isolation and characterization of dna sequences that are
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, Mar. 1993, p. 1378-13840270-7306/93/031378-07$02.00/0Copyright ©) 1993, American Society for Microbiology
Isolation and Characterization of DNA Sequences That AreSpecifically Bound by Wild-Type p53 Protein
ORIT FOORD, NIR NAVOT, AND VARDA ROTTER*
Department of Cell Biology, The Weizmann Institute of Science, Rehovot, Israel 76100
Received 1 June 1992/Returned for modification 25 June 1992/Accepted 8 December 1992
Wild-type p53 was shown to function as a transcription factor. The N-terminal region of the protein containsthe transcription activation domain, while the C terminus is responsible for DNA binding. Localization of theDNA-binding domain of the p53 protein to the highly conserved carboxy-terminal region suggests that theinteraction of p53 with DNA is important for its function. We have developed a strategy for studying the DNAsequence specificity of p53-DNA binding that is based on random sequence selection. We report here on theisolation of marine genomic DNA clones that are specifically bound by the wild-type p53 protein but are notbound by mutant p53 protein forms. The isolated p53 target gene contains the unique DNA-binding sequenceGACACTGGTCACACTTGGCTGCTTAGGAAT. This fragment exhibits promoter activity as measured byits capacity to activate transcription of the chloramphenicol acetyltransferase reporter gene. Our resultssuggest that p53 directly binds DNA and functions as a typical transcription factor.
Wild-type p53 is a growth regulator that functions as asuppressor gene (32, 33, 56). Its inactivation through dele-tion or mutation plays a critical role in malignant transfor-mation, probably by allowing the cell to escape normalgrowth control (2, 4, 5, 26, 27, 32, 39, 42, 44, 60). The notionthat p53 functions as a suppressor gene was substantiated byvarious experimental approaches. Wild-type p53 expressioninhibited malignant transformation of primary embryonic ratfibroblasts induced by cotransfection with activated onco-genes (15, 16, 18). Furthermore, expression of wild-type p53induced direct growth arrest of proliferating cells in vitro (10,13, 36, 37, 50) and tumor development in vivo (8, 9, 51).The molecular mechanisms that underlie the functions of
the wild-type p53 protein are still largely unclear. Still, it wasfound that p53 is a nuclear protein, that it is spatiallyregulated during the cell cycle, and that it functions as thecell advances from the GJG1 to the S phase (12, 13, 23, 35,36, 47, 49). The wild-type p53 protein was suggested to playa direct role in DNA replication. Mouse p53 competed withDNA polymerase a in binding to the simian virus 40 (SV40)large T antigen and inhibited SV40 origin-dependent DNAreplication under in vitro conditions (8, 21, 54, 57). p53 was
also found to relocate to the same sites as known DNAreplication proteins in herpesvirus-infected cells (58). Recentstudies have shown that wild-type p53 binds directly tospecific DNA sequences such as those found in the SV40origin of replication (6) or in other genes involved in DNAreplication (29). Furthermore, p53 was shown to function as
a transcription factor (7, 17, 40, 43) and to activate theexpression of genes adjacent to a p53-DNA binding site (19,30). Mutant p53 proteins on the other hand, were inactive inthe latter two functional assays. Comparison between wild-type and mutant p53 protein forms showed variations in theirDNA-binding affinities. Wild-type p53 protein exhibits a highDNA-binding activity, whereas mutant p53 protein formsshow low DNA-binding capacities (28, 53).On the bases of the predicted structure of the p53 protein
and accumulated data, it was shown that the acidic Nterminus of the wild-type p53 contains a transcriptional
* Corresponding author.
domain (7, 17, 40, 43), whereas the basic C terminus isresponsible for DNA binding (53). p53 is a nuclear protein,and its migration to this cellular compartment is dictated bythree defined nuclear localization signals inherent in theprimary structure of the protein and clustered at its Cterminus (1, 11, 48). Once the wild-type p53 reaches the cellnucleus, it may bind to specific target genes, and subse-quently exert its transcriptional activity (52).As a step towards understanding the biochemical pathway
in which wild-type p53 protein functions in the normal cell,we conducted experiments aimed at isolating the potentialDNA target site(s) of p53. We show here that wild-type, butnot mutant, murine p53 specifically recognized a nucleotidesequence of 30-bp isolated from the murine genome. Thistarget sequence has a promoter activity, demonstrated by itscapacity to drive the expression of a reporter chloramphen-icol acetyltransferase (CAT) gene. We suggest that p53, likeother nuclear proteins, is a member of the class of sequence-specific DNA-binding factors that are involved in the regu-lation of specific target genes.
MATERIALS AND METHODS
Plasmids. p53 bacterial expression plasmids were con-structed by insertion of p53 cDNAs into the pET-8c plasmid(45). Expression of the inserted p53 cDNA is controlled bythe T7 RNA polymerase promoter. All cDNAs were clonedas NcoI-BamHI fragments into the same sites in the pET-8Cvector. The first ATG, the start codon of the native proteins,lies within the NcoI cloning site (53). pET-p53M8 andpET-pS3M11 code for full-length mutant p53 proteins, andpET-pS3cD contains a full-length wild-type p53 cDNA (3,59). pJflp53cD plasmid contains the long terminal repeat(LTR) promoter that directs the expression of wild-type p53protein (3, 52).
Synthesis of bacterial p53 proteins. Escherichia coli BL21(DE3) cells containing the different plasmids were grown at370C in Luria-Bertani medium containing ampicillin (50mg/ml). Isopropyl-p-D-thiogalactopyranoside at a final con-centration of 0.4 mM was added when the cells reached an
optical density of 1. Addition of isopropyl-o-D-thiogalacto-pyranoside to growing cultures of BL21 (DE3) lysogen
1378
Vol. 13, No. 3

p53-DNA BINDING SEQUENCES 1379
induces T7 RNA polymerase, which in turn transcribes thetarget gene in the plasmid. Three hours after the induction,cells were harvested by centrifugation, and the pellet waswashed and resuspended in Tris-EDTA containing 50 mMNaCl and 10 mg of lysozyme per ml. The lysate wassonicated three times for 15 s (each) and subjected tolow-speed centrifugation. Insoluble material containing thep53 protein was washed three times with 4 M urea-0.1 MTris-HCl (pH 8.5) and solubilized by being dissolved in 7 Mguanidine-HCI-50 mM Tris-HCI (pH 9.0)-2 mM EDTA. Thedenatured material was clarified by centrifugation and dia-lyzed against 50 mM NaCl-10 mM Tris-HCl (pH 7.8)-i mMEDTA.
