jayne cartmell et al- the metabotropic glutamate 2/3 receptor agonists ly354740 and ly379268...
TRANSCRIPT

8/3/2019 Jayne Cartmell et al- The Metabotropic Glutamate 2/3 Receptor Agonists LY354740 and LY379268 Selectively Attenu…
http://slidepdf.com/reader/full/jayne-cartmell-et-al-the-metabotropic-glutamate-23-receptor-agonists-ly354740 1/10
The Metabotropic Glutamate 2/3 Receptor Agonists LY354740and LY379268 Selectively Attenuate Phencyclidine versusd-Amphetamine Motor Behaviors in Rats
JAYNE CARTMELL, JAMES A. MONN, and DARRYLE D. SCHOEPP
Lilly Research Laboratories, Eli Lilly and Company, Indianapolis, Indiana
Accepted for publication June 3, 1999 This paper is available online at http://www.jpet.org
ABSTRACT
Previous animal studies have indicated that drugs targeted atmetabotropic glutamate (mGlu) receptors may be useful fortreatment of psychosis. In this article, the effects of the novel,
potent, and selective mGlu2/3 receptor agonists LY354740 andLY379268, and the clinically effective agents clozapine andhaloperidol, were investigated using phencyclidine (PCP; 5 mg/ kg)- versus d-amphetamine (AMP; 3 mg/kg)-evoked motor ac-tivities. LY354740 (1–10 mg/kg s.c.), LY379268 (0.3–3 mg/kgs.c.), clozapine (1–10 mg/kg s.c.), and haloperidol (0.03–1mg/kg s.c.) reversed the increases in ambulations, fine motor(nonambulatory) movements, and decreased time at restevoked by PCP. Furthermore, the inhibitions of the PCP re-sponse by the mGlu2/3 agonist LY379268, but not by cloza-pine, were completely reversed by the selective mGlu2/3 re-ceptor antagonist LY341495. Doses of LY354740 andLY379268 that blocked the effects on PCP had no effects on
rotorod performance, and (with the exception of rearing behav-ior) had minimal effects on AMP-evoked motor activities. Clo-zapine blocked AMP-induced rearing but enhanced AMP-in-
duced ambulations and fine movements at the lower doses (1and 3 mg/kg). Unlike the mGlu2/3 agonists, the highest dose ofclozapine tested (10 mg/kg) impaired animals on the rotorod.Haloperidol potently blocked all PCP and AMP effects, but onlyat doses associated with motor impairment. These data dem-onstrate that mGlu2/3 receptor agonists act via a unique mech-anism to selectively block PCP-induced behaviors. Moreover,the marked mGlu2/3 receptor-mediated inhibitions of PCP-evoked behaviors by LY354740 and LY379268, with minimaleffects on AMP, may indicate potential antipsychotic effects inhumans in the absence of dopamine mediated extrapyramidalside effects.
Metabotropic glutamate (mGlu) receptors are a novel fam-ily of at least eight G protein-coupled receptor subtypes,which have been subclassified into three groups based onshared structural homology, coupling mechanisms, and phar-macological properties. Group I mGlu receptors (mGlu1 andmGlu5) are coupled to phospholipase C activation, whereasthe group II (mGlu2 and mGlu3) and group III (mGlu4,mGlu6, mGlu7, and mGlu8) mGlu receptors are coupled neg-atively to adenylate cyclase (Nakanishi, 1992; Pin and Du-
voisin, 1995; Conn and Pin, 1997). Current knowledge of thepharmacology of these receptors, along with studies on their
localizations and physiological/pathological functions in thecentral nervous system (CNS) suggest that mGlu receptorsare novel drug targets for psychiatric and neurological disor-ders (Nicoletti et al., 1996; Knopfel and Gasparini, 1996).
Modulation of glutamatergic synaptic transmission bygroup II (mGlu2/3) receptor agonists is of interest as a novelmechanism with which to regulate neuronal activity withinspecific synapses and neuronal circuits. MGlu2 and mGlu3
receptors are highly expressed in forebrain regions (i.e., pre-frontal cortex and hippocampus; Ohishi et al., 1993a,b; Nekiet al., 1996) where pathologically enhanced glutamate trans-mission has been implicated in a variety of CNS disorders.Interestingly, mGlu2 receptors are located presynapticallybut outside of the active zone of glutamate release (in theperiphery of the synapse) where they function to reduceglutamate excitations in an activity-dependent manner(Ohishi et al., 1994; Shigemoto et al., 1997; Forsythe andBarnes-Davies, 1997). Brain synapses where mGlu2/3 ago-nists have been shown to suppress glutamate transmission
include the mossy fiber (Scanziani et al., 1997) and perforantpathways (Macek et al., 1996; Kilbride et al., 1998) of thehippocampus, the locus ceruleus (Dube and Marshall, 1997),nuclei of the amygdala (Neugebauer et al., 1997), the cortico-striatal pathway (Lovinger and McCool, 1995), and the me-dial prefrontal cortex (Marek and Aghajanian, 1998). Thus,mGlu2/3 receptor activation represents a novel mechanismfor limiting excitatory synaptic transmission under condi-tions of excessive (pathologically) enhanced glutamate trans-mission in a regional, selective, and synapse-specific manner.Received for publication March 5, 1999.
ABBREVIATIONS: mGlu, metabotropic glutamate; PCP, phencyclidine; AMP, d-amphetamine; MCPG, ( RS )--methyl-4-carboxyphenylglycine.
0022-3565/99/2911-0161$03.00/0THE JOURNAL OF PHARMACOLOGY AND E XPERIMENTAL THERAPEUTICS Vol. 291, No. 1Copyright © 1999 by The American Society for Pharmacology and Experimental Therapeutics Printed in U.S.A.JPET 291:161–170, 1999
161

8/3/2019 Jayne Cartmell et al- The Metabotropic Glutamate 2/3 Receptor Agonists LY354740 and LY379268 Selectively Attenu…
http://slidepdf.com/reader/full/jayne-cartmell-et-al-the-metabotropic-glutamate-23-receptor-agonists-ly354740 2/10
LY354740 is a nanomolar potent, receptor-selective, andsystemically active mGlu2/3 receptor agonist (Monn et al.,1997; Schoepp et al., 1997; Battaglia et al., 1997). Thus,LY354740 has served as a useful in vivo pharmacological toolfor investigating the role of mGlu2/3 receptors in multipledisorders of the CNS. LY354740 has been shown to be activein animal models of seizures (Monn et al., 1997), globalischemia (Bond et al., 1998), anxiety (Monn et al., 1997;Helton et al., 1998b), and drug withdrawal states (Helton etal., 1997; Helton et al., 1998a). Each of these conditions havein common pathologically enhanced glutamate transmissionin the CNS (Danysz et al., 1995), and thus provide the the-oretical basis for investigations of the therapeutic prospectsfor mGlu2/3 agonists.
With this in mind, recent animal studies with phencyclid-ine (PCP) have suggested that group II mGlu receptors rep-resent a novel target for the treatment of psychosis. Pharma-cological studies in humans and animals suggest that PCPrepresents a useful model of psychosis, and serves as thebasis for the “glutamate hypothesis of schizophrenia”. PCPhas been reported to produce schizophrenia-like symptoms inhealthy subjects and worsen pyschosis in schizophrenic pa-
tients (Javitt et al., 1991; Halberstadt, 1995; Steinpreis,1996). The mechanism(s) by which PCP induces psychosis inhumans is currently an active area of research, and may bethe key to understanding the actions of current and futureantipsychotic drugs. PCP is open channel blocker of the N -methyl-D-aspartate subclass of ionotropic glutamate recep-tors (Anis et al., 1983) that has multiple behavioral effects inanimals, which include increased motor behaviors, stereo-typy, and cognitive disruptions (Murray and Horita, 1979;Sturgeon et al., 1979; Steinpreis, 1996). These actions of PCPin animals have been linked to enhanced release of a numberof CNS neurotransmitters, including glutamate (Adams andMoghaddam, 1998), as well as dopamine and serotonin (Her-nandez et al., 1988; Hondo et al., 1994; Lillrank et al., 1994).
