identification acyl donor steryl ester biosynthesis ... · acyl donorin steryl ester biosynthesis...
TRANSCRIPT

Plant Physiol. (1978) 62, 348-353
Identification of an Acyl Donor in Steryl Ester Biosynthesis byEnzyme Preparations from Spinach Leaves
Received for publication November 17, 1977 and in revised form January 31, 1978
RAYMOND E. GARCIA"2 AND J. BRIAN MUDDDepartment of Biochemistry and Statewide Air Pollution Research Center, University of California, Riverside,California 92521
ABSTRACT MATERIALS AND METHODS
A pathway for steryl ester biosynthesis in acetone powder preparationsfrom spinach (Spincis olkraces L.) leaves has been elucidated; free steroland 1,2-dilyceride were the substrates. Although animals synthesize cho-lesteryl esters by three distinct biosynthetic pathways, none of thesepathways utilizes 1,2-diglyceride as an acyl donor. Phosphatidylcholine,phosphatidic acid, triglyceride, 1,3-diglyceride, 1-monoglyceride, free fattyacid, and fatty acyl-CoA were not acyl donors for spinach leaf steryl esterbiosynthesis in our assay system. The unstable 2 isomer of monoglyceridewas not tested. It is possible that 1,2-diglyceride and 2-monoglyceride wereboth acyl donors for spinach leaf steryl ester biosynthesis. Acyl-labeledphosphatidylyholine and acyl-labeled phosphatidylethanolamine were rap.idly degraded by acetone powder preparations to 1,2diglyceride via phos-phatidic acid. The 1,2-diglycerides were slowly metabolized to monoglyc-erides, triglycerides, free fatty acids, and steryl esters. The monoglycerideswere rapidly degraded to free fatty acids and glycerol.
The acyl donors and the enzymes involved in steryl esterbiosynthesis in higher plants have not been identified. In animalscholesterol esterase (10), fatty acyl-CoA-cholesterol acyltransfer-ase (9), and phosphatidylcholine-cholesterol acyltransferase (8)catalyze the esterification of a fatty acid from free fatty acid, fattyacyl-CoA, and phosphatidylcholine, respectively, to free choles-terol, forming cholesteryl ester. Bartlett et al. (1) have recentlyshown the biosynthetic pathways for fungal steryl ester biosyn-thesis to be the same as those established for animal cholesterylester biosynthesis. Kolattukudy (13, 14) has shown that the waxester biosynthetic pathway in broccoli leaves are mechanisticallyanalogous to the three pathways catalyzed by animals in choles-teryl ester biosynthesis. In both broccoli leafwax ester biosynthesisand animal cholesteryl ester biosynthesis the acyl donors are freefatty acid, fatty acyl-CoA, and phospholipid. It is not unreasonableto assume, therefore, that higher plants may utilize one or more ofthese acyl donors in the biosynthesis of steryl esters. Our investi-gation indicates, however, that spinach leaves employed a uniquesteryl ester biosynthetic pathway in which 1,2-diglyceride was theacyl donor. Although phospholipids were promising in our initialstudies, an extensive product analysis indicated that they wererapidly metabolized by phospholipase D and phosphatidic acidphosphatase to 1,2-diglycerides. These two enzymes are frequentlyfound in plants (Il, 17, 18).
Supported by a predoctoral National Science Foundation Traineeship.2 Present address: Biomedical Sciences Division, Lawrence Livermore
Laboratory, University of California, Livermore, California 94550.
Materials. Sodium [l,2-'4C]acetate (55 mCi/mmol) was pur-chased from International Chemical and Nuclear Corp., Irvine,Calif. [4-'4C]Cholesterol (56 mCi/mmol), [16-'4Clpalmitic acid(18.7 mCi/mmol), and [1-_4CJpalmitoyl-CoA (57.8 mCi/mmol)were obtained from New England Nuclear. [1-_4C]Palmitate-la-beled dipalmitin (116 mCi/mmol) and [I-_4CJpalmitate-labeledmonopalmitin (58 mCi/mmol) were obtained from Dhom Prod-ucts, Ltd., North Hollywood, Calif. Tripalmitin, 1,3-dipalmitin,1,2-diolein, 1,2-dipalmitin, 1-monopalmitin, cholesterol, palmit-oyl-CoA, cholesteryl palmitate, and BSA3 were obtained fromSigma Chemical Company. All other chemical were reagent gradeor better. Chloroform, methanol, acetone, toluene, and benzenewere glass-distilled. Fresh spinach (Spinacia oleracea L.) waspurchased from the local markets.
Acetone Powder Preparation. In most cases, the acetone powderof the 20,000g pellet fraction from spinach leaves was prepared asdescribed previously (6). The acetone powder used in the [1-14C]palmitate-labeled dipalmitin and [1-'4C]palmitate-labeled mono-palmitin assays was prepared in a slightly different manner. Inthis case, the homogenizing buffer contained 10 mm MES-NaOH(pH 6); the 20,000g subcellular suspension was added to 120 ml ofacetone at 0 C; and the precipitated protein was washed with 120ml of acetone at 0 C. All other procedures were as describedpreviously (6).
Acetone-Ether Powder Preparation. The acetone-ether powderof the 20,000g pellet fraction from spinach leaves was prepared asdescribed previously (7), except the dried acetone powder wasresuspended in water instead of 100 mm MES-NaOH (pH 6).