Electrophoretic protein blotting and subsequent DNA hy-bridization (Southwestern (DNA-protein) blot analysis). Toprepare a crude bacterial extract, pelleted cells from late-logarithmic-phase cultures were boiled for 5 min in samplebuffer. The extracts were subjected to electrophoresis in a10% polyacrylamide-sodium dodecyl sulfate (SDS) gel. ForWestern blotting (immunoblotting) (55), proteins were trans-ferred from 10% acrylamide gels to a nitrocellulose mem-brane (Schleicher & Schuell) for 4 h at 200 mA. The blot waswashed for 2 h in standard binding buffer (10 mM Tris-HCl[pH 7.0], 50 mM NaCl, 1 mM EDTA, 0.02% bovine serumalbumin, 0.02% polyvinylpyrrolidone, 0.02% Ficoll). Hy-bridization was performed in a heat-sealable plastic bag for 3h at room temperature. In addition to the specific radiola-beled DNA probe (2 x 105 cpm/ml), salmon sperm DNA (10,ug/ml) was added as a nonspecific competitor. The blot waswashed three times with standard binding buffer for 20 minand subjected to autoradiography.DNA transfections and CAT assays. SVT-2 cells were
grown in Dulbecco modified Eagle medium-supplemented10% fetal calf serum. Ten micrograms of the various plas-mids was transfected into cells at 40% confluence by calciumphosphate coprecipitation (25). After 5 h in the presence ofthe precipitate, the cells were treated with 20% glycerol incomplete medium for 1 min. The cells were washed andmaintained in complete medium. After 48 to 72 h, extractswere prepared and assayed for CAT activity (24) by usingequivalent amounts of protein. The percent [ 4C]chloram-phenicol acetylated was quantified by eluting the acetylatedand nonacetylated species and measuring the amount ofradioactivity by scintillation counting (percent acetyl coen-zyme A conversion). Control experiments showed that as-says were in the linear range.
Sequencing of isolated clones. DNA sequencing was per-formed by the chain termination procedure by using theSequenase version 2.0 kit (USB).
Gel retardation assay. A double-stranded oligonucleotidecontaining the 30-bp p53 binding site was synthesized. It wasend labeled by using [a-32P]dATP and the Klenow fragmentof DNA polymerase. Binding reactions with this probe andnuclear extracts or renatured recombinant p53 proteins wereperformed in 0.1% Triton X-100, 4% glycerol, 1 mM EDTA,10 mM P-mercaptoethanol, 10 mM Tris-HCl (pH 7.5), and 80mM NaCl. A total of 4 to 6 ,ug of nuclear proteins was addedtogether with 2 ,ug of poly(dI-dC) and end-labeled probe(2,000 cpm per reaction). The reaction mixture was incu-bated for 15 min at 4°C and for another 15 min at roomtemperature. Samples were loaded on 4% polyacrylamidegels and subjected to electrophoresis in 0.5x Tris-borate-EDTA running buffer.DNase I protection assay. The BamHI fragment of 364 bp
was subcloned into the polylinker of the Bluescript sk+plasmid in both orientations. The fragments were released
from the plasmid by EcoRI-XbaI digestion, and labeled with[x-32P]dATP by a filling-in reaction with the Klenow en-zyme, and subsequently purified by elution from an acryla-mide gel. Labeled fragments were incubated with 2 pug ofpoly(dI-dC) and increasing amounts of nuclear extractsobtained from the pre-B 70Z/3 cell line (41). Incubation wasperformed for 15 min on ice and then for 15 min at roomtemperature. The incubation was done in a buffer containing0.1% Triton X-100, 4% glycerol, 1 mM EDTA, 10 mMP-mercaptoethanol, 10 mM Tris-HCl (pH 7.5), and 80 mMNaCl. After adding DNase I (Sigma), the reactions wereallowed to continue for 1 min and terminated by the additionof SDS and EDTA. The reaction products were separated on6% acrylamide-urea gels.
RESULTS
Isolation of genomic DNA fragments bound by wild-typep53 protein. Previous experiments showed that p53 is aDNA-binding protein (28, 53). We found that the C terminusof the wild-type p53 protein is responsible for DNA binding.Those experiments were performed with the wild-type p53expressed in a bacterial expression vector (53). The presentexperiments were aimed at isolating specific genomic DNAfragments that are specifically bound by wild-type p53 pro-tein. For this purpose a high-capacity immunoaffinity matrixwas prepared by binding PAb-421, a monoclonal antibodydirected against the C terminus of the p53 antigen, to anAffi-prep 10 matrix (Bio-Rad). The use of a monoclonalantibody rather than a polyclonal serum means that only asingle type of antibody-antigen interaction must be disruptedto displace the antigen from the column and ensures a moreefficient elution.
Wild-type p53 protein purified from bacterial cells wasloaded on a 1-ml PAb-421 immunoaffinity column. Thecolumn was then washed extensively, until the effluent'sabsorption at 280 nm approached background levels. Totalgenomic DNA extracted from mouse fibroblasts and di-gested with the BamHI-HindIII restriction enzymes wasthen loaded on the column to form a p53 protein-DNAcomplex. The column was then washed with 100 volumes ofTE to deplete nonspecific DNA. p53-DNA complexes werereleased from the PAb-421 column by using an NH4 solutionat pH 11.5. DNA fragments were extracted from the eluentwith phenol-chloroform, precipitated with ethanol, and li-gated to the Bluescript vector linearized by the HindIIl-BamHI restriction enzymes. The ligation mixture was usedto transform the XL1 Blue strain of E. coli. Twenty-fourindependent clones were obtained. Digestion of theseclones, designated MgBH 1 to 24, with HindIII-BamHIrestriction enzymes released inserts of various sizes (Fig. 1).
