induction of hsp70, hsp60, hsp83 and hsp26 and oxidative stress markers in benzene, toluene and...
TRANSCRIPT

Toxicology and Applied Pharmacology 235 (2009) 226–243
Contents lists available at ScienceDirect
Toxicology and Applied Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /ytaap
Induction of hsp70, hsp60, hsp83 and hsp26 and oxidative stress markers in benzene,toluene and xylene exposed Drosophila melanogaster: Role of ROS generation
Mahendra Pratap Singh a, M.M. Krishna. Reddy b, N. Mathur c, D.K. Saxena a, D. Kar Chowdhuri a,⁎a Embryotoxicology Section, Indian Institute of Toxicology Research, P.O. Box No. 80; Mahatma Gandhi Marg, Lucknow- 226 001, Uttar Pradesh, Indiab Analytical Chemistry Section, Indian Institute of Toxicology Research, P.O. Box No. 80; Mahatma Gandhi Marg, Lucknow- 226 001, Uttar Pradesh, Indiac Epidemiology Section, Indian Institute of Toxicology Research, P.O. Box No. 80; Mahatma Gandhi Marg, Lucknow- 226 001, Uttar Pradesh, India(Council of Scientific and Industrial Research, New Delhi, India)
⁎ Corresponding author. Fax: +91 522 2628227/26115E-mail address: [email protected] (D.K
0041-008X/$ – see front matter © 2008 Elsevier Inc. Adoi:10.1016/j.taap.2008.12.002
a b s t r a c t
a r t i c l e i n f oArticle history:
Exposure to benzene, tolue Received 15 July 2008Revised 14 November 2008Accepted 2 December 2008Available online 11 December 2008Keywords:D. melanogasterHeat shock proteinsOxidative stress markersReactive oxygen speciesBenzene, Toluene and xylene
ne and xylene in the human population may pose a health risk. We tested aworking hypothesis that these test chemicals cause cellular toxicity to a non-target organism, Drosophilamelanogaster. Third instar larvae of D. melanogaster transgenic for hsp70, hsp83 and hsp26 and Oregon R+
strain were exposed to 1.0–100.0 mM benzene, toluene and xylene for 2–48 h to examine the heat shockproteins (hsps), ROS generation, anti-oxidant stress markers and developmental end points. The testchemicals elicited a concentration- and time-dependent significant (pb0.01) induction of the hsps in theexposed organism in the order of hsp70Nhsp83≥hsp26 as evident by β-galactosidase activity after 24 h.RT-PCR amplification studies in Oregon R+ larvae revealed a similar induction pattern of these genes alongwith hsp60 in the order of hsp70Nhsp60Nhsp26≥hsp83. Under similar experimental conditions, asignificant induction of ROS generation and oxidative stress markers viz. superoxide dismutase, catalase,glutathione S-transferase, thioredoxin reductase, glutathione, malondialdehyde and protein carbonylcontent was observed. Sub-organismal response was propagated towards organismal response i.e., a delayin the emergence of flies and their reproductive performance. While hsp70 was predominantly induced inthe organism till 24 h of treatment with the test chemicals, a significant or insignificant regression ofHsp70 after 48 h was concurrent with a significant induction (pb0.01) of hsp60Nhsp83≥hsp26 incomparison to the former. A significant positive correlation was observed between ROS generation andthese hsps in the exposed organism till 24 h and a negative correlation between ROS generation and hsp70in them after 48 h indicating a modulatory role of ROS in the induction of hsps. The study suggests thatamong the tested hsps, hsp70 may be used as an early bioindicator of cellular toxicity against benzene,toluene and xylene and D. melanogaster as an alternative animal model for screening the risk posed byenvironmental chemicals.
© 2008 Elsevier Inc. All rights reserved.
Introduction
A variety of chemicals are used in pharmaceutical and chemicalindustries, and among them monocyclic aromatic hydrocarbons likebenzene, toluene and o-, m- and p-xylene isomers are the majorones. These aromatic hydrocarbons are present in gasoline (3–15%),paints, rubber products, plastics, detergents, dyes, pesticides, adhe-sives and several other household products (Derwent et al., 2000;Kim and Kim, 2002; Chang et al., 2007). Thus benzene, toluene andxylene (collectively known as BTX) are harmful substances inoccupational and non-occupation environment and humans arefrequently exposed to low levels of these chemicals for prolongedperiods (McDermott et al., 2008). In recent years, they acquired greatrelevance as one of the major pollutants of the outdoor and indoor
47.. Chowdhuri).
ll rights reserved.
environment (Lee et al., 2002; Srivastava and Devotta, 2007).Benzene is a well known carcinogen that has been causally linkedto leukemia (IARC) (US-EPA, 1996; ATSDR, 2005). However, tolueneand xylene are not currently listed as carcinogens (Gallegos et al.,2007). Benzene, toluene and xylene have been reported to induceoxidative stress, an imbalance of the anti-oxidant system. In thiscontext, alteration of the anti-oxidant system such as SOD, GPx, GSHand MDA content in petrochemical workers was reported (Crouteet al., 2002; Georgieva et al., 2002).
Stress response and anti-oxidant defense system comprising ofstress proteins (also termed as heat shock proteins; Hsps) and anti-oxidants (both enzymatic and non-enzymatic), respectively, are theprimary protective responses that are highly conserved componentsof cellular stress responses found in all phyla from bacteria to man(Lindquist and Craig 1988). Originally discovered as inducibleproteins against heat shock, the hsps counter proteotoxic effectsand play various roles in a cell including chaperoning proteins

227M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
during synthesis, folding, assembly and degradation (Morimoto,1993). Other than heat shock, diverse agents including amino acidanalogues, transition metal ions, sulphydryl reagents, glucosedeprivation, hypoxia etc. have been shown to induce heat shockgenes (Nover, 1991). Among the stress genes family, hsp70 is one ofthe highly conserved genes and first to be induced in Drosophila(Ritossa, 1962, Feder et al., 1992), mammalian cell lines (Farzaneh etal., 2005), parasite and free living protozoa (PeÂrez-Serrano et al.,2000), isopoda (Kohler et al., 1999), freshwater plant species(Ireland et al., 2004), marine teleost (Maradonna and Carnevali,2007) against various physical (Laubitz et al., 2006), physiologicaland chemical stressors (Franzellitti and Fabbri, 2005, Lynes et al.,2007). Apart from hsp70, other hsps also participate in stressresponse in organisms exposed to various stresses. Earlier studieshave shown that Drosophila Hsp83 (homologue of Hsp90 inmammals) and Hsp60 work as chaperone protein refolding systemin coordination with or without Hsp70 (Pratt and Toft, 2003; Younget al., 2004). A study by Bagchi et al. (1996) showed that Hsp89aand Hsp89b were significantly induced in PC-12 cells followingexposure to selected pesticides. A significant induction of bothhsp70 and hsp60 in nematodes and isopods and mites after exposureto heavy metals both under field and laboratory conditionsrespectively were shown indicating their role in biomarker evalua-tion (Arts et al., 2004; Kohler et al., 2005). A study in Trichinellaspiralis muscle larvae after exposure to elevated temperature andhydrogen peroxide showed significantly elevated expression ofHsp70, Hsp60 and Hsp90 (Martinez and Rodrıguez-Caabeiro,1999). Timofeyev et al. (2004) showed that natural organic matterisolated from eutrophic Sanctuary Pond has an adverse impact on anumber of amphipod species tested as evidenced by increasedHsp70 expression. Earlier, small hsps like hsp26 were found to besignificantly induced by a chlorinated pesticide, hexachlorocyclo-hexane in D. melanogaster (Kar Chowdhuri et al., 1999) and by aherbicide 2,4-dichlorophenoxyacetic acid in Saccharomyces cerevisiae(Simoes et al., 2003). The upregulation of these stress genes inhuman population after exposure to various environmental stresseswas shown by Wu and Tanguay (2006).
Molecular oxygen is the key to aerobic life but is also convertedinto cytotoxic byproducts referred to as reactive oxygen species (ROS).In addition to their involvement in the normal metabolic activities,ROS have been reported to play a major role in the toxicity of severalxenobiotics including metals and pesticides (Fortunato et al., 2006).Aerobic organisms have therefore developed multiple defensemechanisms where the cellular anti-oxidant molecules or the anti-oxidant enzymes remove ROS to rescue cells from oxidative stress(Ahmad et al., 2000). The detection of oxidatively modified moleculessuch as protein carbonyl (PC) andmalondialdehyde (MDA) believed tobe specific products of protein and lipid oxidation, respectively havebeen considered to be reliable markers of oxidative stress (Pragasamet al., 2006). Glutathione (γ-glutamyl cysteinyl glycine) is a highlyversatile component of the anti-oxidative system and can directlyreact with a variety of ROS including superoxide anion, hydroxylradical and several organic radicals (Hegde and Verma, 2004). Thusevaluation of anti-oxidant stress markers together with stress geneswill give an account of the chemical stress caused to the exposedorganism.
The present study was aimed to examine the cellular toxicity ofbenzene, toluene and xylene as individual chemicals per se in D.melanogaster with an objective to understand the role of selectedstress genes expression (hsp70, hsp83, hsp60 and hsp26) and anti-oxidant defense systems. The study was further extended to examinethe organismal (emergence of the exposed organisms vis-a-visreproduction) effect of these compounds.
Drosophila as an experimental model has been chosen for itsworldwide acceptability due to its defined genetics and molecularbiology. The model raises few ethical concerns and is approved by
European Centre for the Validation of Alternative Methods (ECVAM)(Festing et al., 1998). Its genome can be easily manipulated by P-element mediated germ line transformation for studying a particulargene of interest under a defined condition (Lis et al., 1983; O'Kane andGehring, 1987; Adams et al., 2000).
Materials and methods
Fly strains. Transgenic Drosophila melanogaster strains viz. (hsp70-lacZ) Bg9 (Lis et al., 1983), (hsp83-lacZ) 83Z-880 (Xiao and Lis, 1989)and (hsp26-lacZ) 351-94A (Glaser and Lis, 1990) that expressbacterial β-galactosidase as a response to stress and wild type D.melanogaster (Oregon R+) were used in the study. Flies and larvaewere reared at 24±1 °C on standard Drosophila diet containing agar-agar, maize powder, sugar, yeast, nepagin (methyl-p-hydroxybenzoate) and propionic acid. Additional yeast suspension wasprovided for healthy growth of the organism.
Treatment schedule. Four different concentrations (1.0, 10.0, 50.0and 100.0 mM) of technical grade benzene (99.7% from Ranbaxy PvtLtd., India), toluene (99.5%, SRL Pvt Ltd., Mumbai, India) and xylene(mixture of o, m and p, 99.8%, SRL Pvt Ltd., Mumbai, India)corresponding to different fractions of the LC50 (48 h) value ofbenzene and toluene and xylene in third instar larvae of bothOregon R+ and transgenic D. melanogaster were used during thepresent study. Larvae were grown on standard Drosophila diet,contaminated with or without different concentrations of three testchemicals in DMSO (99.5%, SRL, Mumbai, India) with finalconcentration of 0.3% (∼42.0 mM) in food (Nazir et al., 2003) for2–48 h. Three control groups were included in the study viz.,negative (without any treatment), positive (temperature shock) andDMSO control (0.3% DMSO).
Quantitative determination of benzene, toluene and xylene by GCMS(SPME-HS). Oregon R+, Bg9 and 83Z-880 third instar larvae weregrown on food contaminated with 100.0 mM benzene, toluene andxylene for 48 h. Three biological replicates (control and treated) wererun for each chemical. A Perkin Elmer gas chromatography (USA)coupled with mass selection detector was used for the analysis.
The injection port was fitted with Solid phase micro-extraction(SPME) inlet guide (Supelco, USA) and pre-drilled thermogreen LB-2 septa to secure the SPME fibre holder. A special inlet linear forSPME was used in this study (Supelco, San Diego, USA). Desorptionof SPME fibre in GC injection port was 5 min at 250 °C with purgegas off. After sample desorption, the fibre was further heated in theinjection port for an additional 30 min at 250 °C with purge gasturned on to remove trace residue in the fibre. The fibre was thenretracted into the protection needle and the needle was insertedinto a clean vial ready for the next sample extraction.
Headspace extraction procedure was followed by using a 2 mllarval homogenate (10% w/v) prepared in deionized water byplacing it in a 10 ml headspace vial. The vials were heated at 65 °Cfor 30 min with magnetic agitation. After the equilibrium, septawere pierced with SPME needle and the SPME polydimethylsiloxane (PDMS) fibre was exposed to headspace for 15 min toaffect the adsorption of the test chemicals in the sample. SPMEfibre was collected and inserted directly into the GC injection at200 °C. Total time of this chromatographic analysis was 27 min.Blank analysis was carried out to avoid any carry over phenomenaand/or external contamination between analyses of samples(Alegretti et al., 2004).
Emergence of flies. Newly eclosed first instar larvae of Oregon R+,Bg9, 83Z-880 and 351-94A (±0.5 h) were transferred to normal food,food containing extracting solvent and to food containing differentconcentrations of test chemicals (50 larvae per vial and 5 vials per