116-4ClPahnitic Acid Reaction Mixtures. The reaction mixtureswere the same as those described for the [14C]acyl-labeled phos-phatidylcholine reaction mixtures, except the assays contained28,800 dpm [16-'4Cjpalmitic acid (18.7 mCi/mmol).
[1-"4ClPahnitoyl-CoA Reaction Mixtures. The unlabeled cho-lesterol was added to the acetone powder in acetone at 0 C. Theacetone powder was dried under vacuum at 0 C before theenzyme-cholesterol mixture was homogenized to uniformity incold 100 mm MES-NaOH (pH 6) with a Potter-Elvehjem homog-enizer. The reaction mixtures contained 44,000 cpm [1-_4CJpal-mitoyl-CoA (57.8 mCi/mmol). All other reaction mixture com-ponents and experimental procedures were as described for the['4Clacyl-labeled phosphatidylcholine reaction mixtures.
14CIAcyl-labeled Phosphatidylchollne Reaction Mixtures. Thelabeled phosphatidylcholine and unlabeled cholesterol were added
Abbreviations: ASG: acylated steryl glucoside; BSA: essentially fattyacid-free bovine albumin; DG: diglyceride (diacylglycerol); EP: ethylpalmitate; FFA: free fatty acid; MG: monoglyceride (monoacylglycerol);0: origin; PA: phosphatidic acid; PC: phosphatidylcholine; PE: phospha-tidylethanolamine; SE: steryl ester; TG: triglyceride (triacylglycerol).
348 www.plantphysiol.orgon March 21, 2020 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.

ACYL DONOR IN STERYL ESTER BIOSYNTHESIS
to the acetone powder in diethyl ether at 0 C. The acetone powderwas dried under vacuum at 0 C before the enzyme-substratemixture was homogenized to uniformity in cold 100 mm MES-NaOH (pH 6) with a Potter-Elvehjem homogenizer. The reactionmixtures contained 34,000 dpm ['4Clacyl-labeled phosphatidyl-choline (0.66 mCi/mmol), 0.2 mm cholesterol, 100 mm MES-NaOH (pH 6), 10 mg of acetone powder of the 20,000g pelletfraction (3 mg of protein), and water to make a final volume of 1ml. The reaction was initiated by the addition of the enzyme-substrate mixture and the samples were incubated in a Dubnoffmetabolic shaking incubator at 30 C for 0, 2.5, 5, 10, 20, 40, 80, or120 min. The reactions were terminated by adding 3 ml of CHC13-CH30H (1:2, v/v). Whenever enzymic catalysis was evident in the0-min incubation, this background was subtracted from the me-tabolites.
114CIAcyl-labeled 1,2-Diglyceride Reaction Mixtures. The re-action mixtures were the same as those described for the [14Clacyl-labeled phosphatidylcholine reaction mixtures, except the assayscontained 20,000 dpm ['4C]acyl-labeled 1,2-diglyceride (<0.66mCi/mmol).
11-_4ClPalmitate-labeled Dipahmitin and 1l-"4CiPalmitate-la-beled Monopalmitin Reaction Mixtures. The reaction mixtureswere the same as those described for the [14Clacyl-labeled phos-phatidylcholine reaction mixtures, except the assays containedeither 100,000 cpm [1-_4C]palmitate-labeled dipalmitin (116mCi/mmol) or 100,000 cpm [1-_4C]palmitate-labeled monopal-mitin (58 mCi/mmol).
Extraction, Isolation, and Counting of Acyl-labeled Lipids(Method A). The lipids were extracted according to the method ofBligh and Dyer (2) and separated into individual components byTLC. The lipid samples were applied in small aliquots of CHC13to Silica Gel G TLC plates that had been activated at 110 C forI hr. Neutral lipids were resolved by developing the plates inhexane-diethyl ether-acetic acid (70:30:1, v/v). Polar lipids wereresolved by developing the plates in chloroform-methanol-aceticacid-water (85:15:3:2, v/v). The radioactive areas were detectedby autoradiography and identified by their co-chromatographywith standards (cholesteryl palmitate, tripalmitin, palmitic acid,1,2-dipalmitin, 1-monopalmitin, methyl stearate, phosphatidyl-choline, phosphatidylethanolamine, and phosphatidic acid). Thestandards were detected by spraying with 1% (w/v) iodine inmethanol. The radioactive areas were scraped into scintillationvials, 10 ml of liquid scintillation fluid (10 g of PPO + 0.6 g ofPOPOP + 2 liters of toluene) were added to each vial, and thesamples were counted for 10 min in a Nuclear-Chicago 720 seriesliquid scintillation system. Quenching was corrected by the chan-nel ratio method.
Extraction, Isolation, and Counting of Acyl-labeled Lipids(Method B). This method was used only for the [1-_4C]palmitate-labeled dipalmitin and [1-_4Cjpalmitate-labeled monopalmitin as-says. The lipids were extracted by a modified Bligh and Dyermethod (2) in which I ml of CHC13 and 0.8 ml of 2 N KCI in 0.5M K-phosphate at pH 7.3 were added to the 4 ml of monophasicCHC13-CH30H-H20 (1:2:1, v/v) solution. This gave a distinctbiphasic CHC13-CH30H-H20 (2:2:1.8, v/v) solution. The CHC13phase was removed, dried under vacuum, and redissolved in asmall aliquot of CHC13. The lipids were separated by one-dimen-sional Silica Gel G TLC using a two-step development system asdescribed by Skipski and Barclay (20). The first development wasdone in isopropyl ether-acetic acid (96:4, v/v), and the seconddevelopment was done in hexane-diethyl ether-acetic acid(90:10:1, v/v). All other procedures were as described in methodA, except the samples were counted for 10 min in a Beckman LS-230 liquid scintillation system.
