identification of root-knot nematodes (meloidogyne spp
TRANSCRIPT

675
http://journals.tubitak.gov.tr/zoology/
Turkish Journal of Zoology Turk J Zool(2016) 40: 675-685© TÜBİTAKdoi:10.3906/zoo-1508-19
Identification of root-knot nematodes (Meloidogyne spp.) fromgreenhouses in the Middle Black Sea Region of Turkey
Gökhan AYDINLI1,*, Sevilhan MENNAN2
1Bafra Vocational High School, Ondokuz Mayıs University, Samsun, Turkey2Department of Plant Protection, Faculty of Agriculture, Ondokuz Mayıs University, Samsun, Turkey
* Correspondence: [email protected]
1. IntroductionGreenhouses, as intensive vegetable productive systems, have favorable environmental conditions for both plants and pathogens, including nematodes. The most important nematode threat causing severe losses in vegetable crop production in greenhouses is the root-knot nematode. Root-knot nematodes belong to the genus Meloidogyne Göldi 1887 and are obligate plant parasites that feed on roots and are able to parasitize almost every species of vascular plants (Jones et al., 2013). Nematode invasion into roots results in altered root architecture. The most obvious morphological change is the characteristic root galling, which affects the uptake of water and nutrients. As a result of the damage to the root system, typical aboveground symptoms of nematode infections include stunting, yellowing, and wilting, and yield reductions occur as a result. Yield losses depend on the nematode species, population level, and crop species and may range from 10% (Collange et al., 2011) to 50%–80% (Stirling, 1991; Siddiqi, 2000). In a study of several particular crops, Sikora and Fernandez (2005) reported that specific losses ranged from 17% to 20% for eggplant, 18% to 33% for melon, and 24% to 38% for tomato.
The success of nematode management tactics depends upon problem recognition, specifically the proper knowledge of nematode species. Accurate diagnosis of the species is essential for the determination of the most appropriate control strategies to be employed (Coyne et al., 2009). Perineal pattern morphology of adult females is most frequently used for the identification of root-knot nematodes (Jepson, 1987; Hunt and Handoo, 2009). This method is often inconclusive for distinguishing closely related Meloidogyne species, because individuals within a population often vary considerably (Zijlstra et al., 2000). The esterase isozyme pattern of females is another tool to identify species and has been considered the most reliable and useful method in nematology (Esbenshade and Triantaphyllou, 1985). Recently, a PCR method based on DNA has also been widely used for the identification of nematodes (Zijlstra et al., 2004; Powers et al., 2005; Adam et al., 2007; Devran and Söğüt, 2009). Species-specific primers were developed and are used routinely on a large number of samples with high sensitivity and specificity. This method is simple and fast, and it can be used by personnel with no nematological expertise (Zijlstra et al., 2004).
Abstract: Root-knot nematodes are among the main important pests causing serious yield losses in greenhouses worldwide, including Turkey. Accurate identification of these pests is essential for determining appropriate control strategies. Identification of Meloidogyne species based on perineal pattern morphology alone is not viable or reliable due to overlaps and high variability among the species. Therefore, our objective was to overcome the difficulties in identifying 90 populations of root-knot nematodes collected from different greenhouses in the Middle Black Sea Region (Samsun, Tokat, Amasya, Çorum, Ordu, and Sinop provinces), using not only morphological (perineal pattern morphology) but also biochemical (esterase phenotype) and molecular (PCR with species-specific primers) techniques. The results indicated that M. arenaria, M. ethiopica, M. javanica, and M. incognita were identified at 42.2%, 41.1%, 12.2%, and 4.4% of the total populations, respectively. Meloidogyne ethiopica was detected for the first time in Turkey on eggplant (Solanum melongena L.) and nightshade (Solanum nigrum L.), and was only found in greenhouses in the coastal provinces (Samsun, Sinop, Ordu) of the Middle Black Sea Region. Our study confirmed that the E3 esterase phenotype is a useful character for distinguishing M. ethiopica from other Meloidogyne species.
Key words: Greenhouse, identification, Meloidogyne spp., Middle Black Sea Region, Turkey
Received: 10.08.2015 Accepted/Published Online: 11.04.2016 Final Version: 24.10.2016
Research Article

676
AYDINLI and MENNAN / Turk J Zool
To date, over 90 named globally distributed root-knot nematode species (Meloidogyne spp.) have been reported (Hunt and Handoo, 2009). In Turkey, eight species (Meloidogyne arenaria, M. artiellia, M. chitwoodi, M. ethiopica, M. exigua, M. hapla, M. incognita, M. javanica) were recorded in different agricultural areas (Yüksel, 1974; Di Vito et al., 1994; Elekçioğlu et al., 1994; Mennan and Ecevit, 1996; Kaşkavalcı and Öncüer, 1999; Söğüt and Elekçioğlu, 2000; Devran et al., 2009; Devran and Söğüt, 2009; Özarslandan et al., 2009; Özarslandan and Elekçioğlu, 2010; Akyazı and Ecevit, 2011; Aydınlı et al., 2013; Kepenekçi et al., 2014). Among them, Meloidogyne incognita, M. arenaria, and M. javanica are regarded as the most common species reported in vegetable-growing areas in Turkey (Elekçioğlu et al., 1994; Mennan and Ecevit, 1996; Kaşkavalcı and Öncüer, 1999; Söğüt and Elekçioğlu, 2000; Devran and Söğüt, 2009).
The Black Sea Region is the third largest regional greenhouse-producing area in Turkey. Most greenhouses are in the Middle Black Sea Region. Prior to this study, only the greenhouses of Samsun and Tokat provinces had been surveyed for root-knot nematodes, and the authors reported that M. incognita was a common species according to perineal pattern morphology and differential host tests (Katı and Mennan, 2006; Akyazı and Ecevit, 2011). During a local survey in the summer of 2009, M. ethiopica was reported for the first time in Turkey on tomato and cucumber in greenhouses located in Samsun (Aydınlı et al., 2013). The perineal pattern morphology of this species was similar to that of M. incognita. Due to variations in perineal pattern morphology, a precise identification based only on this feature is difficult. Thus, M. ethiopica may have already been present in Turkey for many years because of the misinterpretation in identification.
In this study, our objective was to overcome the difficulty of identifying root-knot nematode populations from vegetable-growing greenhouses in the Middle Black Sea Region of Turkey by using morphological (perineal pattern morphology), biochemical (esterase phenotype), and molecular (PCR with species-specific primers) techniques.
