global gene expression in the bovine corpus luteum is...
TRANSCRIPT

Global gene expression in the bovine corpus luteumis altered after stimulatory and superovulatory treatments
LucianaA. FatimaA,E,Pietro S. BaruselliB, LindsayU.GimenesB,Mario BinelliB,Francisco P. RennoC, Bruce D. MurphyD and Paula C. PapaA
ASector of Anatomy,Department of Surgery, Faculty of VeterinaryMedicine andAnimal Sciences,
University of Sao Paulo, Sao Paulo – SP, 05508-270 Brazil.BDepartment of Animal Reproduction, Faculty of Veterinary Medicine and Animal Science,
University of Sao Paulo, Pirassununga – SP, 13635-900, Brazil.CDepartment of Nutrition and Animal Production, Faculty of Veterinary Medicine and Animal
Science, University of Sao Paulo, Pirassununga – SP, 13635-900, Brazil.DAnimal Reproduction Research Centre (CRRA), University of Montreal, St-Hyacinthe,
QC J2S 7C6, Canada.ECorresponding author. Email: [email protected]
Abstract. Equine chorionic gonadotrophin (eCG) has been widely used in superovulation and artificial inseminationprogrammes and usually promotes an increase in corpus luteum (CL) volume and stimulates progesterone production.Therefore, to identify eCG-regulated genes in the bovine CL, the transcriptome was evaluated by microarray analysis and
the expression of selected genes was validated by qPCR and western blot. Eighteen Nelore crossbred cows were dividedinto control (n¼ 5), stimulated (n¼ 6) and superovulated groups (n¼ 7). Ovulation was synchronised using a progester-one device-based protocol. Stimulated animals received 400 IU of eCG at device removal and superovulated animalsreceived 2000 IU of eCG 4 days prior. Corpora lutea were collected 7 days after gonadotrophin-releasing hormone
administration. Overall, 242 transcripts were upregulated and 111 transcripts were downregulated in stimulated cows(P# 0.05) and 111 were upregulated and 113 downregulated in superovulated cows compared to the control animals(1.5-fold,P# 0.05). Among the differentially expressed genes,manywere involved in lipid biosynthesis and progesterone
production, such as PPARG, STAR, prolactin receptors and follistatin. In conclusion, eCG modulates gene expressiondifferently depending on the treatment, i.e. stimulatory or superovulatory. Our data contribute to the understanding of thepathways involved in increased progesterone levels observed after eCG treatment.
Additional keywords: eCG, follistatin, lipid metabolism, progesterone, prolactin receptors, PPARG.
Received 17 May 2012, accepted 5 September 2012, published online 30 October 2012
Introduction
The use of biotechnology to improve reproductive efficiency isprevalent in cattle, especially techniques directed towards
ovarian follicle growth and manipulation of conception rates.Different strategies have been proposed to improve the responsesto timed artificial insemination, such as the use of equine
chorionic gonadotrophin (eCG) to stimulate follicle growth,progesterone (P4) production and conception rates (Bo et al.
2002; Sa Filho et al. 2010; Sales et al. 2011). Furthermore, eCG
has been employed in various superovulatory protocols toincrease the number of embryos produced per donor cow(Baruselli et al. 2011) and this technique renders it possible todisseminate high genetic quality and improve the reproductive
performance in cattle (Ambrose et al. 1999; Hansen et al. 2001;Baruselli et al. 2011).Derived from the equine fetal chorion, eCGis a glycoprotein that binds to both LH and FSH receptors in
species other than the horse (Murphy andMartinuk 1991) and cantherefore activate multiple signal transduction pathways in theovary. Treatments using eCG can improve corpus luteum (CL)
volume, which is usually associated with increases in progester-one concentration (Binelli et al. 2001; Sa Filho et al. 2010;Baruselli et al. 2011; Fields et al. 2012). Progesterone plays a key
role in reproductive events associated with the establishment andmaintenance of pregnancy via P4-induced changes in the endo-metrial transcriptome (Clemente et al. 2009). Luteotrophic hor-
mones can alsomodulate luteal synthesis of cytokines and growthfactors, such as vascular endothelial growth factor (VEGF; Papaet al. 2007), the insulin-like growth factors (IGFs; Juengel et al.1997) and many other elements that can influence luteal cell
function (Devoto et al. 2000; Webb et al. 2002).The differentiation of follicular granulosa cells into large
luteal cells is associated with marked changes in metabolic
CSIRO PUBLISHING
Reproduction, Fertility and Development
http://dx.doi.org/10.1071/RD12155
Journal compilation � CSIRO 2012 www.publish.csiro.au/journals/rfd

function, including an increase in mitochondrial enzymeactivity (Doody et al. 1990; Richards and Almond 1994),
necessary to supply energy for steroid production. The first stepin progesterone synthesis is the conversion of cholesterol topregnenolone by cytochrome P450, family 11, subfamily A,
polypeptide 1 (CYP11A1/P450scc), which is located in the innermitochondrial membrane (Miller 1988; Niswender and Nett1994; Niswender et al. 1994). Pregnenolone can then be con-
verted to progesterone by hydroxy-delta-5-steroid dehydro-genase 3b (HSD3B), which is present in the smoothendoplasmic reticulum (Labrie et al. 1992). The initial chal-lenge for steroidogenesis as well as its rate-limiting step is to
transport the hydrophobic parent molecule cholesterol from theouter to the inner mitochondrial membrane under the influenceof the steroidogenic acute regulatory protein (STAR), which is
acutely induced by luteotrophic hormones (Stocco et al. 2001).Luteal cells can obtain cholesterol through cholesterol-rich low-density lipoprotein (LDL) endocytosis or through selective
uptake of high-density lipoprotein (HDL) cholesterol esters(Brannian and Stouffer 1993). In addition, de novo cholesterolsynthesis contributes to the intracellular cholesterol load andthis synthesis is controlled by luteotrophic hormones (Golos and
Strauss 1988). In cows, fatty-acid precursor availability iscoupled with an increase in sexual steroid levels (Garcia-Bojalilet al. 1998). Cholesterol homeostasis is central to adequate CL
function and might involve several factors, such as peroxisomeproliferator-activated receptors (PPARs), which are key tran-scription factors in lipid metabolism regulation (Komar 2005;
Varga et al. 2011).It is well established that CL function requires coordinated
actions between luteotrophic hormones, growth factors and
steroidogenic proteins (Niswender 2002), as well as cellularlipid biosynthesis and homeostasis (Kliewer et al. 1997;Christenson and Devoto 2003). Because it has been previouslyreported that eCG increased CL volume and P4 production in
cows (Bo et al. 2002; Sa Filho et al. 2010; Sales et al. 2011), wehypothesised that eCG regulates the expression of genesinvolved in luteal development and in the steroidogenic process.
Therefore, this work was designed to analyse the effects of eCGon global gene expression in bovine CL bymicroarray, focusingon steroidogenesis-related protein expression, to gain further
insights into the progesterone increases observed after stimula-tory and superovulatory treatments.
Material and methods
Animals and tissue collection procedures
All procedures were approved by the Committee in Ethics for
the Use of Experimental Animals of the Faculty of VeterinaryMedicine and Animal Sciences, University of Sao Paulo, Brazil,under protocol number 1638/2009. Eighteen Nelore crossbred
cows aged between 2 and 5 years were used. The animals weremaintained on pasture and supplemented with cornmeal(17.03%), soybean meal (4.65%), urea (1.13%), ammonium
sulfate (0.11%), minerals (1.69%), salt (0.53%) and corn silage(74.86%). Before the trial period, cows were evaluated for theirbody condition score on a scale of 1 to 5 (Wildman 1982) and fortheir ovarian status by rectal palpation, as described by
Madureira (2004); animals showing a condition score between2.0 and 3.0 with functional ovaries were selected.
Hormonal treatments
Cows were randomly distributed into three groups (control,n¼ 5; stimulated, n¼ 6 and superovulated, n¼ 7; Fig. 1).
Briefly, on Day 0 (random day of the oestrous cycle) all animalsreceived an intravaginal device containing 1 g of progesterone(Primer, Technopec, Sao Paulo, Brazil) and an intramuscular
injection of 2mg of oestradiol benzoate (Estrogin; Farmavet,Sao Paulo, Brazil). On Day 8, the intravaginal devices wereremoved from the control and stimulated cows and 0.150mg ofd-cloprostenol (prostaglandin (PGF) 2a, Prolise; Arsa, BuenosAires, Argentina) was administered. The control cows did notreceive eCG (Novormon; Syntex, Buenos Aires, Argentina),whereas the stimulated cows received 400 IU on Day 8, which is
considered to be the optimal dose to improve dominant follicles(Sa Filho et al. 2010; Sales et al. 2011). At 48 h following deviceremoval, control and stimulated cows received 0.025mg of
lecirelin (gonadotrophin-releasing hormone (GnRH), GestranPlus; Arsa). Cows submitted to superovulatory treatment, whichaimed to increase the number of ovulated follicles, received
2000 IU of eCG on Day 4 and 0.150mg of PGF2a on Day 6(Baruselli et al. 2011). On Day 7, the devices were removed anda second dose of PGF2a was administered. Twelve hours afterdevice removal, 0.025mg of GnRH was administered to induce
ovulation (Fig. 1) Differences regarding hormonal protocol forsuperovulated cows are based on the achievement of moresynchronous ovulations (Baruselli et al. 2012). The animals
were slaughtered on Day 7 after GnRH administration, theovaries were collected and the CL was dissected and snap-frozen in liquid nitrogen for later mRNA or protein extraction.
