melatonin enhances the sensitivity of circadian pacemakers to light in the nocturnal field mouse mus...
TRANSCRIPT

Melatonin Enhances the Sensitivity of CircadianPacemakers to Light in the Nocturnal FieldMouse Mus booduga
VIJAY KUMAR SHARMA1n, R. CHIDAMBARAM2, andA. KASIN YADUNANDAM2
1Chronobiology Laboratory, Evolutionary and Organismal Biology Unit,Jawaharlal Nehru Centre for Advanced Scientific Research, Jakkur,Bangalore- 560 064, Karnataka, India2Department of Animal Behaviour and Physiology, School of Biological Sciences,Madurai Kamaraj University, Madurai, Tamil Nadu, India
ABSTRACT The effect of exogenous melatonin (1 mg/kg) on light pulse (LP) induced phaseshifts of the circadian locomotor activity rhythm was studied in the nocturnal field mouse Musbooduga. Three phase response curves (PRCs: LP, control, and experimental) were constructed tostudy the effect of co-administration of light and melatonin at various circadian times (CTs). The LPPRC was constructed by exposing animals free-running in constant darkness (DD) to LPs of 100–luxintensity and 15–min duration, at various CTs. The control and experimental PRCs wereconstructed by using a single injection of either 50% DMSO or melatonin (1 mg/kg dissolved in50% DMSO), respectively, administered 5 min before LPs, to animals free-running in DD. A singledose of melatonin significantly modified the waveform of the LP PRC. The experimental PRC hadsignificantly larger areas under advance and delay regions of the PRC compared to the control PRC.This was also confirmed when the phase shifts obtained at various CTs were compared between thethree PRCs. The phase delays at three phases (CT12, CT14, and CT16) of the experimental PRCswere significantly greater than those of the control and the LP PRCs. Based on these results weconclude that phase shifting effects of melatonin and light add up to produce larger responses.J. Exp. Zool. 297A:160–168, 2003. r 2003 Wiley-Liss, Inc.
INTRODUCTION
In nature, a vast majority of biological processesfollow a 24–h pattern. When assayed underconstant conditions of the laboratory, these pro-cesses occur with periodicities close to 24–h(circadian rhythms). The synchronization of thesebiological processes to geophysical cycles is be-lieved to occur due to simultaneous actions of anumber of cyclic biotic and abiotic factors such aslight/darkness (LD), temperature, humidity, cyclicsocial interactions, and food availability cycles.Besides these external factors, a number ofinternal factors could also act as time cues(Zeitgebers) for the circadian clocks (Armstrong,’89). Melatonin, which coordinates various beha-vioral and physiological processes, has beenhypothesized to be one such internal Zeitgeberfor the circadian timing systems of vertebrates(Armstrong, ’89). In mammals, melatonin issynthesized and secreted in a rhythmic manner
by the pineal gland. Its levels begin to rise in theevening, and remain high throughout the night,and are regulated both by the circadian clocks andthe LD cycles (Reiter, ’87). Moreover, the sensi-tivity of circadian clocks, measured as phaseresponse curves (PRC; a plot of phase shifts inthe circadian rhythm as a function of phase atwhich the stimulus is presented) to externallyadministered melatonin, also varies periodically inmammals (for review see Redman, ’97). A periodicvariation in sensitivity to melatonin injectionshas also been reported in a number of species ofmammals (Redman, ’97).
A number of rodents can be entrained to dailymelatonin injections (Redman, ’97). Timely ad-
nCorrespondence to: Vijay Kumar Sharma, Chronobiology Labora-tory, Evolutionary and Organismal Biology Unit, Jawaharlal NehruCentre for Advanced Scientific Research, Jakkur, PO Box-6436,Bangalore- 560 064, Karnataka, India. E-mail: [email protected]
Received 8 October 2002; Accepted 10 February 2003Published online in Wiley InterScience (www.interscience.wiley.
com). DOI:10.1002/jez.a.10265
r 2003 WILEY-LISS, INC.