228 M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
group). The number of flies emerging from different groups wasrecorded until all the flies emerged. The emergence of the flies indifferent groups was evaluated as described earlier (Gayathri andKrishnamurthy, 1981).
Fecundity, fertility and reproductive performance. The method ofGayathri and Krishnamurthy (1981) was followed with somemodifications. Freshly eclosed first instar larvae followingsynchronous egg laying for 0.5 h were transferred to normal foodmedium (control), food mixed with 0.3% DMSO (DMSO control) andto food containing different concentrations of benzene, toluene andxylene (1.0–100.0 mM). They were allowed to grow on themthroughout their development. Virgin male and female flies ofOregon R+, Bg9, 83Z-880 and 351-94A emerging from control, DMSOcontrol and treated food were separated and mated in normal foodvials. For each treatment group, 10 pairs of flies in 10 individual vialswere taken and they were transferred to fresh vials everyday for thenext 10 days.
The number of eggs laid during this period was scored. From thisdata, total fecundity (total no. of eggs laid in 10 days), mean daily eggproduction by a female (total no. of eggs/ female/day) and fertilitypercentage (no. of flies eclosed in 10 days/no. of eggs laid in10 days×100) were calculated. The total number of flies emergingfrom the eggs laid during these 10 days was counted; the meannumber of flies emerged per pair for 10 days gave a measure ofreproductive performance.
Temperature shock in third instar Oregon R+, Bg9, 83Z-880 and 351-94Alarvae. Healthy third instar larvae of Oregon R+, Bg9 and 351-94Awere placed on a Petri dish lined withmoist filter paper and were giventemperature shock at 37±1 °C for 1 h as described previously byLakhotia and Mukherjee (1980). For 83Z-880 larvae, temperature shockwas given at 33±1 °C for 1 h (Xiao and Lis, 1989).
hsp70, hsp83 and hsp26 promoter induction experiments. Bg9, 83Z-880 and 351-94A third instar larvae after feeding on normal diet andthat mixed with different concentrations of benzene, toluene andxylene for 2–48 h, were washed thoroughly with Poels' salt solution(PSS) (Lakhotia and Mukherjee, 1980) and then processed for hsp70,hsp83 and hsp26 expression studies as described below.
Soluble O-Nitrophenyl-β-D-galactopyranoside (ONPG) assay in Bg9, 83Z-880 and 351-94A larvae. A modified version of the assaypreviously described by Stringham and Candido (1994) wasfollowed (Nazir et al., 2001). At the termination of the treatment,larvae after a brief washing, were placed in a microcentrifuge tube(20 larvae/tube, 3 replicates/concentration/time point),permeabilized in acetone for 10 min and incubated overnight indark at 37 °C in 600 μl ONPG (SRL, Mumbai, India) staining buffer.After incubation, the reaction was stopped by adding 300 μl 1 MNa2CO3. The extent of reaction was quantified by measuring theabsorbance at 420 nm on GBC-UV spectrophotometer (GBCScientific Equipment, Melbourne, Australia).
RT-PCR analysis of hsp70, hsp83, hsp60 and hsp26 mRNA in third instarlarval tissues of D. melanogaster (Oregon R+)
RNA isolation. Internal tissues from third instar larvae Oregon R+
were explanted in PSS and total RNA was extracted using TRI reagent,chloroform, isopropanol and ethanol following the manufacturers'instructions (Ambion, USA). The extracted RNAwas finally dissolved indiethyl pyrocarbonate (DEPC) treated water and concentration andquanti ty was checked (A260/A280 rat io) by NanoDropSpectrophotometer (Nano Drop Technologies, Wilmington, DE, USA)followed by agarose gel electrophoresis.
Complementary DNA (cDNA) synthesis. Total RNA from the tissues ofcontrol and treated larvae were used for cDNA synthesis using RevertAid™ H Minus first strand cDNA synthesis kit (Fermentas MD, USA)essentially following the manufacturers' protocol. Each reactionmixture consisted of total RNA (3 μg), 0.5 μg/μl oligo (dT)18 primer(1 μl), 5× reaction buffer (4 μl), 20 U Ribolock™ ribonuclease inhibitor,10 mM dNTP mixture, 200 U Revert Aid™ H Moloney MurineLeukemia Virus reverse transcriptase (M-MuLV RT) and DEPC waterto make a final volume of 20 μl. The synthesized cDNA was stored at−20 °C till further use.
Polymerase chain reaction (PCR). The cDNA synthesized fromcontrol and treated groups were amplified by PCR in a thermocycler(Eppendorf, Hamburg, Germany) using primer pairs specific forinducible form of hsp70, hsp83, hsp60 and hsp26. The primer pairsobtained from Sigma, MO, USA were:
hsp70
Forward (F) 5′GAACGGGCCAAGCGCACACTCTC3′Reverse (R) 5′TCCTGGATCTTGCCGCTCTGGTCTC3′hsp83
Forward (F) 5′CCCGTGGCTTCGAGGTGGTCT3′Reverse (R) 5′TCTGGGCATCGTCGGTAGTCATAGG3′hsp60
Forward (F) 5′ CCTCCGGCGGCATTGTCTTC3′Reverse (R) 5′ AGCGCATCGTAGCCGTAGTCACC3′hsp26
Forward (F) 5′CAAGCAGCTGAACAAGCTAACAATCTG3′Reverse (R) 5′GCATGATGTGACCATGGTCGTCCTGG3′β actin
Forward (F) 5′CCTCCGGCGGCATTGTCTTC3′Reverse (R) 5′GGGCGGTGATCTCCTTCTGC3′PCR reaction mixture (total 25 μl) consisted of 1× Taq buffer,1.5 mM MgCl2, 0.20 mM dNTPs mixture, 0.40 μM each of forward andreverse primer, 1 U Taq DNA polymerase (Fermentas Life Sciences,Hanover, MD, USA), 2 μl cDNA and milli-Q water. Optimized PCRconditions consisted of an initial cycle of 94 °C for 3 min (denatura-tion), followed by 35 cycles (hsp70, hsp83, hsp60 and hsp26) of 95 °Cfor 30 s (denaturation), 55 °C for 30 s (annealing), 70 °C for 1 min(extension) and a final step at 70 °C for 5 min (termination). Theamplicons were separated on a 1.5% agarose gel containing ethidiumbromide at 5 V/cm and visualized with a VERSA DOC Imaging SystemModel 1000 (Bio-Rad, USA). The intensity of the bands was quantifiedby the Quantity One software of Bio-Rad, CA, USA. Each experimentwas carried out with three biological replicates prepared fromindependent pools.
Assay of oxidative stress markers. To evaluate the oxidative damagecaused by test chemicals, ROS generation, superoxide dismutase (SOD),catalase (CAT), glutathione S-transferase (GST), and thioredoxinreductase (TrxR) activities, glutathione (GSH) content, protein carbonyl(PC) content and lipid peroxidation (LPO) product were assayed in bothOregon R+ and transgenic third instar larvae. Except for ROSmeasurement for which single cell suspension was used, the rest ofthe abovementioned assays were carried out in 10% tissue homogenate.
Preparation of homogenate. The internal tissues of control andtreated larvae after removal of cuticle were homogenized in cold 0.1Mphosphate buffer (pH 7.4) containing 0.15 M KCl to obtain 10%homogenate. The supernatant after centrifugation at 10,000 ×g wasused for different assays and protein estimation.
Single cell preparation. Tissues of fifteen larvae from control andtreated groups were incubated in collagenase (0.5 mg/ml) for 15 minat 24±1 °C. The cells were then passed through 85 μm nylon mesh.Collagenase was removed by washing the cell suspension with PBSwith gentle shaking. Finally, the cells were processed for different endpoint measurements as described below.
Measurement of ROS. Intracellular ROS generationwas estimated byflow cytometry using the membrane permeable dye 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA; Sigma, St. Louis,