Tissue samples were fixed in 4% phosphate-buffered formalinsolution and embedded in paraffin for immunohistochemistryand immunofluorescence analyses.
Number and volume of corpora lutea (CLL)
CLLwere counted and volumewas calculated using the formula
for an ellipsoid: (3/4pi (width/2) (height/2) (depth/2)). Forsuperovulated cows three CLL were used to calculate the CLvolume.
Progesterone assay
P4 concentrations were measured by radioimmunoassay (RIA)using a commercial kit (COAT-A-COUNT Progesterone;Siemens Medical Solutions Diagnostics, Los Angeles, CA,
USA). The assay sensitivity was 0.05 ngmL�1, and the high andlow intra-assay coefficients of variation were 2.11% and 5.91%,respectively. The total plasma progesterone concentration was
divided by the number of corpora lutea in the superovulatedanimals.
RNA extraction
Total RNA extraction from superovulated animals wasperformed using a pool of three CLL from each animal.Trizol reagent (Life Technologies, Carlsbad, CA, USA) wasused to extract the total RNA, according to manufacturer’s
B Reproduction, Fertility and Development L. A. Fatima et al.

recommendations. The RNA was further purified using an
RNAeasy kit with DNase treatment to eliminate genomiccontamination (Qiagen, Valencia, CA, USA). The RNA wasmeasured with a NanoDrop 2000 (Thermo Scientific, Hudson,
NH, USA) and the purity and integrity of the samples wasassessed with the RNA 6000 LabChip using an Agilent 2100Bioanalyzer (Agilent Technologies, Santa Clara, CA,USA). For
the next steps, only RNAwith a RNA integrity number (RIN) of9 or above was used.
Microarray
The microarray analysis was performed at the McGill Univer-sity and Genome Quebec Innovation Centre, Montreal, Canada.Briefly, target RNA was reverse transcribed into cDNA using a
T7 oligo(dT) primer (Affymetrix, Santa Clara, CA, USA) tosynthesise cDNA containing a T7 promoter sequence. Then, thepurified cDNA was used as a template for in vitro transcription
to synthesise biotin-modified aRNA with the IVT LabellingMaster Mix (Affymetrix), which produced multiple copies ofbiotin-modified aRNA. The aRNA was purified to removeunincorporated NTPs, salts, enzymes and inorganic phosphate
to improve the stability of the biotin-modified aRNA.
The labelled aRNA was then fragmented and hybridised to an
Affymetrix GeneChip Bovine Genome Array (Affymetrix,Santa Clara, CA,USA) containing 23 000 probesets and scannedin a GeneChip Scanner 3000 (Affymetrix). The experiments
were performed using three samples from each group. Analysisof the data was performed with the FlexArray Software V1.3(Genome Quebec Innovation Center, Montreal, QC, Canada;
http://genomequebec.mcgill.ca/FlexArray/). Raw fluorescencedata were processed and normalised with the affx Bioconductorpackage Version 1.1.0 (http://bioconductor.org/). Lists of geneshaving a fold change of $�1.5 and a P value of #0.05 were
generated, and the results were compared between control andstimulated, control and superovulated and stimulated andsuperovulated cows. Lists of differently expressed genes were
analysed using Ingenuity Pathway Analysis 7 software (IPA;Ingenuity Systems, San Francisco, CA, USA; http://www.ingenuity.com/). The IPA interprets data in the context of
common molecular networks and biological processes usinginformation present in the literature. The software ranks thenetworks and biological function according to the number ofgenes present in each list. Themicroarray data were deposited in
the NCBI gene expression and hybridisation array data
Control (n � 5)
BE (2 mg) �P4 device
BE (2 mg) �P4 device
BE (2 mg) �P4 device
Removal of P4 device � 0.150 mg
of PGF2α
Removal of P4 device � 0.150 mg
of PGF2α �400 IU of eCG
Removal of P4 device �0.150 mg of
PGF2α
Slaughter andmaterial
collection
Slaughter andmaterial
collection
Progesterone
D0 D8 D10
GnRH Ovulation
GnRHOvulation
Slaughter andmaterial
collectionGnRHOvulation
D11 D17
D0 D8 D10 D11 D17
D0 D4 D8 M D9 D15D7 AD6 M
Progesterone
Progesterone
Stimulated (n � 6)
Superovulated (n � 7)
2000 IUof eCG PGF2α
Fig. 1. Hormonal treatment protocol for oestrus synchronisation, stimulation of the dominant follicle or
superovulation showing the timetable for administration of oestradiol benzoate (BE), progesterone (P4)-releasing
device and administration of prostaglandin (PGF2a), gonadotrophin releasing hormone (GnRH) or equine
chorionic gonadotrophin (eCG). D, day after the beginning of the protocol; M, morning; A, afternoon.
eCG alters global gene expression of bovine CL Reproduction, Fertility and Development C

repository (GEO, http://www.ncbi.nlm.nih.gov/geo). The entiremicroarray dataset can be retrieved using the GEO accession
number GSE37844.
Quantitative real-time PCR
For validation of microarray data, quantitative real-time PCR
(qPCR) was performed to assess the abundance of the followinggenes: peroxisome proliferator-activated receptor gamma(PPARG), the CD36 molecule (CD36), fatty-acid binding pro-tein 5 (FABP5), cytochrome P450, family 27, subfamily A,
polypeptide 1 (CYP27A1), acyl-CoA synthetase family member2 (ACSF), neutral cholesterol ester hydrolase 1 (NCEH1),3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR), short
(PRLRS) and long (PRLRL) prolactin receptors, follistatin andtransforming growth factor b2 (TGFB2). Three other candidategenes related to steroidogenesis, STAR, HSD3B and CYP11A1,
were also analysed. All 18 samples were used in those analyses.An aliquot of 1mg of total RNA was submitted to reverse tran-scription using SuperScript III (Life Technologies). Theresulting cDNA was used in subsequent qPCR reactions using a
7500 real-time PCR system (Life Technologies). The TaqManchemistry (TaqMan Universal PCR Master Mix; Life Tech-nologies) was used for three transcripts encoding STAR,HSD3B
and CYP11A1 and the reference gene a-tubulin, while SYBRGreen (SYBR Green; Life Technologies) was used for theremaining transcripts and the reference genes a-tubulin and
glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Theprimers and probes for all transcripts are described in Table 1.Common thermal cycling settings were used to amplify each
transcript (2min at 508C, 10min at 958C then 40 cycles of 15 s at958C and 60 s at 608C). After the SYBRGreen reaction, meltingcurve analyses were performed to verify product identity byadding a dissociation step to the PCR run (15 s at 958C, 60min at
608C, 15 s at 958C and 15 s at 608C). To test the efficiency ofamplification of primer pairs for each gene, a cDNA pool wasmade using equal volumes of the cDNA solution from each
sample. The cDNA pool was serially diluted from 1 : 2 to 1 : 64
in autoclaved RNase-free water. Efficiency of amplification oftarget as well as the reference genes was calculated by
‘LinRegPCR’ (Ramakers et al. 2003) using the slope of thedilution curve. The relative expression was calculated using thePfaffl equation (Pfaffl 2001).
Immunohistochemistry
Tissue sections from one CL per animal were deparaffinised inxylene and rehydrated in a graded series of ethanol. Antigenretrieval was performed by boiling the sections in sodium citrate
buffer (10mM, pH 6.0) in a microwave for 3� 5min, followedby blocking endogenous peroxidase activity with 1% (v/v)hydrogen peroxide in methanol. Nonspecific binding was
blocked by incubating the sections with Protein Block (Dako,Carpinteria, CA, USA) for 30min. The sections were thenincubated with primary antibodies (Table 2) overnight at 48C.For HSD3B and P450scc, antibody dilutions of 1 : 200 and1 : 1000 were used, respectively. Next, the sections were incu-bated with Dako LSAB System-HRP and the DAB ChromogenSystem (Dako) according the manufacturer’s instructions. The
slides were observed with an Olympus BX 50 microscopeequipped with a CCD colour video camera (Olympus DP71;Olympus America Inc, Center Valley, PA, USA), and the ima-
ges were captured using Axio Vision software (Carl Zeiss,Oberkochen, Germany).
Immunofluorescence
The protein localisation of STAR, FABP5, HMGCR and fol-listatin was verified by immunofluorescence in the CL tissue.
Briefly, tissue sections were treated as described above until theblockade step. The sections were blocked in phosphate-bufferedsaline (PBS) containing 5% bovine serum albumin (BSA) for30min at room temperature (RT) followed by incubation with
the primary antibody overnight at 48C (Table 2). The antibodieswere diluted in PBS containing 5% BSA to the following con-centrations: STAR 1 : 300, HMGCR1 : 50, follistatin 1 : 200 and
FABP5 1 : 500. As a negative control, sections were incubated
Table 1. Primers and probes used in qPCR
Gene Forward Reverse Probe GenBank no.