JOURNAL OF EXPERIMENTAL ZOOLOGY 297A:160–168 (2003)

ministration of melatonin to rodents acceleratedre-entrainment to phase shifted LD cycles (Red-man, ’97; Sharma et al., ’99). The suprachiasmaticnucleus (SCN) of mammals regulates a number ofcircadian rhythms such as wheel-running, feeding,drinking, plasma corticosterone and growth hor-mone levels, heart rate, learning, sleep-wake cycle,pineal serotonin, and N-acetyltransferase rhythms(Moore, ’81; Klein et al., ’91). Externally adminis-tered melatonin altered the metabolic and elec-trical activities of the SCN (Cassone et al., ’88;Mason and Brooks, ’88; Shibata et al., ’89) andphase shifted the firing of rat SCN neurons invitro (McArthur et al., ’91). These studies suggestthat the circadian clocks of mammals are sensitiveto melatonin. However, in hamsters, a single doseof melatonin had no effect on the free-runningactivity rhythm when administered without hand-ling the animals (Hastings et al., ’92; Yellon andHilliker, ’94).In the C3H/HeN mice, a single dose of melato-
nin did not alter the phase of the circadianlocomotor activity rhythm (Benloucif and Dubo-covich, ’96). However, when it was administeredalong with a brief exposure to light, the resultantphase delay in the locomotor activity rhythm wasconsistently smaller than the phase delays pro-duced by light alone (Dubocovich et al., ’96;Benloucif et al., ’99). The authors conclude thatcombined administration of melatonin and lightcauses phase shifts in the circadian clocks in amanner that cannot be predicted based on simpleaddition of melatonin and LP PRCs (Benloucifet al., ’99). However, it remains unclear how asingle dose of melatonin, which was unable tocause any measurable phase shift on its own, canreduce the phase shifts evoked by light. In studiesof the effects of melatonin on the locomotoractivity rhythm of the nocturnal field mouse, M.booduga, single injections of melatonin (1 mg/kg)induced phase delays when administered at phasesbetween CT2 and CT19, and induced phaseadvances when given between CT19 and CT2(Sharma et al., ’99). In this species, the LP PRCconstructed using LPs of 1000-lux intensity and15-min duration had regions of phase delaysbetween CT10 and CT18, and phase advancesbetween CT18 and CT10 (Sharma et al., ’97). Itwould therefore be interesting to investigate theeffect of a single dose of melatonin on the lightinduced phase shifts in these animals, as theyrespond to a single dose of melatonin withsignificant non-zero phase shifts in the circadianrhythms. Further, the melatonin PRC of M.
booduga (with a single dose of melatonin) hadsimilar waveform but different time course com-pared to the melatonin PRC of C3H/HeN mice (forthree doses on three successive cycles) (Benloucifand Dubocovich, ’96; Sharma et al., ’99). Unlikethe C3H/HeN mice, the nocturnal mouse M.booduga is a tropical rodent, and experimentalanimals were caught from the wild only a few daysprior to the experiments. Therefore, an extensivestudy of the combined effect of melatonin and lighton the circadian clocks of this species of nocturnalmammal could help us understand the interac-tions between photic and nonphotic Zeitgebers.
In this paper, we report the results of ourexperiments on the nocturnal field mouse M.booduga. We constructed three PRCs; the LPPRC was constructed by administering LPs of100–lux intensity and 15–min, given at variouscircadian times (CTs), to animals free-running inconstant darkness (DD). The other two PRCs wereconstructed using vehicle (50% DMSO) or melato-nin injections (1 mg/kg dissolved in 50% DMSO),administered 5-min before the LPs of 100–luxintensity and 15–min duration. DMSO was used todissolve melatonin because the melatonin PRCsconstructed earlier (Sharma et al., ’99), which willbe used to compare the present PRCs, wereconstructed using melatonin dissolved in 50%DMSO. The results of our experiments suggestthat melatonin enhances the LP induced phaseshifts in the circadian locomotor activity rhythmof the nocturnal field mouse M. booduga.
MATERIALS AND METHODS
Adult male field mice M. booduga (n¼206) werecaptured from the fields surrounding the MaduraiKamaraj University campus (91 58’ N 781 10’ E).At this latitude, most days are sunny with clearskies. The animals were initially maintained in LD12:12 h for about 15 days and then introduced tocages equipped with running wheels. An eccen-trically placed magnet in the running wheelenabled the recording of wheel revolutions on anEsterline Angus event recorder (A 620� ). Micefeed consisting of grains and water were availablead libitum. The animals were housed in DDconditions in a light-tight, adequately ventilatedexperimental room, allowing their circadianrhythms to free-run. Battery-powered torchlightwith a combination of filters transmitting only redlight of wavelength exceeding 650 nm was used forfeeding and cleaning purposes for short intervals.The activity patterns of 20 mice in separate