2. Materials and methods2.1. Nematode populationsSurveys of Meloidogyne were carried out in 2011 and 2012 in the Middle Black Sea Region of Turkey. Ninety populations of root-knot nematodes were collected randomly from different vegetable-growing greenhouse locations in six provinces (Table 1). Some populations were collected from roots of infested weeds near vegetable plants in the same greenhouse, because certain greenhouses possessed resistant tomato plants carrying the Mi gene or rootstocks.
Egg masses were handpicked individually from infested roots in each greenhouse. Each population was established from a single egg mass and reared on susceptible tomato plants (Solanum lycopersicum L.) of cultivar Falcon (May Seed, Turkey). Tomato seedlings at the fourth true leaf stage were transplanted singly into 300-mL plastic pots containing sterilized sandy loam soil and sand (2:1) and were allowed to grow for 5–7 days before being inoculated with single egg masses. All populations obtained from single egg masses were maintained continuously in pot cultures in greenhouses at 24 ± 2 °C and were renewed by transferring 10 egg masses from each plant to new tomato seedlings bimonthly. 2.2. Perineal pattern studiesFemale nematodes obtained from the first individuals of single egg mass cultures of each population were used for perineal pattern studies to minimize genetic variation. Females were dissected from infected tomato roots and fixed in triethanolamine-formaldehyde (TAF) solution until observation. Ten individual females from each population were selected randomly and perineal patterns were cut in 45% lactic acid and mounted in glycerin (Taylor and Netscher, 1974; Hartman and Sasser, 1985). Perineal pattern studies were conducted under a light microscope according to Eisenback et al. (1981) and Jepson (1987).
2.3. Esterase phenotype studiesYoung egg-laying females of each pure population were handpicked from roots under a dissection microscope and collected in 0.9% NaCl (sodium chloride). A single female or three females for each microhematocrit tube with 5 µL of extraction buffer (20% sucrose, 1% Triton X-100) were transferred and macerated with a pestle. The specimens were frozen immediately and stored at –20 °C (Conceição et al., 2012). Electrophoresis using a Mini-Protean Tetra System (Bio-Rad Laboratories, Hercules, CA, USA) was performed at 6 mA/gel during the first 15 min and then at 20 mA/gel for about 45 min. The gels were stained for esterase activity with the substrate α-naphthyl acetate. Protein extracts were obtained from three young egg-laying females of M. javanica (J3) in each gel as a reference. In some gels, protein extracts of M. arenaria (A2) or M. ethiopica (E3) were used as a reference, in addition to M. javanica. Esterase phenotypes were designated with a letter suggesting the nematode species and a number indicating the number of bands, as described by Esbenshade and Triantaphyllou (1985).2.4. Molecular studiesDNA was extracted from the egg masses of each population using the DNeasy Tissue and Blood Kit (QIAGEN, Hilden, Germany) according to the manufacturer’s instructions. DNA samples were stored at –20 °C until used. PCR amplifications were conducted with the species-specific
677
AYDINLI and MENNAN / Turk J Zool
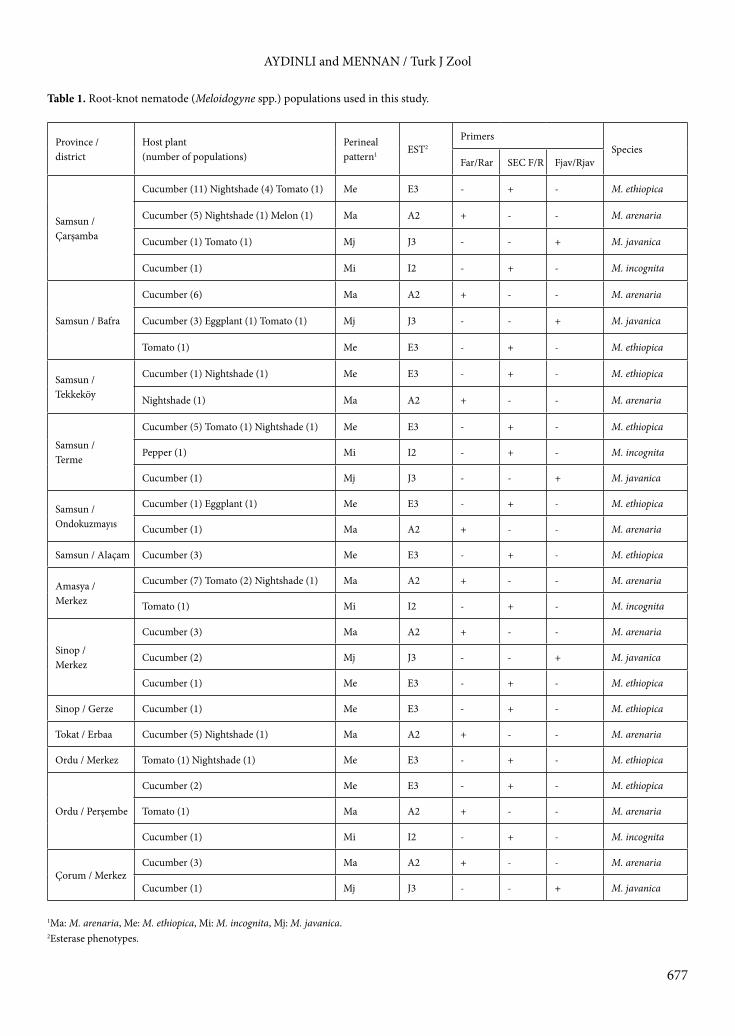
Table 1. Root-knot nematode (Meloidogyne spp.) populations used in this study.