ACSF2 CACAGTCATCTCGGTGGATG GTCATGGCAGGACAGGAACT – NM_001078112.1
CD36 TCAGAAATCAAGTGACTGGGAAAA ACCACACCAACACTGAGCAAGA – NM_174010.2
CYP11A1 CCCTTTCCACCAATCCAGCTA GGACTGAGCGAGCAATGGA TGGCCCCACCCCCTC NC_007319
CYP27A1 AGCTGTTGGTGCAAGGCTAC GTGCATCTGAGGCCCTACTC – NM_001083413.1
FABP5 CACAGCTGATGGCAGAAAGA TCCCATCTTCCAGTTTTCTTG – NM_174315.3
Follistatin TTTCTGTCCAGGCAGCTCTA GTCACTCCATCATTCCCACA – NM_175801.2
GAPDH GCGATACTCACTCTTCTACTTTCGA TCGTACCAGGAAATGAGCTTGAC AB098985
HMGCR GAAATGGAACTCCCTGTGGA TGGGATATGCTTTGCATTGA – NM_001105613.1
HSD3B GCTAGACAAAGTCTTCAGACCAGAA CAGCAGGGTCAGCTTGATCTT CTGGAGCTTAGAAAATT NM_001034696
NCEH1 CCAGAAGAGCCTCTGAAACG TGGTTGTGCACAGTTCATCAT NM_001123034.1
PPARG CACTCCGCACTATGAGGACA ACAGGCTCCACTTTGATTGC – NM_181024.2
PRLRL TTGATGTTCATCTGCTGGAGAAG GCAAGTCCTCGCAGTCAGAA – NM_001039726.1
PRLRS GCCTTCTCGCCTTGTGTCTATG GCAAGTCCTCGCAGTCAGAA – NM_001039726.1
STAR Bt03213114 NM_174189.2
TGFB2 GCCGAGTTCAGAGTCTTTCGTTT GCGCTGGGTTGGAGATGTTA – NM_001113252.1
a-Tubulin TGTTCGCTCAGGTCCTTTTGG CCCTTGGCCCAGTTGTTG CCCGGACTGACCAAAA BT_0323101
D Reproduction, Fertility and Development L. A. Fatima et al.

with BSA instead of the primary antibody. The sections were
washed, incubated with Cy3-conjugated donkey anti-rabbit(Millipore, Temecula, CA, USA) diluted 1 : 300 (STAR,HMGCR and FABP5) or fluorescein isothiocyanate
(FITC)-conjugated anti-rabbit (1 : 300;Millipore) in PBS for 1 h
at RT. The slides were then washed and the sections werecounterstained with 40,6-diamidino-2-phenylindole (DAPI;Sigma-Aldrich, St. Louis, MO, USA) diluted 1 : 1000 in PBS
Table 2. Antibodies used for immunohistochemistry, immunofluorescence and western blotting
Antibodies Isotype Epitope Supplier (order no.)
ß-Actin Mouse monoclonal IgG1 N-terminus human Sigma (1978)
FABP5 Rabbit polyclonal IgG 1–100 human E-FABP Abcam (ab84028)
Follistatin Rabbit polyclonal IgG C-terminus mouse Santa Cruz (sc-315)
HMGCR Rabbit polyclonal IgG Mouse Santa Cruz (sc-33827)
HSD3B Mouse polyclonal IgG1 Full-length human HSD3B Santa Cruz (sc-100466)
P450scc Rabbit polyclonal IgG 421–441 Mouse CYP11A1 Abcam (ab78416)
PPARG Rabbit polyclonal IgG 8–109 Human PPARG Santa Cruz (sc-7196)
Prolactin receptors Mouse monoclonal IgG1 Extracellular portion of rat PRLR Abcam (ab84028)
STAR Rabbit polyclonal IgG 1–285 human STAR Santa Cruz (sc-25806)
Up-regulated Down-regulated
203 38
1
20
135
0
53 108 13
47
186
0
0
53
Stim � Control
Stim � Sup Stim � Sup
Sup � Control Stim � Control Sup � Control
Fig. 2. Venn diagram showing number of genes belonging to the three lists and the total number of
genes exhibiting 1.5-fold change (P# 0.05) in stimulated and superovulated animals in relation to each
other and to control animals. Stim, stimulated group; Sup, superovulated group.
3.0Control
Stimulated
Superovulated
aab
a a a
b b
bb b
b
b b b
a a a
bb
ab
ab
1.0
0CD36 FABP5 PPARG CYP27 ACSF2 HMGCR NCEH1
2.5
qPC
R m
RN
A r
elat
ive
expr
essi
on
0.5
2.0
1.5
Fig. 3. Gene expression ofCD36, FABP5, PPARG;CYP27A, ACSF2,HMGCR andNCEH1 in bovine
CLL from Day 7 after gonadotrophin releasing hormone (GnRH) administration in control, stimulated
and superovulated animals as determined by qPCR (mean � s.e.m.); bars with different superscripts
differ significantly (P, 0.05). The reference genes used were GAPDH and a-tubulin.
eCG alters global gene expression of bovine CL Reproduction, Fertility and Development E

and incubated for 5min. The slides were mounted in Permafluor(Laboratory Vision Corp., Fremont, CA, USA). The CL
distribution of each protein was observed by fluorescencemicroscopy using the Olympus Fluoview 1000 system(Olympus America Inc) and Fluoview Version 1.7 software
(Olympus America Inc).
Protein extraction and purification
Protein extraction from superovulated animals was performedusing a pool of three CLL from each animal. The CL samples
(100mg) were homogenised in buffer containing 50mMpotassium phosphate (pH 7.0), 0.3M sucrose, 0.5mM dithio-threitol (DTT), 1mM ethylenediaminetetraacetic acid (EDTA,
pH 8.0), 0.3mM phenylmethylsulfonyl fluoride (PMSF),10mM NaF and phosphatase inhibitor cocktail (1 : 100; Sigma-Aldrich). After centrifugation (20 000g, 5min, 48C) the
supernatant containing the total protein was isolated. Proteinlevels were determined by the Bradfordmethod (Bradford 1976;
Protein Assay Kit; Bio-Rad Laboratories, Hercules, CA, USA),comparing the measurements obtained for samples with thestandard curve of albumin read at 595 nm.
Western blot
Western blot analysis was performed to detect and compare theprotein content of STAR, HSD3B, FABP5, PPARG, HMGCR,follistatin, PRLRS and PRLRL in all experimental groups.Equal amounts (50mg) of total proteinwere resolved on 10–17%one-dimensional SDS–PAGE minigels depending on theprotein molecular weight. The separated proteins wereelectro-transferred to immunoblot polyvinylidene difluoride
membranes (Bio-Rad Laboratories). After the transfer themembranes were blocked with 5% non-fat dried milk inPBS-Tween 1% (PBS-T) for 2 h and incubated with primary
antibodies (Table 2) overnight at 48C at the following dilutions:STAR 1 : 300, 3HSDB 1 : 200, HMGCR 1 : 200, PPARG 1 : 200,FABP5 1 : 500, follistatin 1 : 800, prolactin receptors 1 : 50.
On the following day the membranes were incubated with thesecondary antibody, either anti-rabbit IgG-peroxidase conjugateor anti-mouse IgG-peroxidase (1 : 5000, ECL; GE Healthcare,Buckinghamshire, UK) for 1 h at room temperature. The pro-
teins were visualised by chemiluminescence (ImmobilonWestern; Millipore Corporation, Billerica, MA, USA). Next themembranes were incubated with a b-actin (ACTB) horseradishperoxidase-conjugated anti-mouse antibody (Santa Cruz Bio-technology, Santa Cruz, CA,USA) diluted 1 : 50 000 in blockingsolution for 30min at room temperature. The optical density of
the bands was quantified using image processing and analysissoftware (ImageJ; NIH, Bethesda, MD, USA). The ratio of thedensity of the target protein to ACTB was obtained foreach sample.
Statistical analysis
The statistical analysis of the microarray data was conducted
according to the random variance model of Wright and Simon
6
5
4
3
a a a a
ab
PRLRS PRLRL Follistatin TGFB2
b
b
b
b
b
ab
ab2
1
0
qPC
R m
RN
A r
elat
ive
expr
essi
on
Control
Stimulated
Superovulated
Fig. 4. Gene expression of PRLRS, PRLRL, follistatin and TGFB2 in
bovine CLL from Day 7 after gonadotrophin releasing hormone (GnRH)
administration in control, stimulated and superovulated animals as deter-
mined by qPCR. (mean � s.e.m.); bars with different superscripts differ
significantly (P, 0.05). The reference genes used were GAPDH and
a-tubulin.
Table 3. Microarray results for genes validated by qPCR
Gene name Gene symbol Fold change P value Fold change P value
Stimulated� control Superovulated� control
Acyl-CoA synthetase family member 2 ACSF2 1.89A 0.0001A �1.02 0.76
CD36 molecule CD36 1.56A 0.002A 1.07 0.49
Cytochrome P450, family 11, subfamily A, polypeptide 1 CYP11A1 �1.02 0.78 �1.04 0.56
Cytochrome P450, family 27, subfamily A, polypeptide 1 CYP27A1 1.60A 0.01A 1.19A 0.04A
Fatty acid-binding protein 5 (psoriasis-associated) FABP5 3.02A 0.01A 2.05A 0.01A
Follistatin FST 2.52A 0.002A 1.50 0.21
3-hydroxy-3-methylglutaryl-CoA reductase HMGCR 1.24 0.06 1.07 0.59
Hydroxy-delta-5-steroid dehydrogenase, 3b and steroid delta-isomerase 1 HSD3B1 �1.06 0.23 �1.10 0.13
Neutral cholesterol ester hydrolase 1 NCEH �3.93A 0.001A �5.57A 0.001A
Peroxisome proliferator-activated receptor gamma PPARG 1.64A 0.01A 1.53A 0.03A
Prolactin receptor PRLR 2.44A 0.01A �1.18 0.19
Steroidogenic acute regulatory protein STAR 1.01 0.89 �1.05 0.57
Transforming growth factor, b 2 TGFB2 1.52 0.03 1.40 0.05
AIndicates that genes were up- or down-regulated (1.5 fold, P, 0.05).
