movement coleoptiles zea mays l. geotropic · pdf fileduring geotropic stimulation. 1 2 ......
TRANSCRIPT

Movement of Auxin in Coleoptiles of Zea mays L.during Geotropic Stimulation. 1 2
Mary Helen M. Goldsmith and Malcolm B. Wilkins3Department of Biology, Yale University, New Haven, Connecticut
The theorv of Cholodny and Went (20) attributesthe tropic curvatures of growing plants to an unequaldistribution of auxin caused by lateral movement ofauxin in stimulated organs. The work of Dolk (3)with tips and isolated sections of coleoptiles of Avenasativa and Zea wnays provided strong support for thistheory as applied to geotropism. Subsequently, otherworkers found an unequal distribution of auxinresulting from geotropic stimulation of organs otherthan coleoptiles. (See reviews by Went and Thi-mann (20) and Anker (1).)
With the advent of radioactive IAA, the distribu-tion of auxin in geotropically stimulated organs was
reinvestigated, and several attempts failed to demon-strate an unequal distribution of radioactivity (2, 14,15). In all these experiments, an exogenous source
of radioactive IAA wvas supplied to the tissue andthen removed before the organ was stimulated bygravity. More recently, however, Gillespie andThimann (5, 6) have obtained positive results.They applied IAA-C14 to horizontal sections of Avenaand Zea coleoptiles. and collected more radioactivityin the lower than the upper block at the base of thesection. XVithin the 7-mm sections themselves, an
asymmetric distribution of radioactivity was detect-able in corn but not in oats. For corn, the ratio ofradioactivity in the upper and lower half of thereceiver was 30: 70, but within the sections only40: 60. On the basis of their observations, Gillespieand Thimann (5, 6) suggest that radioactive IAAwhich is beinig polarly transported can move laterally,but since a large fraction of the activity in the tissueis immobile, a lateral gradient of mobile radioactivitymay be obscured. Thimann and Gillespie's experi-ments differed in 2 significant aspects from theprevious attempts to demiionstrate lateral movement oflabel. First, they looked for an asymmetric distribu-tion in the activity transported through sections as
well as in the total radioactivity in the sections, andsecond, they supplied the IAA-Cl4 during instead of
Received June 17, 1063.2 This research supported in part by a research grant
GM-08886 to M. H. M. and T. H. Goldsmith from theDivision of General Miedical Sciences, Public Health Ser-vice, and by a fellowship to M. B. Wilkins from theRockefeller Foundatioin.
3 Present address: Department of Botany, King's Col-lege, London, WV. C. 2, England.
before stimulation. This second condition of theirexperiment is important because Goldsmith and Thi-mann (8) have shown that after an exogenous sourceof IAA is removed, the rate of transport declinesrapidly and virtually ceases within an hour eventhough much activity still remains in the section.
The results of Dolk and of Gillespie and Thimann,obtained by supplying isolated horizontal sections ofcoleoptiles with symmetric sources of auxin, wereopen to 2 interpretations: the asymmetic distributionof auxin (or radioactivity) is the result of A) lateralmovement of IAA within the sections or B) increaseduptake and transport on the lower as opposed to theupper side of a section. It was necessary, therefore,to design an experiment to determine whether or notlateral movement occurs in an isolated section. BothHertel (10) and Gillespie and Thimann (6) split corncoleoptiles longitudinally, applied IAA-C'4 to theapical cross-section, and looked for radioactivity toappear laterally in a receiver placed against thelongitudinal cut surface of the split sections. Gil-lespie and Thimann collected approximately 3 timesas much radioactivity in the lateral receivers whenmovement was from the upper half of a horizontalcoleoptile than from half of a vertical coleoptile.Hertel found 2.5 times as much radioactivity movedlaterally downward as upward.
In the present investigation another approach wasused to demonstrate lateral movement; both horizontaland vertical sections were supplied with asymmetricsources of IAA-C14 and the subsequent distributionof radioactivity was recorded. Asymmetric sourceshad been used much earlier by Nuernbergk (13) andby Koch (12) in studying lateral movement of auxin;and in the present work offer 2 advantages oversymmetric ones: any radioactivity found in the halfof the section opposite the source must have movedlaterally; and with upper asymmetric sources, a smallincrease in the radioactivity of the lower half of asection will be statistically significant. The presentexperiments have already been briefly reported (9),and together with those of Hertel and of Gillespieand Thimann unequivocably demonstrate lateralmovement of radioactivity in horizontal corn coleop-tiles supplied with IAA-C14.
