generation of tumor-specific cytolytic t lymphocytes from peripheral blood of cervical cancer...
TRANSCRIPT

Generation of tumor,specific cytolytic T lymphocytes from peripheral blood of cervical cancer patients by in vitro stimulation with a synthetic human papillomavirus type 16 E7 epitope
Margaret Alexander , MD, a'b Michael L. Salgaller, PhD, a Esteban Celis, PhD, c
Alessandro Sette, PhD, c WiUard A. Barnes, MD, b Steven A. Rosenberg , MD, PhD, a and
Michae l A. Steller, MD a
Bethesda, Maryland, Washing¢on, D.C., and San Diego, California
OBJECTIVE: Approximately 90% of squamous carcinomas of the cervix harbor the human papillomavirus and type 16 has been detected in nearly 50% of cases. Recent studies in mice have shown that the
• human papillomavirus type 16 E7 oncoprotein contains peptide epitopes that are processed and presented in association with a major histocompatibility antigen for recognition by cytolytic T lymphocytes. We investigated whether an epitope from human papillomavirus type 16 E7 could be used to generate specific human cytolytic T lymphocytes in patients with cervical carcinoma. STUDY DESIGN: After radiation therapy, three patients with antigen HLA-A2 and with locally advanced cervical cancer underwent leukapheresis. Epitope-specific cytolytic T lymphocytes were generated from the peripheral blood mononuclear cells by in vitro stimulation with autologous peripheral blood mononuclear cells pulsed with a human papillomavirus type 16 E7, HLA-A2-restricted, synthetic peptide, E711_2o (YMLDLQPE-I-F). RESULTS: In two patients cytolytic T lymphocytes were capable of E711_2o-specific, HLA-A2-restricted cytolysis of the peptide-pulsed, HLA-matched, T2 target cell line. Cytolytic T lymphocytes from one of these patients also demonstrated specific cytolysis against the HLA-A2 +, HPV-16 + CaSki cervical cancer cell line but did not lyse either HLA-A2 +, HPV-16- MS-751 cells or HLA-A2-, HPV-16- HT-3 cells. CONCLUSIONS: These experiments demonstrate that novel cytolytic T lymphocytes that recognize a human papiltomavirus type 16 E7 epitope can be generated by using the peripheral blood mononuclear cells from irradiated patients with cervical cancer. In addition, because CaSki cells were specifically lysed by the cytolytic T lymphocytes, these data indicate that the peptide E711_2o is endogenously processed and presented on the cell surface of the CaSki cells. The demonstration of epitope-specific lysis of cytolytic T lymphocytes of HPV-16 + cervical cancer cells supports further efforts to develop human papillomavirus peptide-based vaccines or antigen-specific adoptive immunotherapy for the prevention and treatment of cervical carcinoma. (Am J Obstet Gynecol 1996;175:1586-93.)
Key words: Human papillomavirus, vaccine, peptide epitopes, cervical cancer, cytolytic T lymphocytes
Cervical cancer is a leading cause of cancer-related death in women worldwide and accounts for >200,000 deaths annually. 1 On the basis of extensive epidemio- logic data, the human papillomavirus (HPV) is strongly associated with a spectrum of anogenital neoplasias, including condyloma, cervical dysplasia, and cervical cancer. Such studies indicate that nearly 90% of invasive cervical tumors harbor HPV deoxyribonucleic acid, and
From the Surgery Branch, National Cancer Institute, ~ the Division of Gynecologic Oncology, Georgetown University Medical Center, b and Cytel Corp ~ Received for publication March 7, 1996; revised May 15, 1996; accepted June 19, 1996. Reprint requests: Michael A. Steller, MD, National Cancer Institute, Bldg 10, Rm 2B-42, 9000 Rockville Pike, Bethesda, MD 20892-1502. 6/1/75967
type 16 can be detected in approximately 50% of squa- mous cell carcinomas of the cervix. 2
The E6 and E7 oncoproteins of HPV-16 have trans- forming ability and have been implicated in the tumor- igenicity of HPV, and their consistent expression appears necessary to maintain a neoplastic s ta te) Both E6 and E7 can alter cell growth regulation by inactivating the prod- ucts of tumor suppressor genes p53 and pRB, respective- ly. 4 Several lines of evidence suggest the importance of the host's immune response in the pathogenesis of HPV-associated cervical lesions. For example, >60% of HPV-positive, mildly dysplastic lesions resolve spontane- ously 5 and such factors as human immunodeficiency virus infection and exposure to immunosuppressive agents have been strongly associated with HPV-induced
1586

Volume 175, Number 6 Alexander et al. 1587 Am J Obstet G)mecol
carcinogenesis.< 7 Thus these observations form the basis for the development of novel immunotherapies.
