glyc osami nog lycans
DESCRIPTION
glycosaminoglycansTRANSCRIPT

M
Gt
DB
a
ARRA
KPGPTE
1
tio(itmBtg(furgottd
h1
The International Journal of Biochemistry & Cell Biology 68 (2015) 87–91
Contents lists available at ScienceDirect
The International Journal of Biochemistry& Cell Biology
jo ur nal home page: www.elsev ier .com/ locate /b ioce l
olecules in focus
lycosaminoglycans: Sorting determinants in intracellular proteinraffic
eyan Mihov ∗, Martin Spiessiozentrum, University of Basel, Basel, Switzerland
r t i c l e i n f o
rticle history:eceived 26 June 2015eceived in revised form 29 July 2015ccepted 27 August 2015
eywords:
a b s t r a c t
Intracellular transport of proteins to their appropriate destinations is crucial for the maintenance of cel-lular integrity and function. Sorting information is contained either directly in the amino acid sequenceor in a protein’s post-translational modifications. Glycosaminoglycans (GAGs) are characteristic modifi-cations of proteoglycans. GAGs are long unbranched polysaccharide chains with unique structural andfunctional properties also contributing to protein sorting in various ways. By deletion or insertion of
roteoglycanslycosaminoglycansrotein transportGN exportndocytosis
GAG attachment sites it has been shown that GAGs affect polarized sorting in epithelial cells, target-ing to and storage in secretory granules, and endocytosis. Most recently, the role of GAGs as signals forrapid trans-Golgi-to-cell surface transport, dominant over the cytosolic sorting motifs in the core protein,was demonstrated. Here, we provide an overview on existing data on the roles of GAGs on protein andproteoglycan trafficking.
© 2015 Elsevier Ltd. All rights reserved.
. Introduction
Many physiological functions depend on the proper localiza-ion of proteins to specific cellular compartments. The sortingnformation is contained directly within the amino acid sequencer alternatively in the protein’s posttranslational modificationsBonifacino and Traub, 2003; Seaman, 2008). Even glycosylation,.e. modifications in the lumen of the endoplasmic reticulum (ER) orhe Golgi compartments and thus without direct access to cytoplas-
ic machineries, has been shown to act as sorting determinants.est understood is late endosomal/lysosomal transport of pro-eins carrying the mannose-6-phosphate modification of N-linkedlycans by interaction with the mannose-6-phosphate receptorsGhosh et al., 2003). In addition, N- and O-linked glycans have beenound to mediate apical sorting in polarized cells by incompletelynderstood mechanisms most-probably involving lectins as cargoeceptors (Potter et al., 2006; Vagin et al., 2009). Since N- and O-lycosylation are also found on basolateral proteins, only a subsetf carbohydrate chains seems to be responsible for polarized sor-ing (Kitagawa et al., 1994). Finally, glycosaminoglycans (GAGs),
he defining modification of proteoglycans, are emerging as sortingeterminants for various transport steps.∗ Corresponding author.E-mail address: [email protected] (D. Mihov).
ttp://dx.doi.org/10.1016/j.biocel.2015.08.019357-2725/© 2015 Elsevier Ltd. All rights reserved.
2. Structure
Unlike N- and O-linked glycans, GAGs are long unbranchedpolysaccharide chains made of two alternating monosaccharides(Fig. 1A), an uronic acid or galactose, and an amino sugar (Mikamiand Kitagawa, 2013; Sarrazin et al., 2011). Structurally, GAGs areextended semi-rigid polymers adopting helical conformation withan axial rise of about 1 nm per disaccharide unit (Almond andSheehan, 2000; Rodriguez-Carvajal et al., 2003). The length ofGAG chains is variable, typically from 20 to 60 kDa (40–120 dis-accharides) in proteoglycans and 6 to 34 kDa for xyloside-primedprotein-free chains (Victor et al., 2009).
3. Synthesis and degradation
GAG chains synthesis is initiated in the early secretory path-way by formation of a tetra-saccharide linker on serine residues(Fig. 1B), except for keratan sulfate chains which are initiated on N-or O-linked glycans. Chain extension, deacetylation and epimeriza-tion continue during the transport through the Golgi complex. Inthe trans-Golgi and the trans-Golgi network (TGN), GAGs are sul-
fated at specific positions (Mikami and Kitagawa, 2013; Sarrazinet al., 2011). Degradation of GAG chains takes place in the lyso-somes by the help of endo-type hydrolases, exolytic glycosidasesand sulfatases to liberate monosaccharide moieties.
