genotoxical, teratological and biochemical effects of
TRANSCRIPT

139Reprod. Nutr. Dev. 46 (2006) 139–156© INRA, EDP Sciences, 2006DOI: 10.1051/rnd:2006007
Original article
Genotoxical, teratological and biochemical effects of anthelmintic drug oxfendazole Maximum Residue
Limit (MRL) in male and female mice
Aida EL- MAKAWY, Hasnaa A. RADWAN, Inas S. GHALY, A. Abd EL-RAOUF
Cell Biology Department, National Research Center, Dokki, Giza, Egypt
(Received 30 May 2005; accepted 16 November 2005)
Abstract – Oxfendazole, methyl-5 (6)-phenylsulfinyl-2-benzimidazole carbamate, is a member ofthe benzimidazole family of anthelmintics. Anthelmintic benzimidazoles are widely used in meatproducing animals (cattle, sheep and pigs) for control of endoparasites. The extensive use ofveterinary drugs in food-producing animals can cause the presence of small quantities of the drugresidues in food. Maximum residue limit or “MRL” means the maximum concentration of residueresulting from the use of a veterinary medicinal product which may be legally permitted recognizedas acceptable in food. The FAO/WHO Expert Committee on Food Additives (1999) evaluations oftoxicological and residue data, reported that oxfendazole (MRL) has toxicological hazards on humanhealth. The toxicity of oxfendazole (MRL) was tested in male and female mice and their fetuses.Chromosomal aberrations, teratological examination and biochemical analysis were the parametersused in this study. The results show that oxfendazole MRL induced a mutagenic effect in all testedcell types. Also, oxfendazole exhibit embryotoxicity including teratogenicity. The biochemicalresults show that oxfendazole induced a disturbance in the different biochemical contents of all testedtissues. So, we must increase the attention paid to the potential risk of oxfendazole residues in humanbeings and should stress the need for careful control to ensure adherence to the prescribed withdrawaltime of this drug.
anthelmintic / oxfendazole / maximum residue limit / genotoxicity / embryotoxicity /teratogenicity / biochemical changes
1. INTRODUCTION
Antiparasitic agents represent an impor-tant class in veterinary medicine. Amongthem, anthelmintic benzimidazoles are widelyused in meat producing animals (cattle,sheep and pigs) for the control of endopar-asites. Some members of this class of drugshave teratogenic and embryotoxic effects in
animal species. In order to ensure humanfood safety, the European Union has setmaximum residue limits (MRL) for benz-imidazoles and their metabolites in animalproducts [1]. A Maximum Residue Limit or“MRL” means the maximum concentrationof residue resulting from the use of a veter-inary medicinal product which may belegally permitted recognized as acceptable
* Corresponding author: [email protected]
Article published by EDP Sciences and available at http://www.edpsciences.org/rnd or http://dx.doi.org/10.1051/rnd:2006007

140 A. El-Makawy et al.
in food. This limit is either based on the typeand amount of residue considered to bewithout any toxicological hazard for humanhealth as expressed by the acceptable dailyintake (ADI), or on the basis of a temporaryADI that utilizes an additional safety factor[2]. Liver, kidney, muscle and fat are thetarget tissues for residue control. MRLrange from 10 to 1000 µg·kg–1 dependingon compound and matrix [1]. FAO/WHOExpert Committee on Food Additives [3]convened to evaluate the safety of residuesof certain veterinary drugs in food and torecommend maximum levels for such resi-dues. The Committee’s evaluations of tox-icological and residue data, reported thatoxfendazole (MRL) has toxicological hazardson human health. Oxfendazole, methyl-5 (6)-phenylsulfinyl-2-benzimidazole carbamate,is a member of the benzimidazole family ofanthelmintics. It is the sulphoxide metabo-lite of fenbendazole and it is possible thatfenbendazole owes much of its efficacyspectrum to oxfendazole. The probenzimi-dazole, febantel, when it is metabolized,also becomes fenbendazole, and finallyoxfendazole, which is believed to be theactive metabolite. The compound oxfenda-zole reaches peak blood levels more slowlyfollowing absorption from the intestinaltract than many of the older benzimida-zoles, thereby maintaining effective con-centrations for a longer time in both the serumand the intestinal tract increases its efficacy[4]. The heat stability of the anthelminticoxfendazole in water, cooking oil and asincurred residues in cattle liver was inves-tigated. It was found that cooking did notdestroy residues although it may affect thepoint of equilibrium between oxfendazole,oxfendazole sulphone, fenbendazole andsome other metabolites in incurred tissue[5]. The extensive use of veterinary drugsin food-producing animals can cause thepresence of small quantities of the drug res-idues in food. Certain drugs can be carriedover into the milk of lactating cows ordeposited in the tissues of an animal intendedfor slaughter [6, 7]. In recent years, therehas been widespread and increasing con-
cern that drugs, as well as environmentalchemicals, may present a potential hazardto mankind by causing gene mutations orchromosome aberrations. Genetic altera-tions in somatic tissues can have a numberof immediate effects upon the cellsinvolved, including cell death or transfor-mation into malignancy. Genetic alterationin germ cells leads to reproductive failureor genetic disorder in subsequent genera-tions that include heritable mutations low-ering the reproductive output of affectedpopulation [8]. So, the purpose of the presentstudy was to evaluate the competence ofoxfendazole maximum residue limit (MRL)on the incidence of genetic alteration inmale and female mice and their teratogeniceffect in embryos of treated parents.
2. MATERIALS AND METHODS
2.1. Drug
Tyvert, Oxfendazole 2.265% is a broadspectrum worm drench for cattle and sheep,manufactured by Pharmacia au Upjohn,Purs-Belgium. Oxfendazole CAS No.53716-50-0. The dose equal to Oxfenda-zole maximum residue limit in liver(1000 µg·kg–1) was used. Each animal gav-ages orally 0.01 mL daily.
2.2. Assay of genotoxicity in males
Twenty adult male Swiss mice weighing(20–30 g) obtained from the animal houseof the national research center were used toevaluate the oxfendazole (MRL) genotox-icity in male cells. The animals were ran-domly divided into two groups and hadfood and water ad libitum. The first groupwas given distilled water orally and servedas the control. The second group was orallygiven the dose of oxfendazole (MRL) dailyfor eight weeks. At the end of the time ofdrug administration, all animals wereinjected IP with a colchicine solution. Twohours later, the animals were sacrificed bycervical dislocation and the chromosomes