Presentation to T lymphocytes of endogenously pro- cessed antigens involves degradation of the protein into small pept ide epitopes. Intracellularly, peptides bind to nascent class I heavy-chain [32-microglobulin dimers to form a stable trimeric complex that is transported to the cell surface, resulting in recognition by T cells that are specific for structural features of both the class I mole- cule and the particular major histocompatibility (MHC)- bound peptide. So far, all the peptide binding sites of class I molecules examined possess long grooves on the top surface of the molecule. The amino acid residues at the ends of the 8, 9, or 10 amino acid peptides that bind to the groove are highly conserved and are felt to play a role as stabilizing residues that determine specific pep- tide anchoring. 8 Cytolytic T cells recognize antigens by the binding of their clonotypic receptors to the endog- enously processed peptides associated with class I mole- cules. On the basis of these immunologic observations, class I restricted cytolytic T-lymphocyte peptide epitopes have been generated from HPV-16 E7. Results from both in vitro and in vivo experiments suggest that HPV-16 E7 peptides stimulate an immune response in H2-D b trans- genic mice. 9 Because of the consistent expression of E7 in human cervical tumors associated with HPV-16 m and the recognition that E7 contains peptide epitopes that are processed intracellularly and presented in association with the class I molecule HLA-A2, n we investigated whether an HPV-16 E7 epitope may be used to generate cytolytic T lymphocytes in patients with invasive cervical cancer.
Material and methods
Cell lines. CaSki cervical cancer cells (which are HPV- 16 +),12, la HT-3 and MS-751 cervical cancer cells (which are both HPV-16 12, 13), and the Daudi lymphoma cell
line were purchased from American Type Culture Col- lection (Rockville, Md.). The HLA-A2-positive, antigen- processing defective human lymphoblastoid cell line T2 is maintained in the Surgery Branch and has been described elsewhere. 14
Human leukocyte antigen (HI,A) typing. HLA typing of peripheral blood mononuclear cells from patients with cervical cancer was performed by the National Institutes of Health HLA Typing Laboratory with the modified Amos microcytotoxity assay. 15
Peptides. The HPV E711_20 peptide epitope YMLD- LQPETT) was synthesized at Cytel according to standard f-MOC solid-phase synthesis methods and purified with high-pressure liquid chromatography. This epitope was selected because it possesses high-affinity binding to HLA-A2 in vitro I6 and because it is capable of inducing peptide-specific cytolytic T lymphocytes in HLA-A2 trans- genic mice. The influenza matrix peptide M15s_66 (GIL-
GFVFTL), which is highly immunogenic, was synthesized by Multiple Pepdde Systems (San Diego) and was used as a control to assess the specificity of cytolytic T lympho- cytes and the ability to respond to in vitro stimulation.
HLA phenotypic analysis of tumor cells. Flow cytom- etry of CaSki, HT-3, and MS-751 cells was performed with a FACScan 440 (Becton-Dickinson, Mountain View, Cal- if.) as previously descr ibedY Cells were stained with monoclonal antibody MA2.1 (purified from the MA2.1 hybridoma cell line [American Type Culture Collec- tion]) and monoclonal antibody W6/32 (Incstar, Stillwa- ter, Minn.), which recognize t he HLA-A2 molecule and class 1 molecules, respectively. An isotype-matched mu- rine (immunoglobulin G2a ) antibody served as a negative control.
Lymphokine-activated killer cells. Lymphokine-acti- vated killer cells were harvested from the buffy coats of healthy volunteers, as previously described, is These ceils were propagated in media supplemented with 1000 I U / m l human recombinant interleukin-2 (IL-2) (Cetus- Oncology Division, Chiron, Emeryville, Calif.).
Donors. Three HLA A2.1-positive cervical cancer pa- tients treated at the National Cancer Institute underwent leukapheresis 1 month after completion of treatment. Patient 1 had stage IB (bulky) adenosquamous carci- noma, padent 2 had stage IIB squamons cell carcinoma, and patient 3 had stage IB (bulky) adenocarcinoma. All three patients received whole-pelvic radiation therapy and brachytherapy; patients 1 and 3 also underwent adjuvant total abdominal hysterectomy.