88 D. Mihov, M. Spiess / The International Journal of Biochemistry & Cell Biology 68 (2015) 87–91
Fig. 1. Structure and synthesis of glycosaminoglycan chains. GAGs are long unbranched polysaccharide chains made of the following monosaccharides: xylose (Xyl), galac-tose (Gal), glucuronic acid (GlcA), iduronic acid (IdoA), N-acetyl-glucosamine (GlcNAc), N-acetyl-galactosamine (GalNAc). (A) The GAG chains consist of long repeats ofdisaccharide units, whose composition is specific for the different GAG types: CS (–4GlcA�1–3GalNAc�1–), DS (–4IdoA�1–3GalNAc�1–), KS (–3Gal�1–4GlcNAc�1–), HS(–4GlcA�1–4GlcNAc�1–) and heparin (–4IdoA�1–4GlcNAc�1–). GAGs can be sulfated at different sites (orange arrowheads) and the sulfation pattern further increases thec of a tT atan so sion o
4
GDta2alIfiv
4
fmatacaca1m
omplexity of their structures. (B) The synthesis of GAGs is initiated by the generationhis linker is common for CS/DS/HS/heparin proteoglycans with the exception of kerf the references to color in this figure legend, the reader is referred to the web ver
. Biological function: GAGs as sorting determinants
Due to their many carboxylate groups and extensive sulfation,AG chains attract water and cations forming hydrated matrices.istinct saccharide and sulfation patterns allow specific interac-
ions with various ligands, thus regulating cell signaling, migration,nd differentiation (Mikami and Kitagawa, 2013; Sarrazin et al.,011). Proteoglycans are prominent components at the cell surfacend of the extracellular matrix (ECM), but are also found intracellu-arly in secretory granules and in the endosomal–lysosomal system.nitially, the protein core was believed to be solely responsibleor the correct proteoglycan localization. However, several stud-es have demonstrated that GAGs contribute to protein traffic in aariety of ways (Fig. 2).
.1. Polarized surface expression
In polarized epithelial cells, proteoglycans are found to be dif-erentially distributed to the apical and basolateral sides of the cell
onolayer, reflecting their specific roles on either surface. Hep-ran sulfate (HS) modification was proposed to mediate basolateralransport, since glypican, a glycosylphosphatidylinositol-anchorednd predominantly basolateral proteoglycan in CaCo-2 and MDCKells, was transported strictly to the apical surface, when its HSttachment sites were mutated (Mertens et al., 1996). On the
ontrary, chondroitin sulfate (CS) proteoglycans and free xyloside-ttached CS were mainly found to be secreted apically (Kolset et al.,999). Rat growth hormone, normally secreted in a non-polarizedanner from MDCK cells, was found predominantly in the apicaletra-saccharide linked to a serine residue: GlcA�1–3Gal�1–3Gal�1–4Xyl�1–O-Ser.ulfate, which can be generated on both N- and O-linked glycans. (For interpretationf this article.)
medium upon modification with a sequence containing multipleCS attachment sites (Hafte et al., 2011). In the contrary, how-ever, fusion of a single CS attachment site to the asialoglycoproteinreceptor H1 (ASGPR-H1), which is targeted to the basolateral sur-face by a cytoplasmic tyrosine motif interacting with the clathrinadaptor AP-1B (Sugimoto et al., 2002), did not alter its polarity(Kobialka et al., 2009). Similarly, natural splice variants of amy-loid precursor-like protein 2 (APLP2) with and without a singleCS chain were equally targeted to the basolateral surface, both asmembrane-bound and secretory forms (Lo et al., 1995). The baso-lateral sorting information in the cytoplasmic portions of theseproteins thus is dominant over the apical signal of the CS chain.Alternatively, the affinity to the apical transport machinery mightdepend on the number of GAG chains.
