hemopoietic responses in mice injected with purified ... from 12 mice per group ± standard...
TRANSCRIPT

Exp. Hematol. 15:1-9 (1987) © 1987 International Society for Experimental Hematology
Experimental Hematology
Hemopoietic Responses in Mice Injected with Purified Recombinant Murine GM-CSF Donald Metcalf, 1 C. Glenn Begley, 1 D. James Williamson, 1 Edouard C. Nice,2 John De Lamarter,3 JeanJacque Mermod, 3 David Thatcher, 3 and Albert Schmidt3
1Cancer Research Unit, Walter and Eliza Hall Institute ofMedical Research, and 2Ludwig Institute for Cancer Research, Royal Melbourne Hospital, Melbourne, Victoria, Australia; and 3Biogen SA, Geneva, Switzerland
(Received 13 May 1986; in revised form 10 July 1986; accepted 17 July 1986)
Abstract. Normal adult BALB/c, C57BL, and C3H/HeJ mice were injected intraperitoneally three times daily for six days with 6-200 ng purified, bacterially synthesized, murine recombinant GM-CSF. Mice injected with 200 ng rGM-CSF developed a twofold increase in blood neutrophils. In the peritoneal cavity, a dose-related rise was observed in macrophages (up to 15-fold), neutrophils (10- to 100-fold) and eosinophils (10- to 100-fold). Peritoneal macrophages exhibited 15-fold increased mitotic activity (to 7.6/103 cells) and increased phagocytic activity for antibody-coated erythrocytes. Increased numbers of infiltrating neutrophils and monocytes were observed in the liver and lung. Doserelated rises were observed in spleen weight (up to 50%) and the spleen content ofmonocytes (twofold) and nonerythroid progenitor cells (up to fourfold). A dose-related fall occurred in total marrow cellularity (40%) and total nonerythroid progenitor cells (37%-66%), but levels ofneutrophils and monocytes remained constant. The data indicate that the injection ofrGM-CSF to normal mice increases overall numbers of granulocytes and macrophages and the phagocytic activity of macrophages and provides direct evidence for the conclusion that GMCSF is likely to function in vivo as a regulator of these cell populations.
Key words: Colony-stimulating factor - In vivo effects Granulocyte-macrophage production - Phagocytosis
The colony-stimulating factors (CSFs) are a group of specific glycoproteins that, in cultures of hemopoietic cells, are able to control the proliferation and differentiation of granulocytes, macrophages, and some related hemopoietic cell populations. Four murine CSFs have been identified-GM-CSF G-CSF, M-CSF (CSF-1), and Multi-CSF (IL-3)_: -----Address reprint requests to: Dr. D. Metcalf, Cancer Research Unit,
. Walt"r and Eliza Hall Institute of Medical Research, PO 3050, Royal i\1elbourne Hospital, Victoria, Australia.
and each has been purified to homogeneity and its range of actions in vitro characterized (see review by Metcalf [1]).
Studies have succeeded in isolating cDNAs for murine Multi-CSF [2, 3] and GM-CSF [4] and, using each eDNA, biologically active recombinant CSF has been produced in both mammalian and bacterial expression systems [5-10]. Purified, bacterially synthesized GM-CSF has been shown at low concentrations (2-80 pg!ml) to be an effective stimulus for granulocytic and macrophage colony formation, at concentrations above 80 pg!ml to stimulate eosinophil colony formation, and at concentrations above 640 pg!ml to stimulate megakaryocyte and some pure and mixed erythroid colony formation [ 1 0]. These properties were similar to those of native GM-CSF at equivalent concentrations.
The present studies were undertaken to determine whether the injection of purified, bacterially synthesized GM-CSF into normal adult mice was capable of stimulating detectable hemopoietic changes corresponding to the known actions of this regulator in vitro.
Materials and methods
Mice. Mice used were three-month-old males of the strains C57BL/ 6fJ/WEHI, C3H/HeJ, and BALB/c/An/Bradley/WEHI maintained in this institute and were matched for body weight before use. Mice had been reared under specific pathogen-free conditions then conventionalized for five weeks before use.
Preparation and purification of bacterially synthesized murine GM-CSF. A murine GM-CSF clone was isolated from EL-4 cells using oligonucleotide probes based on the previously published eDNA sequence for murine GM-CSF [4]. The production and purification of recombinant murine GM-CSF synthesized by Escherichia coli have been described previously [9, 10]. The purity to homogeneity of the recombinant GM-CSF was verified by the following analytical procedures: refractionation using a TSK HPLC column, peptide mapping, isoelectric focusing, amino acid sequence analysis, and quantitative amino acid analysis . In no instance was evidence obtained of contaminating polypeptides.

2
Based on quantitative amino acid measurements, the recombinant GM-CSF (rGM-CSF) was diluted in 10% syngeneic mouse serum/saline to produce a dilution containing 200 ng/0.2 ml. During this dilution step, the material was processed through a PD 10 column to remove residual acetonitrile. Serial dilutions of this material were then made to contain 100, 50, 25, 12, and 6
ng/0.2 ml. All batches of rGM -CSF prediluted in mouse serum/ saline and
the carrier serum/saline itself were assayed for endotoxin using the Limulus amebocyte lysate assay both before and after courses of in vivo injections and no endotoxin was detected (lower detection limit, 0.04 ng!ml). In previous control studies, mice injected with 0.2 ng endotoxin three times daily for six days failed to develop any of the changes to be described, and a redescription of these negative results [ 11] will not be repeated in the present data although, in many of the experiments, control mice were injected with 0.2 ng endotoxin, again with negative effects.
Injection schedule. Mice were injected intraperitoneally (i.p.) with 0.2 ml of 10% mouse serum/saline containing 0-200 ng rGMCSF three times daily at 8:00A.M., 4:00 P.M. and 10:00 P.M. All analyses were commenced at 9:00 A.M. on the morning following the last (evening) injection.
