investigating the impact of offshore wind farms on ... · observed numbers in sample catches...
TRANSCRIPT

1 June 2012
Investigating the impact of offshore wind
farms on European Lobster (Homarus
gammarus) and Brown Crab (Cancer
pagurus) fisheries
Daniel J. Skerritt1, Clare Fitzsimmons1, Nicholas V.C. Polunin1, Peter Berney1,
Mike H. Hardy2,
1) School of Marine Science & Technology
Newcastle University
Newcastle upon Tyne NE1 7RU
2) Northumberland Inshore Fisheries & Conservation Authority
Unit 60B South Nelson Road
Cramlington
Northumberland
NE23 1WF
Report to
The Marine Management Organisation
June 2012

2 June 2012
Please print this report on recycled paper
Report to be cited as:
Skerritt, D.J., Fitzsimmons, C., Polunin, N.V.C., Berney, P., Hardy, M.H. (2012)
Investigating the impact of offshore wind farms on European Lobster (Homarus gammarus) and
brown Crab (Cancer pagurus) fisheries. Report to the Marine Management Organisation June 2012

3 June 2012
CONTENTS Page
SUMMARY 4
INTRODUCTION AND RATIONALE 5
1.1 Introduction 5
1.2 Rationale 6
1.3 Objectives 7
1.4 Study Area 7
METHODOLOGY 10
2.1 Fieldwork 10
2.2 Site Selection 10
2.3 Data Collection 10
2.4 Field Summary 13
MODELLING METHODOLOGY 14
3.1 Model Framework 14
3.2 Model Fitting 16
3.3 Estimating Population Size 18
3.4 Effort Modelling 18
RESULTS 19
4.1 Catch Data 19
4.2 Site Comparisons 22
4.3 Mark-Recapture Data 25
4.4 Effective Effort 26
4.5 Population size 27
4.6 Movement direction 29
DISCUSSION 31
5.1 Catch characteristics 31
5.2 Additional Outputs 33
5.3 Model Outputs 33
5.3 Model Framework and future development 34
CONCLUSION 36
ACKNOWLEDGEMENTS 36
REFERENCES 37
APPENDIX 41

4 June 2012
SUMMARY
Despite growing demand for offshore wind farms, relatively little is known about their
ecological impacts upon marine benthic fauna. Once constructed they could offer protection
from activities such as trawling and provide new artificial habitats; the degree to which
fishing is displaced and shellfish population dynamics changed is unknown. UK shellfisheries
account for over 35% of total UK landings and the evidence base for possible effects needs
urgently to be expanded. In previous Environmental Statements, methods have scarcely
informed the state of the shellfishery, but to understand future impacts, adequate baseline
data prior to construction are required. This study aims to establish four small (1km2) geo-
referenced study sites and gather baseline data for the proposed Blyth offshore
demonstration wind farm site (BODS) on movement and densities and population dynamics
of European lobster (Homarus gammarus) and brown crab (Cancer pagurus).
Differences in population structure between the areas were found. A smaller population of
lobster was present within the demonstration wind farm site than at the inshore ‘control’,
and the average size of lobster there was greater than inshore. A large population of crab,
with a larger average size was also observed at the wind farm site. However, it remains
unclear whether spatial variations in the shellfish populations are influenced by habitat
differences or other physical properties, such as distance from shore, depth of water or
temperature.
Capture and recapture rates were very low within the wind farm site, which made the
population modelling unfeasible there. The inshore site produced sufficient recaptures, and
a population of 6,163 lobsters per km2 was estimated; this figure is consistent with other
large mobile decapod density estimates. Despite poor weather impacting surveys, resulting
in insufficient data to fully exploit the proposed methodology, this study has attained
replicable and reliable catch data from within and outwith the BODS, to be used as a
baseline for future studies.

5 June 2012
INTRODUCTION AND RATIONALE
1.1 Introduction
The UK leads the world in generating electricity from offshore wind farms (OWF), with more
projects in planning or construction than any other country, and increased political and
public pressure to increase production (Smith et al., 1999). Assessments of possible OWF
locations and subsequent effects of installation must be based on existing knowledge; but,
despite growing demands, relatively little is known about their ecological impacts upon
marine benthic invertebrates and the associated fisheries. Once constructed, OWFs exclude
fishers, altering both perceptions of resource availability and subsequent fishing behaviour
(Dimech et al., 2009). Turbine arrays could provide new habitat for organisms (Landers et
al., 2001; Lacroix and Pioch, 2011; Lindeboom et al., 2011), but can also cause disturbance
to birds (Garthe and Huppop, 2004), fish (Whiteley and Taylor, 1990; Vijayakumaran et al.,
2009) and mammals (Karlsson and Christiansen, 1996; Harding and Mann, 2010); but to
what degree crustacean population dynamics change remains unknown.
Shellfish are economically important throughout Europe, yet most are managed by
comparatively few regulations; UK shellfish management measures are often locally specific,
enforced under IFCA byelaws. Stocks of several shellfish species, including European lobster
(Homarus gammarus) are considered to be fully exploited, and the UK government is
seeking improved management of key shellfish resources (Bannister, 2006; Lake and Utting,
2007). The collapse of some major finfish stocks underscores the immediate importance of
managing remaining shellfish; European Lobster form the most economically valuable part
of the catch for shellfish permit holders in the Northumberland Inshore Fisheries and
Conservation Authority (NIFCA) district (Fig. 1) (Lake and Utting, 2007), with landings of 204
tonnes in 2008, valued at an estimated £2.9 million (Aebischer et al., 1993; MFA, 2007;
Cowan, 2010; Skerritt et al., 2012). Yet knowledge gaps remain and sustainable
management of this valuable resource requires sound understanding of current fishing
activity, population dynamics, accurate and replicable baseline data, and local acceptance
and compliance with regulatory measures. Newcastle University have recently reviewed
these lobster fisheries in the NIFCA district, and have begun to quantify population
dynamics and movements via mark-recapture.

6 June 2012
Population size estimation via mark-recapture (MR) models dates back more than four
decades (Jolly, 1965; Seber, 1965). The method involves estimating probabilities of capture
(ρ) and survival/fidelity (φ), from MR data to calculate the population size implied by
observed numbers in sample catches (Dunnington et al., 2005). This approach is well-suited
to studies of fisheries resources; fishing methods by their nature capture samples of the
available population and direct observation is often impossible. MR allows for population
size estimation in an open population, with estimation of both population inputs
(recruitment & immigration) and outputs (mortality & emigration) simultaneously.
Population estimates, through MR are difficult to derive for shellfish, due to high mobility of
the animals, poorly understood behaviour, and in this instance a multi-species fishery
introducing inter-specific interactions (Cancer pagurus and H. gammarus) (Miller and
Addison, 1995; Williams et al., 2009). Despite difficulties, estimates of population via MR
have been applied to decapod crustaceans (Tremblay and Smith, 2001), for example Cancer
irroratus (Hilborn, 1997), Cancer maenas (Addison, 1997), Cancer pagurus (Bell et al., 2003),
Callinectes sapidus (Fitz and Wiegert, 1992) and Homarus americanus (Cobb et al., 1997;
Dunnington et al., 2005; Bowlby et al., 2008). Most MR approaches (Jolly-Seber and related)
are conducted over long time periods (months-years), where sampling is seen as occurring
at discrete intervals, with population dynamic processes occurring between sampling
occasions (Bell et al., 2003). However, over short time periods, the use of baited traps does
not conform to the Jolly-Seber approach, as the capture process is continuous while the trap
is set (soaking), operating alongside short-term population processes.
This study aims to build upon recent work funded by the Marine Management Organisation,
Natural England and NIFCA to develop robust MR methods (Taylor, 1982; Matsuda and
Yamakawa, 1997) and gather baseline data for European lobster, H. gammarus, prior to the
installation of turbines at the National Renewable Energy Centre’s (NaREC), BODS. This will
provide a basis from which to establish if shellfish populations are altered by OWF
installations in subsequent studies.
1.2 Rationale
Together, Newcastle University and NIFCA aim to describe small-scale European lobster
population dynamics based on a short-term continuous mark-recapture sampling method

7 June 2012
developed in previous collaborations with Natural England. The collaborative nature of the
project offers further benefits to each participating organisation, contributing to NIFCA
objectives ‘to monitor fisheries in the interest of the fishery itself and of the environment’,
while enhancing NIFCA research capacity and developing Newcastle University’s ability to
support local fisheries management.
1.3 Objectives
The objectives are:
to establish geo-referenced monitoring sites within the BODS, and appropriate
control sites outside the area
to determine shellfish distributions and population dynamics within the sites
to determine European lobster movements via tagging
to map substrate hardness within studied areas
1.4 Study Area
In August 2010 NaREC was successful in progressing a proposal for the development of the
BODS to the planning stage. Located between 1.8km and 13.8km off the east coast of
Northumberland, at Blyth, this is a 99.9MW grid-connected demonstration project with a
capacity of up to 20 large-scale prototype turbines (Fig. 1-2). Three arrays consisting of five
turbines each are currently planned, at increasing distances from shore and in depths of
approximately 35, 45 and 55 metres. Turbines will be approximately 1km apart within an
array, and arrays will be 5.5km apart.
Potting within the region is restricted by the available habitat for target species and
potential conflict with other gear types, particularly trawlers. The trap fishery targets four
main species: European lobster (H. gammarus), brown crab (C. pagurus), velvet swimming
crab (Necora puber) and prawns (Nephrops norvegicus). Many fishers use an assortment of
trap type; the majority being multi-purpose, and deployed on various ground types at
different times of year to target particular species. There are ~31 vessels under 10m in
length registered at the Port of Blyth, of which 12 are potting vessels. Each has the ability to
fish up to 800 traps within 6 nm of shore, and 95% of all Blyth potting activity occurs within
the surrounding 190km2 (Turner et al., 2009).