4.2. Biosynthetic export
To analyze the mechanism of proteoglycan export from the Golgito the cell surface, HS proteoglycan-containing post-Golgi vesicleswere isolated from [35S]sulfate-labeled rat hepatocytes (Barthelet al., 1995; Nickel et al., 1994). Surprisingly, other secretoryproteins, such as serum albumin, apolipoprotein E and fibrino-gen, were not detected in this vesicle fraction, indicating thatHS-proteoglycans are sorted into distinct types of constitutivesecretory carriers at the TGN.
Additional evidence for GAG-mediated sorting in biosyntheticexport was obtained by comparison of the transport kinetics of pro-teins with and without GAG modification (Kobialka et al., 2009).The ASGPR-H1 and �1-protease inhibitor (A1Pi) were used as
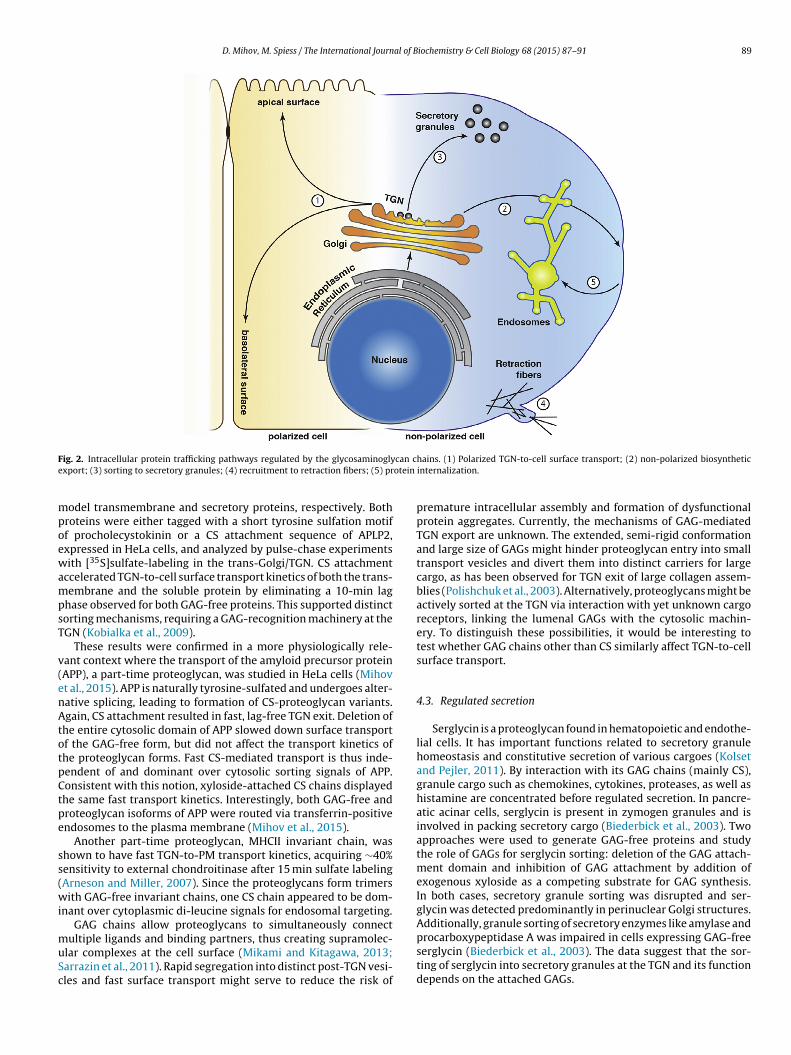
D. Mihov, M. Spiess / The International Journal of Biochemistry & Cell Biology 68 (2015) 87–91 89
F can ce otein
mpoewampsT
v(enAtotpCtpe
ss(wi
muSc
ig. 2. Intracellular protein trafficking pathways regulated by the glycosaminoglyxport; (3) sorting to secretory granules; (4) recruitment to retraction fibers; (5) pr
odel transmembrane and secretory proteins, respectively. Bothroteins were either tagged with a short tyrosine sulfation motiff procholecystokinin or a CS attachment sequence of APLP2,xpressed in HeLa cells, and analyzed by pulse-chase experimentsith [35S]sulfate-labeling in the trans-Golgi/TGN. CS attachment
ccelerated TGN-to-cell surface transport kinetics of both the trans-embrane and the soluble protein by eliminating a 10-min lag
hase observed for both GAG-free proteins. This supported distinctorting mechanisms, requiring a GAG-recognition machinery at theGN (Kobialka et al., 2009).