Province /district
Host plant(number of populations)
Perineal pattern1 EST2
PrimersSpecies
Far/Rar SEC F/R Fjav/Rjav
Samsun / Çarşamba
Cucumber (11) Nightshade (4) Tomato (1) Me E3 - + - M. ethiopica
Cucumber (5) Nightshade (1) Melon (1) Ma A2 + - - M. arenaria
Cucumber (1) Tomato (1) Mj J3 - - + M. javanica
Cucumber (1) Mi I2 - + - M. incognita
Samsun / Bafra
Cucumber (6) Ma A2 + - - M. arenaria
Cucumber (3) Eggplant (1) Tomato (1) Mj J3 - - + M. javanica
Tomato (1) Me E3 - + - M. ethiopica
Samsun / Tekkeköy
Cucumber (1) Nightshade (1) Me E3 - + - M. ethiopica
Nightshade (1) Ma A2 + - - M. arenaria
Samsun / Terme
Cucumber (5) Tomato (1) Nightshade (1) Me E3 - + - M. ethiopica
Pepper (1) Mi I2 - + - M. incognita
Cucumber (1) Mj J3 - - + M. javanica
Samsun / Ondokuzmayıs
Cucumber (1) Eggplant (1) Me E3 - + - M. ethiopica
Cucumber (1) Ma A2 + - - M. arenaria
Samsun / Alaçam Cucumber (3) Me E3 - + - M. ethiopica
Amasya / Merkez
Cucumber (7) Tomato (2) Nightshade (1) Ma A2 + - - M. arenaria
Tomato (1) Mi I2 - + - M. incognita
Sinop / Merkez
Cucumber (3) Ma A2 + - - M. arenaria
Cucumber (2) Mj J3 - - + M. javanica
Cucumber (1) Me E3 - + - M. ethiopica
Sinop / Gerze Cucumber (1) Me E3 - + - M. ethiopica
Tokat / Erbaa Cucumber (5) Nightshade (1) Ma A2 + - - M. arenaria
Ordu / Merkez Tomato (1) Nightshade (1) Me E3 - + - M. ethiopica
Ordu / Perşembe
Cucumber (2) Me E3 - + - M. ethiopica
Tomato (1) Ma A2 + - - M. arenaria
Cucumber (1) Mi I2 - + - M. incognita
Çorum / MerkezCucumber (3) Ma A2 + - - M. arenaria
Cucumber (1) Mj J3 - - + M. javanica
1Ma: M. arenaria, Me: M. ethiopica, Mi: M. incognita, Mj: M. javanica. 2Esterase phenotypes.
678
AYDINLI and MENNAN / Turk J Zool
primers listed in Table 2. All amplification reactions were performed in a final volume of 25 µL containing 5 µL of DNA (20 ng), 2.5 µL of 10X Taq buffer with KCl (1X Taq buffer with KCl), 2 µL of 25 mM MgCl2 (2 mM MgCl2), 0.5 µL of 10 mM dNTPs (0.2 mM dNTPs), 1 µL of 10 µM of each primer (0.4 µM of each primer), and 0.25 µL of 5 U/µL Taq DNA Polymerase (1.25 U of Taq DNA Polymerase) (Thermo Scientific, Waltham, MA, USA). PCR cycling conditions using a MJ Mini Personal Thermal Cycler (Bio-Rad Laboratories) comprised an initial denaturation step at 94 °C for 3 min, followed by 35 cycles of 30 s at 94 °C, 30 s at annealing temperature, and 1 min at 72 °C, with a final extension for 7 min at 72 °C. Annealing temperatures were 56 °C for primers SEC-1F/1R, 61 °C for primers Far/Rar, and 64 °C for primers Fjav/Rjav. PCR products were analyzed by electrophoresis in 2% agarose gel stained with ethidium bromide and visualized by UV illumination using a Gel Doc XR (Bio-Rad Laboratories).
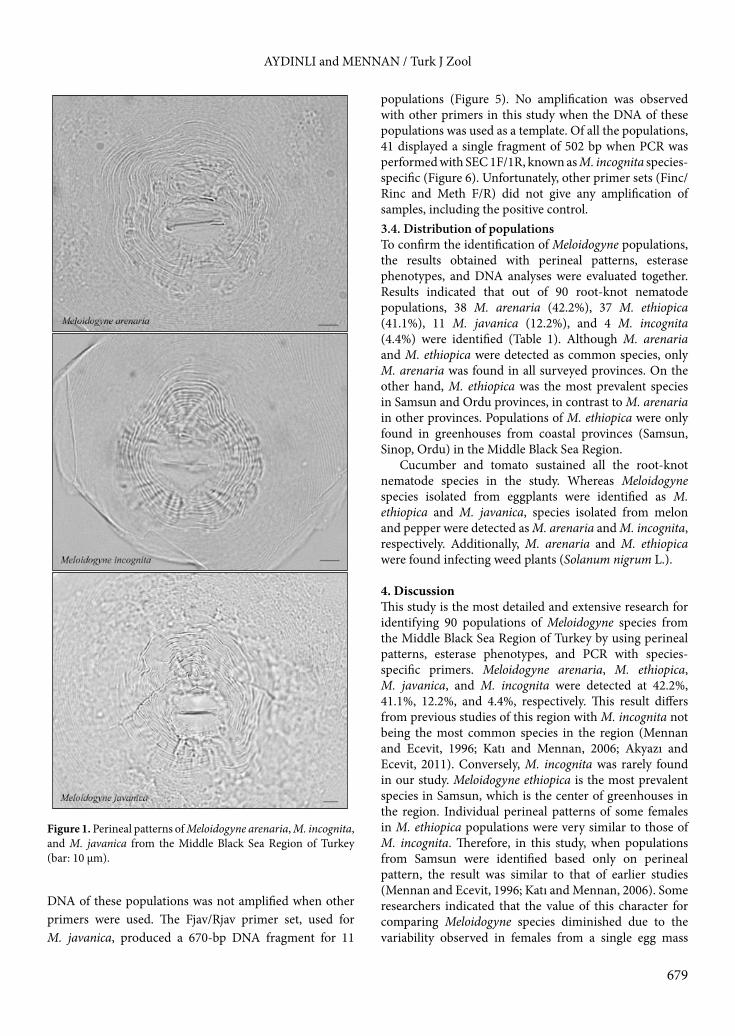
3. Results3.1. Perineal patterns of populationsPerineal pattern variability was observed within and between populations isolated from the Middle Black Sea Region (Figures 1 and 2). Therefore, as a first step, the perineal patterns of females from each Meloidogyne population were grouped into common root-knot species M. arenaria, M. incognita, and M. javanica according to similar morphology by a main diagnostic feature for species identification. Only M. arenaria perineal patterns were detected in 38 of the populations (Table 1). These patterns showed a generally low dorsal arch and forming shoulders. Lateral lines were distinct, dorsal and ventral striae connected with an angle and forked. Eleven populations were perineal patterns of only M. javanica, which were