8 June 2012
Four survey sites were identified based on the proposed location of the BODS turbines,
previous survey sites and NIFCA knowledge; BL1 (55°08’13N, 01°22’95W); BL2 (55°06’70N,
01°23’32W); BL3 (55°04’82N, 01°21’38W) and BL4 (55°07’46N, 01°26’89W) (Fig. 2). Sites
were of differing depths, habitat and distance from turbine and shore. BL1 was located
between the two inner most arrays. Identified as an area not currently heavily fished, this
site was chosen to establish population changes between the two arrays, rather than in the
immediate vicinity, as it the case with BL2; located near to the inshore array, within an area
identified by Turner et al (2009) as an area of high potting intensity. BL3 was intended as a
control site, in similar depth but situated away from the BODS. BL4 is located inshore for
comparison between the current methods used. Six sites were initially proposed, but
weather conditions and winter daylight restricted operations. MB indicates a second site
from the 2010 winter survey, which will also be used for future analysis and comparison.
Figure 1. Northumberland coastline, the NIFCA district boundaries and major fishing ports

9 June 2012
Figure 2. Map showing the location of sampling sites, turbine arrays and ports. Overlaid on map of lobster
distribution based on landings, vessel sightings and average vessel home-range (Turner et al., 2009)
Figure 3. NIFCA Fisheries Patrol Vessel, St Oswald (ZNW04), used for the duration of this project, returning to port, North Shields

10 June 2012
METHODOLOGY
2.1 Fieldwork
All surveys were conducted on the 21m NIFCA fisheries patrol vessel, St Oswald, berthed at
Royal Quays marina, North Shields (Max/Average speed 12.7/10.8 knots) (Fig. 3). This had to
maintain its primary use as a patrol vessel, so reducing the number of days at sea. Fieldwork
commenced on the 29th October and despite two days lost to unfavourable weather was
completed on 15th December 2011.
2.2 Site Selection
Four survey sites were selected within the NaREC indicative layout, using local NIFCA
knowledge to minimise interaction with other fishing gear. One was selected near a random
turbine in array 2 (BL2), one within the BODS site (BL1) and one as a control (BL3). The final
site (BL4) was introduced to ensure the rigour of the methodology by repeating a site that
had been successfully sampled and modelled the previous winter. BL1 was dominated by
‘soft’ sediment, with slightly ‘harder’ areas in the north and west of the site (Fig. 14.a);
average depth of the site was 41.3m (min 38.8m; max 44.1m). BL2 was also dominated by
‘soft’ sediment, but was less homogeneous than BL1, with ‘harder’ areas to the north-west,
and central-southern areas of the site with an average depth 35.2m (min 29.7m; max
39.8m) (Fig. 14b). Habitat at BL3 and BL4 was much ‘harder’, with extensive areas of rock and
cobble (Fig 14c & 14d, respectively); average depth was 38.9m at BL3 (min 34.2m; max
42.8m), and 27.2m at the inshore BL4 site, which also had much more variance in depth, so
forming a more complex habitat (min 16.7m; max 31.8m). The substrate distribution and
depths in the areas outside of the immediate study areas have been interpolated from
known substrate hardness at the sites.
2.3 Data Collection
Standard steel framed parlour traps 0.66m in length were used, with 27mm mesh and
selective grill on the bottom and 130mm fixed diameter, single-side entrance. Traps were
arranged in eight identical fleets of eight traps (A-H; West-East), running North to South at

11 June 2012
each site (1-8; North-South), spaced approximately 40m between traps and 100m between
fleets. Traps were baited with one frozen flatfish per trap (Fig. 4).
On setting the fleets, the vessel was lined up to predetermined fleet positions using the on-
board navigation software, with a due North bearing. Fleets were then set once the vessel
was at the correct position and released at a speed of 3.5 knots.
Figure 4. Images of the indicative trap layout at BL4 (top); one fleet of eight traps prior to being set (bottom-left),
and image of the standard parlour trap, a: length of trap (0.66m), b: width (0.46m), and c: mesh size (27mm),
baited with flatfish (bottom-right)
a
b c
Blyth
¯0 0.5 10.25 Kilometers
Legend
BL4
3_mile_limit_OSGB

12 June 2012
The study consisted of hauling all 64 traps (8 fleets of 8 traps) at three day intervals over a
two week period, so all fleets were hauled four to five times at each site (Average soak time:
3.7 days; Min: 2 days; Max: 6 days) (Fig. 5). Fleets were allowed to soak for four to five days
prior to the first sampling occasion, to generate a sample of animals for marking. Position of
fleet and water depth were recorded for each trip, as fleets and traps can move during
unfavourable weather, shooting and resetting of the fleet, and from interaction with other
sea users. In fleets of traps, catch rates are often highest in the traps at the ends of the
fleets. One possible interpretation of this observation is that the individual trapping areas
interfere with one another (Fig. 7), therefore the distance between traps in this study was
increased from 20m standard commercial distance to 40m.
On hauling, all lobster caught were tagged with a persistent T-bar tag (Hallprint Pty Ltd,
Aus.), inserted just behind the carapace, into the abdominal musculature, offset to avoid the
alimentary canal and vital organs (Fig. 6). Inserted correctly they should remain post-
ecdysis, resulting in an individual lobster being identifiable for several seasons. Each tag has
an individual ID number printed on it making it possible to construct accurate capture and
movement records for each marked lobster (Fig. 6). All captures were released, unless
seriously damaged, in which case they were recorded as removed from the experiment.
Figure 5. The date of setting and hauling of fleets at all sites during the 12 week study period

13 June 2012
Release location was at the approximate location of the trap from which they were
captured. Additionally biometric data was recorded for every individual lobster and crab;
including carapace length/width (CL/CW) respectively (rear of eye socket to base of the
carapace for lobster), sex, species, and presence of eggs, general condition, and their
capture location (site, fleet and trap number).
2.4 Field Summary
The project ran smoothly, in part due to the strength of the working relationship between
Newcastle University, NIFCA, and the local fishing community. The initial start date was
planned for summer, with two sites conducted in July, two in August and two in September,
but delays to the project start date resulted in a winter survey, which had some adverse
implications. Traps were lost on only one occasion; the fleet was re-shot with replacement
traps, minimising impact, and the data removed from future analysis. The main issue was
the cutting of the lines to fleets on sixteen occasions, probably due to sites being located
near busy shipping lanes, which added time constraints. The study consisted of 20 sampling
days split between four sites (Fig. 5). Previous catch data demonstrates that lobster
numbers are lower during winter months, consequently low recaptures will weaken the
modelling methodology and subsequent population estimates. In total 206 European
lobsters were tagged with persistent individual T-bar tags, this will enable the continued
monitoring of their movements via commercial recapture. With such low numbers at BL1,
Figure 6. Images showing lobster with T-bar tag inserted into the abdominal musculature (top left), the tagging gun
and tags (top right), and the printed tags (bottom left)

14 June 2012
BL2 and BL3 analysis of the population at each site will be difficult, but total population
numbers are high enough for strong analysis of the population demographic as a whole. A
successive study period for the summer of 2012 is planned, to make up for the shortfall in
sampling occasions.
MODELLING METHODOLOGY
3.1 Model Framework
A general model framework for MR data was defined (Bell et al., 2003), in which a cohort of
marked lobster was released. Subsequent sampling was used to recapture the survivors
(those that remain in the study area), which were then re-released at approximately the
same place (and time) of capture. Consequently, a marked individual could be captured
several times, generating a capture history (CH) (Burnham et al., 1987); the probability of a
particular CH occurring can be predicted by parameters describing capture, movement, and
survival processes between release occasions. However, Jolly-Seber methods consider the
capture process as discrete; whereas short-term trap fishing is a continuous process
(capture could occur at any point between setting and hauling). Therefore population
processes are described over shorter intervals than the time between sampling occasions
(Dunnington et al., 2005). This can be either tidal or diel intervals, depending on the
sampling intervals. Tidal is a behaviourally meaningful period (Smith et al., 1999) that also
allows for easier specification of models for unequal sampling intervals; however, as all
fleets were hauled on the same day during this study, a diel is considered sufficient.
As with all MR methods, key assumptions made were: (1) tagged individuals mix freely with
the untagged population; (2) tags remain present or the rate of tag-loss is known; (3)
capture and tagging does not alter survival or behaviour that would change capture
probability relative to untagged or non-captured individuals; (4) individuals that leave the
study area do not return to the study area (Lebreton et al., 1992); and (5) species
interactions do not affect capture rate (See 4.8).
For each tagged individual one of three observed states was recorded for each tide after
first release, to generate the CH: 0 not observed; 1 captured and released; -1 captured and