These results were confirmed in a more physiologically rele-ant context where the transport of the amyloid precursor proteinAPP), a part-time proteoglycan, was studied in HeLa cells (Mihovt al., 2015). APP is naturally tyrosine-sulfated and undergoes alter-ative splicing, leading to formation of CS-proteoglycan variants.gain, CS attachment resulted in fast, lag-free TGN exit. Deletion of
he entire cytosolic domain of APP slowed down surface transportf the GAG-free form, but did not affect the transport kinetics ofhe proteoglycan forms. Fast CS-mediated transport is thus inde-endent of and dominant over cytosolic sorting signals of APP.onsistent with this notion, xyloside-attached CS chains displayedhe same fast transport kinetics. Interestingly, both GAG-free androteoglycan isoforms of APP were routed via transferrin-positivendosomes to the plasma membrane (Mihov et al., 2015).
Another part-time proteoglycan, MHCII invariant chain, washown to have fast TGN-to-PM transport kinetics, acquiring ∼40%ensitivity to external chondroitinase after 15 min sulfate labelingArneson and Miller, 2007). Since the proteoglycans form trimersith GAG-free invariant chains, one CS chain appeared to be dom-
nant over cytoplasmic di-leucine signals for endosomal targeting.GAG chains allow proteoglycans to simultaneously connect
ultiple ligands and binding partners, thus creating supramolec-lar complexes at the cell surface (Mikami and Kitagawa, 2013;arrazin et al., 2011). Rapid segregation into distinct post-TGN vesi-les and fast surface transport might serve to reduce the risk of
hains. (1) Polarized TGN-to-cell surface transport; (2) non-polarized biosyntheticinternalization.
premature intracellular assembly and formation of dysfunctionalprotein aggregates. Currently, the mechanisms of GAG-mediatedTGN export are unknown. The extended, semi-rigid conformationand large size of GAGs might hinder proteoglycan entry into smalltransport vesicles and divert them into distinct carriers for largecargo, as has been observed for TGN exit of large collagen assem-blies (Polishchuk et al., 2003). Alternatively, proteoglycans might beactively sorted at the TGN via interaction with yet unknown cargoreceptors, linking the lumenal GAGs with the cytosolic machin-ery. To distinguish these possibilities, it would be interesting totest whether GAG chains other than CS similarly affect TGN-to-cellsurface transport.
4.3. Regulated secretion
Serglycin is a proteoglycan found in hematopoietic and endothe-lial cells. It has important functions related to secretory granulehomeostasis and constitutive secretion of various cargoes (Kolsetand Pejler, 2011). By interaction with its GAG chains (mainly CS),granule cargo such as chemokines, cytokines, proteases, as well ashistamine are concentrated before regulated secretion. In pancre-atic acinar cells, serglycin is present in zymogen granules and isinvolved in packing secretory cargo (Biederbick et al., 2003). Twoapproaches were used to generate GAG-free proteins and studythe role of GAGs for serglycin sorting: deletion of the GAG attach-ment domain and inhibition of GAG attachment by addition ofexogenous xyloside as a competing substrate for GAG synthesis.In both cases, secretory granule sorting was disrupted and ser-glycin was detected predominantly in perinuclear Golgi structures.Additionally, granule sorting of secretory enzymes like amylase and
procarboxypeptidase A was impaired in cells expressing GAG-freeserglycin (Biederbick et al., 2003). The data suggest that the sor-ting of serglycin into secretory granules at the TGN and its functiondepends on the attached GAGs.