White blood cell counts. Mice were anesthetized with ether and orbital plexus blood collected using 50-f.Ll micropipettes. No mouse was examined on more than one occasion to avoid misleading cell counts from previously damaged and/or inflamed vessels. White cell counts were performed using hemocytometers and differential cell counts were performed on blood films stained with May-Griinwald-Giemsa.
1000
e g;_ ~
l:l 100 :i:: '.:
2 =>
0·5 Hours after injection
Fig. 1. Serum GM-CSF levels in BALB/c mice following a single intraperitoneal injection of 65 ng rGM-CSF. Serum GM-CSF levels in uninjected BALB/c mice were undetectable in the assay used ( < 5 U/ml). Mean values ± standard deviations.
Experimental Hematology vol. 15 (1987)
Examination of tissues. After performance of white cell counts, mice were killed with ether. Following death, each mouse was injected i.p. with 2 ml saline and, after gentle massage of the abdominal cavity, the peritoneal wall was opened and as much fluid as possible collected using a sterile pipette.
Bone marrow cells were collected from one complete femur shaft and the spleen was removed aseptically. Portions of the following tissues were taken for histological examination after fixation in 10% formalin in saline-spleen, kidney, liver, mesenteric node, small bowel adjacent to the mesenteric node, lung, heart, skin, and thymus. All sections were prepared in duplicate and stained with hematoxylin-eosin or, for mast cell enumeration, with Alcian blue-safranin. Cytocentrifuge preparations were made of peritoneal, bone marrow, and spleen cell suspensions and stained with May-Griinwald-Giemsa.
Bioassay of rGM-CSF levels. Levels of rGM-CSF in all preparations used for injection were assayed prior to and following each course of injections using serial twofold dilutions of 0.1 ml in 1 ml agar-medium cultures containing 75,000 C57BL bone marrow cells [12]. Units (U) ofCSF per milliliter were calculated from the linear portion of the dose-response curve, 50 U being the concentration stimulating the formation of half-maximal numbers of colonies. With the batches of purified recombinant GM-CSF used, quantitative amino acid analysis indicated that 200 ng ofrGM-CSF assayed as 120,000 bone marrow units (specific activity 6 x 108 U/mg).
Bioassays on GM-CSF levels in the serum of injected mice were performed using microwell cultures of 15 1'1 Dulbecco's modified Eagle's medium containing 5 1'1 of serial serum dilutions and 200 FDC-Pl cells [13]. Cell counts were performed on the wells after 48 h of incubation and observed levels of GM-CSF converted to bone marrow units per milliliter by use of standard control preparations of GM-CSF of known activity.
Assay of progenitor cell levels in injected mice. Dispersed cell suspensions were prepared from femoral marrow and a portion of the spleen. Assays were performed in 1-ml agar cultures in 35-mm petri dishes using 50,000 spleen cells or 25,000 marrow cells per culture. The medium used was Dulbecco's modified Eagle's medium with a final concentration of 20% fetal calf serum (FCS) and 0.3% agar. Colony formation was stimulated by addition of 0.1 ml of a semi purified preparation of pokeweed-mitogen-stimulated spleen-conditioned medium containing 400 U GM-CSF and 400 U Multi-CSF [14]. Cultures were incubated for seven days in a fully humidified atmosphere of 10% C02 in air and colony formation (clones >50 cells) scored at x 35 using a dissection microscope. The whole cultures were fixed with 2.5% glutaraldehyde and then differential colony counts performed af-
Table 1. Peripheral blood white cell levels in BALB/c mice injected with rGM-CSF
Total cells/f.Ll
Injected with Total cells/ 1'1 Neutrophils Lymphocytes Eosinophils Monocytes
9600 ± 3990 1540 ± 590 7500 ± 3260 60 ± 80 500 ± 300
Serum/saline 0.2 ml 8770 ± 2100 1810 ± 770 6450 ± 1470 90 ± 80 420 ± 210
rGM-CSF 200 ng 10,160 ± 2050 2840 ± 800 6740 ± 1730 110 ± 100 400 ± 170
rGM-CSF 100 ng 8860 ± 2170 2690 ± 140 5470 ± 1500 60 ± 60 480 ± 270
BALB/c mice were injected three times daily for six days and then examined on day 7. Data from 12 mice per group ± standard deviations. Neutrophil levels in mice injected with 200 ng rGM-CSF were significantly elevated above levels in either control group (t = 4.5 and 3.4, respectively; p < 0.01). Levels in mice injected with 100 ng were not significantly different from those in mice injected
with serum/saline.
D. Metcalf et al.: Effects in vivo of GM-CSF
ter staining for acetylcholinesterase and counter-staining with Luxol fast blue-hematoxylin.
Quantitative estimation of tissue content of megakaryocytes and mast cells. Megakaryocytes and mast cells were counted in whole spleen sections and then the organ content of these cells was calculated from camera Iucida drawings as described previously [II].
Phagocytosis assays. Pooled peritoneal cells from each treatment group were washed twice in HEPES-buffered RPMI-1640 containing 2% FCS (RPMI-1640) and suspended at 106 cells/ml. Antibody-coated sheep erythrocytes (EA) were prepared by incubating the cells with a pretitrated agglutinating amount of mouse IgG2b anti-E (Sera-Lab MAS 013, Crawley Down, UK) for 40 min at room temperature. The EA were then washed three times to remove unbound antibody and suspended at 5 x I 07 cells/ml in RPMI-FCS. Equal volumes (150 !'\) of peritoneal cells and EA targets were added to Wasserman tubes and the cells were kept at 4°C until the incubation (37°C, 10 min) was started after the cells had been brought into contact by gentle centrifugation (500 g, 20 s). The assay was stopped by adding an excess of ice-cold medium; extracellular erythrocytes were subjected to hypotonic lysis and stained smears of the cells were prepared and scored as previously described [11, 15].