Generation of cytolytic T lymphocytes. After in-
formed consent was obtained, peripheral blood mono- nuclear cells, consisting of macrophages and lympho- cytes, were harvested after leukapheresis of three HLA A2.1-positive cervical cancer patients treated at the Na- tional Cancer Institute. The cells were separated from peripheral blood by centrifugation over lymphocyte sep- aration medium (Organon-Teknika, Durham, N.C.) and then washed with Ca ++-, Mg ++-, and phenol-free phos- phate-buffered saline solution. The samples were stored as cryopreserved aliquots. Peptide-specific cytolytic T- lymphocyte lines were generated as follows. On day 0 peripheral blood mononuclear cells were plated at a concentration of 1.5 × 106 cells/ml in 24-well plates (2 ml/well) in Iscove's complete medium (Biofluids, Rock- ville, Md.) supplemented with 10% human AB serum, 50 m m o l / L HEPES (N-2-hydroxyethylpiperazine-N-2-eth- anesulfonic acid), 10 n m o l / L glutamine, 250 U / m l penicillin-streptomycin, and 0.25 m g / m l gentamicin. All cuhures were established in the presence of 10 txg/ml peptide except cultures with influenza matrix protein
M158_66, in which 1 ~xg/ml was added. On day 3, 24 I U / m l IL-2 was added to the cultures. Lymphocytes were restimulated weekly in the following manner: responder cells were harvested, washed once, and plated in 24-well

1588 Alexander et al. December 1996 Am J Obstet Gynecol
plates at a concentration of 2.5 × 105 cells/ml in lscove's complete medium. Autologous peripheral blood mono- nuclear cells were thawed and rinsed and then resus- pended at 1.5 to 10 × 106 cells/ml in Iscove's complete
medium pulsed with 10 b~g/ml E711_20 or 1 ~xg/ml M158_66 for 3 hours at 37 ° C. The peripheral blood mononuclear cells were irradiated at 3000 fads with a cesium 137-labeled source, washed in phosphate-buff- ered saline solution, and added to the responder cells at stimulator-to-responder ratios ranging from 5:1 to 25:1. The subsequent day 24 I U / m l IL-2 was added to these cultures. The activity of these cytolytic T lymphocytes was tested in cytotoxicity assays after at least 2 rounds (14 days) of peptide stimulation.
Cytotoxicity assays. Samples containing 1 × 10 6 target
cells were labeled with 200 Ci of chromimn 51 (New England Nuclear, Boston) in 0.5 ml of RPMI medium with 2% human AB serum for 2 hours at 37 ° C. Effectors and target ceils were incubated at 37 ° C at various effector/target ratios for 4 hours. The supernatants were collected and counted on a gamma counter. Percentage of specific lysis was calculated according to the following formula:
Specific lysis =
Sample radioactivity - Spontaneous radioactivity
Maximum radioactivity - Spontaneous radioactivity
× 100
Spontaneous radioactivity (in counts per minute) was obtained from targets incubated with 2% sodium dode- cyl sulfate. All assays were performed in triplicate. Spon- taneous 51Cr release did not exceed 30% of the maxi-
mum release, Analysis of peptide recognition by cytolytic T lympho-
eytes. To analyze pepride recognition, the T2 cell line was preincubated overnight at 37 ° C with 1 txg/ml peptide. The T2 cells were washed in RPMI 1640 with 2% AB human serum and used as a target cell in a 51Cr release cytotoxic assay.
Results
HLA typing. The peripheral blood mononuclear cells from all three cervical cancer patients manifested het- erozygous HLA haplotypes for allele A2 (HLA-A2).
Phenotypie analysis. By flow cytometry analysis, CaSki and MS-751 cells were recognized as HLA-A2 positive. No staining was detected on HT-3 cells, indicating that this cell line does not bear the HLA-A2 allele. MHC class I expression was detected in each cell line.