In the previous studies on the effect of geotropicstimulation on the movement of IAA-C14, the neces-sity of demonstrating that the radioactivity is still
151 www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.

associate(l wVithl IAA has been largely neglected.Censequently 1 purpose of the present wvork Nwas todetermine the relation between the distribution ofradioactivity and geotropic curvature.
Materials and Methods
Plant Material. See(ds of Zea inays L. var. Bur-pee Snowvcross were germinated and grown in dark-ness in verimliculite as (lescribe(l elsewhere (21). Thecoleoptiles were excised 6 (lays after soaking, vhenthey were 20 to 30 mmii in length. The leaves werewithdravinI fromii their bases, anid sections 15 mim loingwere cut 3 mim below the apex of the coleoptiles.
Ini(lolc-3 -acetic A cid. Both unlabeled IAA(Hoffmlani-LaRoche) andl C14 carboxyl-labeled IAAwere use(l in these experiments.
The dlesirecl concentratioin of TAA was suppliedl tocoleoptiles in blocks (4 mm1:<) of 1.5% wxashecd agar(8). The labeled IAA wvas synthesized by Dr. BruceB. Stowve (16) and lhas a specific activity of 16.9c/mole. On chromatographic analysis (isopropanol:amnmolnial: water; 8: 1: 1), this material yields asingle radlioactive spot with an RF and color reaction(ferric chloride-perchloric aci(d reagent) i(lentical toa purifie(d samiiple of unlabeled IAA. The ultravioletabsorption of the radioactive IAA obtaine(d wvith aCary recor(linig spectrophotometer coincidles exactlywith that of unlabeled IAA and has remainedl stablefor nmore than a year.
Experimnental Procedufre. In the course of thisstudy 2 (lifferent metlhodls wrere devisecl. In the firstprocedlure both the (listribution of Cl wvithin asectioni and the amount of C14 which passedl througlhthe section into a receiving block of agar weredeterminie(l. Each coleoptile section was slippe(l overa filne glass nlee(dle whlclih wvas in turn securedl in a
block of paraffin. The base of the coleoptile sectionrestedl oni a block of 1.5% agar (20 nimi:m) that wason top of the paraffini block. The fine glass needlerani through1 this agar block, up the central hollowrof the caleoptile, and projectedl half a centinmeter fromthe apical endl of the section to support the agarsources. A sinlgle block containing a knownui anmountof IAA-Cl- wvas appliecl asymmetrically to the ap)icalcut surface or alternatively 2 blocks of the samiie sizeanicl cncentration wvere placedl symmetrically oln op-posite si(les of the coleoptile. Each block covered a
stril) of tissue 1.5 mm wiide and extended fromii thecentratl lhollow to the epidermiiis of the coleoptile.The block was always placed over the vascularbundlle. All manipulations were performedl undler adimii greeni light that did not affect the geotropicresponse of Zea coleoptiles (21).
As soonl as the sections hal(l been preplaredl. tleywere place(l in darkness in a chamber of higlhhumidity. Idlentical sections (5 or 6) were treatedlas a group and were either left vertical or placedlhorizontally. Somle horizontal groups had the asyn-nmetric sources on the upper sidles, others had thenmoni tlle lower sides. In all experimenits, the sectionsrem-naillnel horizontal until the source was remiioved
ani(l the sectiolls xxWrc split ilito upper and lowe.-halves.
At the end(l of the period of translport. a group ofsections was tralnsferredlto white liglht for the sub-se(luent mlianipulations. For identificationi, a markwith india ink was immediately place(l oln the samesi(le of eaclh section in the group. Tlhen all thesources were removed andl placedl togetlher in a singleplanchet. In the case of symmetric sources, theupper and lower sources were placed in separateplancliets. Finally, each section xvas split longitu-dinallv; horizontal sections into uppl)er and lowerhalves; vertical ones into halves un(ler- and oppositethe source. Corresponding halves were placed in thesanme planchet. W\ithin 3 minutes of their transferfrom the dark, all sections were split. Finally theagar receivers were removed from the paraffin blocksand put in still aniotlher planchet. This miietho(d hadlthe advantage that the radioactivity transportedlthrouglh the coleoptiles wvas collecte(d in receivingblocks and could be easily counted, but the obviousdisadvantage that the glass rod runninig up the centerof the coleoptiles restricted the development of thecurvature.
