journal of neurovirology (1997) 3, 1997 journal of ...1)/49-61.pdf · differential tropism of...
TRANSCRIPT

Differential tropism of pseudorabies virus for sensoryneurons in the cat
J Patrick Card1,2, Lynn W Enquist4, Alan D Miller5 and Bill J Yates1,3
Departments of 1Neuroscience, 2Psychiatry, and 3Otolaryngology, University of Pittsburgh, Pittsburgh, Pennsylvania15260; 4Department of Molecular Biology, Princeton University, Princeton, New Jersey 08544; 5Laboratory ofNeurophysiology, The Rockefeller University, New York, NY, 10021, USA
The permissiveness of cat motor and sensory systems to infection bypseudorabies virus (PRV) was examined. Eight adult cats of both sexes receivedinjections of a virulent strain of PRV (PRV-Becker) into either the diaphragm orneck musculature. Temporal analysis of the replication and transynapticpassage of virus in each experimental paradigm revealed that sensory neuronsin the dorsal root ganglia were more susceptible to infection than motorneurons. Only scattered motor neurons displayed productive replication ofvirus at postinoculation intervals extending to 192 h whereas robustreplication of virus in neurons in the dorsal root ganglia and dorsal horn ofthe spinal cord was apparent as early as 96 h post inoculation. The datademonstrate that functionally distinct populations of neurons in the cat aredifferentially permissive to infection and transneuronal transport of PRV.
Keywords: alpha herpesvirus; transynaptic infection; dorsal root ganglia; cat
Introduction
Many alpha herpesviruses are characterized by theirneurotropism and ability to infect and spreadwithin populations of synaptically-linked neurons.Recognition of these properties and the recentdemonstrations of the specificity of transneuronalpassage of these viruses has led their use fordefinition of multisynaptic circuits (Strick andCard, 1992; Card and Enquist, 1994; Enquist,1994; Loewy, 1995; Ugolini, 1995). Thus, themechanisms governing the replication and transportof these pathogenic virions within individualneurons and through a multisynaptic circuit haveimportant implications for interpreting data derivedfrom this experimental approach. To date, themajority of tract tracing studies have used eitherthe human pathogen herpes simplex virus (HSV) ora swine alpha herpesvirus known as pseudorabiesvirus (PRV). Although both viruses have beenshown to replicate in a variety of circuits, severalreports described differential invasiveness of func-tionally distinct components of a circuit. As early as1938, Sabin recognized that PRV spread throughtrigeminal and autonomic circuitry of the mousequite efficiently following nasal inoculation, butwas apparently incapable of invading the olfactory
system (Sabin, 1938). It has subsequently beenshown that some, but not all, strains of PRV havethe capacity to invade the CNS via the olfactorynerve in young pigs (McFerran and Dow, 1965;Sabo et al, 1969; Wittmann et al, 1980; Kritas et al,1995) and other examples of differential invasive-ness of PRV or HSV have been identified. Forexample, Rotto-Percelay and colleagues (1992)reported that neurons in the sympathetic compo-nent of the autonomic nervous system are moresusceptible to infection by an attenuated strain ofPRV than motor neurons innervating somaticmusculature. Similarly, we have demonstrated thatfunctionally distinct classes of retinal ganglion cellsrespond differentially to infection with geneticallydefined isogenic PRV mutants (Card et al, 1991,1992; Whealy et al, 1993; Enquist et al, 1994).Barnett and colleagues (1993) demonstrated thatHSV type 1 and mouse hepatitis virus producedifferent patterns of central infection followingidentical inoculation of the nasal cavity or olfactorybulb, and Ugolini (1992) has shown that the rate ofHSV-1 transport to the spinal cord through periph-eral nerves of adult rats occurs faster in neuronswith smaller caliber axons. In addition, Zemanickand colleagues (1991) have reported that one strainof HSV is transported anterogradely after injectioninto the motor strip of primate cortex while anotherstrain is only transported in the retrograde directionafter identical injection. These and other examples
Correspondence: J Patrick CardReceived 6 June 1996; revised 14 August 1996; accepted31 August 1996
Journal of NeuroVirology (1997) 3, 49 ± 61ãhttp://www.jneurovirol.com
1997 Journal of NeuroVirology, Inc.

of differential viral tropism vividly demonstrate thatneuroinvasiveness is strain dependent and is animportant consideration in the use of alphaherpesviruses for the characterization of multi-synaptic circuits.
Pseudorabies virus is remarkable for the widerange of animals that it can infect. It causes a naturaldisease in some domestic and wild animals(Wittmann and Rziha, 1989) and, if introduced byinjection, can infect a more diverse population ofmammals and some birds (Nara, 1982). It isnoteworthy that it cannot infect humans, apes,chimpanzees, reptiles and insects by any route.Natural infections seem to occur most often inswine, cattle, sheep, dogs and cats (Gustafson,1975). However, in a given species, young animalsare often more susceptible to PRV disease than areadult animals (Wittmann and Rziha, 1989). Sincethe anatomy and physiology of neural systems incats are in many instances well established, it is ofparticular interest to explore the utility of usingPRV for transneuronal analysis in this species.
In the present investigation we have analyzed theinvasiveness and patterns of viral replication thatoccur in the spinal cord following injection of PRV-Becker into the diaphragm or neck musculature ofadult cats. Dobbins and Feldman (1994) havecharacterized the pattern of transneuronal infectionin the rat produced by injection of PRV intodiaphragm and, as noted earlier, Rotto-Percelayand co-workers (1992) have analysed the invasive-ness of this strain of virus in rat motor circuitry.Thus, there is a solid foundation for evaluating theutility of this experimental approach in the catnervous system. Furthermore, the two systems offercomplementary approaches for analysis of factorsinfluencing viral tropism and invasiveness. Thediaphragm is characterized by a diffuse motorinnervation and few sensory afferents (Balkowiecet al, 1995; Corda et al, 1965; Duron et al, 1978;Gottschall, 1981) whereas the neck musculature isrich in sensory afferents and the motor endplatesare more densely concentrated than in the dia-phragm (Richmond and Abrams, 1975). The datademonstrate that functionally distinct componentsof the cat nervous system are differentially suscep-tible to the productive replication of a virulentstrain of PRV following peripheral inoculation.
Results
Peripheral inoculation of adult cats with a virulentstrain of PRV initially isolated by Becker (PRV-Becker; Becker, 1967) produced an immunohisto-chemically detectable productive infection in thespinal cord of all animals. However, the magnitudeand pattern of infection was directly dependentupon the route of inoculation, the amount ofinjected virus and the postinoculation interval.
