miwi associates with translational machinery and … associates with translational machinery and...
TRANSCRIPT

MIWI associates with translational machineryand PIWI-interacting RNAs (piRNAs) inregulating spermatogenesisShane T. Grivna*†, Brook Pyhtila*, and Haifan Lin*‡
Departments of *Cell Biology and †Pharmacology and Molecular Cancer Biology, Duke University Medical School, Durham, NC 27710
Communicated by Brigid L. M. Hogan, Duke University Medical Center, Durham, NC, July 14, 2006 (received for review March 9, 2006)
Noncoding small RNAs have emerged as important regulators ofgene expression at both transcriptional and posttranscriptionallevels. Particularly, microRNA (miRNA)-mediated translational re-pression involving PIWI�Argonaute family proteins has beenwidely recognized as a novel mechanism of gene regulation. Wepreviously reported that MIWI, a murine PIWI family member, isrequired for initiating spermiogenesis, a process that transformsround spermatids into mature sperm. MIWI is a cytoplasmic proteinpresent in spermatocytes and round spermatids, and it is requiredfor the expression of its target mRNAs involved in spermiogenesis.Most recently, we discovered a class of noncoding small RNAscalled PIWI-interacting RNAs (piRNAs) that are abundantly ex-pressed during spermiogenesis in a MIWI-dependent fashion. Here,we show that MIWI associates with both piRNAs and mRNAs incytosolic ribonucleoprotein and polysomal fractions. As polysomesincrease in early spermiogenesis, MIWI increases in polysomefractions. Moreover, MIWI associates with the mRNA cap-bindingcomplex. Interestingly, MIWI is required for the expression of notonly piRNAs but also a subset of miRNAs, despite the presence ofDicer. These results suggest that MIWI has a complicated role in thebiogenesis and�or maintenance of two distinct types of smallRNAs. Together, our results indicate that MIWI, a PIWI subfamilyprotein, uses piRNA as the major, but not exclusive, bindingpartner, and it is associated with translational machinery.
Argonaute � germ line � ribosomes � Dicer � cap binding
Translational regulation is a major mechanism that controls geneexpression in diverse biological processes (1–4). The impor-
tance of such regulation is highlighted in the last phase of sper-matogenesis, known as spermiogenesis, where translation is uncou-pled from transcription as the haploid genome is condensed andrepackaged (5–9). Genes involved in spermiogenesis are tran-scribed during earlier stages of spermatogenesis, and they aretranslationally repressed for several days until their protein prod-ucts are needed. This translational repression has been well studied;its mechanisms include alteration of poly(A) tail length (10) andbinding of particular sequences in the untranslated regions byregulatory proteins (11, 12).
Recently, microRNA (miRNA)-mediated translational repres-sion has emerged as a distinct mechanism of posttranscriptionalregulation (3, 13, 14). Although the precise nature of such regula-tion remains unclear, the miRNA machinery is targeted to anmRNA by target sequences nearly complementary to an �21-ntmiRNA (15, 16). The miRNA machinery consists of the Dicerribonuclease and the effector complex known as RISC, containinga member of the PIWI�Argonaute protein family (17, 18). Evidenceis emerging that miRNAs may constitute a major mechanism oftranslational regulation during spermatogenesis in the mouse.Members of the PIWI protein family are essential for completionof spermatogenesis (19, 20), and miRNAs are expressed in malegerm cells (21). In addition, it has been shown in vitro that transitionprotein 2 (TP2) mRNA, a posttranscriptionally regulated spermio-genic mRNA, is targeted by miR-122a, an miRNA expressed in lategerm cells (21).
In addition to miRNAs, we and others have recently discovereda class of noncoding small RNAs termed PIWI-interacting RNAs(piRNAs) that are expressed during spermatogenesis, mostly inspermatids, at much higher levels than miRNAs (22). piRNAs are�30 nt in length. They tend to cluster, and they are diverselydistributed among exonic, intronic, intergenic, and repeat se-quences in the mouse genome, suggesting their potentially diverseroles in regulating gene expression. We also observe that a fractionof piRNAs are associated with polysomes. Interestingly, piRNAsassociate with MIWI, a murine PIWI�Argonaute protein expressedexclusively in spermatogenic cells and required for initiating thespermiogeneic program (19). Moreover, piRNAs depend on MIWIfor their expression (22).
To explore the functional relationship among MIWI, piRNAs,and miRNAs in spermatogenesis, we have biochemically charac-terized MIWI. We previously showed that MIWI is a cytoplasmicprotein that binds to and maintains the level of its mRNA targets(19), some of which are known to be translationally repressed andinvolved in spermiogenesis (23–25). Spermatogenesis in miwi-nulltestes is arrested uniformly at the onset of spermiogenesis, indicat-ing the crucial role of MIWI in initiating this process. Most recently,MIWI was shown to localize in the chromatoid body, a perinuclearstructure related to the P body of somatic cells, and it was proposedto serve as an RNA storage and processing center (26, 27). Here,we report that MIWI is associated with translational machinery andpiRNAs. MIWI is also required for the expression of a subset ofmiRNAs. These results imply a potentially complicated role forMIWI in small RNA-mediated processes during spermatogenesis.
ResultsMIWI Associates with Polysomes in an RNA-Dependent Manner. Toexplore the biochemical function of MIWI, we first examined thesubcytoplasmic localization of MIWI in spermatogenic cells byimmunofluorescence microscopy. In spermatocytes, MIWI ispresent throughout the cytoplasm (Fig. 1A). In round spermatids,MIWI concentrates in the chromatoid body as reported by Kotajaet al. (27), but it is also present throughout the cytoplasm (Fig. 1A).The presence of MIWI outside the chromatoid body implies anadditional function for the protein beyond its proposed role in RNAstorage and�or processing.