229M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
MO, USA) which is converted by intracellular esterases to 2′,7′-dichlorodihydrofluorescein (DCFH2). The latter is then oxidized by ROSto highly fluorescent 2′,7′-Dichlorofluorescein (DCF) (Royall andIschiropoulos, 1993). Cells from control and treated larvae weresuspended in Schneider's medium (Invitrogen, Paisley, UK) containing10 μM DCFH-DA and incubated for 1 h at 24±1 °C. After incubation,mediawas removed and cellswerewashedwith 0.1MPBS (pH 7.4) andresuspended in PBS for analysis. The level of ROS was measured by anincrease in green fluorescence (DCF fluorescence) at 530 nmwhen thesamplewas excited at 485 nmon BectonDickinsonflowcytometer (BDBiosciences, New Jersey, USA). The datawere analysed using cell questsoftware (Mac OS 8.6) and 10,000 events were counted per sample.
Superoxide dismutase (SOD) (superoxide: superoxide oxidoreductase EC1.15.1.1). The method for estimating cytosolic Cu-Zn SOD describedpreviously by Nishikimi et al. (1972) was followed with minormodification (Gupta et al., 2005). The assay mixture consisted of0.052 M sodium pyrophosphate buffer (pH 8.3), 186 μM phenazinemethosulphate, 300 μM nitroblue tetrazolium, 780 μM reducednicotinamide adenine dinucleotide and the homogenate. One unit ofenzyme activity is defined as enzyme concentration required forinhibiting chromogen production (optical density 560 nm) by 50% in1min under assay condition and expressed as specific activity in units/min/mg larval protein.
Catalase (CAT) (H2O2: H2O2 oxidoreductase EC 1.11.1.6). CAT activityin the control and treated larvae was measured by following theability of the enzyme to split H2O2 within 1 min of incubation time.After incubation, the reaction was stopped by adding dichromate/acetic acid reagent (5% solution of K2Cr2O7: glacial acetic acid, 1:3 byvolume) and the remaining H2O2 was determined by measuringchromic acetate at 570 nm as described previously by Sinha (1972).
Glutathione S-transferase (GST, EC 2.5.1.18). Glutathione S-transferase (GST) activity was determined by the method of Habig etal. (1974) with minor modifications. The reaction mixture consisted of0.2 M sodium phosphate buffer, reduced glutathione (1.0 mM) and 1-chloro 2, 4 dinitrobenzene (CDNB 5.0 mM). An increase in absorbance(340 nm) was measured for 3 min at 30 s interval and the enzymeactivity calculated as nmol CDNB reduced/min/mg larval protein usingmolar extinction coefficient of 6.25×103 M−1 cm−1.
GSH content. GSH in theexposed larvaewasquantifiedusingElmann'sreagent (Elmann, 1959). The assay mixture consisted of 0.2 M phosphatebuffer (pH 8.0), 0.01% 5, 5′-Dithiobis-2-nitro benzoic acid (DTNB) and thelarval homogenate. The reaction was monitored at 412 nm and theamount of GSH was expressed in terms of nmol/mg larval protein.
Thioredoxin reductase (TrxR). We followed the method ofHolmgren and Bjornstedt (1995) with minor modifications (Kanzoket al., 2001). TrxR activity in the exposed larvae was assessedspectrophotometrically following the oxidation of NADPH (decreasein absorbance at 340 nm) and expressed as nmol NADPH oxidized/min/mg larval protein using molar extinction coefficient of6.22×103 M−1 cm−1.
Assay for lipid peroxidation (LPO). We followed the method ofOhkawa et al. (1979) for assaying malondialdehyde (MDA) as ameasurement of LPO using tetraethoxypropane as an externalstandard. The assay mixture consisted of 10% sodium do-decylsulphate (SDS), 0.8% thiobarbituric acid (TBA) and larval tissuehomogenate. Lipid peroxide level was expressed in terms of nmolMDA formed/h/mg larval protein.
Determination of protein carbonyl (PC) content. We followed themethod of Levine et al. (1990) with minor modification for
determination of PC content. Two equal aliquots of supernatantfraction were taken, one treated with equal volume of 2, 4-Dinitrophenyl hydrazine (10 mM dissolved in 2 M HCl) (test sample)and the otherwith 2MHCl (blank). Eachmixturewas incubated for 1 h,followed by precipitation with 20% TCA and subsequently extractedwith ethanol:ethylacetate mixture (1:1). The pellets were thendissolved in 1.0 ml 6 M guanidine hydrochloride. The spectrum ofthe DNPH treated sample versus the HCl blank was determined at370 nm and results were expressed in terms of nmol DNPHincorporated/mg larval protein based on molar absorption coefficientof 22,000 M−1 cm−1.
Protein estimation. Protein concentration was determined by themethod of Lowry et al. (1951) using protein estimation kit (BangaloreGenei, Bangalore, India) essentially following manufacturers' protocoland bovine serum albumin (BSA) as standard.
Statistical analysis. To analyse the effects of test chemicals, theirdoses, and interaction, 4×4 factorial ANOVA (three test chemicalsalong with DMSO control and four concentrations) was usedseparately on daily egg laying and reproductive performance. Thepost hoc analysis was carried out calculating the least significantdifferences at 5% level of significance. Significance of difference inmean values of each test chemical was evaluated separately at eachconcentration level. Similarly differences in mean values of eachconcentration were tested separately for each test chemical by usingone way ANOVAwhen interactions were found to be significant. Priorto applying the ANOVA, homogeneity of variance was ascertainedusing Levene's test of equality error variance. Similar analysis wascarried out for emergence of adult flies considering days as replicates.Two-way analysis of variance was carried out for β-galactosidaseassay, RT-PCR, biochemical parameters (SOD, CAT, GSH, GST, TrxR, LPOand PC), ROS estimation and protein estimation as dependentvariables. (Zar, 1984). SPSS 14.0 (SPSS, Mapinfo Max, Troy, NY, USA)was used to analysis the data.
Results
During the course of study, no overt signs of toxicity were observedin the exposed organisms. However, organisms exposed to 100.0 mMof test chemicals displayed sluggish movement after 48 h exposure.Larvae fed DMSO for 2–48 h did not exhibit any significant change inthe endpoints measured as compared to control. Since the datagenerated for all the endpoints except reporter gene assay werecomparable for Oregon R+ and Bg9, 83Z-880 and 351-94A, datapresented are from D. melanogaster (Oregon R+).
Detection of benzene, toluene and xylene in larvae of D. melanogaster
We detected presence of benzene, toluene and xylene in theexposed larvae of D. melanogaster by GCMS coupled with single phasemicro-extraction (SPME) and observed 80, 82 and 83% recovery ofbenzene, toluene and xylene in the exposed organism respectively inrespect to food (unpublished data). Thus the nominal concentration of100.0 mMbenzene, toluene and xylene exposed organism is 80.0, 82.0and 83.0 mM respectively. The other concentrations of the testchemicals were not measured in the exposed larvae and hence thedata are not given. To maintain uniformity in data representation, weshowed the concentrations of the chemicals mixed in food.
Emergence of adult flies
In control, DMSO control and at the lowest dietary concentration(1.0 mM) of benzene, toluene and xylene, we observed comparableemergence pattern of the flies. No statistically significant changes inemergence pattern were observed among 1.0 mM benzene, toluene

230 M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
and xylene exposed groups. Overall statistically significant reducingtrend was observed in emergence of the flies in the exposed groupswith an increase in concentration of the test chemicals [10.0 mM(pb0.001), 50.0 mM (pb0.001) and 100.0 mM (pb0.001) treatedgroups in comparison to control and DMSO control]. We did notobserve any statistically significant deviation in emergence of flieseither in control or in DMSO control and subsequent data presentation
Fig.1. Effect of benzene (a), toluene (b) and xylene (c) on the emergence pattern of Drosophiladietary concentration of the test chemicals. Significance is ascribed as ⁎⁎pb0.001 vs. control o10.0 and 50.0 mM test chemicals respectively. Con=control, DMC=DMSO control, B1, B10T100=toluene 1.0, 10.0, 50.0 and 100.0 mM respectively; X1, X10, X50 and X100=xylene 1.0
includes control only for comparison. At 10.0 mM dietary concentra-tion of the test chemicals, a delay of one day in the emergence of theflies was concomitant with a significant (pb0.001) reduction in thenumber of flies emerged (43, 54 and 55% flies emerged on day 10 fromthe three test chemicals treated groups as compared to control). Adelay of 2 days in the emergence of flies along with a statisticallysignificant (pb0.001) reduction (20, 23 and 26% flies emerged on day
melanogaster (Oregon R+). Note a significant delay in the emergence of flies at 100.0 mMr DMSO control or 1.0 mM test chemicals. # and $=significance, pb0.01 as compared to, B50 and B100=benzene 1.0, 10.0, 50.0 and 100.0 mM respectively; T1, T10, T50 and, 10.0, 50.0 and 100.0 mM respectively.
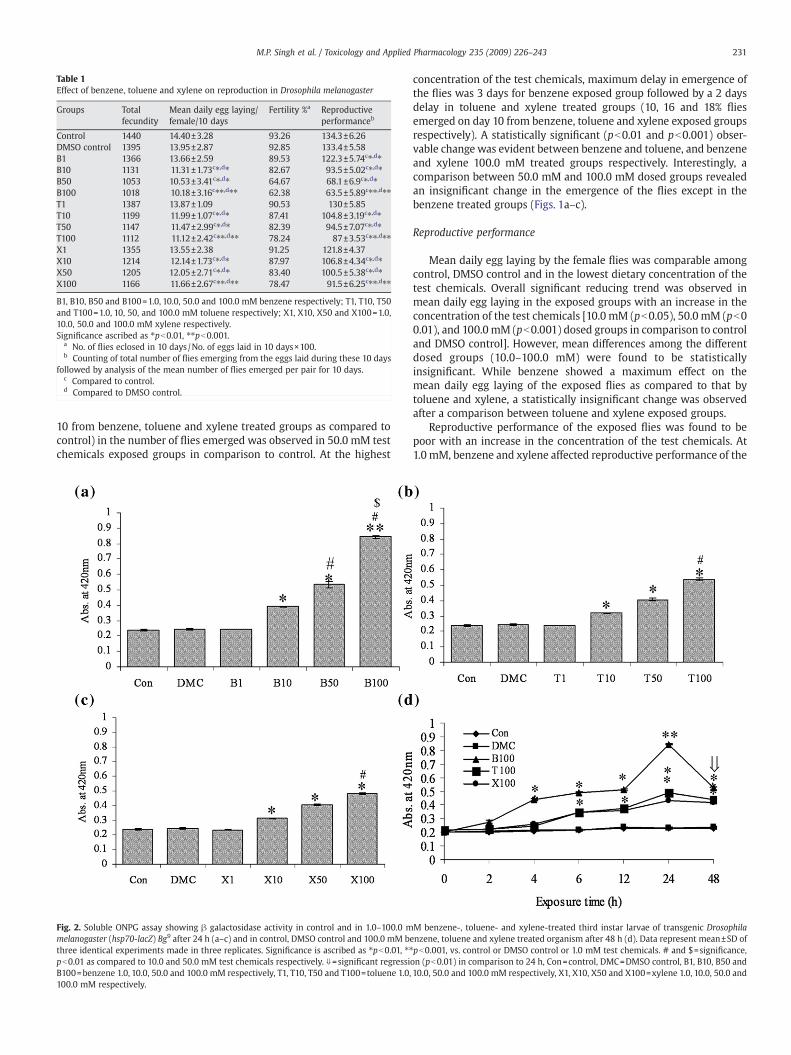
Table 1Effect of benzene, toluene and xylene on reproduction in Drosophila melanogaster
Groups Totalfecundity
Mean daily egg laying/female/10 days
Fertility %a Reproductiveperformanceb
Control 1440 14.40±3.28 93.26 134.3±6.26DMSO control 1395 13.95±2.87 92.85 133.4±5.58B1 1366 13.66±2.59 89.53 122.3±5.74c⁎,d⁎B10 1131 11.31±1.73c⁎,d⁎ 82.67 93.5±5.02c⁎,d⁎B50 1053 10.53±3.41c⁎,d⁎ 64.67 68.1±6.9c⁎,d⁎B100 1018 10.18±3.16c⁎⁎,d⁎⁎ 62.38 63.5±5.89c⁎⁎,d⁎⁎T1 1387 13.87±1.09 90.53 130±5.85T10 1199 11.99±1.07c⁎,d⁎ 87.41 104.8±3.19c⁎,d⁎T50 1147 11.47±2.99c⁎,d⁎ 82.39 94.5±7.07c⁎,d⁎T100 1112 11.12±2.42c⁎⁎,d⁎⁎ 78.24 87±3.53c⁎⁎,d⁎⁎X1 1355 13.55±2.38 91.25 121.8±4.37X10 1214 12.14±1.73c⁎,d⁎ 87.97 106.8±4.34c⁎,d⁎X50 1205 12.05±2.71c⁎,d⁎ 83.40 100.5±5.38c⁎,d⁎X100 1166 11.66±2.67c⁎⁎,d⁎⁎ 78.47 91.5±6.25c⁎⁎,d⁎⁎
B1, B10, B50 and B100=1.0, 10.0, 50.0 and 100.0 mM benzene respectively; T1, T10, T50and T100=1.0, 10, 50, and 100.0 mM toluene respectively; X1, X10, X50 and X100=1.0,10.0, 50.0 and 100.0 mM xylene respectively.Significance ascribed as ⁎pb0.01, ⁎⁎pb0.001.
a No. of flies eclosed in 10 days/No. of eggs laid in 10 days×100.b Counting of total number of flies emerging from the eggs laid during these 10 days
followed by analysis of the mean number of flies emerged per pair for 10 days.c Compared to control.d Compared to DMSO control.
231M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
10 from benzene, toluene and xylene treated groups as compared tocontrol) in the number of flies emerged was observed in 50.0 mM testchemicals exposed groups in comparison to control. At the highest
Fig. 2. Soluble ONPG assay showing β galactosidase activity in control and in 1.0–100.0 mmelanogaster (hsp70-lacZ) Bg9 after 24 h (a–c) and in control, DMSO control and 100.0 mM bethree identical experiments made in three replicates. Significance is ascribed as ⁎pb0.01, ⁎⁎pb0.01 as compared to 10.0 and 50.0 mM test chemicals respectively. ⇓=significant regressiB100=benzene 1.0, 10.0, 50.0 and 100.0 mM respectively, T1, T10, T50 and T100=toluene 1.0,100.0 mM respectively.
concentration of the test chemicals, maximum delay in emergence ofthe flies was 3 days for benzene exposed group followed by a 2 daysdelay in toluene and xylene treated groups (10, 16 and 18% fliesemerged on day 10 from benzene, toluene and xylene exposed groupsrespectively). A statistically significant (pb0.01 and pb0.001) obser-vable change was evident between benzene and toluene, and benzeneand xylene 100.0 mM treated groups respectively. Interestingly, acomparison between 50.0 mM and 100.0 mM dosed groups revealedan insignificant change in the emergence of the flies except in thebenzene treated groups (Figs. 1a–c).
Reproductive performance
Mean daily egg laying by the female flies was comparable amongcontrol, DMSO control and in the lowest dietary concentration of thetest chemicals. Overall significant reducing trend was observed inmean daily egg laying in the exposed groups with an increase in theconcentration of the test chemicals [10.0 mM (pb0.05), 50.0 mM (pb00.01), and 100.0 mM (pb0.001) dosed groups in comparison to controland DMSO control]. However, mean differences among the differentdosed groups (10.0–100.0 mM) were found to be statisticallyinsignificant. While benzene showed a maximum effect on themean daily egg laying of the exposed flies as compared to that bytoluene and xylene, a statistically insignificant change was observedafter a comparison between toluene and xylene exposed groups.
Reproductive performance of the exposed flies was found to bepoor with an increase in the concentration of the test chemicals. At1.0 mM, benzene and xylene affected reproductive performance of the
M benzene-, toluene- and xylene-treated third instar larvae of transgenic Drosophilanzene, toluene and xylene treated organism after 48 h (d). Data represent mean±SD ofpb0.001, vs. control or DMSO control or 1.0 mM test chemicals. # and $=significance,on (pb0.01) in comparison to 24 h, Con=control, DMC=DMSO control, B1, B10, B50 and10.0, 50.0 and 100.0 mM respectively, X1, X10, X50 and X100=xylene 1.0, 10.0, 50.0 and