Recently, Moghaddam and Adams (1998) reported thatLY354740 blocked PCP-evoked increases in prefrontal cortexglutamate levels, while having no effects on basal or PCP-evoked extracellular dopamine levels. This effect of LY354740 was associated with reductions in PCP-evokedlocomotor activity and inhibition of PCP-induced stereotyp-ies. These data indicate that mGlu2/3 receptor agonists mayblock PCP-mediated behaviors via a novel mechanism (re-duction in PCP-evoked glutamate release), which, unlike typ-ical and atypical antipsychotic drugs, might be independentof the dopamine system.
Amphetamine interacts with catecholamine systems (do-pamine, serotonin, and norepinephrine) in the CNS to pro-duce a variety of behaviors in animals and humans (Seiden etal., 1993). In rats, d-amphetamine (AMP) produces increasedlocomotion and stereotypies, the expression of which arehighly dependent on the dose of AMP tested (Kuczenski andSegal, 1989; Seiden et al., 1993). Antipsychotic drugs, includ-ing typical drugs such as haloperidol, and atypical drugs,such as clozapine, have in common postsynaptic dopaminereceptor-blocking properties (Iversen, 1975). Because thesecompounds reverse certain AMP-induced behaviors in ani-mals (Del Rio and Fuentes, 1969; Tschanz and Rebec, 1989;Mueller, 1993), as well as schizophrenic symptoms in hu-mans, a role for an abnormally activated dopamine system inschizophrenia has been proposed. However, motor distur-
bances such as extrapyramidal Parkinsonian signs have alsobeen linked to dopamine blockade by antipsychotic drugs(Seeman, 1995). Thus, the discovery of novel antipsychoticmechanisms (e.g., drugs that directly target glutamate recep-tor mechanisms) may be desirable.
In this article, we further investigated a potential role of mGlu2/3 receptors in the treatment of drug-induced psycho-sis and possibly schizophrenia using both PCP and AMPmodels in rats. The systemically active mGlu2/3 receptoragonist LY354740 (Monn et al., 1997; Schoepp et al., 1997)and a newer structurally different mGlu2/3 receptor agonistcompound, LY379268 (Monn et al., 1999), were examined fortheir abilities to influence PCP- versus AMP-induced motoractivations, by using an automated behavioral system thatmonitors both gross ambulatory locomotion and nonambula-tory fine movements in a home-cage environment. To test forsedation/ataxia and thus determine whether the effects of these agents were related to motor impairment of the ani-mals, a rotorod apparatus was used. The actions of knownantipsychotics, clozapine and haloperidol, were also exam-ined in the same test systems. To further establish that theeffects of mGlu receptor agonists in these test systems were
mGlu2/3 receptor mediated, we used the potent and highlyselective mGlu2/3 receptor antagonist compound, LY341495(Kingston et al., 1998).
Experimental Procedures
All experiments were performed in accordance with Eli Lilly andCompany animal care and use policies. Male Sprague-Dawley rats(250–300 g) were group-housed (maximum of seven rats per cage)under standard laboratory conditions with ad libitum access to foodand water (12 h light/dark cycle), for at least 1 day before use.
Activity Assessment. Behaviors were monitored in transparent,plastic shoebox cages of the dimensions 45 25 20 cm, with 1 cmdepth of wood chips as bedding, and a metal grill on top of the cage.
Motor monitors (Hamilton Kinder, Poway, CA) consisted of a rect-angular rack of 12 photobeams arranged in a 8 4 formation. Shoebox cages were placed inside these racks, enabling the activity of therat to be monitored in a home-cage environment. The lower rack waspositioned at a height of 5 cm, which allowed the detection of PCP-induced head movements in addition to movements of the body of therat. Rearing activity was detected by a second rack placed 10 cmabove the first.
Rats were placed in the cage for an acclimation period of 30 min,then were removed, administered a s.c. injection of LY354740,LY379268, clozapine, haloperidol, or sterile water (1 ml/kg), andthen returned to the same cages. Antagonist studies using the selec-tive group II receptor antagonist LY341495 at a dose of 1 mg/kgconsisted of coinjection of LY341495 with LY379268, clozapine, orsterile water. After 30 min, the rats were given an s.c. injection of
PCP, AMP, or sterile water (1 ml/kg), and once again returned to thecages. Motor activity was monitored for the following 60 min. Soft-ware analysis of beam breaks, under the definitions of HamiltonKinder, resulted in the measurement of three different parameters:ambulations (pattern of beam breaks indicating that the animal hasrelocated its entire body), fine movements (nonambulatory beambreaks), and time at rest (total seconds in a 60-min session in whichno new beams were broken, measured at 1-s intervals). An indicationof rearing activity was determined by the total number of ambula-tions detected in the upper rack of photobeams.
Rotorod Performance. An automated rotorod apparatus (Rotor-Rod; San Diego Instruments Inc., San Diego, CA) was used as a testfor motor impairment/ataxia. Ninety minutes before drug adminis-tration, rats were trained to stay on the rotorod, rotating at 4 rpm,
162 Cartmell et al. Vol. 291
8/3/2019 Jayne Cartmell et al- The Metabotropic Glutamate 2/3 Receptor Agonists LY354740 and LY379268 Selectively Attenu…
http://slidepdf.com/reader/full/jayne-cartmell-et-al-the-metabotropic-glutamate-23-receptor-agonists-ly354740 3/10
over four successive trials. Those rats that remained on the rod for aconsecutive 60-s period were retested 30 min before drug adminis-tration. Rats successful in the retesting session were then given s.c.injections of LY354740, LY369268, clozapine, haloperidol, or sterilewater (1 ml/kg). After an additional 30 min, these rats were againtested on the rotorod for a period of up to 60 s. Data were expressedas the number of seconds in which the animal remained on therotorod apparatus.
Statistical Analysis. Statistical analyses of behaviors were car-
ried out using the GraphPad PRISM statistical program (GraphPad,San Diego, CA). Data were analyzed by a one-way ANOVA, and thenpost hoc comparisons for each dose group versus control or PCP aloneor PCP and test compound were made using Newman-Keuls Multi-ple Comparison Test. A P .05 was considered significant. ED50
values were calculated from mean data using the median effect plotof Chou and Talalay (1983). For this analysis, data at each dose of test drug were converted percentage of inhibition (or percentage of reversal) of PCP or amphetamine-induced changes in activity. ED50
values generally represent three increasing doses of drug producingat least one value with 50% effect and one value with 50% effectand a linear regression coefficient (r2) of 0.9.
Materials. PCP, hydrochloride, and AMP sulfate were obtainedfrom Sigma (St. Louis, MO). Clozapine and haloperidol were pur-chased from Research Biochemicals International (Natick, MA).
LY354740 and LY379268 were synthesized as described previously(Monn et al., 1997, 1999). LY341495 (Ornstein et al., 1998) wasprovided by Dr. Paul L. Ornstein at Lilly Research Laboratories,Indianapolis.