Because the chromatoid body is a nonmembranous structure, todetermine whether cytoplasmic MIWI is associated with mem-branes such as endoplasmic reticulum, we subjected testicularextract to a membrane-flotation assay to separate membranous andcytosolic fractions (illustrated in Fig. 1B). MIWI does not cofrac-tionate with membranes, as marked by TRAP�, an endoplasmicreticulum integral membrane protein (fractions 2 and 7, Fig. 1C);
Conflict of interest statement: No conflicts declared.
Abbreviations: dpp, days postpartum; GSK3�, glycogen synthase kinase 3�; miRNA,microRNA; mRNP, messenger RNP; MVH, mouse vasa homolog; piRNA, PIWI-interactingRNA; RNP, ribonucleoprotein.
‡To whom correspondence should be addressed. E-mail: [email protected].
© 2006 by The National Academy of Sciences of the USA
www.pnas.org�cgi�doi�10.1073�pnas.0605506103 PNAS � September 5, 2006 � vol. 103 � no. 36 � 13415–13420
DEV
ELO
PMEN
TAL
BIO
LOG
Y

but rather, it fractionates to the membrane-free cytosol (fractions3–6, Fig. 1C), suggesting that cytosolic MIWI does not associatewith membranes.
Because some MIWI target mRNAs are known to be transla-tionally regulated, we wanted to determine whether MIWI plays arole in translation. Two Argonaute subfamily proteins, humaneIF2C2 and trypanosome TbAgo, are known to associate withpolysomes (28, 29). To determine whether MIWI, a PIWI subfamilyprotein, also associates with polysomes, we subjected adult testic-ular extracts to sucrose density gradient fractionation. MIWI co-sediments with both free ribonucleoproteins (RNPs) and poly-somes (Fig. 2A). The addition of EDTA to dissociate ribosomalsubunits results in a shift of MIWI to subunit fractions of thegradient, suggesting that a portion of MIWI associates with poly-somes. To ensure that MIWI is not in an EDTA-sensitive complexthat cosediments with polysomes, we digested testicular extracts
with micrococcal nuclease before sedimentation to cleave mRNAsbetween ribosomes. This treatment results in a complete shift ofMIWI to the RNP fractions, away from ribosomal subunits. Thisresult not only confirms the association of MIWI with polysomes,it also reveals that the association of MIWI with ribosomes ismRNA-dependent.
MIWI Associates with Translationally Inactive mRNAs. To determinewhether MIWI associates with mRNAs in the RNP fractions,we performed an RNP capture assay. RNP and polysomalfractions from sucrose gradients were incubated with oli-go(dT)-cellulose to capture poly(A)� RNA and associatedproteins (see Materials and Methods). Both RNP- and poly-some-associated MIWI bind to oligo(dT) (Fig. 2B), as does thegerm-cell RNA-binding protein MSY2 (30). In contrast, thenon-RNA-binding cytosolic protein GSK3� does not bind tooligo(dT). Furthermore, preincubation of the oligo(dT) matrixwith poly(A) significantly reduces both MIWI and MSY2binding in both RNP and polysome fractions, indicating thatMIWI binds oligo(dT) through associated poly(A)� RNAs.Thus, MIWI is associated with mRNAs in both polysomal andRNP fractions.
Global Translation Profile Is Not Altered in miwi�/� Testes. BecauseMIWI associates with polysomes, we examined whether MIWIregulates global translation in spermatogenic cells or plays aunique role in translational regulation. MIWI is first detectablein mid-pachytene spermatocytes, and it becomes much more
Fig. 1. MIWI is present in both the chromatoid body and the cytosol. (A)Cross-section of a 24-days postpartum (dpp) seminiferous tubule costained forMIWI (red) and DNA (green). In a seminiferous tubule, spermatogonia (Sg)contain DAPI-intense nuclei, and they reside in the basal layer where MIWIexpression is absent. Interspersed among the spermatogonia are a smallnumber of somatic supporting cells called Sertoli cells (St), which also reside inthe basal layer. The nuclei of Sertoli cells show diffuse DAPI staining butcontain two distinct nucleoli (DAPI bright spots). Spermatogonia divide toproduce primary spermatocytes (Sc) that contain the largest nuclei and residemostly in the two to three subbasal layers. These primary spermatocytes are atthe prophase of meiosis I. Completion of meiosis then generates roundspermatids (Sd) that contain small nuclei and reside in the luminal layers. Theround spermatids each contain a chromatoid body that accumulates a highconcentration of MIWI (arrowheads). Round spermatids then undergo sper-miogenesis to produce mature sperm (not shown). Surrounding the tubule arethin myoid cells (M), which are separated from the tubule proper by basallamina (not shown). (Scale bar, 20 �m.) (B) Membrane-flotation assay. Testic-ular extract was centrifuged on a sucrose step gradient for isopycnic separa-tion of membranes from cytosol. rER, rough endoplasmic reticulum. (C) West-ern blots of individual sedimentation fractions probed for MIWI and TRAP�.RNA was isolated from 20% of each fraction.
Fig. 2. MIWI associates with mRNA in both RNPs and polysomes. (A) Untreated(�Mg) and EDTA-treated (�EDTA) or micrococcal nuclease-treated (�MNase)postnuclear testicular extract was fractionated on 15–50% sucrose density gra-dients and analyzed by UV spectrometry and Western blotting. RNA was isolatedfrom 20% of every other fraction. Five percent of each fraction was immuno-blotted for MIWI. (B) RNP and polysome fractions from sucrose gradients weresubjected to RNP-capture assay with oligo(dT)-cellulose. Poly(A) sequences wereadded as a competitor. Twenty-five percent of the input and bound proteinswere immunoblotted for MIWI, MSY2, and glycogen synthase kinase 3� (GSK3�).