Toxicity of anthelmintic drug oxfendazole residues 141
of bone marrow cells and spermatocyteswere prepared according to Yosida andAmano [9], Brewen and Preston [10].
2.3. Assay of genotoxicity in females
Adult male and adult virgin females ofSwiss albino mice were maintained on feedand water ad libitum in the animal house ofthe national research center. Both male andfemale animals were divided into twogroups. The animals of the first group weregiven distilled water orally and served asthe control animals. The animals of the sec-ond group were orally given a dose ofoxfendazole (MRL) for four weeks. Afterthat, males and females were caged by ratio(1:1) according to the following groups:
Group I: control males and females. Group II: treated males and control females.Group III: control males and treatedfemales. Group IV: treated males and females.Successful mating was determined by
the presence of vaginal plugs on the nextmorning which was designed as day zero ofpregnancy. Drug oral administration in thesame manner continued until day 18 ofpregnancy.
2.4. Teratological examination
On day 18 of gestation, ten pregnantfemales of each group were sacrificed bycervical dislocation. The uterine horns wereopened and the numbers of alive, dead andresorbed fetuses were recorded accordingto Gleich and Frohberg [11]. The bodyweights of live fetuses were recorded just afterdissection. Also the fetuses were externallyexamined to determine any deformities. Forskeletal examination, fetal skeletons wereprepared and stained in Alizarin red-S accord-ing to the method described by Weesner [12].After staining, the skeletons were examinedunder the dissecting binocular microscope todetect any malformation in the bones of theaxial and appendicular skeletons.
2.5. Chromosome preparation
The chromosomes of bone marrow cellsof pregnant females were prepared by usingthe methodology of Yosida and Amano [9]and the chromosomes of fetal cells wereprepared according to the method ofRomagnano et al. [13].
The slides were stained with 10%Giemsa stain and 100 good metaphasespreads of each cell types for all animalswere examined microscopically to analyze thedifferent types of chromosomal aberrations.
2.6. Biochemical analysis
Total content of DNA, RNA, protein andenzymes in different tissues (liver, kidney,testes and embryos) of male and femalemice were determined. DNA contents weredetermined according to Dische [14] usingthe diphenylamine method; DNA givingblue color was read using a spectrophotom-eter at 600 nm. RNA content was deter-mined by using the Orcinol reagent methodaccording to Schneider [15] and read at660 nm. Protein content was determinedaccording to Gomall et al. [16] and read at545 nm. Enzyme activities of the liver(GOT and GPT) were determined accord-ing to Reitman and Frankel [17] at 545 nm.
2.7. Statistical analysis
Data of biochemical analysis andcytogenetical examination of males wereanalyzed by a paired- t -test. The data of thefetuses morphological examination andchromosome aberrations were analyzedstatistically by using one way analysis ofvariance (ANOVA).
3. RESULTS
3.1. Effect of oxfendazole maximum residue limit (MRL) in males
The data of the present study indicatedthat oxfendazole (MRL) oral administra-tion of male mice for eight weeks inducedchromosome aberrations in bone marrowand spermatocyte cells as shown in Figures 1

142 A. El-Makawy et al.
and 2. Table I and Figure 3 illustrate themean values and standard deviations of dif-ferent types of chromosomal aberrations inbone marrow cells of control and oxfenda-zole (MRL) treated male mice. The datashowed that oxfendazole caused a statisti-cally significant increase in the mean valuesof the gaps, endomitosis, and centric fusionsat (P ≤ 0.05) when compared with the con-trol. The results also revealed that oxfend-azole showed a statistically significantincrease in the mean values of fragments,deletions, and end to end associations, poly-ploidy and total chromosomal aberrations
at (P ≤ 0.01) than the control. However, thestatistical analysis of the other individualchromosomal aberrations showed non sig-nificant differences between the mean val-ues of the different treated groups and thecontrol. Table II and Figure 4 demonstratethat oxfendazole (MRL) treated male miceshowed a statistically significant increasein the mean values of autosomal univalent,x-y univalent, aneuploidy and total chro-mosomal aberrations at (P ≤ 0.01) whencompared with the control group, whereas,translocation, ring and polyploidy showednon significant differences.
Figure 1. Different types ofchromosomal aberrations indu-ced by oxfendazole (MRL) inbone marrow cells: (A) gap (g)and break (b) (B) deletion(C) end to end association(D) centric fusion (E) poly-ploidy (F) endomitosis.