Binding of MgBH fragments to wild-type p53 protein. Tofurther characterize the isolated genomic DNA fragmentscontaining the p53 binding sites, we compared their bindingaffinities with those of wild-type and mutant p53 proteins.Previous studies showed that wild-type and mutant p53protein forms vary in their DNA-binding affinities (28, 53).Bacterial cell extracts expressing mutant or wild-type p53were boiled in sample buffer, subjected to electrophoresis inan SDS-polyacrylamide gel, and transferred to nitrocellu-lose. The blots were incubated with the various isolatedradiolabeled MgBH double-stranded DNA fragments. Afterincubation, the filters were washed to remove unbound DNAand subjected to autoradiography. Figure 2 shows thatinserts of clones MgBH4, MgBH5, and MgBH6 were spe-cifically bound by the wild-type p53 protein (pET-p53cD) but
VOL. 13, 1993

1380 FOORD ET AL. ML EL IL
H
22 32
17
18
261 42019
qHE
H C s B___ ___ __ B
H B B
H SBH BH B
HBH B
H BH S ~~~BH SX
H B)HS S
FIG. 1. Restriction maps of MgBH clones. The various MgBHclones were digested by the BamHI-HindIII restriction enzymes. Apartial restriction map of the different inserts is illustrated. H,HindIII; 5, SacI; B, BamHI; E, EcoRI; C, ClaI. In some of theclones the BamHI site was lost, as indicated.
not by the mutant p53 protein (pET-p53M8). These resultssuggested that Southwestern blot analysis may facilitate theisolation of DNA target sequences that are unique for thewild-type p53 but not mutant p53 protein forms. Inserts ofclones MgBH8 and MgBH13 bind wild-type p53 to a lesserextent, but again no binding is evident with mutant p53.Western blot analysis confirmed that equal amounts ofwild-type and mutant p53 proteins were present in each ofthe lanes containing the bacterial extracts (data are notshown). Although fragments MgBH4, MgBH5, and MgBH6vary in size and have different restriction maps (Fig. 1), theydo share sequence homology, as estimated by direct hybrid-izations between each other (data not shown).
Analysis of the MgBH6 clone. To further define the specificp53 binding domain, we focused on the analysis of cloneMgBH6. Digestion of clone MgBH6 by the BamHI restric-tion enzyme gave rise to two fragments of 1.6 and of 0.36 kbpwhich were subcloned in the Bluescript vector and desig-nated MgBH6.1 and MgBH6.2, respectively. Southwesternanalysis showed that the 0.36-kb BamHI fragment can bebound by the wild-type p53 protein (Fig. 3). In addition tothe specific DNA binding of wild-type p53 protein, some
MgBH4rn wt
a0
MgBH5m wt
MgBH6m wt
MgBH13m wt
MgBHSm wt
FIG. 2. Specific DNA binding to MgBH clones mediated bywild-type p53 protein. Mutant (pET-p53M8) and wild-type (pET-
p53cD) bacterially expressed proteins were transferred onto nitro-
cellulose blots and hybridized with radiolabeled inserts of different
MgBH clones. Arrow indicates the p513 protein.
MgBH6-2m wt
I a
FIG. 3. Specific DNA binding to subcloned fragments of cloneMgBH6 mediated by wild-type p53. Mutant (pET-p53M8) andwild-type (pET-p53cD) bacterially expressed proteins were trans-ferred onto nitrocellulose blots and hybridized with radiolabeledDNA fragments representing the BamHI 0.36-kb insert of cloneMgBH6 (MgBH6.2). Arrow indicates the p53 protein.
bacterial proteins bound the labeled DNA. In the last exper-iment (Fig. 3), crude bacterial extracts containing the mam-malian p53 were used for routine screening of p53 targetsequences. When more purified extracts were used, nobackground of the bacterium-derived products was detected(compare with Fig. 2).The DNA sequence of this clone was determined (Fig. 4)
and used to screen the GenBank and EMBL data bases. Nosignificant homologous sequences were found.DMase I protection experiments. The nucleotide sequence
protected by protein binding can be defined by treatment ofthe radiolabeled DNA fragment with a nonspecific DNase.We used the 70Z/3 cell line as a source of wild-type p53-containing nuclear extract. This is a murine leukemia cellline that was induced with methylnitrosourea in thymecto-mized mice (41); it was recently found to express wild-typep53 protein (2a). These DNase I protection experimentsshowed that by using the 70Z/3 nuclear extract, a 30-nucleotide region was protected on both strands of the364-bp fragment (Fig. 5). As can be seen in Fig. 5A and B,only one window that shared the identical sequence wasprotected in both strands. Nuclear extract obtained fromp53-nonproducing pre-B L12 cells was compared with anuclear extract from a p53-producer stable L12-derivedclone that was generated by transfection with a p53 cDNAcoding for wild-type protein (50). When the two nuclearextracts were compared, we found that nuclear extracts
1 10 20 30 40 50 60
GGATCCTCAC TCTTTGGGGT GGTGACGACA GCACTAGGCG GCAGGGAGCC ACCTACATTT
CCAGCACTAG GGAGGGTGGA TGCTGGGCTC CAGAAGATAG GGTGAGGGTT GTCTCCCACA
GCAGGTCACA AGGAGCTGGT CCCCTGGTGA GCAGCAGTTT CTTCTGGTCC GGTATGAGTG
TAGGATTTCA GCACAGAGAC ACTGAGGACA GGTGGTCTCT TGAGATCAGT AAACCAAGTC
AACCTCACAA GCAGAGGAGA GGGACCTCCT GACACTGGTC ACACTTGCT QCTTAGGAAT
CTATTAGGTA CAGGACCTTG GGATGTGGCC TCATAAATTC TCAACGGA.AA CTGGGAGAGG
ATCC
FIG. 4. Nucleotide sequence of clone MgBH6.2. Nucleotides271 to 300, the protected area in the DNase protection assay, areunderlined.