Only scattered infected motor neurons were ob-served in the ventral horn of the spinal cord at postinoculation intervals extending to 192 h followinginjection of PRV-Becker into the diaphragm. Incontrast, large numbers of neurons displaying viralantigens were observed in the dorsal root gangliaand dorsal horn of the spinal cord followinginjection of the virus into neck musculature asearly as 96 h post inoculation. The permissivenessof animals to infection did not correlate with thesex of the animal.
Temporal aspects of cellular infectionInfected neurons characteristically exhibited mod-erate to dense immunoreactivity associated withboth the cell soma and proximal dendrites, with theintensity and intracellular distribution of viralantigen reflecting the apparent duration of viralreplication. For example, the earliest immunoreac-tivity was observed in the nucleus of infectedneurons, while continued viral replication resultedin the appearance of viral immunoreactivity in thesomata and dendrites of infected cells (e.g.,compare Figure 3b and c). The early appearance ofviral antigen in the cell nucleus correlates withDNA replication and packaging whereas the punc-tate staining that progressively appears across thesomatodendritic compartment denotes sites ofenvelopement, intracellular trafficking, and transy-naptic passage of virions (Card et al, 1993; Whealyet al, 1991).
Diaphragm injectionsOnly scattered infected neurons were observed inthe animals in which virus was injected into thediaphragm and all of these cells were confined tothe ventral horn of the spinal cord. The distributionof infected neurons observed in these animals ismapped on Figure 1 and photomicrographs ofrepresentative examples are illustrated in Figure 2.The largest number of immunoreactive neurons wasobserved in the longest surviving animals (Cases D4& D5; see Tables 1 and 2). These animals wereinoculated with 0.6 ± 1.26107 pfu of virus andallowed to survive 192 h. In both cases, weobserved scattered infected motor neurons thatwere occassionally associated with small immuno-reactive interneurons (Figure 2). The only differ-ence between these cases was the volume of theinoculum; case D4 was injected with 35 ml and caseD5 received 70 ml of virus. At shorter postinocula-tion intervals the number of infected neurons wassubstantially reduced, even though larger concen-trations and volumes of virus were injected into thediaphragm. Only one immunopositive neuron wasobserved in 151 sections from C5 and C6 of theanimal that was inoculated with 172 ml of virus(2.96107 pfu) and allowed to survive 144 h. Sec-tions of C5 and C6 spinal cord from the tworemaining animals injected with PRV-Becker
Pseudorabies virus invasiveness in catJP Card et al
50

revealed a total of 4 and 14 neurons in a total of 85and 158 sections, respectively (Table 2). Thisincluded one animal inoculated with 172 ml(2.96107 pfu) that was sacrificed 96 h post-inocu-lation and another that received 3.46107 in 201 mland was permitted to survive 72 h. Thus, post-inoculation interval was the most important vari-able in eliciting a productive infection in thisexperimental paradigm.
Only 76 neurons were observed in a total of 548coronal sections though the C5 and C6 levels of thespinal cord (Table 2). The majority of these neuronswere confined to the inferior limit of the ventralhorn, midway along its mediolateral axis (Figure 1).They displayed two distinct morphologies and,when more than one cell was present in a singlesection, they were usually found in clusters. Eachcluster consisted of a large multipolar motor neuronsurrounded by small bipolar neurons (Figure 2a ±d). The distribution of immunoreactivity in theseneurons was consistent with intermediate toadvanced stages of viral replication and there wereno cytopathic changes in the infected cells. Norwere there signs of the characteristic infection ofglial that has been shown to be associated with lyticchanges in neurons infected with PRV-Becker andother strains of PRV after chronic infection of rodentCNS (Rinaman et al, 1993). Case D2 demonstratedthat infection of the large motor neurons alwaysoccurred prior to infection of the small neurons andwe never observed infected neurons in the dorsalhorn of spinal cord in any of the animals. Theseobservations suggest that infection of smallerneurons in the ventral horn resulted from retrogradetransynaptic passage of virus from the first orderinfection in motor neurons.
The large neurons displaying viral immunoreac-tivity exhibited a morphology consistent with motorneurons previously shown to innervate the catdiaphragm (Rose et al, 1983). The cells exhibited awidest cross sectional diameter of approximately
Figure 1 The patterns of infection produced in cervical levels 3and 5 (C3 & C5) of the spinal cord following injection of thediaphragm (C5) and neck musculature (C3) are illustrated. Eachillustration reflects the relative distribution of infected neuronsobserved in each paradigm; absolute numbers of infectedneurons in the different cases are presented in Table 2. Opentriangles reflect the position of infected propriospinal neuronsand filled triangles mark the location of infected motor neurons.The filled circles illustrate the distribution of infected neuronsin laminae I and II of the dorsal horn whereas the small opencircles in the ventral horn illustrate the relative position ofinfected interneurons. Diagrams are adapted from those pub-lished by Rexed (1954).
Table 1 Experimental animals
Level of
Vol.Inj Spinal
ml Conc.Inj. Survival Cord
Animal Virus virus/cat pfu6107
(hours) Analyzed
D1
D2
D3
D4
D5
N1
N2
N3
PRV-Becker
PRV-Becker
PRV-Becker
PRV-Becker
PRV-Becker
PRV-Becker
PRV-Becker
PRV-Becker
201
172
172
35
70
172
172
70
3.4
2.9
2.9
0.6
1.2
2.9
2.9
1.2
72
144
96
192
192
144
96
120
C2, C3, C5, C6
C5, C6
C5, C6
C5, C6
C5, C6
C1, C3, C4
C3
C1, C3, C4, C5
Details regarding the injection parameters, postinoculation
survival intervals and regional analysis conducted in each
experiment are provided. D=diaphragm injected animal,
N=neck injected animal; C=cervical spinal cord.
Pseudorabies virus invasiveness in catJP Card et al
51

50 mm (range=43 ± 65 mm) and were multipolar inconformation (Figure 2a). In cases in which viralreplication was more advanced, viral immunoreac-tivity was apparent in primary and secondarydendrites (Figure 2a and b). Sections adjacent tothose exhibiting immunopositive motor neuronsoften exhibited immunoreactive dendrites in regionsapproximating the location of the infected perikarya,indicating that the dendritic arbors of infected cellsextended radially from the parent neurons (Figure 2band c). Nevertheless, all immunoreactive dendriteswere confined to the gray matter of the ventral horn.
The small infected neurons surrounding these motorneurons were the sole evidence for transynapticpassage of virus in the diaphragm-injected animals.These cells were much smaller than the larger motorneurons and exhibited a distinctly different mor-phology (Figure 2c and d). The widest diameter ofindividual cells was approximately 15 mm(range=10 ± 22 mm) and the cell somas were slightlyelongated in conformation. In some instances, thepresence of viral antigen within the proximaldendrites demonstrated that these cells were bipolarin morphology (Figure 2c and d).