14-1'CICholesterol Reaction Mixtures. The [4-"CJcholesteroland unlabeled cholesterol were added to the acetone powder inacetone at 0 C. The acetone powder was dried under vacuum at
0 C before the enzyme-substrate mixture was homogenized touniformity in cold 100 mM MES-NaOH (pH 6) with a Potter-Elvehjem homogenizer. The reaction mixtures contained 44,400dpm [4-'4C]cholesterol (56 mCi/mmol), and the incubation timewas either 20 or 60 min. All other reaction mixture componentsand experimental procedures were as described for the [' C]acyl-labeled phosphatidylcholine reaction mixtures. The 14-'4Clcholes-teryl esters were extracted, isolated, and counted as describedpreviously (6).AgNO3 TLC. The biosynthesized [4-' Clcholesteryl esters were
separated into different fatty acyl groups depending upon theirdegree of unsaturation by AgNO3 TLC as described previously(7).
Preparation of 14CIAcyl-labeled Phosphatidylcholine and 114CIAcyl-labeled Phosphatidylethanolamine. ['4CjAcyl-labeled phos-phatidylcholine and [14Clacyl-labeled phosphatidylethanolaminewere biosynthesized as outlined by Galliard (5), using 87 ,iCi ofsodium [1,2-'4C]acetate (55 mCi/mmol) as the starting materialand 27 g of potato discs from White Rose potato tubers as theenzyme source. The labeled phospholipids were partially purifiedby DEAE-cellulose column chromatography as described by Gal-liard (5). The glycolipid contaminants were removed by silicicacid chromatography as outlined by Vorbeck and Marinetti (23).The purified ["4Clacyl-labeled phosphatidylcholine (0.66mCi/mmol) and [14Clacyl-labeled phosphatidylethanolamine(0.27 mCi/mmol) gave single autoradiography spots when theSilica Gel G TLC plates were developed in chloroform-methanol-acetic acid-water (34:6:4:1, v/v) or chloroform-methanol-aceticacid-water (25:15:2:4, v/v).
Preparation of 14CIAcyl-labeled 1,2-Diglyceride. The ["4Clacyl-labeled 1,2-diglyceride was prepared by incubating 50 mg ofacetone powder preparation with 170,000 dpm ['4C]acyl-labeledphosphatidylcholine (0.66 mCi/mmol) for 10 min. The reactionmixtures were as described for the [14Clacyl-labeled phosphatidyl-choline assays. The [''Clacyl-labeled 1,2-diglyceride was purifiedby preparative Silica Gel G TLC using hexane-diethyl ether-aceticacid (70:30:1, v/v) as the solvent system. The labeled 1,2-diglyc-erides were scraped onto filter paper and eluted with CHC13 intoa drying flask. The sample was then dried under vacuum andredissolved in a small aliquot of CHC13-CH30H (9:1, v/v). SilicaGel G TLC in hexane-diethyl ether-acetic acid (70:30:1, v/v)indicated that the labeled 1,2-diglycerides were 89% pure.
Analytical Method. Protein was determined by the method ofLowry et al. (15).
RESULTS
116-14CIPalmitic Acid. [16-'4C]Palmitic acid was not incorpo-rated into steryl esters when incubated with a spinach leaf acetonepowder preparation (data not shown). Autoradiography analysisof the thin layer chromatograms of the lipid extract revealed thatpalmitic acid was the only labeled lipid in these assays. Controlexperiments indicated that this acetone powder preparation wasable to incorporate [4-'4CJcholesterol into [4-'4CJcholesteryl ester.
[1-14CIPalmitoyl-CoA. A small amount of label from [1-''4Cpalmitoyl-CoA was incorporated into cholesteryl [11-4CJpalmitate,as shown in Table I. It is not clear, however, if the label wasdirectly or indirectly incorporated into steryl ester. The dataindicate that free fatty acid was the most highly labeled product,followed by diglyceride. Phosphatidylcholine and phosphatidyl-ethanolamine were not significantly labeled in the 120-min incu-bation (Table I) nor in shorter incubation times (data not shown).The MG + ASG? + X assignment in Table I is a combination
of at least three lipids that were not successfully resolved on thethin layer chromatograms. Additional TLC indicated that MGwas a very minor component, accounting for only 3% of the labelin MG + ASG? + X. ASG accounted for 51% of the label, but its
349Plant Physiol. Vol. 62, 197 8
www.plantphysiol.orgon March 21, 2020 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.

GARCIA AND MUDDTable I. Product Analysis for the Metabolism of [1-l4C]Palmitoyl-CoA
Reaction mixtures were as described under Materials and Methods.Incubation time was 120 min. Polar lipids (PC, PE, X1, and MG + ASG? + X)and neutral lipids (X2, DG, FFA, TG, and SE) were extracted, isolated, andcounted as described for acyl-labeled lipids under method A of Materialsand Methods. Samples were counted in a Beckman LS-230 liquid scintillationsystem. X1: a polar lipid that chromatographed just above PE in the polarsolvent system; X2: a neutral lipid that chromatographed just below DG inthe neutral solvent system.