232 M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
exposed organism significantly [benzene (pb0.001) and xylene(p≤0.05) respectively] while toluene exposure did not elicit anysignificant change of the same as compared to control and DMSO
Fig. 3. Soluble ONPG assay showing β-galactosidase activity in control and 1.0–100.0 mMmelanogaster (hsp83-lacZ) 83Z-880 (a–c) and hsp26-lacZ-351-94A (e–g) after 24 h, and in contrepresent mean±SD of three identical experiments made in three replicates. Significance is aand $=significance at pb0.01 as compared to 10.0 and 50.0 mM test chemicals respectively100.0 mM respectively, T1, T10, T50 and T100=toluene 1.0, 10.0, 50.0 and 100.0 mM, X1, X1
control. Interestingly, no significant observable change was evidentbetween benzene and xylene treated groups. A significant concentra-tion-dependent effect on reproductive performance of the flies was
benzene-, toluene- and xylene-treated third instar larvae of transgenic Drosophilarol and 100.0 mM benzene-, toluene and xylene-treated organisms after 48 h (d, h). Datascribed as ⁎pb0.01, ⁎⁎pb0.001 vs. control or DMSO control or 1.0 mM test chemicals. #. Con=control, DMC=DMSO control, B1, B10, B50 and B100=benzene 1.0, 10.0, 50.0 and0, X50 and X100=xylene 1.0, 10.0, 50.0 and 100.0 mM respectively.

Fig. 4. RT-PCR analysis of hsp70, hsp60, hsp83 and hsp26 mRNA in 100.0 mM benzene-, toluene- and xylene-treated third instar larvae of D. melanogaster (Oregon R+). Panel (a)represents an agarose gel picture from one of the experiments. Lane 1=control, lane 2=DMSO control, lanes 3–5=benzene, toluene and xylene 6 h respectively, lane 6–8=benzene,toluene and xylene 12 h respectively, lanes 9–11=benzene, toluene and xylene 24 h respectively and lanes 12–14 benzene, toluene and xylene 48 h respectively. Percent change of theabove mentioned hsps against control after benzene (b), toluene (c), and xylene (d) treatment to the larvae representative of three independent experiments. Significance is ascribedas ⁎pb0.01, ⁎⁎pb0.001, vs. control or DMSO control. ⇓=significant regression (pb0.01) as compared to 24 h. B100, T100, X100=benzene, toluene and xylene 100.0 mM respectively.
233M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243

Fig. 5. ROS generation (a–d) in control and in 1.0–100.0 mM benzene-, toluene- and xylene-treated third instar larvae of D. melanogaster (Oregon R+) after 48 h exposure (a–c) and incontrol, DMSO control and 100.0 mM benzene, toluene and xylene treated organisms for 2–48 h (d). Data represent mean±SD of three identical experiments made in three replicatesand significance is ascribed as ⁎pb0.01, ⁎⁎pb0.001, vs. control or DMSO control or 1.0 mM test chemicals. # and $=significance at pb0.01 as compared to 10.0 and 50.0 mM testchemicals respectively. Con=control, DMC=DMSO control, B1, B10, B50 and B100=benzene 1.0, 10.0, 50.0 and 100.0 mM respectively, T1, T10, T50 and T100=toluene 1.0, 10.0, 50.0and 100.0 mM, X1, X10, X50 and X100=xylene 1.0, 10.0, 50.0 and 100.0 mM respectively.
234 M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
observed after 10.0, 50.0 and 100.0 mM benzene, toluene and xylenetreatment respectively with a maximum effect by benzene followedby toluene and xylene. Interestingly, a comparison between 50.0 mMand 100.0 mM dosed groups revealed an insignificant change in theirreproductive performance (Table 1).
Quantitative detection of hsp70, hsp83 and hsp26 expression inD. melanogaster
The extent of induction of one of the major hsps, hsp70 and othermembers of the hsp family, viz., hsp83 and hsp26 were measured byβ-galactosidase activity in the third instar larvae of Bg9, 83Z-880 and351-94A exposed to different concentrations (1.0–100.0 mM) ofbenzene, toluene and xylene.
An insignificant statistical change was observed for all the datapoints at any given time among control, DMSO control and the lowestdietary concentration (1.0 mM) of the test chemicals. In the rest of thegroups, a concentration- (Figs. 2a–c) and time-dependent (Fig. 2d)significant increase in β-galactosidase activity as a measure of hsp70expression was observed in the exposed Bg9 larvae as compared tocontrol and DMSO control. Larvae exposed to 100 mM benzeneexhibited a 1.3-fold increase (pb0.01) in β-galactosidase activity after2 h and a maximum β-galactosidase activity after 24 h (pb0.001; 3.6-
Fig. 6. Cu-Zn SOD (a–d) and CAT (e–h) activities in control and 1.0–100.0 mM benzene-, toexposure (a–c, e–g) and in control and 100.0 mM benzene-, toluene- and xylene-treated orgathree replicates and significance is ascribed as ⁎pb0.01, ⁎⁎pb0.001, vs. control or DMSO con50.0 mM test chemicals respectively. Con=control, DMC=DMSO control, B1, B10, B50 and B1010.0, 50.0 and 100.0 mM, X1, X10, X50 and X100=xylene 1.0, 10.0, 50.0 and 100.0 mM respe
fold) as compared to control followed by a significant (pb0.001)regression (46%) after 48 h in comparison to that observed after 24 h(Fig. 2d). At this concentration, toluene and xylene elicited amaximum 2.0- and 1.8-fold induction of β-galactosidase activity inthe larvae exposed for 24 h and an insignificant (pN0.05) regression inthe activity in the larvae after 48 h as compared that observed after24 h (17 and 13% regression in β-galactosidase activity after tolueneand xylene treatment respectively). A comparison of β-galactosidaseactivity was made among the 100.0 mM test chemicals treated groupsfor a particular exposure time (24 h). A statistically significant changein β-galactosidase activity was observed between benzene andtoluene (pb0.001) and benzene and xylene (pb0.001) and aninsignificant change between toluene and xylene (pN0.05) treatedgroups respectively (Fig. 2d).
83Z-880 larvae when exposed to 1.0 mM of the test chemicals, itwas observed that β-galactosidase activity in these larvae and also inthe control and DMSO control did not showany statistically significantchange. With other concentrations, a time- (Fig. 3d) and concentra-tion-dependent (Figs. 3a–c) significant increase in β-galactosidaseactivity measured for hsp83 promoter induction was observed.However, hsp83 promoter induction in 100.0 mM test chemicalstreated 83Z-880 larvae after 24 h was significantly less (pb0.01) ascompared to peak hsp70 promoter induction in 100mM test chemicals
luene- and xylene-treated third instar larvae of D. melanogaster (Oregon R+) after 48 hnisms for 2–48 h (d, h). Data represent mean±SD of three identical experiments made introl or 1.0 mM test chemicals. # and $=significance at pb0.01 as compared to 10.0 and0=benzene 1.0, 10.0, 50.0 and 100.0 mM respectively T1, T10, T50 and T100=toluene 1.0,ctively.