Table 2 Distribution of infected neurons
Sections
Total # with
Sections infected Total # of Infected Neurons
Animal SC Level Analyzed neurons Laminae I & II PNs Ventral Horn
D1
D2
D3
D4
D5
N1
N2
N3
C5
C6
C5
C6
C5
C5
C5
C3
C4
C3
C3
C4
83
75
73
78
85
56
96
106
45
109
108
41
8
0
1
0
2
10
14
94
26
17
100
21
0
0
0
0
0
0
0
1885
67
18
2052
79
0
0
0
0
0
0
0
5
1
0
9
2
6/8
0
1/0
0
1/3
5/37
9/7
0
0
0
17/119
1/2
Specifics regarding the number and location of infected neurons in each case are provided. The number of neurons in the ventral
horn of each level of spinal cord in represented as follows: motor neurons/interneurons. D=diaphragm, N=neck, SC=spinal cord,
PN=propriospinal neuron.
Figure 2 The morphology of infected neurons in the ventral horn of C5 of the spinal cord produced by injection of PRV-Becker intothe diaphragm is illustrated. First order infection is restricted to large multipolar neurons with diffusely ramifying dendrites (a and b).Portions of the dendritic arbor of these motor neurons are commonly observed in adjacent sections (arrows in b and c). Retrogradetransynaptic passage of virus into the afferents synapsing upon infected motor neurons leads to infection of smaller interneurons in theimmediate vicinity of the motor neurons (c and d). Marker bars for a and b=50 mm; bars for c and d=25 mm.
Pseudorabies virus invasiveness in catJP Card et al
52

Injections of neck musculatureInjection of PRV-Becker into the splenius andbiventer muscles of the neck resulted in a produc-tive infection of neurons at upper levels of cervicalspinal cord in all three experimental animals. Thepatterns of infection in the spinal cord produced byinjection of PRV-Becker into neck musculaturediffered substantially from that produced by dia-phragm injection (Figure 1). All three neck injectedanimals exhibited infected neurons in lamina I andII of the dorsal horn, but rarely exhibited immunor-eactive neurons in the ventral horn (Table 2). Themagnitude of infection differed among animals, butthe pattern was the same in all cases. A summary ofthe distribution of infected neurons observed inthese cases is shown in Figure 1 and the morphologycharacteristically exhibited by these cells is illu-strated in Figures 3 and 5. Absolute numbers ofinfected neurons in these regions of spinal cord are
presented in Tables 2 and 3. After injection of 172 ml(2.96107 pfu) and 96 h survival (case N2) a total of18 infected neurons were present in 17 of 109sections from C3 of the spinal cord and theseneurons were confined to the superficial laminaeipsilateral to the injection. In contrast, 1890 infectedneurons were observed in 94 of 106 sections fromthe same level of spinal cord 144 h followinginjection of an equivalent volume of virus (caseN1, Table 2). The majority of these cells (1885) werefound in the ipsilateral laminae I and II of the dorsalhorn, although five cells with a morphology char-acteristic of propriospinal neurons (see followingsection for description) were present in laminae Vand VI (Figures 3 and 5). The C4 level of the cord inthis animal exhibited the same pattern of infection,but the number of infected neurons was substan-tially reduced (Table 2). No infected neurons wereobserved in the ventral horn of this animal.
Figure 3 Injection of PRV-Becker into the splenius and biventer muscles of the neck led to a preferential infection of neurons inlaminae I, II, V and VII of C3. The general pattern of infection observed in these regions are illustrated in a. Small spherical neurons inlaminae I and II constituted the largest number of infected neurons (a and b), but occasional infected neurons in laminae V and VIwere also observed (c). Marker bar for a=100 mm; bars for b and c=50 mm.
Pseudorabies virus invasiveness in catJP Card et al
53

Case N3, in which the animal was sacrificed120 h after injection of 1.26107 pfu of virus in70 ml, also exhibited a large number of infectedneurons in laminae I and II of the ipsilateral dorsalhorn and occasional infected cells in laminae V andVI; 2052 infected neurons were observed in thesuperficial laminae of C3 and 9 cells exhibitingpropriospinal neuron morphology were present indeeper layers. Again, the pattern of infection in C4was similar to that seen in C3, but the number ofcells was substantially reduced (Table 2). Unlikecase N1, a substantial number of PRV-immunor-eactive neurons were present in the medial seg-ments of laminae I and II and scattered infectedneurons were present in the ventral horn, primarilyat the ventral and medial extent of the gray matter(Figure 1b). The morphology of the latter group ofneurons was similar to both motor neurons andinterneurons, with the number of interneuronsexceeding that of the motor neuron population(Table 2).
The pattern of infection produced in the spinalcord by injection of virus into the splenius andbiventer muscles also exhibited evidence oftopographical segregation of neurons within sub-fields of laminae I and II. This was true of allcases, but was more apparent in the two animalsin which the spinal cord infection was moreadvanced. The principal characteristic of thistopography was the segregation of the majority ofinfected cells within the lateral half of the dorsalhorn (Figures 1, 3a, 5a and b). Furthermore, thepresence of large numbers of infected neurons insome sections, considered with the very lownumber of infected cells in other sections fromthe same segment of spinal cord, suggested thatinfected neurons occurred in groups along thelongitudinal axis of individual spinal cord seg-ments. For example, of the 106 sections from C3that were analyzed in case N1, approximately half(49) had fewer than 10 infected cells per section(Table 3). The remaining sections containing anaverage of 32.33 infected dorsal horn neurons persection and, of that population, less than one cellper section was found in the medial half oflaminae I/II. A similar preponderance of infectedneurons in the lateral half of laminae I and II wasevident in case N3 (Table 3). However, there were
more neurons in the medial portion of laminae Iand II in this case and occasional infected neuronswere observed in the ventral horn (Figure 5a andb; Table 2).
Examination of sections through the C3 dorsalroot ganglion of case N4 ipsilateral to the viralinjection demonstrated numerous infected neurons(Figure 4a). Six hundred and fifty-one infectedneurons were observed in 17 sections through theganglion. These cells exhibited an average crosssectional diameter of 41.37 mm (range=30 ± 61 mm;n=144) and they were distributed throughout theganglion. Photomicrographs illustrating the distri-bution and morphology of the neurons from thiscase are shown in Figure 4a ± c.