9 al of B
4
tgpbdcmcoN2
Eah1smtr
alc2eodHnc
tthaicTfcSwi(
sptpeh(
5
bpn(i
0 D. Mihov, M. Spiess / The International Journ
.4. Sorting at the plasma membrane
In some cases, GAGs are important for proteoglycan recruitmento specific membrane domains at the cell surface. Neural/glial anti-en 2 (NG2 or CSPG4) CS proteoglycan is a membrane-spanningrotein involved in the formation of retraction fibers and expressedy tumors and immature progenitor cells including oligoden-rocyte progenitors, chondroblasts, and pericytes/smooth muscleells (Stallcup and Dahlin-Huppe, 2001). The amount of GAG-freeutant NG2 at cellular retraction fibers was significantly lower
ompared to wild-type NG2. Similarly, chondroitinase treatmentf cells expressing wild-type NG2 resulted in similar reduction ofG2 localization to retraction fibers (Stallcup and Dahlin-Huppe,001).
In addition, GAGs are determinants for protein endocytosis.arly studies showed that GAGs are involved in receptor bindingnd cellular uptake of secreted HS proteoglycans in arterial anduman skin fibroblasts (Kresse et al., 1975; Kruger and Kresse,986). The same phenomenon was later demonstrated for themall DS proteoglycan decorin: its uptake in the mouse skeletaluscle cell line C2C12 is mediated by simultaneous and cumula-
ive interaction of the protein core and the GAGs with lipoproteineceptor-related protein (Brandan et al., 2006).
Transmembrane HS proteoglycans have also been observed toct themselves as uptake receptors for a variety of ligands includingipoproteins, FGF2, anti-HS antibodies, cell-penetrating peptides,ationic polymers and polyplexes (Payne et al., 2007; Ram et al.,008; Tkachenko et al., 2004; Williams and Fuki, 1997; Wittrupt al., 2010). The internalization of proteoglycan–ligand complexesccurs through relatively slow clathrin-independent, but lipid raft-ependent pathways as well as macropinocytosis. Clustering of theS proteoglycans is required for internalization, but the mecha-ism and the precise role of HS chains in the process remain to belarified.
Interestingly, CS chains were found to negatively influencehe rapid clathrin-mediated endocytosis of transmembrane pro-eins. The ASGPR-H1 is a typical recycling receptor, mediatingepatic uptake of galactose-terminal glycoproteins. Insertion of
GAG attachment site and resulting CS addition caused strongnhibition of receptor internalization, which was reversed uponhondroitinase digestion of the cell surface (Kobialka et al., 2009).he inhibitory effect of CS on endocytosis was further demonstratedor APP, which in its GAG-free forms is also actively internalized vialathrin-coated vesicles at a rate of ∼8%/min (Mihov et al., 2015;chneider et al., 2008). Endocytosis of the CS-containing isoformas three-fold reduced. GAG chains thus act dominantly over exist-
ng cytosolic internalization signals in transmembrane proteinsMihov et al., 2015).
How CS chains inhibit endocytosis is not known. They mightterically hinder entry of the proteoglycan into clathrin-coatedits. Alternatively, the CS chain might interact with components ofhe extracellular matrix, thus restricting its lateral diffusion in thelasma membrane and its capture at endocytic sites. Experimentsmploying fluorescence recovery after photobleaching (FRAP),owever, did not detect significant immobilization of CS-ASGPR-H1Kobialka et al., 2009).
. Significance and future development
Proteoglycans participate in a plethora of cellular functions,oth at the cell surface and intracellularly, and are thus indis-
ensable for many essential processes including morphogenesis,euronal development, tumor formation, and pathogen uptakeMikami and Kitagawa, 2013; Sarrazin et al., 2011). Correct local-zation is therefore a prerequisite for their proper cellular functions.iochemistry & Cell Biology 68 (2015) 87–91
Good evidence has been accumulated that GAGs act as sortingdeterminants at different transport steps. However, the mecha-nisms underlying GAG-mediated sorting are still largely unknown.By analogy to established sorting mechanisms, specific cargo recep-tors might specifically recognize GAGs and distinguish betweendifferent types. They could be either transmembrane proteins link-ing the lumenally exposed GAG chains with cytosolic transportmachinery, or lumenal linker proteins sequestering the proteogly-cans into specific lipid domains. Alternatively, GAGs might havea passive effect on protein sorting via their physical and stericalproperties. Unraveling the role of GAGs in protein trafficking is animportant key for a better understanding of proteoglycan biologyin health and disease.
Acknowledgments
Our work was supported by the Swiss National Science Founda-tion (grant 31003A-125423).