F Reproduction, Fertility and Development L. A. Fatima et al.
(2003; significance analysis of microarrays (SAM)). Data fromreal-time PCR and western blots were tested for a normal dis-
tribution of residuals (Anderson–Darling test) and data that didnot follow a normal distribution were transformed into loga-rithms. Subsequently, the effect of treatment was analysed by
one-way ANOVA and treatment means were separated by theTukey test. The effect of treatments andmeans comparison wereconsidered significant when P, 0.05. The data are presented as
the mean� s.e.m. The statistical analyses were performed usingthe GraphPad Prism software (Version 4.00 for Windows;GraphPad Software, San Diego, CA, USA).
Results
Number and volume of CLL
CLwas single in all cows from the control and stimulated groupswhereas in superovulated cows the number ranged from 6 to 36
CLL. CL volume was highest in superovulated (1495.18�137.01mm3) followed by stimulated (1177.37� 167.07mm3)and then control (830.33� 234.99mm3) cows.
Plasma progesterone concentrations
P4 concentrations were higher in stimulated (5.95�0.17 ngmL�1) than in control (3.69� 0.72 ngmL�1, P¼ 0.03)
cows. However, in superovulated cows, total P4 concentrationsranged between 47.3 ngmL�1 and 184.7 ngmL�1 (mean85.12 ngmL�1); when divided by CL number, values were
lower (4.11� 0.63 ngmL�1 per CL, P¼ 0.001) than in stimu-lated cows and equal to control cows.
Microarray
The microarray analysis yielded three lists of genes whoseexpression levels changed at least 1.5-fold (P# 0.05) whengroups were analysed by pairs. Overall, 242 transcripts were
upregulated and 111 transcripts were downregulated in stimu-lated cows (P, 0.05) and 111 were upregulated and 113downregulated in superovulated cows in relation to the control
(1.5 fold, P# 0.05). Genes belonging to the three lists and thetotal number of genes are depicted in aVenndiagram (Fig. 2).Thecomplete lists of the differently expressed genes are in Table S1available as SupplementaryMaterial to this paper. The Ingenuity
Pathway network functions found in the CL of stimulated ani-mals are related to cell death, carbohydrate and lipid metabolismand tissue development. In the case of superovulated animals, the
networkswere related to cell-to-cell signalling, the inflammatoryresponse, cell morphology, the cell cycle, cell growth and pro-liferation and lipid metabolism. The main network functions
involving the genes that were differently expressed between thestimulated and superovulated animals were related to lipidmetabolism, post-translational modification, cell death, cell-to-
cell signalling and protein folding.
Real-time qPCR validation
Most of the genes selected for qPCR analyses were related to
lipid metabolism. Among them, CD36, CYP27 and ASCF2,which participate in PPAR pathways, were upregulated in thestimulated animals comparedwith controls.PPARG andFABP5,
which also participate in PPAR pathways, were upregulated inboth the stimulated and superovulated groups comparedwith thecontrol. HMGCR and NCHI, participants in cholesterol metab-olism were, respectively, up- and down-regulated in stimulated
and superovulated cows in relation to control (Fig. 3). In the nextstep, genes belonging to different pathways related to proges-terone production and CL development were also analysed. The
PRLRs and TGFB2 were upregulated in stimulated cows com-paredwith control and follistatinwas upregulated in both groups,stimulated and superovulated (Fig. 4).
In the stimulated animals, the expression ratios obtained byqPCRwere consistent with themicroarray results (Figs 3 and 4).
25
20
15
10
5
7
6
5
4
3
2
1
0
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0
0Control
(a)
(b)
(c )
qPC
R r
elat
ive
expr
essi
onH
SD
3BqP
CR
rel
ativ
e ex
pres
sion
CY
P11
A1
qPC
R r
elat
ive
expr
essi
onS
TAR
Stimulated Superovulated
Control
a
bb
Stimulated Superovulated
Control Stimulated Superovulated
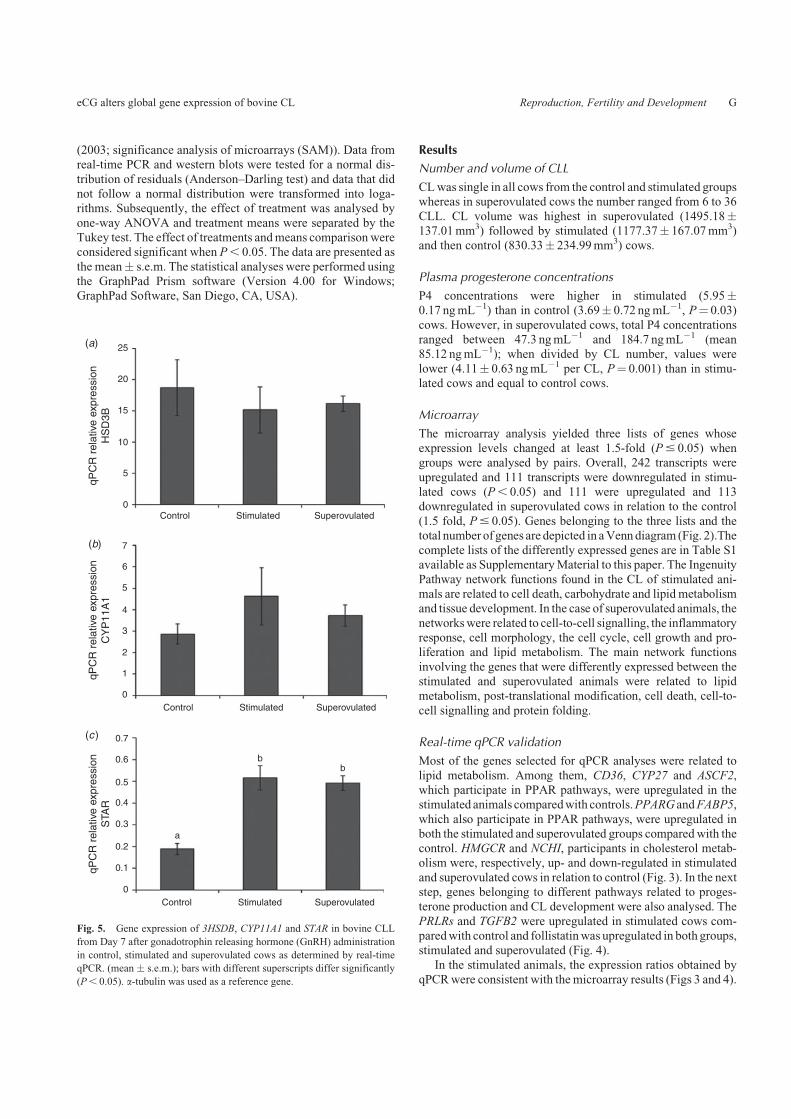
Fig. 5. Gene expression of 3HSDB, CYP11A1 and STAR in bovine CLL
from Day 7 after gonadotrophin releasing hormone (GnRH) administration
in control, stimulated and superovulated cows as determined by real-time
qPCR. (mean � s.e.m.); bars with different superscripts differ significantly
(P, 0.05). a-tubulin was used as a reference gene.
eCG alters global gene expression of bovine CL Reproduction, Fertility and Development G

For superovulated animals, the discrepancy between the results
was greater, most likely due to the large individual variability inresponse to the treatment. With respect to the expression offollistatin, no difference between superovulated and control
animals was observed using microarray analysis, whereas therelative abundance of this transcript was increased in super-ovulated animals compared with control when analysed by
qPCR (P, 0.05). The stimulated and superovulated animalsshowed increased expression of HMGCR (Fig. 3) comparedwith control animals, although according to the microarrayresults, no difference was observed in the superovulated animals
(Table 3).Three important transcripts (HSD3B, CPY11A1 and STAR)
related to luteal steroidogenesis, which showed no significant
differences in expression in the microarray analysis, were alsoanalysed by qPCR. No significant differences in mRNA levelswere detected forHSD3B or CPY11A1 but the expression of the
STAR gene was increased in stimulated and superovulated(P, 0.05; Fig. 5) compared with control animals.
Protein localisation
HSD3B immunostaining was restricted to the cytoplasm ofsmall and large luteal cells, exhibiting the same intensity in allgroups (Fig. 6). For the P450scc protein, expression was alsoobserved in the cytoplasm of both luteal cell populations;
however, its expression seemed to be increased in the smallluteal cells of the treated groups. No positive staining couldbe observed in the endothelial or stroma cells of any of the
studied groups (Fig. 6). For STAR, FABP5 and HMCGR
immunostaining a secondary antibody conjugated with the
fluorophore CY3 was used and positive immunostaining wasobserved in the cytoplasm of small and large luteal cells. STARand FABP5 immunostaining was more evident in luteal cells
from the stimulated and superovulated groups, whereasHMGCR did not show any signal intensity difference amongthe groups (Fig. 7). Follistatin was detected in the small and
large luteal cells from all animals and the staining signalappeared to be greater in the cells from the stimulated andsuperovulated animals in relation to the control group (Fig. 7).No positive stainingwas observed in the negative control for any
of the proteins under study (Figs 6 and 7).
