garrett and grisham, biochemistry, third edition dna structure chapters 10&11 biochemistry by...
TRANSCRIPT

Garrett and Grisham, Biochemistry, Third Edition
DNA Structure Chapters 10&11
Biochemistryby
Reginald Garrett and Charles Grisham
Igor ChesnokovDepartment of Biochemistry and Molecular GeneticsOffice Phone # 934-6974E-mail: [email protected]

Garrett and Grisham, Biochemistry, Third Edition
Central Dogma of Molecular Biology Information Transfer in Cells
• Information encoded in a DNA molecule is transcribed via synthesis of an RNA molecule
• The sequence of the RNA molecule is "read" and is translated into the sequence of amino acids in a protein.

Garrett and Grisham, Biochemistry, Third Edition
DNA is the repository of genetic information (replication), RNA serves in the expression of this information through the processes of transcription and translation.

Garrett and Grisham, Biochemistry, Third Edition
Essential Questions
• What are the structures of the nucleotides?
• How are nucleotides joined together to form nucleic acids?
• What is the higher-order structure of DNA
• What are the biological functions of nucleotides and nucleic acids?

Garrett and Grisham, Biochemistry, Third Edition
Outline1
• What Is the Structure and Chemistry of Nitrogenous Bases?
• What Are Nucleosides? • What Is the Structure and Chemistry of
Nucleotides?• What Are Nucleic Acids? • What Are the Different Classes of Nucleic
Acids?

Garrett and Grisham, Biochemistry, Third Edition
Outline2
• What Sorts of Secondary Structures Can Double-Stranded DNA Molecules Adopt?
• Can the Secondary Structure of DNA Be Denatured and Renatured?
• What is the Tertiary Structure of DNA?
• What Is the Structure of Eukaryotic Chromosomes?

Garrett and Grisham, Biochemistry, Third Edition
Structure and Chemistry of Nitrogenous Bases
The bases of nucleotides and nucleic acids are derivatives of either pyrimidine or purine.
• Pyrimidines – Cytosine (DNA, RNA)– Uracil (RNA)– Thymine (DNA)
• Purines – Adenine (DNA, RNA)– Guanine (DNA, RNA)

Garrett and Grisham, Biochemistry, Third Edition
The pyrimidine ring system (six-membered heterocylic aromatic ring with two nitrogen atoms); by convention, atoms are numbered as indicated. (b) The purine ring system consists of two rings (pyrimidine and imidazole), nine atoms numbered as shown.
Both are relatively insoluble in water due to aromatic character.

Garrett and Grisham, Biochemistry, Third Edition
The common pyrimidine bases – cytosine (DNA and RNA), uracil (RNA), and thymine (DNA).
Garrett and Grisham, Biochemistry, Third Edition
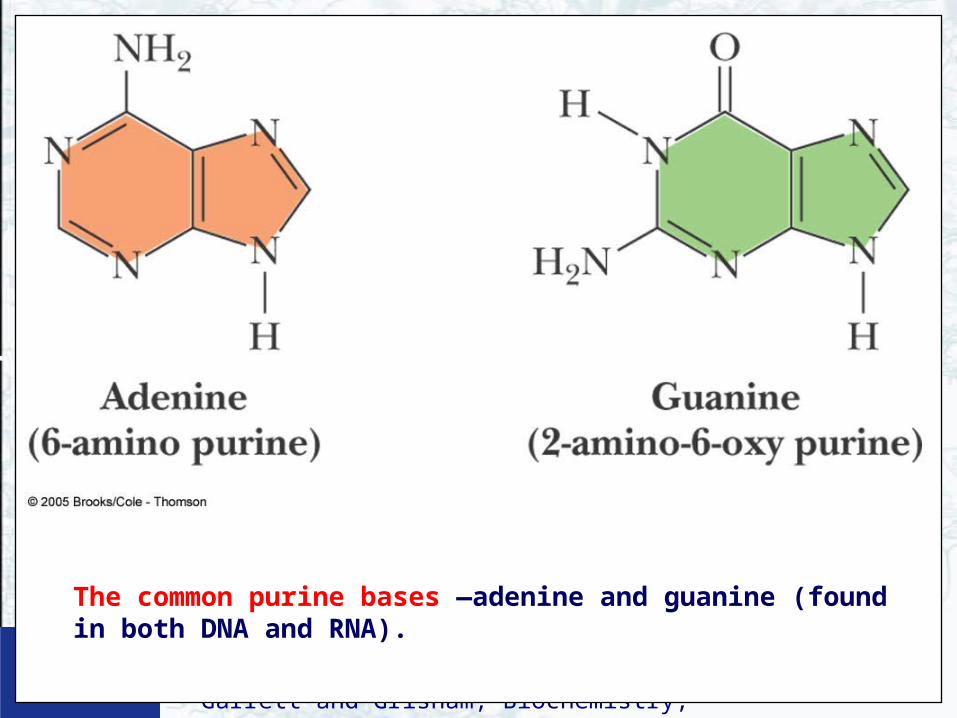
The common purine bases —adenine and guanine (found in both DNA and RNA).