The infected neurons in the spinal cord dorsalhorn exhibited two distinct morphologies. Those inlaminae I and II presented a very homogenousmorphology. They were generally spherical inshape and exhibited a widest diameter of approxi-mately 14 mm (range=10 ± 19 mm) (Figure 3b). Onlyon rare occasions did the viral immunoreactivityextend into proximal dendries of these cells and inthose instances they presented a bipolar conforma-tion. Infected neurons in laminae V and VIpresented a more complex morphology (Figure 5aand b). They were substantially larger than infectedneurons in superficial laminae with a widestdiameter of approximately 60 mm (range=52 ±78 mm). In addition, viral immunoreactivity routi-nely extended well into the dendritic tree of theseneuons revealing three to five primary dendritesthat radiated away from the cell soma and gave riseto smaller branching secondary and tertiary den-drites. Apical branches of the larger dendritesextended dorsolaterally into laminae I and II wherethey were often coextensive with infected neuronswithin these regions. The remaining dendritesextended radially from the multipolar perikarya,but exhibited a preferential trajectory back into thegray matter of the cord (Figure 5a and b).
Discussion
While the cat is reported to be a host for PRV (Nara,1982), our data demonstrate a dramatic differencein the permissiveness of motor and sensory systems
Table 3 Distribution of neurons in dorsal horn of C3 after neck injection
Total # # Sections Average Number of Neurons in Laminae II & III
Sections with 510 in Sections with 510 Infected Neurons
Animals Analyzed cells/section Total Medial Lateral
N2
N4
106
108
57
75
32.33
26.79
0.70
3.65
31.63
23.12
Details regarding the number of infected neurons present in different regions of laminae I and II of the C3 level of spinal cord in two
of the neck injected animals (N2 and N4) are provided.
Pseudorabies virus invasiveness in catJP Card et al
54
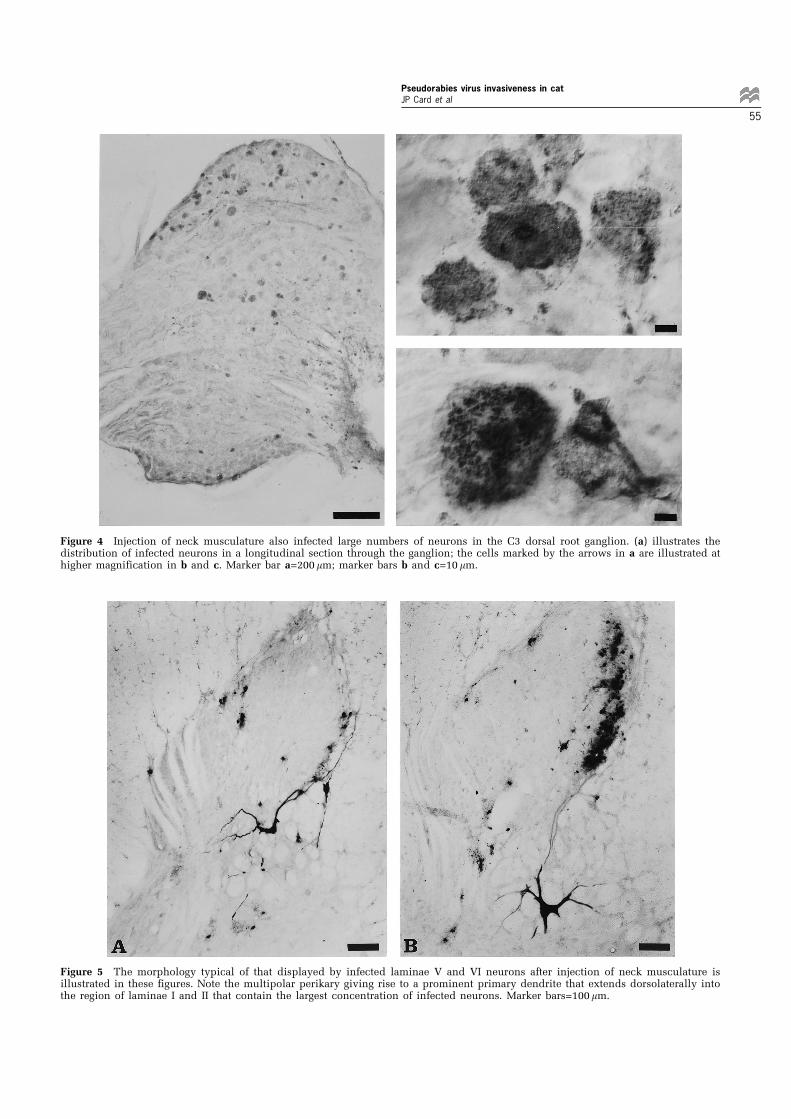
Figure 5 The morphology typical of that displayed by infected laminae V and VI neurons after injection of neck musculature isillustrated in these figures. Note the multipolar perikary giving rise to a prominent primary dendrite that extends dorsolaterally intothe region of laminae I and II that contain the largest concentration of infected neurons. Marker bars=100 mm.
Figure 4 Injection of neck musculature also infected large numbers of neurons in the C3 dorsal root ganglion. (a) illustrates thedistribution of infected neurons in a longitudinal section through the ganglion; the cells marked by the arrows in a are illustrated athigher magnification in b and c. Marker bar a=200 mm; marker bars b and c=10 mm.
Pseudorabies virus invasiveness in catJP Card et al
55

of the cat spinal cord to infection with PRV. Motorneurons innervating both the diaphragm and neckmusculature were relatively resistant to viralinfection following inoculation with large concen-trations of PRV-Becker and long postinoculationintervals. In contrast, small doses of PRV-Beckerand shorter postinoculation intervals produced arobust transneuronal infection of dorsal hornneurons in regions previously shown to receiveprojections from the DRG. Furthermore, the ob-served patterns of viral replication in the DRG andsuperficial laminae of the dorsal horn suggested adifferential tropism of PRV-Becker for DRG neuronswith small caliber axons.
The underlying mechanisms that account for thispreferential tropism of PRV for sensory neurons inthe cat remain to be defined. Nevertheless, someaspects of our findings combined with those ofprevious studies provide interesting insights. PRVinfection of neonatal animals is usually moredevastating than infection of adult animals. More-over, PRV has a predilection to infect sensorypathways in adult animals, often leading to estab-lishment of latency in sensory ganglia (Wittmannand Rziha, 1989). One particularly noteworthyaspect of our data is the fact that productivereplication of virus was largely restricted to sensoryDRG neurons and their synaptic targets in the dorsalhorn of the spinal cord. As noted earlier, followingintranasal inoculation of PRV in adult mice, Sabin(1938) described the preferential spread of virusthrough trigeminal and autonomic pathways, butfailed to observe propagation of the virus inolfactory systems. Although recent studies demon-strate that some strains of PRV can gain access to theCNS via the olfactory nerves, these studies wereconducted in young pigs (McFerran and Dow, 1965;Sabo et al., 1969; Wittmann et al, 1980; Kritas et al,1995). In adult animals, intranasal inoculation ofPRV generally leads to establishment of a latentinfection of sensory trigeminal neurons rather thanproductive replication and invasion of the CNS.Although one must be cautious in generalizingproperties of infection across species and indifferent neuronal systems, our findings are con-sistent with the natural tropism of the virus forsensory fibers in the adult animals. Injection ofvirus into neck musculature produced a robustreplication of virus in the DRG and transynapaticinfection of dorsal horn neurons known to receivesensory input from DRG neurons, whereas onlyoccasional viral replication was observed in motorneurons in these animals and those in which thevirus had been injected into the diaphragm. Theabsence of infection in the dorsal horn followinginjection of the diaphragm is consistent with thedemonstrated sparse distribution of a sensoryinnervation to this muscle. Free sensory endingsand Pacinian corpuscles are widely scattered overthe surface of the muscle and there are few muscle
spindles and Golgi tendon organs that are confinedprimarily to the crural region (Balkowiec et al.,1995; Corda et al., 1965; Duron et al., 1978;Gottschall, 1981). Our paradigm did not involveinjection of the crus.
