inheritance and linkage relationships of morphological … investigated linkage relationships among...
TRANSCRIPT

Inheritance and Linkage Relationships ofMorphological and Isozyme Loci inLentil (Lens Miller)F. J. Muehlbauer, N. F. Weeden, and D. L. Hoffman
Allozyme polymorphisms for 18 loci are described and their monogenic inheritanceis demonstrated. We Investigated linkage relationships among the 18 isozyme locias well as four genes controlling morphological traits. Six small linkage groups wereidentified, containing 14 of the loci analyzed. Several of these groups appear to beconserved between Lens and Plaum, indicating that this intergeneric comparisonmay expedite genetic studies and breeding programs in both crops.
From the USDA-ARS, Washington State University,Pullman, Washington (Muehlbauer); the Departmentof Horticulture, Cornell University, Geneva, New York(Weeden); and the USDA-ARS, Aberdeen, Idaho (Hoff-man). This work is from the Agricultural Research Ser-vice, U.S. Department of Agriculture, In cooperationwtth the College of Agriculture and Home Economics,Agricultural Research Center, Washington State Uni-versity, Pullman, Washington, and the New York StateAgricultural Experiment Station, Cornell University,Geneva, New York. Address reprint requests to Dr.Muehlbauer, USDA-ARS, Washington State University,Pullman, WA 99164.
Journal of Heredity 1989-.80-.298-303; 0022-1503/89/S2.00
Lentil (Lens culinaris Medik.) is an impor-tant food legume in the Middle East, north-ern Africa, central and western Asia, andNorth and South America, where the cropis grown in semiarid regions, usually inrotation with cereals. The crop is valuedas a high-protein food and for its residues,which are used in animal feeding. Germplasm for breeding has been made avail-able from systematic collection of landraces and indigenous wild species in cen-ters of diversity. Ladizinsky et al.5 showedthat the wild species of Lens belong to twobiological species: L. culinaris and L. ni-gricans. Hybridization within L. culinaris,the species containing the cultigen L cu-linaris ssp. culinaris, and two closely re-lated subspecies, odemensis and orientalis,was easily accomplished, leading to fertileprogenies. Hybridization across the bio-logical species could be accomplished onlythrough the use of embryo rescue.3
Even though the lentil has been an im-portant food legume for centuries, geneticstudies have been made only recently.Muehlbauer and Slinkard9 reviewed thegenetics of Lens and listed 12 genes re-sponsible for morphological and seedvariation. Skibinski et al.9 identified allo-zymic variation at an aspartate amino-transferase locus which they designatedAat-1. Zamir and Ladizinsky19 describedthe inheritance of eight allozyme poly-morphisms and the presence of two link-age groups. In one of these linkage groups,the gene for green epicotyl, Gs, was placed14 map units from Got-2 (Aat-1 of Skibinskiet al.9) and 25 map units from a gene cod-ing an isozyme of malic enzyme, Me-1. Morerecently, Tadmor et al." summarized link-age relationships among 15 isozyme loci,
four loci encoding seed proteins, two mor-phological markers, and a translocationbreak point Hoffman et al.4 identified sev-eral other allozyme polymorphisms withinthe L. culinaris subspecies. We report thefurther characterization and genetic anal-ysis of these and other polymorphisms us-ing hybrid F2 populations. Four morpho-logical characters, two of which have notbeen reported in previous studies, alsowere included in our analysis.
Materials and Methods
We used the F2 populations from sevendifferent crosses for this investigation. Fourcrosses had L culinaris ssp. culinaris cv'Brewer' as the maternal parent and oneof four L. culinaris ssp. orientalis acces-sions (Nos. 4, 59, 66, and 77) as the pa-ternal parent. L culinaris ssp. culinaris cv'Giza 9' was used twice as the maternalparent in crosses with L culinaris ssp. ori-entalis No. 78 and L. culinaris ssp. ode-mensis No. 61. The seventh cross was L.culinaris ssp. culinaris cv. 'Red Chief x L.culinaris ssp. orientalis No. 78. The parentsof these crosses were known to differ forseveral morphological genes (Table 1) andseveral isozyme loci. Plants from the sev-en F2 populations were scSfe^vTsTRul^Tormorphological markers, and allozyme seg-regation was determined by means of hor-izontal starch gel electrophoresis.