Results
Serum levels of rGM-CSF in injected mice
BALB/c mice were given a single i.p. injection of 65 ng rGM-CSF and then the serum was collected at intervals up to 6 h and assayed in microwell cultures ofFDC-P1 cells to determine GM-CSF levels. Serum GM-CSF levels in uninjected BALB/c mice were undetectable ( < 5 U/ml). As shown in Figure 1, levels of GM-CSF in the serum rose to a peak value of 500 U/ml 30 min following injection, then fell logarithmically with a half-life of approximately 35 min to 10 U/ml by 3 h after injection. Based on these observations, the range of doses of rGM -CSF chosen for injection was from 6 ng to 200 ng.
Peripheral blood changes
White cell counts were performed on BALB/c mice after six days of injection of 200 or 100 ng rGMCSF. As shown in Table 1, the only significant change observed was a slightly less than twofold elevation in neutrophil levels. No immature granulocytes or nucleated red cells were observed in the blood.
Peritoneal cell changes
Groups of BALB/c mice were injected with 6-200 ng. rGM-CSF three times daily for six days and the m1~e examined on day 7. The data on the peritoneal. cell populations in mice from three such experiments have been pooled in Figure 2. Clear
3
Neutrophlls
30
'§ 0
;2 20
10
GM- CSF per injection ng
Fig. 2. Peritoneal cell population changes in BALB/c mice injected intraperitoneally for six days with varying doses ofrGMCSF: U, uninjected mice; and S, mice injected with 0.2 ml mouse serum/saline. Each point is mean value from 12 mice± standard deviations. Neutrophil levels in mice injected with 50, 100, and 200 ng rGM-CSF were significantly higher than in serum/salineinjected mice (t = 3.2, 3.9, 2.8, respectively; p < 0.01). Macrophage levels in mice injected with 6, 12, 25, 50, 100, and 200 ng rGM-CSF were significantly higher than in serum/saline-injected mice (t = 5.1, 9.0, 5.5, 6.5, 7.9, and 8.0, respectively; p < 0.01). Eosinophil levels in mice injected with 6, 12, 25, 50, 100, and 200 ng rGM-CSF were significantly higher than in serum/ saline-injected mice (t = 5.4, 4.7, 3.8, 5.6, 3.9, and 4.5, respectively; p < 0.01).
dose-related rises were observed in peritoneal macrophages, eosinophils, and neutrophils, with no significant changes in lymphocyte levels. Rises in macrophage numbers were significant even with the lowest dose used (6 ng/injection) and, with the highest dose, macrophage levels reached 50 x 106 cells in some mice. The data suggested that plateau responses had not been achieved even with the highest dose used. The neutrophil and eosinophil rises reached levels 100-fold higher than in uninjected control mice and 10- to 20-fold higher than in mice injected with serum/saline and involved exclusively postmitotic mature cells in both lineages (Fig. 3). The peritoneal macrophages from mice injected with rG M -CSF were larger and more basophilic than resident macrophages and many exhibited cytoplasmic vacuolation. Significant mitotic activity was observed in peritoneal macrophages from mice injected with rGM-CSF, reaching mean levels of 7.6/ 103 cells, although no distinct relationship was observed between the dose of rGM-CSF injected and the frequency of mitotic figures (Table 2).

4
Fig. 3. Changes induced in BALB/c mice following the intraperitoneal injection for six days of 200 ng GM-CSF. (A) Peritoneal cells from mouse injected with rGM-CSF showing increased macrophages (one in mitosis), neutrophils, and eosinophils (arrows). (B) Peritoneal cells from control mouse injected with serum/saline. (C) Liver from mouse injected with rGM-CSF showing increased infiltration of granulocytes and monocytes. (D) Liver from control mouse. (E) Lung from mouse injected with rGM-CSF showing increased cellularity of alveolar sac walls, including neutrophils. (F) Lung from control mouse. Cytocentrifuge preparations stained with May-Griinwald-Giemsa, sections with hematoxylin-eosin.
Observations on groups ofC57BL and C3H/HeJ mice injected with the same dose range ofrGM-CSF gave similar results to those described for BALB/c mice. In both strains, significant rises in peritoneal macrophages and eosinophils were observed with
Experimental Hematology vol. 15 (1987)
Table 2. Elevated mitotic activity in peritoneal macrophages from BALB/c mice injected with rGM-CSF
Injected with
0.2 ml serum/saline
rGM-CSF
6 ng 12 ng 25 ng 50 ng
100 ng 200 ng
Mitoses/ 103 macro phages
0±0 0.2 ± 0.7
6.5 ± 5.4 5.0 ± 3.9 4.0 ± 4.0 4.2 ± 4.5 7.6 ± 7.5 5.1 ± 5.8
Twelve mice per group were injected three times daily for six days and examined on day 7. Mean frequency of mitotic figures per 103 macro phages ± standard deviations (300-600 cells scored in each preparation). Frequency of mitoses in cells from mice injected with 6, 12, 25, 50, and 100 ng rGM-CSF were significantly higher than in cells from control mice injected with serum/ saline (t = 3.8, 4.0, 3.0, 2.9, and 3.3, respectively; p < 0.01).
the lowest dose used (6 ng) and with 200 ng the mean levels of cells were as follows. C57BL: macrophages 43.2 ± 10.2 x 106 , neutrophils 0.4 ± 0.3 x 106 , and eosinophils 4.8 ± 1.7 x 106 (control C57BL mice injected with serum/saline: macrophages 3.1 ± 1.0 x 106 , neutrophils 0.07 ± 0.08 x 106 , and eosinophils 0.01 ± 0.03 x 106 cells). C3H/ HeJ: macrophages 16.4 ± 2.4 x 106 , neutrophils 0.3 ± 0.3 x 106 , and eosinophils 1.3 ± 0.3 x 106
cells (control C3H/HeJ mice injected with serum/ saline: macrophages 2.0 ± 1.1 x 106 , neutrophils 0.05 ± 0.06 x 106 , and eosinophils 0.01 ± 0.02 x 106 cells). In neither strain were significant changes observed in levels of peritoneal lymphocytes.