LIGHT AND MELATONIN PRCS 161

running wheels, placed on open shelves in theexperimental room, could be assayed concurrently.In this setup we have never observed any obviousinteraction between animals with respect toinfluences on circadian rhythms, either in anyLD regime or in DD. Temperature (251C711C)and humidity (70%75%) in the experimentalrooms were recorded with a thermohygrograph(Wilh. Lambrecht KG, Gottingen, Germany) andwere found to be nearly constant throughout theexperiment.Three PRCs (LP PRC, control PRC, experi-
mental PRC) were constructed by exposing adultmale mice (M. booduga) to fluorescent LPs ofapproximately 100–lux intensity and 15–minduration, and LPs in combination with 50%DMSO and melatonin (1 mg/kg), after about 15days of free-run in DD. The LP PRC wasconstructed by exposing (n¼68) animals to LPsat all even CTs between CT2 and CT24. Thecontrol PRC was constructed by exposing (n¼66)animals to LPs in combination with injection of50% DMSO. The experimental PRC was con-structed by exposing (n¼72) animals to LPs incombination with melatonin injection (1 mg/kgdissolved in 50% DMSO). The desired phases (CT)were calculated from the prior stable free-runningrhythm in DD. To construct the control andexperimental PRCs, DMSO and melatonin wereadministered 5–min before the LPs. In all thethree PRC experiments, five to six animals wereused at each CT. To set the light intensity at about100-lux, the light probe of a UDT optometer wasplaced inside the plexiglass box and its experi-mental position relative to the light source wasadjusted. After the administration of the stimuli,cages were returned to the dark rooms. Free-running period (t) was computed using regressionlines through 10–15 onsets of locomotor activitypreceding the stimuli and through 10–15 onsets ofactivity after the stimuli, leaving out the firstthree onsets to allow transients to subside. Phase-shifts and CT are expressed in circadian hours(i.e.,� t/24) throughout the text. Each mouse wasused only once in an experiment. Melatoninobtained from Sigma Chemicals Co., (St Louis,MO) was dissolved in 50% DMSO. Golombek andCardinali’s (’93) protocol for the preparation anddosage of melatonin was followed in the presentstudy.The PRCs were compared against each other
using the phase shift values and the areas underdelay (D) and advance (A) zones of the PRCs. Theareas under advance and delay portions of the
PRCs were chosen to compare the PRCs inaddition to the phase shifts because entrainmentmechanisms can be better reflected by the com-plete time-course and waveform of the PRC(Pittendrigh and Daan, ’76; Johnson, ’95). The Aand D values of the two PRCs (DMSO andmelatonin PRCs) reported in one of our earlierpapers (Sharma et al., ’99) were used in additionto the data on three PRCs obtained in the presentexperiments for the comparison between the fivePRCs (DMSO PRC, melatonin PRC, LP PRC,control PRC, and experimental PRC). The experi-mental conditions in the present study weresimilar to those in our previous study, allowingmeaning comparison of the PRCs (Sharma et al.,’99). The DMSO PRC was obtained by injecting50% DMSO to animals at various CTs, whereasthe melatonin PRC was constructed using mela-tonin (1 mg/kg) dissolved in 50% DMSO (Sharmaet al., ’99). In order to get an estimate of thevariation in D and A within each PRC experiment,the following scheme of resampling was followed(Sharma et al., ’97; Sharma et al., ’99). From thefive or six data points at each phase, available ineach experiment (DMSO PRC, melatonin PRC, LPPRC, control PRC and experimental PRC) one waschosen at random and a PRC was constructedusing the 12 data points chosen. This samplingprocess was repeated, with replacement, twelvetimes, thus generating 12 PRCs per experiment,which, in turn, yielded twelve values of D and Afor each PRC experiment. These twelve values ofD and A per experiment were used to constructconfidence intervals about the estimated mean ofD and A for each experiment. These 60 values of Dand A were used as data for analysis of variance(ANOVA), to test for any significant effect of thePRC type (DMSO PRC, melatonin PRC, LP PRC,control PRC, and experimental PRC). The PRCswere also compared using phase shift dataobtained at various phases of the circadian cycle.Two-way mixed model analysis of variance (ANO-VA), treating PRC experiment and phase as fixedfactors, was used to compare the three PRCs (LPPRC, control PRC, and experimental PRC). Post-hoc multiple comparisons of the phase shifts andA and D values were done using Tukey’s test.
