identification of mouse langerin/cd207 in … national de la sante´ et de la recherche me´dicale...
TRANSCRIPT

Identification of mouse langerin/CD207 in Langerhans cells and somedendritic cells of lymphoid tissues
Valladeau, J., Clair-Moninot, V., Dezutter-Dambuyant, C., Pin, J. J., Kissenpfennig, A., Mattei, M. G., ... Saeland,S. (2002). Identification of mouse langerin/CD207 in Langerhans cells and some dendritic cells of lymphoidtissues. Journal of Immunology, 168(2)(2), 782-792.
Published in:Journal of Immunology
Document Version:Early version, also known as pre-print
Queen's University Belfast - Research Portal:Link to publication record in Queen's University Belfast Research Portal
General rightsCopyright for the publications made accessible via the Queen's University Belfast Research Portal is retained by the author(s) and / or othercopyright owners and it is a condition of accessing these publications that users recognise and abide by the legal requirements associatedwith these rights.
Take down policyThe Research Portal is Queen's institutional repository that provides access to Queen's research output. Every effort has been made toensure that content in the Research Portal does not infringe any person's rights, or applicable UK laws. If you discover content in theResearch Portal that you believe breaches copyright or violates any law, please contact [email protected].
Download date:08. Nov. 2018

Identification of Mouse Langerin/CD207 in Langerhans Cellsand Some Dendritic Cells of Lymphoid Tissues1
Jenny Valladeau,* Valerie Clair-Moninot,* Colette Dezutter-Dambuyant,† Jean-Jacques Pin,*Adrien Kissenpfennig,§ Marie-Genevieve Mattei,‡ Smina Ait-Yahia,* Elizabeth E. M. Bates,*Bernard Malissen,§ Franz Koch,¶ Francois Fossiez,* Nikolaus Romani,¶ Serge Lebecque,* andSem Saeland2*
Human (h)Langerin/CD207 is a C-type lectin of Langerhans cells (LC) that induces the formation of Birbeck granules (BG). Inthis study, we have cloned a cDNA-encoding mouse (m)Langerin. The predicted protein is 66% homologous to hLangerin withconservation of its particular features. The organization of human and mouse Langerin genes are similar, consisting of six exons,three of which encode the carbohydrate recognition domain. The mLangerin gene maps to chromosome 6D, syntenic to the humangene on chromosome 2p13. mLangerin protein, detected by a mAb as a 48-kDa species, is abundant in epidermal LC in situ andis down-regulated upon culture. A subset of cells also expresses mLangerin in bone marrow cultures supplemented with TGF-�.Notably, dendritic cells in thymic medulla are mLangerin-positive. By contrast, only scattered cells express mLangerin in lymphnodes and spleen. mLangerin mRNA is also detected in some nonlymphoid tissues (e.g., lung, liver, and heart). Similarly tohLangerin, a network of BG form upon transfection of mLangerin cDNA into fibroblasts. Interestingly, substitution of a conservedresidue (Phe244 to Leu) within the carbohydrate recognition domain transforms the BG in transfectant cells into structuresresembling cored tubules, previously described in mouse LC. Our findings should facilitate further characterization of mouse LC,and provide insight into a plasticity of dendritic cell organelles which may have important functional consequences. The Journalof Immunology, 2002, 168: 782–792.
D endritic cells (DC)3 are APCs with a key function in theimmune system. These cells are uniquely responsible forthe stimulation of naive T lymphocytes. DC have the
ability to capture and process foreign and self Ag, and to migrateand present processed Ag to T lymphocytes (1). Langerhans cells(LC), located in epidermis and stratified mucosal epithelia, con-stitute a distinct population of immature DC of bone marrow der-ivation that play a sentinel role in the capture of Ags from theexternal environment (2). Following Ag capture, LC differentiateinto mature DC as they migrate via lymphatics to the T cell areasof draining lymph nodes (3, 4). LC are characterized by Birbeckgranules (BG), which are organelles consisting of superimposedpentalamellar and zippered membranes (5, 6). We previously iden-
tified human (h)Langerin/CD207, a transmembrane C-type lectinof LC (7). Langerin is a potent inducer of BG formation and is anonconventional endocytic receptor routing into these organellesin LC, with potentially important consequences for Ag processingand presentation (7).
The DC system is continuously being defined both in human andmouse, and numerous subsets of DC with distinct phenotype andfunction have been described. The finding that murine thymic DCexpressing CD8� derive from a CD4low lymphoid progenitor hasled to definition of the lymphoid DC lineage (8). It has recentlybeen proposed that mouse LC originate from a lymphoid-commit-ted progenitor on the basis of CD8� expression (9, 10). However,to date, CD8� has not been observed on human DC. Furthermore,CD1a, which represents a marker of human LC, is absent in themouse. These examples illustrate the difficulties of translatingfunctional findings between human and mouse DC subsets, whichis key to optimizing preclinical models. In this work, we describethe identification of mouse (m)Langerin/CD207. The molecule dis-plays functional features similar to the human counterpart andshould be highly valuable for studying mouse LC. In addition,whereas wild-type mLangerin induces BG, a single amino acidsubstitution in the extracellular carbohydrate recognition domain(CRD) results in different cytoplasmic structures as observed byelectron microscopy. This finding suggests that different organellesmay form in LC depending on Langerin conformation and ligandinteraction with the sugar-binding region.
Materials and MethodsMice, media, and reagents
Mice of inbred strains C57BL/6 and BALB/c were purchased from CharlesRiver Breeding Laboratories, Iffa-Credo (L’Arbresle, France), and used at8–12 wk of age. All experiments involving mice were conducted according
*Schering-Plough Laboratory for Immunological Research, Dardilly, France; †In-stitut National de la Sante et de la Recherche Medicale Unite 346, CentreHospitalier Edouard Herriot, Lyon, France; ‡Institut National de la Sante et de laRecherche Medicale Unite 491, Faculte de Medecine, Marseille, France; §Centred’Immunologie, Institut National de la Sante et de la Recherche Medicale-CentreNational de la Recherche Scientifique, Marseille, France; ¶Department of Der-matology, University of Innsbruck, Innsbruck, Austria
Received for publication August 9, 2001. Accepted for publication November6, 2001.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 J.V. was the recipient of a grant from the Fondation Marcel Merieux (Lyon, France).2 Address correspondence and reprint requests to Dr. Sem Saeland, Schering-PloughLaboratory for Immunological Research, 27 chemin des Peupliers, 69571 DardillyCedex, France. E-mail address: [email protected] Abbreviations used in this paper: DC, dendritic cell; CRD, carbohydrate recognitiondomain; BG, Birbeck granule; LC, Langerhans cell; KCR, Kupffer cell receptor; m,mouse; h, human; FISH, fluorescence in situ hybridization; EST, expressed sequencetag; BLAST, basic local alignment search tool; DC-SIGN, dendritic cell-specificICAM-3-grabbing nonintegrin; ORF, open reading frame; ASGPR, asialoglycopro-tein receptor.