Protein levels assessed by western blot
Western blot analysis was performed to determine the asso-
ciation between the abundance of STAR, HSD3B, PPARG,follistatin, HMGCR, PRLRS, PRLRL and FABP5 proteins andmRNA abundance. The relative expression levels of these
proteins were calculated by normalising the results to ACTB(42 kDa). For STAR (Fig. 8b), a specific band of 31 kDa wasobserved and its expression was higher in stimulated and
superovulated animals compared with controls (P, 0.05).HSD3B (Fig. 8a) corresponds to a 30-kDa band and its levelswere not different among the groups. The immunoblots forPPARG, HMGCR and FABP5 proteins revealed bands at 52, 55
and 15 kDa, respectively. Densitometric analysis indicated anincrease of the three proteins in the corpora lutea of stimulatedand superovulated cows compared with controls (P, 0.05;
Fig. 9).
Control
HS
D3B
P45
0scc
Stimulated Superovulated
Fig. 6. HSD3B and P450scc expression in bovine CLL from Day 7 after gonadotrophin releasing hormone (GnRH) administration, detected by
immunohistochemistry. Positive signals can be observed as the orange–brown colour in the cytoplasm of the large (full arrow) and small (empty arrow)
luteal cells in the control, stimulated and superovulated animals. NC, negative control. Bars¼ 50mm.
H Reproduction, Fertility and Development L. A. Fatima et al.

PRLRS and PRLRL were detected using the same antibody,
which detected bands at ,40 and 100 kDa, respectively. Therelative expression of PRLRS showed a significant increase inthe stimulated and superovulated groups; however, PRLRL wasonly increased in the stimulated group (P, 0.05; Fig. 10a,b).
Follistatin comprised two bands of,55 kDa showing increasedexpression in stimulated and superovulated groups comparedwith the control (P, 0.05; Fig. 10c).
Discussion
In the present study we successfully identified many differen-tially expressed genes in the CL from stimulated and super-ovulated cows in relation to each other and to control cows.
Based on the biological functions and, in particular, the processof steroidogenesis, genes regulated by eCG treatment wereselected. It is important to highlight that eCG treatments were
Control
STA
RFA
BP
5H
MG
CR
Fol
lista
tin
Stimulated Superovulated
NC
NC
NC
NC
Fig. 7. STAR, FABP5, follistatin and HMGCR expression in bovine CLL from Day 7 after gonadotrophin releasing hormone (GnRH) administration,
detected by immunofluorescence. Positive signals (red for STAR, FABP5 and HMGCR or green for follistatin) can be observed in the cytoplasm of large (full
arrow) and small (empty arrow) luteal cells in control, stimulated and superovulated cows. NC, negative control. Bars¼ 50 mm.
eCG alters global gene expression of bovine CL Reproduction, Fertility and Development I

conducted in the follicle and we analysed the subsequentlyformed CL, indicating that observed alteration in luteal cell
machinery may be due to residual activity of eCG for severaldays and even to the reprogramming of the follicle, which ren-dered a more steroidogenic CL. We observed that CL volume
was higher in superovulated than in stimulated cows and higherin stimulated than in the control cows, and the P4 concentrationwas higher in stimulated animals than in control cows.
Furthermore, protocols using gonadotrophins to stimulate thelargest follicles consistently result in increased plasma proges-terone concentrations (Nogueira et al. 2004; Souza et al. 2009;Sa Filho et al. 2010). Thus, according to the literature and our
own findings, we chose to explore the pathways that mightelucidate the regulatory mechanisms that are affected by eCG inluteal cells.
Considering the augmentation of progesterone levels instimulated animals, i.e. in recipient cows for fixed-time embryotransfer, the first step was to verify the expression of key factors
related to steroidogenic activity. We observed that both STARprotein and mRNA expression were upregulated by eCG instimulated and superovulated animals, which most likely con-
tributed to the increase in P4 concentrations because STAR isconsidered to be the rate-limiting step in steroid biosynthesis(Lin et al. 1995; Stocco and Clark 1996; Strauss et al. 2003;Miller 2007). Furthermore, although small and large luteal cells
show intense P450scc and HSD3B activity (Wiltbank et al.
1993; Belfiore et al. 1994), none of these enzymes appeared tobe altered by eCG.
The main function of any steroid-producing cell is to main-tain a supply of cholesterol, and, in this context, most of thegenes that were selected for qPCR validation are related to lipid
biosynthesis and belong to the PPARG pathway. PPARGregulates the transcription of several target genes involved inovarian functions, such as STAR and steroidogenesis (Seto-Young et al. 2007), ovulation, oocyte maturation and CL
maintenance (Komar 2005). PPARG is active in several meta-bolic pathways, including lipid and cholesterol metabolism, and
it is considered to be the key regulator of lipid cellular homeo-stasis (Varga et al. 2011). In our study, the PPARG gene and
protein expression levels were higher in eCG-treated cows thanin non-treated cows, indicating that this transcription factormight participate in the activation of STAR expression and also
that of genes related to lipid metabolism in the CL from eCG-treated cows. Among the other PPARG target genes involved inlipid metabolism and regulated by eCG treatment are CD36,
FABP5, CYP27A1 and ACSF2. CD36 is a multifunctionalscavenger receptor that mediates endocytosis or selective cho-lesterol uptake from oxidised LDL and HDL lipoproteins. It isupregulated in bovine pre-ovulatory follicles in response to the
LH surge (Li et al. 2009). CD36 gene expression was increasedonly in eCG-stimulated cows and it may increase progesteroneoutput in these animals by increasing the levels of available
substrate. FABP5 was the most extensively upregulated of thedifferentially expressed genes in stimulated animals and it wasalso increased in superovulated CLs. FABP5 is a lipid carrier
related to PPARG. It further transports lipids to droplets forstorage, to the endoplasmic reticulum for signalling or to themitochondria for oxidation (Haunerland and Spener 2004;
Chmurzynska 2006; Furuhashi and Hotamisligil 2008). Addi-tionally, FABP5 gene knockdown results in decreased choles-terol and cholesterol ester levels in ARPE-19 cells (Wu et al.
2010) and FABP5 levels have been shown to increase in rat
ovaries after hCG induction of ovulation (Hennebold 2004).These findings point to new insights into the potential role ofFABP5 in CL function, which might be related to the supply or
transport of cholesterol and other lipids. The other two analysedgenes participating in lipid metabolism were CYP27 andACSF2, and the mRNA abundance of both genes was upregu-
lated in the CLs of stimulated cows compared with control ones.CYP27 catalyses the synthesis of 27-hydroxycholesterol, whichmay be regulated by steroidogenic activity in ovarian cells toensure the availability of steroid hormone precursors (Rennert
et al. 1990). On the other hand, ACSF2 belongs to the long-chainacyl-CoA synthetase family and catalyses the synthesis of
2.5
1.5
0.5
0
Control
Pro
tein
rel
ativ
e de
nsity
HS
D3B
Stimulated Superovulated Control
a
bb
Stimulated Superovulated
Control Stimulated Superovulated Control Stimulated Superovulated
2.0
1.0
1.4
0.2
0
Pro
tein
rel
ativ
e de
nsity
STA
R
1.0
1.2
0.4
0.6
0.8
HSD3B
ACTB
STAR
ACTB
30 kDa
42 kDa
31 kDa
42 kDa
(a) (b)
Fig. 8. Expression levels of steroidogenic proteins (a) HSD3B and (b) STAR in bovine CLL from Day 7 after gonadotrophin releasing hormone (GnRH)
administration in control, stimulated and superovulated animals. Electrophoresis images are representative of three independent experiments. The data were
normalised to ACTB expression and expressed as mean � s.e.m.; bars with different superscripts differ significantly (P, 0.05).
J Reproduction, Fertility and Development L. A. Fatima et al.

acyl-CoA, a substrate entering numerous pathways includingthat of de novo synthesis of fatty acids, synthesis of triacylgly-cerol and phospholipids, b-oxidation, and cholesterol esterifi-
cation (Coleman et al. 2002). Based on these findings, a locallipid metabolism regulatory mechanism may exist in the bovine
Control Stimulated Superovulated
Control Stimulated Superovulated
Control Stimulated Superovulated
Control Stimulated Superovulated
PPARG
ACTB
FABP5
ACTB
HMGCR
ACTB
52 kDa
42 kDa
52 kDa
42 kDa
55 kDa
42 kDa
a
(a)
(b)
(c)
a
b
b
a
b
1.0
1.2
1.0
0.8
0.6
0.4
0.2
0
0.8
0.6
0.4
Pro
tein
rel
ativ
e de
nsity
PPA
RG
Pro
tein
rel
ativ
e de
nsity
FAB
P5
Pro
tein
rel
ativ
e de
nsity
HM
GC
R
0.2
0
b
b
b
1.8
1.5
1.2
0.9
0.6
0.3
0
Fig. 9. Expression levels of the (a) PPARG, (b) FABP5 and (c) HMGCR
proteins in bovine CLL from Day 7 after gonadotrophin releasing
hormone (GnRH) administration in control, stimulated and superovulated
animals. Electrophoresis images are representative of three independent
experiments. The data were normalised to ACTB expression and expressed
as mean� s.e.m.; bars with different superscripts differ significantly
(P, 0.05).