RESULTS
The waveform and time course of all the threePRCs (LP, control, and experimental PRCs)resembled the general shape of a standard LPPRC, with phase delays during the early subjective
V.K. SHARMA ET AL.162

night (about CT12–CT18) and phase advancesduring the late subjective night (about CT18–CT24). The PRCs had no region which could becharacterized as a true ‘‘dead zone.’’ The LP PRCconstructed using fluorescent LPs of 100–luxintensity and 15–min duration had regions ofphase delays ranging between CT10–CT19 andphase advances between CT19–CT2 (Fig. 1).Maximum phase delays and maximum phaseadvances of �1.4670.31 h (mean7SEM) andþ0.8870.31 h were obtained by exposing animalsto LPs at CT14 and CT22 respectively (Fig. 1). Thecontrol PRC was constructed by estimating phaseshifting effects of the vehicle injections (50%DMSO), administered 5–min before LPs (100 luxintensity and 15 min duration) (Fig. 2). Thecontrol PRC had regions of phase delays rangingbetween CT12–CT18 and phase advances betweenCT20–CT24 (Fig. 2).Maximum phase delays and maximum phase
advances of �1.7070.10 h (mean7SEM) andþ0.7070.09 h were obtained at CT14 and CT22
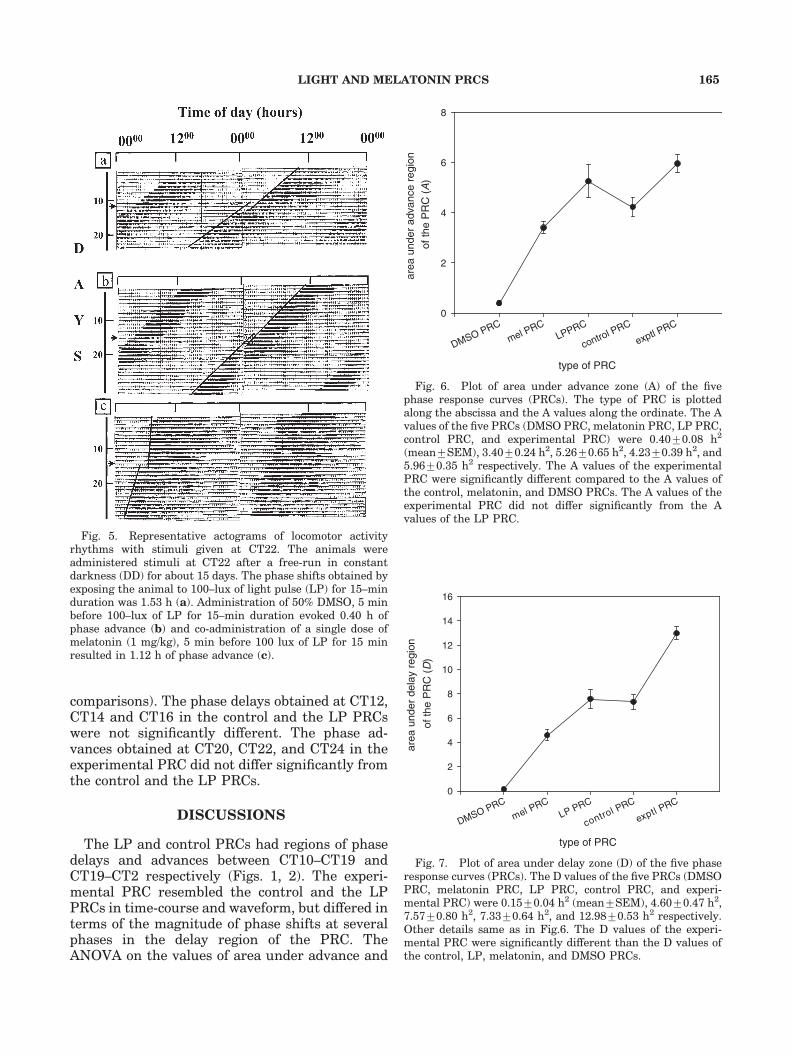
respectively (Fig. 2). The experimental PRC,constructed using melatonin (1 mg/kg dissolvedin 50% DMSO), administered 5-min before LP,had phase delays at phases ranging between CT10and CT20 and phase advances between CT20 andCT10 (Fig. 3). Maximum phase delays and max-imum phase advances of �2.3470.25 h (mean7SEM) and þ0.8270.09 h were obtained at CT14and CT22 respectively (Fig. 3). The activity recordsshowing phase delaying and phase advancingresponses of one animal each for various combina-tions of stimuli at phases of maximum delayand maximum advances are illustrated in Figs. 4and 5.
The areas under advance (A) and delay (D) zonesof the LP PRC were 5.2670.65 h2 (mean7SEM)and 7.5770.80 h2 respectively as compared tothe same for the control PRC (A¼4.2370.39h2;D¼7.3370.64 h2) and experimental PRC (A¼5.9670.35h2; D¼12.9870.53h2). The areas underadvance (A) and delay (D) zones of the DMSO
0 6 12 18 24
phas
e sh
ifts
(hou
rs)
-2
-1
0
1
2
circadian time
Fig. 1. Light pulse phase response curve (LP PRC)constructed using phase shifts induced by a single LP of100–lux intensity and 15–min duration, given at variouscircadian times (CTs). The phases of LP exposure wereestimated using regression lines through the onsets oflocomotor activity (defined as CT12, i.e., onset of subjectivenight). The error bars represent standard error around themean (SEM). Exposure to LPs during early and late subjectivenights evoked phase delays and phase advances respectively,with LPs at CT14 and CT22 causing maximum phase delays of–1.4670.31 h (mean7SEM) and maximum phase advances ofþ0.8870.31 h. The areas under advance (A) and delay (D)zones of the LP PRC were 5.2670.65 h2 (mean7SEM) and7.5770.80 h2 respectively.