References
Almond, A., Sheehan, J.K., 2000. Glycosaminoglycan conformation: do aqueousmolecular dynamics simulations agree with X-ray fiber diffraction?Glycobiology 10, 329–338.
Arneson, L.S., Miller, J., 2007. The chondroitin sulfate form of invariant chaintrimerizes with conventional invariant chain and these complexes are rapidlytransported from the trans-Golgi network to the cell surface. Biochem. J. 406,97–103.
Barthel, A., Nickel, W., Tonko, C., Söling, H.D., 1995. Sorting and budding ofconstitutive secretory vesicles in hepatocytes and hepatoma cells. Adv.Enzyme Regul. 35, 283–292.
Biederbick, A., Licht, A., Kleene, R., 2003. Serglycin proteoglycan is sorted intozymogen granules of rat pancreatic acinar cells. Eur. J. Cell Biol. 82, 19–29.
Bonifacino, J.S., Traub, L.M., 2003. Signals for sorting of transmembrane proteins toendosomes and lysosomes. Annu. Rev. Biochem. 72, 395–447.
Brandan, E., Retamal, C., Cabello-Verrugio, C., Marzolo, M.P., 2006. The low densitylipoprotein receptor-related protein functions as an endocytic receptor fordecorin. J. Biol. Chem. 281, 31562–31571.
Ghosh, P., Dahms, N.M., Kornfeld, S., 2003. Mannose 6-phosphate receptors: newtwists in the tale. Nat. Rev. Mol. Cell Biol. 4, 202–212.
Hafte, T.T., Fagereng, G.L., Prydz, K., Grondahl, F., Tveit, H., 2011. Proteincore-dependent glycosaminoglycan modification andglycosaminoglycan-dependent polarized sorting in epithelial Madin–Darbycanine kidney cells. Glycobiology 21, 457–466.
Kitagawa, Y., Sano, Y., Ueda, M., Higashio, K., Narita, H., Okano, M., et al., 1994.N-glycosylation of erythropoietin is critical for apical secretion byMadin–Darby canine kidney cells. Exp. Cell Res. 213, 449–457.
Kobialka, S., Beuret, N., Ben-Tekaya, H., Spiess, M., 2009. Glycosaminoglycan chainsaffect exocytic and endocytic protein traffic. Traffic 10, 1845–1855.
Kolset, S.O., Pejler, G., 2011. Serglycin: a structural and functional chameleon withwide impact on immune cells. J. Immunol. 187, 4927–4933.
Kolset, S.O., Vuong, T.T., Prydz, K., 1999. Apical secretion of chondroitin sulphate inpolarized Madin–Darby canine kidney (MDCK) cells. J. Cell Sci. 112, 1797–1801.
Kresse, H., Tekolf, W., von Figura, K., Buddecke, E., 1975. Metabolism of sulfatedglycosaminoglycans in cultivated bovine arterial cells. II. Quantitative studieson the uptake of 35SO4-labeled proteoglycans. Hoppe Seylers Z. Physiol. Chem.356, 943–952.
Kruger, U., Kresse, H., 1986. Endocytosis of proteoheparan sulfate by cultured skinfibroblasts. Biol. Chem. Hoppe Seyler 367, 465–471.
Lo, A.C., Thinakaran, G., Slunt, H.H., Sisodia, S.S., 1995. Metabolism of the amyloidprecursor-like protein 2 in MDCK cells polarized trafficking occursindependent of the chondroitin sulfate glycosaminoglycan chain. J. Biol. Chem.270, 12641–12645.
Mertens, G., Van der Schueren, B., van den Berghe, H., David, G., 1996. Heparansulfate expression in polarized epithelial cells: the apical sorting of glypican(GPI-anchored proteoglycan) is inversely related to its heparan sulfate content.J. Biol. Chem. 132, 487–497.
Mihov, D., Raja, E., Spiess, M., 2015. Chondroitin sulfate acceleratestrans-Golgi-to-surface transport of proteoglycan amyloid precursor protein.Traffic.
Mikami, T., Kitagawa, H., 2013. Biosynthesis and function of chondroitin sulfate.Biochim. Biophys. Acta 1830, 4719–4733.