0.30
0.25
0.20
a
a
a
b
Control(a)
(b)
(c)
Pro
tein
rel
ativ
e de
nsity
PR
LRS
Pro
tein
rel
ativ
e de
nsity
PR
LRL
Pro
tein
rel
ativ
e de
nsity
Fol
lista
tin
PRLRSACTB
Stimulated Superovulated
40 kDa42 kDa
b
b
0.15
0.10
0.05
0
2.5
1.5
1.0
0.5
2.0
0
Control
PRLRL
ACTB
Stimulated Superovulated
70 kDa42 kDa
1.0
0.8
0.6
0.4
0.2
Control
ACTB
Stimulated Superovulated
Control Stimulated Superovulated
100 kDa
42 kDa
0
b
a
b
Follistatin
Fig. 10. Expression levels of (a) PRLRS, (b) PRLRL and (c) follistatin
proteins in bovine CLL from Day 7 after gonadotrophin releasing hormone
(GnRH) administration in control, stimulated and superovulated cows.
Electrophoresis runs are representative of three independent experiments.
The data were normalised to ACTB expression and expressed as mean�s.e.m.; bars with different superscripts differ significantly (P, 0.05).
eCG alters global gene expression of bovine CL Reproduction, Fertility and Development K

CL, and it might be enhanced by eCG, improving the efficiencyor magnitude of steroid synthesis.
Additionally or alternatively, de novo synthesis of cholester-ol may be a further means of increasing cholesterol availabilityfor luteal steroidogenesis. HMGCR is the rate-limiting enzyme
in this process (Gwynne and Strauss 1982). Its gene and proteinexpression were both upregulated in the CLL of stimulated andsuperovulated animals and it has been reported that FSH
increases HMGCR expression in granulosa cells (Liu et al.
2009). HMGCR activity is usually regulated by a negativefeedback mechanism and it involves sterol regulation of itsgene promoter (Golos and Strauss 1988). Thus, the regulation of
progesterone production in the CL by eCG treatment appears tobemediated, at least in part, by increases in HMGCR expressionand cholesterol synthesis.
Interestingly, our study showed a lower expression ofNCEH1 in corpora lutea of eCG-treated cows. NCEH1 isimportant for the hydrolysis of cholesterol esters found in lipid
droplets and its activity can be regulated within steroidogenictissues by hormones including FSH, LH and hCG (Trzeciaket al. 1984; Kraemer et al. 1993). However, NCEH1 is notdynamically regulated in luteal cells and therefore it may not
limit steroidogenesis. Targeted deletion of the NCEH gene doesnot result in accumulated cholesterol esters in steroidogeniccells, suggesting the existence of alternative genes encoding
cholesterol esterase activity (Chung et al. 2001).Another upregulated gene of particular interest was PRLR
because prolactin (PRL) can affect luteal progesterone produc-
tion (Stocco 2012) and also regulate cell differentiation andproliferation in diverse tissues (Bole-Feysot et al. 1998). Whilethe evidence for a role for PRL in the bovine ovary is controver-
sial, in rodents, where the requirement for luteal prolactinstimulation is well established, CL cells from PRLR knockoutmice failed to organise appropriately and underwent dramaticapoptosis (Grosdemouge et al. 2003). The PRLRS and PRLRL
mRNA and protein levels were upregulated in stimulatedanimals, whereas only the PRLRS protein was increased insuperovulated animals. These results point towards a possible
role of eCG in the regulation of PRLR expression and theysuggest that the expression of each PRLR isoform can beregulated independently (Picazo et al. 2004). Furthermore, the
increase in PRLRS expression in superovulated animals couldbe related to its action on cell proliferation and angiogenesis(Bole-Feysot et al. 1998; Stocco 2012), whereas the increase ofPRLRL in stimulated animals could be related to prolactin’s
effect on luteal progesterone production (Stocco 2012).To gain more insights regarding the influence of eCG on CL
function, the follistatin and TGFB2 genes were also analysed.
Follistatin canmodulate the function of granulosa cells in favourof luteinisation and may also directly modulate luteal progester-one production (Findlay 1993; Hillier andMiro 1993; Kaipainen
et al. 1995). Moreover, in cows, the dominant follicle containedmore follistatin than the corresponding subordinate follicles,and, in the CL, the highest expression of follistatin was detected
on the day of maximum CL activity (Singh and Adams 1998).The eCG treatment upregulated follistatin gene and proteinexpression in stimulated and superovulated animals, suggestingthat follistatin could be involved in the final stages of follicle and
luteal development in the eCG-treated animals. TGFB2 is amultifunctional growth factor that mediates several physiologi-
cal processes related to cellular function (Roberts and Skinner1991), such as follicle growth, cell proliferation and enhancedformation of the extracellular matrix (Lawrence 1996). In the
CL, TGFB2 is mainly expressed by small luteal cells (Sriper-umbudur et al. 2010) and macrophages (Matsuyama andTakahashi 1995). Moreover, TGFB2 mediates the luteotrophic
action of prolactin in the rat CL (Matsuyama and Takahashi1995) and is able to increase progesterone production by thecacells in cattle (Roberts and Skinner 1991). Indeed, the specificfunction of TGFB2 in the bovine CL is not well established.
Here, TGFB2 gene expression was upregulated in the CL fromeCG-stimulated animals, and these results provide additionalinformation about the possible involvement of TGFB2 in bovine
CL growth and remodelling under regulation of eCG.In summary, our findings provide important insights into
potential eCG targets during bovine CL development, mainte-
nance and progesterone production. Equine CG stimulated theexpression of steroidogenic proteins such as STAR, as well asthe regulation of cholesterol biosynthesis and structural-relatedgenes. Differential expression of these genes is proposed as an
important cause that leads to an increase of CL volume andimprovement in luteal progesterone production.
Acknowledgement
The authors are thankful for FAPESP grants 2008/58837–5 and 2008/
58839–8.
References
Ambrose, J. D., Drost, M., Monson, R. L., Rutledge, J. J., Leibfried-
Rutledge, M. L., Thatcher, M. J., Kassa, T., Binelli, M., Hansen, P. J.,
Chenoweth, P. J., and Thatcher,W.W. (1999). Efficacy of timed embryo
transfer with fresh and frozen in vitro-produced embryos to increase
pregnancy rates in heat-stressed dairy cattle. J. Dairy Sci. 82(11),
2369–2376. doi:10.3168/JDS.S0022-0302(99)75487-1
Baruselli, P. S., Ferreira, R. M., Sales, J. N., Gimenes, L. U., Sa Filho, M. F.,
Martins, C. M., Rodrigues, C. A., and Bo, G. A. (2011). Timed embryo
transfer programs for management of donor and recipient cattle. Ther-
iogenology 76(9), 1583–1593. doi:10.1016/J.THERIOGENOLOGY.
2011.06.006
Baruselli, P. S., Sa Filho, M. F., Ferreira, R. M., Sales, J., Gimenes, L. U.,
Vieira, L., Mendanha,M., and Bo, G. A. (2012).Manipulation of follicle
development to ensure optimal oocyte quality and conception rates in
cattle. Reprod. Domest. Anim. 47(4), 134–141. doi:10.1111/J.1439-
0531.2012.02067.X
Belfiore, C. J., Hawkins, D. E., Wiltbank, M. C., and Niswender, G. D.
(1994). Regulation of cytochrome P450scc synthesis and activity in the
ovine corpus luteum. J. Steroid Biochem. Mol. Biol. 51(5–6), 283–290.
doi:10.1016/0960-0760(94)90041-8
Binelli, M., Thatcher, W. W., Mattos, R., and Baruselli, P. S. (2001).
Antiluteolytic strategies to improve fertility in cattle. Theriogenology
56(9), 1451–1463. doi:10.1016/S0093-691X(01)00646-X
Bo, G. A., Baruselli, P. S., Moreno, D., Cutaia, L., Caccia, M., Trıbulo, R.,
Trıbulo, H., and Mapletoft, R. J. (2002). The control of follicular wave
development for self-appointed embryo transfer programs in cattle.
Theriogenology 57(1), 53–72. doi:10.1016/S0093-691X(01)00657-4
Bole-Feysot, C., Goffin, V., Edery, M., Binart, N., and Kelly, P. A. (1998).
Prolactin (PRL) and its receptor: actions, signal transduction pathways
L Reproduction, Fertility and Development L. A. Fatima et al.

and phenotypes observed in PRL receptor knockout mice. Endocr. Rev.
19(3), 225–268. doi:10.1210/ER.19.3.225
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye
binding.Anal. Biochem. 72, 248–254. doi:10.1016/0003-2697(76)90527-3
Brannian, J. D., and Stouffer, R. L. (1993). Native andmodified (acetylated)
low-density lipoprotein-supported steroidogenesis by macaque granu-
losa cells collected before and after the ovulatory stimulus: correlation
with fluorescent lipoprotein uptake. Endocrinology 132(2), 591–597.
doi:10.1210/EN.132.2.591
Chmurzynska, A. (2006). The multigene family of fatty acid-binding
proteins (FABPs): function, structure and polymorphism. J. Appl. Genet.
47(1), 39–48. doi:10.1007/BF03194597
Christenson, L. K., and Devoto, L. (2003). Cholesterol transport and
steroidogenesis by the corpus luteum. Reprod. Biol. Endocrinol. 1, 90.
doi:10.1186/1477-7827-1-90
Chung, S.,Wang, S. P., Pan, L.,Mitchell,G., Trasler, J., andHermo, L. (2001).