13416 � www.pnas.org�cgi�doi�10.1073�pnas.0605506103 Grivna et al.

abundant in diplotene spermatocytes (19). This enhanced ex-pression persists in meiotic spermatocytes and steps 1–3 roundspermatids. We therefore compared the polysome profiles ofage-matched miwi�/� and miwi�/� testicular extracts isolated atthree key time points of MIWI expression during the first waveof spermatogenesis: (i) at 16 dpp, 2 days after MIWI expressionbegins, when only spermatogonia and primary spermatocyteshave formed (31); (ii) at 20 dpp, when MIWI is abundantlyexpressed in diplotene to secondary spermatocytes, beforeround spermatid formation; (iii) at 24 dpp, when abundantMIWI expression extends to newly formed steps 1–3 roundspermatids. Twenty-four days postpartum is also the age at whichspermatogenesis is arrested in miwi mutants (19). Importantly,at all three ages, the equivalent complement of spermatogeniccells is present in both genotypes. The polysome profiles ofmiwi�/� and miwi�/� testes at all three times are identical (Fig.3 A–C), suggesting that global translation is not affected in miwimutants.
The absorbance profiles (Fig. 3 A–C) also reveal a significantincrease in both the abundance of polysomes and MIWI associationwith polysomes at 24 dpp, when round spermatids have formed. Toensure that the increased association of MIWI with polysomes at 24dpp is not simply the result of increased MIWI expression, weexamined MIWI levels in 16-, 20-, and 24-dpp testes, relative toCDK5 (Fig. 3D). MIWI levels increase �2-fold from 16 to 20 dpp;however, they remain steady through 24 dpp. Thus, more MIWIbecomes associated with mRNAs on polysomes as translationalactivity increases.
To determine whether MIWI associates with elongating poly-somes, we cultured seminiferous tubules under conditions allowingactive translation in germ cells (32). Cultures were treated with anelongation inhibitor, cycloheximide, and they were subjected tosucrose density fractionation. As expected, cycloheximide-treatedtubules show a decrease in the monosomal peak and a correspond-ing increase in the polysomal peak compared with untreated tubules(Fig. 3E). Interestingly, MIWI distribution correspondingly shifts toheavier polysomal fractions (Fig. 3E). The accumulation of poly-somes after cycloheximide treatment is likely caused by a reducedelongation rate, reduced premature termination, or both (32–34).In any case, our results suggest that MIWI associates with activelyelongating polysomes.
The RNP Fraction Contains Chromatoid Body Components. Because alarge portion of MIWI sediments to the RNP fractions of sucrosegradients, we hypothesized that the RNP fractions contain thedissociated elements of the chromatoid body. To test this possibility,we examined the distribution of MVH and GW182 in 24-dppsucrose gradients from Fig. 3C. MHV is present in the cytoplasmof spermatocytes, and it is concentrated in the chromatoid body ofround spermatids (35, 36), whereas GW182, a P body-specificcomponent, also localizes in the chromatoid body (27, 37). In ourexperiments, MVH and GW182 sediment predominantly in theRNP fractions (Fig. 3F), suggesting that chromatoid body compo-nents sediment in the RNP fractions. Hence, it is likely thatchromatoid body-associated MIWI also sediments in the RNPfractions. However, we cannot conclude whether a portion ofcytosolic MIWI also sediments in the RNP fractions.
MIWI Complexes with piRNAs. Although the roles of mammalianArgonaute subfamily proteins in RNAi and miRNA pathways arewell documented, the roles of PIWI subfamily members have notyet been determined (13, 14). We have recently shown that MIWIcomplexes with piRNAs and that a fraction of piRNAs cosedimentwith polysomes (22). To investigate whether MIWI associates withpiRNAs on polysomes, we immunoprecipitated MIWI from threepools of testicular extract created by combining sucrose gradientfractions corresponding to the messenger ribopucleoprotein(mRNP), monosomal, and polysomal fractions. Control precipita-
tions were performed with preimmune serum. CoprecipitatedRNA was end-labeled and separated on denaturing polyacrylamidegels. Small RNA species of expected piRNA size (�30 nt) specif-
Fig. 3. MIWI association with polysomes corresponds to increased transla-tion during spermatogenesis. (A–C) Postnuclear testicular extracts frommiwi�/� and miwi�/� mice at 16 dpp (A), 20 dpp (B), and 24 dpp (C) werefractionated on sucrose density gradients and analyzed by UV spectrometry.Numbers on the left in each panel correspond to A260 maximum and minimum.Five percent of every other fraction was immunoblotted for MIWI, which isshown below the corresponding A260 profile. (D) MIWI expression at 16, 20,and 24 dpp, normalized against CDK5 and set with the expression level at 16dpp as 1. (E) Effect of cycloheximide (CHX) on polysome profile and MIWIdistribution in cultured seminiferous tubules. (F) Immunoblots of fractionsfrom C probed for MIWI, mouse vasa homolog (MVH), and GW182.