We prepared samples for electropho-resis using two extraction buffers, as de-scribed by Weeden and Emmo.12 Samplesextracted in TRIS-maleate (pH 8.0) buffer(Buffer I) were placed on a pH 8.1 TRIS-citrate/lithium borate system.7 The potas-sium phosphate extraction buffer (pH 7.0)
298

Table 1. Morphological markers In parental lines of Lent
Marker genes'
Parent Yc G/i Gs Pi
'Brewer''Giza9''Red ChiefL culinaris ssp. orientalis (No. 4)L culinaris ssp. onentalis (No. 59)L culinaris ssp. onentalis (No. 77)L culinaris ssp. orientalis (No. 78)A. culinans ssp. orientalis (No. 66)i. culinaris spp. odemensis (No. 61)
1 For a description, see Muehlbauer and Slinkard.8
* + — dominance; — — recessiveness.
was used for samples placed on a pH 6.5histidine gel1 and a pH 6.1 citrate/N-(3-aminopropyl)-morpholine gel.2 Slices fromthe TRIS-citrate/lithium borate, pH 8.1, gelwere stained for aspartate aminotrans-ferase (AAT, EC 2.6.1.1), N-acetyl glucos-aminidase (NAG, EC 3.2.1.30), peroxidase(PRX, EC 1.11.1.7), leucine amino-pepti-dase (LAP, EC 3.4.11.1), alcohol dehydro-genase (ADH, EC 1.1.1.1), aldolase (ALDO,EC 4.1.2.13), /9-D-galactosidase (GAL, EC3.2.1., pH 8.5), glucosephosphate isom-erase (GPI, EC 5.3.1.9), and phospho-glucomutase (PGM, EC 2.7.5.1). Slices fromthe histidine gel were stained for fructo-kinase (FK, EC 2.7.1.4), 6-phosphogluco-nate dehydrogenase (6PGD, EC 1.1.1.44),and shikimic dehydrogenase (SKDH, EC1.1.1.25). We assayed the cathodal slide ofthe pH 6.1 gel for GAL, pH 4.5 (EC 3.2.1.23).
For most enzyme systems, we used ex-tracts from approximately 2 cm2 of young,newly expanding leaf tissue from green-house-grown plants for analysis. The leaftissue was ground in approximately 0.2 mlof extraction buffer using a Plexiglas rod.Grinding was performed in 7-ml polysty-rene weigh boats that were kept on iceduring the extraction. Extracts were usedimmediately after grinding. The green-house used to grow the plants was kept at25°C during the day and lcFC at night. Thenatural photoperiod was extended to 16 hwith supplemental cool white fluorescentlighting, which provided photosyntheti-cally active irradiance of at least 145 Wm"2 (400 to 700 mm) at pot level. Extractsfrom root samples were used to assay forNAG, PRX, and ADH. To induce ADH, potsof plants were immersed in water for 18 hat room temperature before sampling. Forthe sampling of young root tissue, the potwas removed from the water and inverted,and the pot was gently removed from theclump of soil to expose young, activelygrowing roots. Approximately 2 g of youngroot tissue was sampled for each plant. For
analysis of PRX isozymes, 2-mercapto-ethanol was omitted from the extractionbuffer.
The assays for ADH, 6PGD, and LAP wereidentical to or slight modifications of thosedescribed by Shaw and Prasad.8 The as-says we used for AAT, ALDO, GPI, PGM,and SKDH were described by Weeden andGottlieb.13 PRX was assayed according tothe method of Weeden and Marx.16 TheNAG assay contained 10 ml 0.1 M sodiumcitrate, pH 4.5, and 5 mg N-acetyl-D-glu-cosaminide. The GAL, pH 4.5, assay con-tained 10 ml 0.1 M sodium citrate, pH 4.5,and 5 mg 4-methylumbelliferyl-D-galac-toside dissolved in 1 ml dimethylformam-ide. Both solutions were mixed togetherjust before staining. The GAL, pH 8.5, assaywas similar to the GAL, pH 4.5, assay ex-cept that the buffer was 10 ml 0.1 M TRIS-HC1, pH 8.5. The NAG, GAL, pH 4.5, andGAL, pH 8.5, assays were poured directlyonto the gel slice on a glass plate. Assaysolutions were poured off after 10 min, andthe activity bands were then visualized un-der UV light (X - 302 nm).