Toxicity of anthelmintic drug oxfendazole residues 143
3.2. Effect of oxfendazole maximum residue limit (MRL) in females
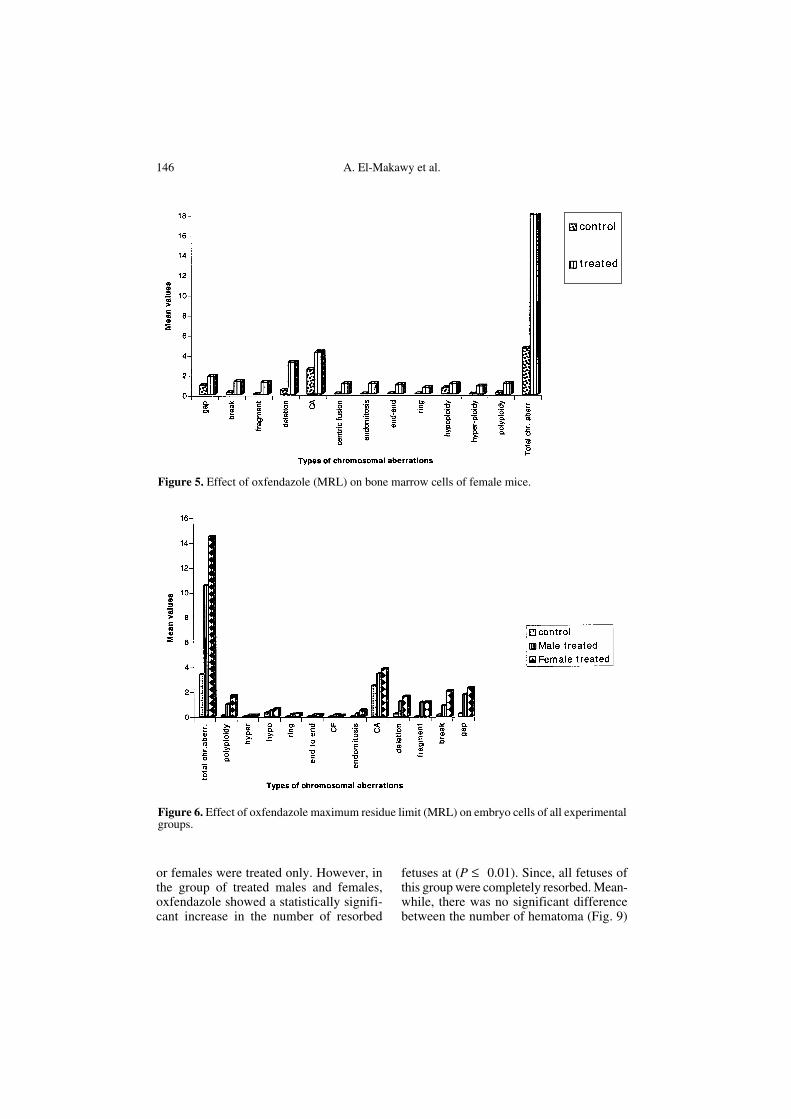
The mean frequencies of different indi-viduals and total chromosomal aberrationsinduced in female bone marrow cells of thetreated group and the standard deviationsare presented in Table III and Figure 5. Thedata showed that oxfendazole caused a sta-tistically significant increase in the meanvalues of endomitosis, centric fusion, frag-ments, deletions, end to end associations,polyploidy and total chromosomal aberra-tions at (P ≤ 0.01) when compared with thecontrol. Meanwhile, it showed a statisti-cally significant increase in the mean values
of the other types of chromosomal aberra-tions at (P ≤ 0.05).
3.3. Effect of oxfendazole maximum residue limit (MRL) on fetal cells
The data of Table IV and Figure 6 rep-resent the mean values of different individ-ual and total chromosomal aberrationsinduced by oxfendazole (MRL) in embryocells. The results indicate that oxfendazoleoral administration for males or femalesonly induced a statistically significantincrease in all individual and total chromo-some aberrations in the cells of theirembryos at (P ≤ 0.01) when compared withthe embryos of control males and females.
Figure 2. Different types ofchromosomal aberrations indu-ced by oxfendazole (MRL) inspermatocytes: (A) x-y univa-lent (B) x-y univalent (C) auto-somal univalent (D) hypoploidy(E) polyploidy.

144 A. El-Makawy et al.
Ta
ble
I. T
he m
ean
valu
es o
f diff
eren
t chr
omos
omal
abe
rrat
ions
indu
ced
by o
xfen
dazo
le m
axim
um re
sidu
e lim
it (M
RL
) in
bone
mar
row
cel
ls o
f mal
e m
ice.
Gro
ups
No. of
anim
als
Gap
Bre
akFr
agm
ent
Del
etio
nC
entr
omer
ic
atte
nuat
ion
End
omito
sis
Cen
tric
fu
sion
End
to
end
Rin
gH
ypo-
ploi
dyH
yper
-pl
oidy
Poly
-pl
oidy
Tota
l ch
rom
osom
alab
erra
tions
Con
trol
10M
± S
D0.
90±
0.8
80.
20±
0.4
20.
00±
0.0
00.
40±
0.5
22.
50±
0.8
50.
00±
0.0
00.
00±
0.0
00.
00±
0.0
00.
00±
0.0
00.
50±
0.7
10.
00±
0.0
00.
20±
0.4
24.
60±
1.0
7
Tre
ated
10M
± S
D1.
80*
± 0
.79
1.00
*±
0.8
20.
90**
± 0
.74
3.10
**±
1.6
04.
00±
1.6
30.
70*
± .8
20.
60*
± 0
.70
0.90
**±
0.7
40.
20±
0.4
20.
60±
0.9
70.
60±
0.9
71.
00**
± 0
.82
15.1
0**
± 2
.47
* Si
gnif
ican
t dif
fere
nce
at (
P ≤
0.0
5).
** S
igni
fica
nt d
iffe
renc
e at
(P
≤ 0
.01)
.
Tabl
e II
. The
mea
n va
lues
of d
iffe
rent
chr
omos
omal
abe
rrat
ions
indu
ced
by o
xfen
dazo
le m
axim
um re
sidu
e lim
it (M
RL
) in
sper
mat
ocyt
es o
f mal
e m
ice.
Gro
ups
No.
of a
nim
als
Aut
osom
alun
ival
ent
x-y
univ
alen
tT
rans
loca
tion
Rin
gH
ypop
loid
yH
yper
ploi
dyPo
lypl
oidy
Tota
l chr
omos
omal
ab
erra
tions
Con
trol
100.
80 ±
0.7
90.
40 ±
0.7
00.
00 ±
0.0
00.
00 ±
0.0
00.
90 ±
0.8
80.
00 ±
0,0
00.
00 ±
0.0
02.
10 ±
0.5
3
Tre
ated
103.
00**
± 0
.94
3,10
** ±
1.4
50.
20 ±
0.4
20.
30 ±
0.4
83.
70**
± 0
.82
1.60
** ±
0.8
40.
60 ±
0.8
412
.50*
* ±
1.7
8
* Si
gnif
ican
t dif
fere
nce
at (
P ≤
0.0
5).
** S
igni
fica
nt d
iffe
renc
e at
(P
≤ 0
.01)
.
Tabl
e II
I. T
he m
ean
valu
es o
f diff
eren
t chr
omos
omal
abe
rrat
ions
indu
ced
by o
xfen
dazo
le m
axim
um r
esid
ue li
mit
(MR
L) i
n bo
ne m
arro
w c
ells
of f
emal
e m
ice.
Gro
ups
No. of
anim
als
Gap
Bre
akFr
agm
ent
Del
etio
nC
entr
omer
ic
atte
nuat
ion
End
o-m
itosi
sC
entr
ic
fusi
onE
nd to
en
dR
ing
Hyp
o-pl
oidy
Hyp
er-
ploi
dyPo
ly-
ploi
dyTo
tal
chro
mos
omal
aber
ratio
ns
Con
trol
10M
± S
D1.
00±
089
0.66
± 0
.81
0.00
± 0
.00
0.50
± 0
.55
2.55
± 0
.85
0.00
± 0
.00
0.00
± 0
.00
0.00
± 0
.00
0.00
± 0
.00
0.60
± 0
.84
0.00
± 0
.00
0.25
± 0
.52
5.56
± 1
.97
Tre
ated
10M
± S
D1.
80*
± 0
.79
1.30
*±
0.9
51.
20**
± 0
.92
3.20
**±
1.4
34.
20*
± 1
.43
1.00
**±
0.9
41.
00**
± 0
.67
0.90
**±
0.7
40.
60*
± 0
.70
1.00
± 1
.05
0.80
*±
0.9
21.
10**
± 0
.74
17.9
0**
± 2
.42
* Si
gnif
ican
t dif
fere
nce
at (
P ≤
0.0
5).
** S
igni
fica
nt d
iffe
renc
e at
(P
≤ 0
.01)
.