Results
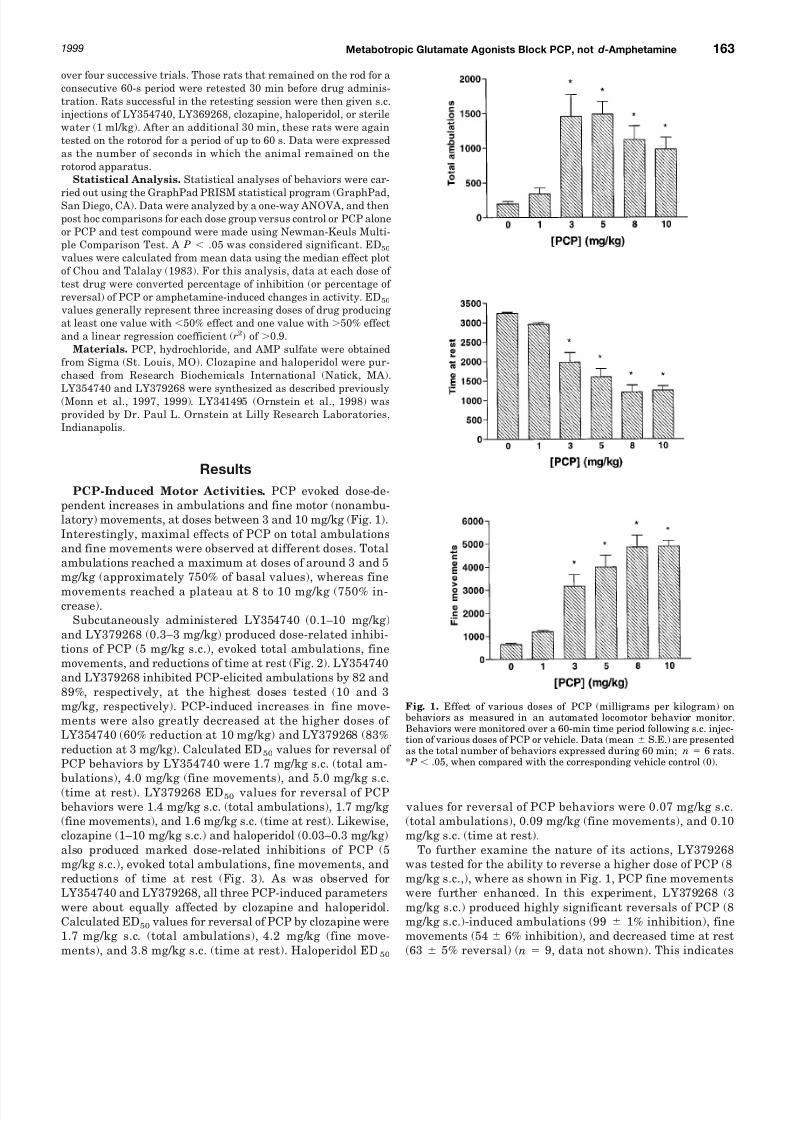
PCP-Induced Motor Activities. PCP evoked dose-de-pendent increases in ambulations and fine motor (nonambu-latory) movements, at doses between 3 and 10 mg/kg (Fig. 1).Interestingly, maximal effects of PCP on total ambulationsand fine movements were observed at different doses. Totalambulations reached a maximum at doses of around 3 and 5mg/kg (approximately 750% of basal values), whereas fine
movements reached a plateau at 8 to 10 mg/kg (750% in-crease).
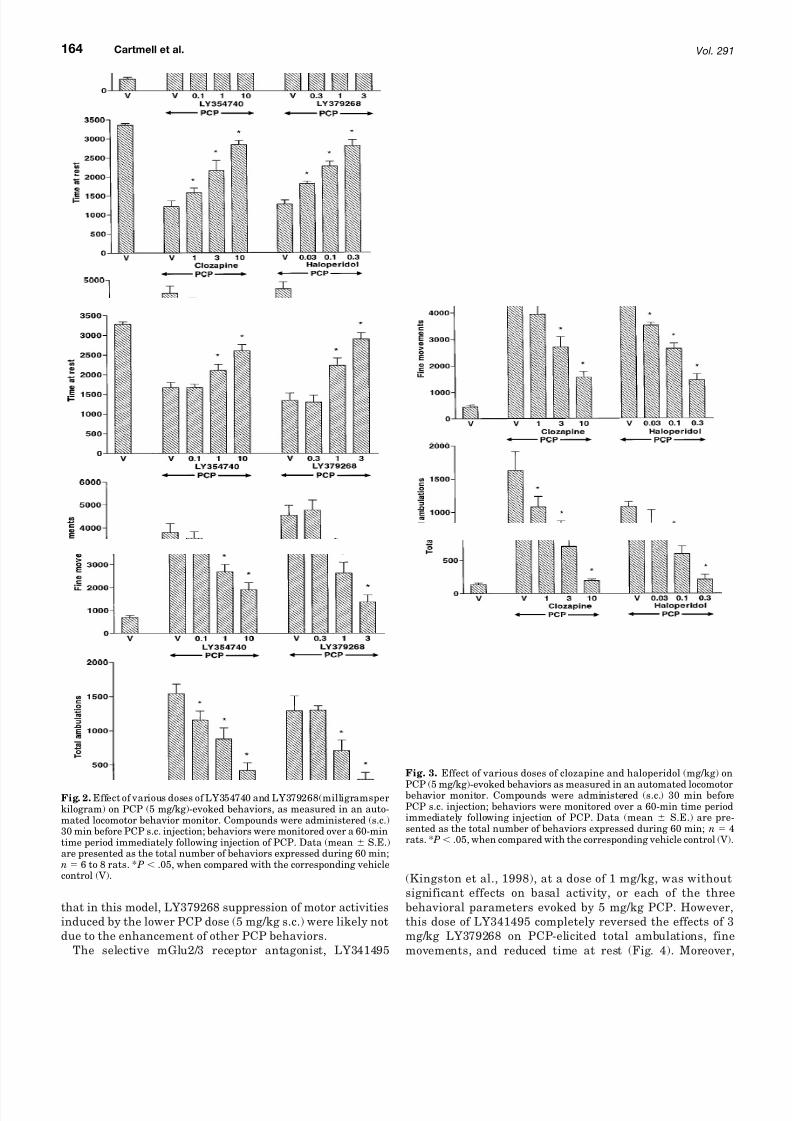
Subcutaneously administered LY354740 (0.1–10 mg/kg)and LY379268 (0.3–3 mg/kg) produced dose-related inhibi-tions of PCP (5 mg/kg s.c.), evoked total ambulations, finemovements, and reductions of time at rest (Fig. 2). LY354740and LY379268 inhibited PCP-elicited ambulations by 82 and89%, respectively, at the highest doses tested (10 and 3mg/kg, respectively). PCP-induced increases in fine move-ments were also greatly decreased at the higher doses of LY354740 (60% reduction at 10 mg/kg) and LY379268 (83%reduction at 3 mg/kg). Calculated ED50 values for reversal of PCP behaviors by LY354740 were 1.7 mg/kg s.c. (total am-bulations), 4.0 mg/kg (fine movements), and 5.0 mg/kg s.c.(time at rest). LY379268 ED50 values for reversal of PCPbehaviors were 1.4 mg/kg s.c. (total ambulations), 1.7 mg/kg(fine movements), and 1.6 mg/kg s.c. (time at rest). Likewise,clozapine (1–10 mg/kg s.c.) and haloperidol (0.03–0.3 mg/kg)also produced marked dose-related inhibitions of PCP (5mg/kg s.c.), evoked total ambulations, fine movements, andreductions of time at rest (Fig. 3). As was observed forLY354740 and LY379268, all three PCP-induced parameterswere about equally affected by clozapine and haloperidol.Calculated ED50 values for reversal of PCP by clozapine were1.7 mg/kg s.c. (total ambulations), 4.2 mg/kg (fine move-ments), and 3.8 mg/kg s.c. (time at rest). Haloperidol ED50
values for reversal of PCP behaviors were 0.07 mg/kg s.c.(total ambulations), 0.09 mg/kg (fine movements), and 0.10mg/kg s.c. (time at rest).
To further examine the nature of its actions, LY379268was tested for the ability to reverse a higher dose of PCP (8mg/kg s.c.,), where as shown in Fig. 1, PCP fine movementswere further enhanced. In this experiment, LY379268 (3mg/kg s.c.) produced highly significant reversals of PCP (8mg/kg s.c.)-induced ambulations (99 1% inhibition), finemovements (54 6% inhibition), and decreased time at rest(63 5% reversal) (n 9, data not shown). This indicates
Fig. 1. Effect of various doses of PCP (milligrams per kilogram) onbehaviors as measured in an automated locomotor behavior monitor.Behaviors were monitored over a 60-min time period following s.c. injec-tion of various doses of PCP or vehicle. Data (mean S.E.) are presentedas the total number of behaviors expressed during 60 min; n 6 rats.* P .05, when compared with the corresponding vehicle control (0).