Copyright © 2002 by The American Association of Immunologists 0022-1767/02/$02.00

to institutional guidelines and all cell types were cultured in RPMI 1640(Life Technologies, Gaithersburg, MD) supplemented with 10% (v/v) heat-inactivated FBS (Flow Laboratories, Irvine, U.K.), 2 mM L-glutamine, 5 �10�5 M 2-ME (Sigma-Aldrich, St. Louis, MO), and gentamicin (Schering-Plough, Kenilworth, NJ) (hereafter referred to as complete medium). Cul-ture supernatant of a cell line transfected with the mouse GM-CSF gene(kind gift of A. Lanzavecchia, Belinzona, Switzerland) was used as asource for this cytokine (0.5% v/v), and human TGF-�1 (specific activity1–2 � 107 U/mg) was purchased from Strathmann Biotech (Hamburg,Germany). Transfections were performed with the murine fibroblasticCOP5 cell line (11) as described (7).
Identification of mLangerin cDNA and cloning
Bioinformatics searches were performed in GenBank EST databases. Twoexpressed sequence tags (ESTs) were used for PCR amplification of cDNAfrom a mouse lung cDNA library (Clontech Laboratories, Palo Alto, CA)using the RACE marathon kit (Clontech Laboratories). Primer sequenceswere for the 5� RACE (GSP1, TTGGAGCCTCAGGGAGAGAGACAT;and NGSP1, GTTGTTCCCTGCGTTGTTGGGTTC), and for the 3�RACE (GSP2, GGAGAATGTCAACAAGCTGCTCAA; and NGSP2,AAAGCTCACCTGACCTCAGTGTCC). PCR products were cloned inthe PCRII TOPO plasmid (Invitrogen, San Diego, CA) and double-stranded plasmid DNA was sequenced on an ABI 373A sequencer (Ap-plied Biosystems, Foster City, CA) using dye terminator technology. Se-quencher (Gene Codes, Ann Arbor, MI) and Lasergene (DNAstar, London,U.K.) software was used to analyze sequences. Comparisons against Gen-Bank databases were made using the basic local alignment search tool(BLAST) algorithm. Construction of Langerin-mutated forms was per-formed with the Gene Editor in vitro Site-Directed Mutagenesis kit (Pro-mega, Madison, WI), according to instruction guidelines and using wild-type Langerin cDNAs as templates and the mutated oligonucleotides 5�-ATCTCCCTCTGGATCCGAGAGCCTCCT (hLangerin WPREPP toWIREPP) and 5�-GAACAAAAGTTACTCTACAAG (mLangerinQKFLYK to QKLLYK).
Northern and Southern analysis
Northern analysis was performed with Multiple Tissue Northern blots pre-pared from high-quality poly(A)� RNA normalized for a �-actin hybrid-ization signal (Clontech Laboratories, catalog no. 7762-1 and 7763-1), asdescribed (12). For Southern analysis, DNA (5 �g) from primary amplifiedcDNA libraries constructed in our laboratory was digested with NotI andSalI (Boehringer Mannheim, Indianapolis, IN) to release the inserts, run ona 1% agarose gel, and transferred to a nylon membrane (Schleicher andSchuell, Keene, NH). Hybridization was conducted using a PCR-generatedprobe labeled with [32P]dCTP using the High Prime kit (Boehringer Mann-heim, Meylan, France). Unincorporated nucleotides were removed by spincolumn chromatography (Chromaspin-100; Clontech Laboratories). Hy-bridization and washes were performed in stringent conditions (0.1� SSC/0.1% SDS at 65°C). Membranes were exposed for 21 days using BiomaxMR film (Kodak, Rochester, NY). Characteristics (number of indepen-dent clones/percentage of clones with inserts) of the cDNA librariesused were as follows: wild-type thymus, 6.6 � 106/43%; rag-1�/� thy-mus, 3.9 � 106/85%; pancreas, 2.2 � 108/97%; skin, 1.4 � 107/90%;spinal cord, 4.4 � 107/95%; spleen, 1.1 � 108/72%; lung, 3.7 � 106/85%; stomach, 4.0 � 106/73%; Peyer’s patch, 8 � 106/72%; mesentericlymph node, 5.8 � 106/83%; and colon, 5.4 � 106/90%.
Genomic organization and chromosomal localization ofLangerin genes
A 900-bp EcoRI fragment of mLangerin cDNA was used as a probe to hy-bridize a 129/Ola mouse spleen genomic cosmid 121 library (Resource Center/Primary Database, Max Planck Institute for Molecular Genetics, Heidelberg,Germany). Hybridization was performed as described (13), and positive cloneswere selected and sequenced. Seven overlapping cosmid clones were found tobe positive for the Langerin gene, as follows: MPMGc121BO4216Q2,MPMGc 121A22222Q2, MPMGc121F01387Q2, MPMGc121K20580Q2,MPMGc 121D19640Q2, MPMGc121K20694Q2, and MPMGc121HO2
719Q2. One cosmid was used for fluorescence in situ hybridization (FISH)mapping of the mouse gene as described (14). The organization of thehLangerin gene was obtained by BLAST nucleotide using the GenBankhigh throughput genomic database. Chromosomal localization of thehLangerin gene was performed with the Stanford G3 RH mediumresolution panel (Research Genetics, Huntsville, AL), using PCR witholigonucleotides that amplify an 850-bp fragment specific for thehLangerin gene (forward, 5�-ACGCCATTCAACAAGGTC; reverse,
5�-CAGCCAAGACAGACGGACTC). The result was scored manuallyand analysis was performed with the RHmapper program(http://shgc-www.stanford.edu).
Generation of Abs against mLangerin
mAbs were produced by immunizing BALB/c mice with three i.p. injec-tions of lysate of COP5 cells transfected with mLangerin cDNA. Spleenswere removed 3 days after a final i.v. injection and splenocytes were fusedwith SP2/0 cells using polyethylene glycol-1000 and cultured in 96-wellplates using standard procedures. Hybridoma supernatants were screenedfor reactivity against mLangerin transfected vs nontransfected cells. Aftercloning of selected hybridomas, mAbs were produced as ascites fluids andpurified. Data presented were obtained with mAb HD24 (306.G9) of IgG1isotype. Polyclonal Ab was obtained by immunizing rabbits with mLan-gerin peptides followed by purification from antiserum using affinity chro-matography (CovalAb, Oullins, France).
Biochemistry
COP5 fibroblastic cells transfected with mLangerin cDNA were washedthree times with ice-cold PBS, pH 8 and then treated in a lysis buffercontaining 50 mM Tris-HCl, pH 8, 1% Nonidet P-40, 150 mM NaCl, andprotease inhibitors (Boehringer Mannheim). Lysates were incubated at 4°Cfor 20 min, and insoluble material was pelleted by centrifugation at12,000 � g for 10 min at 4°C. Soluble extracts were precleared three timeswith control mAb and protein G-agarose (Boehringer Mannheim). Theextract was then incubated with mAb HD24 for 1 h before adding proteinG-agarose for 3 h. Beads were washed three times in lysis buffer, resus-pended in SDS-PAGE sample buffer with or without 5% 2-ME, boiled for3 min, and centrifuged. Immunoprecipitates were separated by SDS-PAGEusing 12% polyacrylamide gel and then transferred to a polyvinylidenedifluoride membrane (Immobilon P; Millipore, Bedford, MA). Blots wereblocked with 1% BSA and 0.1% Tween 20 in PBS, then incubated withHRP-conjugated mAb HD24 for 30 min. Proteins were detected by ECL(Boehringer Mannheim).
Epidermal sheets, epidermal cell suspensions, and bonemarrow-derived dendritic cells
Epidermis from ear skin was separated from dermis by means of ammo-niumthiocyanate (15). Resulting sheets were fixed in acetone for 20 min atroom temperature and rinsed in several changes of PBS and PBS contain-ing 1% BSA. They were then subjected to an immunolabeling procedure asdescribed (16). Bone marrow-derived DC were generated essentially asdescribed by Inaba et al. (17), in the presence of GM-CSF supplemented ornot with TGF-�1. Cultures were analyzed on acetone-fixed cytospins ondays 6, 8, 10, 13, and 19 of culture.