Toxicity of anthelmintic drug oxfendazole residues 145
3.4. Teratogenic effect of oxfendazole maximum residue limits (MRL)
3.4.1. External morphological examination
The mean values of different teratogenicvariables are tabulated in Table V and Fig-
ure 7. The results showed that oxfendazole(MRL) induced a statistically significantdecrease in the number of live fetuses in allgroups at (P ≤ 0.01) when compared withthe control, whereas, it induced a non sig-nificant increase in the number of resorbedfetuses (Fig. 8) in the groups in which males
Figure 3. Effect of oxfendazole (MRL) on bone marrow cells of male mice.
Figure 4. Effect of oxfendazole (MRL) on spermatocytes of male mice.
146 A. El-Makawy et al.
or females were treated only. However, inthe group of treated males and females,oxfendazole showed a statistically signifi-cant increase in the number of resorbed
fetuses at (P ≤ 0.01). Since, all fetuses ofthis group were completely resorbed. Mean-while, there was no significant differencebetween the number of hematoma (Fig. 9)
Figure 5. Effect of oxfendazole (MRL) on bone marrow cells of female mice.
Figure 6. Effect of oxfendazole maximum residue limit (MRL) on embryo cells of all experimentalgroups.

Toxicity of anthelmintic drug oxfendazole residues 147
Tab
le I
V. T
he m
ean
valu
es o
f dif
fere
nt c
hrom
osom
al a
berr
atio
ns in
duce
d by
oxf
enda
zole
max
imum
res
idue
lim
it (M
RL
) in
embr
yo c
ells
of d
iffe
rent
expe
rim
enta
l gro
ups.
Gro
ups
No. of
embr
yos
Gap
Bre
akFr
agm
ent
Del
etio
nC
entr
omer
ic
atte
nuat
ion
End
o-m
itosi
sC
entr
ic
fusi
onE
nd to
en
dR
ing
Hyp
o-pl
oidy
Hyp
er-
ploi
dyP
oly-
ploi
dyTo
tal
chro
mos
omal
ab
erra
tions
Con
trol
50M
± S
D0.
22c
± 0
.51
0.10
c±
0.3
60.
00b
± 0
.00
0.20
c±
0.4
52.
48b
± 0
.95
0.00
c±
0.0
00.
00b
± 0
.00
0.00
b±
0.0
00.
00b
± 0
.00
0.30
b±
0.5
80.
00b
± 0
.00
0.10
c±
0.3
03.
38c
± 1
.60
Mal
e tr
eate
d50
M ±
SD
1.74
b±
0.9
40.
86b
± 1
.01
1.12
a±
1.1
21.
20b
± 1
.18
3.40
a±
1.4
60.
22b
± 0
.46
0.14
a±
0.3
50.
12a
± 0
.33
0.20
a±
0.4
00.
48ab
± 0
.71
0.12
a±
0.3
31.
00b
± 0
.90
10.5
4b±
3.7
2
Fem
ale
trea
ted
50M
± S
D2.
28a
± 0
.90
2.02
a±
0.8
71.
30a
± 0
.54
1.60
a±
0.9
33.
80a
± 1
.29
0.52
a±
0.6
10.
10a
± 0
.30
0.14
a±
0.3
50.
24a
± 0
.48
0.66
a±
0.7
50.
12a
± 0
.33
1.7a
± 0
.95
14.4
6a±
2.7
7
Mea
ns w
ith
diff
eren
t let
ters
wit
hin
each
col
umn
are
sign
ific
ant a
t 5%
leve
l.
Tab
le V
. Eff
ect o
f ox
fend
azol
e m
axim
um r
esid
ue li
mit
(MR
L)
on e
mbr
yos
of d
iffe
rent
exp
erim
enta
l gro
ups.
Gro
ups
No.
of
fem
ales
Aliv
e em
bryo
sR
esor
bed
embr
yos
Hae
mat
oma
Mea
n bo
dy w
eigh
t
Con
trol
10M
± S
D9.
40 ±
1.2
6 a
0.20
± 0
.42
b0.
50 ±
0.5
3 ab
2.70
8 ±
0.3
24 a
Mal
e tr
eate
d10
M ±
SD
7.10
± 3
.03
b1.
50 ±
2.0
1 b
0.80
± 0
.79
a1.
523
± 0
.135
b
Fem
ale
trea
ted
10M
± S
D7.
70 ±
1.9
7 ab
1.40
± 1
.15
b1.
00 ±
1.1
5 a
1.13
7 ±
0.1
15 c
Mal
e +
fem
ale
trea
ted
10M
± S
D0.
00 ±
0.0
0 c
8.70
± 2
.06
a0.
00 ±
0.0
0 a
0.00
± 0
.00
d
Mea
ns w
ith
diff
eren
t let
ters
wit
hin
each
col
umn
are
sign
ific
ant a
t 5%
leve
l.