1999 Metabotropic Glutamate Agonists Block PCP, not d -Amphetamine 163
8/3/2019 Jayne Cartmell et al- The Metabotropic Glutamate 2/3 Receptor Agonists LY354740 and LY379268 Selectively Attenu…
http://slidepdf.com/reader/full/jayne-cartmell-et-al-the-metabotropic-glutamate-23-receptor-agonists-ly354740 4/10
that in this model, LY379268 suppression of motor activitiesinduced by the lower PCP dose (5 mg/kg s.c.) were likely notdue to the enhancement of other PCP behaviors.
The selective mGlu2/3 receptor antagonist, LY341495
(Kingston et al., 1998), at a dose of 1 mg/kg, was withoutsignificant effects on basal activity, or each of the threebehavioral parameters evoked by 5 mg/kg PCP. However,this dose of LY341495 completely reversed the effects of 3mg/kg LY379268 on PCP-elicited total ambulations, finemovements, and reduced time at rest (Fig. 4). Moreover,
Fig. 2. Effect of various doses of LY354740 and LY379268(milligramsperkilogram) on PCP (5 mg/kg)-evoked behaviors, as measured in an auto-mated locomotor behavior monitor. Compounds were administered (s.c.)30 min before PCP s.c. injection; behaviors were monitored over a 60-mintime period immediately following injection of PCP. Data (mean S.E.)are presented as the total number of behaviors expressed during 60 min;n 6 to 8 rats. * P .05, when compared with the corresponding vehiclecontrol (V).
Fig. 3. Effect of various doses of clozapine and haloperidol (mg/kg) on
PCP (5 mg/kg)-evoked behaviors as measured in an automated locomotorbehavior monitor. Compounds were administered (s.c.) 30 min beforePCP s.c. injection; behaviors were monitored over a 60-min time periodimmediately following injection of PCP. Data (mean S.E.) are pre-sented as the total number of behaviors expressed during 60 min; n 4rats. * P .05, when compared with the corresponding vehicle control (V).
164 Cartmell et al. Vol. 291

8/3/2019 Jayne Cartmell et al- The Metabotropic Glutamate 2/3 Receptor Agonists LY354740 and LY379268 Selectively Attenu…
http://slidepdf.com/reader/full/jayne-cartmell-et-al-the-metabotropic-glutamate-23-receptor-agonists-ly354740 5/10
when tested against a dose of clozapine (10 mg/kg) thatproduced the same extent of inhibition of the PCP responseas 3 mg/kg 379268, 1 mg/kg LY341495 was without signifi-cant effect on each of the three parameters measured (Fig. 4).
AMP-Induced Motor Activities. AMP produced a cleardose-related biphasic effect on total ambulations (Fig. 5). Adose of 3 mg/kg AMP maximally increased total ambulationsby 590% of basal, a value comparable with the maximal effectinduced by PCP (Fig. 1). The higher doses of AMP (10 and 30mg/kg) produced significantly smaller increases in total am-bulations when compared with the 3 mg/kg dose ( P .05). Infact, at the highest dose tested (30 mg/kg), significant AMP-induced increases in total ambulations were no longer ob-served (Fig. 5). Similar to ambulations, AMP also increasedrearing activity in a biphasic manner, peaking at a dose of 3mg/kg, with no significant increases at the higher doses of 10or 30 mg/kg (Fig. 5). At the 3 mg/kg dose, rearing activity wasincreased over 30-fold the basal values (Fig. 5). In contrast,
AMP-evoked increases in fine movements were maximal at 1mg/kg and remained significantly elevated at doses up to 30mg/kg AMP. The maximal increases in AMP-evoked finemovements were lower than those observed for PCP (330% of basal at 3 mg/kg s.c. AMP). These behavioral effects alsoresulted in a dose-related reductions in time spent at restover a range of 1 to 30 mg/kg AMP.
Overall, doses of LY354740 and LY379268, which signifi-
cantly reduced PCP-evoked motor activities produced little orrelatively modest reductions in AMP (3 mg/kg s.c.)-evokedtotal ambulations, fine movements, or reduced time at rest(Fig. 6). Even at the dose of LY354740, which almost com-pletely abolished actions of PCP (10 mg/kg), no significanteffects on AMP-induced total ambulations, fine movements,or reduced time at rest were observed. Certain doses of LY379268 produced statistically significant decreases in
AMP-evoked total ambulations (29 and 42% reductions at 1and 3 mg/kg, respectively) or fine movements (30 and 33%reductions at 1 and 3 mg/kg, respectively). However, thesesame doses had no effect AMP-evoked reductions in time atrest (Fig. 6). In contrast, both LY354740 and LY379268greatly reduced AMP-elicited rearing activity. At 10 mg/kg of
LY354740, AMP-induced rearing activity was inhibited by55%, whereas a dose of 3 mg/kg LY379268 produced 91%inhibition of AMP-rearing activity (Fig. 6).
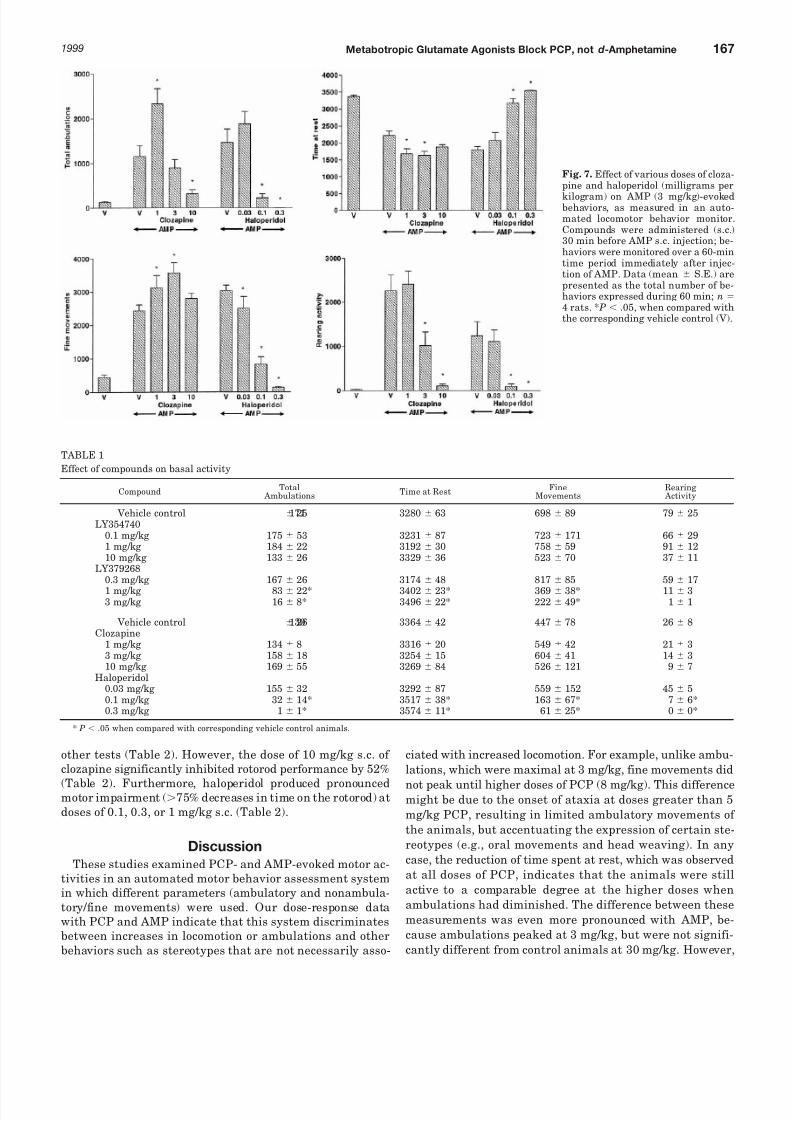
Similar to mGlu2/3 agonists, clozapine failed to inhibit finemovements elicited by AMP, but almost completely abolishedrearing activity at 10 mg/kg. Interestingly, fine movementswere actually enhanced by the lower dose (1 mg/kg s.c.) of clozapine, to 160% of the control value. Likewise, clozapine(1 mg/kg s.c.) significantly increased total ambulations (to215% of basal). Doses of 1 and 3 mg/kg clozapine also signif-icantly enhanced both AMP-evoked increases in fine move-ments and decreases in time at rest. However, the highestdose of clozapine (10 mg/kg s.c.) reduced AMP-evoked ambu-lations by 81%, but had no effects on either fine movementsor time at rest (Fig. 7). In contrast, rearing activity evoked by
AMP was reduced by clozapine in a dose-related manner overthe range of 1 to 10 mg/kg clozapine (ED50 4.0 mg/kg).