easy to separate from those of other populations with the presence of distinct lateral lines. The overall shape of these patterns was ovoid or rounded and the dorsal arch was generally low. Only four populations showed the typical M. incognita pattern, which is characterized by the presence of a high and squarish dorsal arch and indistinct lateral lines. Thirty-seven populations showed patterns that combined features of M. arenaria and M. incognita. These perineal patterns showed a rounded to squarish arch and indistinct forming shoulders. Lateral lines were mostly indistinct. The patterns were similar to M. ethiopica, with some variations, and overlapped with those of M. incognita and M. arenaria (Figure 2).3.2. Esterase phenotypes of the populationsTen bands for esterase activity and four distinct esterase phenotypes were detected among 90 populations of Meloidogyne from different provinces (Figure 3). Meloidogyne javanica was used as a reference for each gel, and 11 populations exhibited the species-specific phenotype J3 (Rm: 35, 41, 45). Phenotypes A2 (Rm: 40, 44) and E3 (Rm: 35, 39, 42), as the most prevalent esterase phenotypes in this study, were detected in 38 and 37 populations, respectively (Table 1). On the other hand, phenotype I2 (Rm: 35, 37) was rarely shown with 4 populations. A2, E3, and I2 are species-specific for M. arenaria, M. ethiopica, and M. incognita, respectively. 3.3. DNA analysis of populationsSpecies-specific primers were used for molecular characterization of populations. Three (SEC 1F/1R; Far/Rar; Fjav/Rjav) pair primers tested on DNA isolated from egg masses resulted in consistent amplifications. PCR with M. arenaria species-specific primers Far/Rar produced a single band of 420 bp for 38 populations (Figure 4). The
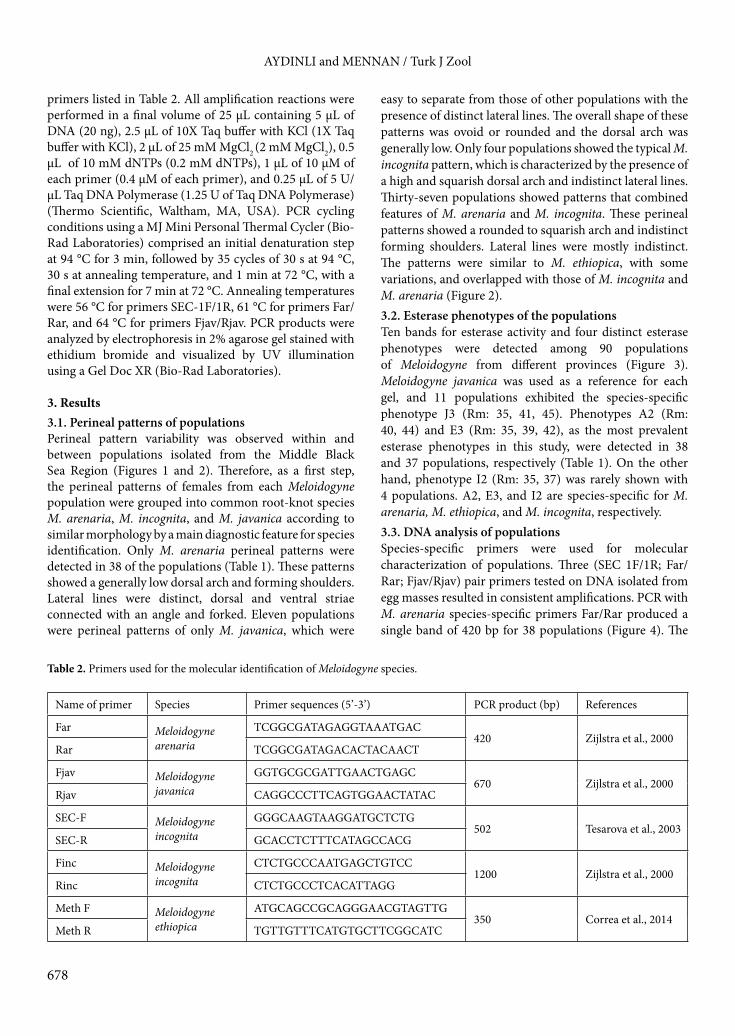
Table 2. Primers used for the molecular identification of Meloidogyne species.
Name of primer Species Primer sequences (5’-3’) PCR product (bp) References
Far Meloidogyne arenaria
TCGGCGATAGAGGTAAATGAC420 Zijlstra et al., 2000
Rar TCGGCGATAGACACTACAACT
Fjav Meloidogyne javanica
GGTGCGCGATTGAACTGAGC670 Zijlstra et al., 2000
Rjav CAGGCCCTTCAGTGGAACTATAC
SEC-F Meloidogyne incognita
GGGCAAGTAAGGATGCTCTG502 Tesarova et al., 2003
SEC-R GCACCTCTTTCATAGCCACG
Finc Meloidogyne incognita
CTCTGCCCAATGAGCTGTCC1200 Zijlstra et al., 2000
Rinc CTCTGCCCTCACATTAGG
Meth F Meloidogyne ethiopica
ATGCAGCCGCAGGGAACGTAGTTG350 Correa et al., 2014
Meth R TGTTGTTTCATGTGCTTCGGCATC
679
AYDINLI and MENNAN / Turk J Zool
DNA of these populations was not amplified when other primers were used. The Fjav/Rjav primer set, used for M. javanica, produced a 670-bp DNA fragment for 11
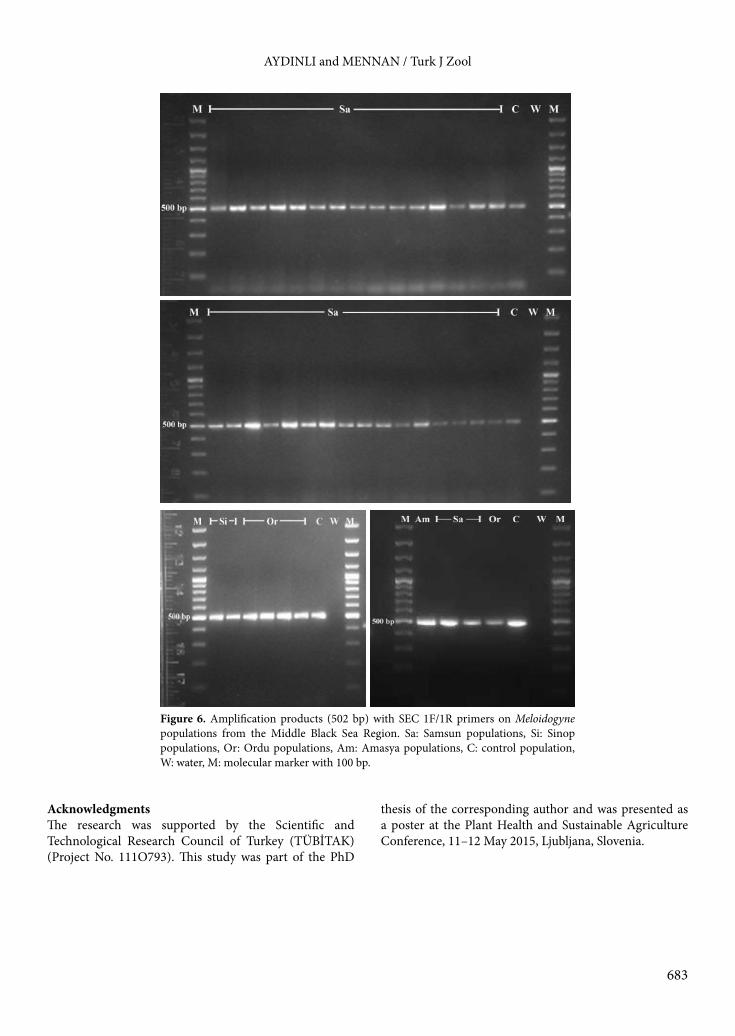
populations (Figure 5). No amplification was observed with other primers in this study when the DNA of these populations was used as a template. Of all the populations, 41 displayed a single fragment of 502 bp when PCR was performed with SEC 1F/1R, known as M. incognita species-specific (Figure 6). Unfortunately, other primer sets (Finc/Rinc and Meth F/R) did not give any amplification of samples, including the positive control.3.4. Distribution of populationsTo confirm the identification of Meloidogyne populations, the results obtained with perineal patterns, esterase phenotypes, and DNA analyses were evaluated together. Results indicated that out of 90 root-knot nematode populations, 38 M. arenaria (42.2%), 37 M. ethiopica (41.1%), 11 M. javanica (12.2%), and 4 M. incognita (4.4%) were identified (Table 1). Although M. arenaria and M. ethiopica were detected as common species, only M. arenaria was found in all surveyed provinces. On the other hand, M. ethiopica was the most prevalent species in Samsun and Ordu provinces, in contrast to M. arenaria in other provinces. Populations of M. ethiopica were only found in greenhouses from coastal provinces (Samsun, Sinop, Ordu) in the Middle Black Sea Region.
Cucumber and tomato sustained all the root-knot nematode species in the study. Whereas Meloidogyne species isolated from eggplants were identified as M. ethiopica and M. javanica, species isolated from melon and pepper were detected as M. arenaria and M. incognita, respectively. Additionally, M. arenaria and M. ethiopica were found infecting weed plants (Solanum nigrum L.).