148 A. El-Makawy et al.
in all treated groups and the control. How-ever, oxfendazole (MRL) showed a statis-tically significant decrease in the fetal bodyweights of all treated groups at (P ≤ 0.01)when compared with the untreated group.
3.4.2. Skeletal examination
Alizarin red–S stained 18th day of ges-tation oxfendazole treated male or femalefetal skeletons showed an obvious decrease
Figure 7. Teratogenic effect of oxfendazole (MRL) in all experimental groups.
Figure 8. Photograph of the uterus of a control pregnant female (A) and a uterus with resorbedembryos of an oxfendazole (MRL) treated female (B) on day 18 of gestation.

Toxicity of anthelmintic drug oxfendazole residues 149
in the size and reduction in the ossificationof fetal skeletons as compared to those ofthe control group. Sacral vertebrae showedsevere lack of ossification and caudal ver-tebrae were completely non ossified. Thebones of the pelvic girdle of the controlfetuses, illium, ischium and pubis were wellossified, whereas in the skeletons of theoxfendazole (MRL) treated fetuses, thepubis bones were completely non-ossifiedand the ischium bones showed a lesserdegree of ossification (Fig. 10).
3.5. Biochemical results
The data of biochemical studies on dif-ferent parameters in all experimental tis-sues figure in Tables VI, VII and VIII. Thedata show that oxfendazole (MRL) affected
Figure 10. Photograph of fetal skeletons at day 18 of gestation showing (A) control fetus with wellossified bones (B) oxfendazole (MRL) maternally treated fetus showing complete non- ossificationof sacral and caudal vertebrae (ca.v) and pubis (pb) bones of the pelvic girdle (alizarine S- red stain).
Figure 9. Photograph of oxfendazole (MRL)maternally treated embryo on day 18 of gesta-tion showing an external Haematoma.

150 A. El-Makawy et al.
Tabl
e V
I. E
ffec
t of
oxfe
ndaz
ole
on to
tal p
rote
in, t
otal
DN
A, t
otal
RN
A a
nd li
ver
enzy
me
activ
ity o
f m
ale
mic
e.
Gro
ups
Pro
tein
mg·
g–1
DN
A m
g·g–
1R
NA
mg·
g–1
Liv
er e
nzym
es u
·L–1
Liv
erK
idne
yTe
stes
Liv
erK
idne
yTe
stes
Liv
erK
idne
yTe
stes
GO
TG
PT
Con
trol
9.66
± 0
.09
8.12
± 0
.32
7.12
± 0
.317
0.46
± 0
.017
0.37
± 0
.02
0.34
± 0
.02
0.28
3±
0.1
70.
26±
0.0
20.
23±
0.0
026
.20
± 1
.49
30.2
0±
1.7
1
Tre
ated
11.9
8**
± 6
.24
9.27
**±
0.1
09.
45**
± 0
.292
0.40
**±
0.0
100.
28**
± 0
.03
0.27
*±
0.1
60.
22*
± 0
.01
0.21
*±
0.0
20.
17*
± 0
.00
34.2
0*±
2.3
939
.40*
± 3
.18
* Si
gnif
ican
t dif
fere
nce
at (
P ≤
0.0
5). *
* S
igni
fica
nt d
iffe
renc
e at
(P
≤ 0
.01)
.
Tabl
e V
II. E
ffec
t of
oxfe
ndaz
ole
on to
tal p
rote
in, t
otal
DN
A, t
otal
RN
A a
nd li
ver
enzy
me
activ
ity o
f fe
mal
e m
ice.
Gro
ups
Pro
tein
mg·
g–1
DN
A m
g·g–
1R
NA
mg·
g–1
Liv
er e
nzym
es u
·L–1
Liv
erK
idne
yL
iver
Kid
ney
Liv
erK
idne
yG
OT
GP
T
Con
trol
9.01
± 0
.44
7.19
± 0
.10
0.45
± 0
.02
0.38
± 0
.02
0.26
± 0
.01
0.24
± 0
.01
24.6
0 ±
2.0
423
.40
± 2
.04
Tre
ated
11.7
28**
± 6
.24
9.09
**±
0.1
10.
28**
± 0
.02
0.28
0**
± 0
.01
0.19
**±
0.0
10.
20±
0.0
133
.20*
± 2
.39
35.8
0*±
3.1
8
* Si
gnif
ican
t dif
fere
nce
at (
P ≤
0.0
5). *
* S
igni
fica
nt d
iffe
renc
e at
(P
≤ 0
.01)
.
Tabl
e V
III.
Eff
ect o
f ox
fend
azol
e on
tota
l pro
tein
, tot
al D
NA
, tot
al R
NA
and
live
r en
zym
e ac
tivit
y of
par
enta
lly tr
eate
d em
bryo
s.
Prot
ein
mg·
g–1
DN
A m
g·g–
1R
NA
mg·
g–1
Liv
er e
nzym
es u
·L–1
GO
TG
PT
Con
trol
7.28
± 0
.27
0.32
± 0
.01
0.23
± 0
.01
20.8
0 ±
1.9
021
.00
± 1
.74
Fem
ale
trea
ted
8.06
* ±
0.1
20.
24 ±
0.0
16.
20 ±
0.0
121
.40
± 0
.98
22.6
0 ±
2.0
4
Mal
e tr
eate
d8.
98**
± 0
.25
0.22
** ±
0.0
10.
16 ±
0.0
127
.800
* ±
1.4
931
.20*
* ±
2.4
2
Mal
e F
emal
e tr
eate
d0.
00 ±
0.0
00.
00 ±
0.0
00.
00 ±
0.0
00.
00 ±
0.0
00.
00 ±
0.0
0
* Si
gnif
ican
t dif
fere
nce
at (
P ≤
0.0
5). *
* S
igni
fica
nt d
iffe
renc
e at
(P
≤ 0
.01)
.