In common with its effects on PCP-evoked behaviors, hal-operidol (0.03–0.3 mg/kg) produced dose-related reductionsin all AMP-induced behaviors, including ambulations (ED50
0.1 mg/kg), fine movements (ED50 0.05 mg/kg), reduced time atrest (ED50 0.05 mg/kg), and rearing activity (ED50 0.05 mg/kg)(Fig. 7).
Effect on Basal Motor Activities. The effects of LY354740, LY379268, clozapine, and haloperidol on basalactivities were determined under the same conditions used
Fig. 4. Effect of the selective mGlu2/3 receptor antagonist, LY341495 (1mg/kg) on the reversal of PCP (5 mg/kg)-evoked behaviors by 3 mg/kgLY379268 and 10 mg/kg clozapine. LY341495 or vehicle were coadminis-tered (s.c.) with LY379268, clozapine or vehicle 30 min before PCPor vehicles.c. injection; behaviors were monitored over a 60-min time period immedi-ately after injection of PCP. Data (mean S.E.) are presented as the totalnumber of behaviors expressed during 60 min; n 6 to 8 rats. * P .05,when compared with corresponding LY341495 vehicle control (V).
1999 Metabotropic Glutamate Agonists Block PCP, not d -Amphetamine 165

8/3/2019 Jayne Cartmell et al- The Metabotropic Glutamate 2/3 Receptor Agonists LY354740 and LY379268 Selectively Attenu…
http://slidepdf.com/reader/full/jayne-cartmell-et-al-the-metabotropic-glutamate-23-receptor-agonists-ly354740 6/10
when studying AMP- or PCP-evoked increases in motor ac-tivities (Table 1). Animals were adapted to the cage environ-ment for 30 min, drugs or saline vehicle were then adminis-tered, and motor activities were monitored 30 min later for a60-min period (Table 1). LY354740 and clozapine (up to dosesof 10 mg/kg) did not significantly affect any basal activities of the animals. In contrast, haloperidol significantly reducedtotal ambulations, fine movements, and rearing activity atdoses of 0.1 mg/kg and greater; total ambulations and rears
were completely abolished at 0.3 mg/kg. In contrast toLY354740, LY379268 at 1 and 3 mg/kg significantly reducedbasal total ambulations and fine movements, and increasedthe animals’ time at rest (Table 1).
Performance on Rotorod. LY354740 and LY379268,tested at doses up to 10 and 3 mg/kg, respectively, did notsignificantly affect the performance of rats on a rotorod ap-paratus, indicating that these compounds did not impair themotor abilities of the animals at all of the doses examined in
Fig. 5. Effect of various doses of AMP(mg/kg) on behaviors as measured inan automated locomotor behaviormonitor. Behaviors were monitoredover a 60-min time period after s.c.injection of various doses of AMP or
vehicle. Data (mean S.E.) are pre-sented as the total number of behav-iors expressed during 60 min; n 3rats. * P .05, when compared withthe corresponding vehicle control (V).
Fig. 6. Effect of various doses of LY354740 and LY379268 (mg/kg) on
AMP (3 mg/kg)-evoked behaviors, asmeasured in an automated locomotorbehavior monitor. Compounds wereadministered (s.c.) 30 min before AMPs.c. injection, behaviors were moni-tored over a 60-min time period imme-diately after injection of AMP. Data(mean S.E.) are presented as thetotal number of behaviors expressedduring 60 min; n 4 to 6 rats. * P .05, when compared with AMP alone.
166 Cartmell et al. Vol. 291
8/3/2019 Jayne Cartmell et al- The Metabotropic Glutamate 2/3 Receptor Agonists LY354740 and LY379268 Selectively Attenu…
http://slidepdf.com/reader/full/jayne-cartmell-et-al-the-metabotropic-glutamate-23-receptor-agonists-ly354740 7/10
other tests (Table 2). However, the dose of 10 mg/kg s.c. of clozapine significantly inhibited rotorod performance by 52%
(Table 2). Furthermore, haloperidol produced pronouncedmotor impairment (75% decreases in time on the rotorod) atdoses of 0.1, 0.3, or 1 mg/kg s.c. (Table 2).
Discussion
These studies examined PCP- and AMP-evoked motor ac-tivities in an automated motor behavior assessment systemin which different parameters (ambulatory and nonambula-tory/fine movements) were used. Our dose-response datawith PCP and AMP indicate that this system discriminatesbetween increases in locomotion or ambulations and otherbehaviors such as stereotypes that are not necessarily asso-
ciated with increased locomotion. For example, unlike ambu-lations, which were maximal at 3 mg/kg, fine movements did
not peak until higher doses of PCP (8 mg/kg). This differencemight be due to the onset of ataxia at doses greater than 5mg/kg PCP, resulting in limited ambulatory movements of the animals, but accentuating the expression of certain ste-reotypes (e.g., oral movements and head weaving). In anycase, the reduction of time spent at rest, which was observedat all doses of PCP, indicates that the animals were stillactive to a comparable degree at the higher doses whenambulations had diminished. The difference between thesemeasurements was even more pronounced with AMP, be-cause ambulations peaked at 3 mg/kg, but were not signifi-cantly different from control animals at 30 mg/kg. However,
TABLE 1
Effect of compounds on basal activity
Compound Total Ambulations Time at Rest Fine
MovementsRearing Activity
Vehicle control 171 25 3280 63 698 89 79 25LY354740
0.1 mg/kg 175 53 3231 87 723 171 66 291 mg/kg 184 22 3192 30 758 59 91 1210 mg/kg 133 26 3329 36 523 70 37 11
LY3792680.3 mg/kg 167 26 3174 48 817 85 59 17
1 mg/kg 83 22* 3402 23* 369 38* 11 33 mg/kg 16 8* 3496 22* 222 49* 1 1
Vehicle control 130 26 3364 42 447 78 26 8Clozapine
1 mg/kg 134 8 3316 20 549 42 21 33 mg/kg 158 18 3254 15 604 41 14 310 mg/kg 169 55 3269 84 526 121 9 7
Haloperidol0.03 mg/kg 155 32 3292 87 559 152 45 50.1 mg/kg 32 14* 3517 38* 163 67* 7 6*0.3 mg/kg 1 1* 3574 11* 61 25* 0 0*
* P .05 when compared with corresponding vehicle control animals.
Fig. 7. Effect of various doses of cloza-pine and haloperidol (milligrams per
kilogram) on AMP (3 mg/kg)-evokedbehaviors, as measured in an auto-mated locomotor behavior monitor.Compounds were administered (s.c.)30 min before AMP s.c. injection; be-haviors were monitored over a 60-mintime period immediately after injec-tion of AMP. Data (mean S.E.) arepresented as the total number of be-haviors expressed during 60 min; n 4 rats. * P .05, when compared withthe corresponding vehicle control (V).