The infection of the DRG and dorsal horn ofspinal cord in neck injured animals was furtherdistinguished by the differential concentration ofviral antigens in a subset of sensory neurons.Examination of the DRG in case N3 demonstratedthat only a subpopulation of DRG neurons exhibitedviral immunoreactivity and that these cells aredistinguished by their small cross-sectional dia-meter compared to uninfected perikarya. In addi-tion, the transneuronal infection of the synaptictarget of these neurons in the dorsal horn of spinalcord was largely confined to the lateral half oflaminae I and II, and further appeared to besegregated within groups distributed along therostrocaudal extent of the C3 level of spinal cord.All of these observations are consistent with viruspreferentially infecting small caliber sensory axonsinnervating the region of the axial neck musculaturenear the multiple sites of inoculation. Small caliberAd and C fibers arising from small DRG sensoryneurons are known to arborize within the super-ficial laminae of this region of spinal cord in apattern that conforms to the distribution of infectedneurons observed in our analysis (Willis andCoggeshall, 1991). The infected neurons in laminaeV, VI and VII of the C3 level of spinal cord aresimilar in morphology to propriospinal neuronsthought to play an important role in posturaladjustments associated with targeted movements(Alstermark et al, 1987, 1991). These neurons areknown to receive descending inputs from thecortex, tectum and brainstem as well as afferentinput from the forelimb (Illert et al., 1978; Alster-mark et al., 1984). Since these neurons exhibiteddendrites that extended into the lateral extent oflaminae I and II it is probable that they becomeinfected via monosynaptic transneuronal passage ofvirus from sensory afferents.
Although productive replication of PRV-Beckerwas largely confined to a subset of sensory neurons,we did observe a limited number of infected motorneurons. This raises the issue of why we did notobserve more extensive infection of motor andsensory systems following inoculation of eitherdiaphragm or neck musculature. Particularly ger-mane to this discussion are recent studies of thereplication of HSV-1 (Ugolini, 1992) and PRV-Bartha (Rotto-Perceley et al., 1992) in motorcircuitry. Ugolini demonstrated extensive infectionof both sensory, autonomic and motor systemsfollowing injection of HSV-1 into crushed mixednerves innervating the forelimb on hindlimb of rats(Ugolini, 1992). However, the data from that studyalso revealed temporal differences in the rate ofinfection of functionally distinct components of
Pseudorabies virus invasiveness in catJP Card et al
56

these mixed nerves. Productive replication of virusin sensory and autonomic neurons with smallcaliber axons preceded that of motor neurons andsensory neurons with large myelinated axons. Thisfinding suggests that the restricted pattern ofinfection observed in our analysis may reflectdiffering rates of viral replication and/or transportin different types of neurons. However, our findingssuggest that species specificity and a restrictedpermissiveness to infection may have also contri-buted to the preferential infection of sensorysystems in the cat. In both of our paradigms, themajority of motor neurons were consistently resis-tent to productive replication of PRV-Becker evenin circumstances in which animals were injectedwith large concentrations of virus and permitted tosurvive as long as 8 days. While one might arguethat the sparseness of the terminal distributionacross the surface of the diaphragm contributed toour failure to infect substantial numbers of phrenicmotor neurons (i.e., neurons did not accumulateenough virus to elicit a productive infection), thisexplanation does not explain the small number ofinfected motor neurons resulting from injection oflarge amounts of virus into neck musculature.Unfortunately, the sensory irritation that accompa-nied the early infection of small DRG neuronsprecluded anlaysis of longer surviving animals inthis paradigm. Thus, we cannot exclude thepossibility that large numbers of motor neuronswould have become infected at longer post inocula-tion intervals. However, it is also important toconsider the differences in the method of inocula-tion in comparing our findings and those of Ugolini(1992). Whereas Ugolini injected virus directly intocrushed peripheral nerves, we injected virus intoperipheral targets. This suggets that the peripher-ally projecting axons of motor neurons may not beas permissive to virion invasion as those of sensoryterminals. Alternatively, the induction of a produc-tive replication of virus in motor neurons mayrequire higher intracellular concentrations of virus.Resolution of this issue would require injection ofvirus into the neck musculature of animals that hadpreviously received dorsal root section to eliminatetransport of virus into the spinal cord throughsensory fibers.
The possibility that motor neurons are moreresistant to infection by PRV than other systems isalso supported by the findings of Rotto-Percelay andcolleagues (1992). These investigators reported thatthe ability of PRV-Bartha to infect motor neuronsinnervating the gastrocnemius muscle of the rat wasless efficient than the ability of the virus to invadeand replicate within autonomic circuitry. Theexperimental paradigm employed by these investi-gators was similar to that used in our study in thatthey injected virus into multiple sites in the muscle.However, the amount of injected virus was far lessthan used in the present study (16104 pfu versus
1.2 ± 2.96107 pfu) and they used the attenuatedBartha strain rather than the virulent Becker strainof PRV. Using this approach these authors reportedthat they were successful in infecting sympatheticpreganglionic neurons in approximately 90% ofanimals, but only infected ventral horn motorneurons in about 60% of the injected rats. Analysisof the DRGs and sectioning of the dorsal roots in thatstudy also revealed that virus did not repliciate inDRG sensory neurons and they did not observe thepattern of infection in the dorsal horn that we havedocumented in this study. Certainly, some of thedifferences observed in our findings and those ofRotto-Perceley and co-workers can be attributed tothe different strains of virus employed, and thisimportant difference should be carefully consideredin interpreting the data derived from both studiesgiven the results from prior investigations that havedocumented differences in the neuroinvasivenessand replication of PRV-Becker and PRV-Bartha(Card and Enquist, 1995). Neverthelesss, the find-ings of both studies provide compelling evidencethat the tropism of PRV for motor systems issubstantially less than it is for other neuronalsystems.