MOL. CELL. BIOL.
I
p53-DNA BINDING SEQUENCES 1381
B.
5' GACACTGGTCACACTTGGCTGCTTAGGAAT 3'CTGTGAGGAGTGTGAACCGACGAATCCTTA
FIG. 5. DNase I footprints of MgBH6.2. BamHI fragments la-beled at the 5' (A) or the 3' (B) end were exposed to DNase I in thepresence of increasing amounts of nuclear extracts of 70Z cells. Theleftmost lane in each panel is the control without any nuclear extractadded. The protected window is marked, and its sequence ispresented below.
from the pre-B L12 p53 producer protected the same regionas did the 70Z nuclear extract. The parental L12 p53-nonproducer cells did not show any protection at that region(data not shown). It should be noted that all three cell linesrepresent cells of the early B-cell differentiation pathway.
Mobility shift assay. To further establish our conclusionthat the protected region in MgBH6.2 is a result of specificbinding to the p53 protein, we utilized the gel shift assay withsynthetic oligonucleotides corresponding to the protectedregion. To that end, the various nuclear extracts wereincubated with the synthetic radiolabeled oligonucleotidewhich contains the p53 binding site. As can be seen in Fig. 6,the nuclear extract obtained from L12 cells showed nospecific band shifts upon incubation with a variety of anti-bodies. However, when a nuclear extract of the p53-pro-ducer line (L12-cD10) was preincubated for 20 min on icewith PAb-421 (an anti-p53 monoclonal antibody) beforemixing with the specific probe, a supershift was induced(Fig. 6, lane b, see arrow). When the PAb-246 anti-p53monoclonal antibody was incubated instead (Fig. 6, lanes c)no supershift was evident; instead, binding seemed to be
L12 L12-cD1O
a b c d a b c d
4-
FIG. 6. Mobility shift mediated by nuclear extracts obtainedfrom p53-producing or nonproducing cells. Nuclear extracts ob-tained from L12, a p53 nonproducer cell line or from a stably derivedp53 producer clone, L12-cD1O, were incubated with labeled oligo-nucleotide (lanes a) and preincubated with PAb-421 (lanes b),PAb-246 (lanes c), or anti-mouse immunoglobulin G (lanes d).
A B C D EF G H
G..H I
70Z Baculovirus Bacterial
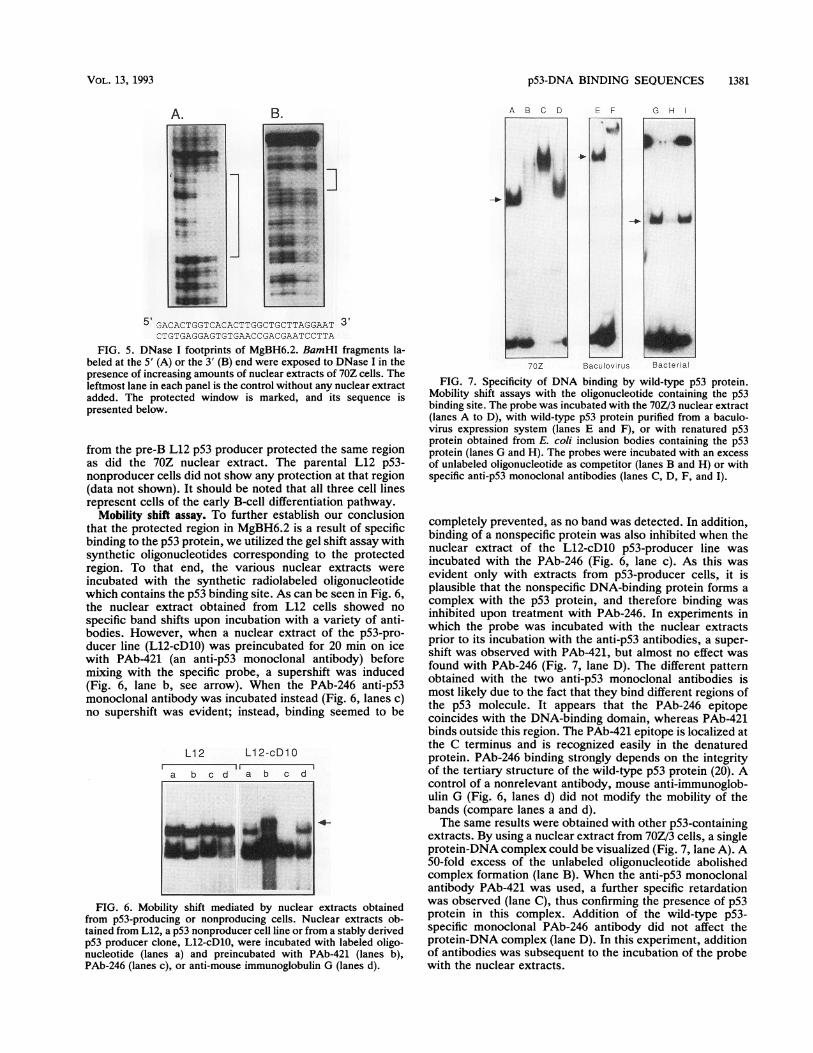
FIG. 7. Specificity of DNA binding by wild-type p53 protein.Mobility shift assays with the oligonucleotide containing the p53binding site. The probe was incubated with the 70Z/3 nuclear extract(lanes A to D), with wild-type p53 protein purified from a baculo-virus expression system (lanes E and F), or with renatured p53protein obtained from E. coli inclusion bodies containing the p53protein (lanes G and H). The probes were incubated with an excessof unlabeled oligonucleotide as competitor (lanes B and H) or withspecific anti-p53 monoclonal antibodies (lanes C, D, F, and I).