15 June 2012
removed. The 0 state was recorded if no traps were hauled that day or if the tagged lobster
was not observed on a haul occasion.
The overall probability of a particular CH occurring is the product of a series of probabilities
of the possible fate of the individual over each day following marking and first release. Given
its availability in the ‘capture area’ (area over which experimental traps exercise an influence
and the area in between and around traps from which a lobster could potentially return to
the area of influence during the study (Bell et al., 2003) (Fig. 20), three possible fates could
be defined: (1) the lobster does not enter a trap, but remains in the capture area; (2) the
lobster enters a trap and is observed; (3) the lobster does not enter a trap and permanently
emigrates from the capture area.
The respective probabilities of each fate can be described by the following three equations:
The probability that a lobster is present in the capture area but not caught between times t1
and t2 can be calculated as:
[Eq. 1]
Where ρi is probability of capture per unit of effort over tide i; φi the fidelity to the capture
area over tide i; and fij the fishing effort exerted by fleet j over tide i. ‘Fidelity to capture
area’ is defined here as remaining available for capture: not emigrating, dying or losing the
tag.
The probability a lobster enters a trap in fleet j between times t1 and t2 can be calculated as:
[Eq. 2]
Equation [1], refers to the probability of not being captured in any fleet, so the effort is
summed across all fleets , whereas equation [2] demonstrates the probability of being
captured in one particular string, j; .
The probability of a lobster permanently leaving the capture area between times t1 and t2
can be calculated as;
[Eq. 3]
2
11 1
2
11
t
ti
i
J
j
iji
t
tfpU
2
11
1
1
2
1)(
t
ti
iji
i
t
t
tfpUjV
2
11 1
1
1
2
111
t
ti
i
J
j
iji
i
t
t
tfpUW
J
j
ijf1
ijf

16 June 2012
Given values of ρi, φi and fij, the probability of any CH can be calculated, from a combination
of equations [1-3].
Each CH probability takes into account the history of setting and hauling of each fleet during
the experiment, which allows for difficulties encountered from incomplete sampling of the
sites, especially during winter conditions. The approach taken assumed that average capture
probabilities in the fleets hauled on any given occasion were consistent with average
capture probabilities across all fleets (i.e. ρij = ρi for all j).
Given pre-determined values of fishing effort, f, (see section 2.6) for each hauling occasion i
and fleet of traps j, the model framework can be used to obtain estimates of ρi and φi from
MR data. Initial values of ρ and φ were constrained to 1, as no trap effects would be
observed in the first instance. Ultimately, it is the ρ and φ values that are the fundamental
quantities from which the CH probabilities are constructed (Lebreton et al., 1992).
3.2 Model Fitting
The probabilities of each observed CH were combined with the CH frequencies to calculate
the likelihood function (likelihood of a set of parameter values given observed outcomes are
equal to the probability of those observed outcomes given those parameter values). The
goal of the model fitting was to maximize this likelihood by adjusting the survival and
recapture probabilities. Twenty-five possible models were defined for the MR data, from
the most complex model (ρsex*time, φsex*time), to the simplest possible model with parameters
constant across time and between sexes (ρ, φ). The most parsimonious model and robust
basis for inference about population size was selected by minimum value of the Akaike
Information Criterion (AIC), in its bias-adjusted form, AICc (Burnham et al., 1987), calculated
as;
[Eq. 4]
[Eq. 5]
kLLAIC 2ln2
12ln2
kn
nLLAIC
C

17 June 2012
Where lnL is the log-likelihood for the model, k is the number of separately identifiable
model parameters and n is the sample size (number of marked individuals) (Burnham &
Anderson 2002).
MR data were presented in reduced m-array format (Table 1), observed and expected m-
array totals with χ2 contributions for a goodness-of-fit (GoF) test for all suitable models to
find the most parsimonious. Because low numbers of captures and few recaptures might
have yielded low power for the GoF tests, pooling by column (grey cells) was used. Total
numbers released on each occasion included recaptured, previously marked lobster added
to the newly marked lobster caught for the first time. Expected values were generated by
multiplying the model output probability of a particular capture history by the number of
new released individuals. Degrees of freedom were calculated as number of cells that
contribute to the χ2 value, minus number of model parameters (i.e. 11 – 2 = 9), the
contribution of each pooled cell to the GoF test statistic is shown in table 1. The P value at
the bottom shows a particularly good fit of the model with the data, therefore we accept
that the model is suitable to continue with population estimates.
Table 1. Reduced m-array (Burnham et al., 1987) showing observed, expected and χ2 values for the model (ρ, φ) at the BL4 site, used for GoF testing. Each row represents first recapture for a given release cohort.
Recaptures are added to the release totals, so that multiple recaptures are pooled with first recaptures from a new release cohort.

18 June 2012
3.3 Estimating Population Size
ρ and φ allow for appropriate scaling of catch data to obtain estimates of the size of the
local population from which the catches were drawn. These catch by fleet data were used to
obtain a separate estimate of population size from each fleet on each haul, thus, treating
each fleet catch as a realisation of average fleet catches over the whole capture area; this
avoided problems of definition of capture areas for individual fleets.
To calculate the population N, of a given fleet j, over a given tide t, we used the following
formula:
[Eq. 6]
Equation 6 was used to give separate population estimates from each observation of the
catch by a fleet, thus, treating each fleet’s catch as an average of all fleet catches over the
whole capture area.
Bootstrapping was not conducted on these data in the current report, instead variation
between individual fleet hauls was considered, and produced much larger confidence
intervals (Bell et al., 2003). Because spatial heterogeneity would inflate the variance of the
overall population estimate combined over all catches, the median of all estimates was
taken to reduce the impact of outliers.
3.4 Effort Modelling
Given unequal soak times and capture probabilities defined at the temporal scale of
individual tides, it was essential to adjust the effective fishing effort applied by each fleet on
each tide of its soak. Catches are commonly assumed to have asymptotic relationships with
soak time, due to trap saturation and declines in attractiveness of bait (Briand et al., 2001):
[Eq. 6]
Where C∞ is the asymptotic catch (maximum possible catch), Ct the catch for soak time of t
and b the rate at which the increase in catch declines over time.
If the effective effort exerted by a fleet of traps j is set equal to 1 over the first tide of soak
time, the effective effort on any subsequent tide is given as:
fffp
CN
jij
tj
tj**
bteCCt
1

19 June 2012
[Eq. 7]
No independent estimates of b exist for European Lobster, so the catch data from this
experiment were used to infer a value. The approach to assume a value of C∞ and estimate
b from the relationship by simple linear regression without an intercept, for effective effort
is unlikely to introduce serious biases into the population estimates.
The approach was:
[Eq. 8]
RESULTS
4.1 Catch Data
To test for interactions between fleets and traps the difference in catch rates of total
animals was tested between outside fleets (A and H) and inside fleets (B to G), and between
end traps (1 and 8) and inside traps (3 to 6). Average total catch rates did not differ
significantly between outside and inside fleets at BL2 (t-test: Total12 = P=0.559), BL3 (t-test:
Total18 = P=0.115) or BL4 (t-test: Total13 = P=0.733), but did significantly differ at BL1 (t-test:
Total24 = P=0.028). However, to minimise effects of unusually high catches that occurred on
inside fleets at BL1, the average catch of each fleet over the whole study period was used to
test for differences between outside and inside fleets at BL1; like the other sites, there was
no significant difference (t-test: Total6 = P=0.092). No significant difference between the
average catch of end traps and middle traps was observed at any site (BL1, BL2, BL3 and BL4
t-test: Total46 = P=0.246, 0.208, 0.162, and 0.343 respectively).
b
jiijeff
,1
t
tjb
C
CLn
1
Figure 7. Trap interaction visualisations (Bell et al., 2001); a. circles represent overlap of trapping areas, of radius
r, for three baited traps; b. overlap relating to capture probability, Pcapture, to distance from trap.

20 June 2012
In total 200 individual lobsters were caught (BL1 10; BL2 43; BL3 12; BL4 135) in 19 hauls at
the four sites; 91 were male and 115 were female, (M:F =1:1.26). The size distributions of
male and female lobsters were normally distributed around ~80mm (Fig. 8a). There was a
decrease in observations at 85-95mm, when the lobsters reach legal landing size (MLS =
87mm), 74% of the lobster caught being below MLS (Fig. 9). A second size peak ~93mm was
observed, that was not as obvious in 2010. There was no significant difference in mean CL
between male and female lobsters (t-test; Sex198, P=0.70). Fewer undersized lobsters were
observed in 2011 than 2010 (Fig. 8b), but no significant difference in the size distribution of
lobster between 2010 and 2011 (t-test: Year552 P<0.05). The distributions between 80 and
90mm were precisely matched between the years. Approximately 15% of the observed
female population had eggs present (ovigerous). The average CL of ovigerous females was
89.6 mm, however 41% of the ovigerous females were under the MLS of 87mm, with the
smallest size recorded at 79mm.
0
5
10
15
20
25
30
45
50
55
60
65
70
75
80
85
90
95
10
0
10
5
11
0
11
5
Mo
re
CL (mm)
a. Male and female lobster size distribution 2011
Male = 91
Female = 115
0
2
4
6
8
10
12
14
16
35
40
45
50
55
60
65
70
75
80
85
90
95
10
0
10
5
11
0
11
5
12
0
12
5
CL (mm)
b. Population size distribution of lobster 2010/2011
3 per. Mov. Avg. (2010)
3 per. Mov. Avg. (2011)
Figure 8. a. Size distribution of all female and male lobster caught throughout the survey period, 2011. b. Size distribution of all
lobster caught during the 2011 study (Red) against the size distribution of all lobster caught in the 2010 survey, MLS of 87mm
is represented by orange vertical line.
MF
120
110
100
90
80
70
60
CL (
mm
)
Lobster Sex Distribution
Figure 9. Distribution of CL between the sexes of lobster.