Although our injections of diaphragm onlyinfected a small number of phrenic motor neuronsthere was limited transynaptic passage of virus fromthese neurons into local circuit premotor neurons.Interneurons with a variety of respiratory-relateddischarge patterns have been recorded duringelectrophysical studies at the same spinal level asthe phrenic motor pool in cats and rabbits (Belling-ham and Lipski, 1990; Douse and Duffin, 1993;Grelot et al, 1993; Palisses et al, 1989). Inhibitoryconnections have been observed between C5 ex-piratory interneurons and phrenic motoneurons(Douse and Duffin, 1993), but the possible connec-tions between other subtypes of C4 ± 6 respiratoryinterneurons and phrenic motoneurons have notbeen examined. Bellington and Lipski (1990) havereported respiratory interneurons are widely dis-tributed around the phrenic nucleus while Dousseand Duffin (1993) described the C5 expiratoryinterneurons being located predominately dor-somedial to the phrenic motor pool. In our analysis,infected ventral horn interneurons were found inthe immediate vicinity of the motor neurons fromwhich they were presumably infected via retrogradetransynaptic passage of virus, a finding that isconsistent with the distribution characterized inrat by Dobbins and Feldman (1994) using transneur-onal tracing with PRV.
In conclusion, we have demonstrated that theneurotropism of PRV places a number of constraintsupon its use for transneuronal analysis in the cat.The initial goal of our study was to examinethe morphological substrates of vestibular influ-ences upon respiratory outflow and of non-respiratory inputs to the phrenic motor pool.
Pseudorabies virus invasiveness in catJP Card et al
57

Previous anaylsis by Yates and colleagues (1993)demonstrated vestibulo-respiratory and vestibulo-sympathetic responses in decerebrate cats, but theprecise organization of neurons that mediate theseresponses has not been defined in this species.Studies of this circuitry in rats by Dobbins andFeldman (1994) had shown that rat phrenic motorneurons are capable of replicating and transportingPRV-Bartha, but did not reveal a monosynapticprojection to these cells from either the medial orlateral vestibular nuclei. Thus, the apparent resis-tance of cat motor neurons to PRV infection,combined with the permissiveness of sensoryneurons to the replication and spread of the virulentBecker strain of PRV was an unexpected findingthat further emphasizes the need for thoroughparametric analysis before using these neurotropicpathogens for transneuronal analysis of neuralcircuitry. The data also provide a vivid example ofhow the tropism of this virus changes in differentspecies.
Materials and methods
VirusThe virulent Becker strain of pseudorabies virus(PRV-Becker; Becker, 1967) was used in thisanalysis. Virus was propagated on PK15 cells aspreviously described (Card and Enquist, 1994) to atiter of 1.76108 plaque forming units per milliliter(pfu/ml).
AntiseraTissue from animals injected with PRV wasprocessed using rabbit polyvalent antisera raisedagainst acetone inactivated virus (Card et al, 1990).In prior studies we demonstrated that these antiseraproduce robust staining of virally infected neuronsand are specific for PRV (Card and Enquist, 1994). Inthe present investigation we incubated sections in a1 : 10 000 dilution of primary antiserum for 24 ±48 h and localized viral immunoreactivity with theavidin-biotin modification (Hsu et al., 1981) ofSternberger's peroxidase-antiperoxidase precedure(Sternberger, 1979). The reagents included affinitypurified donkey antirabbit secondary antisera(Jackson ImmunoResearch Laboratories) and theVectastain Elite Kit (Vector Laboratories).
Experimental animalsEight adult antibody-free cats of both sexes wereused in the analysis. These `Antibody ProfileDefined (APD) Felines' were purchased from HarlanSprague Dawley, Incorporated (Indianapolis, Indi-ana). They are unvaccinated and barrier-reared toassure freedom from disease or humoral antibodiesaffecting disease susceptibility and progression.Upon delivery to the laboratory the animals weremaintained in isolation for a brief period ofacclimation prior to injection of virus. Injected
animals recovered in individual hepa-filtered cagesmaintained within an isolated room equipped witha dedicated air supply. The laboratory and proce-dures used for these experiments met the criteria forBiosafety Level 2 experiments as stipulated inHealth and Human Services Publication #88-8395entitled Biosafety in Microbiological and Biomedi-cal Laboratories. The experimental protocol wasapproved by the Animal Care and Use Committeesat Rockefeller University and the University ofPittsburgh.
Animal surgeryAll surgery was performed under aseptic conditionsin a sterile operating room. Four animals weresedated by intramuscular injection of 0.3 mg/kgacepromazine and 0.15 mg/kg butorphanol. Anintravenous catheter was then inserted and theanimals were maintained under deep sodiumpentobarbital induced anaesthesia. A 25 mg/kgdose was delivered immediately following insertionof the canula; supplementary doses were adminis-tered as needed to maintain areflexia. The otheranimals were anaesthetized using isofluorane va-porized in oxygen; the anesthetic concentration wasadjacent to maintain areflexia during the surgicalprocedures.
Five animals received injections into the leftcostal region of the diaphragm through an incisionin the abdominal wall. Three animals receivedinjections into the splenius and biventer muscles,exposed by an incision through the skin on thedorsal aspect of the left side of the neck at levels C1to C3 of the vertebral column. All animals receivedmultiple injections of 1 to 2 ml into each target tissueto achieve final injected volumes ranging from 35 ±201 ml of virus per animal (5.96106 pfu to3.46107 pfu). This amount of injected virus pro-duces a productive infection in 100% of rodents(Card et al., 1995). After inoculation the skin wassutured and the animals were returned to their cagefor the balance of the experiment. Further informa-tion regarding the amount and type of injectedvirus, the site of injection, the postinoculationinterval, and the regions of spinal cord analyzedin each animal is indicated in Table 1.
Tissue fixation and processingAt the conclusion of each experient animals weredeeply anesthetized with an intraperitoneal injec-tion of sodium pentobarbital (40 mg/kg) and per-fused transcardially with phosphate-buffered salinefollowed by the paraformaldehyde-lysine-sodiummetaperiodate (PLP) fixative developed by McLeanand Nakane (1974). Segments of cervical spinal cordand, in some cases, their associated dorsal rootganglia (DRG) were postfixed for 1 h at 48C, washedin 0.1 M sodium phosphate buffered saline (PBS),cryoprotected by immersion in phosphate buffered20% sucrose solution, and sectioned in the coronal
Pseudorabies virus invasiveness in catJP Card et al
58

plane with a sliding microtome equipped with atissue freezing unit. The spinal cord was cut at 35 mmand dorsal root ganglia (DRG) were cut in thelongitudinal plane at 40 mm per section. Tissue wascollected serially in 12 bins of PBS and stored at 48C.One bin of tissue from each level of spinal cord wasimmediately processed for immunohistochemicallocalization of viral immunoreactivity to determinethe extent of viral replication and transport throughmotor and sensory circuitry in the spinal cord. Theremaining bins were transferred to cryoprotectantand stored at 7208C to preserve antigenicity(Watson et al., 1986). Some of these bins weresubsequently processed for viral immunoreactivityto permit analysis of the distribution of infected cellsin a more frequent series of sections. The minimumfrequency of analysis of each level of spinal cord was420 mm and, in most cases, a more frequent series ofsections was examined.
