidopsis wastransformed by floral dip (1) and transgenic plants were selectedon kanamycin-containing (50 μg/mL) medium. Mesophyll androot protoplasts were isolated from 3-wk-old seedlings or roottissues of 2- to 3-wk-old plants grown vertically in tissue culture,respectively. Protoplast transformation followed previously de-scribed procedures (2).
Molecular, Biochemical, and Histochemical Analysis. All recombinantDNAprocedures were performed according to standard and PCR-based methodology. Table S1 shows a lists of primers used forcloning of cDNAs and genomic fragments and PCR amplification.Basic cloning strategies were used; sequences compatible withrestriction enzyme recognition sites were included at the end ofprimers shown in Table S1, indicating cloning strategies. FERpromoter (pFER)::FER-GFP containing 2,007 bp upstream of theFER coding region was cloned in Agrobacterium Ti plasmid in-termediate vector pAC1352 (3), introduced into AgrobacteriumGV2260 (4) and used to transform wild type (WT), fer-4, and fer-5.CaMV35S–FER–GFP, GFP–ROP2 (5), and NtRAC1(CA) (2)were similarly transformed into WT and fer plants as indicated inthe text. pFER::GUS was derived from pBI121 (Clontech) andtransformed into WT Arabidopsis. Plasmids used in protoplasttransient transfection assays, 35S–FER–HA and 35S–ROPGEF4–HA were constructed in Bluescript pSK (Strategene) using full-length FER and ROPGEF1 cDNAs. ICR1–MBP and ROP2–MBP were derived from pMALC (NEB) for Escherichia coli ex-pression. Genomic DNA was used for PCR analysis of T-DNAinserts. RNA from 10-d-old seedlings isolated by the PrepEaseRNA isolation kit (USB/Affymetrix) was used in RT-PCR analysisfor gene expression. The Clontech MATCHMAKER 2 protocolwas used for yeast two-hybrid assays (6). ROPGEF1 cDNA wascloned into the BD vector described in ref. 7 except that the genefor ampicillin resistance was replaced by NPTII to facilitate sub-sequent recovery of kanamycin-resistant plasmids in E. coli. Thekinase domain of FER was tested both as fusion with BD and ADwith GEF1–AD and GEF1–BD, accordingly. GUS staining oftransgenic seedlings followed standard procedure (8) in 0.2mg/mLX-gluc at 37 °C for 16 h.
Microscopy. Fluorescence and DIC microscopy were carried outon a NIKON Eclipse E800 microscope equipped with a SPOTcamera (Molecular Diagnostic). In comparative studies, imageacquisition conditions are detailed in the figure legends. AnOlympus SZ61 dissection microscope with a Q imaging camerawas used to acquire seedling, inflorescence, and trichome images.
Pulldown Assays. For ICR1 and PBD pulldown of activated RAC/ROPs, ICR1–MBP, and PBD–GST (2) was expressed in E. coliBL21 by IPTG (0.5 mM) induction. Cells were resuspended inbinding buffer [40 mM Tris-HCl (pH 7.5), 100 mM NaCl, 1 mMNa2-EDTA] and sonicated (six 10-s pulses at 1-min intervals) forprotein extraction. After removal of cell debris by centrifugation
at 4 °C, proteins were applied to amylose resin (NEB) or GSTresin in the same buffer and the ICR1–MBP or PBD–GST-boundresin was used to pull down activated RAC/ROPs. For targetproteins, the roots of 7-d-old WT and fer-4, -5 seedlings werehomogenized in liquid N2 and 100 mg of frozen powderwas extracted in pulldown buffer [binding buffer, plus 5% glyc-erol, 1 mM PMSF, protease inhibitor mixture (Calbiochem)]supplemented with 0.75% Triton X-100. Proteins were extractedat 4 °C with mixing for 15 min. The debris was removed by cen-trifugation at top speed in a microcentrifuge for 10 min at 4 °C.A total of 90% of the supernatant was applied to the ICR1–MBP-bound or PBD–GST-bound resin for pulldown assays. The re-maining 10% of the supernatant was saved for protein quantifi-cation. Pulldown was carried out by binding extracted proteins tothe amylose resin at 4 °C for 2 h. The resin was washed five times inbinding buffer. Proteins remained bound to the resin were elutedin SDS/PAGE loading buffer, boiled for 5 min, and applied to 15%SDS/PAGE for protein blot analysis. Protein blots were stained byPonçeauRed to ensure comparable loading of samples. RAC/ROPdetection was carried out by binding with anti-NtRac1 antibodies(2), followed by horseradish peroxidase-conjugated secondary an-tibodies and chemiluminescence detection (for ICR–MBP pull-down) or alkaline phosphatase-conjugated secondary antibodiesand colorimetric detection (for PBP–GST pulldown). Data fromimmunoblot were quantified by Adobe Photoshop. Mean histo-gram values were quantified for ROIs of identical area from eachband after background subtraction from blank region of the blots.For ROP2 pulldown of FER and GEF4–HA, ROP2–MBP-