21 June 2012
In total 5,736 individual brown crabs were caught throughout the duration of the study (BL1
3,596; BL2 872; BL3 461; BL4 807) in 19 hauls at the four sites; 3,950 were male and 1,786
were female (Fig. 11a; F:M =1:2.22). There is a significant difference between the CW of
male crabs and females crab (t-test, Sex2811, P<0.05), with the male mean being slightly
lower than the female; however, female distribution was slightly skewed towards the
smaller size range (Fig. 10 and 11a). There was no obvious decrease in observations as the
crab become legal to land (MLS>130mm). In 2010, most crab observed were well below the
MLS, in part due to large numbers of juvenile crab observed at a homogeneous sandy
inshore site. When the offshore sites are excluded from the 2011 population (Fig. 11b, green
line), the graph more closely resembles that of 2010 (Fig. 11b).
0 50
100 150 200 250 300 350 400 450
65
75
85
95
10
5
11
5
12
5
13
5
14
5
15
5
16
5
17
5
18
5
19
5
20
5
Mo
re
CW (mm)
a. Female and male total Crab population
Female = 1,786
Male = 3,950
0
5
10
15
20
25
30
35
0
20
40
60
80
100
120
140
160
54
63
72
81
90
99
10
8
11
7
12
6
13
5
14
4
15
3
16
2
17
1
18
0
18
9
19
8
20
7
21
6
CW (mm)
b. Population size distribution of lobster 2010/2011
3 per. Mov. Avg. (2010)
3 per. Mov. Avg. (2011)
3 per. Mov. Avg. (2011 BL4 ONLY)
Figure 11. a. Size distribution of all female and male brown crab caught throughout the survey period, 2011. b. Size distribution of
all brown crab caught during the 2011 study (Red) against the size distribution of all in the 2010 survey, and 2011 inshore site BL4
only. MLS of 130mm is represented by orange vertical line.
MF
225
200
175
150
125
100
75
50
CW
(m
m)
Crab Sex Distribution
Figure 10. Distribution of CW between the sexes of crab.

22 June 2012
4.2 Site Comparison
Significantly more lobster were caught at the inshore site BL4 (catch per unit effort (CPUE) =
7.95 ±0.51) (Fig. 13 g) in the same period as at the three offshore sites: BL3 (CPUE = 0.91
±0.02); BL2 (CPUE = 2.95 ±0.12); BL1 (CPUE = 0.86 ±0.03) (Fig. 13 a, c, e). Mean CL of lobster
differed among sites (one-way ANOVA3; P<0.05); BL1, BL2, and BL3 were not significantly
different from each other (Fisher, post-hoc) but BL4 had lower mean CL than both BL2 and
BL3 (Fisher post-hoc; -6.52, 95% confidence range for mean difference (CR)) (Fig. 12b).
Highest observed crab catches, however, were offshore: BL1 (CPUE = 309.71 ±12.17), BL2
(CPUE = 59.75 ±2.49), and BL3 (CPUE = 34.85 ±0.95) (Fig. 13b, d & f). Inshore site BL4 crabs
were negatively skewed towards the smaller sizes (Fig. 13h), rather than normally
distributed as at BL1 and BL2 (CPUE = 46.52 ±2.99).
Mean CW of crabs also differed among sites (one-way ANOVA3; P<0.05). BL4 had a
significantly lower mean CW than the other sites (Fisher’s post-hoc, -19.31 95% CR), and BL3
was slightly lower than BL2; however BL1, BL2, and BL3 had similar mean CW (Fisher post-
hoc,-4.30 95% CR) (Fig. 12a). BL1 and BL2 had the largest crab population size and CW values
(Fig. 13b, d & Fig. 14a, b). BL1 (Fig. 14a) and BL2 (Fig. 14b) were similar in substrate and
depth although BL2 had more areas of hardness, distributed to the south of the study area.
BL3 (Fig. 14c) and BL4 (Fig. 14d) had much larger proportions of rock and cobble.
BL4BL3BL2BL1
225
200
175
150
125
100
75
50
CW
(m
m)
a. Crab Size Distribution
BL4BL3BL2BL1
120
110
100
90
80
70
60
CL (
mm
)
b. Lobster Size Distribution
Figure 12. a) Size distribution of crab caught at each site. b) Size distribution of lobster caught at each site

23 June 2012
0
5
10
15
20
25
30
35
45
50
55
60
65
70
75
80
85
90
95
10
0
10
5
11
0
11
5
Mo
re
Fre
qu
en
cy
CL (mm)
c. BL2 lobster population
n = 44
0
5
10
15
20
25
30
35
45
50
55
60
65
70
75
80
85
90
95
10
0
10
5
11
0
11
5
Mo
re
Fre
qu
en
cy
CL (mm)
g. BL4 lobster population
n = 135
0
20
40
60
80
100
120
140
65
75
85
95
10
5
11
5
12
5
13
5
14
5
15
5
16
5
17
5
18
5
19
5
20
5
Mo
re
Fre
qu
en
cy
CW (mm)
f. BL3 crab population
n = 947
0
20
40
60
80
100
120
140
65
75
85
95
10
5
11
5
12
5
13
5
14
5
15
5
16
5
17
5
18
5
19
5
20
5
Mo
re
Fre
qu
en
cy
CW (mm)
d. BL2 crab population
n = 1,218
0
20
40
60
80
100
120
140
65
75
85
95
10
5
11
5
12
5
13
5
14
5
15
5
16
5
17
5
18
5
19
5
20
5
Mo
re
Fre
qu
en
cy
CW (mm)
h. BL4 crab population
n = 1,048
0
5
10
15
20
25
30
35
45
50
55
60
65
70
75
80
85
90
95
10
0
10
5
11
0
11
5
Mo
re
Fre
qu
en
cy
CL (mm)
e. BL3 lobster population
n = 12
Figure 13 a-h. Observed population size-frequency distribution of European lobster (a, c, e, g) and brown crab (b, d, f, h)
0
50
100
150
200
250
300
65
75
85
95
10
5
11
5
12
5
13
5
14
5
15
5
16
5
17
5
18
5
19
5
20
5
Mo
re
Fre
qu
en
cy
CW (mm)
b. BL1 crab population
n = 2,450
0
5
10
15
20
25
30
35 4
5
50
55
60
65
70
75
80
85
90
95
10
0
10
5
11
0
11
5
Mo
re
Fre
qu
en
cy
CL (mm)
a. BL1 lobster population
n = 10