An agarose overlay was used for the FKassay. The FK assay was prepared by mix-ing 9 ml 0.1 M TRIS-HC1, pH 7.1, and 1 ml0.1 M MgCl2. Low-gelling-temperature aga-rose (SeaPlaque, FMC, Rockland, Maine,80 mg) was added to 6 ml of buffer thatwas brought to a boil over low heat andheld in a 37°C water bath. To the remaining4 ml of buffer, 25 mg /S-D-fructose, 12 mgadenosine 5'-triphosphate, 30 units phos-phoglucose isomerase, 10 units glucose6-phosphate dehydrogenase, 5 mg nico-tinamide-adenine dinucleotide phosphate(NADP), 0.5 mg MTT, and 0.5 mg meldolablue were added. The two solutions werecombined and immediately poured ontothe gel slice on a glass plate.
The subcellular compartmentalizationof the individual isozymes of AAT, ALDO,GPI, 6PGD, PGM, and SKDH was deter-mined by the use of pollen extracts and
isolation of cell fractions greatly enrichedfor plastids or mitochondria. Pollen leach-ates were obtained from freshly collectedpollen, as described by Weeden and Gott-lieb.14 Chloroplast and mitochondrial pel-lets were obtained by differential centrif-ugation of leaf extracts on a Sorvall RC-2Brefrigerated centrifuge. Isolation of thechloroplast pellet was performed as de-scribed by Weeden and Robinson." Thesupernatant from the initial 1,000 g cen-trifugation step in the isolation of the chlo-roplast pellet was centrifuged for 2 min at5,000 g, transferred to a clean tube, andcentrifuged for 5 min at 20,000 g. The re-sulting mitochondrial pellet was resus-pended in 200 n\ of 50 mM TRIS/HC1, pH8.0, containing 14 mM 2-mercaptoethanoland 0.3% Triton X-100.
Data from the F2 plants in each of sixpopulations were tested for single-locusgoodness of fit using chi-square. Devia-tions from random assortment and recom-bination frequencies for pairs of genes weredetermined using the LINKAGE-1 programof Suiter et al.10
Results
In all seven F2 populations, segregation ofmorphological markers and isozyme phe-notypes generally fit the expected 3:1 or1:2:1 ratios (Table 2). Significant devia-tions from expected ratios were observedat several isozyme loci and one morpho-logical trait However, for each of the locithat displayed aberrant segregation withinan F2 family, ratios within the normal range(/> > .05) were obtained in at least twoother F2 families. Some partial sterility wasobserved in the Giza 9/LO61 population,possibly reflecting chromosomal rear-rangement.