0 6 12 18 24
phas
e sh
ifts
(hou
rs)
-2
-1
0
1
2
circadian time
Fig. 2. Phase response curve constructed using 50% DMSOand light pulse (LP) (control PRC). Animals free-running inconstant darkness (DD) were injected 50% DMSO, 5–minbefore the LP, given at various phases of the circadian cycles.Co-administration of DMSO and LP evoked PRC of similartime course and waveform as the LP PRC. The areas underadvance (A) and delay (D) zones of the control PRC were4.2370.39 h2 (mean7SEM) and 7.3370.64 h2 respectively.The A and D of the control PRC did not differ significantlyfrom the A and D of the LP PRC. Maximum phase delays andmaximum phase advances of �1.7070.10 h (mean7SEM)and þ0.7070.09 h were obtained by exposing the animals toLPs at CT14 and CT22 respectively. Other details same asin Fig. 1.
LIGHT AND MELATONIN PRCS 163

and melatonin PRCs were (A¼0.4070.08h2;D¼0.1570.04h2) and (A¼3.4070.24h2; D¼4.6070.47h2) respectively (Sharma et al., ’99a). TheANOVA on the A values of the five PRCs revealed asignificant main effect of the PRC type (F4,55¼30.29, po0.0001; Fig. 6). Post-hoc multiple com-parisons using Tukey’s test showed that the A ofthe experimental PRC was significantly greaterthan the A of the control PRC (po0.03). The Aof the LP PRC was not significantly different fromthe A of the experimental and the control PRCs.The ANOVA on the D values of the five PRCsrevealed a significant main effect of the PRC type(F4,55¼70.39, po0.0001; Fig. 7). Multiple compar-isons using Tukey’s test showed that the D of theexperimental PRC was significantly greater thanthe D of the control and the LP PRCs (po0.0001,for both comparisons, Fig. 7). The D of the LP PRCwas not significantly different from the D of thecontrol PRC (p¼0.99), thus the A and D values ofthe experimental PRC were significantly greater
than A and D values of the DMSO, melatonin, andcontrol PRCs, and the D of the LP PRC (po0.0001for all comparisons).
The ANOVA on the phase shift data obtained inthe LP, the control, and the experimental PRCexperiments revealed significant effects of phase(F11,144¼16.24, po0.0001; Figs. 1–3) and interac-tion between PRC type and phase (F22,144¼1.81,po0.02). Post-hoc multiple comparisons usingTukey’s test revealed that phase delays obtainedin the experimental PRC at CT12 (�2.1670.14 h:mean7SEM), CT14 (�2.3470.25 h) and CT16(�1.6370.38 h) were significantly larger in mag-nitude compared to the phase delays obtainedat corresponding phases in the control (�0.3270.29 h, �1.7070.10 h, �0.9770.33 h respectively)and the LP (�0.2570.25h, �1.4670.31h, �1.1470.44 h respectively) PRCs ( po0.01 for all the
0 6 12 18 24
phas
e sh
ifts
(hou
rs)
-2
-1
0
1
2
circadian time
Fig. 3. Phase response curve evoked by co-administrationof melatonin and light pulse (LP) (experimental PRC).Animals free-running in constant darkness (DD) wereadministered a single dose of melatonin (1 mg/kg), 5 minbefore LP, at various circadian times (CTs). The area underphase advance (A) and phase delay (D) regions of theexperimental PRC were 5.9670.35 h2 (mean7SEM) and12.6270.53 h2 respectively. The A and the D of theexperimental PRC were significantly greater than the A andD of the control PRC. The maximum phase delay andmaximum phase advances of �2.3470.25 h (mean7SEM)and þ0.8270.09 h were obtained by exposing the animals tostimuli between CT14 and CT22 respectively. Other detailssame as in Figure 1.
Fig. 4. Representative actograms of locomotor activityrhythm with stimuli given at CT14. The animals wereadministered stimuli at CT14 after a free-run in DD for about15 days. The phase shifts obtained by exposing the animal to100–lux of light pulse (LP) for 15–min duration was �1.16 h(a). Administration of 50% DMSO, 5 min before LP of 100 luxintensity for 15 min duration evoked a phase delay of �1.62 h(b) and co-administration of a single dose of melatonin (1 mg/kg), 5 min before LP of 100 lux intensity for 15 min resulted in�2.14 h of phase delay (c).
V.K. SHARMA ET AL.164
comparisons). The phase delays obtained at CT12,CT14 and CT16 in the control and the LP PRCswere not significantly different. The phase ad-vances obtained at CT20, CT22, and CT24 in theexperimental PRC did not differ significantly fromthe control and the LP PRCs.