bound amylose resin was generated as described above. For pro-toplast-expressed FER and GEF4, mesophyll or root protoplastswere transfected by 5–10 μg CaMV35S::FER-HA or CaMV35S::GEF4-HA. After overnight culture, protoplasts were collected andsonicated in pulldown buffer. After sonication, Triton X-100 wasadded to the extract to a final concentration of 0.4%, followed by15 min. of shaking at 4 °C. Pulldown reactions and subsequentanalyses were as described above. Five-day-old pFER–FER–GFPseedlings were also used for ROP2–MBP pulldown analysis.Seedlings were ground in liquid nitrogen and resuspended in ex-traction buffer [40 mM Tris-HCl (pH 7.5), 100 mM NaCl, 5%glycerol, 1 mMNa2-EDTA, PMSF, and protease inhibitor mixture(Calbiochem)]. Microsomes were extracted at 4 °C with mixing for30 min. Cell debris was removed by centrifugation at 10,000 g at 4 °C and the resulting supernatant was centrifuged at 100,000 g for 3h to pellet the microsomal fraction. The pellet was resuspended inextraction buffer supplemented with 0.3% Triton X-100 and thencentrifuged again at 2,000 g to remove insoluble material. Themicrosomal extract was then used for pulldown analysis. HA an-tibody (Santa Cruz) was used for detection.
Mutant Screening. One to three T-DNA insertion mutants forseveral vegetative tissue-expressed FER-related genes, At2g23200(HERK4), At5g24010 (HERK5), At5g54380 (THE1), At3g46290(HERK1), At5g61350, At3g51550 (FER), At1g30570 (HERK2),At2g39360 (HERK3), At5g39000, At5g38990, At5g39030, andAt5g39020 were obtained from the Salk collection and screenedfor root hair defects in 3- to 4-d-old seedlings. Only insertions inAt3g51550 (FER) induced readily noticeable root hair defects andwere studied in detail. The bulk of the screening was carried out in2007 and should be considered preliminary.
Statistics. Fig. 3B shows the difference between WT and fer-5 wassignificant, P < 0.05, 10−4, 10−4 at 0, 50, and 100 nM NAA.
Duan et al. www.pnas.org/cgi/content/short/1005366107 1 of 7

Difference between WT and complemented fer-5 was in-significant under all conditions (P > 0.1). Difference betweenuntreated and NAA treated fer-5 was insignificant (P > 0.05).Fig. 3C shows the difference between WT and fer-4 was sig-
nificant, P < 10−4 at 0 and 100 nM NAA. Difference betweenuntreated and NAA-treated WT was significant (P < 10−6).Difference between untreated and NAA-treated fer-4 was in-significant (P ≥ 0.5).Fig. 3E shows the difference between WT and fer-5 was signif-
icant, P < 0.05, 10−4, 10−4 at 0, 50, 100 nM NAA. Differencebetween WT and ROP2-rescued fer-5 was insignificant under allconditions (P > 0.1). Difference between untreated and NAA-treated fer-5 was insignificant (P > 0.05). Difference betweenuntreated and NAA-treated GFP–ROP2-rescued fer-5 was sig-nificant (P < 10−4).
Fig. 4 E and G shows the difference between WT and fer-4, -5and srn was significant (P < 10−6) under both conditions. Dif-ference between untreated and NAA-treated WT was significant(P < 10−5). Difference between untreated and NAA-treated fer-4, srn was insignificant (P > 0.05), but significant for fer-5 (P <0.05), reflecting a weaker phenotype than the two null mutants.Fig. 4I shows the difference between WT and mutants was
significant (P < 10−6), between WT and complemented mutantswas insignificant (P > 0.1).Fig. 5B shows the difference was significant (P < 10−4) be-
tween WT and fer-5, insignificant (P > 0.1) between WT andGFP–ROP2-rescued fer-5, and significant (P < 10−2) betweenfer-5 and GFP–ROP2-rescued fer-5. Note signal in GFP–ROP2-rescued fer-5 without incubation with the ROS substrateH2DCFDA (fourth data bar) was negligible.