1999 Metabotropic Glutamate Agonists Block PCP, not d -Amphetamine 167

8/3/2019 Jayne Cartmell et al- The Metabotropic Glutamate 2/3 Receptor Agonists LY354740 and LY379268 Selectively Attenu…
http://slidepdf.com/reader/full/jayne-cartmell-et-al-the-metabotropic-glutamate-23-receptor-agonists-ly354740 8/10
AMP increases in fine movements and reductions in time atrest were still present at the 30 mg/kg dose. In our drugstudies, we generally used doses of 5 mg/kg PCP and 3 mg/kg
AMP, because these doses gave robust increases in each of the three parameters measured for both agents (ambula-tions, fine movements, and time at rest).
It should be noted that our study used an automated motorbehavioral system that may not fully discriminate betweenthe specific motor behaviors induced by PCP or AMP. Fur-thermore, in addition to stimulation of motor activities inrodents, PCP also induces nonstimulant drug actions, whichinclude disruption of cognition and sensory gating functions
(prepulse inhibition of startle reflexes; Geyer and Braff,1987), and these effects of PCP are thought to be relevant toantipsychotic drug actions. Moreover, the acute actions of PCP in animals, although suggested as a useful model forstudying mechanisms of antipsychotic drug actions, have notyet been validated as a model of human schizophrenia (Halber-stadt, 1995; Thornberg and Saklad, 1996; Jentsch and Roth,1999). However, LY354740 (10 mg/kg i.p.) has been reported toreverse PCP-induced disruption of working memory and stereo-typy, as well as locomotion in rats (Moghaddam and Adams,1998). This indicates that suppression of PCP-induced motoractivations may be a useful and predictive test system for initialevaluation of novel agents against PCP behaviors. In any case,this requires further investigation of these agents in other PCPmodels, and, ultimately, clinical studies are required to validateany animal model.
All compounds examined in this study reduced ambula-tions and fine motor movements evoked by 5 mg/kg PCP withthe relative potency order of haloperidol LY379268 clo-zapine LY354740. Thus, in the present model, haloperidolwas highly potent, and our work supports earlier reports byothers (Del Rio and Fuentes, 1969; Ljungberg and Unger-stedt, 1985) who showed that similar doses of haloperidolreduced locomotor activity and stereotypies evoked by AMP.Haloperidol also potently blocked the actions of PCP in thisstudy. However, these doses of haloperidol greatly impaired
animals, as indicated by both decreased spontaneous basalmotor activities and severe impairments on the rotorod.Thus, the nonselective properties of haloperidol on differentaspects of stimulated and basal motor functions make itdifficult to ascribe antipsychotic activities to haloperidolbased on these data.
In contrast to haloperidol, clozapine selectively inhibitedthe actions of PCP at doses that did not produce motorimpairment. Interestingly, AMP fine movements were en-hanced by lower doses of clozapine. Clozapine augmentationof AMP behaviors has been reported in other studies as well.Robertson and MacDonald (1984) showed that clozapine, atdoses of 5 and 10 mg/kg, reduced locomotor activity in re-sponse to 2.5 mg/kg AMP, but increased the expression of stereotypies. Similarly, in another study, although haloperi-dol reduced all behaviors evoked by 2 mg/kg AMP, clozapineantagonized AMP-induced locomotion, but increased certainstereotypic behaviors (Ljungberg and Ungerstedt, 1985). Incontrast, Tschanz and Rebec (1989) showed that clozapine, atdoses of 1 and 5 mg/kg, did block some types of stereotypies,such as sniffing and oral movements, evoked by 1 mg/kg
AMP. Thus, it would seem that the effects of clozapine on
AMP-evoked stereotypic behaviors are dependent on the typeof stereotypy assessed. Although clozapine failed to reducefine motor movements in the current model, it is possible thatthe compound could have reduced oral stereotypic behaviorsthat were not specifically differentiated by our monitoringsystem. In any case, the lack of pronounced dose-relatedclozapine inhibition of fine movements evoked by AMP werein contrast to the potent reduction of behaviors by haloperi-dol. It has been suggested that the occurrence of extrapyra-midal side effects of neuroleptics in the clinic can be predictedby their ability to inhibit certain AMP-induced behaviorsindicative of interference of striatal dopaminergic neuro-transmission. Thus, the selective actions of mGlu2/3 agonistsand clozapine against AMP suggests that mGlu2/3 agonists,
like clozapine, may have a low risk of extrapyramidal sideeffects.
Furthermore, like clozapine, LY354740 and LY379268 hadselective actions on PCP-evoked motor activations. LY379268was more potent when compared with LY354740 in these be-havioral studies. At doses that greatly reduced the actions of PCP, LY354740 was without effect on either spontanous motoractivities or rotorod performance. However, the more potentcompound LY379268 did reduce basal motor activities at 1 and3 mg/kg, doses that were also highly effective against PCP. Thereasons for this difference between LY354740 and LY379268are not clear, but might be related to subtle differences inmGlu2 versus mGlu3 receptor selectivity. In binding and func-tional studies in vitro, LY354740 has a 2- to 6-fold selectivity formGlu2 versus mGlu3 receptors, whereas LY379268 has aboutequal activity at both receptors (Monn et al., 1999). Neverthe-less, at these doses of LY379268, the animals’ motor abilitieswere not impaired, as demonstrated by normal performance onthe rotorod and the ability of AMP to stimulate motor activity,albeit to a somewhat lesser degree, in the presence of the drug.Importantly, the effects of LY379268 (but not clozapine) on PCPwere completely reversed by the selective mGlu2/3 receptorantagonist, LY341495, clearly indicating that, unlike clozapine,the effects of LY379268 in the PCP model are mediated viagroup II mGlu receptors.
Previous studies have reported that direct intracerebral
TABLE 2
Effect of compounds on rotorod performance
Compound Seconds onRotorod
Vehicle control 60 0LY354740
0.01 mg/kg 60 00.1 mg/kg 60 01 mg/kg 60 03 mg/kg 59 1
10 mg/kg 60 0LY379268
0.003 mg/kg 49 110.03 mg/kg 60 00.3 mg/kg 55 41 mg/kg 57 33 mg/kg 53 7
Clozapine0.3 mg/kg 60 01 mg/kg 57 33 mg/kg 56 410 mg/kg 29 11*
Haloperidol0.03 mg/kg 60 00.1 mg/kg 13 9*0.3 mg/kg 8 3*1 mg/kg 4 2*
* P .05 when compared with vehicle control animals.
168 Cartmell et al. Vol. 291

8/3/2019 Jayne Cartmell et al- The Metabotropic Glutamate 2/3 Receptor Agonists LY354740 and LY379268 Selectively Attenu…
http://slidepdf.com/reader/full/jayne-cartmell-et-al-the-metabotropic-glutamate-23-receptor-agonists-ly354740 9/10
injections of less selective mGlu2/3 receptor agonists canproduce motor impairments. Kronthaler and Schmidt (1996,1998) showed that infusion of the nonselective mGlu receptoragonists, 1 S,3 R-1-aminocyclopentane-1,3-dicarboxylic acid(1 S,3 R-ACPD) or (2 S,3 S,4 S)--carboxycyclopropylglycine,into the lateral ventricle of rats resulted in a moderate levelof motor impairment described as catalepsy. However, wefound no evidence for catalepsy in LY354740- or LY379268-treated animals. Furthermore, LY354740 has been reportedto block haloperidol-induced catalepsy in rats at doses of 5and 10 mg/kg (Konieczny et al., 1998), suggesting thatmGlu2/3 agonists reverse rather than induce drug-inducedParkinsons symptoms. Collectively, these data suggest that1 S,3 R-ACPD and (2 S,3 S,4 S)--carboxycyclopropylglycine in-duction of catalepsy is mediated by actions at mGlu subtypesother than mGlu2/3 receptors.