DISCUSSIONS
The LP and control PRCs had regions of phasedelays and advances between CT10–CT19 andCT19–CT2 respectively (Figs. 1, 2). The experi-mental PRC resembled the control and the LPPRCs in time-course and waveform, but differed interms of the magnitude of phase shifts at severalphases in the delay region of the PRC. TheANOVA on the values of area under advance and
Fig. 5. Representative actograms of locomotor activityrhythms with stimuli given at CT22. The animals wereadministered stimuli at CT22 after a free-run in constantdarkness (DD) for about 15 days. The phase shifts obtained byexposing the animal to 100–lux of light pulse (LP) for 15–minduration was 1.53 h (a). Administration of 50% DMSO, 5 minbefore 100–lux of LP for 15–min duration evoked 0.40 h ofphase advance (b) and co-administration of a single dose ofmelatonin (1 mg/kg), 5 min before 100 lux of LP for 15 minresulted in 1.12 h of phase advance (c).
type of PRC
DMSO PRCmel PRC
LPPRC
control PRC
exptl PRC
area
und
er a
dvan
ce r
egio
n of
the
PR
C (
A)
0
2
4
6
8
Fig. 6. Plot of area under advance zone (A) of the fivephase response curves (PRCs). The type of PRC is plottedalong the abscissa and the A values along the ordinate. The Avalues of the five PRCs (DMSO PRC, melatonin PRC, LP PRC,control PRC, and experimental PRC) were 0.4070.08 h2
(mean7SEM), 3.4070.24 h2, 5.2670.65 h2, 4.2370.39 h2, and5.9670.35 h2 respectively. The A values of the experimentalPRC were significantly different compared to the A values ofthe control, melatonin, and DMSO PRCs. The A values of theexperimental PRC did not differ significantly from the Avalues of the LP PRC.
type of PRC
DMSO PRCmel PRC
LP PRC
control PRC
exptl PRC
area
und
er d
elay
reg
ion
of th
e P
RC
(D
)
0
2
4
6
8
10
12
14
16
Fig. 7. Plot of area under delay zone (D) of the five phaseresponse curves (PRCs). The D values of the five PRCs (DMSOPRC, melatonin PRC, LP PRC, control PRC, and experi-mental PRC) were 0.1570.04 h2 (mean7SEM), 4.6070.47 h2,7.5770.80 h2, 7.3370.64 h2, and 12.9870.53 h2 respectively.Other details same as in Fig.6. The D values of the experi-mental PRC were significantly different than the D values ofthe control, LP, melatonin, and DMSO PRCs.
LIGHT AND MELATONIN PRCS 165

delay regions of the PRCs revealed that the A andD values of the experimental PRC were signifi-cantly greater than the A and D of the control PRC(Figs. 2, 3). In contrast, the A and D values of theLP and the control PRCs were not significantlydifferent, which suggests that DMSO did notmodify the LP PRC (Figs. 1, 2). A single dose ofmelatonin (1 mg/kg), when administered at phasesbetween CT2 and CT19, to animals free-runningin DD, resulted in phase delays and between CT19and CT2, in phase advances (Sharma et al., ’99).The phase shifts to LP (Fig. 1) and melatonin(Sharma et al., ’99) had similar signs betweenCT10 and CT18, and CT19 and CT2. The LP PRChad non-zero phase shifts during subjective day(i.e. between CT4–CT12). The melatonin PRC(Sharma et al., ’99), however, had non-zero phaseshifts during the subjective day except at CT4,when a small phase delay was recorded. Theexperimental PRC however had phase shiftsindistinguishably closer to zero during the sub-jective day (between CT0–CT10), which suggeststhat the light pulse following the melatonininjections at CT4 suppressed the phase delayingeffects of melatonin. However, except at CT4, itappears that phase shifts evoked by both melato-nin and LPs possibly add up to produce largeradvances and delays (Fig. 3). ANOVA on the phaseshift values revealed a significant effect on phaseand phase�PRC type interaction. The delayphase shifts in the experimental PRC weresignificantly greater compared to the delay phaseshifts of the control and LP PRCs. These resultsclearly suggest that melatonin enhances both theadvancing and delaying effects of LPs. The resultsof our experiments, though, contradict some of therecent findings on interaction between non-photiczeitgebers such as melatonin, serotonin, andactivity with photic zeitgeber (light). These studiesreported that interactions between non-photic andphotic stimuli induce effects smaller in magnitudethan the effects produced by light alone (Glasset al., ’95; Dubocovich et al., ’96; Pickard et al., ’96;Penev et al., ’97; Benloucif et al., ’99). Further, inone of these studies significant effect of melatoninon the LP phase shifts were observed only whenLPs of higher intensities were used (Benloucifet al., ’99). This however suggests that sensitivityto melatonin needs higher intensity of light tosuppress the endogenous melatonin. Moreover,it appears that sensitivity of melatonin in theabsence of endogenous melatonin possibly hasphase advancing effect on the circadian pacemakerof C3H/HeN mice during the early subjective