Garrett and Grisham, Biochemistry, Third Edition
What Are Nucleosides?
• When a base is linked to a sugar it forms a Nucleoside.• The sugars are Pentoses• D-ribose (in RNA) • 2-deoxy-D-ribose (in DNA) • The difference - 2'-OH vs 2'-H • This difference affects secondary structure and stability
of nucleic acids

Garrett and Grisham, Biochemistry, Third Edition
Pentose in a five membered ring is known as furanose. Furanose structures (ribose and deoxyribose) are presented above. Presence of a hydroxyl group at the 2-position has dramatic effect on secondary structures available to DNA and RNA as well as their susceptibilities to hydrolysis. DNA is more stable.

Garrett and Grisham, Biochemistry, Third Edition
continued …• Base in nucleosides is linked to a sugar via a
glycosidic bond • 1’C of sugar links to 9 N of purine or to the 1 N of
pyrimidine base• Nucleosides named by adding –idine (ur-idine)
to the root name of a pyrimidine or –osine (aden-osine) to the root name of a purine
• Sugars make nucleosides more water-soluble than free bases

Garrett and Grisham, Biochemistry, Third Edition
-Glycosidic bonds link nitrogenous bases and sugars to form nucleosides.
1’C links to 9 N of purine and to the 1 N of pyrimidine base.

Garrett and Grisham, Biochemistry, Third Edition
The common ribonucleosides—cytidine, uridine, adenosine, and guanosine. Also, inosine (uncommon nucleoside) is drawn.

Garrett and Grisham, Biochemistry, Third Edition
Structure and Chemistry of Nucleotides
Nucleotides or Nucleoside phosphates result when phosphoric acid is esterified to a sugar-OH group of nucleoside (at C-5)•Most nucleotides are ribonucleotides •Nucleotides have acidic properties•Nucleic acids, which are polymers of nucleosides derive their names from the acidity of phosphate groups.

Garrett and Grisham, Biochemistry, Third Edition
Structures of the four common ribonucleotides —AMP, GMP, CMP, and UMP—together with their two sets of full names, for example, adenosine 5'-monophosphate and adenylic acid. Also shown is the nucleoside 3'-AMP (uncommon, product of hydrolysis).

Garrett and Grisham, Biochemistry, Third Edition
Functions of Nucleotides• NTPs and dNTPs are substrates for nucleic acids• Nucleoside 5'-triphosphates are carriers of energy.
Energy is stored in phosphoric bonds. • Bases serve as recognition units or information symbol
but not involved in the biochemistry of metabolism• ATP is central to energy metabolism (energy currency) • GTP drives protein synthesis • CTP drives lipid synthesis • UTP drives carbohydrate metabolism• Cyclic nucleotides are signal molecules and regulators
of cellular metabolism and reproduction

Garrett and Grisham, Biochemistry, Third Edition
Phosphoryl and pyrophosphoryl group transfer, the major biochemical reactions of nucleotides. Phosphoric bonds are prime source of chemical energy to do biological work (ATP, GTP, CTP and UTP, also deoxy- counterparts).

Garrett and Grisham, Biochemistry, Third Edition
Cyclic nucleotides are cyclic phosphodiesters.
Structures of the cyclic nucleotides cAMP and cGMP. Phosphoric acid is esterified to two of the available ribose hydroxyl groups.
Important! They are regulators of cellular metabolism and are found in all cells.

Garrett and Grisham, Biochemistry, Third Edition
What are Nucleic Acids? Polynucleotides!
•Facts to remember:•Linear polymers of nucleotides linked 3‘C to 5‘C by phosphodiester bridges •Ribonucleic acid and Deoxyribonucleic acid •Know the shorthand notations •Sequence is always read 5' to 3' •In terms of genetic information, this corresponds to "N to C" in proteins

Garrett and Grisham, Biochemistry, Third Edition
3'-5' phosphodiester bridges link nucleotides together to form polynucleotide chains.