At doses that greatly suppressed PCP-induced behaviors,LY354740 and LY379268 had minimal effects on PCP-evokedincreases in ambulations and fine movements. However, likeclozapine, LY354740 and LY379268 greatly reduced AMP-evoked rearing activity. This suggests that this behaviormight be mediated by a mechanism distinct from that under-
lying AMP-evoked locomotion. Interestingly, Attarian and Amalric (1997) have shown that the less selective mGlu ag-onist 1 S,3 R-ACPD potentiates locomotor hyperactivityevoked by amphetamine. Kim and Vezina (1997) have re-ported that injection of relatively low doses of the group I/IImGlu receptor antagonist, ( RS)--methyl-4-carboxyphenylg-lycine (MCPG) into the rat nucleus accumbens augmentsamphetamine-induced locomotion. In contrast, higher con-centrations of MCPG reversed the increase in locomotion byamphetamine. These biphasic effects of MCPG might be in-dicative of the dose-dependent recruitment of distinct sub-groups of mGlu receptors. Our data would suggest that thisincrease in AMP-evoked locomotor activity might be medi-ated via group I, rather than group II mGlu receptors.
In summary, LY354740 and LY379268 effectively reversedPCP-evoked motor activations, at doses that had minimaleffects on AMP-induced locomotor behaviors, and no grossimpairment of the animals’ motor abilities. Because the ef-fects of LY354740 and LY379268 in this study more closelyresemble clozapine than haloperidol, this indicates thatmGlu2/3 receptor agonists, like clozapine, may possess anti-psychotic actions with a low risk of extrapyramidal side ef-fects. Furthermore, the robust inhibitions of PCP behaviorsby these novel compounds, but their more selective effects on
AMP behaviors, provides a basis for further investigation of the neurochemical mechanism(s) by which mGlu2/3 agonistsexert their behavioral actions.
Acknowledgments
We thank Dr. Joseph P. Tizzano for his helpful discussions in thecourse of this work, Bryan G. Johnson and Craig R. Salhoff forperforming the high-dose PCP experiment, and Michael Kinder(Hamilton Kinder Behavioral Testing Systems) for technical exper-tise and advice.
References
Adams B and Moghaddam B (1998) Corticolimbic dopamine neurotransmission istemporally dissociated from the cognitive and locomotor effects of phencyclidine. J Neurosci 18:5545–5554.
Anis NA, Berry SC, Burton NR and Lodge D (1983) The dissociative anesthetics,ketamine and phencyclidine, selectively reduce excitation of central mammalianneurones by N -methyl-aspartate. Br J Pharmacol 79:565–575.
Attarian S and Amalric M (1997) Microinfusion of the metabotropic glutamatereceptor agonist 1 S,3 R-1-aminocyclopentane-1,3-dicarboxylic acid into the nucleusaccumbens induces dopamine-dependent locomotor activation in the rat. Eur J Neurosci 9:809–816.
Battaglia G, Monn JA and Schoepp DD (1997) In vivo inhibition of veratridine-evoked release of striatal excitatory amino acids by the group II metabotropicglutamate receptor agonist LY354740 in rats. Neurosci Lett 229:161–164.
Bond A, O’Neill MJ, Hicks CA, Monn JA and Lodge D (1998) Neuroprotective effectsof a systemically active group II metabotropic glutamate receptor agonistLY354740 in a gerbil model of global ischemia. Neuroreport 9:1191–1193.
Chou T-C and Talalay P (1983) Analysis of combined drug effects: A new look at a very old problem. Trends Pharmacol Sci 11:450–454.
Conn PJ and Pin J-P (1997) Pharmacology and functions of metabotropic glutamatereceptors. Annu Rev Pharmacol Toxicol 37:205–237.Danysz W, Parsons CG, Bresink I and Quack G (1995) Glutamate in CNS disorders.
Drug News Perspect 8:261–277.Del Rio J and Fuentes JA (1969) Further studies on the antagonism of stereotyped
behaviour induced by amphetamine. Eur J Pharmacol 8:73–78.Dube GR and Marshall KC (1997) Modulation of excitatory synaptic transmission in
locus coeruleus by multiple presynaptic metabotropic glutamate receptors. Neu-roscience 80:511–521.
Forsythe ID and Barnes-Davies M (1997) Synaptic transmission: Well placed mod-ulators. Curr Biol 7:R362–R365.
Geyer MA and Braff DL (1987) Startle habituation and sensorimotor gating inschizophrenia and related animal models. Schizophr Bull 13:643–668.
Halberstadt AL (1995) The phencyclidine-glutamate model of schizophrenia. Clin Neuropharmacol 18:237–249.
Helton DR, Schoepp DD, Monn JA, Tizzano JP and Kallman MJ (1998a) A role formetabotropic glutamate receptors in drug withdrawal states, in MetabotropicGlutamate Receptors and Brain Function (Moroni F, Nicoletti F, Pellegrini-Giampietro DE eds) pp 305–314, Portland Press Ltd, London, UK.
Helton DR, Tizzano JP, Monn JA, Schoepp DD and Kallman MJ (1997) LY354740: A
potent, orally active, highly selective metabotropic glutamate receptor agonistwhich ameliorates symptoms of nicotine withdrawal. Neuropharmacology 36:1511–1516.
Helton DR, Tizzano JP, Monn JA, Schoepp DD and Kallman MJ (1998b): Anxiolyticand side-effect profile of LY354740: A potent, highly selective, orally active agonistfor group II metabotropic glutamate receptors. J Pharmacol Exp Ther 284:651–660.
Hernandez L, Auerbach S and Hoebel, BG (1988) Phencyclidine (PCP) injected in thenucleus accumbens increases extracellular dopamine and serotonin as measuredby microdialysis. Life Sci 42:1713–1723.
Hondo H, Yonezawa Y, Nakahara T, Nakamura K, Hirano M, Uchimura H andTashiro N (1994) Effect of phencyclidine on dopamine release in the rat prefrontalcortex; an in vivo microdialysis study. Brain Res 633:337–342.
Iversen LL (1975) Dopamine receptors in the brain. Science (Wash DC) 188:1084–1089.
Javitt DC and Zukin SR (1991) Recent advances in the phencyclidine model of schizophrenia. Am J Psychiatry 148:1301–1308.
Jentsch JD and Roth RH (1999) The neuropsychopharmacology of phencyclidine:From NMDA receptor hypofunction to the dopamine hypothesis of schizophrenia. Neuropsychopharmacology 20:201–225.
Kilbride J, Huang L-Q, Rowan MJ and Anwyl R (1998) Presynaptic inhibitory actionof group II metabotropic glutamate receptor agonists, LY354740 and DCG-IV. Eur J Pharmacol 356:149–157.
Kim JH and Vezina P (1997) Activation of metabotropic glutamate receptors in therat nucleus accumbens increases locomotor activity in a dopamine-dependentmanner. J Pharmacol Exp Ther 283:962–968.
Kingston AE, Ornstein PL, Wright RA, Johnson BG, Mayne NG, Burnett JP, Bel-agaje R, Wu S and Schoepp DD (1998) LY341495 is a nanomolar potent andselective antagonist of group II metabotropic glutamate receptors. Neuropharma-cology 37:1–12.
Knopfel T and Gasparini F (1996) Metabotropic glutamate receptors: Potential drugtargets. Drug Dev Today 1:103–108.
Kronthaler UO and Schmidt WJ (1996) 1S,3R-ACPD has cataleptogenic effects andreverses MK-801-and less pronounced, D,L-amphetamine-induced locomotion. Eur J Pharmacol 316:129–136.
Kronthaler UO and Schmidt WJ (1998) The mGluRs group II agonist (2 S,3 S,4 S)-alpha-carboxycyclopropyl-glycine induces catalepsy in the rat, which is pro-nouncedly anatgonised by dizoclipine and D,L-amphetamine. Neurosci Lett 253:25–28.
Kuczenski R and Segal D (1989) Concomitant characterization of behavioral and
striatal neurotransmitter response to amphetamine using in vivo microdialysis. J Neurosci 9:2051–2065.Lillrank SM, O’Connor WT, Saransaari P and Ungerstedt U (1994) In vivo effects of
local and systemic phencyclidine on the extracellular levels of catecholamines andtransmitter amino acids in the dorsolateral striatum of anaesthitized rats. Acta Physiol Scand 150:109–115.