Immunohistochemistry and cytofluorometry
For immunohistochemistry of sheets and cytospins, the following mAbswere used: mouse IgG1 anti-mLangerin, clone HD24 (used at final con-centrations of 1–2 �g/ml), mouse anti-I-Ek,d (clone 14-4-4S/HB32, mouseIgG2a, American Type Culture Collection, Manassas, VA), anti-I-Adiverse
(clone 2G9, fluoresceinated, BD PharMingen, San Diego, CA) and controlmouse IgG1 (DAKO, Glostrup, Denmark) used at 5–10 �g/ml. Stainingwas visualized by immunofluorescence using species-specific biotinylatedanti-mouse Ig (Amersham, Little Chalfont, U.K.) followed by streptavidin-FITC (Amersham). For double-labeling, streptavidin-Texas Red (Amer-sham) was used (instead of streptavidin-FITC), followed by rat Ig (100�g/ml) for blocking residual binding sites of preceding Abs, and FITC-conjugated anti-MHC class II mAb 2G9. Immunolabeled specimens weremounted in Vectashield (Vector Laboratories, Burlingame, CA) and viewed ona conventional fluorescence microscope (Olympus, Melville, NY).
For immunohistology, microscope slides of acetone-fixed cryocutmouse tissue sections were incubated with polyclonal anti-mLangerin for60 min, subsequently with biotinylated goat anti-rabbit Ig (Vector Labo-ratories) for 30 min, and finally with streptavidin coupled to peroxidase(Sigma-Aldrich) for 30 min. Anti-DEC205 mAb (BMA, Bubendorf, Swit-zerland) was revealed with peroxidase-conjugated affinity purified goat anti-rat IgG (Jackson ImmunoResearch Laboratories, West Grove, PA). En-zyme activity was developed using 3-amino-9 ethylcarbazol highsensitivity (DAKO) and slides were counterstained with Harris hematox-ylin (Sigma-Aldrich).
783The Journal of Immunology

Flow cytometric analysis on COP5 fibroblasts was performed on aFACSCalibur (BD Biosciences, Mountain View, CA), following stainingwith 10 �g/ml anti-mLangerin mAb HD24 or anti-hLangerin mAb(DCGM4) (18) revealed by PE- or FITC-conjugated F(ab�)2 goat anti-
mouse Ig (DAKO). Nonspecific staining was determined using isotype-matched control mAb and all procedures were performed in the presence of0.5 mM EDTA to avoid cell aggregation. Intracytoplasmic staining wasperformed in saponin (0.1% v/v).
FIGURE 1. mLangerin cDNA sequence and alignments. A, Nucleotide and predicted amino-acid sequences of mLangerin cDNA (accession no.AJ302711). The boxed sequence (hydrophobic amino acids 47–67) forms a signal anchor expected to represent the transmembrane domain. The C-typelectin domain with two cysteines forming a disulfide bridge (rounded) is shown in single underline. Broken and double underlines (positions 26–31, 90–92,and 116–118) indicate the proline-rich domain and potential N-glycosylation sites, respectively. B, Alignments of mouse and human Langerin. Conservedamino acids are shaded and the boxed sequence forms a signal anchor expected to represent the transmembrane domain.
784 IDENTIFICATION OF mLANGERIN

Electron microscopy
Staining was performed as described (18). Briefly, COP5 transfectedwith mLangerin cDNA were fixed for 18 h with 2% glutaraldehyde incacodylate buffer, followed by washing for at least 24 h in cacodylatebuffer with sucrose. Samples were postfixed for 1 h with 1% osmium incacodylate buffer with sucrose, dehydrated, and embedded in epoxyresin. Ultrathin sections were poststained with uranyl acetate and leadcitrate and examined on a JEOL 1200 EX electron microscope (JEOL,Tokyo, Japan) (7).
ResultsIdentification of mLangerin/CD207 cDNA
Public databases were screened using a translated BLAST nucle-otide algorithm with the amino acid sequence of hLangerin/CD207as query. Two mouse EST (AA76540 and AA423304) were iden-tified and this sequence used in a 3� and 5� RACE PCR on a mouselung cDNA library. A 1486-bp cDNA sequence was isolated, con-taining a 981-bp open reading frame (ORF) but lacking a startmethionine. Using this cDNA as a probe, two cosmid clones wereisolated and sequenced. Upstream of the 5� end of the cDNA se-quence, a potential initiation site (YCANTYY) followed by twomethionines was identified. Thus, we isolated a 1565-bp full-length cDNA sequence encoding mLangerin/CD207 (accessionno. AJ302711). It contains a 58-bp 5� untranslated region, a 993-bpORF, a 3� untranslated region of 514 nt, and an AATAAA poly-adenylation signal at position 1511 bp. The ORF predicts a novelpolypeptide of 331 aa (Fig. 1A), with a molecular mass of 37.5kDa. As described for hLangerin, the presence of a hydrophobicsignal anchor (residues 47–67) indicates a type-II transmembraneprotein. mLangerin belongs to the Ca2�-dependent (C-type) lectinfamily with a single CRD (Fig. 1A) featuring a glutamate-proline-asparagine motif (residues 288–290), predicting mannose-typespecificity (19). Two potential N-glycosylation sites are present atpositions 90–92 and 116–118 aa. mLangerin contains an intracel-lular domain of 46 aa with a proline-rich motif (PREPPP) as po-tential signal-transduction site (20, 21). mLangerin displays 66%overall amino acid sequence identity with its human counterpart(Fig. 1B).
As described for hLangerin, amino acid sequence comparison inthe European Molecular Biology Laboratory database revealedmarked homology between mLangerin and several type-II lectins,mainly in their CRD. The mLangerin CRD displays the highestamino acid homology with hLangerin (75.8%), followed by therodent Kupffer cell receptors (KCR; 46.1%) (22), chicken hepaticlectin (34.0%) (23), and the human asialoglycoprotein receptors(ASGPR) (24–26) (Fig. 2).
Characterization of Langerin/CD207 genes
A genomic cosmid library from mouse spleen was hybridized byusing the ESTs of mLangerin cDNA as a probe. Positive cloneswere propagated, digested, and hybridized to identify cosmids con-taining the entire mLangerin gene. Two cosmid clones of �40 kbpwere used for subcloning and direct sequencing. The mLangeringene spans 6.7 kbp and consists of six exons varying in lengthbetween 117 and 624 bp (Fig. 3, A and B) (accession no.AJ313164) The first exon encodes half of the intracytoplasmicdomain, the second exon encodes the end of the intracytoplasmicdomain and the transmembrane domain, the third exon encodes thelong neck domain, and three exons encode the CRD. Notably, theproline-rich motif is encoded by two exons. The size of the fiveintrons is highly variable, ranging from 92 to 1.4 kbp (intron 4).Using FISH, the mLangerin gene was mapped to chromosome 6D(Fig. 3C).