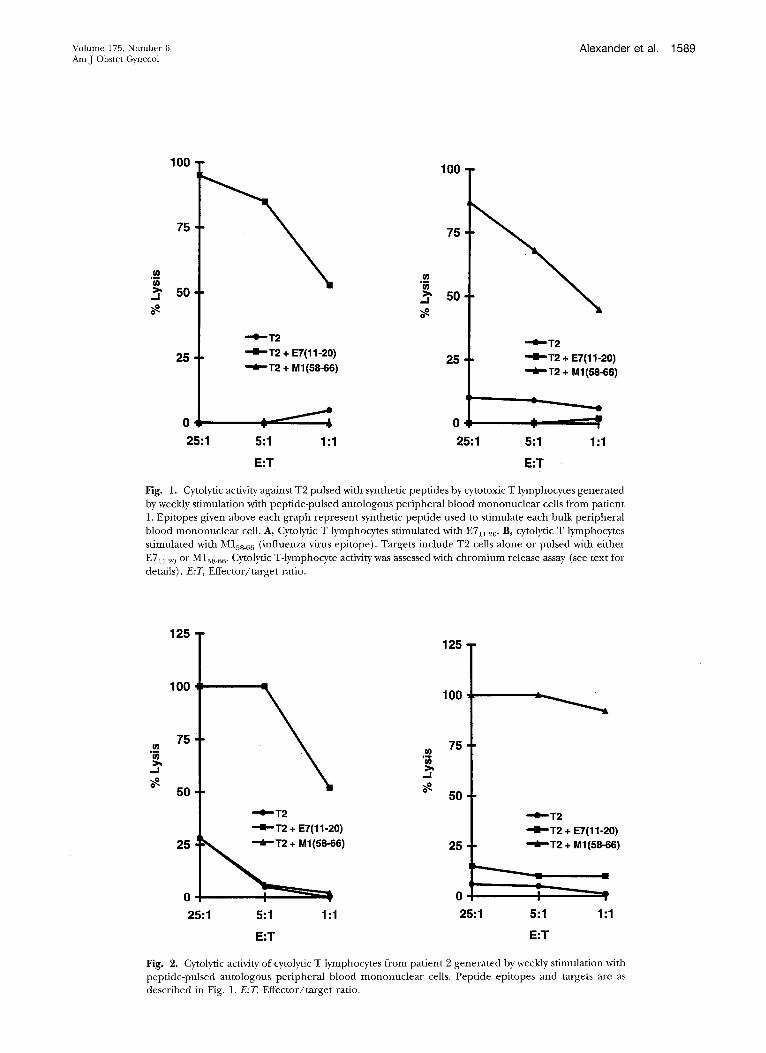
Specificity of eytolytic T lymphocytes generated with E7n.20. The E7~1_20 epitope was evaluated for its ability to induce specific in vitro cytolytic T-lymphocyte re- sponses with peripheral blood mononuclear cells from each patient. Chromium release cytotoxicity assays were
conducted after 4 and 2 weeks of cytolytic T lymphocyte
culture for patients 1 and 2, respectively. The antigen- processing defective T2 cells; which express HLA-A2
molecules but do not process intracellular antigens, were
used as targets. These cells were pulsed with E711.20 and then coincubated with cytolyric T lymphocytes. For pa- tients 1 and 2, cytolytic T lymphocytes induced with HPV
E711_20 lysed only T2 cells pulsed with E7al_z0 but not unpulsed target cells or cells with M158_66 (Figs. 1, A, and 2, A). Because virtually all humans are exposed to the
influenza matrix protein, we used M158_66 as a control epitope to assess each patient 's capacity to generate
anti-MIss_66 cytolytic T lymphocytes. Cytolyric T lympho-
cytes induced with M158_66 recognized only T2 cells
pulsed with M158_66 but not unpulsed target cells or cells
with E711_20 (Figs. 1, B, and 2, B). The experiment was repeated 1 week later (after 5 and 3 weeks of cytolyric T-
lymphocyte culture for patients 1 and 2, respectively)
and yielded similar results (data not shown). These
results demonstrate the efficiency and epitope specificity of the cytolyric T lymphocytes for inducing HLA-A2-
restricted target cell lysis. The E7~ 1_20-induced cytolyric T lymphocytes from patient 3 did not generate a specific
lytic response, although this patient 's M15s_66-induced cytolytic T lymphocytes specifically lysed T2 cells pulsed
with M158_66 (Fig. 3). Assessment of lymphokine-activated killer (nonspecific
cell lysis. To further assess the specificity of cytolytic T
lymphocyte lysis, lymphokine-acrivated killer cells, which
were propagated after IL-2 stimulation, were assessed for their ability to lyse various target cells. In these assays all
cells tested (including Daudi, T2, CaSki, HT-3, and MS-751 cells) were susceptible to lymphokine-activated
killer cell lysis. When the E711_2o-induced cytolytic T lymphocytes were reacted with Daudi cells, <17% lysis
was observed, indicating that nonspecific lyric activity was
negligible (data not shown). These results indicate that the lyric activity of the anti E71 >20 cyt01ytic T lymphocytes is epitope specific and HLA-A2 restricted.