Analysis of tissueEach case was analyzed to determine both thedistribution and extent of viral replication incoronal sections through the appropriate level ofspinal cord or in longitudinal sections through theDRG. Each section of tissue was examined with aNikon Optiphot photomicroscope to determine therelative distribution and extent of infection. Aprecise map of the distribution and number ofinfected neurons in each section was obtained byrecording the positions of infected neurons on acamera lucida schematic map produced with aNikon Optiphot photomicroscope equipped with adrawing tube. The number of neurons in specificregions of the dorsal and ventral horns were thencounted. The distribution and size of infected
neurons in the C3 dorsal root ganglion of one ofthe long surviving PRV-Becker injected animals(N4; see Table 1) were also determined. Thelocation of infected neurons in seventeen 40 mmsections through the ganglion were mapped using a46objective and the perimeter of each cell wastraced.
The widest diameter of a subset of neurons in theDRG and spinal cord was measured using amicrometer scale and a drawing tube. The scalewas projected through the drawing tube using a206 objective and recorded on a piece of paper.Infected neurons were then identified on thephotomicroscope at the same magnification andthe scale was superimposed upon the cell somausing the drawing tube to determine the widestdiameter. Only infected cells displaying viralimmunoreactivity throughout the cell soma wereincluded in this quantitative analysis. Cells dis-playing pathological changes were excluded.
For the purposes of illustration, the distributionof infected neurons was mapped upon schematicdiagrams of the appropriate level of spinal cordadapted from Rexed (Rexed, 1954). Photomicro-graphs of infected neurons were taken with a ZeissAxiophot photomicroscope equipped with differ-ential interference contrast optics.
Acknowledgements
This work was supported by NIH RO1s MH53574(JPC), NINDS 33506 (LWE), DC00693 (BJY),NS20585 (ADM) and DC02644 (ADM). We grate-fully acknowledge the expert technical assistanceof Jen-Shew Yen.
References
Alstermark B, Isa T, Tantisira B (1991). Integration indescending motor pathways controlling the forelimbin the cat. Experimental Brain Research 84: 561 ± 568.
Alstermark B, Kummel H, Pinter MJ, Tantisira B (1987).Branching and termination of C3-C4 propriospinalneurones in the cervical spinal cord of the cat.Neuroscience Letters 74: 291 ± 296.
Alstermark B, Lundberg A, Sasaki S (1984). Integrationin descending motor pathways controlling the forelimb in the cat. II. Inhibitory pathways from highermotor centres and forelimb afferents to the C3-C4propriospinal neurones. Experimental Brain Research56: 308 ± 322.
Balkowiec A, Kukula K, Szulczyk P (1995). Functionalclassification of afferent phrenic nerve fibers anddiaphragmatic receptors in cats. Journal ofPhysiology (London) 483: 759 ± 768.
Barnett EM, Cassell MD, Perlman S (1993). Twoneurotropic viruses, herpes simplex virus type 1 andmouse hepatitis virus, spread along different neuralpathways from the main olfactory bulb. Neuroscience57: 1007 ± 1025.
Becker CH (1967). Zur primaren Schadingung vegetativerGanglien nach Infektion mit dem Herpes suis Virusbei verschiedenen Tierarten. Experentia 23: 209 ± 217.
Bellingham MC, Lipski J (1990). Respiratory interneuronsin the C5 segment of the spinal cord of the cat. BrainResearch 533: 141 ± 146.
Card JP, Dubin JR, Whealy ME, Enquist LW (1995).Influence of infectious dose upon productivereplication and transynaptic passage of pseudorabiesvirus in rat central nervous system. Journal ofNeuroVirology 1: 349 ± 358.
Card JP, Enquist LW (1994). Use of pseudorabies virusfor definition of synaptically linked populations ofneurons. Methods in Moleculan Genetics 4: 363 ± 382.
Card JP, Enquist LW (1995). Neurovirulence ofpseudorabies virus. Critical Reviews in Neurobiology9: 137 ± 162.
Pseudorabies virus invasiveness in catJP Card et al
59

Card JP, Rinaman L, Lynn RB, Lee B-H, Meade RP,Miselis RR, Enquist LW (1993). Pseudorabies virusinfection of the rat central nervous system:Ultrastructural characterization of viral replication,transport, and pathogenesis. Journal of Neuroscience13: 2515 ± 2539.
Card JP, Rinaman L, Schwaber JS, Miselis RR, WhealyME, Robbins AK, Enquist LW (1990). Neurotropicproperties of pseudorabies virus: Uptake andtransneuronal passage in the rat neuronal system.Journal of Neuroscience 10: 1974 ± 1994.
Card JP, Whealy ME, Robbins AK, Enquist LW (1992).Pseudorabies virus envelope glycoprotein gIinfluences both neurotropism and virulence duringinfection of the rat visual system. Journal of Virology66: 2032 ± 3041.
Card JP, Whealy ME, Robbins AK, Moore RY, EnquistLW (1991). Two alpha-herpesvirus strains aretransported differentially in the rodent visual system.Neuron 6: 957 ± 969.
Corda M, von Euler C, Lennerstrand G (1965).Proprioceptive innervation of the diaphragm. Journalof Physiology (London) 178: 161 ± 177.
Dobbins EG, Feldman JL (1994). Brainstem networkcontrolling descending drive to phrenic motoneuronsin rat. Journal of Comparative Neurology 347: 64 ± 86.
Douse MA, Duffin J (1993). Axonal projections andsynaptic connections of C5 segment expiratoryinterneurones in the cat. Journal of Physiology(London) 470: 431 ± 444.
Duron B, Jung-Caillol MC, Marlot D (1978). Myelinatednerve fiber supply and muscle spindles in therespiratory muscles of the cat: a quantitative study.Anatomy and Embryology 152: 171 ± 192.
Enquist LW (1994). Infection of the mammalian nervoussystem by pseudorabies virus (PRV). Seminars inVirology 5: 221 ± 231.
Enquist LW, Card JP (1996). Pseudorabies virus: A toolfor tracing neuronal connections. In: Protocols forGene Transfer in Neuroscience. Towards GeneTherapy of Neurological Disorders, Lowenstein PR,Enquist LW (eds) John Wiley & Sons: Chichester.pp{ 333 ± 348.
Enquist LW, Dubin J, Whealy ME, Card JP (1994).Complementation analysis of pseudorabies virus gEand gI mutants in retinal ganglion cell neurotropism.Journal of Virology 68: 5275 ± 5279.