235M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243

236 M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
exposed Bg9 larvae after 24 h (29, 25 and 24% less β-galactosidaseactivity measured for hsp83 induction in 100.0 mM benzene, tolueneand xylene exposed larvae respectively). 83Z-880 larvae exposed to100.0mM test chemicals for 48 h did not exhibit any regression in theirβ-galactosidase activity unlike that observed in Bg9 larvae. Instead,benzene and toluene evoked a significant increase β-galactosidaseactivity (19.5 and 26%) and xylene an insignificant increase in theenzyme activity (15.0%) in these exposed larvae respectively incomparison to that observed after 24 h.
In 351-94A larvae, where in hsp26 promoter induction wasmeasured by β-galactosidase activity, a trend similar to that observedin 83Z-880 larvae was found after exposure to the test chemicals (Figs.3e–h). An overall comparison of β-galactosidase activity as a measureof hsp70, hsp83 and hsp26 induction after 100.0 mM benzene, tolueneand xylene treatment to the organism for 24 h revealed the inductionof stress genes in the order of hsp70N, hsp83≥hsp26 with benzenebeing the highest inducer of these genes followed by toluene andxylene. However, an insignificant changewas observed between hsp83and hsp26 expression measured by β-galactosidase activity in the100.0 mM test chemicals treated organisms for 24 h. A furthercomparison among the stress genes in the larvae exposed to the testchemicals for 48 h revealed a trend in the order ofhsp83≥hsp26Nhsp70 in themwith benzene being the highest inducerfollowed by the rest (Figs. 2 and 3).
mRNA expression of different stress genes in the exposedD. melanogaster (Oregon R+) larvae
To examine whether the reporter gene activity in the testchemicals exposed Bg9, 83Z-880 and 351-94A larvae mimics theresident gene expression, D. melanogaster (Oregon R+) larvaeexposed to 100.0 mM test chemicals for 6, 12, 24 and 48 h wereassayed for hsp70, hsp83 and hsp26 mRNA expression by RT-PCR.While a significant (pb0.01) increase (185.4%) in hsp70 mRNA wasobserved in larvae treated with benzene for 6 h, toluene and xylenetreated larvae exhibited significant induction (pb0.01) of this geneafter 12 h in comparison to control and DMSO control. While amaximum induction (pb0.001; 442% increase) of hsp70 mRNA wasobserved in benzene treated larvae after 24 h, toluene and xyleneexposure resulted in a 407% and 407% increase in hsp70 mRNArespectively in the exposed organism. At this exposure time,benzene, toluene and xylene treated organisms elicited a 255, 245and 243% increase in hsp83 mRNA respectively and a 205, 179, and192% increase in hsp26 mRNA expression respectively. In addition tothe above, mRNA expression of another stress gene, hsp60 wasexamined under similar experimental condition. While benzeneexposed organisms after 12 h elicited a significant increase (204%;pb0.01) of hsp60 mRNA, toluene and xylene exposed organismsonly after 24 h elicited a 355 and 389% increase of this mRNArespectively as compared to control. After 48 h of 100.0 mM testchemicals treatment, when hsp70 expression was found to besignificantly regressed in the exposed organism in comparison tothat observed after 24 h, a significant (pb0.01) upward trend ofhsp60 and hsp26 mRNA and an insignificant (pN0.05) upward trendof hsp83 mRNA expression were observed in them. Hereafter, acomparison among these four tested stress gene mRNA expressionin the organism exposed to the test chemicals for 24 h indicated atrend of hsp70Nhsp60Nhsp26≥hsp83 and after 48 h a trendhsp60Nhsp26Nhsp83Nhsp70 respectively (Fig. 4).
Fig. 7. GSH (a–d) and TrxR activities (e–h) in control and in 1.0–100.0 mM benzene-, tolexposure (a–c, e–g) and in control and in 100.0mMbenzene-, toluene andxylene-treated orgthree replicates. Significance is ascribed as ⁎pb0.01, ⁎⁎pb0.001, vs. control or DMSO control ortest chemicals respectively. Con=control, DMC=DMSO control, B1, B10, B50 and B100=benzenand 100.0 mM, X1, X10, X50 and X100=xylene 1.0, 10.0, 50.0 and 100.0 mM respectively.
ROS generation in benzene, toluene and xylene treated third instarD. melanogaster (Oregon R+) larvae
Fig. 5 shows ROS generation measured in benzene, toluene andxylene treated larvae by flow cytometry. ROS generation incontrol, DMSO control and 1.0 mM test chemicals treatedorganism were comparable without any significant statisticalchange (pN0.05). In the rest of the groups, a significantconcentration- (Figs. 5a–c) and time-dependent (Fig. 5d) increasein ROS generation was observed. A maximum increase in ROSgeneration was observed in the larvae exposed to 100.0 mMbenzene in comparison to control or DMSO control (pb0.001; a4.6- and 7.9-fold increase in ROS generation in 100.0 mM benzenetreated groups after 4 and 48 h in comparison to controlrespectively). Larvae exposed to 100.0 mM toluene and xyleneexhibited a significant 2.9- and 2.7-fold increase in ROS generationafter 6 h and a maximum induction after 48 h (6.7- and 5.9-foldincrease in ROS generation for toluene and xylene respectively). Acomparison of induced ROS generation among the 100.0 mM testchemicals treated groups for an exposure period (24 h) revealed astatistically significant change between benzene and toluene(pb0.001) and benzene and xylene (pb0.001) and an insignificantchange between toluene and xylene (pN0.05) treated groupsrespectively.
Measurement of anti-oxidant markers in benzene, toluene and xylenetreated third instar D. melanogaster (Oregon R+) larvae
Cu-Zn SOD activityFig. 6 shows Cu-Zn SOD activity in test chemicals treated larvae.
The enzyme activity in control, DMSO control and 1.0 mM testchemicals treated larvae were comparable without any statisticallysignificant change (pN0.05). In the rest of the groups, a time- (Fig.6d) and concentration-dependent (Figs. 6a–c) significant increasein SOD activity was observed. A maximum 2.9-fold (pb0.001)increase in enzyme activity was observed in 100.0 mM benzeneexposed organism after 48 h. A similar trend was observed in100.0 mM toluene and xylene treated larvae with a maximum2.5- and 2.4-fold (pb0.001) increase in SOD activity respectivelyafter 48 h. A comparison of SOD activity among 100.0 mM testchemicals treated groups for an exposure period (24 h) revealed astatistically significant change between benzene and toluene(pb0.001) and benzene and xylene (pb0.001) and an insignificantchange between toluene and xylene (pN0.05) treated groupsrespectively.
Catalase activityLike SOD activity observed in control and test chemicals treated
groups, a time- (Fig. 6h) and concentration-dependent (Figs. 6e–g)increase in CAT activity was observed in these groups. While100.0 mM benzene induced significant (pb0.001) CAT activity in thelarvae exposed for 4 h in comparison to control or DMSO control(4.2-fold increase as compared to control), toluene and xyleneinduced significant (pb0.001) enzyme activity in the exposedorganism after 6 h (a 3.4- and 3.4-fold increase in enzyme activityafter toluene and xylene treatment respectively as compared tocontrol). A comparison of CAT activity among the three testchemicals treated groups showed a trend similar to that observedfor SOD activity as stated above.
uene- and xylene-treated third instar larvae of D. melanogaster (Oregon R+) for 48 hanisms for 2–48h (d, h). Data representmean±SDof three identical experimentsmade in1.0mM test chemicals. # and $=significance at pb0.01 as compared to 10.0 and 50.0mMe 1.0,10.0, 50.0 and 100.0mM respectively; T1, T10, T50 and T100=toluene 1.0,10.0, 50.0

237M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243

Fig. 8. Glutathione S-transferase (GST) activities in control and in 1.0 mM–100.0 mM benzene-, toluene- and xylene-treated third instar larvae of D. melanogaster (Oregon R+) for 48 hexposure (a–c) and in normal and in 100.0 mM benzene-, toluene- and xylene-treated organisms for 2–48 h (d). Data represent mean±SD of three identical experiments made inthree replicates. Significance is ascribed as ⁎pb0.01, ⁎⁎pb0.001, vs. control or DMSO control or 1.0 mM test chemicals. # and $=significance at pb0.01 as compared to 10.0 and50.0 mM test chemicals respectively. Con=control, DMC=DMSO control, B1, B10, B50 and B100=benzene 1.0, 10.0, 50.0 and 100.0 mM respectively; T1, T10, T50 and T100=toluene1.0, 10.0, 50.0 and 100.0 mM, X1, X10, X50 and X100=xylene 1.0, 10.0, 50.0 and 100.0 mM respectively.
238 M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
GSH content and TrxR activityFig. 7 shows GSH content and TrxR activity in the control,
DMSO control and test chemicals treated larvae. No significantchange (pN0.05) in GSH content was observed in 1.0 mM benzene,toluene and xylene treated larvae throughout the exposure period(2–48 h) in comparison to control or DMSO control. Larvaeexposed to 10.0 mM benzene showed a significant (pb0.01)depletion in their GSH content after 12 h (13.2% depletion ascompared to control). Toluene and xylene at this concentrationcaused a significant (pb0.01) depletion in GSH content in theexposed larvae after 24 h (19.7 and 21.0% depletion as comparedto control respectively) (figure not shown). A maximum depletionin GSH content was observed at the highest dietary concentrationof benzene, toluene and xylene exposed organism after 48 h(pb0.001; a 70.2, 50.8 and 48.3% depletion in GSH content afterbenzene, toluene and xylene treatment respectively as comparedto control) (Figs. 7a–d).
TrxR activity was significantly (pb0.001) enhanced in the larvaeexposed to 10.0 mM benzene for 24 h while the same wassignificantly induced in the toluene and xylene treated larvae after48 h in comparison to control or DMSO control. A maximuminduction in TrxR activity was observed in 100.0 mM test chemicalstreated larvae after 48 h (pb0.001, a 5.2-, 3.6- and 3.5-fold increasein TrxR activity after benzene, toluene and xylene treatmentrespectively as compared to control) (Figs. 7e–h). A comparison ofGSH content and TrxR activity among the three test chemicalstreated groups revealed a trend similar to that observed for SOD andCAT activities as stated above.
GST activityFig. 8 shows GST activity in the test chemicals treated larvae. A
statistically significant (pb0.01) increase in the enzyme activity wasobserved in 10.0 and 50.0 mM benzene exposed larvae after 12 h ascompared to control or DMSO control (figure not shown). Larvaeexposed to 100.0 mM benzene for 6 h exhibited a significantlyenhanced (pb0.001) enzyme activity (a 3.6-fold increase in GSTactivity as compared to control). At this concentration, toluene andxylene elicited a significant increase in the enzyme activity in theexposed larvae after 12 h (pb0.001; a 3.4- and 3.4-fold increase in GSTactivity after toluene and xylene treatment respectively as comparedto control). A maximum induction in GST activity was observed in thelarvae exposed to 100.0 mM benzene, toluene and xylene for 48 h (a6.2-, 5.4- and 5.1-fold increase in GST activity after benzene, tolueneand xylene treatment respectively as compared to control respec-tively) (Fig. 8d). A comparison of GST activity among the three testchemicals treated groups revealed a trend similar to that observed forSOD and CAT activities as stated above.
Lipid peroxidation (MDA content) and protein carbonyl content (PC content)
Fig. 9 shows the MDA content and PC content in control, DMSOcontrol and treated larvae. In 1.0 mM benzene, toluene and xylenetreated larvae, MDA content was comparable to control and DMSOcontrol throughout the exposure period (2–48 h). A statisticallysignificant increase (pb0.01; 1.7-fold) in MDA content was evident in10.0 mM benzene dosed group after 12 h while those exposed totoluene and xylene, exhibited significantly increased MDA content

Fig. 9.MDA (a–d) and PC content (e–h) in control and in 1.0–100.0 mM benzene-, toluene- and xylene-treated third instar larvae of D. melanogaster (Oregon R+) for 48 h exposure(a–c, e–g) and in control and 100.0 mM benzene-, toluene- and xylene-treated organisms for 2–48 h (d, h). Data represent mean±SD of three identical experiments made in threereplicates. Significance is ascribed as ⁎pb0.01, ⁎⁎pb0.001, vs. control or DMSO control or 1.0 mM test chemicals. # and $=significance at pb0.01 as compared to 10.0 and 50.0 mM testchemicals respectively. Con=control, DMC=DMSO control, B1, B10, B50 and B100=benzene 1.0,10.0, 50.0 and 100.0mM respectively, T1, T10, T50 and T100=toluene 1.0,10.0, 50.0 and100.0 mM, X1, X10, X50 and X100=xylene 1.0, 10.0, 50.0 and 100.0 mM respectively.
239M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243

Fig. 10. Total protein content in third instar larvae of D. melanogaster (Oregon R+) exposed to 100.0 mM benzene, toluene and xylene for 6–48 h. Data represent mean±SD of threeidentical experiments made in three replicates and significance is ascribed as ⁎pb0.01, ⁎⁎pb0.001 vs. control or DMSO control.▿=significance at pb0.01 as compared to 100.0 mM oftest chemicals after 6 h. Con=control, DMC=DMSO control, B100, T100 and X100=100.0 mM benzene, toluene and xylene respectively.
240 M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
after 24 h (pb0.01; 2.0- and 1.7-fold increase in MDA content intoluene and xylene treated groups respectively as compared tocontrol) (figure not shown). Larvae exposed to 100.0 mM benzenefor 6–48 h exhibited a significant increase in MDA content ascompared to control [a 2.2-fold increase in MDA content after 6 h(pb0.01) and a 3.1-fold increase in MDA content after 48 h (pb0.001)respectively as compared to control] (Figs. 9a–d). At this concentra-tion, toluene and xylene induced MDA content reached its maximum(2.6- and 2.5-fold respectively) after 48 h in the exposed larvae.
PC content in the test chemicals exposed larvae was significantlyinduced in a manner similar to that observed for MDA content asstated above (Figs. 9d–f) except that in 100.0 mM xylene treatedlarvae, significant induction in PC content was evident only after 12 h(Figs. 9e–h). A comparison of MDA and PC content among the threetest chemicals treated groups revealed a trend similar to that observedfor SOD and CAT activities as stated above.
Protein estimation
Third instar larvae exposed to 100.0 mM test chemicals exhibited astatistically significant decrease (pb0.01) in the total protein contentin their tissues. Larvae exposed to 100.0 mM benzene showed asignificant decrease in total protein content only after 12 h where asthose exposed to 100.0 mM toluene and xylene showed a significantlyless (pb0.01) total protein content after 24 h in comparison to controlor DMSO control (Fig. 10).
Discussion
The present study shows benzene, toluene and xylene inducedcellular and organismal toxicity in D. melanogaster by the expressionof stress, oxidative stress genes and organismal assays.
Benzene, toluene and xylene are a group of special lipophilicsubstances that have higher capability to permeate the skin orsubcutaneous fat (Adami et al., 2006). These chemicals have diversering and side chain structures that could significantly, and indepen-dently affect their absorption characteristics and toxicity (Chou et al.,2003).
Development of the organism was affected, as evident by delay inthe emergence of the adult flies and a significant decrease in thenumber of flies that emerged at the tested concentrations of benzene,toluene and xylene. This raises a possibility that such a developmentalchange may be due to environmental and genetic factors as revealed
earlier (Gayathri and Krishnamurthy, 1981). In another study,embryo–larval and adult life stages of sea urchin were shown to beaffected by paper mill effluent containing organic solvents throughgeneration of ROS (Oakes et al., 2004). In this context, an earlier studyfrom this laboratory showed similar developmental changes in D.melanogaster exposed to an organophosphate pesticide, dichlorvos(Gupta et al., 2007). Concurrently, reproductive performance of theexposed organism also showed a poor performance with an increasein the test chemicals concentrations. Benzene, among the three testchemicals, affected the reproductive competency most followed byxylene and toluene indicating the organismal effect by thesechemicals which may be due to damage caused by increased ROSgeneration by the chemicals. However, we are unable to extrapolateuncritically the present findings to field conditions and this standsvalid only to this organism under the present experimental laboratoryconditions.
The heat shock response is induced in a protective way throughnon-specific mechanism of toxicity that involves generation ofabnormal proteins and alteration of cellular functions. As a conse-quence, maximum hsp70 induction is often related to early cellulartoxicity which is reversible (Goering et al., 1993). It has been reportedthat stress induced generation of misfolded proteins constitutes thetrigger signal that upregulates the heat shock response (Hightower,1991). Thus, it is possible that stress induced by the test chemicals inthe organism reach specific levels and enables the organism towithstand the insult.
Hsp70 expression in the organisms exposed to benzene, tolueneand xylene was found to increase in a concentration- and time-dependent manner. Damaging signals allow Hsp70 to take on newroles to provide defense in the cell. It has been reported that hsp70induction by certain environmental chemicals is generally correlatedwith early cytotoxic events and is a secondary consequence of damagesthat affect cellular integrity. The lipophilic nature of benzene, tolueneand xylene easily permit them to pass plasma membranes, alter vitalcellular functions either themselves or through metabolites beforeinteracting with cellular proteins, denaturing them and triggeringstress protein induction (Neuhaus-Steinmetz and Rensing , 1997). Apossibility that benzene, toluene and xylene are lipophilic in naturecould evoke hsp70 induction inD. melanogaster in a similarway cannotbe ruled out. The present study is consistent with the earlier reports ofWu et al. (1998) and Croute et al. (2002) showing higher Hsp70expression in the exposed workers and in cultured mammalian cellsrespectively after benzene or benzene, toluene and xylene exposure.