Lysis of HPV-16-positive tumor cells by E711.20-in- duced cytolytic T lymphocytes. To assess whether the
cytolyric T lymphocytes induced with E711_20 could lyse a rumor cell harboring HPV-16, HLA-A2-positive CaSki cervical cancer cells were used as targets in a chromium-
release assay (Fig. 4). After 4 weeks of culture cytolyric T lymphocytes from patient 1 were incubated with the following target cells alone or pulsed with E71>20: T2, CaSki (which are HLA-A2 + [matched] and HPV-16+), MS-751 (HLA-A2 + and HPV-16 ), and HT-3 (HPV-16- and HLA-A2 ). There was an insufficient quantity of cytolytic T lymphocytes from patient 2 for this analysis. At an effector/target ratio of 25:1 the cytolytic T lympho- cytes induced with E7~1_20 demonstrated cytolysis of CaSki cells alone and lysis of both T2 cells and MS-751
Volume 175, Number 6 Alexander et al. 1589 Am J Obstet Gyaaecol
100
75
5o
25
- ' e " T 2
--D--T2 + E7(11-20)
~ T 2 + M1(58-66)
100
75
w
. j 50
25 --,e-,,T2
- • - ' T 2 + E7(11-20) T2 + M1(58-66)
o... ; + 0 ¢ ~ ' 25:1 5:1 1:1 25:1 5:1 1:1
E:T E:T
Fig. 1. Cytolytic activity against T2 pulsed with synthetic peptides by cytotoxic T lymphocytes generated by weekly stimulation with peptide-pulsed autologous peripheral blood mononuclear cells from patient 1. Epitopes given above each graph represent synthetic peptide used to stimulate each bulk peripheral blood mononuclear cell. A, Cytolytic T lymphocytes stimulated with E711_20. B, cytolytic T lymphocytes stimulated with M158_66 (influenza virus epitope). Targets include T2 cells alone or pulsed with either E711 20 or M158_66. Cytolytic T-lymphocyte activity was assessed with chromium release assay (see text for details). E:T, Effector/target ratio.
u~
.J
12s I 100
75
50
25
0
25:1
- ' e - -T2
- -B ' -T2 + E7(11-20) -66)
5:1 1:1
w >, -,I
o~
125
100
75
50
25
- ' e ' - T 2 - -B--T2 + E7(11-20) " ~ " T 2 + M1(58-66)
A
• i
25:1 5:1 1:1
E:T E:T
Fig. 2. Cytolytic activity of cytolytic T lymphocytes from patient 2 generated by weekly stimulation with peptide-pulsed autologous peripheral blood mononuclear cells. Peptide epitopes and targets are as described in Fig. 1. E:T, Effector/target ratio.

1590 Alexander et al. December 1996 Am J Obstet Gynecol
100
75
.--e--T2 m • ~ "-E--T2 + E7(11-20) ._~ ~>' 50 ~ T 2 + M1(58-66) u~
,,,J
100
75
50
25 ~ 25
0 0 25:1 5:1 1:1 25:1 5:1 1:1
E:T E:T
Fig. 3. Cytolytic activity of cytolytic T lymphocytes from patient 3 generated by weekly stimulation with peptide-pulsed autologous peripheral blood mononuclear cells. Peptide epitopes and targets are as described in Fig. 1. E:T, Effector/target ratio.
80
60
.o_ if) > ~ 4 0 - i
20
0 ¸
t t l A 01
Fig. 4. Cross-reactive lysis of CaSki cervical carcinoma ceils by peptide-specific cytolytic T-lymphocyte clones from patient 1 stimulated in vitro with HPV-16 E711_20 epitope. Lysis assay" was performed by incubating cytolytic T lymphocytes with indicated target cells alone or in presence of relevant (E7] 1-20) or irrelevant (M158~66) peptide. Lysis of unpulsed CaSki cells, which harbor HPV-16, indicates that these cells endogenously processed E71i_20 epitope for cytolytic T-lymphocyte recognition and lysis.
cells after they were pulsed with the relevant peptide.
E71>20-specific cytolytic T lymphocytes did no t demon- strate lysis of T2 cells pulsed with the irrelevant peptide
M158_66. Fur thermore , the E7n_2o-specific cytolytic T lymphocytes lysed ne i ther unpulsed T2 or MS-751 cells no r unpulsed or pulsed HT-3 cells. The cytolytic T
lymphocytes induced with M158_66 specifically lysed T2 cells pulsed with M 108_66 but had no appreciable effect on
the T2 cells a lone or on the T2 cells pulsed with E71>2o (Fig. B). In addition, M15s_66-specific cytolytic T lympho- cytes did no t lyse unpulsed CaSki cells or CaSki cells pulsed with E711_2o but were able to recognize and lyse CaSki cells pulsed with the relevant epitope (M15s.66). These results demonst ra ted that cytolytic T lymphocyte recogni t ion is epitope d e p e n d e n t and HLA-A2 re- stricted. Importantly, because the E7n_20-specific cyto-

Volume 175, Number 6 Alexander et al. 1591 Am J Obstet Gynecol
50
40
• = ao
o. ~ 2 0
10 -
t A
CO
Fig. 5. Epitope-specific lysis by cytolytic T-lymphocyte clones from patient 1 stimulated in vitro with M158_66 epitope. Lysis assay was performed by incubating cytolytic T lymphocytes with indicated target cells alone or in presence of relevant (M158_66) or irrelevant (E7n_20) peptide.