Sequential studies on C57BL mice injected with 200 ng rGM-CSF showed that, by 3 h after a single injection, no significant changes had occurred in peritoneal cell levels, indicating that no rapid immigration of cells to the peritoneal cavity was induced by rGM-CSF. In mice injected three times daily, minor elevations of peritoneal macrophages were observed at 24 h and levels rose progressively thereafter (Fig. 4). A similar pattern was observed for neutrophils and eosinophils.
Phagocytic activity of peritoneal macro phages
Assays were performed on the ability of peritoneal macrophages to phagocytose antibody-coated sheep erythrocytes using cells harvested from BALB/c, C57BL, and C3H/HeJ mice injected for six days with varying doses ofrGM-CSF. Typical results from one such experiment with cells of each strain are shown in Figure 5. The absolute levels of phagocytic activity in each of the three experiments are not
D. Metcalf et al.: Effects in vivo of GM-CSF
Peritoneal Macrophages 36
32
28
rGM-CSF
·""
3 4 Days of Injection
Fi~. 4: . Progressive rise in total peritoneal macrophages in C57BL m1ce Injected three times daily with 200 ng rGM-CSF or 0.2 ml mouse serum/saline. Each point represents an individual mouse.
directly comparable because of the use of different batches of antibody. Clear evidence was obtained with cells from each strain that rGM-CSF induced dose-related rises both in the percentage of macrophages with phagocytosed erythrocytes (up to eightfold) and in the average number of erythrocytes per p~ag~cytically active cell (up to sevenfold). Combmatwn of these data with the absolute rises in total macrophages induced by rGM-CSF indicated that the overall level of phagocytic capacity m the total peritoneal cavity population had been increased in excess of 100-fold.
Liver changes
In BALB/c mice injected for six days with varying doses ofrGM-CSF, a dose-related rise was observed m th~ number ofnonparenchymal cells in the liver. The mfiltrating cells were mainly neutrophils and macrophages with some eosinophils and occasional megakaryocytes (Fig. 3). The infiltrating cells were usually dispersed between the parenchymal cords but were also present as focal aggregates not nee~ essari.ly adjacent to portal vessels. No mitoses were seen m these cells. Counts on the number of nonparenchymal cells were performed avoiding focal a.ggre~ates but, even with this deliberate underestlm~tl?n, a rise of at least 50% was observed in mice receiVmg the higher doses ofrGM-CSF (Fig. 6).
Lung changes
In BALB/c mice injected with the higher doses of rG?vf-CSF, it was common for the alveolar sac walls to b · fi em ltrated by increased numbers of cells the most readily identifiable of which were neutro~hils.
5
rGM·CSF per 1nJect1on ng
Fig. 5. Phagocytic activity for antibody-coated erythrocytes of pooled _veri.t~neal macro phages from BALBI c, C5 7BL, and C3H/ HeJ mice Injected three times daily for six days with varying doses of rGM-CSF: U, uninjected; and S, injected with 0.2 ml mouse serum:s~l.ine. Note progressive rise in percentage ofmacroph~ges exhibitmg phagocytic activity (e----e) and the rGMCS~-mduce~ rise in the average number of erythrocytes in phagocytically active macrophages (0--0). Each point is mean value from duplicate preparations. The absolute values for each mouse str~in are not directly comparable because of the use of differing antibody preparations.
18
16
14 ~
'" i 12
H ~
~ 10 d E
f 8
~
~ 6
§ z
u s 6 12 25 50 100200
rGM-CSF per injection ng
Fig. 6. Dose-related rise in nonparenchymal cells in the liver of B~LB/c mice injected three times daily for six days with :arymg d~ses of rGM-CSF: U, uninjected mice; and S, mice InJected With 0.2 ml mouse serum/saline. Each point represents mean cell counts per six high-power fields from each of 12 mice + standard deviations. Numbers in mice injected with 100 and 200 ~g rGM~CSF were significantly higher than in control mice inJected With serum/saline (t = 4.7 and 5.0, respectively;p < 0.01).
It proved impossible to quantify this increased cell~lar infiltration because of the irregular compres~lOn of the alveolar sacs in the sections, but the I~creased numbers of infiltrating neutrophils per high-power field approximated two- to fivefold.