24 June 2012
Figure 14 a-d. Screenshots of Olex data for BL1 (a), BL2 (b), BL3 (c), and BL4 (d). The substrate hardness is mapped via a colour scale
from purple (0% hardness) through blue, yellow to red (100% hardness). Inset (upper) depth transect of fleet A (westerly fleet) from
each site.
a.
.
b.
c. d.
Figure 15. Lobster population size estimates for individual fleets from model φ (), ρ (), at BL4, showing mean estimates (circles), median,
and interquartile range (box).
HGFEDCBA
12000
10000
8000
6000
4000
2000
0
Fleet
Popula
tion S
ize
25 June 2012
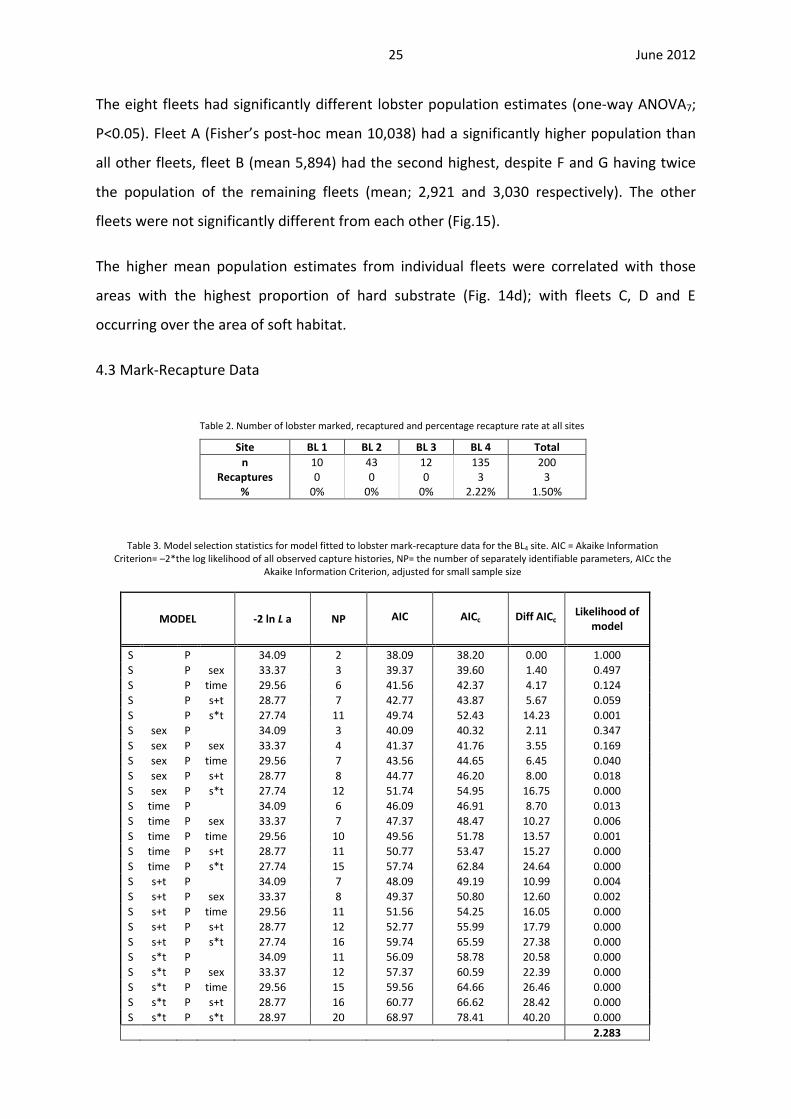
The eight fleets had significantly different lobster population estimates (one-way ANOVA7;
P<0.05). Fleet A (Fisher’s post-hoc mean 10,038) had a significantly higher population than
all other fleets, fleet B (mean 5,894) had the second highest, despite F and G having twice
the population of the remaining fleets (mean; 2,921 and 3,030 respectively). The other
fleets were not significantly different from each other (Fig.15).
The higher mean population estimates from individual fleets were correlated with those
areas with the highest proportion of hard substrate (Fig. 14d); with fleets C, D and E
occurring over the area of soft habitat.
4.3 Mark-Recapture Data
Table 2. Number of lobster marked, recaptured and percentage recapture rate at all sites
MODEL -2 ln L a NP AIC AICc Diff AICc Likelihood of
model
S
P 34.09 2 38.09 38.20 0.00 1.000
S
P sex 33.37 3 39.37 39.60 1.40 0.497
S
P time 29.56 6 41.56 42.37 4.17 0.124
S
P s+t 28.77 7 42.77 43.87 5.67 0.059
S
P s*t 27.74 11 49.74 52.43 14.23 0.001
S sex P 34.09 3 40.09 40.32 2.11 0.347
S sex P sex 33.37 4 41.37 41.76 3.55 0.169
S sex P time 29.56 7 43.56 44.65 6.45 0.040
S sex P s+t 28.77 8 44.77 46.20 8.00 0.018
S sex P s*t 27.74 12 51.74 54.95 16.75 0.000
S time P 34.09 6 46.09 46.91 8.70 0.013
S time P sex 33.37 7 47.37 48.47 10.27 0.006
S time P time 29.56 10 49.56 51.78 13.57 0.001
S time P s+t 28.77 11 50.77 53.47 15.27 0.000
S time P s*t 27.74 15 57.74 62.84 24.64 0.000
S s+t P 34.09 7 48.09 49.19 10.99 0.004
S s+t P sex 33.37 8 49.37 50.80 12.60 0.002
S s+t P time 29.56 11 51.56 54.25 16.05 0.000
S s+t P s+t 28.77 12 52.77 55.99 17.79 0.000
S s+t P s*t 27.74 16 59.74 65.59 27.38 0.000
S s*t P 34.09 11 56.09 58.78 20.58 0.000
S s*t P sex 33.37 12 57.37 60.59 22.39 0.000
S s*t P time 29.56 15 59.56 64.66 26.46 0.000
S s*t P s+t 28.77 16 60.77 66.62 28.42 0.000
S s*t P s*t 28.97 20 68.97 78.41 40.20 0.000
2.283
Table 3. Model selection statistics for model fitted to lobster mark-recapture data for the BL4 site. AIC = Akaike Information Criterion= –2*the log likelihood of all observed capture histories, NP= the number of separately identifiable parameters, AICc the
Akaike Information Criterion, adjusted for small sample size
Site BL 1 BL 2 BL 3 BL 4 Total
n 10 43 12 135 200 Recaptures 0 0 0 3 3
% 0% 0% 0% 2.22% 1.50%

26 June 2012
Recapture rates were exceptionally low; of the 200 individual lobster marked, only three
(1.5%) were recaptured. No recaptures occurred at the offshore sites; BL4 had a recapture
rate of 2.2% (Table 2). Due to the lack of recaptures, movement and fidelity to the capture
area cannot be estimated for sites BL1, BL2, and BL3.
Catch data from BL4 were used to estimate fidelity (φ) and capture (ρ). The minimum AICc
value, indicating the most parsimonious model of fidelity and capture probabilities for the
data, was for the simplest of models φ (), ρ () (Table 3). The model indicated probability of
capture and fidelity to the capture area was influenced neither by sex or time.
The probability of capture was low (ρ = 0.0036), did not change over time or between sexes.
Fidelity to the capture area also remained constant over time and between sexes, and was
very high (φ = 0.9999908).
With some reservations about heterogeneity of capture probabilities and fidelity, the model
φ (), ρ () sufficiently fits the data (GoF, BL49, χ2 = 4.73, d.f. = 9, P>0.05), and is an adequate
basis for inference about population size at BL4.
4.4 Effective Effort
As there is a lack of variation in soak time, no clear signal about the effect of soak time on
catch rates could be derived. The estimates of b show relatively narrow ranges of variation
(Fig. 16 a-d), but inferences from this relationship can only be weak. There was insufficient
information to quantify the maximum possible catch C∞, although catch data per day
declined with increased soak time and can be used to estimate C∞. The effort adjustment
was found to be relatively insensitive to choice of C∞ over a range of possible values (Bell et
al., 2003), so a value of 216 was used (C∞= highest observed n in trap+1 * number of traps
((26 +1) * 8 traps = 216)). The rate of increase in effective effort over time was calculated for
each site.

27 June 2012
4.5 Population size
Although recaptures were low at BL4, the GoF test suggests the chosen model is a good fit
for the data. From Eq. 6, using probabilities of ρ from model φ (), ρ (), and assuming that ρ
remains constant temporally, the catch data at BL4 suggest a population of about 1,730
lobsters within the 0.28 km2 capture area of the study (Fig. 17), a density of 6,163
lobsters/km2.
The large variation in the population estimate (Fig. 17) is attributable to the high catch rates
of fleets A and B (one-way ANOVA; P<0.05; Fig. 15). The wide confidence limits reflect the
high spatial variation (Fig. 18), but population estimates did not vary among occasions (one-
way ANOVA; P>0.05). CPUE show a similar pattern over time to that of population estimates
(Fig. 19), indicating that estimates may be driven largely by differences of catch rather than
ρ and φ.
The model output gives an estimate of lobster density in the 0.28 km2 area surrounding the
experimental fleets of traps (Fig. 20). Although this population is not closed, average
turnover (= 1- φ) was exceptionally low, <1% of the population emigrating during each study
day, with no difference observed between males and females. Most recaptures were also
0
2
4
6
8
10
0 5 10 15 20
Effe
ctiv
e Ef
fort
Soak Time
BL4 - b
0
2
4
6
8
10
0 5 10 15 20
Effe
ctiv
e Ef
fort
Soak Time
BL1 - b
0
2
4
6
8
10
0 5 10 15 20
Effe
ctiv
e Ef
fort
Soak Time
BL2 - b
0
2
4
6
8
10
0 5 10 15 20
Effe
ctiv
e Ef
fort
Soak Time
BL3 - b
Figure 16. Relationships between effective effort and soak time from catch data at each site, calculated from point estimate and 95% CI for b.

28 June 2012
within close proximity of their first capture site (<2km), with very few individuals migrating
greater distances.
The capture area, over which this population estimate applies, must then be calculated, to
determine the density. As boundaries of the study site are theoretical, and little information
exists on movements of European lobster, it was assumed that the population of lobster
was drawn from the area between and surrounding the traps; a minimum convex polygon
50m in diameter being assumed (Fig. 20). The minimum estimate was generated by taking
the only reported home-range estimation of H. gammarus, (Moland et al., 2011), and the
area of bait influence (Bell et al., 2001). It is unlikely the catch is drawn from a much greater
area than this, given the low population turnover and as no overlap was observed between
catches of fleets or traps (Section 4.1).
14131211109876543210
12000
10000
8000
6000
4000
2000
0
Haul Occassion
Popula
tion S
ize
6000
5000
4000
3000
2000
1000
Po
pu
lati
on
Siz
e
Figure 17. Lobster population estimates, with 95% CI quartiles displayed by the box
Figure 18. Lobster population size estimates for individual capture occasion.