4. DiscussionThis study is the most detailed and extensive research for identifying 90 populations of Meloidogyne species from the Middle Black Sea Region of Turkey by using perineal patterns, esterase phenotypes, and PCR with species-specific primers. Meloidogyne arenaria, M. ethiopica, M. javanica, and M. incognita were detected at 42.2%, 41.1%, 12.2%, and 4.4%, respectively. This result differs from previous studies of this region with M. incognita not being the most common species in the region (Mennan and Ecevit, 1996; Katı and Mennan, 2006; Akyazı and Ecevit, 2011). Conversely, M. incognita was rarely found in our study. Meloidogyne ethiopica is the most prevalent species in Samsun, which is the center of greenhouses in the region. Individual perineal patterns of some females in M. ethiopica populations were very similar to those of M. incognita. Therefore, in this study, when populations from Samsun were identified based only on perineal pattern, the result was similar to that of earlier studies (Mennan and Ecevit, 1996; Katı and Mennan, 2006). Some researchers indicated that the value of this character for comparing Meloidogyne species diminished due to the variability observed in females from a single egg mass
Figure 1. Perineal patterns of Meloidogyne arenaria, M. incognita, and M. javanica from the Middle Black Sea Region of Turkey (bar: 10 µm).

680
AYDINLI and MENNAN / Turk J Zool
population (Karssen and van Aelst, 2001; Carneiro et al., 2004b; Garcia and Sanchez-Puerta, 2012). However, it can be useful to separate populations into species groups as a first step towards identification (Munera et al., 2010). Consequently, the perineal patterns of populations in this study were grouped according to the most common root-knot species in both Turkey and the world. To examine the whole genus, Jepson (1987) separated the species into 6 distinct groups according to morphological similarity. Meloidogyne ethiopica was classified into Group 3, which is different from M. arenaria (Group 4) and M. incognita (Group 6). At the same time, Jepson (1987) reported that the characterization of Group 3 is very broad, and M. ethiopica was sometimes with a high, squared dorsal arch similar to the species in Group 6, including M. incognita. Moreover, before M. ethiopica was described as a new species, some individuals were identified as M. arenaria (Whitehead, 1968) due to their low, rounded arch (Jepson, 1987). Previous studies also indicated that the perineal pattern of M. ethiopica is highly variable and varies from similar to M. arenaria to similar to M. incognita (Carneiro et al., 2004a, 2007; Strajnar et al., 2009; Conceição et al., 2012). These results are in agreement with our previous study, which was the first report on M. ethiopica in Turkey (Aydınlı et al., 2013).
Meloidogyne populations in the study exhibited four esterase phenotypes associated with M. arenaria, M. ethiopica, M. javanica, and M. incognita. All the esterase phenotypes, except E3, were reported as the most common phenotypes from a different area of the world (Esbenshade and Triantaphyllou, 1985). Esterase phenotype E3 is M. ethiopica species-specific and is the most useful character for distinguishing it from other Meloidogyne species (Carneiro et al., 2004b, 2007). In a previous study, we found some populations with three esterase bands from cucumber and tomatoes in greenhouses in Samsun, and we considered that this could be an atypical phenotype of M. incognita according to the results of the perineal pattern and species-specific primer (Aydınlı and Mennan, 2011). However, when additional studies were performed,
it became evident that these populations were M. ethiopica. This was the first report of M. ethiopica in Turkey (Aydınlı et al., 2013). Previous studies from various geographical regions around the world have indicated the utility of this enzyme (Esbenshade and Triantaphyllou, 1985; Fargette, 1987; Pais and Abrantes, 1989; Carneiro et al., 2000; Brito et al., 2008). In Turkey, the use of esterase phenotypes for the identification of Meloidogyne species was published for the first time in 2011 (Aydınlı and Mennan, 2011; Çetintaş and Çakmak, 2011; Mennan et al., 2011). These previous studies were conducted on limited populations from a local area. Conversely, this is the first comprehensive study on the identification of the Meloidogyne species from Turkey with esterase analysis.