completely prevented, as no band was detected. In addition,binding of a nonspecific protein was also inhibited when thenuclear extract of the L12-cD1O p53-producer line wasincubated with the PAb-246 (Fig. 6, lane c). As this wasevident only with extracts from p53-producer cells, it isplausible that the nonspecific DNA-binding protein forms acomplex with the p53 protein, and therefore binding wasinhibited upon treatment with PAb-246. In experiments inwhich the probe was incubated with the nuclear extractsprior to its incubation with the anti-p53 antibodies, a super-shift was observed with PAb-421, but almost no effect wasfound with PAb-246 (Fig. 7, lane D). The different patternobtained with the two anti-p53 monoclonal antibodies ismost likely due to the fact that they bind different regions ofthe p53 molecule. It appears that the PAb-246 epitopecoincides with the DNA-binding domain, whereas PAb-421binds outside this region. The PAb-421 epitope is localized atthe C terminus and is recognized easily in the denaturedprotein. PAb-246 binding strongly depends on the integrityof the tertiary structure of the wild-type p53 protein (20). Acontrol of a nonrelevant antibody, mouse anti-immunoglob-ulin G (Fig. 6, lanes d) did not modify the mobility of thebands (compare lanes a and d).The same results were obtained with other p53-containing
extracts. By using a nuclear extract from 70Z/3 cells, a singleprotein-DNA complex could be visualized (Fig. 7, lane A). A50-fold excess of the unlabeled oligonucleotide abolishedcomplex formation (lane B). When the anti-p53 monoclonalantibody PAb-421 was used, a further specific retardationwas observed (lane C), thus confirming the presence of p53protein in this complex. Addition of the wild-type p53-specific monoclonal PAb-246 antibody did not affect theprotein-DNA complex (lane D). In this experiment, additionof antibodies was subsequent to the incubation of the probewith the nuclear extracts.
_o
_ - _ -~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.
VOL. 13, 1993
w

1382 FOORD ET AL.
c p5'MgBH6-2CAT p3'Mg B H6.2CAT
a b c a b c
FIG. 8. CAT activity derived from the 5'MgBH6.2-CAT and3'MgBH6.2-CAT plasmids transfected into SVT-2 cells. SVT-2 cellswere transfected with 10 jig of 5'MgBH6.2-CAT or 3'MgBH6.2-CAT in three independent experiments (lanes a, b, and c), and CATactivity was determined after 48 h. LTR-CAT, positive controlcontaining the LTR promoter.
When a crude protein extract of Sf9 cells overexpressingthe wild-type p53 from the baculovirus vector was used, acomplex that migrated higher than the one observed with the70Z/3 nuclear extract was seen (Fig. 7, lane E). Further-more, a supershift was seen upon addition of the anti-p53monoclonal antibody PAb-421 to the baculovirus p53-DNAmixture (lane F). Additional proof of the identity of theprotein in the shifted complex was achieved by using purifiedbacterial p53 protein (lane G). Complex formation wasgreatly reduced by adding a 50-fold excess of unlabeledoligonucleotide (lane H). Moreover, the complex was shiftedby the addition of a p53 monoclonal antibody (lane I).The differences in the migration patterns of the labeled
oligonucleotide found when protein extracts from varioussources were used could have resulted from interaction ofp53 protein present in those extracts with different accom-panying proteins. The fact that all types of complexes weresupershifted upon addition of the anti-p53 monoclonal anti-body PAb-421 coupled with the observation that PAb-246seems to prevent complex formation strongly suggests thatin all cases binding to the target DNA is due to complexformation with the p53 protein.
Characterization of the nature of clone MgBH6.2. As in thecase of other transactivators, we expected p53 to bind to theregulatory regions of the specific target gene. We assayed forpossible promoter activity directed by the MgBH6.2 clone.To that end we subcloned it upstream to the CAT reportergene in two orientations and assayed for CAT enzymaticactivity in three independent transfections into SVT-2 cells.Figure 8 shows CAT activity measured 48 h after transfec-tion. The p5'MgBH6.2-CAT construct induced in all threetransfections (lanes a, b, and c) significant CAT activity. Thep3'MgBH6.2-CAT construct did not induce any CAT activ-ity. This strongly suggests that the isolated fragment,MgBH6.2, exhibits a promoter activity in an orientation-specific manner.
In the next experiment we tested whether this sequencecan be transactivated by the p53 protein. To that end, wecotransfected cells with pS'MgBH6.2-CAT and pJflp53cDthat codes for the wild-type p53 protein (3, 52) and measured
60
0)
o 40
0
30-
0)
10
0
pJDcD- + - + - + LTR-CATSVT-2 HeLa SaOs-2 SVT-2
FIG. 9. Transactivation of the 5'MgBH6.2-CAT by wild-typep53 protein encoded by pJflp53cD. The various cell lines (SVT-2,HeLa, and SaOs-2) were transfected with 5'MgBH6. 2-CAT (10 ,ug)(-) or cotransfected with 5'MgBH6.2-CAT and the pJflp53cDplasmids (+). CAT activity is presented as percent acetyl coenzymeA (CoA) conversion (see Materials and Methods). LTR-CAT-transfected SVT-2 served as a positive control.
changes in CAT activity. We repeated the experiments withseveral cell types, and the results obtained are summarizedin Fig. 9. As can be seen, CAT activity measured in SVT-2cells that express endogenously the wild-type p53 was notincreased following cotransfection of pS'MgBH6.2 (10 pLg)with pJflp53cD (10 ,ug). However, when HeLa cells, whichexpress low levels of p53 protein, or SaOs-2 cells, which arep53 nonproducers, were cotransfected with pS'MgBH6.2and pJflpS3cD at the same concentrations, an increasedenzymatic activity was observed. The level of CAT activityexpressed by pS'MgBH6.2 (10 ,ug) in HeLa or SaOs-2 wasvery low compared with that found in the SVT-2 cells. Theselow levels were significantly augmented upon cotransfectionwith pJflp53cD. These experiments suggest that the targetgene isolated here can be transactivated by wild-type p53protein.
DISCUSSION
It is well accepted today that the activity of p53 ismediated through its direct binding to specific DNA se-quences. Some of these sequences were already defined andcharacterized (6, 14, 19, 30, 53).