29 June 2012
4.6 Movement direction
General movement vectors were inferred via tagging of individual lobsters and mapping the
vector formed between site of first capture and subsequent recapture locations, either
within the study area, or via commercial recapture. The mean linear directional movement
of all recaptures was 62o which is approximately North-East-East, but has a large variation
(0.76) and the mean movement distance was 510m. Mean directional movements for
female lobster were 91o, due East, again with large variation, and a mean length of 354m.
Male lobster mean directional movement was 41o, North East, with similar variation (0.73),
and mean distance of 633m (Fig. 21). No significant difference between distance moved
between sexes was observed (t-test: P= 0.437). Individuals recaptured during study periods
¯0 0.5 10.25 Kilometers
Legend
Pots
Capture_Area
Figure 20. Illustration of the capture area 299975 m2, formed from a minimum convex polygon of diameter 55m.
1
1.2
1.4
1.6
1.8
2
2.2
2000
2500
3000
3500
4000
4500
5000
0 5 10 15
Ave
rage
CP
UE
Po
pu
lati
on
Siz
e
Soak time
Population Size
CPUE
Figure 19. Average lobster population size for each capture occasion (square) and CPUE (diamonds)

30 June 2012
in 2010 and 2011 and from subsequent commercial sector recapture show predominantly
south-east movements from their initial release location.
DISCUSSION
5.1 Model outputs
Compared with other studies of decapod species (Eggleston et al., 1999; Dunnington et al.,
2005; Bowlby et al., 2008; Agnalt et al., 2009) our estimate of 6,163 lobsters per km2 at BL4,
is relatively low. A MR approach showed the un-fished northern fjords of Norway to hold a
population of 15,576 H. gammarus per km of shoreline (Agnalt et al., 2009). Population
estimates for H. americanus were generally higher, Rowe (2002) finding on average 10,000-
20,000 per km2, falling as low as 2,500 per km2 depending on substrate type
(Newfoundland, Canada). In contrast Dunnington et al. (2005) found 65,000 per km2 at their
summer peak via MR (Maine, US), however Bowlby (2008) found as few as 400-450 per km2
(Northumberland Strait, Canada). Previous studies conducted by Newcastle University using
similar methods to this study revealed densities of 2,359 per km2 and 9,096 per km2 for H.
gammarus at a study site ~750m South-west of BL4 and at MB (Fig.2) respectively (Skerritt et
al., 2011; Skerritt et al., 2012).
H. gammarus is perhaps more suited to MR methodologies than H. americanus, as the latter
are thought to disperse further (Smith et al., 1998; Frusher and Hoenig, 2003), increasing
turnover rate at a site and perhaps also the likelihood of overestimating populations. The
fidelity estimate (p) of 0.99 shows little or no turnover of population at the timescale
involved and one order of magnitude lower than previous years, probably due to the low
number of recaptures. Movement rates were lower than 2010 data where fidelity was 0.97
and 0.93 for Blyth and MB respectively. This would be expected during the winter months,
when activity levels of lobster are at their lowest (Smith et al., 1999; Moland et al., 2011).
Despite uncertainties, the data derived here and previously are winter estimates that are
considered to be on the low side (Skerritt et al., 2012). However, taking into account the
generally higher recorded catch rates of H. americanus, these estimates given here appear
to be reasonable, and are the first estimates of this kind for UK H. gammarus.

31 June 2012
5.2 Catch characteristics
The size distribution of total lobster was skewed towards smaller sizes (<MLS), due to
mortality and removal of larger individuals by fishing (Fig. 8a). There were fewer large male
lobster than females (Fig. 8a & 9, Q3=89mm) (Lizarraga-Cubedo et al., 2003), male C.
pagurus were also significantly smaller than females (Karlsson and Christiansen, 1996; Woll
et al., 2006; Harding and Mann, 2010). Male crab catch rates were over twice as high as
those of females throughout the study period; unlike lobster, berried crabs are rarely seen
in traps, which could account for some reduction in observations (Howard, 1982; Woll,
2003). A repeat study in the summer months would highlight any seasonal variation in sex
proportion of C. pagurus or H. gammarus. Reduced observations of undersized lobsters in
2011 compared with 2010, and a second peak around ~93mm (Fig. 7b) is attributed to a
spatial influence between inshore and offshore sites. Catch is lowest for lobster further
offshore with greater average depths. For crab, the offshore sites had higher catch rates
than inshore.
Lobsters were smaller at BL4 possibly due to interactions with high catch of crabs at offshore
sites, but more likely a difference in habitat type (Fig. 13d), (Tremblay and Smith, 2001) as
juvenile lobsters rely more heavily on habitat cover for predation avoidance than adults
(Herrnkind et al., 1997; Selgrath et al., 2007; Hovel and Wahle, 2010). The predominance of
hard habitat inshore could explain the slightly lower mean crab CW at BL3 (Figs. 13) and BL4
(Fig. 11a). Offshore sites in and around the BODS support a much larger average size of C.
pagurus and H. gammarus than inshore sites; this could be due to a natural size-led spatial
distribution, increased fishing pressure at inshore sites resulting in the removal of animals
>MLS, or the influence of habitat type.
Refuges are important to lobster, as they spend most of the day in shelter (Karnofsky et al.,
1989; Jensen et al., 1994); so structures providing this such as vegetation, rocks and
boulders can influence survival, distribution and abundance. Although numbers of lobster
caught over deeper, soft sediment are low in comparison to inshore, hard substrate, as
feeding activity of lobster might also decrease on homogeneous habitats. Instead lobsters
may use the soft habitat as a link between foraging grounds, reducing the likelihood of
observation (Addison and Bannister, 1994; Micheli and Peterson, 1999; Hanson et al., 2008).

32 June 2012
Increased knowledge of catch rates, spatial and temporal distribution of European lobster
using physical and habitat variables would benefit management. Studies conducted at small
scales, can note the shifts in distribution and abundance that patchiness of habitat
resources can produce (Eggleston et al., 1999), too large a scale and these nuances may be
overlooked.
The number of movements recorded was influenced by the variable distribution of
commercial fishing effort, and the small scale of the study site, constraining recapture
observations to within the trap array. In a very tentative way, the average net movement
was eastwards and offshore (Smith et al., 2001), with males moving on average twice as far
as females; only further research will highlight any clear patterns. Most lobsters move <1km
from first recapture location, but as yet no relationship has been found between movement,
sex, size or time (Rowe, 2001; Agnalt et al., 2007; Moland et al., 2011).
Trap saturation can impact conclusions drawn from catch data with unequal sampling time.
Therefore, it remains important that the traps were hauled regularly; a three day soak
period was sufficient for this study, but may need to be reduced for future summer surveys.
It is possible that larger individuals are more catchable than smaller (Gendron, 2005),
implying that exploitation rates are higher on larger animals and that larger animals should
therefore be rarer in areas with higher fishing pressure. Furthermore if large lobsters inhibit
smaller ones from entering traps the overall catch-rate will depend on population size
distribution and not just on overall abundance (Ihde et al., 2006).
The understanding of behaviours such as changes in catchability, feeding frequency, and
interspecific interactions are beyond the scope of this particular study, but is important in
the development of future work.
5.3 Additional Observations
As the T-bar tags should persist past ecdysis, it is possible that with the continued effort of
commercial fishermen, large scale, temporal and spatial movements may be inferred by
subsequent commercial capture. To date (01/06/2012), five tagged lobster have been
reported by commercial fishermen. Although capture-recapture only offers discrete point
data, this can still be very useful in determining movements in a cost effective manner.

33 June 2012
The presence of the black spot shell disease, the visible degradation of the shell of C.
pagurus by bacteria, was also noted throughout the study. From the total population in the
four sites at Blyth, only 2% of the population was observed to suffer from the disease; which
is very low compared with studies based in Langland Bay (Gower Peninsula, South Wales),
where high prevalence and severity of shell disease was noted in 49-61% of the population
(Vogan et al., 1999; Vogan and Rowley, 2002).
5.4 Model framework and its future development
The trapping methodology used in this study was developed by CEFAS and modified by
Newcastle University (Matsuda and Yamakawa, 1997; Bell et al., 2003; Skerritt et al., 2012).
Changes to Bell et al. (2003) included: increasing distance between traps to 40m to avoid
trap interaction; complete sampling of all fleets at the site to avoid fleets being at different
stages of their soak time and thus homogenising effort across fleets; and increasing the
number of fleets fished from four to eight, giving greater replication and statistical power.
Four key uncertainties were identified in the methodology.
(1) Value of b. Quantifying the decline in fishing effort over time is one of the weaker
aspects of the analyses, however Bell et al. (2003) found the method used here gave results
equivalent to those from more extensive methodologies. There was relatively little variation
in the relationship of effective effort with soak time (Fig. 9), but inferences about this
relationship can only be weak, as the ‘real’ value of b is impacted by numerous factors,
which will vary spatially and temporally. Generating a unique value of b for each study site,
from real data, is more suitable than finding a generic value. The estimate here extracted
from the catch data appears sound for the estimation of population size.
(2) Differences in catch rates of traps between fleets and haul occasions. Variation in catch
among traps and fleets are inevitable, but understanding micro-scale population changes,
and triggers for these changes would aid the introduction of a specific parameter in the
model for this impact.
(3) Small sample size and low recapture rates. The sparse data make it less likely that the
model will reflect any changes in catchability or fidelity, making it more likely that the