Toxicity of anthelmintic drug oxfendazole residues 151
the liver, kidney, testes and embryo tissuesand caused changes in all measured bio-chemical parameters of mice.
3.5.1. Biochemical changes in males
The data presented in Table VI and Fig-ure 11 showed that oral administration ofoxfendazole (MRL) caused a statisticallysignificant increase in the total protein con-tent of the liver, kidney and testes at (P ≤0.01) when compared with the control.However, the total content of DNA of thetissues was significantly decreased at (P ≤0.01) when compared to the control. Mean-while, a statistically significant decrease inthe level of total RNA content at (P ≤ 0.05)was observed. However, the levels of liverenzymes (GOT and GPT) of oxfendazole(MRL) male mice tissues were statisticallysignificantly higher (P ≤ 0.05) than thecontrol.
3.5.2. Biochemical changes in pregnant females and their embryos
The data of Tables VII and VIII,Figures 12 and 13 showed that the total
DNA and RNA content of the liver ofoxfendazole (MRL) pregnant females andtheir embryos, were significantly decreasedat (P ≤ 0.01) when compared with the con-trol. The DNA content of the kidney tissueswas statistically significant decrease at(P ≤ 0.05) when compared with the con-trol. Whereas, RNA content did not show asignificant change. A significant increase at(P ≤ 0.01) in the protein content of treatedpregnant female tissues and in embryosobtained from them was detected. Also,oxfendazole (MRL) significantly increasedthe liver enzyme (GOT and GPT) contentof females and their embryos at (P ≤ 0.01)as compared with the control.
4. DISCUSSION
The extensive use of veterinary drugs infood producing animals can cause the pres-ence of drug residues in food [7]. Certaindrugs can be carried over into the milk oflactating animals or deposited in the tissuesof animals intended for slaughter [6]. AMaximum Residue Limit is an amount of
Figure 11. Effect of oxfendazole (MRL) on different biochemical parameters in male mice.

152 A. El-Makawy et al.
residue that could remain in the tissue orfood product derived from a food-produc-ing animal that has been treated with a vet-erinary drug. This residue is considered topose no adverse health effects if ingesteddaily by humans over a lifetime [18]. In
recent years, there has been increasing con-cern that drugs may present a potential haz-ard to mankind by causing gene mutation orchromosome aberrations. Genetic altera-tions in somatic cells can include cell deathor transform into malignancy. Genetic
Figure 12. Effect of oxfendazole maximum residue limit (MRL) on different biochemical parame-ters in female mice.
Figure 13. Effect of oxfendazole maximum residue limit (MRL) on different biochemical parame-ters in embryo tissues.

Toxicity of anthelmintic drug oxfendazole residues 153
alterations in germ cells lead to reproductivefailure or genetic disorder in subsequentgenerations [8]. In the present study, oraladministration of oxfendazole (MRL)induced a statistically significant increasein the frequencies of chromosomal aberra-tions in all tested cell types. This means thatanthelmintic oxfendazole has a possiblegenotoxic activity. Crebelli [19] reportedthat genotoxicity of benzimidazoles mayappear in DNA damage or involving mul-tiple interactions with non-DNA targets.Ardito et al. [20]) indicated that higher nontoxic concentrations of Thiabendazole causean increase in the sister chromatid exchangefrequency in lymphocyte culture. Theresults of the present study indicate thatoxfendazole induced a statistically signifi-cant increase in the frequencies of poly-ploidy in bone marrow and spermatocytecells of male mice and also in embryo cells. Theliterature reported that several anthelminticdrugs have been evaluated for genotoxicity.Crebelli et al. [21], Leopardi et al. [22],Natarajan et al. [23], Parry and Sors [24],Mailhes et al., [25] and Adler et al. [26]demonstrated that thiabendazole is capableof inducing numerical chromosomal changessuch as aneuploidy and polyploidy. At toxicdoses, thiabendazole increases the fre-quency of aneuploid in oocytes that impairovulation. Also, Schmid et al. [27] showedthat thiabendazole increases the frequencyof aneuploidy induction in the sperm oftreated male mice. Barale et al. [28] studiedthe cytogenetic effects of three benzimida-zoles, i.e., benomyl, methyl thiophanate andmethyl 2-benzimidazolecarbamate (MBC), inmouse bone marrow cells. Benomyl and MBCsignificantly induce micronuclei and aneu-genic flubendazole causes a dose and timedependent induction of polyploidy inChinese hamster lung cells. Mebendazoleinteracting with tubulin interferes with theassemblage of the mitotic apparatus in theparasite cells, thus causing a possible gen-otoxic activity leading to chromosomalmalsegregation [29]. Also, Okamura et al.[30] suggested that oxfendazole inducesapoptosis of meiotic spermatocytes, most
probably due to disruption of the microtu-bules, and degeneration of the Sertoli cells.Uppala et al. [31] indicate that chemicalscan induce cellular and chromosomal alter-ations, inducing early events in carcinogen-esis. This means that oxfendazole geneticalteration in somatic cells may lead tomalignancy. Our findings indicate that oraladministration of oxfendazole (MRL) toparents for eight weeks produced develop-mental toxicity in their embryos. This tox-icity appeared in the decrease in embryobody weights and the less ossified skele-tons. These results were in agreement withDelatour et al. [32] and Delatour [33] indi-cating that benzimidazole anthelminticoxfendazole, the active metabolite of fenb-antel, is embryotoxic and teratogenic in therat. Scholz and Baeder [34] demonstratedthat Fenbendazole showed fetotoxicity inrats and rabbits dosed by gavage. Embryo-toxicity was observed in the form of anincrease in the frequency of occurrence ofthe 13th ribs and delayed ossification of cra-nial bones in rabbits given a dose of63 mg·kg–1 of body weight/day. Lankaset al. [35] reported that thiabendazole isfoetotoxic in mice. Delatour et al. [36]reported that there is possible correlationbetween embryotoxicity and the plasmalevels of identified embryotoxic metabo-lites, but not with the levels of fetal tissuebound drug metabolites. Whittaker andFaustman [37, 38] reported that the benz-imidazole anthelmintic should be consid-ered as potential developmental toxicants.Since, they inhibit cell growth and differ-entiation of micromass culture of rat embryomidbrain and limb bud cells. In agreementwith our result, Capece et al. [39], Navarroet al. [40] reported that benzimidazoleinduces a significant increase of resorptions,a decrease of fetal body weights and anincrease in skeletal malformation. Mantovni[41] and Teruel et al. [42] indicated thatthe benzimidazole anthelmintic, albenda-zole, shows a dose related increase inembryolethality, growth reduction and areduction in the ossification process inthe rat. Cristofol et al. [43] studied the