The Mendelian segregation ratios shownby each of the allozyme polymorphismsindicated that each was coded by a singlenuclear gene. In some cases these resultsrepresented confirmation of previousstudies; however, other results were noveland are described in detail.
The four AAT isozymes (Figure 1) werelocated in different subcellular compart-ments, and each was encoded by a distinctlocus. The fastest migrating isozyme, AAT-1, exhibited low activity relative to the oth-er three isozymes. It was weakly associ-ated with the mitochondrial pellet and,similar to all the other isozymes, was ob-served in the supernatant. Based on pre-vious results in pea" " and Cucurbita,ls itwas postulated to be of microbody origin.The coding locus was therefore designat-
Muertbauer et al • Genetics of Lentl (Lens Mffler) 299
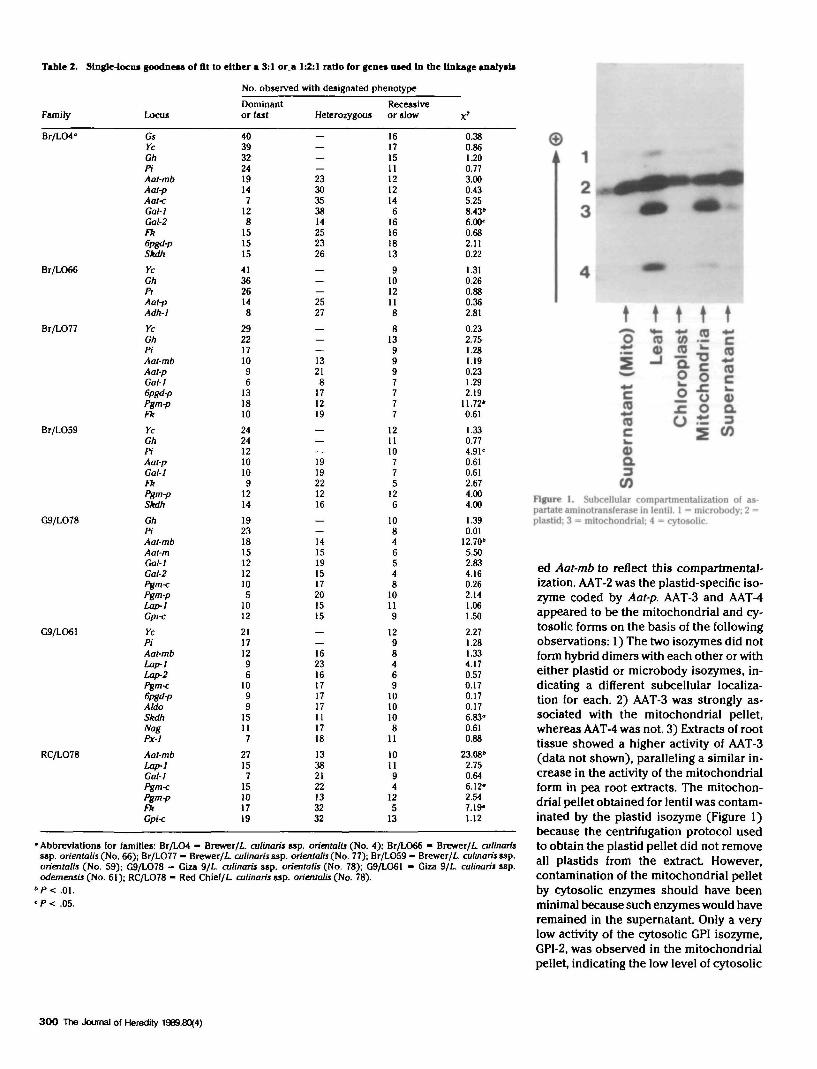
Table 2. Single-locus goodness of fit to either a 3:1 or a 1:2:1 ratio for genes nsed In the linkage analysis
No. observed with designated phenotype
Family Locus
Br/LO4°
Br/LO66
Br/LO77
Br/LO59
G9/LO78
G9/LO61
RC/LO78
GsYcGhPiAat-mbAat-pAat<Gal-IGal-2FkSpgd-PSkdh
YcGhPiAat-pAdh-I
YcGhPiAat-mbAat-pGal-1
Pgm-pFk
YcGhPiAat-pGal-1FkPgm-pSkdh
GhPiAat-mbAat-mGal-1Gal-2Pgm<Pgm-pLap-1Cpi-c
YtPiAat-mbLap-1Lap-2PgfTKSpgd-pAldoSkdhNagPx-1
Aat-mbLap-1Gal-1Pgm<Pgm-pFkGpi-c
Dominantor fast
4039322419147128151515
413626148
2922171096131810
242412101091214
1923181512121051012
21171296109915117
2715715101719
Heterozygous
——
2330353814252326
W
.,—ta21&17If
w
—1919221216^
1415191517201515.
162316171717111718
13382122133232
Recessiveor slow
16171511121214616161813
91012118
8139997777121110775126
1084654810119
1298469101010811
10119412513
X2
0J80.861.200.773.000.435.258.43s
6.00<0.682.110.22
1.310.260.880.362.81
0.232.751.281.190.231.292.1911.72'0.61
1.330.774.91C
0.610.612.674.004.00
1.390.0112.70*5.502.834.160.262.141.061.50
2.271.281.334.170.570.170.170.176.83-0.610.88
23.08*2.750.646.12-2.547.19-1.12
• Abbreviations for families: Br/LO4 - Brewer/i. ailinaris ssp. oriental!* (No. 4); Br/LO66 - Brewer/L culinarisssp. orientalis (No. 66); Br/LO77 - Brewer/L. ailinaris ssp. orientalis (No. 77); Br/LO59 - Brewer/L ailinaris ssp.orientalis (No. 59); G9/LO78 - Giza 9/L. culinaris ssp. orientalis (No. 78); G9/LO61 - Giza 9/L. culinaris ssp.odemensis (No. 61); RC/LO78 - Red Chief/L culinaris ssp. orientalis (No. 78).
*/>< .01.
<P< .05.