241M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
Interestingly, a significant regression in hsp70 expressionwas observedin the larvae treated with 100.0 mM benzene for 48 h. Toluene andxylene exposure also resulted in a regression of hsp70 expression inthe exposed organism thatwas either significant (evident frommRNA)or insignificant (evident from β-galactosidase activity). Such aregression of Hsp70 may be due to attainment of its threshold limitin the cell and is supported by earlier studies in Caenorhabditis elegansbymancozeb and in D. melanogaster by dichlorvos (Easton et al., 2001;Gupta et al., 2007).
To get an overall view of the defense system of the organism, it ispertinent to examine the role of other hsps along with Hsp70 sincescreening for one particular class of stress proteins only may notprovide a reasonably sensitive bioindicator for a wide range ofpollutants because different agents induce different families of stressproteins with different efficiencies (De Pomerai 1996; Kohler et al.,2005). Croute et al. (2002) showed a significant induction of glucoseregulated stress protein (GRP78) and an insignificant induction of hsp70and hsp90 in mammalian cell lines after benzene, toluene and xylenetreatment for 4 days suggesting increased oxidative stress leading toincreased levels of abnormal proteins in the cytoplasm. They opinedthat oxidative attack leads to the formation of electrophilic inter-mediates which are able to react with sulfhydryl group leading todepletion in reduced glutathione. However, we observed a significantinduction of hsp83, hsp60 and hsp26 in concurrence with hsp70induction in larvae of D. melanogaster exposed to these chemicals.Interestingly, these stress genes (hsp60Nhsp26≥hsp83) were signifi-cantly more induced than hsp70 (significantly regressed hsp70) inlarvae exposed to 100.0 mM benzene for 48 h indicating a steady stateof ROS generation and increased abnormal proteins and therebyinvolvement of these genes in cellular defense after chronic toxicantexposure. A similar situation may be possible for 100 mM toluene andxylene treated groups for 48 hwhere in a significant induction of hsp60,hsp83 and hsp26 in comparison to hsp70 was observed. A negativecorrelation observed between the tested hsps and proteotoxicity in theexposed organisms (YPROTEIN=−11.76XHSP70+10.36; r=−0.906, ‘t’=7.7,pb0.001; YPROTEIN=−9.299XHSP83+10.78 r=−0.965, ‘t’=13.3; YPROTEIN=−57.202XHSP26+493.12, r=0.99, ‘t’=28.3; YPROTEIN=−158.74XHSP60
+1172.7, r=0.95, ‘t’=17.7, pb0.001 respectively for benzene exposure)is further relevant in the above context.
A number of recent studies (Verma and Rana, 2008; Chen et al.,2008) documented that the toxicity of many organic compounds ismediated by reactive oxygen species and thereby induce the activitiesof anti-oxidant enzymes. In this study, significant changes in theactivities of SOD, CAT, GST, and TrxR activity and contents of GSH,MDAand PC in the test chemicals exposed organisms confirmed significantinduction of ROS by them. While MDA content and PC contentindicates the generation of ROS, significant augmentation in SOD, CATand GST activities observed in the exposed organisms, may be aresponse to abate the adverse effect of free radicals generated by thetest chemicals (Dolai et al., 2007). This is consistent with a previousstudy of Lee et al. (2006) showing significant SOD activity and ROSgeneration in the organism exposed to benzene. In another study,Georgieva et al. (2002) observed elevated oxidative stress markers viz.SOD, GPx and LPO and depletion in GSH content in occupationallyexposed petrochemical workers where benzene was found as themost hazardous substance in comparison to toluene and xylene inworkplace air. Their study and that by Parke (1996) suggested thatmetabolism of benzene leads to formation of phenol and othermetabolites thatmay have increased ROS production in theseworkers.Further, benzene metabolite, hydroquinone, may be oxidized to p-benzoquinone, a highly reactive intermediate which can bindcovalently to cellular macromolecules (DNA) or to GSH. This canlead to GSH depletion and generation of ROS and GSH depletioncontributes to oxidative damage and lipid peroxidation. The lipidperoxide radical can subsequently propagate into malondialdehyde(Georgieva et al., 2002; Parke, 1996; Snyder, 2007). Although we have
not measured the metabolites of the test chemicals a possibility thatincreased oxidative stress in the exposed organism due to interactionof the metabolites of the test chemicals with various cellularcomponents cannot be ruled out.
A significant increase in ROS generation in concurrence with asignificant increase in Hsp70 expression along with hsp83, hsp60, andhsp26 induction was observed in benzene, toluene and xylene treatedlarvae. In this context, Gillis et al. (2007) showed benzene inducedglobal gene expression in human peripheral blood mononuclear cellswith significant activation of especially Hsp90, Hsp70 and Hsp40 andcytochrome P450 family members suggesting a role of Hsp90 andHsp70 in antigen presentation by the MHC class I complexes towardscellular immunity. ROS has been reported previously to play animportant role in the regulation of gene expression by activatingtranscription factors that, in turn, mediate induction of proteinsinvolved in cellular response to environmental conditions (Villaverdeet al., 2004). That the ROS may modulate the stress gene expressionfollowing the test chemicals exposure to the organism is furtherrelevant due to significant positive correlation observed between theformer and the tested hsps (YHSP70=0.0068XROS+0.3089, r=0.834,‘t’=5.5, pb0.01; YHSP83 =0.0087XROS +0.2272, r=0.884, ‘t’=6.8,pb0.001; YHSP60=0.0445XROS+0.6523, r=0.983; ‘t’=19.2, pb0.001;YHSP26=0.0091XROS+0.2353, r=0.898, ‘t’=9.1, pb0.001 respectivelyfor benzene). Hence, a possibility that ROS generated followingchemical exposure could activate transcription factors that mayultimately trigger the tested stress genes in the exposed organismscannot be ruled out.
Among the test chemicals, benzene is the smallest and most stablearomatic hydrocarbon. Earlier studies on benzene metabolism haveidentified a number of intermediates like phenol, hydroquinone andcatechol (Medinsky et al., 1996 and Snyder, 2007). Several studies havefocused on the possibility that benzene toxicity is the result ofsynergistic interaction among metabolites (Smith et al., 1989; Irons etal., 1992; Kolachana et al., 1993; Chapman et al., 1994). Toluene andxylene too have been suggested to exert their toxicity by interactionsamong their metabolites (Shen, 1998; Khan, 2007; Wan and Winn,2008; Wetmore et al., 2008). Thus, the present observations do notexclude the possibility that these test chemicals exert their effects as aconsequence of synergistic interaction among metabolites generatedthrough their metabolism in the exposed organisms. Similarly, DMSO,used as a solvent in this study, has been shown to be either effective orineffective on the toxicity of a number of chemicals. A study by Anwaret al. (1989) showed that reduction of the clastogenic effect of benzenewas achieved by using 1.25–12.5% DMSO in the medium. In anotherstudy, Haap et al. (2008) showed very little compounding effect of thissolvent on the toxicity of diclofenac in Daphnia magna. Other studiesshowed that the solvent if used N0.5% in the treatment medium caneither exert an inhibitory effect on the toxicity of the chemicals testedin the organisms or the solvent itself could produce toxicity (Kim et al.,2007; Fossum et al., 2008; Qi et al., 2008). We used 0.3% DMSO basedon our previous study (Nazir et al., 2003). However, this does notexclude the possibility of the modulatory effect of DMSO on thetoxicity of these test chemicals which warrants further investigation.
It is pertinent to ask about the toxicity of these chemicals whenthey exist in a mixture. Though, it is beyond the scope of the presentinvestigation, it attains importance due to relevance of mixtureexposure in environment. Earlier it was shown that toluene, acompetitive inhibitor of benzene metabolism, protected male miceagainst benzene toxicity (Andrews et al., 1977). Medinsky et al., (1994)suggested that in laboratory animals and in humans, benzenemetabolism is modulated by coexposure or prior exposure to otherchemicals. Baggi (2000) showed that in toluene-degrading Pseudo-monas strain, the presence of toluene enhanced degradation ofbenzene probably due the enzymes induced by toluene and xyleneretarded both benzene and toluene degradation suggesting synergis-tic or antagonistic effects of one or other chemicals in the mixture.