Lipid cpm
PC 0 0.0
PE 36 0.1
XI 184 0.6
MG + ASG? + X 3,350 11.0
X2 633 2.1
DG 4,750 15.7FFA 20,100 66.3
TG 591 1.9
SE 695 2.3
E
18
6
4
2
10
8
6
4
2
0
5 10 20 40 80TIME (minutes)
FIG. 1. ['4ClAcyl-labeled phosphatidylcholine time course. Reactimixtures were as described under "Materials and Methods." Reactilwere terminated by adding 0.15 ml of 80%o trichloroacetic acid, follomby 3 ml of CHC13-CH30H (1:2, v/v). Acyl-labeled lipids were extractisolated, and counted as described under method A of "Materials aMethods."
identification is tentative. The remaining 46% has not been idetified.
Steryl ester biosynthesis was relatively insensitive to 5 mgBSA when cholesterol was the labeled precursor but was almcompletely inhibited by it when palmitoyl-CoA was the labelprecursor (data not shown). Serum albumin tightly binds lochain fatty acids (21) and assumedly binds the CoA derivativesfatty acids as well (12, 22). The binding of palmitoyl-CoA by B'would make it unavailable to enzymic attack.
I14CIAcyl-labeled Pbospholipids. Figure I is a time course I
the degradation of ['4Clacyl-labeled phosphatidylcholine inspinach leaf acetone powder preparation. Labeled phosphatizacid was detected but only when the protein was precipitatedeither trichloroacetic acid, as in the case of Figure 1, or by boilimethanol (data not shown). Control experiments indicated titrichloroacetic acid was not hydrolyzing phosphatidylcholinephosphatidic acid nor to 1,2-diglyceride. Furthermore, the labelphosphatidylcholine was not degraded if the acetone powcpreparation had been boiled for 5 min or omitted from the reactimixture, indicating that enzymic reactions were involved.There was a rapid disappearance of label from phosphatid
choline and a rapid incorporation of label into 1,2-diglycenr(Fig. 1). Only a trace of 1,3-diglyceride was detected. Label a]appeared in steryl ester and free fatty acid but less rapidly thinto 1,2-diglyceride. After the label in phosphatidylcholine hbecome depleted, the label in 1,2-diglyceride slowly disappear(
Plant Physiol. Vol. 62, 1978
suggesting that 1,2-diglyceride may have been responsible for thecontinued incorporation of label into steryl ester and free fattyacid. A preliminary investigation with ["Clacyl-labeled phospha-tidylethanolamine indicated that a similar product distributionwas obtained with this phospholipid.
Small amounts oflabeled triglycerides and monoglycerides werealso detected in the labeled phosphatidylcholine reaction mixtures(data not shown). Labeled lysophosphatidylcholine was not de-tected in these assays.
I''CIAcyl-labeled 1,2-Diglyceride. Figure 2 compares the rate ofsteryl ester biosynthesis in a [14Clacyl-labeled phosphatidylcholineassay with an assay containing only the labeled 1,2-diglyceridethat was generated in a [14Clacyl-labeled phosphatidylcholineassay. The rate of steryl ester biosynthesis in Figure 2 was thesame whether phosphatidylcholine or 1,2-diglyceride was the la-beled precursor. All of the fatty acids that were esterified tocholesterol in the phosphatidylcholine assays, therefore, cameeither directly from 1,2-diglyceride or from a catabolite of 1,2-diglyceride, such as monoglyceride, and little, if any, came directlyfrom either phosphatidylcholine or phosphatidic acid.The disappearance of label from 1,2-diglyceride was accompa-
nied by the appearance of label in steryl ester and free fatty acid(Fig. 3). Steryl ester was the most heavily labeled product through-out the time course. Monoglycerides and triglycerides were alsodetected (data not shown) but were minor components.
11-_4CPalmitate-labeled Dipahnitin and 11-_4CIPahnitate-la-beled Monopalmitin. Table II shows a product analysis for themetabolism of [1-_4C]palmitate-labeled dipalmitin and [l-'_4Cpal-mitate-labeled monopalmitin. Free fatty acid, monoglyceride, tri-glyceride, and steryl ester were all major products in the metabo-lism of dipalmitin. Label, however, was more rapidly incorporatedinto free fatty acid and monoglyceride than into triglyceride andsteryl ester. Monopalmitin was rapidly degraded into free fattyacid and glycerol. In this case, free fatty acid and diglyceride werethe major products, whereas triglyceride and steryl ester were
tion minor products. Monopalmitin was more susceptible to degrada-Mns tion than was dipalmitin.wvedted,and
en-
, ofostledngofSA
fora
dicbyinghattoledderion
yl-ideLisoianlad-ed,
3
°) 2
E
(I)
0
I II
0
0
0 U
0
0
II I I0 510 20 30 40 50 60 70
TI ME (minutes)
FIG. 2. Incorporation of label from ['"Clacyl-labeled phosphatidylcho-line and ['4Ciacyl-labeled 1,2-diglyceride into steryl ester. Reaction mix-tures were as described under "Materials and Methods." After 10 min ofincubation, [14C]acyl-labeled phosphatidylcholine assays contained 14,000*dpm (U) and 19,000 dpm 1,2-diglyceride (0). In ['4Clacyl-labeled 1,2-diglyceride assays, 20,000 dpm 1,2-diglyceride (0) from a 10-min ['4Clacyl-labeled phosphatidylcholine assay was added to 10 mg of acetonepowder. Acetone powders used in all of these assays were different aliquotsfrom the same acetone powder preparation. In order to make a validcomparison between ['4C]acyl-labeled phosphatidylcholine and [14Ciacyl-labeled 1,2-diglyceride assays, only the rate of steryl ester biosynthesisafter the first 10 min of incubation in the ['4Clacyl-labeled phosphatidyl-choline assays was plotted. Labeled steryl esters were isolated by SilicaGel G TLC in hexane-diethyl ether-acetic acid (70:30:1, v/v), detected byautoradiography, and counted as described for acyl-labeled lipids undermethod A of "Materials and Methods." ['4C]Acyl-labeled phosphatidyl-choline assays (U, 0); ['4C]acyl-labeled 1,2-diglyceride assays (0).