©4 1
t t t t t.t; Q) co CO
•s "o ~2 -1 "5. s **O O c
^ B ^ ^ " ^ * 1 ^
i O CD£ o a
Figure 1. Subcellular compartmentalization of as-partate aminotransferase in lentil. 1 = microbody; 2 =plastid; 3 = mitochondria!; 4 = cytosolic.
ed Aat-mb to reflect this compartmental-ization. AAT-2 was the plastid-specific iso-zyme coded by Aat-p. AAT-3 and AAT-4appeared to be the mitochondrial and cy-tosolic forms on the basis of the followingobservations: 1) The two isozymes did notform hybrid dimers with each other or witheither plastid or microbody isozymes, in-dicating a different subcellular localiza-tion for each. 2) AAT-3 was strongly as-sociated with the mitochondrial pellet,whereas AAT-4 was not. 3) Extracts of roottissue showed a higher activity of AAT-3(data not shown), paralleling a similar in-crease in the activity of the mitochondrialform in pea root extracts. The mitochon-drial pellet obtained for lentil was contam-inated by the plastid isozyme (Figure 1)because the centrifugation protocol usedto obtain the plastid pellet did not removeall plastids from the extract However,contamination of the mitochondrial pelletby cytosolic enzymes should have beenminimal because such enzymes would haveremained in the supernatant Only a verylow activity of the cytosolic GPI isozyme,GPI-2, was observed in the mitochondrialpellet, indicating the low level of cytosolic
3 0 0 "Rie Journal of Heredity 1969.80(4)
F H F H S S H H F H S
px-1
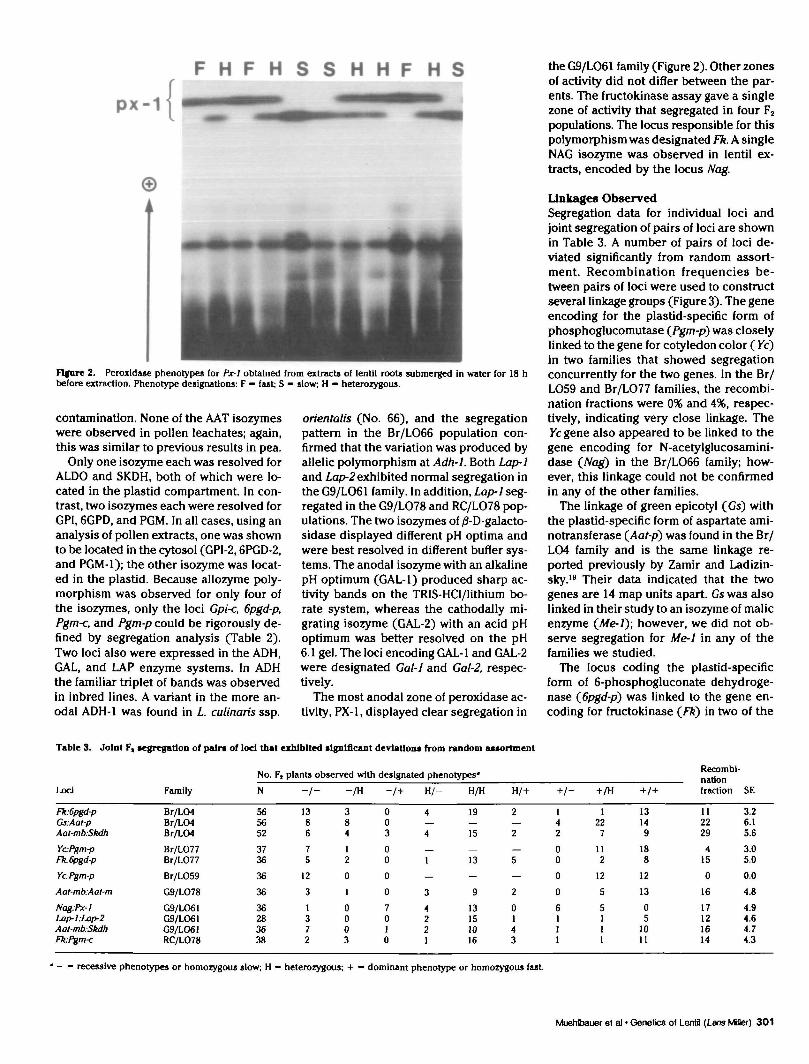
Flfore 2. Peroxldase phenotypes for Px-1 obtained from extracts of lentil roots submerged In water for 18 hbefore extraction. Phenotype designations: F - fast; S - slow; H - heterozygous.
contamination. None of the AAT isozymeswere observed in pollen leachates; again,this was similar to previous results in pea.