lytic T lymphocytes lysed unpulsed CaSki cells, these results indicate that E7n_20 is endogenously processed and presented onto the surface of CaSki cells for recognition and lysis by the anti-E71>20 cytolytic T lymphocytes.
C o m m e n t
In this study we demonstrated that a synthetic peptide epitope of HPV-16 E7 can elicit a specific HLA-restricted cytolytic T lymphocyte response in peripheral blood mononuclear cells from patients with cervical cancer. Several recent reports concur with our findings and provide additional support for further investigations in this area. For example, Kawakami et al. m' 2o have identi- fied several HLA-A2-restricted epitopes of two distinct tumor rejection antigens that are consistently expressed in melanomas and that can elicit specific cytolytic T lymphocytes capable of recognizing and lysing tumors. Cytolytic T lymphocytes with specific ant imelanoma re- activity have also been generated with an HLA-A1- restricted MAGE-1 synthetic epitope, m By selecting high- affinity MHC binding epitopes from another known tumor-associated melanoma antigen, Cells et al. 22 was also able to generate cytolytic T lymphocytes that could specifically recognize and lyse tumor cells in vitro. This strategy clearly has relevance for cervical cancer because vaccination with an HPV-16 E7 epitope has been shown to confer protection against lethal tumor inoculation in mice. 9 Combined with our data, these findings illustrate the feasibility of peptide epitope-based vaccines or anti- gen-specific adoptive immunotherapies for the preven- tion and treatment of cervical cancer.
The recent work of Ressing et al. 23 also supports that peptide epitopes of HPV-16 E7 may be effective in generating specific cytolytic T lymphocytes. By use of peripheral blood mononuclear ceils from healthy do- nors, antigen-presenting cells were generated with Staph- ylococcus aureus Cowan I, and the resulting ceils were
loaded with peptide in the presence of [32-microglobulin.
This culturing technique also resulted in the induction of peptide-specific cytolytic T lymphocytes and empha- sizes that these epitopes are effective under substantially
different culturing conditions. In addition, induction of cytolytic T lymphocytes from the heavily irradiated cervi-
cal cancer patients in this study suggests that the capacity
to recognize these epitopes is retained even after sub- stantial bone marrow ablation.
The effectiveness of the E7n_z0-induced cytolytic T lymphocytes to recognize and lyse unpulsed CaSki cells
represents evidence that the E711_20 is an epitope that may be exploited for therapeutic applications. The deg- radation of intracellular proteins into peptide epitopes is
a vital metabolic function of normal, vitally infected, or tumor cells. Because a target cell presents on its surface
cleaved protein fragments complexed with class I MHC molecules, cytolytic T lymphocytes can monitor the tar-
get cell's surface and initiate lysis of a virally infected or tumor cell. Generation of cytolytic T lymphocytes capa-
ble of epitope-specific recognition and lysis depends in large part on the frequency of precursor cytolytic T
lymphocytes available for clonotypic expansion. Differ- ences in precursor cytolytic T-lymphocyte frequencies
from one patient to another might explain the different clinical consequences that result from exposure to HPV-
16. Therefore, when patients are vaccinated with "immu- nodominant" epitopes, cytolytic T-lymphocyte precursor frequencies might be enhanced to permit rejection of virally infected cells.
The inability to generate HPV-16, E7n_20-specific cyto- lytic T lymphocytes in one of the three patients suggests t h a t a single peptide epitope alone may not be effective in all HLA-matched patients. A memory response may
also be involved in the generation of epitope-specific cytolytic T lymphocytes and may explain why peptide-

1592 Alexander eta[. December 1996 Am J Obstet Gynecol
specific lysis occurred when cytolytic T lymphocytes f rom
pat ient 3 were induced with M158_66 but did not occur
when cytolytic T lymphocytes were induced with E711-90.