Garrett and Grisham, Biochemistry, Third Edition
Shorthand notations for polynucleotide structures.
Furanoses are represented by vertical lines; phosphodiesters are represented by diagonal slashes in this shorthand notation for nucleic acid structures. Bases serve as distinctive side chains and give the polymer it’s unique identity.

Garrett and Grisham, Biochemistry, Third Edition
What Are the Different Classes of Nucleic Acids?
• DNA - one type, one purpose
• RNA - 3 (or 4) types, 3 (or 4) purposes
– ribosomal RNA - the basis of structure and function of ribosomes
– messenger RNA - carries the message
– transfer RNA - carries the amino acids– Small nuclear RNA– Small non-coding RNAs

Garrett and Grisham, Biochemistry, Third Edition
DNA & RNA Differences?
Why does DNA contain thymine?
• Cytosine spontaneously deaminates to form uracil (C-G pair could result in U-A)
• Repair enzymes recognize these "mutations" and replace these Us with Cs
• But how would the repair enzymes distinguish natural U from mutant U
• Nature solves this dilemma by using thymine (5-methyl-U) in place of uracil

Garrett and Grisham, Biochemistry, Third Edition
Deamination of cytosine forms uracil.

Garrett and Grisham, Biochemistry, Third Edition
The 5-methyl group on thymine labels it as a special kind of uracil.

Garrett and Grisham, Biochemistry, Third Edition
DNA & RNA Differences?
Why is DNA 2'-deoxy and RNA is not?
• Vicinal -OH groups (2' and 3') in RNA make it more susceptible to hydrolysis
• DNA, lacking 2'-OH is more stable • This makes sense - the genetic material
must be more stable • RNA is designed to be used and then
broken down

Garrett and Grisham, Biochemistry, Third Edition
How Do Scientist Determine the Primary Structure of Nucleic Acids?
Sequencing Nucleic Acids • Chain termination method (dideoxy method),
developed by F. Sanger • Base-specific chemical cleavage, developed
by Maxam and Gilbert • Both use autoradiography - X-ray film
develops in response to presence of radioactive isotopes in nucleic acid molecules

Garrett and Grisham, Biochemistry, Third Edition
DNA Replication• Chain termination method is based on biochemistry
of DNA replication • Each strand of the double-helical DNA molecule
must be copied in complementary fashion by DNA polymerase
• Each strand can serve as a template for copying • DNA polymerase requires template and primer • Primer: an oligonucleotide that pairs with the end of
the template molecule to form dsDNA • DNA polymerases add nucleotides in 5'-3' direction

Garrett and Grisham, Biochemistry, Third Edition
DNA polymerase copies ssDNA in vitro in the presence of the four deoxynucleotide monomers. A double-stranded region of DNA must be artificially generated by adding a primer, an oligonucleotide capable of forming a short stretch of dsDNA by base pairing with the ssDNA. The primer must have a free 3'-OH end from which the new polynucleotide chain can grow as the first residue is added in the initial step of the polymerization process.

Garrett and Grisham, Biochemistry, Third Edition
Chain Termination Method
Based on DNA polymerase reaction • Run four separate reactions • Each reaction mixture contains dATP,
dGTP, dCTP and dTTP, one of which is P-32-labelled
• Each reaction also contains a small amount of one dideoxynucleotide: either ddATP, ddGTP, ddCTP or ddTTP

Garrett and Grisham, Biochemistry, Third Edition
Chain Termination Method
• Most of the time, the polymerase uses normal nucleotides and DNA molecules grow normally
• Occasionally, the polymerase uses a dideoxynucleotide, which adds to the chain and then prevents further growth in that molecule
• Random insertion of dd-nucleotides leaves (optimally) at least a few chains terminated at every occurrence of a given nucleotide

Garrett and Grisham, Biochemistry, Third Edition
Chain Termination Method
• Run each reaction mixture on electrophoresis gel • Short fragments go to bottom, long fragments on
top • Read the "sequence" from bottom of gel to top • Convert this "sequence" to the complementary
sequence

Garrett and Grisham, Biochemistry, Third Edition
The chain termination or dideoxy method of DNA sequencing. (a) DNA polymerase reaction. (b) Structure of dideoxynucleotide. (c) Four reaction mixtures with nucleoside triphosphates plus one dideoxynucleoside triphosphate. (d) Electrophoretogram. Note that the nucleotide sequence as read from the bottom to the top of the gel is the order of nucleotide addition carried out by DNA polymerase.