242 M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
Therefore, an altogether different kind of adverse affects by the testchemicals individually and when present together remains a possi-bility and warrants further investigation.
The test chemicals individually were found to elicit suborganismalresponses in the exposed organism. However, an organismal responseat the higher dietary concentrations of the test chemicals indicates thepropagation of the suborganismal responses towards organismalresponse which is in accordance with a recent study by Siddique et al.(2007) in D. melanogaster using complex chemical mixture. We arguein favour of in vivo assay using D. melanogaster as an alternativeanimal model for generating information about the adverse effects ofenvironmental chemicals on the exposed organisms to elucidatemolecular and mechanistic studies. A pertinent question is whetherflies are less/or more sensitive to harmful substances as compared tothat observed in higher animals at cellular level. An earlier study fromthis laboratory showed that fly and higher mammals have a similardose–response relationship with four monofunctional alkylatingagents (Siddique et al., 2005). In this context, Hirsch et al. (2003)also demonstrated that fly and mammals have similar dose–responserelationship following lead exposure.
In conclusion, the study suggests that stress gene expression maybe regulated by the cellular redox conditions i.e. altered levels of ROSgeneration against the volatile organic compounds like benzene,toluene and xylene. In this context, while Hsp70 may act as an earlysensor for the transmission of redox changes in the life of cells, otherhsps can be useful components of the defense system for screening awide variety of pollutants.
Acknowledgments
The authors are grateful to the Director, IITR for his keen interest inthe present study. We thank Prof. S.C. Lakhotia, Banaras HinduUniversity, Varanasi for Bg9, and Dr. J.T. Lis, Cornell University, USA for83Z-880 and 351-94A respectively. We also thank Dr. Madhu Dixit andMr. A.L. Viswakarma, Central Drug Research Institute, Lucknow forflow cytometry facility and Mr. B.D. Bhattacharji, Indian Institute ofToxicology Research, Lucknow for editorial assistance. MPS wassupported by UGC-SRF, New Delhi (F.15-9/June 2003). Financialassistance to DKC from CSIR (NWP-34) is thankfully acknowledged.IITR communication number 2691.
References
Adams, M.D., Celniker, S.E., Holt, R.A., Evans, C.A., Gocayne, J.D., Amanatides, P.G.,Scherer, S.E., et al., 2000. The genome sequence of Drosophila melanogaster. Science24, 287, 2185–2195.
Adami, G., Larese, F., Venier, M., Barbieri, P., Lo Coco, F., Reisenhofer, E., 2006. Penetrationof benzene, toluene and xylenes contained in gasolines through human abdominalskin in vitro. Toxicol. in Vitro 20, 1321–1330.
Ahmad, I., Hamid, T., Fatima, M., Chand, H.S., Jain, S.K., Athar, M., Raisuddin, S., 2000.Induction of hepatic antioxidants in freshwater catfish (Channa punctatus Bloch)is a biomarker of paper mill effluent exposure. Biochim. Biophys. Acta 1523,37–48.
Alegretti, A.P., Thiesen, F.V., Maciel, G.P., 2004. Analytical method for evaluation ofexposure to benzene, toluene, xylene in blood by gas chromatography preceded bysolid phase microextraction. J. Chromatography B 809, 183–187.
Andrews, L.S., Lee, E.W., Witmer, C.M., Kocsis, J.J., Snyder, R., 1977. Effects of toluene onthe metabolism, disposition and hemopoietic toxicity of [3H] benzene. Biochem.Pharmacol. 26, 293–300.
Anwar, W.A., Au, W.W., Legator, M.S., Sadagopa Ramanujam, V.S., 1989. Effect ofdimethyl sulfoxide on the genotoxicity and metabolism of benzene in vivo.Carcinogenesis 10, 441–445.
Arts, M.J., Schill, R.O., Knigge, T., Eckwert, H., Kammenga, J.E., Köhler, H.R., 2004. Stressproteins (hsp70, hsp60) induced in isopods and nematodes by field exposure tometals in a gradient near Avonmouth, UK. Ecotoxicology 13, 739–1755.
ATSDR, 2005. Priority List of Hazardous Substances. Agency for Toxic Substances andDisease Registry.
Baggi, G., 2000. Ecological implication of synergistic and antagonistic interaction amonggrowth and non growth analogs present in mixture. Annal. Microbiol. 50, 103–115.
Bagchi, D., Bhattacharya, G., Stohs, S.J., 1996. In vitro and in vivo induction of heat shock(stress) protein (Hsp) gene expression by selected pesticides. Toxicology 112,57–68.
Chang, F.K., Chen, M.L., Cheng, S.F., Shih, T.S., Mao, I.F., 2007. Dermal absorption ofsolvents as a major source of exposure among shipyard spray painters. J. Occu.Environ. Med. 49, 430–436.
Chapman, D.E., Namkung, M.J., Juchau, M.R., 1994. Benzene and benzene metabolites asembryotoxic agents: effects on cultured rat embryos. Toxicol. Appl. Pharmacol. 128,129–137.
Chen, Y., McMillan, W.E., Kong, J., Israels, S.J., Gibson, S.B., 2008. Oxidative stress inducesautophagic cell death independent of apoptosis in transformed and cancer cells.Cell Death Differ. 15, 171–182.
Chou, C.C., Riviere, J.E., Monteiro-Riviere, N.A., 2003. The cytotoxicity of jet fuel aromatichydrocarbons and dose-related interleukin-8 release from human epidermalkeratinocytes. Arch. Toxicol. 77, 384–391.
Croute, F., Poinsot, J., Gaubin, Y., Beau, B., Simon, V., Murat, J.C., Soleilhavoup, J.P., 2002.Volatile organic compounds cytotoxicity and expression of HSP72, HSP90 andGRP78 stress proteins in cultured human cells. Biochim. Biophys. Acta 1591,147–155.
De Pomerai, D.I., 1996. Transgenesis in animal systems: a view from within. Sci. Christ.Belief 1, 39–60.
Derwent, R.G., Davies, T.J., Delaney, M., Dollard, G.J., Field, R.A., Dumitrean, P., Nason, P.D., Jones, B.M.R., Pepler, S.A., 2000. Analysis and interpretation of the continuoushourly monitoring data for 26 C2–C8 hydrocarbons at 12 United Kingdom sitesduring 1996. Atmos. Environ. 34, 297–312.
Dolai, S., Yadav, R.K., Datta, A.K., Adak, S., 2007. Effect of thiocyanate on the peroxidaseand psuedocatalase activities of Leishmania major ascorbate peroxidase. Biochim.Biophys. Acta 1770, 247–256.
Easton, A., Guven, K., de Pomerai, D.I., 2001. Toxicity of the dithiocarbamate fungicidemancozeb to the non target soil nematode, Caenorhabditis elegans. J. Biochem.Molec. Toxicol. 15, 15–25.
Elmann, G.L., 1959. Tissue sulfhydryl groups. Arch. Bichem. Biophys. 82, 70–77.Farzaneh, P., Allameh, A., Pratt, S., Moore, N., Travis, L., Gottschalg, E., Kind, C., Fry, J.,
2005. Increased heat shock protein 70 expression following toxicant-mediatedcytotoxicity: a ubiquitous marker of toxicant exposure? Altern. Lab Anim. 33,105–110.
Feder, J.H., Rossi, J.M., Solomon, J., Solomon, N., Lindquist, S., 1992. The consequences ofexpressing hsp70 in Drosophila cells at normal temperatures. Genes Dev. 6,1402–1413.
Festing, M.F.W., Baumans, V., Combes, D.R., Halder, M., Hendriksen, F.M., Howard, B.R.,Lovell, D.P., Moore, G.J., Overend, P., Wilson, M.S., 1998. Reducing the use oflaboratory animals in biomedical research: problems and possible solutions. Altern.Lab. Anim. 26, 283–301.
Fortunato, J.J., Feier, G., Vitali, A.M., Petronilho, F.C., Dal-Pizzol, F., Quevedo, J., 2006.Malathion-induced oxidative stress in rat brain regions. Neurochem. Res. 31, 671–678.
Fossum, F.N., Lisowski, M.J., Macey, T.A., Ingram, T.A., Morgan, M.M., 2008. Microinjec-tion of the vehicle dimethyl sulfoxide (DMSO) into the periaqueductal graymodulates morphine antinociception. Brain Res 1204, 53–58.
Franzellitti, S., Elena Fabbri, E., 2005. Differential HSP70 gene expression in theMediterranean mussel exposed to various stressors. Biochem Biophys. Res.Commun. 336, 1157–1163.
Gallegos, P., Lutz, J., Markwiese, J., Ryti, R., Mirenda, R., 2007. Wildlife ecologicalscreening levels for inhalation of volatile organic chemicals. Environ. Toxicol. Chem.26, 1299–1303.
Gayathri, M.V., Krishnamurthy, N.B., 1981. Studies on the toxicity of mercurial fungicideAgallol3 in Drosophila melanogaster. Environ. Res. 24, 89–95.
Georgieva, T., Michailova, A., Panev, T., Popov, T., 2002. Possibilities to control the healthrisk of petrochemical workers. Int. Arch. Occup. Environ. Hlth. 75, S21–S26.
Gillis, B., Gavin, I.M., Arbieva, Z., King, S.T., Jayaraman, S., Prabhakar, B.S., 2007.Identification of human cell responses to benzene and benzene metabolites.Genomics 90, 324–333.
Glaser, R.L., Lis, J.T., 1990. Multiple, compensatory regulatory elements specifyspermatocyte-specific expression of the Drosophila melanogaster hsp26. GeneMol. Cell Biol. 10, 131–137.
Goering, P.L., Fisher, B.R., Kish, C.L., 1993. Stress protein synthesis induced in rat liver bycadmium precedes hepatotoxicity. Toxicol. Appl. Pharmacol. 22, 139–148.
Gupta, S.C., Siddique, H.R., Mathur, N., Vishwakarma, A.L., Mishra, R.K., Saxena, D.K.,Kar Chowdhuri, D., 2007. Induction of hsp70, alterations in oxidative stressmarkers and apoptosis against dichlorvos exposure in transgenic Drosophilamelanogaster: modulation by reactive oxygen species. Biochim. Biophys. Acta1770, 1382–1394.
Gupta, S.C., Siddique, H.R., Saxena, D.K., Kar Chowdhuri, D., 2005. Hazardous effect oforganophosphate compound, dichlorvos in transgenic Drosophila melanogaster(hsp70-lacZ): induction of hsp70, antioxidant enzymes and inhibition of acetylcho-linesterase. Biochim. Biophys. Acta 1725, 81–92.
Haap, T., Triebskorn, R., Kohler, H.R., 2008. Acute effects of diclofenac and DMSO toDaphnia magna: immobilization and Hsp70 induction. Chemosphere 73,353–359.
Habig, W.H., Pabst, M.J., Jakoby, W.B., 1974. Glutathione S-transferase, the firstenzymatic step in mercapturic acid formation. J. Biol. Chem. 249, 7130–7139.
Hegde, K.R., Verma, S.D., 2004. Protective effects of ascorbate against oxidative stress inthe mouse lens. Biochim. Biophys. Acta 1670, 12–18.
Hightower, L.E., 1991. Heat shock, stress proteins, chaperones, and proteotoxicity. Cell66, 191–197.
Hirsch,H.V.,Mercer, J., Sambaziotis, H., Huber,M., Stark, D.T., Torno-Morley, T.,Hollocher, K.,Ghiradella,H., Ruden,D.M., 2003.Behavioral effectsof chronic exposure to low levels oflead in Drosophila melanogaster. Neurotoxicology 3, 435–442.
Holmgren, A., Bjornstedt, M., 1995. Thioredoxin and thioredoxin reductase. MethodsEnzymol. 252, 199–208.