The purpose of the second proce(dure was to allowcurvature to develop unhinderedl. Ten decapitated15-nmmi sections were secured by pushing their basesinto a trough fille(d with 1.5%l agar (21). The agarsource was supporte(l by a short lengtlh of prinmaryleaf insertedl ilnto the hollowv center- of the apicall endof the section. In other respects. the coleoptiles \x erehand(lle(d as in the first imiethodl.
After the (lesire(l timie, the sections wxere remove(lfrom the troughs of agar alndl sha(lowgraphed torecor(l their curvature. The curlatures were inlea-sure(l Nith a protraLctor. The axera-e cutrvatul-e an(lstandard error of the mean of 10 sectionis xvere (leter-mine(l.
Immediately after shadoxvgraph i ng, radioacti Vesections wvere split ilnto 2 halves. In this procedlure,a group of 10 sections could be shadowgraphed an(lsplit in 8 minutes. 'Usually 5 sections gave samplesxvith sufficient radioactivity, but at the lowest coni-centration all 10 sections were combined in 1 sample.Witlh this method, both curvature and (listribution ofra(lioactivity were determiiined with the same sections;hoxvever, because the volume of agar in contact wviththe base of the coleoptiles was large, the radioactivitytranslocated throtuglh the coleoptiles w-as not re-
covered.Samiple Preparation anid Counting. The samples
of tissue andl agar were dried anclprelpared for count-ing as previously (lescribed (8). A Nuclear-Chicago(M\odel D-47) win(loless, gas flow couniting systemn(letected radioactiv ity. The counltinlg efficiency wasapproximately 5W0C and background xvas 20 cpm.With this system, the 1AA-C14 gave ap)proximately1.9 X 101" cpmii per mmlilole. The observed activityof each samlple wxas correctedl for backgroundl and fol-self absorption as previously (lescribe(d (8). Eachexperimilent recOr(le(l in this paper wvas repeate I oiiseveral different occasiolls.
152 1'LIAN.-T PHYSIOLOGY'
www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.
GOLDSMITH AND WILKINS-AUXIN MOVEMENT DURING GEOTROPISM
ResultsBasipetal Movement and the Distribution of
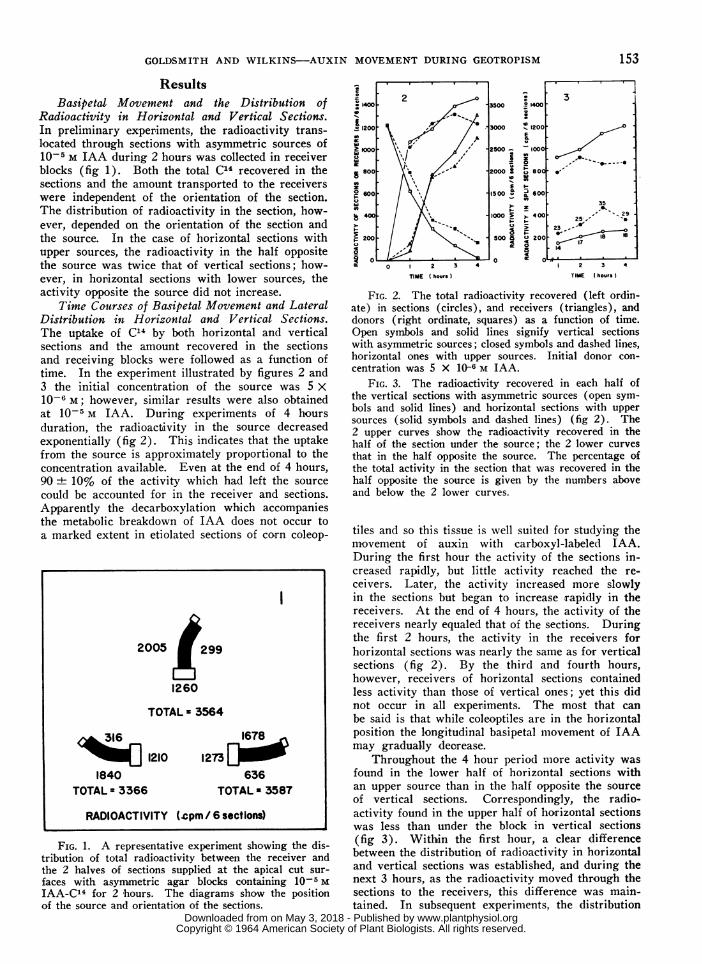
Radioactivity in Horizontal and Vertical Sections.In preliminary experiments, the radioactivity trans-located through sections with asymmetric sources of10-5 M IAA during 2 hours was collected in receiverblocks (fig 1). Both the total C14 recovered in thesections and the amount transported to the receiverswere independent of the orientation of the section.The distribution of radioactivity in the section, how-ever, depended on the orientation of the section andthe source. In the case of horizontal sections withupper sources, the radioactivity in the half oppositethe source was twice that of vertical sections; how-ever, in horizontal sections with lower sources, theactivity opposite the source did not increase.