Only one isozyme each was resolved forALDO and SKDH, both of which were lo-cated in the plastid compartment In con-trast, two isozymes each were resolved forGPI, 6GPD, and PGM. In all cases, using ananalysis of pollen extracts, one was shownto be located in the cytosol (GPI-2,6PGD-2,and PGM-1); the other isozyme was locat-ed in the plastid. Because allozyme poly-morphism was observed for only four ofthe isozymes, only the loci Gpi-c, 6pgd-p,Pgm<, and Pgm-p could be rigorously de-fined by segregation analysis (Table 2).Two loci also were expressed in the ADH,GAL, and LAP enzyme systems. In ADHthe familiar triplet of bands was observedin inbred lines. A variant in the more an-odal ADH-1 was found in L. culinaris ssp.
orientalis (No. 66), and the segregationpattern in the Br/LO66 population con-firmed that the variation was produced byallelic polymorphism at Adh-1. Both Lap-]and Lap-2 exhibited normal segregation intheG9/LO61 family. In addition, Lap-1 seg-regated in the G9/LO78 and RC/LO78 pop-ulations. The two isozymes of /3-D-galacto-sidase displayed different pH optima andwere best resolved in different buffer sys-tems. The anodal isozyme with an alkalinepH optimum (GAL-1) produced sharp ac-tivity bands on the TRIS-HCl/lithium bo-rate system, whereas the cathodally mi-grating isozyme (GAL-2) with an acid pHoptimum was better resolved on the pH6.1 gel. The loci encoding GAL-1 and GAL-2were designated Gal-1 and Gal-2, respec-tively.
The most anodal zone of peroxidase ac-tivity, PX-1, displayed clear segregation in
the G9/LO61 family (Figure 2). Other zonesof activity did not differ between the par-ents. The fructokinase assay gave a singlezone of activity that segregated in four F2
populations. The locus responsible for thispolymorphism was designated Fk. A singleNAG isozyme was observed in lentil ex-tracts, encoded by the locus Nag.
Linkages ObservedSegregation data for individual loci andjoint segregation of pairs of loci are shownin Table 3. A number of pairs of loci de-viated significantly from random assort-ment. Recombination frequencies be-tween pairs of loci were used to constructseveral linkage groups (Figure 3). The geneencoding for the plastid-specific form ofphosphoglucomutase {Pgm-p) was closelylinked to the gene for cotyledon color ( Yc)in two families that showed segregationconcurrently for the two genes. In the Br/LO59 and Br/LO77 families, the recombi-nation fractions were 0% and 4%, respec-tively, indicating very close linkage. TheYc gene also appeared to be linked to thegene encoding for N-acetylglucosamini-dase {Nag) in the Br/LO66 family; how-ever, this linkage could not be confirmedin any of the other families.
The linkage of green epicotyl QGs) withthe plastid-specific form of aspartate ami-notransferase {Aat-p) was found in the Br/LO4 family and is the same linkage re-ported previously by Zamir and Ladizin-sky.19 Their data indicated that the twogenes are 14 map units apart. Gswas alsolinked in their study to an isozyme of malicenzyme (Me-/); however, we did not ob-serve segregation for Me-1 in any of thefamilies we studied.
The locus coding the plastid-specificform of 6-phosphogluconate dehydroge-nase (6pgd-p) was linked to the gene en-coding for fructokinase (/ft) in two of the
Table 3. Joint F, segregation of palra of lod that exhibited (Ignlflcant devlatloiu from random auortment
Loci
Fk:6pgd-pGsiAal-pAat-mb.Skdh
YcPgm-pFk.6pgd-p
YcPgm-p
Aat-mb:Aat-m
Nag.Px-1Lap-l:Lap-2Aat-mb:SkdhFkPgm<
Family
00 0
0 00
Br/LO77Br/LO77
Br/LO59
G9/LO78
G9/LO61G9/LO61G9/LO61RC/LO78
No. F,
N
565652
3736
36
36
36283638
plants observed with designated phenotypes'
- / -
1386
75
12
31372
- / H
384
12
0
1
0003
- / +
O O
CO
00
0
0
7010
H/-
4
4
1
—
3
4221
H/H
19
15
13
—
9
13151016
H/+
2
2
5
—
2
0143
+ / -
142
00
0
0
6111
+/H
122
7
112
12
5
5111
+/+
13149
00
00
12
13
05
1011
Recombi-nationfraction
112229
415
0
16
17121614
SE
3.26.15.6
3.05.0
0.0
4.8
4.94.64.74.3
1 — ~ recessive phenotypes or homozygous slow; H — heterozygous; + — dominant phenotype or homozygous fast
Muerdbauer et a) • Genetics of Lenta (Lens Mffler) 301

15±5.0
(Pgm-c)
-• Fk
•• 6pgd-p
22±6.1
-• Gs
-• Aat-p
4±3.0•• Pgm-p
-• Yc
17±4.9
-• Nag
•- Px-1
29±5.616±4.8
-• Aat-mb
•• Skdh
12±4.6
- • Lap -1
-• Lap-2
(Aat-m)Figure 3. Six linkage groups in lentil as determined by joint segregation of isozyme and morphological markers.