34 June 2012
simplest model would fit the data. Increasing the number of fishing occasions at each site to
increase the proportion of population tagged might increase the number of subsequent
recaptures.
(4) Estimate of capture area, and population from which catches were drawn. As
movement rates for size, sex, site and season specific are uncertain it is difficult to
accurately estimate capture area. The area of bait influence will vary between sites due to
hydrodynamics, as lobster locate bait by odour, and bottom complexity influences the
hydrodynamics of bait plumes (McLeese, 1973; Weissburg and Zimmerfaust, 1993). The
population estimate is derived via the ‘visible’ catch or individuals available to capture; for
example, lobster undergoing ecdysis or near to releasing eggs may not be captured, as
would be the case for all lobster <50mm and >150mm (Miller, 1989; Addison and Bannister,
1994; Watson et al., 2009).
The probability of a particular CH occurring can be predicted by parameters describing
capture, movement, and survival processes between release occasions. Therefore increasing
understanding of these three processes is paramount if the accuracy of MR population
estimates is to be increased, particularly those derived from the short periods mandated by
UK weather conditions. It is very difficult to increase capture rates without increasing the
fishing effort exerted, trap interaction, and cost. In any case this may not solve the issue of
subsequent recaptures; if it is assumed European lobster remain resident in an area (Smith
et al., 2001; Debuse et al., 2003; Bowlby et al., 2008), and turnover is slow, as highlighted in
this study, increasing the study period would increase subsequent recapture rates.
However, time constraints and delays to the start date of the project meant this could not
be attempted for this particular study.
CONCLUSION
This study aimed to apply MR methods to population estimates of lobster from short-term
continuous trapping data. Due to low catch rates, much analysis is based on catch data
alone. The population of lobster within the immediate BODS is low, but generally consists of
larger lobster than more heavily-fished shallow inshore sites. Crab catch rates within BODS
were generally greater than inshore sites and consisted of larger individuals, but this could
be an artefact of the inhibitive effect of the higher lobster numbers at inshore sites.

35 June 2012
Through this collaborative study with NIFCA, it was possible to identify significant
differences in population size and structure between the complex, inshore habitats and
those within the BODS. This study also produced one of the first density estimates of UK
Homarus gammarus via short-term trapping data and mark-recapture data. It also provides
replicable baseline data from within and outwith the BODS, which will inform management
of the Homarus gammarus and Cancer pagurus fishery by NIFCA. Effects of offshore wind
farm installation should be subject to on-going research.
ACKNOWLEDGEMENTS
Funding for this study came from the MMO via the Fisheries Challenge Fund. Special thanks
are due to A. Browne and the crew of the St Oswald for their commitment to the fieldwork,
and all those that helped and supported with the counting, measuring and tagging of catch.
Thanks to Mike Bell, Aileen Mill, and Steve Rushton for statistical assistance.

36 June 2012
REFERENCES
Addison, J.T. (1997) 'Lobster stock assessment: report from a workshop; I', Marine and Freshwater Research, 48(8), pp. 941-944.
Addison, J.T. and Bannister, R.C.A. (1994) 'Re-stocking and enhancement of clawed lobster stocks - A Review', Crustaceana, 67, pp. 131-155.
Aebischer, N.J., Robertson, P.A. and Kenward, R.E. (1993) 'Compositional analysis of habitat use form animalradio-tracking data', Ecology, 74(5), pp. 1313-1325.
Agnalt, A.-L., Farestveit, E., Gundersen, K., Jorstad, K.E. and Kristiansen, T.S. (2009) 'Population characteristics of the world's northernmost stocks of European lobster (Homarus gammarus) in Tysfjord and Nordfolda, northern Norway', New Zealand Journal of Marine and Freshwater Research, 43(1), pp. 47-57.
Agnalt, A.L., Kristiansen, T.S. and Jorstad, K.E. (2007) 'Growth, reproductive cycle, and movement of berried European lobsters (Homarus gammarus) in a local stock off southwestern Norway', Ices Journal of Marine Science, 64(2), pp. 288-297.
Bannister, R.C.A. (2006) 'Towards a National Development Strategy for Shellfish in England, Report for the Sea Fish Industry Authority'.
Bell, M.C., Addison, J.T. and Bannister, R.C.A. (2001) 'Estimating trapping areas from trap-catch data for lobsters and crabs', Marine and Freshwater Research, 52(8), pp. 1233-1242.
Bell, M.C., Eaton, D.R., Bannister, R.C.A. and Addison, J.T. (2003) 'A mark-recapture approach to estimating population density from continuous trapping data: application to edible crabs, Cancer pagurus, on the east coast of England', Fisheries Research, 65(1-3), pp. 361-378.
Bowlby, H.D., Hanson, J.M. and Hutchings, J.A. (2008) 'Stock structure and seasonal distribution patterns of American lobster, Homarus americanus, inferred through movement analyses', Fisheries Research, 90(1-3), pp. 279-288.
Briand, G., Matulich, S.C. and Mittelhammer, R.C. (2001) 'A catch per unit effort - soak time model for the Bristol Bay red king crab fishery, 1991-1997', Canadian Journal of Fisheries and Aquatic Sciences, 58(2), pp. 334-341.
Burnham, K.P., Anderson, D.R., White, G.C., Brownie, C. and Pollock, K.H. (1987) 'Design and analysis methods for fish survival experiments based on release-recapture', American Fisheries Society Monograph, No. 5, pp. American Fisheries Society, Bethesda, MD.
Cobb, J.S., Booth, J.D. and Clancy, M. (1997) 'Recruitment strategies in lobsters and crabs: a comparison', Marine and Freshwater Research, 48(8), pp. 797-806.
Cowan, D.F. (2010) 'Lobsters on the Move', The Lobster Newsletter, 23(1), pp. 14-17.
Debuse, V.J., Addison, J.T. and Reynolds, J.D. (2003) 'Effects of breeding site density on competition and sexual selection in the European lobster', Behavioral Ecology, 14(3), pp. 396-402.
Dimech, M., Darmanin, M., Smith, I.P., Kaiser, M.J. and Schembri, P.J. (2009) 'Fishers' perception of a 35-year old exclusive Fisheries Management Zone', Biological Conservation, 142(11), pp. 2691-2702.
Dunnington, M.J., Wahle, R.A., Bell, M.C. and Geraldi, N.R. (2005) 'Evaluating local population dynamics of the American lobster, Homarus americanus, with trap-based mark-recapture methods and seabed mapping', New Zealand Journal of Marine and Freshwater Research, 39(6), pp. 1253-1276.
Eggleston, D.B., Elis, W.E., Etherington, L.L., Dahlgren, C.P. and Posey, M.H. (1999) 'Organism responses to habitat fragmentation and diversity: Habitat colonization by estuarine macrofauna', Journal of Experimental Marine Biology and Ecology, 236(1), pp. 107-132.
Fitz, H.C. and Wiegert, R.G. (1992) 'Local populations dynamics of esturine blue crabs - Abundance, recruitment and loss', Marine Ecology-Progress Series, 87(1-2), pp. 23-40.

37 June 2012
Frusher, S.D. and Hoenig, J.M. (2003) 'Recent developments in estimating fishing and natural mortality and tag reporting rate of lobsters using multi-year tagging models', Fisheries Research, 65(1-3), pp. 379-390.
Garthe, S. and Huppop, O. (2004) 'Scaling possible adverse effects of marine wind farms on seabirds: developing and applying a vulnerability index', Journal of Applied Ecology, 41(4), pp. 724-734.
Gendron, L. (2005) 'Impact of minimum legal size increases on egg-per-recruit production, size structure, and ovigerous females in the American lobster (Homarus americanus) population off the Magdalen Islands (Quebec, Canada): a case study', New Zealand Journal of Marine and Freshwater Research, 39(3), pp. 661-674.
Hanson, K.C., Arrosa, S., Hasler, C.T., Suski, C.D., Philipp, D.P., Niezgoda, G. and Cooke, S.J. (2008) 'Effects of lunar cycles on the activity patterns and depth use of a temperate sport fish, the largemouth bass, Micropterus salmoides', Fisheries Management and Ecology, 15(5-6), pp. 357-364.
Harding, J.M. and Mann, R. (2010) 'Observations of distribution, size and sex ration of mature blue crabs, Callinectes Sapidus, from a Chesapeke Bay tributary in relation to oyster habitat and environmetnal factors', Bulletin of Marine Science, 86(1), pp. 75-91.
Herrnkind, W.F., Butler, M.J., Hunt, J.H. and Childress, M. (1997) 'Role of physical refugia: implications from a mass sponge die-off in a lobster nursery in Florida', Marine and Freshwater Research, 48(8), pp. 759-769.
Hilborn, R. (1997) 'Lobster stock assessment: report from a workshop; II', Marine and Freshwater Research, 48(8), pp. 945-947.
Hovel, K.A. and Wahle, R.A. (2010) 'Effects of habitat patchiness on American lobster movement across a gradient of predation risk and shelter competition', Ecology, 91(7), pp. 1993-2002.
Howard, A.E. (1982) 'The distribution and behaviour of ovigerous edible crabs (Cancer pagurus), and consequent sampling bias', Journal Du Conseil, 40(3), pp. 259-261.
Ihde, T.F., Frusher, S.D. and Hoenig, J.M. (2006) 'Do large rock lobsters inhibit smaller ones from entering traps? A field experiment', Marine and Freshwater Research, 57(7), pp. 665-674.
Jensen, A.C., Collins, K.J., Free, E.K. and Bannister, R.C.A. (1994) 'Lobster (Homarus Gammarus) movement on an artificial reef - The potential use of artificial reefs for stock enhancement', Crustaceana, 67, pp. 198-211.
Jolly, G.M. (1965) 'Explicit estimates from capture-recapture data with both death and immigration-stochastic model', Biometrika, 52, pp. 225-&.
Karlsson, K. and Christiansen, M.E. (1996) 'Occurrence and population composition of the edible crab (Cancer pagurus) on rocky shores of an islet on the south coast of Norway', Sarsia, 81(4), pp. 307-314.
Karnofsky, E.B., Atema, J. and Elgin, R.H. (1989) 'Field observations of social-behaviour, shelter use, and foraging in the lbster Homarus Americanus', Biological Bulletin, 176(3), pp. 239-246.
Lacroix, D. and Pioch, S. (2011) 'The multi-use in wind farm projects: more conflicts or a win-win opportunity?', Aquatic Living Resources, 24(2), pp. 129-135.
Lake, N. and Utting, S. (2007) 'English Shellfish Industry Development Strategy: 'Securing the industry's future'', Shellfish Association of Great Britain, Seafish.
Landers, D.F., Keser, M. and Saila, S.B. (2001) 'Changes in female lobster (Homarus americanus) size at maturity and implications for the lobster resource in Long Island Sound, Connecticut', Marine and Freshwater Research, 52(8), pp. 1283-1290.
Lebreton, J.D., Burnham, K.P., Clobert, J. and Anderson, D.R. (1992) 'Modeling survival and testing biological hypotheses using marked animals - a unified approach with case studies', Ecological Monographs, 62(1), pp. 67-118.