In this study, we used primers found as effective in previous studies related to common Meloidogyne species, and our results showed agreement with the findings of other authors (Ziljstra et al., 2000; Tzortzakakis et al., 2005; Adam et al., 2007; Devran and Söğüt, 2009; Oh et al., 2009; Özarslandan and Elekçioğlu, 2010; Mennan et al., 2011; Garcia and Sanchez-Puerta, 2012). Two different primer sets (SEC 1F/1R and Finc/Rinc) for M. incognita were used in our studies. The Finc/Rinc primer set did not give reproducible amplifications for the populations. Similar results with this primer set were obtained by Adam et al. (2007) and Devran and Söğüt (2009). SEC 1R/1F gave the expected 502-bp fragment in 4 populations that have M. incognita species-specific esterase phenotypes (I2). However, this primer set also gave positive results for 37 populations with E3 esterase phenotypes. Therefore, the SEC 1R/1F primer set is not specific for M. incognita due to the amplification product in the DNA of M. ethiopica. The Far/Rar primer set successfully gave PCR products in 38 populations with the A2 esterase phenotype. Interestingly, when this primer set was used at 56 °C annealing temperature, as indicated by Devran and Söğüt (2009), some populations with E3 gave the amplified product, but these results were not reproducible. When the primer set was used at 64 °C annealing temperature as suggested by Ziljstra et al. (2000), we obtained the
Figure 2. Perineal pattern variability observed in Meloidogyne ethiopica from a single egg mass population (bar: 10 µm).

681
AYDINLI and MENNAN / Turk J Zool
Figure 3. Esterase phenotypes observed in the Meloidogyne populations of the Middle Black Sea Region of Turkey (*: reference population).
1 7 10
6 9
2 5 8
3 4
20
30
40
50
60
Rm
J3 I2 E3 A2
Band No
Rm (x100)
1 35,1
2 35,2
3 35,4
4 37,4
5 38,7
6 40,1
7 41,4
8 42,2
9 43,7
10 45,2
,
682
AYDINLI and MENNAN / Turk J Zool
expected band only in populations with A2. The annealing temperature for species-specific primers should be high enough to enhance specificity (Blok and Powers, 2009). In this study, the Meth F/R primer set developed by Correa et al. (2014) was also tested on all populations but did not give an amplification product, not even in the DNA of the reference populations. In addition, the described SCAR methods for these primer sets were tested using European M. ethiopica populations by Slovenian nematology groups (Sirca and Geric Stare, personal communication), but the method failed to obtain positive results. Amplification could have failed due to changes in the priming sites (Adam et al., 2007; Devran and Söğüt, 2009), because all populations, except one (from Kenya) used by Correa et al. (2014), were from Brazil and Chile, not from a wide geographical origin. Identification of M. ethiopica in this study was confirmed by Geric Stare (Agricultural Institute of Slovenia) using the mtDNA sequence (data not shown).
M. ethiopica is a new record for eggplant (Solanum
melongena L.) and nightshade (S. nigrum) in Turkey. Some populations of M. ethiopica were found infecting weed (S. nigrum) in greenhouses with resistant tomato plants carrying the Mi gene. This result showed that the Mi gene was also resistant to M. ethiopica and thus agreed with the previous data (Strajnar and Sirca, 2011; Conceição et al., 2012).
In conclusion, it is difficult to distinguish all Meloidogyne species by perineal patterns alone due to the considerable variation between and within populations. Conversely, perineal patterns are valuable features to provide support or confirmation for results obtained by biochemical and molecular methods (Carneiro et al., 2004b; Hernandez et al., 2004). Esterase enzyme phenotypes are species-specific and can be used as a reliable tool to identify Meloidogyne in the Middle Black Sea Region. Since perineal patterns of M. ethiopica are close to those of M. incognita, correct identification based only on perineal pattern is difficult, and there might be misidentification (Aydınlı et al., 2013). Carneiro et al. (1996) reported that using only the perineal pattern is not enough for the accurate identification of root-knot nematodes, since M. paranaensis was misidentified as M. incognita for 22 years in Brazil. Esbenshade and Triantaphyllou (1985) reported one population with three esterase bands from Turkey. This esterase phenotype, designated as M3, could not be identified to the species level by the authors. Therefore, M. ethiopica may have been present in Turkey for many years, because the M3 phenotype is very similar to E3. PCR with species-specific primers have been employed to rapidly detect and identify Meloidogyne species using DNA extracted from every stage of the nematode (Devran and Söğüt, 2009). Thus, it is necessary to generate favorable results in terms of the molecular identification of M. ethiopica. Therefore, future research should focus on developing species-specific primers for the reliable differentiation of M. ethiopica from other species.
Figure 4. Amplification products (420 bp) with Far/Rar primers on Meloidogyne populations from the Middle Black Sea Region. Sa: Samsun populations, Si: Sinop populations, Or: Ordu population, Co: Çorum populations, To: Tokat populations, Am: Amasya populations, C: control population, W: water, M: molecular marker with 100 bp.
Figure 5. Amplification products (670 bp) with Fjav/Rjav primers on Meloidogyne populations from the Middle Black Sea Region. Sa: Samsun populations, Si: Sinop populations, Co: Çorum population, C: control population, W: water, M: molecular marker with 100 bp.
683
AYDINLI and MENNAN / Turk J Zool
Acknowledgments The research was supported by the Scientific and Technological Research Council of Turkey (TÜBİTAK) (Project No. 111O793). This study was part of the PhD
thesis of the corresponding author and was presented as a poster at the Plant Health and Sustainable Agriculture Conference, 11–12 May 2015, Ljubljana, Slovenia.
Figure 6. Amplification products (502 bp) with SEC 1F/1R primers on Meloidogyne populations from the Middle Black Sea Region. Sa: Samsun populations, Si: Sinop populations, Or: Ordu populations, Am: Amasya populations, C: control population, W: water, M: molecular marker with 100 bp.