Furthermore, we obtained the organization of the hLangeringene. A BAC clone (NH0504001) containing the entire hLan-gerin gene was identified by BLAST nucleotide analysis of theGenBank high throughput genomic database, and introns weremapped for this gene. The hLangerin gene, spanning 5.6 kbp,also consists of six exons and has a structure comparable to themouse (Fig. 3, A and B). For human and mouse Langerin allintrons (except the first one) use consensus GT-AG type-IIsplice sites (27, 28), indicating that some form of transcriptionalcontrol may regulate these genes. Computer analysis of theupstream regions of the human and mouse Langerin genesrevealed that these sequences do not contain consensus pro-moter elements. However, regions of homology between thetwo genes may delineate binding sites for novel LC-specifictranscription factors and thus explain the extremely restrictedexpression of these genes. Finally, by radiation hybrid mapping,the hLangerin gene was assigned to chromosome 2p13 syntenicto the localization of the mouse gene (data not shown).
Distribution of mLangerin/CD207 mRNA
By Northern blot analysis, a unique band at the expected size of�2.0 kb was observed in spleen, lung, liver, and heart (Fig. 4A).Southern blot analysis of cDNA libraries from various mouse tis-sues (Fig. 4B) indicated the predominant presence of mLangerinmRNA in skin, in accordance with the previously reported expres-sion of hLangerin (18). Consistent with Northern blot analysis, asignal was also detected in spleen and lung. In addition, mLangerinmRNA was found in mesenteric lymph node and thymus, where itwas markedly enhanced in Rag1�/� mice (Fig. 4B). In contrast tothe above findings, a number of tissues did not reveal mLangerin
FIGURE 2. Analysis of the C-type lectin domain of mLangerin. Alignments of the lectin domain of mLangerin with other C-type lectins are indicatedas follows: conserved amino acids are shaded, residues of the consensus motif of this family are boxed, and open arrows show the part of the Ca2� bindingsite involved in sugar specificity.
785The Journal of Immunology

mRNA (Fig. 4). These results indicate that mLangerin mRNA ismostly expressed in skin and hematopoietic tissues but is also de-tectable in lung, liver, and heart.
Characterization of mLangerin/CD207 protein and mAbs
By immunization of BALB/c mice with a murine cell line (COP5)transfected with mLangerin cDNA, we obtained a panel of mAbsagainst mLangerin (scoring selectively positive on the mLangerintransfectant vs mock-transfected cells). One mAb, named HD24(clone 306G9, IgG1), was chosen for further studies. Immunopre-cipitation revealed that HD24 mAb selectively recognizes a gly-coprotein of �48 kDa in mLangerin-transfected COP5 cells, inaccordance with the molecular mass predicted from the amino-acidsequence (Fig. 5A). Flow cytometric analysis on transfectants ob-
tained with different constructs of mouse and human Langerindemonstrated that mAb HD24 cross-reacts with hLangerin and isdirected against an intracellular epitope containing the proline-richmotif as reactivity is abolished following a site-directed mutationof WPREPP to WIREPP (Fig. 5B).
Expression of mLangerin/CD207 protein in vivo
Because hLangerin is selectively expressed by LC, we first per-formed immunohistology on mouse skin sections. Expression ofmLangerin was observed by cells with LC morphology in the epi-dermis but not by cells in the dermis (Fig. 6A). As shown in Fig.6B, bright staining was obtained on ear epidermal sheets ofBALB/c (n � 3), C57BL/6 (n � 2), and 129 (n � 1) mice. Stainingwas highly specific to the LC, and keratinocytes were completely
FIGURE 3. Organization of human and mouse Langerin genes. A, Schematic representation of the genomic organization. Accession numbers: mouse,AJ313164; human, NT005420. B, Positions of introns and exons. C, FISH of the mLangerin gene showing localization on chromosome 6D.
786 IDENTIFICATION OF mLANGERIN

negative. Under the fluorescence microscope, Langerin expres-sion was visible as granules scattered internally throughout thecell, and no staining was observed on the cell surface. Thegranules were found even in the most distant parts of thedendrites where MHC class II molecules were too sparse to bevisualized (Fig. 6B). Finally, this double-labeling revealed thatvirtually all LC (as identified by their MHC class II expression)were also Langerin positive. Inversely, all Langerin-positivecells were MHC class II positive, emphasizing the specificity ofthe Ab. In corresponding dermal sheets we observed only veryfew Langerin-positive cells, presumably LC in transit (data notshown).
Because mLangerin mRNA was also found in other lymphoidtissues, we analyzed the protein expression in thymus, spleen, andlymph nodes. Of interest, many cells expressing Langerin wereobserved in the thymic medulla (Fig. 6, C and E), localized by aserial staining with DEC205 (Fig. 6D; DEC205low areas). The
Langerin� cells in the medulla had a characteristic dendritic mor-phology (Fig. 6E). Some Langerin� cells, with characteristic den-dritic morphology, were occasionally found in the cortex (Fig. 6F).In spleen (Fig. 6G) and lymph node (Fig. 6H), only few Langerin�
cells were observed in perifollicular areas, expected to be marginalDC or interdigitating cells, as some of them possess dendrites. Wenever observed Langerin� cells in germinal centers. Taken to-gether, mLangerin is expressed by LC and a subset of DC in lym-phoid tissues including thymic medulla.
mLangerin/CD207 is expressed in bone marrow culturessupplemented with TGF-�
DC can be obtained from mouse bone marrow progenitor cellsin culture with GM-CSF (17). By FACS analysis of suchpermeabilized bone marrow-derived DC, only very weak if anyLangerin staining could be detected. Indeed, inspection of
FIGURE 4. mLangerin mRNA expression in a panel of lymphoid and nonlymphoid tissues. A, Northern blot analysis. B, Southern blot analysis.
FIGURE 5. Characterization of anti-mLangerin mAb and mLangerin protein. A, Immunoprecipitation with mAb HD24 on mLangerin-transfectedfibroblasts. B, Staining on COP5 fibroblasts transfected with various forms of mouse and human Langerin cDNA. Cells were permeabilized with saponin,stained with DCGM4 or HD24, and analyzed by FACS. Filled histograms were obtained with control fibroblasts (COP mock). M, Molecular mass; COPmock, control fibroblasts; COP mLg, COP transfected with mLangerin cDNA; WT, wild type.