154 A. El-Makawy et al.
disposition of albendazole and its metabo-lites, albendazole sulfoxide and sulfone inthe pregnant rat. The results indicated thatthe presence of albendazole and its metab-olites in the amniotic sacs and embryos atconcentration were higher than that in theplasma at the same times. A significant cor-relation was found between the rate ofdevelopmental toxicity and metabolite con-centration. Also, El-Makawy [44] men-tioned that embryotoxicity is mainly theresult of chromosome damage leading to aloss of genetic material. The data of the bio-chemical study in the present work showthat oral administration of oxfendazole(MRL) caused a significant increase in thetotal protein content of the liver, kidney andtestes and decrease in the total content ofDNA and RNA of tissues. Baliharova et al.[45] indicated that all benzimidazoles pro-voke an increase in the protein level of rathepatocytes. Also, oxfendazole (MRL) sig-nificantly increased the liver enzyme (GOTand GPT) content. This result was in agree-ment with that of Tada et al. [46] whoreported that thiabendazole showed anincrease in concentrations of GOT and GPTin male and female mice. Karmakar et al.[47] mentioned that hepatic enzymes sig-nificantly increased along with an increasedpercentage of chromosome aberrations inthe bone marrow. The results thereforestrongly suggest that oxfendazole exertsgenotoxicity in male and female mice.Also, oxfendazole exhibits embryotoxicityincluding teratogenicity. We can concludethat embryo developmental toxicity ofoxfendazole may be a result to the increasein the frequencies of different chromosomalaberrations and biochemical changes. So,we must increase the attention to the poten-tial risk of oxfendazole residues on humanbeings and should stress the need for carefulcontrol to ensure the adherence to the pre-scribed withdrawal time of this drug.
REFERENCES[1] Delatour P, Parish R. Drug residues in ani-
mals. Ed Rico AG, Academic Press, NewYork, USA 1986.
[2] Klee S, Baumung I, Kluge K, Ungemach FR,Horne E, O Keeffe M, De Angelis I, VignoliAL, Zucco F, Stammati A. A contribution tosafety assessment of veterinary drug residues:in vitro/ex vivo studies on the intestinal tox-icity and transport of covalently bound resi-dues. Xenobiotica 1999, 29: 641–654.
[3] FAO/WHO Expert Committee on Food Addi-tives, Evaluation of certain veterinary drugresidues in food, Fiftieth report of the jointWorld Health Organ Tech Rep Ser 1999, 888:1–95.
[4] Prichard RK, Ranjan S. Anthelmintics. VetParasitol 1993, 46: 113.
[5] Rose MD, Bygrave J, Farrington WH, ShearerG. The effect of cooking on veterinary drugresidues in food, Part 8. Benzylpenicillin.Analyst 1997, 122: 1095–1099.
[6] McGrane M, Okeeffe M, Smyth MR. Multi-residue analysis of penicillin residues in por-cine tissue using matrix solid phase disper-sion. Analyst 1998, 123: 2779–2783.
[7] Galer DM, Monro AM. Veterinary drugs nolonger need testing for carcinogenicity inrodent bioassays. Regulatory Toxicology andPharmacology 1998, 28: 115–123.
[8] Nagalakshmi K, Tong-man O. Occupationalexposure to genotoxic agents. Mut Res 1999,437: 175–194.
[9] Yosida TH, Amano K. Autosomal polymor-phic in laboratory bred and Wild Norway ratsRattus norvegicus, found in Misima. Chromo-soma 1965, 16: 658–666.
[10] Brewen GJ, Preston JR. Analysis of chromo-some aberrations in mammalian germ cells.Chemical Mutagenesis 1978, 5: 127–150.
[11] Gleich J, Frohberg H. General teratologicaltechnique. In: Methods in prenatal toxicology,George Thieme Publishers, Stuttgart, 1977.
[12] Weesner FM. General zoological microtech-niques. Scientific Book Indian Ed, AgencyCalcutta, 1968, p 100–101.
[13] Romagnano A, King AW, Richer CL, PerroneMA. A direct technique for the preparation ofchromosomes from early equine embryos. JGent Cytol 1985, 27: 365–369.
[14] Dische Z. Color reaction of nucleic acid com-ponents. In: Chargaff E, Davidson JM (Eds),The nucleic acids, vol 1, Academic press, NewYork, 1955, 270–284.
[15] Schneider WC. Determination of acid in tis-sues by pentose analysis. In: Colowick SP,Kaplan NO (Eds), Method enzymology, Aca-demic press, New York, 1957, p 680–684.