1. Clough SJ, Bent AF (1998) Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16:735–743.
2. Tao LZ, Cheung AY, Wu HM (2002) Plant Rac-like GTPases are activated by auxin andmediate auxin-responsive gene expression. Plant Cell 14:2745–2760.
3. Rodermel SR, Abbott MS, Bogorad L (1988) Nuclear-organelle interactions: Nuclearantisense gene inhibits ribulose bisphosphate carboxylase enzyme levels intransformed tobacco plants. Cell 55:673–681.
4. Deblaere R, et al. (1985) Efficient octopine Ti plasmid-derived vectors forAgrobacterium-mediated gene transfer to plants. Nucleic Acids Res 13:4777–4788.
5. Jones MA, Raymond MJ, Yang Z, Smirnoff N (2007) NADPH oxidase-dependentreactive oxygen species formation required for root hair growth depends on ROPGTPase. J Exp Bot 58:1261–1270.
6. Gietz D, St Jean A, Woods RA, Schiestl RH (1992) Improved method for high efficiencytransformation of intact yeast cells. Nucleic Acids Res 20:1425.
7. James P, Halladay J, Craig EA (1996) Genomic libraries and a host strain designed forhighly efficient two-hybrid selection in yeast. Genetics 114:1425–1436.
8. Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: β-Glucuronidase asa sensitive and versatile gene fusion marker in higher plants. EMBO J 6:3901–3907.
9. Lavy M, et al. (2007) A novel ROP/RAC effector links cell polarity, root-meristemmaintenance, and vesicle trafficking. Curr Biol 17:947–952.
10. Nibau C, Wu H-M, Cheung AY (2006) RAC/ROP GTPases: ‘Hubs’ for signal integrationand diversification in plants. Trends Plant Sci 11:309–315.
11. Yang Z, Fu Y (2007) ROP/RAC GTPase signaling. Curr Opin Plant Biol 10:490–494.
12. Carol RJ, Dolan L (2006) The role of reactive oxygen species in cell growth: Lessonsfrom root hairs. J Exp Bot 57:1829–1834.
13. Swanson S, Gilroy S (2010) ROS in plant development. Physiol Plant 138:384–392.14. Jaspers P, Kangasjärvi J (2010) Reactive oxygen species in abiotic stress signaling.
Physiol Plant 138:405–413.15. Torres MA (2010) ROS in biotic interactions. Physiol Plant 138:414–429.16. Wong HL, et al. (2007) Regulation of rice NADPH oxidase by binding of Rac GTPase to
its N-terminal extension. Plant Cell 19:4022–4034.17. Kaothien P, et al. (2005) Kinase partner protein interacts with the LePRK1 and LePRK2
receptor kinases and plays a role in polarized pollen tube growth. Plant J 42:492–503.18. Zhang Y, McCormick S (2007) A distinct mechanism regulating a pollen-specific
guanine nucleotide exchange factor for the small GTPase Rop in Arabidopsis thaliana.Proc Natl Acad Sci USA 104:18830–18835.
19. Lemichez E, et al. (2001) Inactivation of AtRac1 by abscisic acid is essential forstomatal closure. Genes Dev 15:1808–1816.
20. Foreman J, et al. (2003) Reactive oxygen species produced by NADPH oxidase regulateplant cell growth. Nature 422:442–446.
21. Carol RJ, et al. (2005) A RhoGDP dissociation inhibitor spatially regulates growth inroot hair cells. Nature 438:1013–1016.
22. Berken A, Thomas C, Wittinghofer A (2005) A new family of RhoGEFs activates theRop molecular switch in plants. Nature 436:1176–1180.
23. Gu Y, Li S, Lord EM, Yang Z (2006) Members of a novel class of Arabidopsis Rhoguanine nucleotide exchange factors control Rho GTPase-dependent polar growth.Plant Cell 18:366–381.