Nickel, W., Huber, L.A., Kahn, R.A., Kipper, N., Barthel, A., Fasshauer, D., et al., 1994.ADP ribosylation factor and a 14-kD polypeptide are associated with heparan
sulfate-carrying post-trans-Golgi network secretory vesicles in rathepatocytes. J. Cell Biol. 125, 721–732.Payne, C.K., Jones, S.A., Chen, C., Zhuang, X., 2007. Internalization and trafficking ofcell surface proteoglycans and proteoglycan-binding ligands. Traffic 8,389–401.

al of B
P
P
R
R
S
S
S
D. Mihov, M. Spiess / The International Journ
olishchuk, E.V., Di Pentima, A., Luini, A., Polishchuk, R.S., 2003. Mechanism ofconstitutive export from the golgi: bulk flow via the formation, protrusion, anden bloc cleavage of large trans-golgi network tubular domains. Mol. Biol. Cell14, 4470–4485.
otter, B.A., Hughey, R.P., Weisz, O.A., 2006. Role of N- and O-glycans in polarizedbiosynthetic sorting. Am. J. Physiol.: Cell Physiol. 290, C1–C10.
am, N., Aroui, S., Jaumain, E., Bichraoui, H., Mabrouk, K., Ronjat, M., et al., 2008.Direct peptide interaction with surface glycosaminoglycans contributes to thecell penetration of maurocalcine. J. Biol. Chem. 283, 24274–24284.
odriguez-Carvajal, M.A., Imberty, A., Perez, S., 2003. Conformational behavior ofchondroitin and chondroitin sulfate in relation to their physical properties asinferred by molecular modeling. Biopolymers 69, 15–28.
arrazin, S., Lamanna, W.C., Esko, J.D., 2011. Heparan sulfate proteoglycans. ColdSpring Harb. Perspect. Biol., 3.
chneider, A., Rajendran, L., Honsho, M., Gralle, M., Donnert, G., Wouters, F., et al.,
2008. Flotillin-dependent clustering of the amyloid precursor protein regulatesits endocytosis and amyloidogenic processing in neurons. J. Neurosci. 28,2874–2882.eaman, M.N., 2008. Endosome protein sorting: motifs and machinery. Cell Mol.Life Sci. 65, 2842–2858.
iochemistry & Cell Biology 68 (2015) 87–91 91
Stallcup, W.B., Dahlin-Huppe, K., 2001. Chondroitin sulfate and cytoplasmicdomain-dependent membrane targeting of the NG2 proteoglycan promotesretraction fiber formation and cell polarization. J. Cell Sci. 114, 2315–2325.
Sugimoto, H., Sugahara, M., Folsch, H., Koide, Y., Nakatsu, F., Tanaka, N., et al., 2002.Differential recognition of tyrosine-based basolateral signals by AP-1B subunitmu1B in polarized epithelial cells. Mol. Biol. Cell 13, 2374–2382.
Tkachenko, E., Lutgens, E., Stan, R.V., Simons, M., 2004. Fibroblast growth factor 2endocytosis in endothelial cells proceed via syndecan-4-dependent activationof Rac1 and a Cdc42-dependent macropinocytic pathway. J. Cell Sci. 117,3189–3199.
Vagin, O., Kraut, J.A., Sachs, G., 2009. Role of N-glycosylation in trafficking of apicalmembrane proteins in epithelia. Am. J. Physiol.: Renal Physiol. 296, F459–F469.
Victor, X.V., Nguyen, T.K., Ethirajan, M., Tran, V.M., Nguyen, K.V., Kuberan, B., 2009.Investigating the elusive mechanism of glycosaminoglycan biosynthesis. J.Biol. Chem. 284, 25842–25853.
Williams, K.J., Fuki, I.V., 1997. Cell-surface heparan sulfate proteoglycans: dynamic
molecules mediating ligand catabolism. Curr. Opin. Lipidol. 8, 253–262.Wittrup, A., Zhang, S.H., Svensson, K.J., Kucharzewska, P., Johansson, M.C.,Morgelin, M., et al., 2010. Magnetic nanoparticle-based isolation of endocyticvesicles reveals a role of the heat shock protein GRP75 in macromoleculardelivery. Proc. Natl. Acad. Sci. U. S. A. 107, 13342–13347.