Infertility and testicular defects in hormone-sensitive lipase-deficient
mice. Endocrinology 142(10), 4272–4281. doi:10.1210/EN.142.10.4272
Clemente, M., de La Fuente, J., Fair, T., Al Naib, A., Gutierrez-Adan, A.,
Roche, J. F., Rizos, D., and Lonergan, P. (2009). Progesterone and
conceptus elongation in cattle: a direct effect on the embryo or an
indirect effect via the endometrium? Reproduction 138(3), 507–517.
doi:10.1530/REP-09-0152
Coleman, R. A., Lewin, T. M., Van Horn, C. G., and Gonzalez-Baro, M. R.
(2002). Do long-chain acyl-CoA synthetases regulate fatty acid entry
into synthetic versus degradative pathways? J. Nutr. 132(8), 2123–2126.
Devoto, L., Vega, M., Kohen, P., Castro, A., Castro, O., Christenson, L. K.,
Carvallo, P., and Strauss, J. F., 3rd (2000). Endocrine and paracrine–
autocrine regulation of the human corpus luteum during the mid-luteal
phase. J. Reprod. Fertil. Suppl. 55, 13–20.
Doody, K. J., Lorence, M. C., Mason, J. I., and Simpson, E. R. (1990).
Expression of messenger ribonucleic acid species encoding steroidogenic
enzymes in human follicles and corpora lutea throughout the menstrual
cycle. J. Clin. Endocrinol. Metab. 70(4), 1041–1045. doi:10.1210/
JCEM-70-4-1041
Fields, S. D., Gebhart, K. L., Perry, B. L., Gonda, M. G.,Wright, C. L., Bott,
R. C., and Perry, G. A. (2012). Influence of standing oestrus before an
injection of GnRH during a beef cattle fixed-time AI protocol on LH
release, subsequent concentrations of progesterone and steroidogenic
enzyme expression. Domest. Anim. Endocrinol. 42(1), 11–19.
doi:10.1016/J.DOMANIEND.2011.08.002
Findlay, J. K. (1993). An update on the roles of inhibin, activin and follistatin
as local regulators of folliculogenesis. Biol. Reprod. 48(1), 15–23.
doi:10.1095/BIOLREPROD48.1.15
Furuhashi, M., and Hotamisligil, G. S. (2008). Fatty acid-binding proteins:
role in metabolic diseases and potential as drug targets. Nat. Rev. Drug
Discov. 7(6), 489–503. doi:10.1038/NRD2589
Garcia-Bojalil, C.M., Staples, C. R., Risco, C. A., Savio, J. D., and Thatcher,
W.W. (1998). Protein degradability and calcium salts of long-chain fatty
acids in the diets of lactating dairy cows: reproductive responses. J.
Dairy Sci. 81(5), 1385–1395. doi:10.3168/JDS.S0022-0302(98)75702-9
Golos, T. G., and Strauss, J. F. (1988). 8-bromoadenosine cyclic 30,50-phosphate rapidly increases 3-hydroxy-3-methylglutaryl coenzyme A
reductasemRNA in human granulosa cells: role of cellular sterol balance
in controlling the response to tropic stimulation. Biochemistry 27(9),
3503–3506. doi:10.1021/BI00409A056
Grosdemouge, I., Bachelot, A., Lucas, A., Baran, N., Kelly, P. A., and Binart,
N. (2003). Effects of deletion of the prolactin receptor on ovarian gene
expression. Reprod. Biol. Endocrinol. 1, 12. doi:10.1186/1477-7827-1-12
Gwynne, J. T., and Strauss, J. F. (1982). The role of lipoproteins in
steroidogenesis and cholesterol metabolism in steroidogenic glands.
Endocr. Rev. 3(3), 299–329. doi:10.1210/EDRV-3-3-299
Hansen, P. J., Drost, M., Rivera, R. M., Paula-Lopes, F. F., al-Katanani,
Y. M., Krininger, C. E., 3rd, and Chase, C. C., Jr (2001). Adverse impact
of heat stress on embryo production: causes and strategies formitigation.
Theriogenology 55(1), 91–103. doi:10.1016/S0093-691X(00)00448-9
Haunerland, N. H., and Spener, F. (2004). Fatty acid-binding proteins –
insights from genetic manipulations. Prog. Lipid Res. 43(4), 328–349.
doi:10.1016/J.PLIPRES.2004.05.001
Hennebold, J.D. (2004).Characterizationof the ovarian transcriptome through
the use of differential analysis of gene expression methodologies. Hum.
Reprod. Update 10(3), 227–239. doi:10.1093/HUMUPD/DMH017
Hillier, S. G., and Miro, F. (1993). Inhibin, activin and follistatin. Potential
roles in ovarian physiology. Ann. N. Y. Acad. Sci. 687, 29–38.
doi:10.1111/J.1749-6632.1993.TB43850.X
Juengel, J. L., Nett, T. M., Anthony, R. V., and Niswender, G. D. (1997).
Effects of luteotrophic and luteolytic hormones on expression of mRNA
encoding insulin-like growth factor I and growth hormone receptor in the
ovine corpus luteum. J. Reprod. Fertil. 110(2), 291–298. doi:10.1530/
JRF.0.1100291
Kaipainen, A., Korhonen, J., Mustonen, T., van Hinsbergh, V. W., Fang,
G. H., Dumont, D., Breitman, M., and Alitalo, K. (1995). Expression of
the fms-like tyrosine kinase 4 gene becomes restricted to lymphatic
endothelium during development. Proc. Natl. Acad. Sci. USA 92(8),
3566–3570. doi:10.1073/PNAS.92.8.3566
Kliewer, S. A., Sundseth, S. S., Jones, S. A., Brown, P. J., Wisely, G. B.,
Koble, C. S., Devchand, P., Wahli, W., Willson, T. M., Lenhard, J. M.,
and Lehmann, J. M. (1997). Fatty acids and eicosanoids regulate gene
expression through direct interactions with peroxisome proliferator-
activated receptors alpha and gamma. Proc. Natl. Acad. Sci. USA 94,
4318–4323.
Komar, C. M. (2005). Peroxisome proliferator-activated receptors (PPARs)
and ovarian function – implications for regulating steroidogenesis,
differentiation and tissue remodelling. Reprod. Biol. Endocrinol. 3, 41.
doi:10.1186/1477-7827-3-41
Kraemer, F. B., Patel, S., Singh-Bist, A., Gholami, S. S., Saedi, M. S., and
Sztalryd, C. (1993). Detection of hormone-sensitive lipase in various
tissues. II. Regulation in the rat testis by human chorionic gonadotro-
phin. J. Lipid Res. 34(4), 609–616.
Labrie, F., Simard, J., Luu-The,V., Pelletier, G., Belanger, A., Lachance, Y.,
Zhao, H.F., Labrie, C., Breton, N., de Launoit, Y., et al. (1992). Structure
and tissue-specific expression of 3 beta-hydroxysteroid dehydrogenase/
5-ene-4-ene isomerase genes in human and rat classical and peripheral
steroidogenic tissues. J. Steroid Biochem. Mol. Biol. 41(3–8), 421–435.
doi:10.1016/0960-0760(92)90368-S
Lawrence, D. A. (1996). Transforming growth factor-beta: a general review.
Eur. Cytokine Netw. 7(3), 363–374.
Li, Q., Jimenez-Krassel, F., Ireland, J. J., and Smith, G. W. (2009). Gene
expression profiling of bovine preovulatory follicles: gonadotrophin
surge and prostanoid-dependent up-regulation of genes potentially
linked to the ovulatory process. Reproduction 137(2), 297–307.
doi:10.1530/REP-08-0308
Lin, D., Sugawara, T., Strauss, J. F., Clark, B. J., Stocco, D. M., Saenger, P.,
Rogol, A., and Miller, W. L. (1995). Role of steroidogenic acute
regulatory protein in adrenal and gonadal steroidogenesis. Science 267
(5205), 1828–1831. doi:10.1126/SCIENCE.7892608
Liu, Z., Rudd, M. D., Hernandez-Gonzalez, I., Gonzalez-Robayna, I., Fan,
H. Y., Zeleznik, A. J., and Richards, J. S. (2009). FSH and FOXO1
regulate genes in the sterol/steroid and lipid biosynthetic pathways in
granulosa cells. Mol. Endocrinol. 23(5), 649–661. doi:10.1210/
ME.2008-0412
Madureira, E. H. (2004). Sincronizacao com progestagenos. Biotecnologia
da reproducao em bovinos 1, 117–128.
Matsuyama, S., andTakahashi,M. (1995). Immunoreactive (ir)-transforming
growth factor (TGF)-beta in rat corpus luteum: ir-TGF beta is expressed
eCG alters global gene expression of bovine CL Reproduction, Fertility and Development M

by luteal macrophages. Endocr. J. 42(2), 203–217. doi:10.1507/
ENDOCRJ.42.203
Miller, W. L. (1988). Molecular biology of steroid hormone synthesis.
Endocr. Rev. 9(3), 295–318. doi:10.1210/EDRV-9-3-295
Miller,W. L. (2007). Steroidogenic acute regulatory protein (StAR), a novel
mitochondrial cholesterol transporter. Biochim. Biophys. Acta 1771(6),
663–676. doi:10.1016/J.BBALIP.2007.02.012
Murphy, B.D., andMartinuk, S.D. (1991). Equine chorionic gonadotrophin.