Grivna et al. PNAS � September 5, 2006 � vol. 103 � no. 36 � 13417
DEV
ELO
PMEN
TAL
BIO
LOG
Y
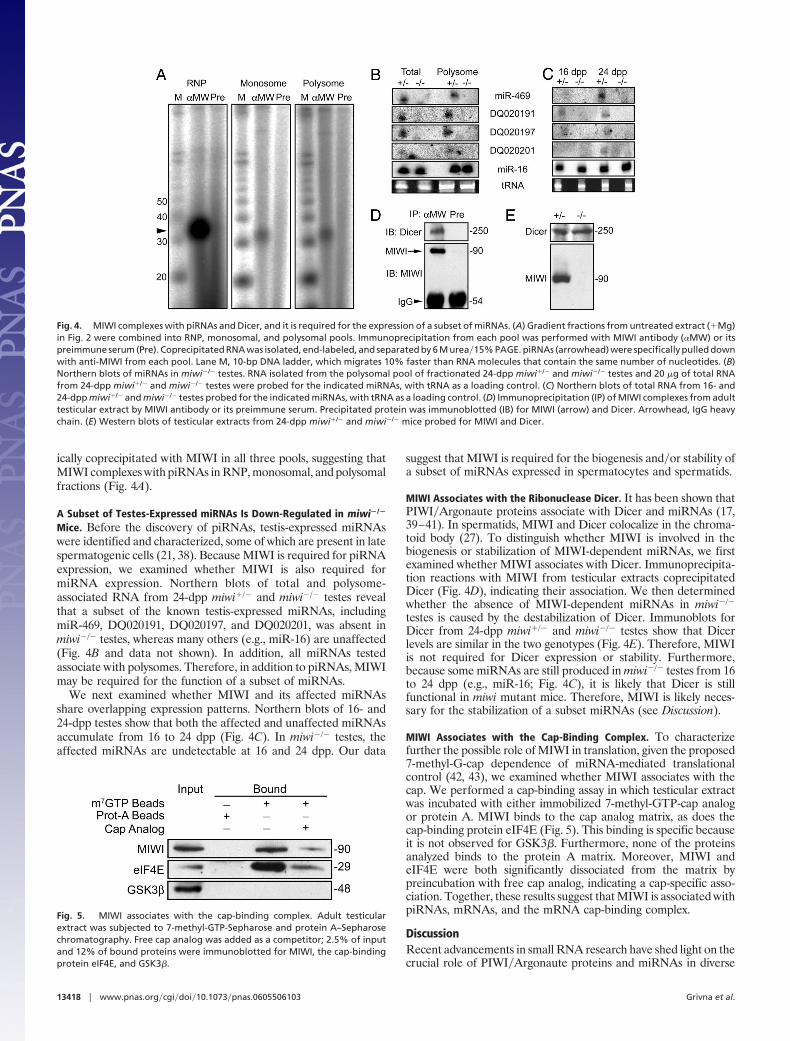
ically coprecipitated with MIWI in all three pools, suggesting thatMIWI complexes with piRNAs in RNP, monosomal, and polysomalfractions (Fig. 4A).
A Subset of Testes-Expressed miRNAs Is Down-Regulated in miwi�/�
Mice. Before the discovery of piRNAs, testis-expressed miRNAswere identified and characterized, some of which are present in latespermatogenic cells (21, 38). Because MIWI is required for piRNAexpression, we examined whether MIWI is also required formiRNA expression. Northern blots of total and polysome-associated RNA from 24-dpp miwi�/� and miwi�/� testes revealthat a subset of the known testis-expressed miRNAs, includingmiR-469, DQ020191, DQ020197, and DQ020201, was absent inmiwi�/� testes, whereas many others (e.g., miR-16) are unaffected(Fig. 4B and data not shown). In addition, all miRNAs testedassociate with polysomes. Therefore, in addition to piRNAs, MIWImay be required for the function of a subset of miRNAs.
We next examined whether MIWI and its affected miRNAsshare overlapping expression patterns. Northern blots of 16- and24-dpp testes show that both the affected and unaffected miRNAsaccumulate from 16 to 24 dpp (Fig. 4C). In miwi�/� testes, theaffected miRNAs are undetectable at 16 and 24 dpp. Our data
suggest that MIWI is required for the biogenesis and�or stability ofa subset of miRNAs expressed in spermatocytes and spermatids.
MIWI Associates with the Ribonuclease Dicer. It has been shown thatPIWI�Argonaute proteins associate with Dicer and miRNAs (17,39–41). In spermatids, MIWI and Dicer colocalize in the chroma-toid body (27). To distinguish whether MIWI is involved in thebiogenesis or stabilization of MIWI-dependent miRNAs, we firstexamined whether MIWI associates with Dicer. Immunoprecipita-tion reactions with MIWI from testicular extracts coprecipitatedDicer (Fig. 4D), indicating their association. We then determinedwhether the absence of MIWI-dependent miRNAs in miwi�/�
testes is caused by the destabilization of Dicer. Immunoblots forDicer from 24-dpp miwi�/� and miwi�/� testes show that Dicerlevels are similar in the two genotypes (Fig. 4E). Therefore, MIWIis not required for Dicer expression or stability. Furthermore,because some miRNAs are still produced in miwi�/� testes from 16to 24 dpp (e.g., miR-16; Fig. 4C), it is likely that Dicer is stillfunctional in miwi mutant mice. Therefore, MIWI is likely neces-sary for the stabilization of a subset miRNAs (see Discussion).
MIWI Associates with the Cap-Binding Complex. To characterizefurther the possible role of MIWI in translation, given the proposed7-methyl-G-cap dependence of miRNA-mediated translationalcontrol (42, 43), we examined whether MIWI associates with thecap. We performed a cap-binding assay in which testicular extractwas incubated with either immobilized 7-methyl-GTP-cap analogor protein A. MIWI binds to the cap analog matrix, as does thecap-binding protein eIF4E (Fig. 5). This binding is specific becauseit is not observed for GSK3�. Furthermore, none of the proteinsanalyzed binds to the protein A matrix. Moreover, MIWI andeIF4E were both significantly dissociated from the matrix bypreincubation with free cap analog, indicating a cap-specific asso-ciation. Together, these results suggest that MIWI is associated withpiRNAs, mRNAs, and the mRNA cap-binding complex.