Previously we showed that the 80 most C-terminal aminoacids of the p53 protein contain a DNA-binding region (53).p53 protein deleted of its C terminus lost its DNA-bindingactivity. Using the Southwestern technique, we concludedthat the binding of DNA to this domain is an intrinsicproperty of the p53 polypeptide and that it does not dependon factors other than p53. Furthermore, that mutant p53exhibited no binding activity was the basis for our assump-tion that this assay was a reliable method for isolating naturaltargets for the wild-type p53 protein. In our experiments weisolated one such potential target sequence that was notidentical with any known p53-DNA binding site.
Recently, a consensus binding site for p53 with an internalsymmetry was identified by El-Deiry et al. (14). Althoughour sequence does not match that consensus sequence, it
MOL. CELL. BIOL.

p53-DNA BINDING SEQUENCES 1383
shows homology in 8 of the 10 residues. Another consensussequence that consists of a palindrome of 20 bp, identified byFunk et al. (19), is completely different from the one de-scribed here. This palindrome is bound by in vitro-translatedp53 protein only when mixed with nuclear extract (19). Thissuggests that the latter sequence required additional proteinsin complex with p53 for binding to occur, whereas thesequence reported here requires no additional factors. Thebinding analysis by means of a Southwestern technique ofvarious MgBH clones showed that wild-type p53 proteinbinds to different sequences with different affinities. Sup-portive evidence comes from the work of Bargonetti et al.(6), showing that wild-type p53 binds less efficiently to theSV40 promoter sequences which do not contain a perfectconsensus sequence (14). On the basis of the fact that theconsensus sequence leads to a strong binding affinity, onecan explain the decrease of binding affinities by an increasingnumber of mismatches. That our p53-DNA binding site is ofmurine origin while that described by others (14, 19) is ofhuman origin can also account for the different characteris-tics of these DNA-binding domains.
In a search for possible target genes for p53 interactions,several candidates involved in normal growth control weresuggested. fos and jun promoters were shown to be down-regulated by wild-type p53 protein. Indeed, cotransfection ofwild-type p53 coding plasmids reduced CAT activity drivenby the fos or jun promoter sequence and the interleukin-6promoter sequences (22, 46). Furthermore, by using thesame assay, wild-type p53 was found to down-regulate theexpression of the multidrug-resistant gene (MDR1), whilemutant p53 was found to upregulate the activity of itspromoter (31). It should be noted that the promoter region ofthe human MDR1 gene shares homology with the p53-DNAbinding site characterized in the present study.The isolated p53 target sequence was capable of promot-
ing the expression of the reporter CAT gene upon coexpres-sion with the wild-type p53 protein. Sequence analysis ofclone MgBH6.2 showed that this fragment contains commonsequence motifs such as promoter-specific sequences andtranscription factor binding sites. These findings are inagreement with recent reports by Kern et al. (30) and byFunk et al. (19), who showed that several genes containingp53 binding sequences can be transactivated by wild-typep53 protein.
Future experiments involving binding of wild-type p53 todifferent promoter sequences will help to determine the roleof p53 in gene transcription and to define the critical ele-ments that are essential for its specific activity. Our resultssupport the conclusion that trans-activation by wild-type p53protein involves recognition of this DNA element, in amanner similar to that of a transcription enhancer. However,on the basis of the involvement of p53 in DNA replication,we cannot exclude the possibility that p53 also functions inDNA synthesis (6, 29) by means of a common DNA element.This is in agreement with the recent suggestion that sometranscription factors activate eukaryotic cellular origins ofDNA replication and transcriptional promoters by means ofthe same mechanism (34). Another possibility is that thetranscription activation function directly interacts with DNAreplication proteins and targets them to their respectiveorigins (38).
ACKNOWLEDGMENTSThis work was supported in part by grants from the Leo and Julia
Forchheimer Center for Molecular Genetics, the Rockefeller-Weiz-mann Fund, and the German-Israeli foundation (GIF). V.R. is the
incumbent of the Norman and Helen Asher Professorial Chair inCancer Research at the Weizmann Institute and the recipient of aCareer Development Award from the Israel Cancer Research Fund(ICRF). N.N. is supported by an Israel Cancer Research Fund(ICRF) postdoctoral fellowship.M. Baer prepared and edited the manuscript.
REFERENCES1. Addison, C., J. R. Jenkins, and H. W. Sturzbecher. 1990. The
p53 nuclear localisation signal is structurally linked to a p34cdc2kinase motif. Oncogene 5:423-426.
2. Ahuja, H., M. Bar-Eli, S. H. Advani, S. Benchimol, and M. J.Cline. 1989. Alterations in the p53 gene and the clonal evolutionof the blast crisis of chronic myelocytic leukemia. Proc. Natl.Acad. Sci. USA 86:6783-6787.
2a.Aloni-Grinstein, R., et al. Unpublished data.3. Arai, N., D. Nomura, K. Yokota, D. Wolf, E. Brill, 0. Shohat,
and V. Rotter. 1986. Immunologically distinct p53 moleculesgenerated by alternative splicing. Mol. Cell. Biol. 6:3232-3239.
4. Baker, J. S., E. R. Fearon, J. M. Nigro, S. R. Hamilton, A. C.Preisinger, J. M. Jessup, P. van Tuinen, D. H. Ledbetter, D. F.Barker, Y. Nakamura, R. White, and B. Vogelstein. 1989.Chromosome 17 deletions and p53 gene mutations in colorectalcarcinomas. Science 244:217-221.
5. Baker, S. J., S. Markowitz, E. R. Fearon, J. K. V. Willson, andB. Vogelstein. 1990. Suppression of human colorectal carcinomacell growth by wild-type p53. Science 249:912-915.
6. Bargonetti, J., P. N. Friedman, S. E. Kern, B. Vogelstein, and C.Prives. 1991. Wild-type but not mutant p53 immunopurifiedproteins bind to sequences adjacent to the SV40 origin ofreplication. Cell 65:1083-1091.
7. Bischoff, J. R., D. Casso, and D. Beach. 1992. Human p53inhibits growth in Schizosaccharomyces pombe. Mol. Cell.Biol. 12:1405-1411.