243M.P. Singh et al. / Toxicology and Applied Pharmacology 235 (2009) 226–243
Ireland, H.E., Harding, S.J., Bonwick, G.A., Jones, M., Smith, C.J., Williams, J.H.H., 2004.Evaluation of heat shock protein 70 as a biomarker of environmental stress in Fucusserratus and Lemna minor. Biomarkers 9, 139–155.
Irons, R.D., Stillman, W.S., Colagiovanni, D.B., Henry, V.A., 1992. Synergistic action ofbenzene metabolite hydroquinone on myelopoietic stimulating activity of granu-locyte/macrophage colony-stimulating in vitro. Proc. Natl. Acad. Sci. USA 89,3691–3695.
Kanzok, S.M., Fechner, A., Baur, H., Ulschmid, J.K., Muller, H.M., Botella-Munoz, J.,Schneuwly, S., Schirmer, R.H., Becker, K., 2001. Substitution of the thioredoxinsystem for glutatione reductase in Drosophila melanogaster. Science 291, 643–646.
Kar Chowdhuri, D., Saxena, D.K., Vishwanathan, P.N., 1999. Effect of hexachlorocyclo-hexane (HCH), its isomers, and metabolites on Hsp70 expression in transgenicDrosophila melanogaster. Pestic. Biochem. Physiol. 63, 15–25.
Khan, A.H., 2007. Benzene's toxicity: a consolidated short review of human and animalstudies. Hum. Exp. Toxicol. 26, 677–685.
Kim, K.H., Kim, M.Y., 2002. The distributions of BTEX compounds in the ambientatmosphere of the Nan-Ji-Do abandoned landfill site in Seoul. Atmos. Environ. 36,2433–2446.
Kim, S.J., Jung, Y.S., Yoon, M.Y., Kim, Y.C., 2007. Comparative effects of dimethylsulfoxideon metabolism and toxicity of carbontetrachloride and dichloromethane. J. Appl.Toxicol. 27, 25–31.
Kohler, H.R., Alberti, G., Seniezak, A., 2005. Lead induced hsp70 and hsp60 patterntransformation and leg malformation during postembryonic development inorbatid mite, Archegozetes longisetosus Aoki. Comp. Biochem. Physiol. 141, 398–405.
Kohler, H.R., Knodler, C., Zanger, M., 1999. Divergent kinetics of hsp70 induction inOniscus asellus (Isopoda) in response to four environmentally relevant organicchemicals (B[a]P, PCB52, g-HCH, PCP): suitability and limits of a biomarker. Arch.Environ. Contam. Toxicol. 36, 179–185.
Kolachana, P., Subrahmanyam, V.V., Meyer, K., Zhang, L., Smith, M.,1993. Benzene and itsphenolic metabolites produce oxidative DNA damage in HL-60 cells in vitro and inthe bone marrow in vivo. Cancer Res. 53, 1023–1026.
Lakhotia, S.C., Mukherjee, T., 1980. Specific activation of puff 93D of Drosophilamelanogaster by benzamide and the effect of benzamide on the heat shock inducedpuffing activity. Chromosoma 81, 125–136.
Laubitz, D., Jankowska, A., Sikora, A., Wolinski, J., Zabielski, R., Grzesiuk, E., 2006. Gutmyoelectrical activity induces heat shock response in Escherichia coli and Caco-2cells. Exp. Physiol. 91, 867–875.
Lee, S.C., Chiu, M.Y., Ho, K.F., Zou, S.C., Wang, X., 2002. Volatile organic compounds(VOCs) in urban atmosphere of Hong Kong. Chemosphere 48, 375–382.
Lee, S.M., Lee, S.B., Park, C.H., Choj, J., 2006. Expression of heat shock protein andhemoglobin genes in Chironomus tentans (Diptera, chironomidae) larvae exposed tovarious environmental pollutants: a potential biomarker of freshwater monitoring.Chemosphere 65, 1074–1081.
Levine, R.L., Garland, D., Oliver, C.N., 1990. The determination of carbonyl content inoxidatively modified proteins. Methods Enzymol. 186, 467–477.
Lindquist, S., Craig, E.A., 1988. The heat shock proteins. Ann. Rev. Genet. 22, 631–677.Lis, J.T., Simon, J.A., Sutton, C.A., 1983. New heat shock puffs and β-galactosidase activity
resulting from transformation of Drosophila with an hsp70-lacZ hybrid gene. Cell 35,403–413.
Lowry, O.H., Rosenbrough, N.J., Farr, A.A., Randall, R.J., 1951. Protein measurement withthe folin phenol reagent. J. Biol. Chem. 193, 265–275.
Lynes, M.A., Kang, J., Sensi, S.L., Perdrized, G.A., Hightower, L.E., 2007. Heavy metalions in normal physiology, toxic stress, and cytoprotection. N.Y. Acad. Sci. 1113,159–172.
Maradonna, F., Carnevali, O., 2007. Vitellogenin, zona radiata protein, cathepsin D and heatshock protein 70 as biomarkers of exposure to xenobiotics. Biomarkers 12, 240–255.
Martınez, J., Rodrıguez-Caabeiro, F., 2005. Relationship between heat shock proteinlevels and infectivity in Trichinella spiralis larvae exposed to different stressors.Parasitol. Res. 97, 213–218.
McDermott, C., Allshire, A., Van Pelt, F., Heffron, J.J., 2008. In vitro exposure of Jurkat T-cells to industrially important organic solvents in binary combination: interactionanalysis. Toxicol. Sci. 101, 263–274.
Medinsky, M.A., Schlosser, P.M., Bond, J.A., 1994. Critical issues in benzene toxicity andmetabolism: the effect of interactions with other organic chemicals on riskassessment. Environ. Hlth. Perspect. 102, 119–124.
Medinsky, M.A., Kenyon, E.M., Seaton, M.J., Schlosser, P.M., 1996. Mechanisticconsiderations in benzene physiological model development. Environ. Hlth.Perspect. 104, 1399–1404.
Morimoto, R.I., 1993. Cells in stress: transcriptional activation of heat shock genes.Science 259, 1409–1410.
Nazir, A., Mukhopadhyay, I., Saxena, D.K., Kar Chowdhuri, D., 2001. Chlorpyrifosinduced hsp70 expression and effects on reproductive performance in transgenicDrosophila melanogaster (hsp70-lacZ) Bg9. Arch. Environ. Contam. Toxicol. 41,443–449.
Nazir, A., Mukhopadhyay, I., Saxena, D.K., Kar Chowdhuri, D., 2003. Evaluation of the Noobserved adverse effect level of solvent dimethyl sulphoxide in Drosophilamelanogaster. Toxicol. Mech. Methods 3, 147–152.
Neuhaus-Steinmetz, U., Rensing, L., 1997. Heat shock protein induction by certainchemical stressors is correlated with their cytotoxicity, lipophilicity and protein-denaturing capacity. Toxicology 123, 185–195.
Nishikimi, M., Rao, N.A., Yagi, K., 1972. The occurrence of superoxide anion in thereaction of reduced phenazine methosulphate and molecular oxygen. Biochem.Biophys. Res. Commun. 48, 849–854.
Nover, L., 1991. Heat shock proteins. In: Nover, L. (Ed.), Heat Shock Response. CRC Press,Boca Raton, FL, pp. 41–128.
O'Kane, C.J., Gehring, W.J., 1987. Detection in situ of genomic regulatory elements inDrosophila. Proc. Natl. Acad. Sci. U. S. A. 84, 9123–9127.
Oakes, K.D., McMaster, M.E., Kraak, G.J.V.D., 2004. Oxidative stress responses in longnose sucker (Catostomus catostomus) exposed to pulp and paper mill and municipalsewage effluents. Aquatic Toxicol. 67, 255–271.
Ohkawa, H., Ohishi, N., Yagi, K., 1979. Assay for lipid peroxides in animal tissues bythiobarbituric acid reaction. Anal. Biochem. 95, 351–358.
Parke, D.V., 1996. Personal reflection on 59 years of study of benzene toxicology.Environ. Hlth. Perspect. 104, 1123–1128.
PeÂrez-Serranoa, J., MartõÂneza, J., PeÂreza, B., Bernadinab, W.E., RodrõÂguez-Caabeiroa, F., 2000. In vitro shock response to different stressors in free livingand pathogenic Acanthamoeba Inter. J. Parasitol. 30, 829–835.
Pragasam,V., Kalaiselvi, P., Sumitra, K., Srinivasan, S., Anandkumar, P., Varalakshmi, P., 2006.Immunological detection of nitrosative stress-mediated modified Tamm-Horsfallglycoprotein (THP) in calcium oxalate stone formers. Biomarkers 11, 153–163.
Pratt, W.B., Toft, D.O., 2003. Regulation of signaling protein function and trafficking bythe hsp90/hsp70-based chaperone machinery. Exp. Biol. Med. 228, 111–133.
Qi, W., Ding, D., Salvi, R.J., 2008. Cytotoxic effects of dimethyl sulphoxide (DMSO) oncochlear organotypic cultures. Hear Res. 236, 52–60.
Ritossa, F., 1962. A new puffing pattern induced by temperature shock and DNP inDrosophila. Experientia 18, 571–573.
Royall, J.A., Ischiropoulos, H., 1993. Evaluation of 2′,7′-dichlorofluorescin and dihy-drorhodamine 123 as fluorescent probes for intracellular H2O2 in culturedendothelial cells. Arch. Biochem. Biophys. 302, 348–355.
Shen, Y., 1998. In vitro cytotoxicity of BTEX metabolites in HeLa cells. Arch. Environ.Contam. Toxicol. 34, 229–234.
Siddique, H.R., Gupta, S.C., Mitra, K., Murthy, R.C., Saxena, D.K., Kar Chowdhuri, D., 2007.Induction of biochemical stress markers and apoptosis in transgenic Drosophilamelanogaster against complex chemical mixtures: role of reactive oxygen species.Chem. Biol. Interact. 169, 171–188.
Siddique, H.R., Kar Chowdhuri, D., Saxena, D.K., Dhawan, A., 2005. Validation of Dro-sophila melanogaster as an in vivo model for genotoxicity assessment usingmodified alkaline Comet assay. Mutagenesis 4, 285–290.
Simoes, T., Teixeira, M.C., Fernandes, A.R., Sá-Correia, I., 2003. Adaptation of Saccharo-myces cerevisiae to the herbicide 2, 4-dichlorophenoxyacetic acid, mediated byMsn2p- and Msn4p-regulated genes: important role of SPI1. Appl. Environ.Microbiol. 69, 4019–4028.
Sinha, A.K., 1972. Colorimetric assay of catalase. Anal. Biochem. 47, 389–394.Smith, M.T., Yager, J.W., Steinmetz, K.L., Eastmond, D.A., 1989. Peroxidase-dependent
metabolism of benzene's phenolic metabolites and its potential role in benzenetoxicity and carcinogenicity. Environ. Hlth. Perspect. 82, 23–29.
Snyder, R., 2007. Benzene's toxicity: a consolidated short review of human and animalstudies by HA Khan. Hum. Exp. Toxicol. 26, 687–696.
Srivastava, A., Devotta, S., 2007. Indoor air quality of public places in Mumbai, India interms of volatile organic compounds. Environ. Monit. Assessment 133, 127–138.
Stringham, E.G., Candido, E.P.M., 1994. Transgenic hsp16-lacZ strains of the soilnematode Caenorhabditis elegans as biological monitors of environmental stress,Environ. Toxicol. Chem. 13, 1211–1220.
Timofeyev, M.A., Wiegand, C., Kent Burnison, B., Shatilina, Z.M., Pflugmacher, S.,Steinberg, C.E., 2004. Impact of natural organic matter (NOM) on freshwateramphipods. Sci. Total Environ. 319, 115–120.
US EPA, 1996. Priority Pollutants, Code of Federal Regulations. Title 40; U.S.Environmental Protection Agency. U.S. Government Printing Office, Washington,DC. Part 423, App. A (Chapter 1).
Verma, Y., Rana, S.V., 2008. Effects of progesterone on benzene toxicity in rats. Arh. Hig.Rada. Toksikol. 59, 1–9.
Villaverde, M.S., Hanzel, C.E., Verstraeten, S.V., 2004. In vitro interactions of thalliumwith components of the glutathione-dependent antioxidant defense system. FreeRadical Res. 38, 977–984.
Wan, J., Winn, L.M., 2008. In utero exposure to benzene increases embryonic c-Myb andPim-1 protein levels in CD-1 mice. Toxicol. Appl. Pharmacol. 228, 326–333.
Wetmore, B.A., Struve, M.F., Gao, P., Sharma, S., Allison, N., Roberts, K.C., Letinski, D.J.,Nicolich,M.J., Bird,M.G., Dorman,D.C., 2008.Genotoxicityof intermittent co-exposureto benzene and toluene in male CD-1 mice. Toxicol. Appl. Pharmacol. 228, 326–333.
Wu, T., Tanguay, R.M., 2006. Antibodies against heat shock proteins in environmentalstresses and diseases: friend or foe? Cell Stress Chaperones 11, 1–12.
Wu, T., Wu, Y., Yuan, Y., He, H., Zhang, G., 1998. Study on amino acid composition ofHSP70 and the level of plasma free amino acids of workers with long-termexposure to harmful factors. J. Tongji. Med. Univ. 18, 204–207.
Xiao, H., Lis, J.T., 1989. Heat shock and developmental regulation of the Drosophilamelanogaster hsp83 gene. Mol. Cell Biol. 4, 1746–1753.
Young, J.C., Agashe, V.R., Siegers, K., Fu, H., 2004. Pathways of chaperone-mediatedprotein folding in the cytosol. Nat. Rev. Mol. Cell Biol. 5, 781–791.
Zar, H.J., 1984. Biostatistical Analysis, 2nd ed. Prentice hall, Inc, Englewood Cliffs, NJ, pp.194–195.