Garrett and Grisham, Biochemistry, Third Edition
Chemical Cleavage MethodNot used as frequently as Sanger's
• Start with ssDNA labelled with P-32 at one end
• Strand is cleaved by chemical reagents
• Assumption is that strands of all possible lengths will be produced, each cleaved at just one of the occurrences of a given base.
• Fragments are electrophoresed and sequence is read

Garrett and Grisham, Biochemistry, Third Edition
A photograph of the autoradiogram from an actual sequencing gel. A portion of the DNA sequence of nit-6, the Neurospora gene encoding the enzyme nitrite reductase.

Garrett and Grisham, Biochemistry, Third Edition
Structure of DNAsummary
The fundamental structure of DNA is a Double Helix stabilized by hydrogen bonds!
•DNA consists of two polynucleotide strands wound together to form DNA double helix•Strands run in opposite direction (antiparallel)•Two strands are held together through inter-chain hydrogen bonds•These H bonds pair the bases of nucleotides in one chain to complementary bases in the other – base pairing.

Garrett and Grisham, Biochemistry, Third Edition
The DNA Double Helix
•Erwin Chargaff had the pairing data, but didn't understand its implications. Chargaff rule – the number of purine residues equals the number of pyrimidine residues in all organisms (A=T, G=C). •Rosalind Franklin's X-ray fiber diffraction data was crucial (Helix!)•Watson-Crick model of the DNA double helix.

Garrett and Grisham, Biochemistry, Third Edition
(a) Double-stranded DNA as an imaginary ladderlike structure. (b) A simple right-handed twist converts the ladder to a helix.

Garrett and Grisham, Biochemistry, Third Edition
A model of DNA double helix.
The nucleotides are linked covalently by phoshodiester bonds through the3’-hydroxil (-OH) group of one sugar and the 5’-phosphate (P) of the next. Two DNA strands are held togetherby hydrogen bonds between the paired bases. Two hydrogen bonds form between A and T, while three form between G and C. The bases can pair in this way only if the two polynucleotide chains that contain them are antiparallel to each other.
The coiling of the two strands around each other creates two groves in the double helix.
Consequences – each strand of DNA contains a sequence of nucleotides that are exactly complementary tothe sequence of its partner strand.

Garrett and Grisham, Biochemistry, Third Edition
The bases in a base pair are not directly across the helix axis from one another but rather are slightly displaced. This displacement, and the relative orientation of the glycosidic bonds linking the bases to the sugar–phosphate backbone, leads to differently sized grooves in the cylindrical column created by the double helix, the major groove and the minor groove, each coursing along its length.
How grooves are formed?

Garrett and Grisham, Biochemistry, Third Edition
Comparison of A, B, Z DNAABZs of DNA Secondary Structure
• A: right-handed, short and broad, 2.3 Å, 11 bp per turn (dehydrated DNA, probably does not exist in vivo)
• B: right-handed, longer, thinner, 3.32 Å, 10 bp per turn, (most common)
• Z: left-handed, longest, thinnest, 3.8 Å, 12 bp per turn (G-C rich regions)

Garrett and Grisham, Biochemistry, Third Edition
Comparison of the A-, B-, and Z-forms of the DNA double helix. The distance required to complete one helical turn is shorter in A-DNA than it is in B-DNA. The alternating pyrimidine–purine sequence of Z-DNA is the key to the “left-handedness” of this helix.

Garrett and Grisham, Biochemistry, Third Edition
Can the Secondary Structure of DNA Be Denatured and Renatured?
Important for Study of Genome complexity• When DNA is heated to 80+ degrees Celsius,
its UV absorbance increases by 30-40% • This hyperchromic shift reflects the unwinding of
the DNA double helix • Stacked base pairs in native DNA absorb less
light • When temperature is lowered, the absorbance
drops, reflecting the re-establishment of stacking

Garrett and Grisham, Biochemistry, Third Edition
Steps in the thermal denaturation and renaturation of DNA. Renaturation (re-annealing) depends on DNA concentration and time. The nucleation phase of the reaction is depending on sequence alignment of the two strands. This process takes place slowly because it takes time for complementary sequences to encounter one another in solution and then align themselves in register. Once the sequences are aligned, the strands zipper up quickly.