DiscussionRecent advancements in small RNA research have shed light on thecrucial role of PIWI�Argonaute proteins and miRNAs in diverse
Fig. 5. MIWI associates with the cap-binding complex. Adult testicularextract was subjected to 7-methyl-GTP-Sepharose and protein A–Sepharosechromatography. Free cap analog was added as a competitor; 2.5% of inputand 12% of bound proteins were immunoblotted for MIWI, the cap-bindingprotein eIF4E, and GSK3�.
Fig. 4. MIWI complexes with piRNAs and Dicer, and it is required for the expression of a subset of miRNAs. (A) Gradient fractions from untreated extract (�Mg)in Fig. 2 were combined into RNP, monosomal, and polysomal pools. Immunoprecipitation from each pool was performed with MIWI antibody (�MW) or itspreimmune serum (Pre). Coprecipitated RNA was isolated, end-labeled, and separated by 6 M urea�15% PAGE. piRNAs (arrowhead) were specifically pulled downwith anti-MIWI from each pool. Lane M, 10-bp DNA ladder, which migrates 10% faster than RNA molecules that contain the same number of nucleotides. (B)Northern blots of miRNAs in miwi�/� testes. RNA isolated from the polysomal pool of fractionated 24-dpp miwi�/� and miwi�/� testes and 20 �g of total RNAfrom 24-dpp miwi�/� and miwi�/� testes were probed for the indicated miRNAs, with tRNA as a loading control. (C) Northern blots of total RNA from 16- and24-dpp miwi�/� and miwi�/� testes probed for the indicated miRNAs, with tRNA as a loading control. (D) Immunoprecipitation (IP) of MIWI complexes from adulttesticular extract by MIWI antibody or its preimmune serum. Precipitated protein was immunoblotted (IB) for MIWI (arrow) and Dicer. Arrowhead, IgG heavychain. (E) Western blots of testicular extracts from 24-dpp miwi�/� and miwi�/� mice probed for MIWI and Dicer.
13418 � www.pnas.org�cgi�doi�10.1073�pnas.0605506103 Grivna et al.

biological processes. Although Argonaute subfamily proteins havebeen implicated in siRNA-mediated mRNA degradation and epi-genetic silencing as well as miRNA-mediated translational repres-sion, the precise underlying mechanisms remain unclear (13, 14).Even less is known about the molecular processes mediated by thePIWI subfamily proteins. Recently, we have shown that MIWI bindsto a class of small RNAs called piRNAs (22). In addition, MIWI andDicer were shown to colocalize in the chromatoid body in sperma-tids, and they were speculated to be involved in mRNA storageand�or processing mediated by miRNAs (27). We show here thatMIWI also exists outside the chromatoid body and is associatedwith the translational machinery. Moreover, MIWI associates withpiRNAs in RNPs and on polysomes. In addition, MIWI is alsorequired for the expression of a subset of miRNAs. Therefore, it istempting to speculate that mammalian PIWI proteins might beinvolved in translational regulation, possibly through more than onetype of small RNA.
Is MIWI Involved in Translational Regulation? Current analysis indi-cates that various members of the Argonaute subfamily havedistinct biochemical activities (44). Likewise, the siRNA andmiRNA pathways are different in that the former leads to eithermRNA degradation or epigenetic silencing, whereas the latter isthought to lead to translational repression (13, 14). Despite thesedifferences, a common feature among PIWI�Argonaute proteins,as well as siRNA and miRNA pathways, is that they all negativelyregulate gene expression. Our results demonstrate that MIWIbecomes associated with polysomes at 20 dpp, when translation isreinitiated in late spermatocytes; and the association is significantlyincreased at 24 dpp, when round spermatids undergo a major phaseof translation (6, 31). MIWI is also associated with both thecap-binding complex and with piRNAs on elongating polysomes,although these associations alone are not sufficient to suggest a roleof MIWI in translational regulation. Further functional assay isrequired to conclude definitively whether MIWI regulates transla-tion or whether piRNAs play an miRNA-like role in translationalregulation.
MIWI in Chromatoid Bodies vs. Polysomes. The presence of MIWI inboth the chromatoid body and the free cytosol is consistent with atwo-phase distribution of MIWI between RNP and ribosomalfractions in sucrose gradients (Fig. 2). The RNP fractions alsocontain MVH and GW182, two other chromatoid body compo-nents, suggesting that chromatoid body-associated MIWI likelysediments in the RNP fractions. This conclusion is supported by theobservation that ribosomes are primarily excluded from the chro-matoid body (27), a phenomenon that was also observed in P bodies(45). The association of mRNA with MIWI in the RNP fractionsfurther supports the observations that the chromatoid body con-tains mRNAs (26) and appears to be related to the somatic P body,a structure involved in mRNA storage�degradation (27).
Does MIWI Partner with piRNA for Its Function? An exciting findingin our study is that MIWI is associated with piRNAs and mRNAsin both RNP and ribosomal fractions. This discovery raises thepossibility that MIWI partners with piRNAs for its function. Suchpartnership might be essential for initiating spermiogenesis becausespermatogenesis in the miwi mutant is arrested at the beginning ofspermiogenesis (19).