Recogni t ion of the E711_20 epi tope may reqUire a larger
n u m b e r of cytolytic T-lymphocyte precursor cells to
permi t clonotypic expansion with IL-2, or E711 20 may be
subdominant in some patients. The use of a subdomi-
nant epi tope to immunize against a tumor is analogous
to the clinical situation in which women with invasive
cervical cancer are unlikely to have genera ted a strong
i m m u n e response against any dominan t HPV cytolytic T
lymphocyte epitope. If patients are anergic to HPV
epitopes, lack of T-cell reactivity may lead to progressive
disease. Vaccination with a strong adjuvant a long with a
cytolytic T lymphocyte epi tope pept ide might pr ime
cytolytic T lymphocytes against a weakly immunogen ic
epi tope and result in clearance of HPV.
Currently, epitope-based immuno the rapy for cervical
cancer faces several impor tan t limitations. For instance,
class I H L A haplotypes are extremely diverse. Initial
efforts to develop epi tope vaccines have focused primar-
ily on class I HLA-A2-restr icted models because this
allele is the one most commonly expressed. 24 It is con-
ceivable that vaccines will ultimately contain a library of
epi topes capable of b inding to a variety of HLA class I
molecules. The use of mult iple epitopes might also
augment tumor reject ion because several different im-
munological ly relevant peptides may result f rom the
lysosomal cleavage of a tumor-associated antigen. It will
also be necessary to genera te immunological ly relevant
pept ide epitopes f rom other HPV types that are associ-
ated with anogeni ta l neoplasias.
Clinically effective tumor vaccines will probably be of
greatest use for the prevention, ra ther than the treat-
ment, of virally induced neoplasias. Evidence of an
effective peptide-based preventive immuniza t ion is pro-
vided by exper iments in mice in which epitopes of E7
induced lasting immuni ty to subsequent chal lenge with
HPV-16 t ransformed tumor cells. 9 The protective effects
of a peptide-based vaccine has also been demons t ra ted
against hepatitis B virus in humans. It is conceivable that
synthetic HPV epi topes might be used for adoptive
immunotherapy , where specific cytolytic T lymphocytes
f rom cervical cancer patients are expanded ex vivo and
then .reinfused into the patient. However, cytolytic T
lymphocyte recogni t ion of tumor cells harbor ing HPV
will requi re that the cell 's epi tope present ing machinery
remains intact, a process that can be subverted dur ing
mal ignant degenera t ion . 25 In the future the develop-
men t of a prophylactic or therapeutic vaccine for cervical
cancer may offer an attractive and cost-effective immuno-
logic approach to reduce the need for expensive screen-
ing and surveillance prevent ion programs and substan-
tially decrease the worldwide morbidi ty f rom this disease.
REFERENCES
1. Pisani P, Parkin DM, Ferlay J. Estimates of the worldwide mortality from eighteen major cancers in 1985: implications for prevention and projections of future burden. Int J Cancer 1993;55:891-903.
2. Bosch FX, Manes MM, Munoz N, et aI. Prevalence of human papillomavirus in cervical cancer: a worldwide perspective. J Natl Cancer Inst 1995;87:796-802.
3. Crook T, MorgensternJP, Crawford L, Banks L. Continued expression of HPV-16 E7 protein is required for mainte- nance of the transformed phenotype of cells co-transformed by HPV-16 plus EJ-ras. EMBO J 1989;8:513-9.
4. Munger K, Scheffner M, Huibregtse JM, HoMey PM. Inter- actions of HPV E6 and E7 oncoproteins with tumour suppressor gene products. Cancer Surv 1992;12:197-217.
5. Nasiell K, Roger V, Nasiell M. Behavior of mild cervical dysplasia during long-term follow-up. Obstet Gynecol 1986; 67:665-9.
6. Maiman M, Fruchter RG, Serur E, RemyJC, Feuer G, Boyce J. Human immunodeficiency virus infection and cervical neoplasia. Gynecol Oncol 1990;38:377-82.
7. Shamanin V, Glover M, Rausch C, et al. Specific types of hmnan papillomavirus found in benign proliferations and carcinomas of the skin in immunosuppressed patients. Cancer Res 1994;54:4610-3.
8. Madden DR, Gorg a JC, Strominger JL, Wiley DC. The structure of HLA-B27 reveals nonamer self-peptides bound in an extended conformation. Nature 1991;353:321-5.
9. Feltkamp MC, Sinits HL, Vierboom MP, et al. Vaccination with cytotoxic T lymphocyte epitope-containing peptide protects against a tumor induced by human papillomavirus type 16-transformed cells. EurJ Immunol 1993;23:2242-9.