787The Journal of Immunology

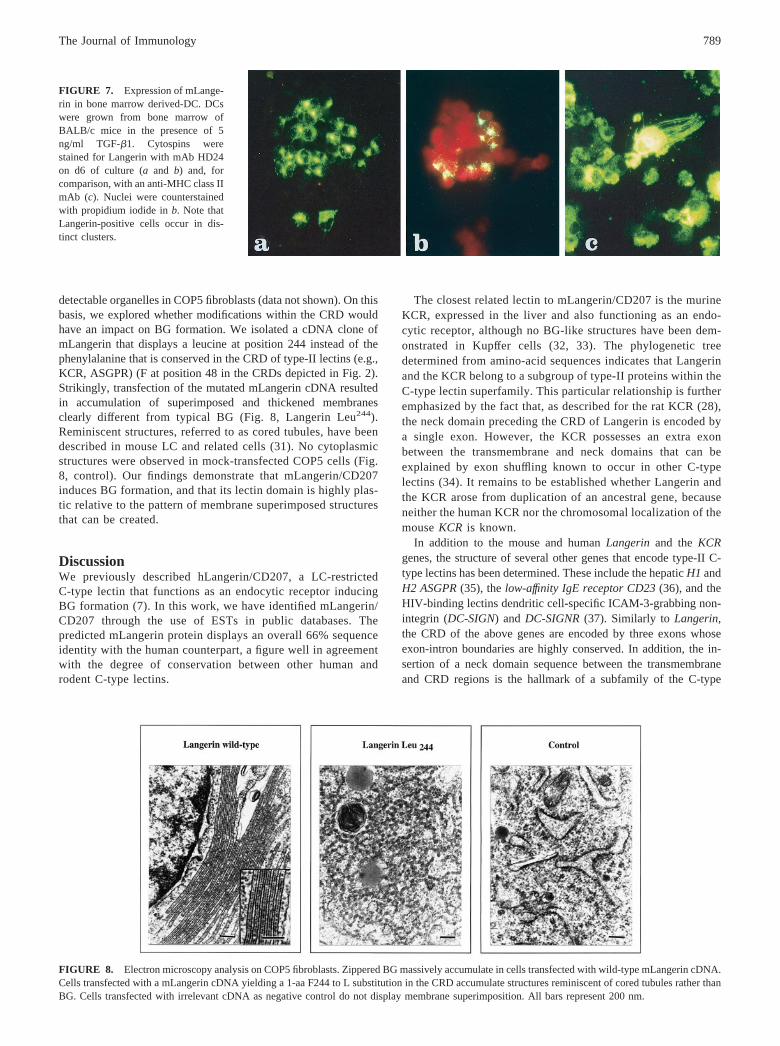
cytospins from standard cultures in the presence of GM-CSFrevealed only a very small number (1% of all cells) of brightlyLangerin-positive cells (data not shown). TGF-� is a cytokineknown to promote the differentiation of LC (29). When TGF-�was added to the cultures (at both 0.5 and 5 ng/ml), the numberof Langerin-positive cells increased up to nearly 10% (Fig. 7).Of note, Langerin-expressing cells occurred preferably in cellaggregates. No obvious differences in the percentages of Lan-gerin-positive cells were noted between various time points(day 6, 8, 10, 13, and 19).
LC from epidermal bulk cultures mature into potent immuno-stimulatory DC in 3 days and typically lose their BG, as demonstratedby electron microscopy (30). While fresh mouse LC expressed Lan-gerin, 3 day-cultured epidermal cells expressed markedly reduced lev-els, and many cells became Langerin negative (data not shown), fur-ther confirming that Langerin is restricted to immature DC. Taken
together, these results demonstrate that mLangerin is regulated in thesame manner as its human counterpart, increased by TGF-� and de-creased by the LC maturation process.
Transfection of mLangerin/CD207 cDNA induces BG, whereas aPhe244 to Leu replacement induces structures resembling coredtubules
Because transfection of hLangerin cDNA induced the formation ofBG in fibroblasts (7), we analyzed by electron microscopy COP5mouse fibroblasts transfected with mLangerin cDNA. In a mannerreminiscent of hLangerin, COP5 cells transfected with the murinecDNA massively accumulated pentalamellar membranes with acentral zippering typical of BG (Fig. 8, Langerin wild-type). Fur-thermore, a truncated hLangerin cDNA construct entirely devoidof the C-type lectin domain (CRD) did not induce BG or any
FIGURE 6. Expression of mLangerinprotein in vivo. mLangerin expression byLC on frozen skin sections with anti-mLangerin polyclonal Ab (A), or doublestaining with anti-mLangerin mAb HD24(red) and FITC-anti-MHC class II mAb2G9 (green) on epidermal sheet (B). In thy-mus, polyclonal anti-mLangerin (red) re-veals mLangerin� DC in the medulla (Cand E) and some in the cortex as localizedby a DEC205 serial staining (D) revealingcortical epithelial cells and indigitatingcells. At higher magnification, the mLange-rin� DC show dendritic morphology (F). Inspleen, some mLangerin� cells (G) are ob-served in perifollicular areas, as in lymphnode (H). No staining was detected withcontrol Abs (not shown). Magnification: A,�200; B, �1000; C and D, �40; E, �200;F, �400; G, �100; and H, �400.
788 IDENTIFICATION OF mLANGERIN
detectable organelles in COP5 fibroblasts (data not shown). On thisbasis, we explored whether modifications within the CRD wouldhave an impact on BG formation. We isolated a cDNA clone ofmLangerin that displays a leucine at position 244 instead of thephenylalanine that is conserved in the CRD of type-II lectins (e.g.,KCR, ASGPR) (F at position 48 in the CRDs depicted in Fig. 2).Strikingly, transfection of the mutated mLangerin cDNA resultedin accumulation of superimposed and thickened membranesclearly different from typical BG (Fig. 8, Langerin Leu244).Reminiscent structures, referred to as cored tubules, have beendescribed in mouse LC and related cells (31). No cytoplasmicstructures were observed in mock-transfected COP5 cells (Fig.8, control). Our findings demonstrate that mLangerin/CD207induces BG formation, and that its lectin domain is highly plas-tic relative to the pattern of membrane superimposed structuresthat can be created.
DiscussionWe previously described hLangerin/CD207, a LC-restrictedC-type lectin that functions as an endocytic receptor inducingBG formation (7). In this work, we have identified mLangerin/CD207 through the use of ESTs in public databases. Thepredicted mLangerin protein displays an overall 66% sequenceidentity with the human counterpart, a figure well in agreementwith the degree of conservation between other human androdent C-type lectins.
The closest related lectin to mLangerin/CD207 is the murineKCR, expressed in the liver and also functioning as an endo-cytic receptor, although no BG-like structures have been dem-onstrated in Kupffer cells (32, 33). The phylogenetic treedetermined from amino-acid sequences indicates that Langerinand the KCR belong to a subgroup of type-II proteins within theC-type lectin superfamily. This particular relationship is furtheremphasized by the fact that, as described for the rat KCR (28),the neck domain preceding the CRD of Langerin is encoded bya single exon. However, the KCR possesses an extra exonbetween the transmembrane and neck domains that can beexplained by exon shuffling known to occur in other C-typelectins (34). It remains to be established whether Langerin andthe KCR arose from duplication of an ancestral gene, becauseneither the human KCR nor the chromosomal localization of themouse KCR is known.
In addition to the mouse and human Langerin and the KCRgenes, the structure of several other genes that encode type-II C-type lectins has been determined. These include the hepatic H1 andH2 ASGPR (35), the low-affinity IgE receptor CD23 (36), and theHIV-binding lectins dendritic cell-specific ICAM-3-grabbing non-integrin (DC-SIGN) and DC-SIGNR (37). Similarly to Langerin,the CRD of the above genes are encoded by three exons whoseexon-intron boundaries are highly conserved. In addition, the in-sertion of a neck domain sequence between the transmembraneand CRD regions is the hallmark of a subfamily of the C-type
FIGURE 8. Electron microscopy analysis on COP5 fibroblasts. Zippered BG massively accumulate in cells transfected with wild-type mLangerin cDNA.Cells transfected with a mLangerin cDNA yielding a 1-aa F244 to L substitution in the CRD accumulate structures reminiscent of cored tubules rather thanBG. Cells transfected with irrelevant cDNA as negative control do not display membrane superimposition. All bars represent 200 nm.
FIGURE 7. Expression of mLange-rin in bone marrow derived-DC. DCswere grown from bone marrow ofBALB/c mice in the presence of 5ng/ml TGF-�1. Cytospins werestained for Langerin with mAb HD24on d6 of culture (a and b) and, forcomparison, with an anti-MHC class IImAb (c). Nuclei were counterstainedwith propidium iodide in b. Note thatLangerin-positive cells occur in dis-tinct clusters.