Although it is accepted that PIWI�Argonaute proteins playconserved roles in small RNA biology, it remains unclear what, ifanything, distinguishes the two subfamilies functionally. Our recentstudy (22) and results reported here demonstrate that MIWI, amammalian PIWI subfamily member, associates mainly, althoughnot exclusively, with piRNAs, and it is responsible for the expressionof piRNAs and a small subset of miRNAs. The association of MIWIwith piRNAs on polysomes provides correlative evidence that PIWI
subfamily members and piRNAs might be partners in the transla-tional machinery.
Interestingly, a subset of miRNAs known to be expressed inlate-stage spermatogenic cells is also down-regulated in miwi mice,which suggests that MIWI is also required, either directly orindirectly, for the expression of these miRNAs. We favor thepossibility that MIWI is required to maintain the stability of thesemiRNAs because Dicer is present at normal levels and is functionalin the miwi mutant. It remains possible, however, that MIWIinteraction with Dicer or a Dicer-like enzyme is required for theproduction of piRNAs and particular miRNAs. In any case, becauseMIWI associates with piRNAs and mRNAs both in RNPs and onribosomes, it is possible that piRNAs are involved in the regulationof both translation and mRNA stability control by MIWI.
Materials and MethodsMembrane-Flotation Assay. This assay was performed as describedby Lerner et al. (46), with testes from adult CD-1 mice. Fractions(0.5 ml) were collected manually, and 5% of each fraction wasimmunoblotted for MIWI and TRAP�. RNA was extracted from20% of each fraction with TRIzol (Invitrogen, Carlsbad, CA).
Immunofluorescence Microscopy of MIWI. Testes from 24-dppmiwi�/� mice were prepared and sectioned as described by Dengand Lin (19). Fixed sections were blocked in PBS supplementedwith 10% cold fish skin gelatin and 10% normal goat serum for 1 hat room temperature. Sections were stained with MIWI antibody(19) and Alexa Fluor 568-labeled secondary antibody (MolecularProbes, Eugene, OR) and counterstained for DNA with DAPI.
Sucrose Density Gradient Fractionation. Decapsulated testes fromadult CD-1 mice were Dounce-homogenized in 150 mM KOAc�50mM Hepes, pH 7.5�5 mM Mg(OAc)2�2 mM DTT�0.5% TritonX-100�1� protease inhibitors (complete EDTA-free; Roche,Indianapolis, IN)�80 units/ml RNaseOUT (Invitrogen)�0.25 Msucrose. Postnuclear supernatants were loaded onto continuous15–50% sucrose gradients prepared in lysis buffer without deter-gent. Gradients were centrifuged for 3 h at 150,000 � g in an SW41rotor (Beckman, Mountain View, CA). Fractions (0.5 ml) werecollected manually, and the A260 was measured. RNA was isolatedfrom 20% of gradient fractions with TRIzol. For EDTA-treatedsamples, Mg(OAc)2 in all buffers was replaced with 20 mM EDTA.For nuclease treatment, lysates were supplemented with 1 mMCaCl2 and 200 �M cycloheximide and incubated with 30 units�mlStaphylococcus micrococcal nuclease for 15 min at room temper-ature. Digestion was stopped with 3 mM EGTA. Five percent ofeach fraction was immunoblotted with MIWI antibody.
For developmental polysome analysis, testes from 16-, 20-, and24-dpp miwi�/� and miwi�/� mice were prepared and fractionatedas above. Four testes were used for 16- and 20-dpp analysis, and twowere used for 24-dpp analysis. Five percent of every other fractionwas immunoblotted for MIWI, MVH, or GW182.
For total-protein analysis, 50 �g of total testicular extract from16-, 20-, and 24-dpp miwi�/� mice was probed for MIWI and CDK5and quantified with ImageQuant TL software (GE Healthcare,Amersham, U.K.).
Seminiferous-Tubule Culture. Seminiferous-tubule culture of singleadult testes was performed as described by Kleene (32), with thefollowing modifications. Cultures were supplemented with 0 or 200�M cycloheximide for 2 h at 34°C in 5% CO2; they were washed incold RPMI medium and subjected to sucrose density gradientfractionation as above. Four percent of every other fraction wasimmunoblotted for MIWI.
mRNP-Capture Assay. Fractions from sucrose gradient fractionscorresponding to the RNP and polysomal pools were subjected toRNP capture with oligo(dT)-cellulose. Briefly, 10 mg of oligo(dT)-
Grivna et al. PNAS � September 5, 2006 � vol. 103 � no. 36 � 13419
DEV
ELO
PMEN
TAL
BIO
LOG
Y

cellulose (Sigma, St. Louis, MO) was blocked in 1 ml of bindingbuffer [50 mM Hepes, pH 7.5�150 mM KOAc�5 mM Mg(OAc)2�2mM DTT] containing 5% BSA for 1 h at 4°C; 1.5% of four RNPfractions or 2% of 10 polysome fractions was mixed with 0.5 ml ofbinding buffer and 25 �l of blocked matrix. For competition assays,200 �g of (A)25 (Integrated DNA Technologies, Coralville, IA) wasadded 10 min before the addition of extracts. After being rocked for30 min at 4°C, beads were spun down and washed twice with bindingbuffer; bound proteins were eluted by boiling in 40 �l of 2�Laemmli buffer. Twenty-five percent of input and bound proteinwas immunoblotted for MIWI, MSY2, and GSK3�.