families. The recombination fractions were15% and 11% in the Br/LO77 and Br/LO4families, respectively. Pgm-c also dis-played linkage with Fk (Table 3) and be-longs in this linkage group. Unfortunately,none of the families had all three loci seg-regating simultaneously, and so the se-quence of the loci could not be determinedunambiguously.
The loci Aat-mb, Aat-m, and Skdh con-stitute another set of linked loci for whichthe relative sequence could not be deter-mined. Aat-mb and Skdh displayed linkagein two segregating families (Br/LO4 andG9/LO61), with recombination fractions of29% and 16%, respectively. Aat-mb andAat-m showed a 16% recombination in theG9/LO78 family.
Two small linkage groups each contain-ing two isozyme loci were identified in theG9/LO61 family (Table 3). The loci Nagand Px-1 showed a 17% recombination fre-quency, and Lap-1 and Lap-2 were ap-proximately 12 recombinant units apart
Discussion
The genetics of lentil has only recentlyreceived much attention; however, thesestudies have revealed a large number ofmorphological genes and polymorphicisozyme loci.4'619 We used several of thesegenes as well as some undescribed vari-ants in our study to identify six linkagegroups, four of which have not been re-ported previously. Comparison of the iso-zyme phenotypes and linkages among locifound in Lens with those described in thegarden pea (Pisum sativum L.) revealed anumber of parallels, providing additionalsupport for our linkage results.15-18 FourAAT isozymes have been identified in the
garden pea, including one each in the mi-crobody, the mitochondrion, the plastid,and the cytosol.1115 Because the organelleisolation was more extensively performedin the pea studies, we used the results inpea to support our subcellular assign-ments of lentil AAT isozymes. In both gen-era the plastid isozyme is the most in-tensely staining isozyme in leaf extracts,whereas the microbody isozyme is themost anodal and displays the weakest ac-tivity. The relative mobilities of the AATisozymes change slightly between the len-til and the pea. In the lentil the mitochon-drial allozymes migrated slightly slowerthan or comigrated with the plastid allo-zymes, whereas in the pea the mitochon-drial allozymes were always cathodal tothe plastid forms and one allozyme comi-grated with a cytosolic allozyme.
Other lentil enzyme systems, includingADH, ALDO, FK, GPI, 6PGD, NAG, PGM,and SKDH, are very similar to those in thepea, with homologous forms being easilyidentified.1518 Similarly, the anodal Px-1 hasits counterpart in Pisum (Figure 2C in ref-erence 16). An anodal /3-galactosidase withan alkaline pH optimum is also found inthe pea, but the pea has two cathodal iso-zymes with acid pH optima.16 We have notfound in lentil an isozyme correspondingto the Gal-3 of the pea.
The ease with which homologous iso-zyme loci can be identified between Lensand Pisum makes it possible to look forlinkage groups that may be conserved be-tween these two members of the Viceae.For instance, the locus coding the cyto-solic PGM (Pgm-c) was linked to that cod-ing the plastid 6PGD (6pgd-p) in the lentil,as it is in the pea.16 Tadmor et al.11 dem-onstrated that the locus coding the plas-tid-specific 6PGD is linked to a locus cod-
ing a peptidase. In the pea 6pgd-p alsoshows linkage to a peptidase locus.16
Other linkages in the pea parallelingthose reported here for lentil include thatbetween Aat-m and Skdhis and one be-tween Px-1 and Nag.16 Finally, in the peaLap-1 and Lap-2 are both on chromosome3 but are too far apart to display nonran-dom assortment15 The significant dispar-ity in linkage distance between the two Laploci in the two species would indicate thatthe parallel linkages are coincidental rath-er than conserved.