Duan et al. www.pnas.org/cgi/content/short/1005366107 2 of 7

Fig. S1. (A) Schematic summary of the RAC/ROP signaling pathway showing the key elements involved in this study. ROPGEFs stimulate GDP/GTP exchange toactivate RAC/ROPs. GDI and GAP (GTPase activating protein) are negative regulators, one inhibiting GDP dissociation and the other accelerating GTP hydrolysis,respectively (10). Activated RAC/ROPs interact directly with downstream effectors RICs and ICR1 to mediate diverse cellular pathways (9–11). RAC/ROP-me-diated NADPH-oxidase–dependent ROS production is well established in mediating root hair growth and several biotic and abiotic stress-induced responses(12–16). In rice, NADPH oxidase has also been shown to be an immediate effector for OsRac1 to mediate pathogen-elicited defense responses (16). A largenumber of surface regulators (designated as ? in the figure) may be involved in mediating diverse signals to RAC/ROP-regulated pathways, but only two areknown (17, 18). Hormones, such as auxin (2) and ABA (19) and pathogen elicitors (16) have been shown to regulate RAC/ROP signaling. Using a ROPGEF asa molecular link, we report here identification of a surface regulator, FER, for the RAC/ROP-regulated pathway that stimulates NADPH oxidase-dependent ROSproduction to mediate polarized root hair growth (20, 21). (B) Yeast two-hybrid assays showing FER kinase domain [FER(K)] interact with multiple ArabidopsisROPGEFs. The originally isolated ROPGEF1-interacting fragments spanned half the RLKs’ kinase domain till the end of their coding region. (C) Yeast two-hybridassays showing ROPGEF1 interact with NtRAC1 (2), similar to previously shown for ROPGEFs and RAC/ROPs from Arabidopsis (22, 23).
Duan et al. www.pnas.org/cgi/content/short/1005366107 3 of 7

Fig. S2. Additional characterization of fer and complemented fer mutants. (A) Genomic DNA PCR analysis for fer-4 and fer-5. T-DNA insert was present in thefer mutants but not in WT (T-DNA) and genomic FER DNA fragments were present in WT but not in fer mutants [FER(Ex) and FER(K)]. For fer-4, primers 1 andGABI(R) were used for the T-DNA insert and primers 1 and 2 were used for the WT FER DNA fragments; for fer-5, primers 3 and LB1 and primers 3 and 4 wereused, respectively. See Fig. 2A and SI Materials and Methods for primer information. (B) FER–GFP expression in pFER::FER-GFP transformed fer-4 and fer-5mutants correlated with complemented root hair phenotype. (Scale bar, 100 μm.) (C) Quantitative data for fer-4–induced trichome defects. Data show themean % of each category of trichomes averaged from four comparably aged true leaves from 3-wk-old seedlings (a total of ∼350 trichomes were counted persample). Differences between WT and fer-4 are significant, Student’s t-tests showed P < 10−7 for trichomes with three or fewer branches, P = 1.15 × 10−3 forthose with more than four branches, and P = 4.7 × 10−3 for collapsed trichomes. (D) Root hair length comparison between WT and srn. Root hair lengths shownfor srn were overestimations because only hairs with measurable lengths but not the most severe ones that never elongated were included in the data set. (E)Relative levels of ROP–MBP pulled down fer-4 and ROP–GST pulled down fer-5. Data were averaged from quantification of data obtained from three in-dependent experiments. 1, significant difference (P < 0.05); 2, insignificant difference (P > 0.05).
Fig. S3. Analysis of CaMV35S::GFP-ROP2 transformed fer-5. (A) Genomic PCR analysis confirms presence of the original T-DNA insert (fer-5 T-DNA) usingprimers LB1 and 3 (see Fig. 2A), absence of the WT FER [FER(K)] using primers 3 and 4, confirming homozgyosity of the T-DNA insertion locus, and presence ofGFP DNA in the CaMV35S::GFP-ROP2 transformed fer-5. (B) Root hairs in fer-5 were restored to normal in GFP–ROP2-expressing CaMV35S::GFP-ROP2 trans-formed fer-5. (Scale bar, 200 μm.)
Duan et al. www.pnas.org/cgi/content/short/1005366107 4 of 7

Fig. S4. Comparison of root hair phenotypes in fer-4 and rhd2. (A) Root segments from 4-d-old fer-4 and rhd2 mutant seedlings. Arrows point to collapsedroot hairs. (Scale bar, 100 μm.) (B) Quantitative comparison of root hair defects. Data show that collapsed root hairs predominated in fer-4, whereas rhd2 roothair defects were more broadly distributed among different severity classes (see Fig. 2D for definition), indicating fer-4 induced more severely defective roothairs. Each data bar represents the mean ± SD where n = 600 root hairs sampled from 12 four-day-old seedlings.