350
www.plantphysiol.orgon March 21, 2020 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.
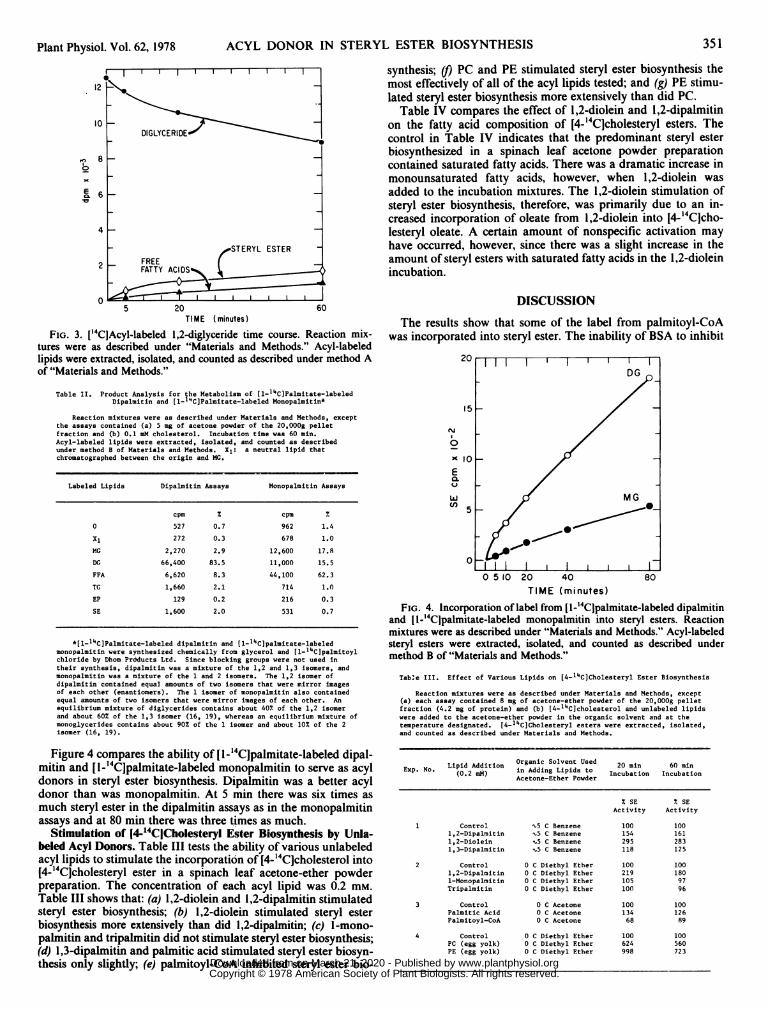
5 20 60TIME (minutes)
FIG. 3. I'4ClAcyl-labeled 1,2-diglyceride time course. Reaction mix-tures were as described under "Materials and Methods." Acyl-labeledlipids were extracted, isolated, and counted as described under method Aof "Materials and Methods."
Table II. Product Analysis for the Metabolism of [1-14C]Palmitate-labeledDipalmitin and [1-14C]Palmitate-labeled Monopalmitin*
Reaction mixtures were as described under Materials and Methods, exceptthe assays contained (a) 5 mg of acetone powder of the 20,OOOg pelletfraction and (b) 0.1 mM cholesterol. Incubation time was 60 min.Acyl-labeled lipids were extracted, isolated, and counted as describedunder method B of Materials and Methods. X1: a neutral lipid thatchromatographed between the origin and MG.
Labeled Lipids Dipalmitin Assays Monopalmitin Assays
cpm % cpm %0 527 0.7 962 1.4
X1 272 0.3 678 1.0
MG 2,270 2.9 12,600 17.8
DG 66,400 83.5 11,000 15.5
FFA 6,620 8.3 44,100 62.3
TG 1,660 2.1 714 1.0
EP 129 0.2 216 0.3
SE 1,600 2.0 531 0.7
*[1-14C]Palmitate-labeled dipalmitin and [1-14C]palmitate-labeledmonopalmitin were synthesized chemically from glycerol and [1-14C]palmitoylchloride by Dhom Prc6ducts Ltd. Since blocking groups were not used intheir synthesis, dipalmitin was a mixture of the 1,2 and 1,3 isomers, andmonopalmitin was a mixture of the 1 and 2 isomers. The 1,2 isomer ofdipalmitin contained equal amounts of two isomers that were mirror imagesof each other (enantiomers). The 1 isomer of monopalmitin also containedequal amounts of two isomers that were mirror images of each other. Anequilibrium mixture of diglycerides contains about 40% of the 1,2 isomerand about 60X of the 1,3 isomer (16, 19), whereas an equilibrium mixture ofmonoglycerides contains about 90% of the 1 isomer and about 10% of the 2isomer (16, 19).