night, which together with phase delay responses(Benloucif and Dubocovich, ’96) causes reducedphase delays in the LP PRC. In humans, the phaseshifts obtained using both melatonin and LPtogether was smaller than the phase shifts evokedby LP alone (Arendt et al., ’97). Such an interac-tion was also observed in our study when DMSOwas administered along with LPs. The combinedstimuli (50% DMSOþLP) did not cause statisti-cally significant changes in the A and D of the LPPRC, but reduced the A of the LP PRC (Fig. 1, 2).However, there are many important differencesbetween the experimental conditions used in ourstudies and studies reported by others, such asspecies, light intensities, experimental design, andvehicle type, which may have resulted in thedifferences between the results reported here andin some previous studies. The effects of melatoninseen in the present and several similar previousstudies, could very well be ‘‘pharmacological’’actions, since the dose of melatonin used is knownto produce blood levels in orders of magnitude inexcess of those experienced during the normalnocturnal melatonin secretion. In rats daily injec-tions of melatonin entrains the free-runninglocomotor activity rhythm, but rats entrain onlywhen melatonin is administered at a time beforethe normal time of pineal melatonin secretion (forreview see Redman, ’97). Furthermore, pinealec-tomized rats exhibit normal entrainment to LDcycles. These observations could suggest that theobserved effects of melatonin injections to entrainrats are pharmacological and that melatonin is notinvolved in normal entrainment. It seems unlikelyin our experiments, as in a recent study we havedemonstrated that daily injections of melatonin(1 mg/kg) entrained the circadian locomotoractivity rhythm of M. booduga with the phase ofentrainment determined partly by the pre-en-trainment t, suggesting a functional relationshipbetween the circadian clocks and the melatonin asa Zeitgeber (Sharma and Chidambaram, 2003).
If interactions between light and melatonin arefunctionally important in evoking phase shifts,then pinealectomized animals should show mea-surable changes in the PRCs. Attempts have beenmade to investigate the effect of melatonin on theLP induced phase shifts after the replacement ofendogenous melatonin. The results based on offsetof melatonin and temperature as marker rhythmssuggested that either melatonin had no effect orhad a counteracting effect on the LP inducedphase shifts (Hatonen et al., ’96; Cagnacci et al.,’97; Krauchi et al., ’97). However, careful exam-
V.K. SHARMA ET AL.166

ination of the onset of melatonin rhythm demon-strated an increase in the magnitude of both LPinduced phase delays and phase advances, whichwas due to melatonin (Hatonen et al., ’96;Cagnacci et al., ’97). These results are in con-cordance with our findings on the locomotoractivity rhythm in M. booduga. Studies based oninteraction between melatonin and light suggestthat long lasting effects of light, such as anincrease in melatonin receptor expression arelikely to mediate the interaction between melato-nin and light (Gauer et al., ’94).There have been only a few studies on the
effects of interaction of melatonin and light on thephase of the circadian clocks, despite the impor-tance of the role of melatonin and light in phaseresetting of mammalian circadian clocks.Although the results of our experiments basedon the analysis of five PRCs on a tropical rodentcaught from the wild clearly suggest that co-administration of melatonin and light can enhancethe phase shifting capacity of light, more studieswith various levels of melatonin and light admi-nistered in various phases of the circadian cycleare required to unequivocally establish the role ofmelatonin in presence of light.
ACKNOWLEDGEMENTS
This work was financially supported by theJawaharlal Nehru Centre for Advanced ScientificResearch, Bangalore, India. We thank the anon-ymous reviewers for carefully reading the manu-script and suggesting improvements.
LITERATURE CITED
Arendt J, Skene DJ, Middleton B, Lockey SW, Deacon S. 1997.Efficacy of melatonin treatment in jet lag, shift work andblindness. J Biol Rhythms 12:604–617.
Armstrong SM. 1989. Melatonin: The internal zeitgeber ofmammals? Pineal Res Rev 7:157–202.
Benloucif S, Dubocovich ML. 1996. Melatonin and light inducephase shifts of circadian activity rhythms in the C3H/HeNmouse. J Biol Rhythms 11:113–125.
Benloucif S, Masana MI, Yun K, Dubocovich ML. 1999.Interaction between light and melatonin on the circadianclock of mice. J Biol Rhythm 14:281–289.
Cagnacci A, Soldani R, Yen SS. 1997. Contempotaneousmelatonin administration modifies the circadian responseto nocturnal bright light stimuli. Am J Physiol 272:R482–R486.