Toxicity of anthelmintic drug oxfendazole residues 155
[16] Gomall AC, Bardaawill CJ, David MM. Pro-tein clometric method. J Biol Chem 1949,177: 751–760.
[17] Reitman S, Frankel S. Got and Gpt colometricmethod. Am J Clin Path 1957, 28: 26–30.
[18] Kirkpatrick D. The veterinary drugs directo-rate to discuss EXTRA-LABEL drug use inHalifax. Can Vet J 2002, 43: 425–426.
[19] Crebelli R, Thershold-mediated mechanismsin mutagenesis: implications in the classifica-tion and regulation of chemical mutagens.Mut Res 2000, 464: 129–135.
[20] Ardito G, Bramanti B, Bigatti P, Lamberti L,Dolara P. Cytogenetic effect of thiabendazoleand diphenylammine on cultured human lym-phocytes; sister chromatid exchanges and cellcycle delay. Boll Soc Ital Biol Sper 1996, 72:171–178.
[21] Crebelli R, Conti G, Conti L, Carere A. Invitro studies with nine known or suspectedspindle poisons: results in tests for chromo-some malsegregation in Aspergillus nidulans.Mutagenesis 1991, 6: 131–136.
[22] Leopardi P, Zijno A, Bassani B, PacchierottiF. In vivo studies on chemically induced ane-uploidy in mouse somatic and germinal cells.Mut Res 1993, 287: 119–130.
[23] Natarajan AT, Duivenvoorden WC, MeijersM, Zwanenburg TS. Induction of mitotic aneu-ploidy using chinese hamster primary embry-onic cells. Test results of 10 chemicals. MutRes 1993, 287: 47–56.
[24] Parry J.M, Sors A. The detection and assess-ment of the aneugenic potentials of environ-mental chemicals. The European CommunityAneuploidy project. Mut Res 1993, 287: 3–15.
[25] Mailhes JB, Young D, Aardema MJ, LondonSN. Thiabendazole-induced cytogenetic abnor-malities in mouse oocytes. Environ Mol Muta-gen 1997, 29: 367–371.
[26] Adler ID, Schmid TE, Baumgartner A. Induc-tion of aneuploidy in male mouse germ cellsdetected by the sperm-FISH assay: a review ofthe present data base. Mutat Res 2002, 504:173–182.
[27] Schmid TE, Xu W, Adler ID. Detection ofaneuploidy by multicolor FISH in mousesperm after in vivo treatment with acrylamide,colchicine, diazepam or thiabendazole. Muta-genesis 1999, 14: 173–179.
[28] Barale R, Scapoli C, Meli C, Casini D,Minunni M, Marrazzini A, Loprieno N, BarraiI. Cytogenetic effects of benzimidazoles inmouse bone marrow. Mut Res 1993, 300: 15–28.
[29] De la Torre RA, Espinosa-Aguirre JJ, Cortinasde Nava C, Izquierdo T, Moron F. Genotoxicactivity of menbendazole in Aspergillus nidu-lans. Mut Res 1994, 305: 139–144.
[30] Okamura M, Watanabe T, Kashida Y, MachidaN, Mitsumori K. Possible mechanisms under-lying the testicular toxicity of oxfendazole inrats. Toxicol Pathol 2004, 32: 1–8.
[31] Uppala PT, Roy SK, Tousson A, Barnes S,Uppala GR, Eastmond DA. Induction of cellproliferation, micronuclei and hyperdiploidy/polyploidy in the mammary cells of DDT- andDMBA-treated pubertal rats. Environ MolMutagen 2005, 46: 43–52.
[32] Delatour P, Parish RC, Gyurik RJ. Albenda-zole: a comparison of relay embryotoxicitywith embryotoxicity of individual metabo-lites. Ann Rech Vet 1981, 12: 159–167.
[33] Delatour P. Some aspects of the teratogenicityof veterinary drugs. Vet Res Commun 1983,7: 125–131.
[34] Scholz D, Baeder O. A teratogenicity test ofHOE 881 with oral administration in yellow-silver rabbits. Hoechst-Roussel unpublishedreport, submitted to WHO by Hoechst AG,Frankfurt am Main, Germany, 1973.
[35] Lankas GR, Nakatsuka T, Komatsu T,Matsumoto H. Developmental toxicity oforally administered thiabendazole in ICRmice. Food Chem Toxicol 2001, 39: 367–374.
[36] Delatour P, Garnier F, Benoit E, Longin C. Acorrelation of toxicity of albendazole andoxfendazole with their free metabolites andbound residues. J Vet Pharmacol Ther 1984,7: 139–145.
[37] Whittaker SG, Faustman EM. Effects ofalbendazole and albendazole sulfoxide on cul-tures of differentiating rodent embryonic cells.Toxicol Appl Pharmacol 1991, 109: 73–84.
[38] Whittaker SG, Faustman EM. Effects of ben-zimidazole analogs on cultures of differenti-ating rodent embryonic cells. Toxicol ApplPharmacol 1992, 113: 144–151.
[39] Capece BP, Navarro M, Arcalis T, Castells G,Toribio L, Perez F, Carretero A, Ruberte J,Arboix M, Cristofol C. Albendazole sulphox-ide enantiomers in pregnant rats embryo con-centrations and developmental toxicity. Vet J2003, 165: 266–275.
[40] Navarro M, Canut L, Carretero A, Cristofol C,Perez-Aparicio FJ, Arboix M, Ruberte J.Developmental toxicity in rat fetuses exposedto the benzimidazole netobimin. Reprod Tox-icol 1999, 13: 295–302.

156 A. El-Makawy et al.
[41] Mantovani A. The role of multigenerationstudies in safety assessment of residues of vet-erinary drugs and additives. Ann Ist SuperSanita 1992, 28: 429–435.
[42] Teruel MT, Felipe AE, Solana HD, SallovitzJM, Lanusse CE. Placental and fetal toxicityof albendazole sulphoxide in Wistar rats. VetHum Toxicol 2003, 45: 131–136.
[43] Cristofol C, Navarro M, Franquelo C, ValladaresJE, Carretero A, Ruberte J, Arboix M. Dispo-sition of netobimin, albendazole and itsmetabolites in the pregnant rats developmen-tal toxicity. Toxicol Appl Pharmacol 1997,144: 56–61.
[44] El-Makawy A. Mutagenic and teratogenicstudies induced by anticancer (cisplatinum) in
rat embryos. A Thesis submitted for the ful-fillment of PhD degree in science, 1998.
[45] Baliharova V, Velik J, Lamka J, Balarinova R,Skalova L. The effects of albendazole and itsmetabolites on hepatic cytochromes P450activities in mouflon and rat. Res Vet Sci2003, 75: 231–239.
[46] Tada Y, Fujitani T, Yoneyama M. Subchronictoxicity of thiabendazole (TBZ) in ICR mice.Food Chem Toxicol 1996, 34: 709–716.
[47] Karmakar R, Banerjee A, Datta S, ChatterjeeM. Influence of cadmium intoxication onhepatic lipid peroxidation, glutathione level,and glutathione S-transferase and gamma-glutamyl transpeptidase activities: correlationwith chromosome aberrations in bone marrowcells. J Environ Pathol Toxicol Oncol 1999,18: 277–287.
To access this journal online: www.edpsciences.org