Garrett and Grisham, Biochemistry, Third Edition
These c0t curves show the rates of re-association of denatured DNA from various sources and illustrate how the rate of re-association is inversely proportional to genome complexity. The DNA sources are as follows: poly A+poly U, a synthetic DNA duplex of poly A and poly U polynucleotide chains; mouse satellite DNA, a fraction of mouse DNA in which the same sequence is repeated many thousands of times; MS-2 dsRNA, the double-stranded form of RNA of MS-2, a simple bacteriophage; T4 DNA, the
DNA of a more complex bacteriophage; E. coli DNA, bacterial DNA; calf DNA (nonrepetitive fraction), mammalian DNA (calf) from which the highly repetitive DNA fraction (satellite DNA) has been removed. Arrows indicate the genome size (in bp) of the various DNAs.

Garrett and Grisham, Biochemistry, Third Edition
Tertiary Structure of DNA
• In duplex DNA, ten bp per turn of helix • Circular DNA sometimes has more or less
than 10 bp per turn - a supercoiled state, underwound (-) or overwound (+).
• Enzymes called topoisomerases or gyrases can introduce or remove supercoils

Garrett and Grisham, Biochemistry, Third Edition
There are toroidal and interwound varieties of DNA supercoiling. (a) The DNA is coiled in a spiral fashion about an imaginary toroid. (b) The DNA interwinds and wraps about itself. (c) Supercoils in long, linear DNA arranged into loops whose ends are restrained—a model for chromosomal DNA.

Garrett and Grisham, Biochemistry, Third Edition
A simple model for the action of bacterial DNA gyrase (topoisomerase II).
The A-subunits cut the DNA duplex (1) and then hold onto the cut ends (2). Conformational changes occur in the enzyme that allow a continuous region of the DNA duplex to pass between the cut ends and into an internal cavity of the protein. The cut ends are then re-ligated (3), and the intact DNA duplex is released from the enzyme. The released intact circularDNA now contains two negative supercoils as a consequence of DNA gyrase action (4).

Garrett and Grisham, Biochemistry, Third Edition
Supercoiled DNA in a toroidal form wraps readily around protein “spools.” A twisted segment of linear DNA with two negative supercoils (a) can collapse into a toroidal conformation if its ends are brought closer together (b) Wrapping the DNA toroid around a protein “spool” stabilizes this conformation of supercoiled DNA (c)

Garrett and Grisham, Biochemistry, Third Edition
Structure of Eukaryotic Chromosomes
• Human DNA’s total length is ~2 meters!
• This must be packaged into a nucleus that is about 5 micrometers in diameter
• This represents a compression of more than 100,000!
• It is made possible by wrapping the DNA around protein spools called nucleosomes and then packing these in helical filaments

Garrett and Grisham, Biochemistry, Third Edition
Nucleosome Structure
• Chromatin, the nucleoprotein complex, consists of histones and non-histone chromosomal proteins
• Histone octamer structure has been solved (without DNA by Moudrianakis, and with DNA by Richmond)
• Non-histone proteins are regulators of gene expression

Garrett and Grisham, Biochemistry, Third Edition
A diagram of the histone octamer.
Nucleosomes consist of two turns of DNA supercoiled about a histone “core” octamer.

Garrett and Grisham, Biochemistry, Third Edition
Nucleosomes as seen in the electron microscope.“Beads on a string” – partially unfolded chromatin.

Garrett and Grisham, Biochemistry, Third Edition
The 2-nm DNA helix is wound twice around histone octamers to form 10-nm nucleosomes.
These nucleosomes are then wound in solenoid fashion with six nucleosomes per turn to form a 30-nm filament.
The 30-nm filament forms long DNA loops, each containing about 60,000 bp, which are attached at their base to the nuclear matrix.
Eighteen of these loops form a miniband unit of a chromosome.
Approximately 106 of these minibands occur in each chromatid of human chromosome 4 at mitosis.

Garrett and Grisham, Biochemistry, Third Edition
Each DNA molecule that forms a linear chromosome must contain a Centromere, two Telomeres and Replication Origins
Sequence of events a typical chromosome follows during the cell cycle:The DNA replicates in interphase beginning at the origins of replication and proceeding bidirectionally from theorigins across the chromosome. Many origins are required to ensure that the entire chromosome can be replicated rapidly. In M phase, the centromere attaches the duplicated chromosome to the mitotic spindleso that one copy is distributed to each daughter cell during mitosis. The centromere also helps to hold theduplicated chromosomes together until they are ready to be moved apart. The telomeres form special capsat each chromosome end.