Figure 4 compares the ability of [1-_4C]palmitate-labeled dipal-mitin and [1-_4C]palmitate-labeled monopalmitin to serve as acyldonors in steryl ester biosynthesis. Dipalmitin was a better acyldonor than was monopalmitin. At 5 min there was six times asmuch steryl ester in the dipalmitin assays as in the monopalmitinassays and at 80 min there was three times as much.
Stimulation of 14-4CICholesteryl Ester Biosynthesis by Unla-beled Acyl Donors. Table III tests the ability of various unlabeledacyl lipids to stimulate the incorporation of [4-'4CJcholesterol into[4-'4CJcholesteryl ester in a spinach leaf acetone-ether powderpreparation. The concentration of each acyl lipid was 0.2 mm.Table III shows that: (a) 1,2-diolein and 1,2-dipalmitin stimulatedsteryl ester biosynthesis; (b) 1,2-diolein stimulated steryl esterbiosynthesis more extensively than did 1,2-dipalmitin; (c) I-mono-palmitin and tripalmitin did not stimulate steryl ester biosynthesis;(d) 1,3-dipalmitin and palmitic acid stimulated steryl ester biosyn-thesis only slightly; (e) palmitoyl-CoA inhibited steryl ester bio-
351
synthesis; (f) PC and PE stimulated steryl ester biosynthesis themost effectively of all of the acyl lipids tested; and (g) PE stimu-lated steryl ester biosynthesis more extensively than did PC.
Table IV compares the effect of 1,2-diolein and 1,2-dipalmitinon the fatty acid composition of [4-'4CJcholesteryl esters. Thecontrol in Table IV indicates that the predominant steryl esterbiosynthesized in a spinach leaf acetone powder preparationcontained saturated fatty acids. There was a dramatic increase inmonounsaturated fatty acids, however, when 1,2-diolein wasadded to the incubation mixtures. The 1,2-diolein stimulation ofsteryl ester biosynthesis, therefore, was primarily due to an in-creased incorporation of oleate from 1,2-diolein into [4-'4CJcho-lesteryl oleate. A certain amount of nonspecific activation mayhave occurred, however, since there was a slight increase in theamount of steryl esters with saturated fatty acids in the 1,2-dioleinincubation.
DISCUSSION
The results show that some of the label from palmitoyl-CoAwas incorporated into steryl ester. The inability of BSA to inhibit
20
15
N
x 10
E
w
(I)
0 510 20 40 80TIME (minutes)
FIG. 4. Incorporation of label from Il-'4CJpalmitate-labeled dipalmitinand [1-'4Clpalmitate-labeled monopalmitin into steryl esters. Reactionmixtures were as described under "Materials and Methods." Acyl-labeledsteryl esters were extracted, isolated, and counted as described undermethod B of "Materials and Methods."
Table III. Effect of Various Lipids on [4-14C]Cholesteryl Ester Biosynthesis
Reaction mixtures were as described under Materials and Methods, except(a) each assay contained 8 mg of acetone-ether powder of the 20,000g pelletfraction (4.2 mg of protein) and (b) [4-14C]cholesterol and unlabeled lipidswere added to the acetone-ether powder in the organic solvent and at thetemperature designated. [4-14C]Cholesteryl esters were extracted, isolated,and counted as described under Materials and Methods.
Lipid Addition Organic Solvent Used 20 min 60 minExp. No. (0.2 mM) in Adding Lipids to Incubation IncubationAcetone-Ether Powder
% SE xSEActivity Activity
1 Control \,5 C Benzene 100 1001,2-Dipalmitin %5 C Benzene 154 1611,2-Diolein 05 C Benzene 295 2831,3-Dipalmitin '.5 C Benzene 118 125
2 Control 0 C Diethyl Ether 100 1001,2-Dipalmitin 0 C Diethyl Ether 219 1801-Monopalmitin 0 C Diethyl Ether 105 97Tripalmitin 0 C Diethyl Ether 100 96
3 Control 0 C Acetone 100 100Palmitic Acid 0 C Acetone 134 126Palmitoyl-CoA 0 C Acetone 68 89
4 Control 0 C Diethyl Ether 100 100PC (egg yolk) 0 C Diethyl Ether 624 560PE (egg yolk) 0 C Diethyl Ether 998 723
Plant Physiol. Vol. 62, 1978 ACYL DONOR IN STERYL ESTER BIOSYNTHESIS
12
10
0
E
www.plantphysiol.orgon March 21, 2020 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.