38 June 2012
Lindeboom, H.J., Kouwenhoven, H.J., Bergman, M.J.N., Bouma, S., Brasseur, S., Daan, R., Fijn, R.C., de Haan, D., Dirksen, S., van Hal, R., Lambers, R.H.R., Ter Hofstede, R., Krijgsveld, K.L., Leopold, M. and Scheidat, M. (2011) 'Short-term ecological effects of an offshore wind farm in the Dutch coastal zone; a compilation', Environmental Research Letters, 6(3).
Lizarraga-Cubedo, H.A., Tuck, I., Bailey, N., Pierce, G.J. and Kinnear, J.A.M. (2003) 'Comparisons of size at maturity and fecundity of two Scottish populations of the European lobster, Homarus gammarus', Fisheries Research, 65(1-3), pp. 137-152.
Matsuda, H. and Yamakawa, T. (1997) 'Effects of temperature on growth of the Japanese spiny lobster, Panulirus japonicus (V. Siebold) phyllosomas under laboratory conditions', Marine and Freshwater Research, 48(8), pp. 791-796.
McLeese, D.W. (1973) 'Orientation of lobsters (Homarus Americanus) to odour', Journal of the Fisheries Research Board of Canada, 30(6), pp. 838-840.
MFA (2007) 'UK Sea Fisheries Statistics 2007', Marine and Fisheries Agency, Defra, London. Available online: http://www.marinemanagement.org.uk/fisheries/statistics/documents/ukseafish/2007/final.pdf.
Micheli, F. and Peterson, C.H. (1999) 'Estuarine vegetated habitats as corridors for predator movements', Conservation Biology, 13(4), pp. 869-881.
Miller, R.J. (1989) 'Catchability of American lobsters (Homarus Americanus) and rock crabs (Cancer irroratus) by traps', Canadian Journal of Fisheries and Aquatic Sciences, 46(10), pp. 1652-1657.
Miller, R.J. and Addison, J.T. (1995) 'Trapping interactions of crabs and American lobster in laboratory tanks', Canadian Journal of Fisheries and Aquatic Sciences, 52(2), pp. 315-324.
Moland, E., Olsen, E.M., Andvord, K., Knutsen, J.A. and Stenseth, N.C. (2011) 'Home range of European lobster (Homarus gammarus) in a marine reserve: implications for future reserve design', Canadian Journal of Fisheries and Aquatic Sciences, 68(7), pp. 1197-1210.
Rowe, S. (2001) 'Movement and harvesting mortality of American lobsters (Homarus americanus) tagged inside and outside no-take reserves in Bonavista Bay, Newfoundland', Canadian Journal of Fisheries and Aquatic Sciences, 58(7), pp. 1336-1346.
Seber, G.A.F. (1965) 'A note on multiple-recapture census', Biometrika, 52, pp. 249-&.
Selgrath, J.C., Hovel, K.A. and Wahle, R.A. (2007) 'Effects of habitat edges on American lobster abundance and survival', Journal of Experimental Marine Biology and Ecology, 353(2), pp. 253-264.
Skerritt, D., Fitzsimmons, C., Polunin, N.V.C. and Scott, C. (2011) 'Establishment of local Lobster (Homarus gammarus) population', Interim Report to Natural England.
Skerritt, D.J., Scott, C., Bell, M., Polunin, N.V.C. and Fitzsimmons, C. (2012) 'Estimating European Lobster (Homarus gammarus) densities from continuous, short-term mark-recapture potting data.', Report to Natural England.
Smith, I.P., Collins, K.J. and Jensen, A.C. (1998) 'Movement and activity patterns of the European lobster, Homarus gammarus, revealed by electromagnetic telemetry', Marine Biology, 132(4), pp. 611-623.
Smith, I.P., Collins, K.J. and Jensen, A.C. (1999) 'Seasonal changes in the level and diel pattern of activity in the European lobster Homarus gammarus', Marine Ecology-Progress Series, 186, pp. 255-264.
Smith, I.P., Jensen, A.C., Collins, K.J. and Mattey, E.L. (2001) 'Movement of wild European lobsters Homarus gammarus in natural habitat', Marine Ecology-Progress Series, 222, pp. 177-186.
Taylor, E.W. (1982) 'Control and coordination of ventilation and circulation in crustaceans - responses to hypoxia and exercise', Journal of Experimental Biology, 100(OCT), pp. 289-319.
Tremblay, M.J. and Smith, S.J. (2001) 'Lobster (Homarus americanus) catchability in different habitats in late spring and early fall', Marine and Freshwater Research, 52(8), pp. 1321-1331.

39 June 2012
Turner, R.A., Hardy, M.H., Green, J. and Polunin, N.V.C. (2009) 'Defining the Northumberland Lobster Fishery', Report to the Marine and Fisheries Agency, London.
Vijayakumaran, M., Venkatesan, R., Murugan, T.S., Kumar, T.S., Jha, D.K., Remany, M.C., Thilakam, J.M.L., Jahan, S.S., Dharani, G., Kathiroli, S. and Selvan, K. (2009) 'Farming of spiny lobsters in sea cages in India', New Zealand Journal of Marine and Freshwater Research, 43(2), pp. 623-634.
Vogan, C.L., Llewellyn, P.J. and Rowley, A.F. (1999) 'Epidemiology and dynamics of shell disease in the edible crab Cancer pagurus: a preliminary study of Langland Bay, Swansea, UK', Diseases of Aquatic Organisms, 35(2), pp. 81-87.
Vogan, C.L. and Rowley, A.F. (2002) 'Dynamics of shell disease in the edible crab Cancer pagurus: a comparative study between two sites on the Gower Peninsula, South Wales, UK', Diseases of Aquatic Organisms, 52(2), pp. 151-157.
Watson, W.H., Golet, W., Scopel, D. and Jury, S. (2009) 'Use of ultrasonic telemetry to determine the area of bait influence and trapping area of American lobster, Homarus americanus, traps', New Zealand Journal of Marine and Freshwater Research, 43(1), pp. 411-418.
Weissburg, M.J. and Zimmerfaust, R.K. (1993) 'Life and death in moving fluids - Hydrodynamic effects on chemosensory-mediated predation', Ecology, 74(5), pp. 1428-1443.
Whiteley, N.M. and Taylor, E.W. (1990) 'The acid base consequences of aerial exposure in the lobster, Homarus gammarus (L) at 10-degrees-C and 20-degrees-C', Journal of Thermal Biology, 15(1), pp. 47-56.
Williams, P.J., MacSween, C. and Rossong, M. (2009) 'Competition between invasive green crab (Carcinus maenas) and American lobster (Homarus americanus)', New Zealand Journal of Marine and Freshwater Research, 43(1), pp. 29-33.
Woll, A.K. (2003) 'In situ observations of ovigerous Cancer pagurus Linnaeus, 1758 in Norwegian waters (Brachyura, Cancridae)', Crustaceana, 76, pp. 469-478.
Woll, A.K., van der Meeren, G.I. and Fossen, I. (2006) 'Spatial variation in abundance and catch composition of Cancer pagurus in Norwegian waters: biological reasoning and implications for assessment', Ices Journal of Marine Science, 63(3), pp. 421-433.

40 June 2012
APPENDIX
Table showing the total catch of lobster and crab for each fleet for each day, and
the average catch for each fleet over the whole study period

41 June 2012
Table showing the complete catch of Lobster and Crab and average catch for each trap at BL 1

42 June 2012
Table showing the complete catch of Lobster and Crab and average catch for each trap at BL2

43 June 2012
Table showing the complete catch of Lobster and Crab and average catch for each trap at BL3

44 June 2012
Table showing the complete catch of Lobster and Crab and average catch for each trap at BL4

45 June 2012
Table of approximate first and last trap coordinates (WGS ’84 UTM 30 N), for all fleets at all sites.