8. Braithwaite, A. W., H. W. Sturzbecher, C. Addison, C. Palmer,K. Rudge, and J. R. Jenkins. 1987. Mouse p53 inhibits SV40origin-dependent DNA replication. Nature (London) 329:458-460.
9. Chen, P.-L., Y. Chen, R. Bookstein, and W.-L. Lee. 1991.Genetic mechanisms of tumor suppression by the human p53gene. Science 250:1576-1580.
10. Chen, Y., P.-L. Chen, N. Arnaiz, D. Goodrich, and W.-H. Lee.1991. Expression of wild-type p53 in human A673 cells sup-presses tumorigenicity but not growth rate. Oncogene 6:1799-1805.
11. Dang, C. V., and W. M. F. Lee. 1989. Nuclear and nucleolartargeting sequences of c-erb, c-myc, N-myc p53 HSP70 andHIV tat proteins. J. Biol. Chem. 264:18019-18023.
12. Deppert, W., G. Buschhausen-Denker, T. Patschinsky, and K.Steinmeyer. 1990. Cell cycle control of p53 in normal (3T3) andchemically transformed (Meth A) mouse cells. II. Requirementfor cell cycle progression. Oncogene 5:1701-1706.
13. Diller, L., J. Kassel, E. N. Camille, M. A. Gryka, G. Litwak, M.Gebhardt, B. Bressac, M. Ozturk, S. J. Baker, B. Vogelstein, andS. H. Friend. 1990. p53 functions as a cell cycle control proteinin osteosarcomas. Mol. Cell. Biol. 10:5772-5781.
14. El-Deiry, W. S., S. E. Kern, J. A. Pietenpol, K. W. Kinzler, andB. Vogelstein. 1992. Definition of a consensus binding site forp53. Nature Genet. 1:45-49.
15. Eliyahu, D., N. Goldfinger, 0. Pinhasi-Kimhi, G. Shaulsky, Y.Skurnik, N. Arai, V. Rotter, and M. Oren. 1988. Meth-Afibrosarcoma cells express two transforming mutant p53 spe-cies. Oncogene 3:313-321.
16. Eliyahu, D., D. Michalovitz, S. Eliyahu, 0. Pinhasi-Kimhi, andM. Oren. 1989. Wild-type p53 can inhibit oncogene-mediatedfocus formation. Proc. Natl. Acad. Sci. USA 86:8763-8767.
17. Fields, S., and S. K. Jang. 1990. Presence of potent transcriptionactivating sequences in the p53 protein. Science 249:1046-1049.
18. Finlay, C. A., P. W. Hinds, and A. J. Levine. 1989. The p53proto-oncogene can act as a suppressor of transformation. Cell57:1083-1093.
19. Funk, D. W., T. D. Pak, R. H. Kara, W. E. Wright, and J. W.Shay. 1992. A transcriptional active DNA-binding site for hu-
VOL. 13, 1993

1384 FOORD ET AL.
man p53 protein complexes. Mol. Cell. Biol. 12:2866-2871.20. Gannon, J. V., R. Greaves, R. Iggo, and D. P. Lane. 1990.
Activating mutations in p53 produce a common conformationaleffect. A monoclonal antibody specific for the mutant form.EMBO J. 9:1595-1602.
21. Gannon, J. V., and D. P. Lane. 1987. p53 and DNA polymeraseoa compete for binding to SV40 T antigen. Nature (London)329:456-458.
22. Ginsberg, D., F. Mechta, M. Yaniv, and M. Oren. 1991. Wild-type p53 can down-modulate the activity of various promoters.Proc. Natd. Acad. Sci. USA 88:9979-9983.
23. Ginsberg, D., D. Michalovitz, D. Ginsberg, and M. Oren. 1991.Induction of growth arrest by a temperature-sensitive p53mutant is correlated with increased localization and decreasedstability of the protein. Mol. Cell. Biol. 11:582-585.
24. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982.Recombinant genomes which express chloramphenicol acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051.
25. Graham, F. L., and A. J. Van der Eb. 1973. A new technique forthe assay of infectivity of human adenovirus DNA. Virology52:456-467.
26. Hollstein, M., D. Sidransky, B. Vogelstein, and C. C. Harris.1991. p53 mutations in human cancers. Science 253:49-53.
27. Kelman, Z., M. Prokocimer, S. Peller, Y. Kahn, G. Rechavi, Y.Manor, A. Cohen, and V. Rotter. 1989. Rearrangements in thep53 gene in Philadelphia chromosome positive chronic myelog-enous leukemia. Blood 74:2318-2324.
28. Kern, S. E., K. W. Kinzler, S. J. Baker, J. M. Nigro, V. Rotter,A. J. Levine, P. Friedman, C. Prives, and B. Vogelstein. 1991.Mutant p53 proteins bind DNA abnormally in vitro. Oncogene6:131-136.
29. Kern, S. E., K. W. Kinzler, A. Bruskin, D. Jarosz, P. Friedman,C. Prives, and B. Vogelstein. 1991. Identification of p53 as asequence-specific DNA-binding protein. Science 252:1708-1710.
30. Kern, S. E., J. A. Pietenpol, S. Thiagalingam, A. Seymour,K. W. Kinzler, and B. Vogelstein. 1992. Oncogenic forms of p53inhibit p53-regulated gene expression. Science 256:827-830.
31. Khew-Voon, C., U. Kazumitsu, I. Pastan, and M. M. Gottesman.1992. Modulation of activity of the promoter of the humanMDR1 gene by ras and p53. Science 255:459-462.
32. Lane, D., and S. Benchimol. 1990. p53, oncogene or anti-oncogene. Genes Dev. 4:1-8.
33. Levine, A. J., J. Momand, and C. A. Finlay. 1991. The p53tumour suppression gene. Nature (London) 351:453-456.
34. Marahrens, Y., and B. Stillman. 1992. A yeast chromosomalorigin of DNA replication defined by multiple functional ele-ments. Science 255:817-823.
35. Martinez, J., I. Georgoff, J. Martinez, and A. J. Levine. 1991.Cellular localization and cell cycle regulation by a temperaturesensitive p53 protein. Genes Dev. 5:151-159.