789The Journal of Immunology

lectins (38). The neck domain of Langerin features a characteristicheptad repeat producing an �-helical coiled-coil stalk (38). Heptadrepeats are important for dimerization/oligomerization, as illus-trated for CD23 (39, 40) and the H1 and H2 subunits of the hepaticASGPR (41). This raises the possibility that Langerin occurs as adimer/oligomer, although we have no experimental evidence sup-porting this hypothesis.
The gene encoding mLangerin maps to chromosome 6D, in aregion syntenic to the hLangerin gene on chromosome 2p13. TheLangerin genes do not belong to any of the known C-type lectingene clusters, such as the human DC-SIGN and CD23 genes (chro-mosome 19p13) (37), the human ASGPR (chromosome 17p11-13)(42), and the human NKR complex (chromosome 12p13-p12) (43).However, due to their particularly close relationship the KCRgenes may turn out to cluster with Langerin.
One of the characteristic features of DC is the expression ofmany C-type lectins. DC express endocytic receptors for cap-turing exogenous carbohydrate-bearing material for pathogenrecognition and Ag routing. Endocytic receptors are exempli-fied by the type-I lectin DEC205 (44, 45) and the type-II humanlectins Langerin (18) and DC-SIGN, which sequesters HIVthrough binding to gp120 (46). However, a given lectin mayhave more than one function, as demonstrated for DC-SIGN,which binds ICAM-3 and ICAM-2 and thus has an impact onDC/T cell interactions and DC migration (47, 48). In addition,type-II lectins such as DCIR (49) and dectin-1 (50) containintracellular immunoreceptor sequences predictive of a role ininhibition or activation of DC function. Consequently, it is animportant issue to determine whether multiple function is ageneral feature of DC lectins and whether lectins on a given DCsubset have overlapping functions.
Langerin is abundantly expressed by mouse epidermal LC, bothat mRNA and protein levels. Using mAb HD24, Langerin couldnot be detected in other epidermal cells, and could only be detectedin scattered DC in the dermis. Langerin should thus be a highlyuseful molecular marker for identification and functional studies ofmouse LC. This has been particularly hampered by paucity of re-agents, because the mouse lacks CD1a, a useful marker of humanLC. Mouse LC do express DEC205, but this molecule is alsowidely expressed by mature DC in lymphoid tissue (45). The co-expression of DEC205 and Langerin on mouse LC raises the pos-sibility of some shared function on this DC subset. Langerin isdown-regulated upon culture of mouse epidermal LC (data notshown), in line with the notion that it represents a feature of im-mature cells. Decreased expression of Langerin agrees with earlierobservations describing the maturation of mouse LC in cultureresulting in disappearance of BG (30). Scattered Langerin� cellsare observed in lymph nodes and spleen. Langerin�CD11c� cellsexpressing high levels of DEC205 are particularly conspicuous inskin-draining lymph nodes, where they represent a mature form ofLC immigrated from the epidermis (51). mLangerin� cells inspleen have recently been shown to represent CD11c�CD8�� DC(K. Inaba, personal communication). Langerin is also present onDC within the mouse thymic medulla. This finding warrants sev-eral comments and questions. First, the possibility that Langerin�
thymic DC and LC have a common origin. Mouse thymic DC,which include a major CD8�� subset (52), are essentially of lym-phoid origin (8). Recently, lymphoid-committed CD4low precur-sors were shown to generate LC upon adoptive transfer (9). Al-though CD8� is not a marker of lymphoid origin (53), it is strikingthat mouse LC can be induced to express this marker (9, 10).Second, the expression of Langerin by thymic DC is consistentwith the description of BG in these cells, in both mouse (54) andhuman (55). Third, the presence of Langerin on thymic DC opens
the interesting perspective that Langerin may play a role in T cellselection, possibly by endocytosis of self-derived mannosylatedAgs. The fact that DEC205 is also expressed by thymic DC (52)further points to overlapping function between these two lectins. Inaddition to skin, mLangerin mRNA is detected in some nonlym-phoid organs. In particular, lung tissue was positive, similar to theexpression of hLangerin in epithelium lining the human airways(18). The significance of mLangerin mRNA in liver and heart re-mains to be explored.
A subset (�10%) of Langerin� cells was generated in cul-tures of mouse bone marrow in the presence of GM-CSF andTGF-�1. The role of TGF-� in expanding the Langerin� subsetis in line with the effects of this cytokine in up-regulation ofhLangerin in human LC cultures (18) and its critical function inmouse LC development (29, 56). Langerin should be a highlyuseful marker to optimize conditions for in vitro generation ofmouse LC that is currently a limitation to functional studies onthis cell type.
Mouse and human Langerin share the capacity to induce theformation of pentalamellar membranes typical of BG. Thisproperty is conserved through evolution and emphasizes thepotential importance of the molecule in LC function. Althoughintroduction of Langerin cDNA does not create “tennis racket”structures sometimes observed in LC, it should be noted thatthese particular structures likely represent only a minority of theBG, probably resulting from fusion with endocytic vesicles thatis not likely to occur in the transfected fibroblasts because mostof the Langerin molecules will superimpose before reaching thecytomembrane. The prediction of our previous (7) and presentfindings is that Langerin plays a particular role in endocytosisof foreign Ags (e.g., mannosylated microorganisms) in theperiphery, and of self Ags in the thymus. Routing of Ag into BGcould represent the feature of a distinctive DC sublineage foundin epidermis, stratified epithelia, and thymus. It would be ofinterest to examine the BG-inducing potential of other lectins(e.g., CD23, DEC205) expressed by the same cells. Deletion ofthe entire lectin domain of Langerin totally abolished themembrane superimposition effect in transfectants, emphasizingthe role of carbohydrate ligand binding in BG formation.However, the nature of the physiological ligand(s) of Langerinremains to be determined. Strikingly, replacement of the highlyconserved F244 by L in the �2 helix of the mLangerin CRDabrogated formation of the rod-shaped zippered membrane super-impositions typical of BG. The L244 form of mLangerin gave riseto structures reminiscent of cored tubules described in mouseLC-type cells (57, 58). As viewed by electron microscopy, coredtubules do have an inner central line, but their structure is differentfrom the disc-shaped BG. Cored tubules have been observed inmouse LC and related cells (31), although their relationship withBG is unknown. We propose that the relationship between thesedifferent organelles is determined by the conformation of Lange-rin. Replacement of F244 by L, both hydrophobic residues, is notexpected to disrupt the �2 helix of the Langerin CRD (59). Rather,a three-dimensional model using the recently determined crystalstructure of the H1 subunit of the ASGPR (60) indicates thatabsence of the aromatic ring of F244 is likely to abolish itsinteraction with the parallel and tightly juxtaposed aromatic ring ofF205. The substitution of F244 with L may thus open the hydro-phobic pocket in this region of the CRD, thus contributing to achange in the domain associated with receptor multimerization orwith ligand binding. It has been suggested that CRD sequencesflanking the coiled-coil stalk may also contribute to C-type lectinsubunit assembly (41). The transition from BG to cored tubulescould thus be a consequence of altered conformation of the
790 IDENTIFICATION OF mLANGERIN

receptor-ligand complex. However, it remains to be determinedwhether the F244 to L substitution mimics a conformation changethat occurs naturally in Langerin as a result of interaction withparticular types of ligands.