Cassone VM, Roberts MH, Moore RY. 1988. Effects ofmelatonin on 2–deoxy-[1–14C] glucose uptake within ratsuprachiasmatic nucleus. Am J Physiol 255:R332–R337.
Dubocovich ML, Benloucif S, Masana MI. 1996. Melatoninreceptors in the mammalian suprachiasmatic nucleus.Behav Brain Res 73:141–147.
Gauer F, Masson-Pevet M, Pevet P. 1994. The NMDAantagonist MK–801 blocks light effect on melatonin receptordensity in rat suprachiasmatic nuclei. Neurosci Lett 171:24–26.
Glass JD, Selim M, Srkalovic G, Rea MA. 1995. Tryptophanloading modulates light-induced responses in the mamma-lian circadian system. J Biol Rhythms 10:80–90.
Golombek DA, Cardinali DP. 1993. Melatonin accelerates re-entrainment after phase advance of the light/dark cycle inSyrian hamsters: Antagonism by flumazenil. ChronobiolInternat 10:435–441.
Hastings MH, Mead SM, Vindlacheruvu RR, Ebling FJP,Maywood ES, Grosse J. 1992. Non-photic phase shifting ofthe circadian activity rhythm of Syrian hamsters: Therelative potency of arousal and melatonin. Brain Res591:20–26.
Hatonen T, Alila A, Laakso ML. 1996. Exogenous melatoninfails to counteract the light-induced phase delay of humanmelatonin rhythm. Brain Res 710:125–130.
Johnson CH. 1995. Phase response curves: what they can tellus about circadian clocks? In: Hiroshige T, Honma K,editors. Fourth Sapporo Symposium on biological rhythms.Sapporo: Hokkaido University Press. p 2–48.
Klein DC, Moore RY, Reppert SM. 1991. SuprachiasmaticNucleus: The mind’s clock. New York: Oxford UniversityPress.
Krauchi K, Cajochen C, Danilenko KV, Wirz-Justice A.1997. The hypothermic effect of late evening melatonindoes not block the phase delay induced by concurrentbright light in human subjects. Neurosci Lett 232:57–61.
Mason R, Brooks A. 1988. The electrophysiological effects ofmelatonin and a putative melatonin antagonist (N-acetyl-tryptamine) on rat suprachiasmatic neurons in vitro.Neurosci Lett 95:296–301.
McArthur AJ, Gillette MU, Prosser RA. 1991. Melatonindirectly rests the rat suprachiasmatic circadian clock invitro. Brain Res 565:158–161.
Moore RY. 1981. The suprachiasmatic nucleus, circadianrhythms, and regulation of brain peptides. In: Martin JB,Reichlin S, Bick KL, editors. Neurosecretion and BrainPeptides. New York: Raven p 449–458.
Penev PD, Zee PC, Turek FW. 1997. Serotonin in thespotlight. Nature 385:123.
Pickard GE, Weber ET, Scott PA, Riderdy AF, Rea MA. 1996.5HT1B receptor agonists inhibit light-induced phase-shiftsof behavioral circadian rhythms and expression of immedi-ate-early genes c-fos in the suprachiasmatic nucleus.J Neurosci 16:8208–8220.
Pittendrigh CS, Daan S. 1976. A functional analysis ofcircadian pacemakers in nocturnal rodents: IV. Entrain-ment: Pacemaker as clock. J Comp Physiol A 106:291–331.
Redman JR. 1997. Circadian entrainment and phaseshifting in mammals with melatonin. J Biol Rhythms12:581–587.
Reiter RJ. 1987. Pineal melatonin message: Duration versuscoincidence hypotheses. Life Sci 40:2119–2131.
Sharma VK, Chidambaram R. 2003. Entrainment of circadianlocomotor activity rhythm of the nocturnal field mouse Musbooduga using daily injections of melatonin. J Exp Zool296A:30–35.
Sharma VK, Chandrashekaran MK, Nongkynrih P. 1997.Daylight and artificial light phase response curves for the
LIGHT AND MELATONIN PRCS 167

circadian rhythm in locomotion activity of the field mouseMus booduga. Biol Rhythm Res 28:39–49.
Sharma VK, Singaravel M, Subbaraj R, ChandrashekaranMK. 1999. Timely administration of melatonin acceleratesre-entrainment to phase shifted light dark cyclesin the field mouse Mus booduga. Chronobiol Internat16:163–170.
Shibata S, Cassone VM, Moore RY. 1989. Effects of melatoninon neuronal activity in the rat suprachiasmatic nucleus invitro. Neurosci Lett 97:140–144.
Yellon SM, Hilliker S. 1994. Influence of acute melatonintreatment and light on the circadian melatonin rhythm inthe Djungarian hamster. J Biol Rhythms 9:71–81.
V.K. SHARMA ET AL.168