Spleen changes
Injection of BALB/c mice for six days with rGMCS~ resulted in a moderate dose-related rise in spleen weight approaching 50% (Table 4). Because of the
Experimental Hematology vol. 15 (1987)
6
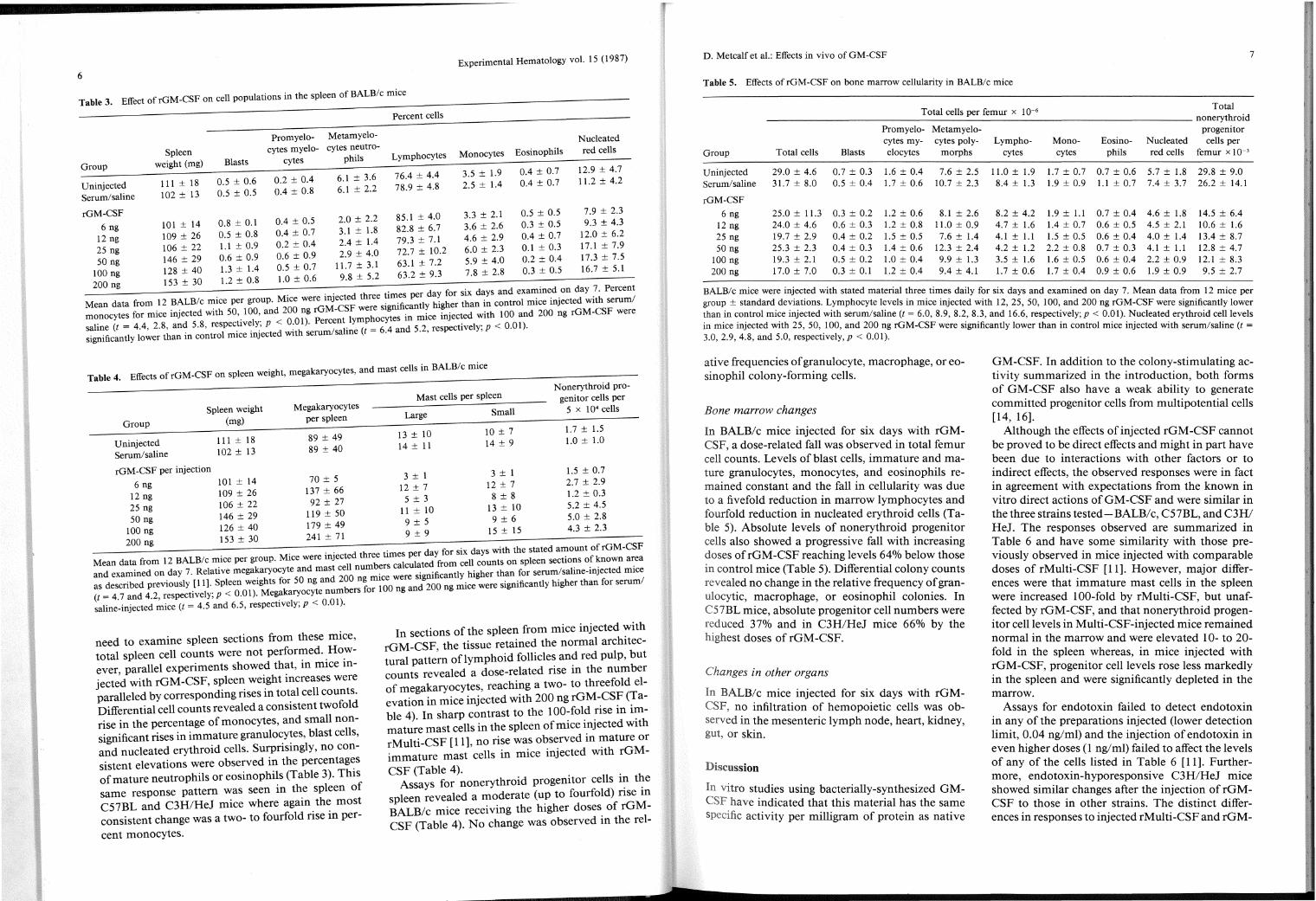
Table 3. Effect of rGM-CSF on cell populations in the spleen of BALB/c mice
Promyelo- Metamyelo-
Spleen cytes myelo- cytes neutro-
weight (mg) Blasts cytes phils Group
6.1 ± 3.6 Uninjected 111 ± 18 0.5 ± 0.6 0.2 ± 0.4
0.5 ± 0.5 0.4 ± 0.8 6.1 ± 2.2 Serum/saline 102 ± 13
rGM-CSF 101 ± 14 0.8 ± 0.1 0.4 ± 0.5 2.0 ± 2.2
6 ng 0.4 ± 0.7 3.1 ± 1.8 12 ng 109 ± 26 0.5 ± 0.8
0.2 ± 0.4 2.4 ± 1.4 25 ng 106 ± 22 1.1 ± 0.9 9 0 6 + 0 9 2.9 ± 4.0
5o ng 146 ± 29 0.6 ± 0. · - · 128 +40 1.3±1.4 0.5±0.7 1!.7±3.1
100 ng - 8 52
Percent cells
Lymphocytes
76.4 ± 4.4 78.9 ± 4.8
85.1 ± 4.0 82.8 ± 6.7 79.3 ± 7.1 72.7 ± 10.2 63.1 ± 7.2 63.2 ± 9.3
Monocytes
3.5 ± 1.9 2.5 ± 1.4
3.3 ± 2.1 3.6 ± 2.6 4.6 ± 2.9 6.0 ± 2.3 5.9 ± 4.0 7.8 ± 2.8
Eosinophils
0.4 ± 0.7 0.4 ± 0.7
0.5 ± 0.5 0.3 ± 0.5 0.4 ± 0.7 0.1 ± 0.3 0.2 ± 0.4 0.3 ± 0.5
Nucleated red cells
12.9 ± 4.7 11.2 ± 4.2
7.9 ± 2.3 9.3 ± 4.3
12.0 ± 6.2 17.1 ± 7.9 17.3 ± 7.5 16.7 ± 5.1
200 ng 153 ± 30 1.2 ± 0.8 1.0 ± 0.6 9. ± · . . . . ed three times per day for six days and examin~d _on day ? . Percent
Mean data from 12 BALB/c mlce per group. Mlce were mject . 'fi ntly higher than in control mice mJected wlth serum/ monocytes for mice injected with 50, 100, and 200 ng rGM-CSlF we: Slytg~~ ~:mice injected with 100 and 200 ng rGM-CSF were
8 d 58 spectively- P < 0 01) Percent ymp oc 01) saline (t = 4.4, 2. , an · • re . . .' : · 1 r e (t = 6 4 and 5.2, respectively; P < 0. · significantly lower than in control m1ce mjected Wlth serum sa m ·
. d mast cells in BALB/c mice Table 4. Effects ofrGM-CSF on spleen we1ght, megakaryocytes, an
Group
Uninjected Serum/saline
Spleen weight (mg)
111 ± 18 102 ± 13
rGM-CSF per injection
Megakaryocytes per spleen
89 ± 49 89 ± 40
101 ± 14 70 ± 5 6 ng 137 ± 66
12 109 ± 26 -ng 92 + 27
25 106 ± 22 -ng 119 + 50
0 146 ± 29 -5 ng 179 ± 49
100 ng 126 ± 40 + 71
Mast cells per spleen
Large
13 ± 10 14 ± 11
3 ± 1 12 ± 7
5 ± 3 11 ± 10
9 ± 5 9±9
Small
10 ± 7 14 ± 9
3 ± 1 12 ± 7
8 ± 8 13 ± 10 9±6
15 ± 15
Nonerythroid progenitor cells per
5 x 104 cells
!.7±1.5 1.0 ± 1.0
1.5 ± 0.7 2.7 ± 2.9 1.2 ± 0.3 5.2 ± 4.5 5.0 ± 2.8 4.3 ± 2.3
200 ng 153 ± 30 241 -. . . d three times per day for six days with the stated amount of rGM-CSF
Mean data from 12 BALB/c mice per group. Mlce were mJecte b 1 lated from cell counts on spleen sections of known area and examined on day 7. Relative megakaryocyte and mast cell num ~rs ca cu . nificantly higher than for serum/saline-injected mice
l S 1 . hts for 50 ng and 200 ng mlce were Slg . h h f, m/ as described previously [11 . Peen welg f, 100 and 200 ng mice were significantly hlg er t an or seru (t = 4.7 and 4.2, respectively; p < 0.01). Meg~aryocyte numbers or ng saline-injected mice (t = 4.5 and 6.5, respectively; P < 0.01).