In summary, our present findings identify mouse Langerin/CD207, which should be a useful molecule both as a marker andfor functional studies of mouse LC and related cells. In addition,we have demonstrated a plasticity of the types of organelles thatcan form in DC, and that may have important consequences for thefunction of these cells.
AcknowledgmentsWe thank Dr. T. McClanahan for cDNA libraries; Drs. A. Vicari and C.Caux for helpful discussion and critical review of the manuscript; Dr. G.Trinchieri for support; I. Durand and E. Garcia for flow cytometry analysis;M. Vatan for editorial assistance; and O. Clear and B. Michat for technicalsupport.
References1. Bell, D., J. W. Young, and J. Banchereau. 1999. Dendritic cells. Adv. Immunol.
72:255.2. Stingl, G. 1993. The skin: initiation and target site of immune responses. Recent
Res. Cancer Res. 128:45.3. Macatonia, S. E., S. C. Knight, A. J. Edwards, S. Griffiths, and P. Fryer. 1987.
Localization of antigen on lymph node dendritic cells after exposure to the con-tact sensitizer fluorescein isothiocyanate. J. Exp. Med. 166:1654.
4. Larsen, C. P., R. M. Steinman, M. Witmer-Pack, D. F. Hankins, P. J. Morris, andJ. M. Austyn. 1990. Migration and maturation of Langerhans cells in skin trans-plants and explants. J. Exp. Med. 172:1483.
5. Birbeck, M. S., A. S. Breathnach, and J. D. Everall. 1961. An electron micro-scope study of basal melanocytes and high-level clear cells (Langerhans cells) invitiligo. J. Invest. Dermatol. 37:51.
6. Wolff, K. 1967. The fine structure of the Langerhans cell granule. J. Cell Biol.35:468.
7. Valladeau, J., O. Ravel, C. Dezutter-Dambuyant, K. Moore, M. Kleijmeer,Y. Liu, V. Duvert-Frances, C. Vincent, D. Schmitt, J. Davoust, et al. 2000. Lan-gerin, a novel C-type lectin specific to Langerhans cells, is an endocytic receptorthat induces the formation of Birbeck granules. Immunity 12:71.
8. Ardavin, C., L. Wu, C. L. Li, and K. Shortman. 1993. Thymic dendritic cells andT cells develop simultaneously in the thymus from a common precursor popu-lation. Nature 362:761.
9. Anjuere, F., G. M. del Hoyo, P. Martin, and C. Ardavin. 2000. Langerhans cellsdevelop from a lymphoid-committed precursor. Blood 96:1633.
10. Anjuere, F., G. Martinez del Hoyo, P. Martin, and C. Ardavin. 2000. Langerhanscells acquire a CD8� dendritic cell phenotype on maturation by CD40 ligation.J. Leukocyte Biol. 67:206.
11. Tyndall, C., G. La Mantia, C. M. Thacker, J. Favaloro, and R. Kamen. 1981. Aregion of the polyoma virus genome between the replication origin and late pro-tein coding sequences is required in cis for both early gene expression and viralDNA replication. Nucleic Acids Res. 9:6231.
12. Halfon, S., J. Ford, J. Foster, L. Dowling, L. Lucian, M. Sterling, Y. Xu,M. Weiss, M. Ikeda, D. Liggett, et al. 1998. Leukocystatin, a new class II cystatinexpressed selectively by hematopoietic cells. J. Biol. Chem. 273:16400.
13. Bates, E. E., A. Kissenpfennig, C. Peronne, M. G. Mattei, F. Fossiez,B. Malissen, and S. Lebecque. 2000. The mouse and human IGSF6 (DORA)genes map to the inflammatory bowel disease 1 locus and are embedded in anintron of a gene of unknown function. Immunogenetics 52:112.
14. Mattei, M. G., J. Matterson, J. W. Chen, M. A. Williams, and M. Fukuda. 1990.Two human lysosomal membrane glycoproteins, h-lamp-1 and h-lamp-2, are en-coded by genes localized to chromosome 13q34 and chromosome Xq24–25,respectively. J. Biol. Chem. 265:7548.
15. Juhlin, L., and W. B. Shelley. 1977. New staining techniques for the Langerhanscell. Acta Derm. Venereol. 57:289.
16. Weinlich, G., M. Heine, H. Stossel, M. Zanella, P. Stoitzner, U. Ortner, J. Smolle,F. Koch, N. T. Sepp, G. Schuler, and N. Romani. 1998. Entry into afferent lym-phatics and maturation in situ of migrating murine cutaneous dendritic cells.J. Invest. Dermatol. 110:441.
17. Inaba, K., M. Inaba, N. Romani, H. Aya, M. Deguchi, S. Ikehara, S. Muramatsu,and R. M. Steinman. 1992. Generation of large numbers of dendritic cells frommouse bone marrow cultures supplemented with granulocyte/macrophage colo-ny-stimulating factor. J. Exp. Med. 176:1693.
18. Valladeau, J., V. Duvert-Frances, J. J. Pin, C. Dezutter-Dambuyant, C. Vincent,C. Massacrier, J. Vincent, K. Yoneda, J. Banchereau, C. Caux, et al. 1999. Themonoclonal antibody DCGM4 recognizes Langerin, a protein specific of Lang-erhans cells, and is rapidly internalized from the cell surface. Eur. J. Immunol.29:2695.
19. Weis, W. I., M. E. Taylor, and K. Drickamer. 1998. The C-type lectin super-family in the immune system. Immunol. Rev. 163:19.
20. Ren, R., B. J. Mayer, P. Cicchetti, and D. Baltimore. 1993. Identification of aten-amino acid proline-rich SH3 binding site. Science 259:1157.
21. Cohen, G. B., R. Ren, and D. Baltimore. 1995. Modular binding domains insignal transduction proteins. Cell 80:237.
22. Hoyle, G. W., and R. L. Hill. 1988. Molecular cloning and sequencing of a cDNAfor a carbohydrate binding receptor unique to rat Kupffer cells. J. Biol. Chem.263:7487.
23. Bezouska, K., G. V. Crichlow, J. M. Rose, M. E. Taylor, and K. Drickamer. 1991.Evolutionary conservation of intron position in a subfamily of genes encodingcarbohydrate-recognition domains. J. Biol. Chem. 266:11604.
24. Suzuki, N., K. Yamamoto, S. Toyoshima, T. Osawa, and T. Irimura. 1996. Mo-lecular cloning and expression of cDNA encoding human macrophage C-typelectin: its unique carbohydrate binding specificity for Tn antigen. J. Immunol.156:128.
25. Spiess, M., and H. F. Lodish. 1985. Sequence of a second human asialoglyco-protein receptor: conservation of two receptor genes during evolution. Proc. Natl.Acad. Sci. USA 82:6465.
26. Spiess, M., A. L. Schwartz, and H. F. Lodish. 1985. Sequence of human asialo-glycoprotein receptor cDNA: an internal signal sequence for membrane insertion.J. Biol. Chem. 260:1979.
27. Burge, C. B., R. A. Padgett, and P. A. Sharp. 1998. Evolutionary fates and originsof U12-type introns. Mol. Cell 2:773.
28. Hoyle, G. W., and R. L. Hill. 1991. Structure of the gene for a carbohydrate-binding receptor unique to rat Kupffer cells. J. Biol. Chem. 266:1850.
29. Borkowski, T. A., J. J. Letterio, A. G. Farr, and M. C. Udey. 1996. A role forendogenous transforming growth factor �1 in Langerhans cell biology: the skinof transforming growth factor �1 null mice is devoid of epidermal Langerhanscells. J. Exp. Med. 184:2417.