36. Mercer, W. E., M. T. Shields, M. Amin, G. J. Sauve, E. Appella,J. W. Romano, and S. J. Ullrich. 1990. Negative growth regu-lation in a glioblastoma tumor cell line that conditionally ex-presses human wild-type p53. Proc. Natl. Acad. Sci. USA87:6166-6170.
37. Michalovitz, D., 0. Halevy, and M. Oren. 1990. Conditionalinhibition of transformation and of cell proliferation by a tem-perature-sensitive mutant of p53. Cell 62:671-680.
38. Mohr, I. J., R. Clark, S. Sun, E. J. Androphy, P. MacPherson,and M. R. Botchan. 1990. Targeting the El replicon protein tothe papillomavirus origin of replication by complex formationwith the E2 transactivator. Science 250:1694-1699.
39. Nigro, J. M., J. S. Baker, A. C. Preisinger, J. M. Jessup, K.Hostetter, K. Cleary, S. H. Bigner, N. Davidson, S. Baylin, P.Devilee, T. Glover, F. S. Collins, A. Weston, R. Modali, C. C.Harris, and B. Vogelstein. 1989. p53 gene mutations occur indiverse human tumor types. Nature (London) 342:705-708.
40. O'Rourke, R. W., C. W. Miller, G. J. Kato, K. J. Simon, D. L.Chen, C. V. Dang, and H. P. Koeffler. 1990. A potentialtranscriptional activation element in the p53 protein. Oncogene5:1829-1832.
41. Paige, C. J., P. W. Kincade, and P. Ralph. 1978. Murine B cellleukemia line with inducible surface immunoglobulin expres-sion. J. Immunol. 121:641-647.
42. Prokocimer, M., M. Shaklai, H. Ben-Bassat, D. Wolf, N. Gold-finger, and V. Rotter. 1986. Expression of p53 in human leuke-mia and lymphoma. Blood 68:113-119.
43. Raycroft, L., H. Wu, and G. Lozano. 1990. Transcriptionalactivation by wild type but not transforming mutants of the p53anti oncogene. Science 249:1049-1051.
44. Rodrigues, N. R., A. Rowan, M. E. F. Smith, I. B. Kerr, W. F.Bodmer, J. V. Gannon, and D. P. Lane. 1990. p53 mutations incolorectal cancer. Proc. Natl. Acad. Sci. USA 87:7555-7559.
45. Rosenberg, A. H., B. N. Lade, D.-S. Chui, S.-W. Lin, J. J. Dunn,and F. W. Studier. 1987. Vectors for selective expression ofcloned DNAs by T7 RNA polymerase. Gene 56:125-135.
46. Santhanam, U., A. Ray, and P. B. Sehgal. 1991. Repression ofthe interleukin 6 gene promoter by p53 and the retinoblastomasusceptibility gene product. Proc. Natl. Acad. Sci. USA 88:7605-7609.
47. Shaulsky, G., A. Ben-Ze'ev, and V. Rotter. 1990. Subcellulardistribution of the p53 protein during the cell cycle of Balb/c 3T3cells. Oncogene 5:1707-1711.
48. Shaulsky, G., N. Goldfinger, A. Ben-Ze'ev, and V. Rotter. 1990.Nuclear accumulation of p53 protein is mediated by severalnuclear localization signals and plays a role in tumorigenesis.Mol. Cell. Biol. 10:6565-6577.
49. Shaulsky, G., N. Goldfinger, A. Peled, and V. Rotter. 1991.Involvement of wild type p53 protein in the cell cycle requiresnuclear localization. Cell Growth Diff. 2:661-667.
50. Shaulsky, S., N. Goldfinger, A. Peled, and V. Rotter. 1991.Involvement of wild type p53 in pre-B cell differentiation invitro. Proc. Natl. Acad. Sci. USA 88:8982-8986.
51. Shaulsky, G., N. Goldfinger, and V. Rotter. 1991. Alteration intumor development in vivo mediated by expression of wild typeor mutant p53 proteins. Cancer Res. 51:5232-5237.
52. Shaulsky, G., N. Goldfinger, M. S. Tosky, A. J. Levine, and V.Rotter. 1991. Nuclear localization is essential for the activity ofp53 protein. Oncogene 6:2055-2065.
53. Shohat-Foord, O., P. Bhattacharya, Z. Reich, and V. Rotter.1991. A DNA binding domain in contained in the C-terminus ofwild type p53 protein. Nucleic Acids Res. 19:5191-5198.
54. Sturzbecher, H. W., R. Brain, T. Maimets, C. Addison, K.Rudge, and J. R. Jenkins. 1988. Mouse p53 blocks SV40 DNAreplication in vitro and downregulates T antigen DNA helicaseactivity. Oncogene 3:405-413.
55. Towbin, H. H., T. Staehelin, and J. Gordon. 1979. Electro-phoretic transfer of proteins from polyacrylamide gels to nitro-cellulose sheets: procedure and some applications. Proc. Natl.Acad. Sci. USA 76:4350-4354.
56. Vogelstein, B. 1990. A deadly inheritance. Nature (London)348:681-682.
57. Wang, E. H., P. N. Friedman, and C. Prives. 1989. The murinep53 protein blocks replication of SV40 DNA in vitro by inhib-iting the initiation function of SV40 large T antigen. Cell57:379-392.
58. Wilcock, D., and D. P. Lane. 1991. Localization of p53, retino-blastoma and host replication proteins at sites of viral replica-tion in herpes-infected cells. Nature (London) 349:429-431.
59. Wolf, D., N. Harris, N. Goldfinger, and V. Rotter. 1985. Isola-tion of a full-length mouse cDNA clone coding for an immuno-logically distinct p53 molecule. Mol. Cell. Biol. 5:127-132.
60. Wolf, D., and V. Rotter. 1985. Major deletions in the geneencoding the p53 antigen cause lack of p53 expression in HL-60cells. Proc. Natl. Acad. Sci. USA 82:790-794.
MOL. CELL. BIOL.