need to examine spleen sections from these mice, total spleen cell counts were not perfor~ed. _Ho_wever parallel experiments showed that, m m1ce mject~d with rGM-CSF, spleen weight increases were paralleled by corresponding rises in total cell counts. Differential cell counts revealed a consistent twofold rise in the percentage of monocytes, and small nonsignificant rises in immature granulo~y~es, blast cells, and nucleated erythroid cells. Surpnsmgly, no consistent elevations were observed in the percentag~s of mature neutrophils or eosinophils (Table 3). Th1s same response pattern was seen in th.e spleen of C57BL and C3H/HeJ mice where agam the most consistent change was a two- to fourfold rise in per-
cent monocytes.
In sections of the spleen from mice injected :-'ith rGM-CSF, the tissue retained the normal architectural pattern oflymphoid follicles and red pulp, but counts revealed a dose-related rise in the number of megakaryocytes, reaching a two- to threefold elevation in mice injected with 200 ng rGM~CS~ <!able 4). In sharp contrast to the IOO~fol~ :1se m 1~mature mast cells in the spleen ofm1ce l_nJected w1th rMulti-CSF [11], no rise was observed m t_nature or immature mast cells in mice injected w1th rGM-
CSF (Table 4). . . Assays for nonerythroid progemtor cells 1~ t~e
spleen revealed a moderate (u~ to fourfold) nse m BALB/c mice receiving the h1gher doses_ of rGMCSF (Table 4). No change was observed m the rei-
D. Metcalf et al.: Effects in vivo of GM-CSF 7
Table 5. Effects ofrGM-CSF on bone marrow cellularity in BALB/c mice
Total cells per femur x IQ-6 Total nonerythroid
Promyelo- Metamyelo- progenitor cytes my- cytes poly- Lympho- Mono- Eo sino- Nucleated cells per
Group Total cells Blasts elocytes morphs cytes cytes phils red cells femur x IQ-3
Uninjected 29.0 ± 4.6 0.7 ± 0.3 1.6 ± 0.4 7.6 ± 2.5 11.0 ± 1.9 1.7 ± 0.7 0.7 ± 0.6 5.7 ± 1.8 29.8 ± 9.0 Serum/saline 31.7 ± 8.0 0.5 ± 0.4 1.7 ± 0.6 10.7 ± 2.3 8.4 ± 1.3 1.9 ± 0.9 l.l ± 0.7 7.4 ± 3.7 26.2 ± 14.1
rGM-CSF
6 ng 25.0 ± 11.3 0.3 ± 0.2 1.2 ± 0.6 8.1 ± 2.6 8.2 ± 4.2 1.9 ± l.l 0.7 ± 0.4 4.6 ± 1.8 14.5 ± 6.4 12 ng 24.0 ± 4.6 0.6 ± 0.3 1.2 ± 0.8 11.0 ± 0.9 4.7 ± 1.6 1.4 ± 0.7 0.6 ± 0.5 4.5 ± 2.1 10.6 ± 1.6 25 ng 19.7 ± 2.9 0.4 ± 0.2 1.5 ± 0.5 7.6 ± 1.4 4.1 ± 1.1 1.5 ± 0.5 0.6 ± 0.4 4.0 ± 1.4 13.4 ± 8.7 50 ng 25.3 ± 2.3 0.4 ± 0.3 1.4 ± 0.6 12.3 ± 2.4 4.2 ± 1.2 2.2 ± 0.8 0.7 ± 0.3 4.1 ± l.l 12.8 ± 4.7
100 ng 19.3 ± 2.1 0.5 ± 0.2 1.0 ± 0.4 9.9 ± 1.3 3.5 ± 1.6 1.6 ± 0.5 0.6 ± 0.4 2.2 ± 0.9 12.1 ± 8.3 200 ng 17.0±7.0 0.3 ± 0.1 1.2 ± 0.4 9.4 ± 4.1 1.7 ± 0.6 1.7 ± 0.4 0.9 ± 0.6 1.9 ± 0.9 9.5 ± 2.7
BALB/c mice were injected with stated material three times daily for six days and examined on day 7. Mean data from 12 mice per group± standard deviations. Lymphocyte levels in mice injected with 12, 25, 50, 100, and 200 ng rGM-CSF were significantly lower than in control mice injected with serum/saline (t = 6.0, 8.9, 8.2, 8.3, and 16.6, respectively; p < 0.01). Nucleated erythroid cell levels in mice injected with 25, 50, 100, and 200 ng rGM-CSF were significantly lower than in control mice injected with serum/saline (t =
3.0, 2.9, 4.8, and 5.0, respectively, p < 0.01).
ative frequencies of granulocyte, macrophage, or eosinophil colony-forming cells.