30. Schuler, G., and R. M. Steinman. 1985. Murine epidermal Langerhans cellsmature into potent immunostimulatory dendritic cells in vitro. J. Exp. Med.161:526.
31. Kobayashi, M., and T. Hoshino. 1983. Distribution of cored tubule-containingLangerhans cells in the skin and lymph nodes of mice. J. Electron Microsc.(Tokyo) 32:197.
32. Naito, M., G. Hasegawa, and K. Takahashi. 1997. Development, differentiation,and maturation of Kupffer cells. Microsc. Res. Tech. 39:350.
33. Toth, C. A., and P. Thomas. 1992. Liver endocytosis and Kupffer cells. Hepa-tology 16:255.
34. Drickamer, K. 1999. C-type lectin-like domains. Curr. Opin. Struct. Biol. 9:585.35. Leung, J. O., E. C. Holland, and K. Drickamer. 1985. Characterization of the gene
encoding the major rat liver asialoglycoprotein receptor. J. Biol. Chem. 260:12523.
36. Suter, U., R. Bastos, and H. Hofstetter. 1987. Molecular structure of the gene andthe 5�-flanking region of the human lymphocyte immunoglobulin E receptor.Nucleic Acids Res. 15:7295.
37. Soilleux, E. J., R. Barten, and J. Trowsdale. 2000. DC-SIGN; a related gene,DC-SIGNR; and CD23 form a cluster on 19p13. J. Immunol. 165:2937.
38. Beavil, A. J., R. L. Edmeades, H. J. Gould, and B. J. Sutton. 1992. �-Helicalcoiled-coil stalks in the low-affinity receptor for IgE (Fc�RII/CD23) and relatedC-type lectins. Proc. Natl. Acad. Sci. USA 89:753.
39. Dierks, S. E., W. C. Bartlett, R. L. Edmeades, H. J. Gould, M. Rao, andD. H. Conrad. 1993. The oligomeric nature of the murine Fc�RII/CD23: impli-cations for function. J. Immunol. 150:2372.
40. Beavil, R. L., P. Graber, N. Aubonney, J. Y. Bonnefoy, and H. J. Gould. 1995.CD23/Fc�RII and its soluble fragments can form oligomers on the cell surfaceand in solution. Immunology 84:202.
41. Bider, M. D., J. M. Wahlberg, R. A. Kammerer, and M. Spiess. 1996. The oli-gomerization domain of the asialoglycoprotein receptor preferentially forms 2:2heterotetramers in vitro. J. Biol. Chem. 271:31996.
42. Sanford, J. P., R. L. Eddy, D. Doyle, and T. B. Shows. 1991. Assignment ofhuman asialoglycoprotein receptor gene (ASGR1) to chromosome 17p11-13.Genomics 11:779.
43. Lanier, L. L. 1998. NK cell receptors. Annu. Rev. Immunol. 16:359.44. Mahnke, K., M. Guo, S. Lee, H. Sepulveda, S. L. Swain, M. Nussenzweig,
and R. M. Steinman. 2000. The dendritic cell receptor for endocytosis, DEC-205, can recycle and enhance antigen presentation via major histocompati-bility complex class II-positive lysosomal compartments. J. Cell Biol. 151:673.
45. Jiang, W., W. J. Swiggard, C. Heufler, M. Peng, A. Mirza, R. M. Steinman,and M. C. Nussenzweig. 1995. The receptor DEC-205 expressed by dendriticcells and thymic epithelial cells is involved in antigen processing. Nature375:151.
46. Geijtenbeek, T. B., D. S. Kwon, R. Torensma, S. J. van Vliet,G. C. van Duijnhoven, J. Middel, I. L. Cornelissen, H. S. Nottet,V. N. KewalRamani, D. R. Littman, et al. 2000. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell100:587.
47. Geijtenbeek, T. B., D. J. Krooshoop, D. A. Bleijs, S. J. van Vliet,G. C. van Duijnhoven, V. Grabovsky, R. Alon, C. G. Figdor, and Y. van Kooyk.2000. DC-SIGN-ICAM-2 interaction mediates dendritic cell trafficking. Nat. Im-munol. 1:353.
48. Geijtenbeek, T. B., R. Torensma, S. J. van Vliet, G. C. van Duijnhoven,G. J. Adema, Y. van Kooyk, and C. G. Figdor. 2000. Identification of DC-SIGN,a novel dendritic cell-specific ICAM-3 receptor that supports primary immuneresponses. Cell 100:575.
49. Bates, E. M., N. Fournier, E. Garcia, J. Valladeau, I. Durand, J. J. Pin, S. M.Zurawski, S. Patel, J. S. Abrams, S. Lebecque, et al. 1999. Antigen-presenting
791The Journal of Immunology

cells express DCIR, a novel C-type lectin surface receptor containing animmunoreceptor tyrosine-based inhibitory motif. J. Immunol. 163:1973.
50. Ariizumi, K., G. L. Shen, S. Shikano, S. Xu, R. Ritter, T. Kumamoto,D. Edelbaum, A. Morita, P. R. Bergstresser, and A. Takashima. 2000. Identifi-cation of a novel, dendritic cell-associated molecule, dectin-1, by subtractivecDNA cloning. J. Biol. Chem. 275:20157.
51. Henri, S., D. Vremec, A. Kamath, J. Waithman, S. Williams, C. Benoist,K. Burnham, S. Saeland, E. Handman, and K. Shortman. 2001. The dendritic cellpopulations of mouse lymph nodes. J. Immunol. 167:741.
52. Vremec, D., J. Pooley, H. Hochrein, L. Wu, and K. Shortman. 2000. CD4 andCD8 expression by dendritic cell subtypes in mouse thymus and spleen. J. Im-munol. 164:2978.
53. Traver, D., K. Akashi, M. Manz, M. Merad, T. Miyamoto, E. G. Engleman, andI. L. Weissman. 2000. Development of CD8�-positive dendritic cells from acommon myeloid progenitor. Science 290:2152.
54. Ardavin, C. 1997. Thymic dendritic cells. Immunol. Today 18:350.
55. Hoshino, T., A. Kukita, and S. Sato. 1970. Cells containing Birbeck granules(Langerhans cell granules) in the human thymus. J. Electron Microsc. (Tokyo)19:271.
56. Zhang, X., L. Giangreco, H. E. Broome, C. M. Dargan, and S. L. Swain. 1995.Control of CD4 effector fate: transforming growth factor �1 and interleukin 2synergize to prevent apoptosis and promote effector expansion. J. Exp. Med.182:699.
57. Kobayashi, M., and T. Hoshino. 1978. Occurrence of “cored tubule” in the Bir-beck granule-containing cells of mice. J. Electron Microsc. (Tokyo) 27:199.
58. Kobayashi, M., and T. Hoshino. 1987. The three-dimensional structure of coredtubules in the dermal Langerhans cells of mice. J. Electron Microsc. (Tokyo)36:411.
59. Drickamer, K. 1993. Evolution of Ca2�-dependent animal lectins. Prog. NucleicAcid Res. Mol. Biol. 45:207.
60. Meier, M., M. D. Bider, V. N. Malashkevich, M. Spiess, and P. Burkhard. 2000.Crystal structure of the carbohydrate recognition domain of the H1 subunit of theasialoglycoprotein receptor. J. Mol. Biol. 300:857.
792 IDENTIFICATION OF mLANGERIN