352 GARCIA AND MUDD Plant Physiol. Vol. 62, 1978
[4-'4C]cholesteryl ester biosynthesis and its ability to inhibit theincorporation of I1-'4C]palmitate from [_-'4CJpalmitoyl-CoA intosteryl ester suggest that fatty acyl-CoA was not the acyl donor inthese assays. The predominant reaction in the palmitoyl-CoAassays was its hydrolysis to palmitic acid and CoA, but palmiticacid was not the acyl donor since [16-'4Cjpalmitic acid was notincorporated into steryl ester. Significant amounts of labeled 1,2-diglycerides, however, were produced, and they may have beenthe actual acyl donors.The results suggest that the 1,2-diglyceride generated in the
degradation of phosphatidylcholine was the acyl donor involvedin spinach leaf steryl ester biosynthesis. The ability of [14Clacyl-labeled 1,2-diglyceride and [I-' Clpalmitate-labeled dipalmitin toincorporate label into steryl ester as well as the ability of unlabeled1,2-dipalmitin and 1,2-diolein to stimulate [4-"4C]cholesteryl esterbiosynthesis verify the 1,2-diglyceride acyl donor assignment.The theoretical maximum amount of labeled monoglyceride
produced by the end of the 60-min [l-14C]palmitate-labeled di-palmitin assay was approximately 9,000 cpm (monoglyceride +free fatty acid in Table II). This is less than the 90,000 cpm of 1-monopalmitin (45,000 cpm for each enantiomer) and about equalto the 10,000 cpm of 2-monopalmitin at the beginning of the [1-14Cjpalmitate-labeled monopalmitin assay. If either 1- or 2-mono-palmitin were the exclusive or preferred acyl donor for steryl esterbiosynthesis, then the initial rate of incorporation of label frommonopalmitin into cholesteryl palmitate should have been muchgreater than the rate of incorporation of label from dipalmitin intocholesteryl palmitate. Since this was not the case (Fig. 4), it is mostunlikely that 1- and 2-monopalmitin were the exclusive or pre-ferred acyl donors.Of the various acyl-labeled glycerides tested, ['4Clacyl-labeled
1,2-diglyceride was the most effective in terms oftotal radioactivityadded in incorporating label into steryl ester. [I-_4CjPalmitate-labeled dipalmitin was the next most effective. [1_-4C]Palmitate-labeled monopalmitin was the least effective. The heterogeneousfatty acid composition of [14C]acyl-labeled 1,2-diglyceride com-pared to the homogeneous fatty acid composition of [l-"CJpal-mitate-labeled dipalmitin may have accounted for the preferenceof [14Clacyl-labeled 1,2-diglyceride. The predominance of the 1,3isomer as well as the presence of 1,2-dipalmitin enantiomers in [I -4C]palmitate-labeled dipalmitin may also have been importantfactors.The ability of 1,2-dipalmitin and the inability of 1-monopal-
mitin and tripalmitin to stimulate steryl ester biosynthesis suggestan acylglycerol specificity (Table III). Since 1,2-dipalmitin stim-ulated steryl ester biosynthesis more effectively than did 1,3-dipalmitin, the enzyme exhibited a positional specificity. The
Table IV. Effect of 1,2-Diolein and 1,2-Dipalmitin on the Fatty AcidComposition of [4-14C]Cholesteryl Esters
Reaction mixtures were as described under Materials and Methods, except(a) [4-14C]cholesterol and unlabeled lipids were added to the acetone
powder in benzene at approximately 5 C and (b) benzene was removed byfreeze-drying. Incubation time was 60 min. [4-14C]Cholesteryl esters were
separated into various fatty acyl groups depending on their degree ofunsaturation by AgNO3 TLC as described under Materials and Methods.Samples were counted in a Nuclear-Chicago 720 series liquid scintillationsystem for 10 min and quenching was corrected by the channel ratio method.
No. of Dotuble AdditionsBonds in AcylMoiety of SE 0.2 mM 0.2 mM
Control 1,2-Dipalmitin 1,2-Diolein
SE dpm % SE dpm I SE dpm %
0 2,260 83 3,320 84 2,840 621 163 6 250 6 1,400 302 169 6 273 7 261 63 125 5 114 3 104 2
Total dpm 2,717 3,957 4,605
Recovery 88 91 90
PHOSPHATI DYLCHOLINE
phosphol/lpose D 1*.CHOLINE
PHOSPHATIDIC ACID
phosphc/tdic acid kphosphctose PHOSPH ATE
+ ucy//runsfercseSTEROL 1,2-DIGLYCERIDE -TTRRIGLYCERIDE
Gcyltfonsferose hydro/lse KFATTY ACID GCyltrGnsferOSe
STERYL ESTER iMONOGLYCER IDE
hydro/GseFATTY ACID
GLYCEROL
FIG. 5. Pathways involved in metabolism of phospholipids, glycerides,and steryl esters in spinach leaves.
ability of 1,2-diolein to stimulate steryl ester biosynthesis moreeffectively than 1,2-dipalmitin could mean that the enzyme ex-hibited a fatty acid specificity, but it could also mean that 1,2-diolein was more suitably dispersed in the aqueous solution and,therefore, more available for enzymic attack.
Since PC and PE were rapidly catabolized to 1,2-diglyceride byspinach leaf acetone powder preparations (Fig. 1), the PC (eggyolk) and PE (egg yolk) should also have been rapidly catabolizedto 1,2-diglyceride (Table III). The greater stimulation with the eggyolk phsopholipids could be explained if the heterogeneous fattyacid composition of their 1,2-diglyceride catabolites more nearlyapproximated the fatty acid specificity requirement of the enzymethan did 1,2-dipalmitin or 1,2-diolein. A second possibility is thatthe enzyme, like many membrane-bound enzymes (4), had aphospholipid cofactor requirement. A third possibility is that thegenerated diglyceride was the preferred isomer.