Bone marrow changes
In BALB/c mice injected for six days with rGMCSF, a dose-related fall was observed in total femur cell counts. Levels of blast cells, immature and mature granulocytes, monocytes, and eosinophils remained constant and the fall in cellularity was due to a fivefold reduction in marrow lymphocytes and fourfold reduction in nucleated erythroid cells (Table 5). Absolute levels of nonerythroid progenitor cells also showed a progressive fall with increasing doses ofrGM-CSF reaching levels 64% below those in control mice (Table 5). Differential colony counts revealed no change in the relative frequency of granulocytic, macrophage, or eosinophil colonies. In C57BL mice, absolute progenitor cell numbers were reduced 37% and in C3H/HeJ mice 66% by the highest doses ofrGM-CSF.
Changes in other organs
In BALB/c mice injected for six days with rGMCSF, no infiltration of hemopoietic cells was observed in the mesenteric lymph node, heart, kidney, gut, or skin.
Dl:scussion
In vitro studies using bacterially-synthesized GMCSF have indicated that this material has the same specific activity per milligram of protein as native
GM-CSF. In addition to the colony-stimulating activity summarized in the introduction, both forms of GM-CSF also have a weak ability to generate committed progenitor cells from multipotential cells [14, 16].
Although the effects of injected rGM-CSF cannot be proved to be direct effects and might in part have been due to interactions with other factors or to indirect effects, the observed responses were in fact in agreement with expectations from the known in vitro direct actions ofGM-CSF and were similar in the three strains tested-BALBI c, C5 7BL, and C3H/ HeJ. The responses observed are summarized in Table 6 and have some similarity with those previously observed in mice injected with comparable doses of rMulti-CSF [11]. However, major differences were that immature mast cells in the spleen were increased 100-fold by rMulti-CSF, but unaffected by rGM-CSF, and that nonerythroid progenitor cell levels in Multi-CSF-injected mice remained normal in the marrow and were elevated 10- to 20-fold in the spleen whereas, in mice injected with rGM-CSF, progenitor cell levels rose less markedly in the spleen and were significantly depleted in the marrow.
Assays for endotoxin failed to detect endotoxin in any of the preparations injected (lower detection limit, 0.04 ng/ml) and the injection of endotoxin in even higher doses (1 ng/ml) failed to affect the levels of any of the cells listed in Table 6 [11]. Furthermore, endotoxin-hyporesponsive C3H/HeJ mice showed similar changes after the injection of rGMCSF to those in other strains. The distinct differences in responses to injected rMulti-CSF and rGM-

8
Table 6. Summary of changes induced by the injection of rGM
CSF for 6 days•
Parameter
Peripheral blood
Neutrophils
Peritoneal cavity
Macro phages Neutrophils Eosinophils Macrophage mitotic
activity Macrophage phagocytic
activity
Liver Hemopoietic cells
Lung Neutrophils
Spleen
Weight Monocytes Megakaryocytes Nonerythroid progenitor
cells
Bone marrow
Total cells Nucleated erythroid cells Lymphocytes Nonerythroid progenitor
cells
Rises
2-fold
15-fold l 0- to l 00-fold l 0- to l 00-fold
40-fold
100-fold
50%
2- to 5-fold
50% 2- to 4-fold 2- to 4-fold
2- to 5-fold
Falls
40% 3-fold 5-fold
37%-66%
• Relative to levels in control mice injected with serum/saline
alone.
CSF (both produced by the same strain of E. coli) in themselves constitute strong evidence that the responses observed were not ascribable to undetected contaminants of bacterial origin.
A single i.p. injection of65 ng rGM-CSF achieved peak serum GM-CSF levels of 500 U/ml 30 min after injection which fell by 3 h to levels of 10 U /ml, a concentration in vitro that would only have minimal effects on hemopoiesis. Even with the highest doses used (200 ng), the interrupted injection schedule used would only have achieved a serum concentration of GM-CSF considered from in vitro studies to be significant (greater than 50 U/ml) for periods of up to 3 h, following which a period of 5-7 h would then elapse with no significant elevation of CSF levels. The transient nature of these elevations probably reduced the magnitude of the changes observed.
Quantitatively, the most striking response to in-jected rGM-CSF involved cells of the peritoneal cavity, the site ofCSF injection. The elevated numbers of neutrophils and eosinophils exclusively in-
Experimental Hematology vol. 15 (1987)
valved cells that were postmitotic and these must have slowly accumulated in the cavity from the circulation. Human GM-CSF has been shown to inhibit neutrophil migration [17] and GM-CSF may possess previously unsuspected direct or indirect chemotactic effects. In the case of the peritoneal macrophages, clear mitotic activity was observed (mitotic index up to 0.7%). Depending on the duration of the mitotic event, this frequency of mitoses could represent local proliferative activity sufficient to account for at least half of the rise in macrophages (if mitosis lasted 30 min), but would account for a smaller proportion of the observed rise if mitosis
lasted longer. The peritoneal macrophages in mice injected with
rGM -CSF exhibited clearly increased phagocytic activity for antibody-coated sheep erythrocytes. This observation was similar to that made previously in mice injected with rMulti-CSF [11] and supports in vitro evidence that the CSFs are able to stimulate the functional activation of mature granulocytes and macrophages [1].
There was a moderate increase in the relative and absolute numbers of granulocyte-macrophage progenitors in the spleen, but this was more than balanced by a fall in the absolute number of progenitor cells in the marrow of approximately 60%. Thus the increased numbers of granulocytes and macrophages in mice injected with rGM-CSF appear to have been achieved at the expense of some depletion of the total population of granulocyte-macrophage progenitor cells. It will require studies oflonger duration to determine whether this depletion process would continue if injections ofrGM-CSF were continued for longer periods or whether stem cell activation would be triggered with some restoration of progenitor cell levels. To clarify this situation, observations are also needed on stem and multipotential cell levels in mice injected with rGM-CSF.