684
AYDINLI and MENNAN / Turk J Zool
References
Adam MAM, Phillips MS, Blok VC (2007). Molecular diagnostic key for identification for single juveniles of seven common and economically important species of root-knot nematode (Meloidogyne spp.). Plant Pathol 56: 190-197.
Akyazı F, Ecevit O (2011). Identification and distribution of root knot nematode species (Meloidogyne spp.) in vegetable fields in Tokat Province. Anadolu J Agr Sci 26: 1-9. (in Turkish with abstract in English).
Aydınlı G, Mennan S (2011). Atypical esterase phenotype from root-knot nematodes in vegetable greenhouse in Samsun. In: Proceedings of the 4th Plant Protection Congress of Turkey, 28–30 June 2011, Kahramanmaraş, Turkey, p. 236.
Aydınlı G, Mennan S, Devran Z, Širca S, Urek G (2013). First report of the root-knot nematode Meloidogyne ethiopica on tomato and cucumber in Turkey. Plant Dis 97: 1262.
Blok VC, Powers TO (2009). Biochemical and molecular identification. In: Perry RN, Moens M, Starr JL, editors. Root-Knot Nematodes. 1st ed. Wallingford, UK: CAB International, pp. 98-118.
Brito JA, Kaur R, Cetintas R, Stanley JD, Mendes ML, McAvoy EJ, Powers TO, Dickson DW (2008). Identification and isozyme characterization of Meloidogyne spp. infecting horticultural and agronomic crops, and weed plants in Florida. Nematology 10: 757-766.
Carneiro RMDG, Almeida MRA, Cofcewicz ET, Magunacelaya JC, Aballay E (2007). Meloidogyne ethiopica, a major root-knot nematode parasitising Vitis vinifera and other crops in Chile. Nematology 9: 635-641.
Carneiro RMDG, Almeida MRA, Queneherve P (2000). Enzyme phenotypes of Meloidogyne spp. populations. Nematology 2: 645-654.
Carneiro RMDG, Carneiro RG, Abrantes IMO, Santos MSNA, Almeida MRA (1996). Meloidogyne paranaensis n. sp. (Nemata: Meloidogynidae), a root-knot nematode parasitizing coffee in Brazil. J Nematol 28: 177-189.
Carneiro RMDG, Randig O, Almeida MRA, Gomes ACMM (2004a). Additional information on Meloidogyne ethiopica Whitehead, 1968 (Tylenchida: Meloidogynidae), a root-knot nematode parasiting kiwi fruit and grape-vine from Brazil and Chile. Nematology 6: 109-123.
Carneiro RMDG, Tigano MS, Randig O, Almeida MRA, Sarah JL (2004b). Identification and genetic diversity of Meloidogyne spp. (Tylenchida: Meloidogynidae) on coffee from Brazil, Central America and Hawaii. Nematology 6: 287-298.
Collange B, Navarrete M, Peyre G, Mateille T, Tchamitchian M (2011). Root-knot nematode (Meloidogyne) management in vegetable crop production: the challenge of an agronomic system analysis. Crop Prot 30: 1251-1262.
Conceição IL, Tzortzakakis EA, Gomes P, Abrantes I, da Cunha MJ (2012). Detection of the root-knot nematode Meloidogyne ethiopica in Greece. Eur J Plant Pathol 134: 451-457.
Correa VR, Mattos VS, Almeida MRA, Santos MFA, Tigano MS, Castagnone-Sereno P, Carneiro RMDG (2014). Genetic diversity of the root-knot nematode Meloidogyne ethiopica and development of a species-specific SCAR marker for its diagnosis. Plant Pathol 63: 476-483.
Coyne DL, Fourie HH, Moens M (2009). Current and future management strategies in resource-poor farming. In: Perry RN, Moens M, Starr JL, editors. Root-Knot Nematodes. 1st ed. Wallingford, UK: CAB International, pp. 444-475.
Çetintaş R, Çakmak B (2011). Diagnosis of Meloidogyne species found on tomatoes, eggplants and cucumbers grown in Kahramanmaraş province by PAGE (Polyacrylamide Gel Elektrophoresis) and perineal patterns. In: Proceedings of the 4th Plant Protection Congress of Turkey, 28–30 June 2011, Kahramanmaraş, Turkey, p. 48.
Devran Z, Mutlu N, Özarslandan A, Elekçioğlu IH (2009). Identification and genetic diversity of Meloidogyne chitwoodi in potato production areas of Turkey. Nematropica 39: 75-83.
Devran Z, Söğüt MA (2009). Distribution and identification of root-knot nematodes from Turkey. J Nematol 41: 128-133.
Di Vito M, Greco N, Oreste G, Saxena MC, Singh KB, Kusmenoglu I (1994). Plant parasitic nematodes of legumes in Turkey. Nematologia Mediterranea 22: 245-251.
Eisenback JD, Hirschmann H, Sasser JN, Triantaphyllou AC (1981). A Guide to the Four Most Common Species of Root-Knot Nematodes (Meloidogyne species) with a Pictorial Key. Raleigh, NC, USA: North Carolina State University.
Elekçioğlu İH, Ohnesorge B, Lung G, Uygun N (1994). Plant parasitic nematodes in the Mediterranean region of Turkey. Nematologia Mediterranea 22: 59-63.
Esbenshade PR, Triantaphyllou AC (1985). Use of enzyme phenotypes for identification of Meloidogyne species (Nematoda: Tylenchida). J Nematol 17: 6-20.
Fargette M (1987). Use of the esterase phenotype in the taxonomy of the genus Meloidogyne. 2. Esterase phenotypes observed in West African populations and their characterization. Revue de Nématologie 10: 45-56.
Garcia LE, Sanchez-Puerta MV (2012). Characterization of a root-knot nematode population of Meloidogyne arenaria from Tupungato (Mendoza, Argentina). J Nematol 44: 291-301.
Hartman KM, Sasser JN (1985). Identification of Meloidogyne species on the basis of differential host test and perineal pattern morphology. In: Barker KR, Carter CC, Sasser JN, editors. Advanced Treatise on Meloidogyne. Volume II: Methodology. Raleigh, NC, USA: North Carolina State University, pp. 69-77.
Hernandez A, Fargette M, Sarah JL (2004). Characterisation of Meloidogyne spp. (Tylenchida: Meloidogynidae) from coffee plantations in Central America and Brazil. Nematology 6: 193-204.