Figure 5 summarizes the various enzyme activities encounteredin spinach leaf acetone powder preparations during this investi-gation. The appearance of [I4C]acyl-labeled phosphatidic acidfrom ['4C]acyl-labeled phosphatidylcholine suggests that phospho-lipase D was present. The disappearance of label from phospha-tidic acid and the appearance of label in 1,2-diglyceride indicatethe presence of phosphatidic acid phosphatase. Cheniae (3) haspreviously shown that spinach leaf microsomes synthesize phos-phatidic acid, monoglyceride, diglyceride, and triglyceride whenincubated with labeled sn-glycerol-3-P. He has also shown thatphosphatidic acid in the precursor for these glycerides. Thisinvestigation has extended this earlier work by studying the me-tabolism of diglyceride and monoglyceride individually in spinachleaf acetone powder preparations and showing that their fattyacids are incorporated into several different metabolic pathways.One involved the transfer of an acyl moiety from 1,2-diglycerideto a free sterol. Although this is a new steryl ester biosyntheticpathway, its universal occurrence in plants requires further testing.
LITERATURE CITED
1. BARTLETT K, MJ KEAT, El MERCER 1974 Biosynthesis of sterol esters in Phycomyces blakes-leeanus. Phytochemistry 13: 1107-1113
2. BLIGH EG, WJ DYER 1959 A rapid method of total lipid extraction and purification. Can JBiochem Physiol 37: 911-917
3. CHENIAE GM 1965 Phosphatidic acid and glyceride synthesis by particles from spinach leaves.Plant Physiol 40: 235-243
4. COLEMAN R 1973 Membrane-bound enzymes and membrane ultrastructure. Biochim. Biophys www.plantphysiol.orgon March 21, 2020 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 62, 1978 ACYL DONOR IN STERYL ESTER BIOSYNTHESIS 353
Acta 300: 1-305. GALIIARI) T 1972 A simple biochemical method for the preparation of high specific activity
'4Clacyl-labeled phosphatidylcholine and other phospholipids. Biochim Biophys Acta 260:541-546
6. GAR(IA RE, JB Mum) 1978 Partial characterization of steryl ester biosynthesis in spinachleaves. Plant Phtiol 61: 354-356
7. GCARIA RE, JB Mum) 1978 Subcellular distribution of steryl ester biosynthesis in spinachleaves. Plant Physiol 61: 357-360
8. Gl.OMSET JA 1968 The plasma lecithin:cholesterol acyltransferase reaction. J Lipid Res 9:155-167
9. GOOoviDMAN DS, D DFYKIN, T SIIIRATIORI 1964 The formation of cholesterol esters with rat liverenzymes. J Biol Chenm 239: 1335-1345
10. HYUN J, H KOTIIARI, E HIERM, J MORTEINSON, CR TRE.ADWE.I.1., GV VAII()UNY 1969 Purificationand properties of pancreatic juice cholesteral esterase. J Biol ('hem 244: 1937-1945
I 1. KATES M 1954 Lecithinase systems in sugar beet, spinach, cabbage, and carrot. Can J BiochemPhysiol 32: 571-583
12. KNOC(ItH H, TW EsI)IRs, K Koriis, K Bt.o(-tt 1973 Palmityl coenzyme A inhibition of fattyacid synthesis. J Biol Chem 248: 2317-2322
13. Kot.ArFuKLJDY PE 1967 Mechanisms of synthesis of waxy esters in broccoli ( Brassica oleracea).
Biochemistry 6: 2705 271714. KOI.AITrruKUDY PE 1970 Plant waxes. Lipids 5: 259- 27515. LOWRY OH, NJ RoSEBtROMMtt, AL FARR, RJ RANDAL.L 1951 Protein measurement with the
Folin phenol reagent. J Biol Chem 193: 265-27516. MATITSON FH, RA VOItPENIIEI.N 1962 Synthesis and properties of glycerides. J Lipid Res 3:
281 -29617. MOORI. TS. JM LORI), T KAGAWA, H BEEVERS 1973 Enzymes of phospholipid metabolism in
the endoplasmic reticulum of castor bean endosperm. Plant Physiol 52: 50-5318. QUARitIS RH, RMC DAWSON 1969 The distribution of phospholipase D in developing and
mature plants. Biochem J 112: 787-79419. St:RDAREVtctII B 1967 Glyceride isomerization in lipid chemistry. J Am Oil Chem Soc 44:
381-39320. SKIt-SKI VP. M BARCLAY 1969 Thin-layer chromatography of lipids. Methods Enzymol 14:
530-59821. SPEC( IOR AA 1975 Fatty acid binding to plasma albumin. J Lipid Res 16: 165-17922. TAKETA K, BM POGEI.I 1966 The effect of palmityl coenzyme A on glucose 6-phosphate
dehydrogenase and other enzymes. J Biol Chem 241: 720-72623. VORRICK ML. GV MARINEI-II 1965 Separation of glycosyl diglycerides from phosphatides
using silicic acid column chromatography. J Lipid Res 6: 3-6
www.plantphysiol.orgon March 21, 2020 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.