Small RNA Immunoprecipitation. Fractions from sucrose gradientswere combined to create mRNP, monosomal, and polysomal pools.One-half of each pool was combined with an equal volume of NT2buffer (50 mM Tris, pH 7.4�150 mM NaCl�1 mM MgCl2�0.05%Nonidet P-40) and incubated with protein A–Sepharose beadscoated with either anti-MIWI or preimmune serum. To recovercoprecipitated RNA, 200 �l of TE (10 mM Tris/1 mM EDTA, pH7.5)�1% SDS was added to the precipitate and heated to 95°C for3 min, followed by phenol�chloroform extraction and isopropylalcohol precipitation. Precipitated RNA was dephosphorylatedwith calf intestinal phosphatase (New England Biolabs, Ipswich,MA) and 5�-end-labeled with [�-32P]ATP and T4 polynucleotidekinase (New England Biolabs). An end-labeled 10-bp DNA ladder(Invitrogen) was used as a marker. Labeled RNA and marker wereseparated on denaturing 15% polyacrylamide gels and exposed toa PhosphorImager (Molecular Dynamics, Sunnyvale, CA).
miRNA Northern Blotting. RNA from 24-dpp mouse testes andpooled polysome fractions was extracted with TRIzol. Twentymicrograms of total testicular RNA and one testis equivalent ofpolysomal RNA were separated by 6 M urea�15% PAGE andtransferred to a Hybond-N� membrane (Amersham Biosciences,Piscataway, NJ). Hybridizations were performed at 42°C in hybrid-ization buffer (1� SSC�7% SDS�20 mM Na2HPO4, pH 7.2�1�Denhardt’s solution�0.1 mg/ml salmon sperm DNA). Blots wereprobed with radiolabeled DNA oligonucleotides (Integrated DNA
Technologies) complementary to mature miRNA sequences.Probes were labeled with [�-32P]ATP and T4 polynucleotide kinase.Blots were washed twice in 1� SSC�0.1% SDS at 42°C for 10 minand exposed a PhosphorImager. Two duplicate blots were strippedand reprobed as necessary.
Dicer-MIWI Coimmunoprecipitation. Immunoprecipitations from 2mg of adult testis extract were performed as described by Deng andLin (19). Ten percent of each pellet was immunoblotted for MIWI,and the remainder was probed with Dicer antibody (Imgenex, SanDiego, CA). For total-protein analysis, 200 �g and 50 �g of totaltestis extract from 24-dpp mice was immunoblotted for Dicer andMIWI, respectively.
Cap-Binding Assay. Adult CD-1 testes were homogenized in bindingbuffer [50 mM Hepes, pH 7.4�100 mM KOAc�2 mM Mg(OAc)2�1mM DTT�0.1% Triton X-100�1� protease inhibitors (completeEDTA-free; Roche)]. Fifty microliters of a 1:1 suspension of either7-methyl-GTP-Sepharose 4B (Amersham Biosciences) or proteinA–Sepharose 4B (Sigma) was blocked in binding buffer containing3% BSA for 1 h at 4°C. Two milligrams of postnuclear extract wasadded to blocked beads in 500 �l of binding buffer for 2 h at 4°Cwith rocking. For competition assays, 200 �g�ml m7G(5�)ppp(5�)Gcap analog (Ambion, Austin, TX) was added to the extract andincubated for 1 h at 4°C before the addition of beads. Beads werewashed with binding buffer, and bound proteins were eluted byboiling in Laemmli buffer; then 2.5% of input and 12% of elutedproteins were immunoblotted for MIWI, eIF4E (Santa Cruz Bio-technology, Santa Cruz, CA), and GSK3�.
We thank Dr. C. Nicchitta for advice and help; members of the Lin andNicchitta laboratories for stimulating discussions; Dr. N. Hecht (Uni-versity of Pennsylvania, Philadelphia, PA) for MSY2 antibody; Dr. T.Noce (Mitsubishi Institute, Japan) for MVH antibody; Dr. M. Fritzler(University of Calgary, Alberta, Canada) for GW182 antibody; and Drs.B. Hogan, J. Keene, A. Fedoriw, S. Findley, and Y. Unhavaithaya as wellas Ms. H. Megosh and Mr. H. Yin for valuable comments on themanuscript. This work was supported by National Institutes of Health(NIH) Grant HD42012 (to H.L.) and NIH Predoctoral Training GrantT32 CA59365 (to S.T.G).
1. Kozak, M. (1992) Annu. Rev. Cell Biol. 8, 197–225.2. Keene, J. D. (2001) Proc. Natl. Acad. Sci. USA 98, 7018–7024.3. Storz, G., Altuvia, S. & Wassarman, K. M. (2005) Annu. Rev. Biochem. 74, 199–217.4. Wilusz, C. J. & Wilusz, J. (2004) Trends Genet. 20, 491–497.5. Kleene, K. C. (2003) Cytogenet. Genome Res. 103, 217–224.6. Hecht, N. B. (1986) in Experimental Approaches to Mammalian Embryonic Development, eds.
Rossant, J. & Pedersen, R. A. (Cambridge Univ. Press, New York), pp. 151–193.7. Braun, R. E. (1998) Semin. Cell. Dev. Biol. 9, 483–489.8. Braun, R. E. (2000) Int. J. Androl. 23, 92–94.9. Hecht, N. B. (1998) BioEssays 20, 555–561.