10. Schwarz E, Freese UK, Gissmann L, et al. Structure and transcription of human papillomavirus sequences in cervi- cal carcinoma cells. Nature 1985;314:111-4.
11. Kast WM, Brandt RM, Drijfhout JW, Melief CJ. Human leukocyte antigen-A2.1 restricted candidate cytotoxic T lym- phocyte epitopes of human papillomavirus type 16 E6 and E7 proteins identified by using the processing-defective human cell line T2. J Immunother 1993;14:115-20.
12. Yee C, Krishnan-Hewlett I, Baker CC, Schlegel R, HoMey PM. Presence and expression of human papillomavirus sequences in human cervical carcinoma cell lines. Am J Pathol 1985;119:361-6.
13. Pater MM, Pater A. Human papillomavirus types 16 and 18 sequences in carcinoma cell lines of the cervix. Virolog T 1985;145:313-8.
14. Salter RD, Cresswell P. Impaired assembly and transport of HLA-A and -B antigens in a mutant TxB cell hybrid. EMBO . J 1986;5:943-9.
15. Hopkins KA. Basic microlymphocytotoxicity test. In: ASHI laboratory manual. Lenexa (KS): American Society for Histocompatibility and Immunogenetics, 1990:195-201.
16. Kast WM, Brandt RM, Sidney J, et al. Role of HLA-A motifs in identification of potential CTL epitopes in human pap- illomavirus type 16 E6 and E7 proteins. J Immunol 1994; 152:3904-12.
17. Topalian SL, Muul LM, Solomon D, Rosenberg SA. Expan- sion of human tumor infiltrating lymphocytes for use in immunotherapy trials. J lmmunol Methods 1987;102:127- 41.
18. Rosenberg SA, Lotze MT, Muul LM, et al. A progress report on the treatment of 157 patients with advanced cancer using lymphokine-activated killer cells and interleukin-2 or high- dose interleukin-2 alone. N Engl J Med 1987;316:889-97.
19. Kawakami Y, Eliyahu S, Sakaguchi K, et al. Identification of the immunodominant peptides of the MART-1 human melanoma antigen recognized by the majority of HLA-A2- restricted tumor infiltrating lymphocytes. J Exp Med 1994; 180:347-52.
20. Kawakami Y, Eliyahu S, Jennings C, et al. Recognition of multiple epitopes in the human melanoma antigen gpl00

Volume 175, Number 6 Alexander et al, 1593 AmJ Obstet Gynecol
by tumor-infiltrating T lymphocytes associated with in vivo tumor regression. J Immunol 1995;154:3961-8.
21. Salgaller ML, Weber JS, Koenig S, Yannelli JR, Rosenberg SA. Generation of specific anti-melanoma reactivity by stim- ulation of human tumor-infiltrating lymphocytes with MAGE-1 synthetic peptide. Cancer Immunol Immunother 1994;39:105-16.
22. Celis E, Tsai V, Crimi C, et al. Induction of anti-tumor cytotoxic T lymph0cytes in normal humans using primary cultures and synthetic peptide epitopes. Proc Natl Acad Sci U S A 1994;91:2105-9.
23. Ressing ME, Sette A, Brandt RM, et al. Human CTL epitopes
24.
25.
encoded by human papillomavirus type 16 E6 and E7 identified through in vivo and in vitro immunogenicity studies of HLA-A*0201-binding peptides. J Immunol 1995; 154:5934-43. Lee TD. Distribution of HLA antigens in North American caucasions, North American blacks and orientals. In: Lee J, editor. The HLA system: a new approach. New York: Spring- er-Verlag, 1990:141-78. Restifo NP, Esquivel F, Kawakami Y, et al. Identification of human cancers deficient in antigen processing. J Exp Med 1993;177:265-72.
Availability of Journal back issues As a service to our subscribers, copies of back issues of the Amer ican Journa l of Obstetrics
and Gynecology for the preced ing 5 years are main ta ined and are available for purchase f rom Mosby until inventory is deple ted at a cost of $14.00 per issue. The following quantity discounts are available: 25% off on quantit ies of 12 to 23 and one third off on quantit ies of 24 or more. Please write to Mosby-Year Book, Inc., Subscription Services, 11830 Westline Industrial Drive, St. Louis, MO 63146-3318, or call (800)453-4351 or (314)453-4351 for informat ion on availability of particular issues. If back issues are unavailable f rom the publisher, photocopies of comple te issues may be purchased f rom UMI, 300 N. Zeeb Road, Ann Arbor, MI 48106-1346. Tel.: (313)761-4700.