Endocr. Rev. 12(1), 27–44. doi:10.1210/EDRV-12-1-27
Niswender, G. D. (2002). Molecular control of luteal secretion of progester-
one. Reproduction 123(3), 333–339. doi:10.1530/REP.0.1230333
Niswender, G. D., and Nett, T. M. (1994). The corpus luteum and its control
in infraprimate species. In ‘The Physiology of Reproduction’. (Eds
E. Knobil and J. D. Neill.) pp. 781–816. (Roven Press 1: New York.)
Niswender,G. D., Juengel, J. L.,McGuire,W. J., Belfiore,C. J., andWiltbank,
M.C. (1994).Luteal function: the oestrous cycle and earlypregnancy.Biol.
Reprod. 50(2), 239–247. doi:10.1095/BIOLREPROD50.2.239
Nogueira, M. F., Melo, D. S., Carvalho, L. M., Fuck, E. J., Trinca, L. A., and
Barros, C. M. (2004). Do high progesterone concentrations decrease
pregnancy rates in embryo recipients synchronized with PGF2alpha and
eCG? Theriogenology 61(7–8), 1283–1290. doi:10.1016/J.THERIO
GENOLOGY.2003.07.012
Papa, P. C., Moura, C. E., Artoni, L. P., Fatima, L. A., Campos, D. B.,
Marques, J. E., Baruselli, P. S., Binelli, M., Pfarrer, C., and Leiser, R.
(2007). VEGF system expression in different stages of oestrous cycle in
the corpus luteum of non-treated and superovulated water buffalo.
Domest. Anim. Endocrinol. 33(4), 379–389. doi:10.1016/J.DOMA
NIEND.2006.07.003
Pfaffl, M. W. (2001). A new mathematical model for relative quantification
in real-time RT-PCR. Nucleic Acids Res. 29(9), e45. doi:10.1093/NAR/
29.9.E45
Picazo, R. A., Garcıa Ruiz, J. P., Santiago Moreno, J., Gonzalez de Bulnes,
A.,Munoz, J., Silvan, G., Lorenzo, P. L., and Illera, J. C. (2004). Cellular
localization and changes in expression of prolactin receptor isoforms in
sheep ovary throughout the oestrous cycle. Reproduction 128(5),
545–553. doi:10.1530/REP.1.00343
Ramakers, C., Ruijter, J. M., Deprez, R. H., and Moorman, A. F. (2003).
Assumption-free analysis of quantitative real-time polymerase chain
reaction (PCR) data. Neurosci. Lett. 339(1), 62–66. doi:10.1016/S0304-
3940(02)01423-4
Rennert, H., Fischer, R. T., Alvarez, J. G., Trzaskos, J. M., and Strauss, J. F.
(1990). Generation of regulatory oxysterols: 26-hydroxylation of cho-
lesterol by ovarian mitochondria. Endocrinology 127(2), 738–746.
doi:10.1210/ENDO-127-2-738
Richards, R. G., and Almond, G. W. (1994). Tumour necrosis factor-alpha
differentially alters progesterone and prostaglandin F2 alpha production
by porcine luteal cells. J. Endocrinol. 143(1), 75–83. doi:10.1677/JOE.0.
1430075
Roberts, A. J., and Skinner,M. K. (1991). Transforming growth factor-alpha
and -beta differentially regulate growth and steroidogenesis of bovine
thecal cells during antral follicle development. Endocrinology 129(4),
2041–2048. doi:10.1210/ENDO-129-4-2041
Sa Filho, M. F., Torres-Junior, J. R., Penteado, L., Gimenes, L. U., Ferreira,
R. M., Ayres, H., Castro, E. P. L. A., Sales, J. N., and Baruselli, P. S.
(2010). Equine chorionic gonadotrophin improves the efficacy of a
progestin-based fixed-time artificial insemination protocol in Nelore
(Bos indicus) heifers. Anim. Reprod. Sci. 118(2–4), 182–187.
doi:10.1016/J.ANIREPROSCI.2009.10.004
Sales, J. N., Crepaldi, G. A., Girotto, R.W., Souza, A. H., andBaruselli, P. S.
(2011). Fixed-timeAI protocols replacing eCGwith a single dose of FSH
were less effective in stimulating follicular growth, ovulation and
fertility in suckled–anoestrus Nelore beef cows. Anim. Reprod. Sci.
124(1–2), 12–18. doi:10.1016/J.ANIREPROSCI.2011.02.007
Seto-Young,D., Avtanski, D., Strizhevsky,M., Parikh, G., Patel, P., Kaplun,
J., Holcomb, K., Rosenwaks, Z., and Poretsky, L. (2007). Interactions
among peroxisome proliferator-activated receptor-gamma, insulin sig-
nalling pathways and steroidogenic acute regulatory protein in human
ovarian cells. J. Clin. Endocrinol. Metab. 92(6), 2232–2239.
doi:10.1210/JC.2006-1935
Singh, J., and Adams, G. P. (1998). Immunohistochemical distribution of
follistatin in dominant and subordinate follicles and the corpus luteum of
cattle.Biol. Reprod. 59(3), 561–570. doi:10.1095/BIOLREPROD59.3.561
Souza, A. H., Viechnieski, S., Lima, F. A., Silva, F. F., Araujo, R., Bo, G. A.,
Wiltbank, M. C., and Baruselli, P. S. (2009). Effects of equine chorionic
gonadotrophin and type of ovulatory stimulus in a timed AI protocol on
reproductive responses in dairy cows. Theriogenology 72(1), 10–21.
doi:10.1016/J.THERIOGENOLOGY.2008.12.025
Sriperumbudur, R., Zorrilla, L., and Gadsby, J. E. (2010). Transforming
growth factor-beta (TGFbeta) and its signalling components in peri-
ovulatory pig follicles. Anim. Reprod. Sci. 120(1–4), 84–94.
doi:10.1016/J.ANIREPROSCI.2010.03.003
Stocco, C. (2012). The long and short of the prolactin receptor: the corpus
luteum needs them both! Biol. Reprod. 86(3), 1–2. doi:10.1095/
BIOLREPROD.111.098293
Stocco, D. M., and Clark, B. J. (1996). Role of the steroidogenic acute
regulatory protein (StAR) in steroidogenesis. Biochem. Pharmacol.
51(3), 197–205. doi:10.1016/0006-2952(95)02093-4
Stocco, C. O., Chedrese, J., and Deis, R. P. (2001). Luteal expression of
cytochrome P450 side-chain cleavage, steroidogenic acute regulatory
protein, 3beta-hydroxysteroid dehydrogenase and 20alpha-
hydroxysteroid dehydrogenase genes in late pregnant rats: effect of
luteinizing hormone and RU486. Biol. Reprod. 65(4), 1114–1119.
doi:10.1095/BIOLREPROD65.4.1114
Strauss, J. F., Kishida, T., Christenson, L. K., Fujimoto, T., and Hiroi, H.
(2003). START domain proteins and the intracellular trafficking of
cholesterol in steroidogenic cells. Mol. Cell. Endocrinol. 202(1–2),
59–65. doi:10.1016/S0303-7207(03)00063-7
Trzeciak, W. H., Sonnenborn, U., Balkow, C., and Kunau, W. H. (1984).
Regulation of steroidogenesis in rat adrenal gland: identification of the
bifunctional, hormone-sensitive cholesterol esterase–triacylglycerol
lipase enzyme protein and its discrimination from hormone-insensitive
lipases. Mol. Cell. Endocrinol. 35(2–3), 131–141. doi:10.1016/0303-
7207(84)90009-1
Varga, T., Czimmerer, Z., and Nagy, L. (2011). PPARs are a unique set of
fatty acid-regulated transcription factors controlling both lipid metabo-
lism and inflammation. Biochim. Biophys. Acta 1812(8), 1007–1022.
doi:10.1016/J.BBADIS.2011.02.014
Webb, R., Woad, K. J., and Armstrong, D. G. (2002). Corpus luteum (CL)
function: local control mechanisms. Domest. Anim. Endocrinol.
23(1–2), 277–285. doi:10.1016/S0739-7240(02)00163-7
Wildman, E. E. (1982). Dairy cow body-condition scoring system and its
relationship to selected production characteristics. J. Dairy Sci. 65,
495–501. doi:10.3168/JDS.S0022-0302(82)82223-6
Wiltbank,M. C., Belfiore, C. J., andNiswender, G. D. (1993). Steroidogenic
enzyme activity after acute activation of protein kinase (PK) A and PKC
in ovine small and large luteal cells.Mol. Cell. Endocrinol. 97(1–2), 1–7.
doi:10.1016/0303-7207(93)90205-X
Wright, G. W., and Simon, R. M. (2003). A random variance model for
detection of differential gene expression in small microarray experi-
ments. Bioinformatics 19(18), 2448–2455. doi:10.1093/BIOINFOR
MATICS/BTG345
Wu, T., Tian, J., Cutler, R. G., Telljohann, R. S., Bernlohr, D. A., Mattson,
M. P., and Handa, J. T. (2010). Knockdown of FABP5mRNA decreases
cellular cholesterol levels and results in decreased apoB100 secretion
and triglyceride accumulation in ARPE-19 cells. Lab. Invest. 90(6),
963–965. doi:10.1038/LABINVEST.2010.87
www.publish.csiro.au/journals/rfd
N Reproduction, Fertility and Development L. A. Fatima et al.