Gottschall J (1981). The diaphragm of the rat and itsinnervation. Muscle fiber composition, perikarya andaxons of efferent and afferent neurons. Anatomy andEmbryology 161: 405 ± 417.
Grelot L, Milano S, Portillo F, Miller AD (1993).Respiratory interneurons of the lower cervical (C4-C5) cord: membrane potential change during fictivecoughing, vomiting, and swallowing in thedecerebrate cat. Pflugers Archives 425: 313 ± 320.
Gustafson DP (1975). Pseudorabies. In: Diseases ofSwine, Dunne HW, Leman AD (eds) Ames: The IowaState University Press. pp 391 ± 410.
Hsu SM, Raine L, Fanger H (1981). The use of avidin-biotin-peroxidase complex (ABC) in immuno-peroxidase techniques: a comparison between ABCand unlabeled antibody (PAP) procedures. Journal ofHistochemistry and Cytochemistry 29: 577 ± 580.
Illert M, Lundberg A, Padel Y, Tanaka R (1978).Integration in descending motor pathways controllingthe forelimb in the cat. 5. Properties of andmonosynaptic excitatory convergence on C3-C4propriospinal neurones. Experimental Brain Research33: 101 ± 130.
Kritas SK, Nauwynck JJ, Pensaert MB (1995).Dissemination of wild-type and gC-, gE- and gI-deleted mutants of Aujeszky's disease virus in themaxillary nerve and trigeminal ganglion of pigs afterintranasal inoculation. Journal of General Virology 76:2063 ± 2066.
Loewy AD (1995). Pseudorabies virus: A transneuronaltracer for neuronanatomical studies. In: Viral Vectors,Kaplitt MG, Loewy AD (eds) Academic Press: SanDiego. pp. 349 ± 366.
McFerran JB, Dow C (1965). The distribution of the virusof Aujeszky's disease (pseudorabies virus) inexperimentally infected swine. Americal Journal ofVeterinary Research 26: 631 ± 635.
McLean IW, Nakane PK (1974). Periodate-lysine-paraformaldehyde fixative. A new fixative forimmunoelectron microscopy. Journal ofHistochemistry and Cytochemistry 22: 1077 ± 1083.
Nara PL (1982). Porcine herpesvirus. I. In: ComparativePathobiology of Viral Disease, Olson JR, Krakown S,Blakeslee JJR (eds) CRC Press: Boca Raton. pp 90 ±113.
Palisses R, Persegol L, Viala D (1989). Evidence forrespiratory interneurones in the C3-C5 cervical spinalcord in the decorticate rabbit. Experimental BrainResearch 78: 624 ± 632.
Rexed B (1954). A cytoarchitctonic atlas of the spinalcord in the cat. Journal of Comparative Neurology100: 297 ± 380.
Richmond FJR, Abrams VC (1975). Morphology anddistribution of muscle spindles in dorsal muscles ofthe cat neck. Journal of Neurophysiology 38: 1322 ±1339.
Rinaman L, Card JP, Enquist LW (1993). Spatiotemporalresponses of astrocytes, ramified microglia, and brainmacrophages to central neuronal infection withpseudorabies virus. Journal of Neuroscience 13:685 ± 702.
Rose D, Larnicol N, Marlot D, Duron B (1983).Quantitative morphological changes in phrenic andintercostal motor columns and their respective spinalcord segments during postnatal development in thekitten. Neuroscience Letters, 40, 119 ± 125.
Rotto-Perceley DM, Wheeler JG, Osorio FA, Platt KB,Loewy AD. (1992). Transneuronal labeling of spinalinterneurons and sympathetic preganglionic neuronsafter pseudorabies virus injections in the rat medialgastrocnemius muscle. Brain Research 574, 291 ± 306.
Sabin AB (1938). Progression of different nasally instilledviruses along different nervous system pathways inthe same host. Proceedings of the Society forExperimental Biology and Medicine 38: 270 ± 275.
Sabo A, Rajcani J, Blaskovic D (1969). Studies on thepathogenesis of Aujesky's disease. III. The distributionof virulent virus in piglets after intranasal infection.Acta Virologica, 13: 407 ± 414.
Pseudorabies virus invasiveness in catJP Card et al
60

Sternberger LA (1979). Immunohistochemistry, 2ndedition, New York: Wiley and Sons.
Strick PL, Card JP (1992). Transneuronal mapping ofneural circuits with alpha herpesviruses. In:Experimental Neuroanatomy. A Practical ApproachOxford University Press: Oxford, pp. 81 ± 101.
Ugolini G (1992). Transneuronal transfer of herpessimplex virus type 1 (HSV 1) from mixed limbnerves to the CNS. I. Sequence of transfer fromsensory, motor, and sympathetic nerve fibres to thespinal cord. Journal of Comparative Neurology 326:527 ± 548.
Ugolini G (1995). Transneuronal tracing with Alpha-herpesviruses: A review of the methodology. In: ViralVectors. Gene Therapy and NeuroscienceApplications, pp. 293 ± 318.
Willis WD, Coggeshall RE (1991). Sensory Mechanisms ofthe Spinal Cord Plenum Press: New York.
Watson RE, Wiegand ST, Clough RW, Hoffman GE(1986). Use of cryoprotectant to maintain long-termpeptide immunoreactivity and tissue morphology.Peptides 7: 155 ± 159.
Whealy ME, Card JP, Meade RP, Robbins AK, EnquistLW (1991). Effect of Brefeldin A on alphaherpesvirusmembrane protein glycosylation and virus egress.Journal of Virology 65: 1066 ± 1081.
Whealy ME, Card JP, Robbins AK, Dubin JR, Rziha H-J,Enquist LW (1993). Specific pseudorabies virusinfection of the rat visual system requires both gIand gp63 glycoproteins. Journal of Virology 67: 3786 ±3797.
Wittmann G, Jakubik J, Ahl R (1980). Multiplication anddistribution of Aujesky's disease (pseudorabies) virusin vaccinated and non-vaccinated pigs after intranasalinfection. Journal of Virology 66: 227 ± 240.
Wittmann G, Rziha HJ (1989). Aujesky's disease(pseudorabies) in pigs. In: Herpesvirus Diseases ofCattle, Horses and Pigs, Wittman G (ed) Kluwer:Boston. pp. 230 ± 325.
Yates BJ, Jakus J, Miller AD (1993). Vestibular effects onrespiratory outflow in the decerebrate cat. BrainResearch 629: 209 ± 217.
Zemanick MC, Strick PL, Dix RD (1991). Direction oftransneuronal transport of herpes simplex virus 1 inthe primate motor system is strain independent. PNASUSA 88: 8048 ± 8051.
Pseudorabies virus invasiveness in catJP Card et al
61