Ljungberg T and Ungerstedt U (1985) A rapid and simple behavioural screeningmethod for simultaneous assessment of limbic and striatal blocking effects of neuroleptic drugs. Pharmacol Biochem Behav 23:479–485.
Lovinger DM and McCool BA (1995) Metabotropic glutamate receptor-mediatedpreysnaptic depression at corticostriatal synapses involves mGluR2 or 3. J Neu-rophysiol 73:1076–1083.
Macek TA, Winder DG, Gereau RW, Ladd CO and Conn PJ (1996) Differentialinvolvement of group II and group III mGluRs as autoreceptors at lateral andmedial perforant path synapses. J Neurophysiol 76:3798–3806.
Marek GJ and Aghajanian GK (1998) The electrophysiology of prefrontal serotoninsystems: Therapeutic implications for mood and psychosis. Biol Psychiatry 44:1118–1127.
1999 Metabotropic Glutamate Agonists Block PCP, not d -Amphetamine 169

8/3/2019 Jayne Cartmell et al- The Metabotropic Glutamate 2/3 Receptor Agonists LY354740 and LY379268 Selectively Attenu…
http://slidepdf.com/reader/full/jayne-cartmell-et-al-the-metabotropic-glutamate-23-receptor-agonists-ly354740 10/10
Moghaddam B and Adams BW (1998) Reversal of phencyclidine effects by a group IImetabotropic glutamate receptor agonists in rats. Science (Wash DC) 281:1349–1352.
Monn JA, Valli MJ, Massey SM, Hansen MM, Kress TJ, Wepsiec JP, Harkness AR,Grutsch JL, Wright RA, Johnson BG, Andis SL, Kingston A, Tomlinson R, LewisR, Griffey KR, Tizzano JP and Schoepp DD (1999) Synthesis, pharmacologicalcharacterization and molecular modeling of heterobicyclic amino acids related toLY354740: Identification of two new potent, selective and systemically activeagonists for group II metabotropic glutamate receptors. J Med Chem 42:1027–1040.
Monn JA, Valli MJ, Massey SM, Wright RA, Salhoff CR, Johnson BG, Howe T, AltCA, Rhodes GA, Robey RL, Griffey KR, Tizzano JP, Kallman MJ, Helton DR andSchoepp DD (1997) Design, synthesis and pharmacological characterization of ()-2-aminobicyclo[3.1.0]hexane-2,6-dicarboxylate: A potent, selective, systemi-cally active group 2 metabotropic glutamate receptor agonist possessing anticon-
vulsant and anxiolytic properties. J Med Chem 40:528–537.Mueller K (1993) Locomotor stereotypy is produced by methylphenidate and am-
fonelic acid and reduced by haloperidol but not clozapine or thioridazine. Phar-macol Biochem Behav 45:71–76.
Murray TF and Horita A (1979) Phencyclidine-induced stereotyped behavior in rats:Dose response effects and antagonism by neuroleptics. Life Sci 24:2217–2226.
Nakanishi S (1992) Molecular diversity of glutamate receptors and implications forbrain function. Science (Wash DC) 258:597–603.
Neki A, Ohishi H, Kaneko T, Shigemoto R, Nakanishi S and Mizuno N (1996) Pre-and postsynaptic localization of a metabotropic glutamate receptor, mGluR2, inthe rat brain: An immunohistochemical study with a monoclonal antibody. Neu-rosci Lett 202:197–200.
Neugebauer V, Keele NB and Shinnick-Gallagher P (1997) Epileptogenesis in vivoenhances the sensitivity of inhibitory presynaptic metabotropic glutamate recep-tors in basolateral amygdala neurons in vitro. J Neurosci 17:983–995.
Nicoletti F, Bruno V, Copani A, Casabona G and Knopfel T (1996) Metabotropicglutamate receptors: A new target for the therapy of neurodegenerative disorders?Trends Neurosci 19:267–271.
Ohishi H, Ogawa-Meguro R, Shigemoto R, Kaneko T, Nakanishi S and Mizuno N(1994) Immunohistochemical localization of metabotropic glutamate receptors,mGluR2 and mGluR3, in rat cerebellar cortex. Neuron 13:55–66.
Ohishi H, Shigemoto R, Nakanishi S and Mizuno N (1993a) Distribution of themessenger RNA for a metabotropic glutamate receptor, mGluR2, in the centralnervous system of the rat. Neuroscience 53:1009–1018.
Ohishi H, Shigemoto R, Nakanishi S and Mizuno N (1993b) Distribution of themRNA for a metabotropic glutamate receptor (mGluR3) in the rat brain: An in situhybridization study. J Comp Neurol 335:252–266.
Ornstein PL, Bleisch TJ, Arnold MB, Wright RA, Johnson BG and Schoepp DD(1998) 2-Substituted 2 SR-2-amino-2-(1 SR,2 SR-2-carboxycyclopropan-1-yl)gly-
cines as potent and selective antagonists of group 2 metabotropic glutamatereceptors. Part 1: Effects of alkyl, arylalkyl and diarylalkyl substitution. J MedChem 41:346–357.
Konieczny J, Ossowska K, Wolfarth S and Pilc A (1998) LY354740, a group IImetabotropic glutamate receptor agonist with potential antiparkinsonian proper-ties in rats. Naunyn-Schmiedeberg’s Arch Pharmacol 358:500–502.
Pin JP and Duvoisin R (1995) The metabotropic glutamate receptors: Structure andfunctions. Neuropharmacology 34:1–26.
Robertson A and MacDonald C (1984) Atypical neuroleptics clozapine and thiorida-zine enhance amphetamine-induced stereotypy. Pharmacol Biochem Behav 21:97–101.
Scanziani M, Salin PA, Vogt KE, Malenka RC and Nicoll RA (1997) Use-dependent
increases in glutamate concentration activate presynaptic metabotropic glutamatereceptors. Nature (Lond) 385:630–634.Schoepp DD, Johnson BG, Wright RA, Salhoff CR, Mayne NG, Wu S, Cockerham SL,
Burnett JP, Belagaje R, Bleakman D and Monn JA (1997) LY354740 is a potentand highly selective group II metabotropic glutamate receptor agonist in cellsexpressing human glutamate receptors. Neuropharmacology 36:1–11.
Seeman P (1995) Dopamine receptors: Clinical correlates, in Psychopharmacology:The Fourth Generation of Progress (Bloom FE and Kupfer DJ eds) pp 295–302,Raven Press, New York.
Seiden LS, Sabol KE and Ricaurte GA (1993) Amphetamine: Effects on catechol-amine systems and behavior. Annu Rev Pharmacol Toxicol 32:639–677.
Shigemoto R, Kinoshita A, Wada E, Nomura S, Ohishi H, Takada M, Flor PJ, Neki A, Abe T, Nakanishi S and Mizuno N (1997) Differential presynaptic localizationsof metabotropic glutamate receptor subtypes in the rat hippocampus. J Neurosci17:7503–7522.
Steinpreis RE (1996) The behavioral and neurochemical effects of phencyclidine inhumans and animals: Some implications for modeling psychosis. Behavioral Brain Res 74:45–55.
Sturgeon RD, Fessler RG, Meltzer HY (1979) Behavioral rating scales for assessingphencyclidine-induced locomotor activity, stereotyped behavior and ataxia in rats.
Eur J Pharmacol 59:169–179.Thornberg SA and Saklad SR (1996) A review of NMDA receptors and the phencyc-
lidine model of schizophrenia. Pharmacotherapy 16:82–93.Tschanz JT and Rebec GV (1989) Atypical antipsychotic drugs block selective com-
ponents of amphetamine-induced stereotypy. Pharmacol Biochem Behav 31:519–522.
Send reprint requests to: Dr. Darryle D. Schoepp, Neuroscience Research,Lilly Research Laboratories, Eli Lilly and Co., Lilly Corporate Center, Drop0510, Indianapolis, IN 46285. E-mail: [email protected]
170 Cartmell et al. Vol. 291