Hunt DJ, Handoo ZA (2009). Taxonomy, identification and principal species. In: Perry RN, Moens M, Starr JL, editors. Root-Knot Nematodes. 1st ed. Wallingford, UK: CAB International, pp. 55-97.

685
AYDINLI and MENNAN / Turk J Zool
Jepson SB (1987). Identification of Root-Knot Nematodes (Meloidogyne Species). 1st ed. Wallingford, UK: CAB International.
Jones TJ, Haegeman A, Danchin EG, Gaur HS, Helder J, Jones MGK, Kikuchi T, Manzanilla-Lopez R, Palomares-Rius JE, Wesemael WML et al. (2013). Top 10 plant-parasitic nematodes in molecular plant pathology. Mol Plant Pathol 1: 946-961.
Karssen G, van Aelst AC (2001). Root-knot nematode perineal pattern development: a reconsideration. Nematology 3: 95-111.
Kaşkavalcı G, Öncüer C (1999). Investigations on distribution and economic importance of Meloidogyne Goeldi, 1887 (Tylenchida: Meloidogynidae) species found in the major areas of hot climate vegetables in Aydın province. Turk J Entomol 23: 149-160 (in Turkish with abstract in English).
Katı T, Mennan S (2006). Researches on species and race determination of root-knot nematodes (Meloidogyne spp.) found in greenhouse of Samsun, Turkey. In: Proceedings of the 28th International Symposium of the European Society of Nematologists, 5–9 June 2006, Blagoevgrad, Bulgaria, p. 130.
Kepenekçi İ, Evlice E, Öztürk G (2014). Taxonomic characteristics of Meloidogyne exigua Goeldi which is a new root-knot nematodes for Turkey and other root-knot nematode species. Bitki Koruma Bülteni 54: 1-9 (in Turkish with abstract in English).
Mennan S, Aydınlı G, Katı T (2011). First report of root-knot nematode (Meloidogyne arenaria) infecting parsley in Turkey. J Phytopathol 159: 694-696.
Mennan S, Ecevit O (1996). Studies on biology, distribution and the ratio of infestation of root-knot nematodes (Meloidogyne spp.) in summer vegetable growing area in Bafra and Çarşamba plains. In: Proceedings of the 3rd Turkish National Congress of Entomology, 24–28 September 1996, Ankara, Turkey, pp. 700-705 (in Turkish with abstract in English).
Munera GE, Bert W, Karssen G, Couvreur M, Waeyenberge L, Vierstraete A, Decraemer W (2010). Morphological and molecular characterisation of Meloidogyne populations associated with Andean fruits and Musa plants in Colombia. Nematropica 40: 121-162.
Oh HK, Bae CH, Kim MI, Wan X, Oh SH, Han YS, Lee HB, Kim I (2009). Molecular biological diagnosis of Meloidogyne species occurring in Korea. Plant Pathol J 25: 247-255.
Özarslandan A, Devran Z, Mutlu N, Elekcioğlu IH (2009). First report of Columbia root-knot nematode (Meloidogyne chitwoodi) in potato in Turkey. Plant Dis 93: 316.
Özarslandan A, Elekçioğlu İH (2010). Identification of the root-knot nematode species (Meloidogyne spp.) (Nemata: Meloidogynidae) collected from different parts of Turkey by molecular and morphological methods. Turk J Entomol 34: 323-335 (in Turkish with abstract in English).
Pais CS, Abrantes IMO (1989). Esterase and malate dehydrogenase phenotypes in Portuguese populations of Meloidogyne species. J Nematol 21: 342-346.
Powers TO, Mullin PG, Harris TS, Sutton LA, Higgins RS (2005). Incorporating molecular identification of Meloidogyne spp. into large-scale regional nematode survey. J Nematol 37: 226-235.
Siddiqi MR (2000). Tylenchida Parasites of Plants and Insects. 2nd ed. Wallingford, UK: CAB International.
Sikora RA, Fernandez E (2005). Nematode parasites on vegetables. In: Luc M, Sikora RA, Bridge J, editors. Plant Parasitic Nematodes in Subtropical and Tropical Agriculture. 2nd ed. Wallingford, UK: CAB International, pp. 319-392.
Söğüt MA, Elekçioğlu IH (2000). Determination of Meloidogyne Goeldi, 1892 (Nemata: Heteroderidae) species races found in vegetable growing areas of the Mediterranean region of Turkey. Turk J Entomol 24: 33-40 (in Turkish with abstract in English).
Stirling GR (1991). Biological Control of Plant Parasitic Nematodes: Progress, Problems and Prospects. 1st ed. Wallingford, UK: CAB International.
Strajnar P, Sirca S (2011). The effect of some insecticides, natural compounds and tomato cv. Venezia with Mi gene on the nematode Meloidogyne ethiopica (Nematoda) reproduction. Acta Agr Slov 97: 5-10.
Strajnar P, Sirca S, Geric Stare B, Urek G (2009). Characterization of the root-knot nematode, Meloidogyne ethiopica Whitehead, 1968, from Slovenia. Russ J Nematol 17: 135-142.
Taylor DP, Netscher C (1974). An improved technique for preparing perineal patterns of Meloidogyne spp. Nematologica 20: 268-269.
Tesarova B, Zouhar M, RyŠanek P (2003). Development of PCR for specific determination of root-knot nematode Meloidogyne incognita. Plant Protact Sci 39: 23-28
Tzortzakakis EA, Adam MAM, Blok VC, Paraskevopoulos C, Bourtzis K (2005). Occurrence of resistance-breaking populations of root-knot nematodes on tomato in Greece. Eur J Plant Pathol 113: 101-105.
Whitehead AG (1968). Taxonomy of Meloidogyne (Nematoda: Heteroderidae) with descriptions of four new species. T Zool Soc London 31: 263-401.
Yüksel H (1974). The distribution of root-knot nematodes (Meloidogyne spp.) in Turkey. Atatürk Üniversitesi Ziraat Fakültesi Dergisi 5: 83-105 (in Turkish with abstract in English).
Zijlstra C, Donkers-Venne DTHM, Fargette M (2000). Identification of Meloidogyne incognita, M. javanica and M. arenaria using sequence characterised amplified region (SCAR) based PCR assays. Nematology 2: 847-853.
Zijlstra C, van Hoof R, Donkers-Venne D (2004). A PCR test to detect the cereal root-knot nematode Meloidogyne naasi. Eur J Plant Pathol 110: 855-860.