10. Kleene, K. C. (1989) Development (Cambridge, U.K.) 106, 367–373.11. Lee, K., Fajardo, M. A. & Braun, R. E. (1996) Mol. Cell. Biol. 16, 3023–3034.12. Giorgini, F., Davies, H. G. & Braun, R. E. (2001) Mol. Cell. Biol. 21, 7010–7019.13. Tomari, Y. & Zamore, P. D. (2005) Genes Dev 19, 517–529.14. Valencia-Sanchez, M. A., Liu, J., Hannon, G. J. & Parker, R. (2006) Genes Dev 20, 515–524.15. Reinhart, B. J., Slack, F. J., Basson, M., Pasquinelli, A. E., Bettinger, J. C., Rougvie, A. F.,
Horvitz, H. R. & Ruvkun, G. (2000) Nature 403, 901–906.16. Tang, G. (2005) Trends Biochem. Sci. 30, 106–114.17. Hammond, S. M., Boettcher, S., Caudy, A. A., Kobayashi, R. & Hannon, G. J. (2001) Science
293, 1146–1150.18. Jaronczyk, K., Carmichael, J. B. & Hobman, T. C. (2005) Biochem. J. 387, 561–571.19. Deng, W. & Lin, H. (2002) Dev. Cell 2, 819–830.20. Kuramochi-Miyagawa, S., Kimura, T., Ijiri, T. W., Isobe, T., Asada, N., Fujita, Y., Ikawa,
M., Iwai, N., Okabe, M., Deng, W., et al. (2004) Development (Cambridge, U.K.) 131,839–849.
21. Yu, Z., Raabe, T. & Hecht, N. B. (2005) Biol. Reprod. 73, 427–433.22. Grivna, S. T., Beyret, E., Wang, Z. & Lin, H. (2006) Genes Dev. 20, 1709–1714.23. Yelick, P. C., Kwon, Y. H., Flynn, J. F., Borzorgzadeh, A., Kleene, K. C. & Hecht, N. B.
(1989) Mol. Reprod. Dev. 1, 193–200.24. Sibony, M., Segretain, D. & Gasc, J. M. (1994) Biol. Reprod. 50, 1015–1026.25. Morales, C. R., Oko, R. & Clermont, Y. (1994) Mol. Reprod. Dev. 37, 229–240.26. Parvinen, M. (2005) Int. J. Androl 28, 189–201.27. Kotaja, N., Bhattacharyya, S. N., Jaskiewicz, L., Kimmins, S., Parvinen, M., Filipowicz, W.
& Sassone-Corsi, P. (2006) Proc. Natl. Acad. Sci. USA 103, 2647–2652.
28. Nelson, P. T., Hatzigeorgiou, A. G. & Mourelatos, Z. (2004) RNA (New York) 10, 387–394.29. Shi, H., Ullu, E. & Tschudi, C. (2004) J. Biol. Chem. 279, 49889–49893.30. Yang, J., Medvedev, S., Reddi, P. P., Schultz, R. M. & Hecht, N. B. (2005) Proc. Natl. Acad.
Sci. USA 102, 1513–1518.31. Bellve, A. R., Cavicchia, J. C., Millette, C. F., O’Brien, D. A., Bhatnagar, Y. M. & Dym, M.
(1977) J. Cell Biol. 74, 68–85.32. Kleene, K. C. (1993) Dev. Biol. 159, 720–731.33. Spirin, A. S. & Ryazanov, A. G. (1991) in Translation in Eukaryotes, ed. Trachsel, H. (CRC,
Boca Raton, FL), pp. 325–350.34. Kleene, K. C., Cataldo, L., Mastrangelo, M.-A. & Tagne, J.-B. (2003) Exp. Cell Res. 291,
101–110.35. Toyooka, Y., Tsunekawa, N., Takahashi, Y., Matsui, Y., Satoh, M. & Noce, T. (2000) Mech.
Dev. 93, 139–149.36. Noce, T., Okamoto-Ito, S. & Tsunekawa, N. (2001) Cell Struct. Funct. 26, 131–136.37. Eystathioy, T., Chan, E. K., Tenenbaum, S. A., Keene, J. D., Griffith, K. & Fritzler, M. J.
(2002) Mol. Biol. Cell 13, 1338–1351.38. Lagos-Quintana, M., Rauhut, R., Yalcin, A., Meyer, J., Lendeckel, W. & Tuschl, T. (2002)
Curr. Biol. 12, 735–739.39. Tahbaz, N., Kolb, F. A., Zhang, H., Jaronczyk, K., Filipowicz, W. & Hobman, T. C. (2004)
EMBO Rep. 5, 189–194.40. Doi, N., Zenno, S., Ueda, R., Ohki-Hamazaki, H., Ui-Tei, K. & Saigo, K. (2003) Curr. Biol.
13, 41–46.41. Meister, G., Landthaler, M., Peters, L., Chen, P. Y., Urlaub, H., Luhrmann, R. & Tuschl,
T. (2005) Curr. Biol. 15, 2149–2155.42. Pillai, R. S., Bhattacharyya, S. N., Artus, C. G., Zoller, T., Cougot, N., Basyuk, E., Bertrand.
E. & Filipowicz, W. (2005) Science 309, 1573–1576.43. Humphreys, D. T., Westman, B. J., Martin, D. I. K. & Preiss, T. (2005) Proc. Natl. Acad. Sci.
USA 102, 16961–16966.44. Liu, J., Carmell, M. A., Rivas, F. V., Marsden, C. G., Thomson, J. M., Song, J. J., Hammond,
S. M., Joshua-Tor, L. & Hannon, G. J. (2004) Science 305, 1437–1441.45. Teixeira, D., Sheth, U., Valencia-Sanchez, M. A., Brengues, M., Parker, R. (2005) RNA (New
York) 11, 371–382.46. Lerner, R. S., Seiser, R. M., Zheng, T., Lager, P. J., Reedy, M. C., Keene, J. D. & Nicchitta,
C. V. (2003) RNA (New York) 9, 1123–1137.
13420 � www.pnas.org�cgi�doi�10.1073�pnas.0605506103 Grivna et al.