The clear parallel between enzyme sys-tems in the lentil and the pea provides anopportunity to establish a consistent no-menclature for isozyme loci at least in theViceae. Thus, we have used locus desig-nations in the lentil that will clearly iden-tify the homologous locus in Pisum. Wefeel that such a practice will promote in-teraction between geneticists and breed-ers working on these crops as well as onsweet pea (Lathyrus ssp.) and faba bean(Vicia faba), all members of the Viceae. Inour system the Aat-1 of Skibinski et al.9
becomes Aat-p, and the Got-1, Got-2, andGot-3 of Tadmor et al.11 are equivalent toAat-mb, Aat-p, and Aat-m, respectively. Thepracticality of this convention is particu-larly apparent in cases in which commer-cially important genes have been taggedby isozyme loci in one of the species. Forinstance, in the pea Pgm-p has been shownto be a closely linked marker for the Mogene, which confers resistance to beanyellow mosaic virus.17 Such closely linkedmarkers can be used as tags for importantgenes in breeding programs.
The formulation of detailed gene mapsshould facilitate marker-assisted selectionmethods for these crops. It may also bepossible to analyze the genetic variationpresent in germ plasm collections to syn-thesize segregating populations withunique combinations of traits.
Reference*1. Cardy BJ, Stuber CW, and Goodman MM. Techniquesfor starch gel electrophoresis of enzymes from maize(Zea maysL.'). Raleigh, North Carolina: North CarolinaState University Department of Statistics Mimeo SeriesNo. 1317. 1980.
2. Clayton JW, and Tretlak DN. Amlne-citrate buffersfor pH control in starch geJ electrophoresis. J Fish ResBd Can 1972; 29:1169-1172.
3. Cohen D, Ladizinsky G, ZW M, and Muehlbauer FJ.Rescue of interspecific Lens hybrids by means of em-bryo culture. Plant Cell Tissue Organ Culture 1984; 3:343-347.
4. Hoffman DL, Soltis DF, Muehlbauer FJ, and Ladizin-sky G. Isozyme polymorphism in Lens (Leguminosae).SystBot 1986; 11:392^02.
302 The Journal of Heredity 1969.80(4)

5. Ladizinsky G, Braun D, Goshen D, and MuehlbauerFJ. The biological species of the genus Lens. Bot Gaz1984; 145:253-261.
6. Muehlbauer FJ, and Sllnkard AE. Genetics andbreeding methodology. In. Lentils (Webb C and HawtinG, eds). London: Commonwealth Agricultural Bureau;1981*9-90.
7. Selander RK, Smith MH, Yang, SY, Johnson WE, andGentry JB. Biochemical polymorphism and systemat-ics In the genus Peromyscus. I. Variation in the old-field mouse (Peromyscus potionotus). Austin: Univer-sity of Texas Publication 7103. 1971:49-90.
8. Shaw CR, and Prasad R. Starch gel electrophoresls—a compilation of recipes. Biochem Genet 1970, 4:297-320.
9. Sklblnsld DOF, Rassol D, and Ersldne W. Aspartateamino transferase allozyme variation in a germplasmcollection of the domesticated lentil {Lens culinans).Theor Appl Genet 1984; 68:441-448.
10. Suiter KA, Wendel JF, and Case JS. J Hered 1983;74:203-204.
11. Tadmor Y, Zamir D, and Ladizinsky G. Genetic map-ping of an ancient translocation in the genus Lens.Theor Appl Genet 1987; 73:883-892.
12. Weeden NF, and Emmo AC. Isozyme characteriza-tion of Kentucky bluegrass cultivars. Can J Plant Sci1985; 65:985-994.
13. Weeden NF, and Gottlieb LD. The genetics of chlo-roplast enzymes. J Hered 1980; 71:392-396.
14. Weeden NF, and Gottlieb LD. Isolation of cyto-plasmic enzymes from pollen. Plant Physlol 1980; 66:400-403.
15. Weeden NF, and Marx GA. Chromosomal locationsof twelve isozyme loci in Pisum sativum J Hered 1984;75:365-370.
16. Weeden NF, and Marx GA. Further genetic analysisand linkage relationships of isozyme loci in the pea. JHered 1987; 78:153-159.
17. Weeden NF, Prowidenti R, and Marx GA. An iso-zyme marker for resistance to bean yellow mosaic vi-rus in Pisum sativum J Hered 1984; 75:411-412.
18. Weeden NF, and Robinson RF. Allozyme segrega-tion ratios in the interspecific cross Cucurbita maximax C ecuadorensis suggest that hybrid breakdown isnot caused by minor alteration in chromosome struc-ture. Genetics 1987; 114:593-609.
19. Zamlr D, and Ladizinsky G. Genetics of allozymevariants and linkage groups In lentil. Euphytica 1984;33:329-336.
Muenfoauer et al • Genetics of Lentil {Lens MBer) 303