The spleen is the characteristic organ in adult mice exhibiting increased hemopoiesis in response to stress or stimulation. One curious aspect of the response pattern to injected rGM-CSF was the small change in granulocytic or monocytic cells in the spleen in contrast to the marked rises in these populations in the peritoneal cavity, liver, and lungs. Even in the marrow, absolute numbers of granulocytes remained constant. Thus the stimulation of granulocytic and monocytic populations by rGMCSF, at least at the time points sampled, did not increase these populations in the organs generating these cells (the marrow and spleen) but, instead, resulted in clear increases of these cells in peripheral tissues such as the peritoneal cavity, liver, and lung.
D. Metcalf et al.: Effects in vivo of GM-CSF
In the context of the possible use of rGM-CSF in an animal suffering from a refractory infection, this pattern of the response is quite advantageous in increasing the numbers of protective granulocytes and monocytes in locations that might be sites of a widespread infection.
With the lower doses of rGM-CSF used, the expected serum CSF levels would at maximum still be in the linear portion of the in vitro dose-response curve. The ability of these relatively small doses of rGM-CSF, even when separated by relatively long intervals, to induce detectable changes suggests that the responsiveness to rGM-CSF of at least some in vivo parameters may not be dissimilar from that exhibited by hemopoietic populations in vitro. The range of serum concentrations achieved by the injection of rGM-CSF are within those actually observed in the serum during responses to natural viral and bacterial infections [12].
These present responses induced in normal mice injected rGM-CSF strongly support the conclu
sion reached from extensive earlier indirect observations [12] that GM-CSF is likely to function in vivo as a genuine regulator of granulocyte-macrophage populations and possibly some related hemopoietic populations. In the clinical context the administration ofrGM-CSF can be expected t~ result in useful stimulation of hemopoietic populations in patients with subnormal hemopoiesis, e.g., following cytotoxic therapy, and in patients with refractory infections.
Acknowledgments
The authors are indebted to the careful technical assistance of Mrs. C. Quilici and Miss Y. Wiluszynski. This work was supported by the Carden Fellowship Fund of the Anti-Cancer Council of Victoria, The National Health and Medical Research Council, Canberra, and the National Institutes of Health, Bethesda, grants CA-22556 and 25972.
References
l. MetcalfD (1986) The molecular biology and functions of the granulocyte-macrophage colony-stimulating factors. Blood 67:257
2. F ung MC, Hapel AJ, Ymer S, Cohen DR, Johnson RM, CampbellHD, YoungiG (1984) MolecularcloningofcDNA for mouse interleukin-3. Nature 307:233
9
3. Yokota T, Lee F, Rennick D, Hall C, Arai N, Mosmann T, Nabel IG, Cantor H, Arai K (1984) Isolation and characterization of a full-length eDNA for mast cell growth factor from a mouse T -cell clone: expression in monkey cells. Proc Natl Acad Sci USA 81:1070
4. Gough NM, Gough J, MetcalfD, Kelso A, Grail D, Nicola NA, Burgess A W, Dunn AR (1984) Molecular cloning of eDNA encoding a murine haemopoietic growth regulator, granulocyte-macrophage colony stimulating factor. Nature 309:763
5. Hapel AJ, Fung MC, Johnson RM, Young IG, Johnson G, Metcalf D (1985) Biologic properties of molecularly cloned and expressed murine interleukin 3. Blood 65:453
6. Dunn AR, MetcalfD, Stanley E, Grail D, King J, Nice EC, Burgess A W, Gough NM (1985) Biological characterization of regulators encoded by cloned hematopoietic growth factor gene sequences. In: Seramisco J, Ozanne B, Stiles C (eds) Cancer cells 3: growth factors and transformation. Cold Spring Harbor NY: Cold Spring Harbor Laboratory, p 227
7. Greenberger JS, Humphries RK, Messner H, Reed DA, Sakakeeny AA (1985) Molecularly cloned and expressed T -cell gene product is biologically similar to interleukin-3. Exp Hematol 13:249
8. Kindler V, Thorens B, De Kossodo S, Allet B, Eliason JF, Thatcher D, Vassalli P (1986) Stimulation ofhematopoiesis in vivo by recombinant bacterial murine interleukin 3. Proc Nat! Acad Sci USA 83:1001
9. De Lamarter JF, Mermod J -J, Liang CM, Eliason JF, Thatcher D (1985) Recombinant murine GM-CSF from E. coli has biological activity and is neutralized by a specific antiserum. EMBO J 4:2275
10. MetcalfD, Burgess AW, Johnson GR, Nicola NA, Nice EC De Lamarter J, Thatcher DR, Mermod J-J (1986) In vitro actions on hemopoietic cells of recombinant murine GMCSF purified after production in E. coli: comparison with purified native GM-CSF. J Cell Physiol 128:421
11. Metcalf D, Begley CG, Johnson GR, Nicola NA, Lopez AF, Will_iamson ~J (1986) Effects of purified bacterially-synthesized munne Multi-CSF (IL-3) on hemopoiesis in normal adult mice. Blood 68:46
12. MetcalfD ( 1984) The hemopoietic colony stimulating factors. Amsterdam: Elsevier
13. Dexter TM, Garland J, Scott D, Scolnick E, MetcalfD (1980) Growth of factor-dependent hemopoietic precursor cell lines. J Exp Med 152:1036
14. <=:utle~ RL, MetcalfD, Nicola NA, Johnson GR (1983) Punficatwn of a multipotential colony stimulating factor from pokeweed mitogen-stimulated spleen conditioned medium. J Bioi Chern 260:6579
15. Lopez AF, Strath M, Sanderson CJ (1981) IgG and complement receptors on purified mouse eosinophils and neutrophils. Immunology 43:779
16. Iscove NN, Keller G, Roitsch C (1985) Factors required by pluripotential hemopoietic stem cells in culture. Prog Clin Bioi Res 184: 105
17. Gasson JC, Weisbart RK, Kaufman SE, Clark SC, Hewick RM, Wong GG, Golde DW (1984) Purified human granulocyte-macrophage colony-stimulating factor: direct action on neutrophils. Science 226: 13 3 9