Fig. S5. GFP signal from pFER::FER-GFP and CaMV35S::GFP-ROP2–transformed fer seedlings was negligible under H2DCF–DA-detected ROS imaging condition.(A) pFER::FER-GFP transformed fer-4 seedlings observed with (+) or without (−) H2DCF–DA treatment. The + H2DCF–DA image was acquired by autoexposure(373 ms); when exposed under the same condition, GFP signal from the − H2DCF–DA seedling was not detectable. Autoexposure of the same − H2DCF–DAseedling required a 10-s exposure, suggesting FER–GFP signal could not interfere with ROS detection. (B) CaMV35S::GFP-ROP2 transformed fer-5 seedlingsobserved with (+) or without (−) H2DCF–DA treatment. (Left) The WT seedling was treated with H2DCF–DA for ROS analysis and imaged by autoexposure(Upper; exposure time was 0.687 s). Lower panel shows a DIC image of the WT seedling. (Right) The fer-5 + GFP–ROP2 seedling was not treated for ROSdetection and imaged using 0.687-s exposure time (Upper) to detect contribution to the fluorescence signal by GFP–ROP2, which was negligible. Lower panelshows an autoexposed image of the same fer-5 + GFP–ROP2 seedling root to reveal the GFP signal. The long autoexposure time (9.27 s) signified a very lowfluorescence level from GFP–ROP2 under ROS detection condition.
Duan et al. www.pnas.org/cgi/content/short/1005366107 5 of 7

Fig. S6. (A and B) Dihydroethidium (DHE) detection of ROS shows strong signals in WT (Left) and CaMV35S::GFP-ROP2–rescued fer-5 (Right), whose root hairsshowed normal growth, but not in fer-5 mutant, whose root hairs showed aborted growth (Middle). Lower panels show higher magnification images acquiredusing the WT imaging condition for quantification of signals at root hair tips in boxed ROIs of identical areas. (B) Average ROS level (n = 43 root hairs persample) quantified from ROI of identical areas (boxes in A). Difference was significant (P < 0.05) between WT and fer-5, between fer-5 and GFP–ROP2-rescuedfer-5, and insignificant (P > 0.05) between WT and GFP–ROPs-rescued fer-5. (C) Protein blot analysis for CaMV35S::FER-HA overexpression lines. FERox1 wasrepresentative of lines with detectable FER–HA and showed enhanced root ROS accumulation relative to WT (see Fig.5 C and D). FERox2 was representative ofthe majority of similarly transformed lines; they did not accumulate detectable levels of FER–HA and their ROS levels were not distinguishable from controls.We could not recover transgenic lines expressing levels of FER–HA higher than that observed in FERox1. (D) RT-PCR for FER mRNA levels in WT and FERox1seedlings (7 d). Primers 1 and 2 (see Fig. 2A) were used for FER DNA amplification.
Duan et al. www.pnas.org/cgi/content/short/1005366107 6 of 7

Table S1. List of primers
Primers used for cloning purposes
cDNA/genomic DNA Primers (top row forward; bottom row reverse) RE* siteFER cDNA CGGATCCATGAAGATCACAGAGGGACGATTC BamHI/EcoRI
CGAATTCACGTCCCTTTGGATTCATGATCTGGEF1 cDNA GAGATCTATGGGGAGCTTATCTTC BglII/NcoI
CCCATGGCATCTCTTTCCGGCGTCACTCCGEF4 cDNA CGGATCCATGGAGAGTTCTTCGAATTCC BamHI/NcoI
GCCATGGCATCATCTCTGTTTCTCACTGFER promoter CGCGGCCGCCGAGTTGTAAAAGGCCTGGCTAAAG NotI/BamHI
CGGATCCCGATCAAGAGCACTTCTCCGGGICR1 cDNA CAGATCTATGCCAAGACCAAGAGTTTCAGAG BglII/EcoRI
CGAATTCCTTTTGCCCTTTCTTCCTCCACROP2 cDNA CGGATCCATGGCGTCAAGGTTTATTAAG BamHI/SalI
CGTCGACTCACAAGAACGCGCAACGGTTCPrimers used for PCR analysis
Primer 1† CGGATCCATGAAGATCACAGAGGGACGATTCPrimer 2† CGCAGATCTAGCACCAAACACACAAAACCCPrimer 3† CGGATCCATGGCTTACCGCAGACGTAAGCGTGGPrimer 4† CGAATTCACGTCCCTTTGGATTCATCATCTGLB1† GCGTGGACCGCTTGCTGCAACTGAB1 (R)† GTGGATTGATGTGATATCTCCGFP CGGATCCATGGTGAGCAAGGGCGAGGAG
CGTCGACTTACTTGTACAGCTCGTCCATGCCActin2 CGTACAACCGGTATTGTGCTGG
GGAGATCCACATCTGCTGGAATG
*Restriction enzyme.†See Fig. 2 for primer designation.
Duan et al. www.pnas.org/cgi/content/short/1005366107 7 of 7