7ime Courses of Basipetal Movement and LateralDistribution in Horizontal and Vertical Sections.The uptake of C14 by both horizontal and verticalsections and the amount recovered in the sectionsand receiving blocks were followed as a function oftime. In the experiment illustrated by figures 2 and3 the initial concentration of the source was 5 X10-6 M; however, similar results were also obtainedat 10-5 M IAA. During experiments of 4 hoursduration, the radioactivity in the source decreasedexponentially (fig 2). This indicates that the uptakefrom the source is approximately proportional to theconcentration available. Even at the end of 4 hours,90 10% of the activity which had left the source
could be accounted for in the receiver and sections.Apparently the decarboxylation which accompaniesthe metabolic breakdown of IAA does not occur toa marked extent in etiolated sections of corn coleop-
FIG. 1. A representative experiment showing the dis-tribution of total radioactivity between the receiver andthe 2 halves of sections supplied at the apical cut sur-
faces with asym,metric agar blocks containing 10-5 MIAA-C14 for 2 hours. The diagrams show the positionof the source and orientation of the sections.
0 2 3
TIME (hours)
2500 -
I
2000
E
1500&
1000 >
500 5
e 1400
E
100
z
0 Boo
F 8
3
r-
35
>. 400 "5 -29-
25,I*s> .23I 2
I. 0~03to
TIME (hours)
FIG. 2. The total radioactivity recovered (left ordin-ate) in sections (circles), and receivers (triangles), anddonors (right ordinate, squares) as a function of time.Open symbols and solid lines signify vertical sectionswith asymmetric sources; closed symbols and dashed lines,horizontal ones with upper sources. Initial donor con-
centration was 5 X 10-6 M IAA.FIG. 3. The radioactivity recovered in each half of
the vertical sections with asymmetric sources (open sym-bols and solid lines) and horizontal sections with uppersources (solid symbols and dashed lines) (fig 2). The2 upper curves show the radioactivity recovered in thehalf of the section under the source; the 2 lower curvesthat in the half opposite the source. The percentage ofthe total activity in t,he section that was recovered in thehalf opposite the source is given by the numbers aboveand below the 2 lower curves.
tiles and so this tissue is well suited for studying themovement of auxin with carboxyl-labeled IAA.During the first hour the activity of the sections in-creased rapidly, but little activity reached the re-
ceivers. Later, the activity increased more slowlyin the sections but began to increase rapidly in thereceivers. At the end of 4 hours, the activity of thereceivers nearly equaled that of the sections. Duringthe first 2 hours, the activity in the receivers forhorizontal sections was nearly the same as for verticalsections (fig 2). By the third and fourth hours,however, receivers of horizontal sections containedless activity than those of vertical ones; yet this didnot occur in all experiments. The most that can
be said is that while coleoptiles are in the horizontalposition the longitudinal basipetal movement of IAAmay gradually deorease.
Throughout the 4 hour period more activity was
found in the lower half of horizontal sections withan upper source than in the half opposite the sourceof vertical sections. Correspondingly, the radio-activity found in the upper half of horizontal sectionswas less than under the block in vertical sections(fig 3). Within the first hour, a clear differencebetween the distribution of radioactivity in horizontaland vertical sections was established, and during thenext 3 hours, as the radioactivity moved through thesections to the receivers, this difference was main-tained. In subsequent experiments, the distribution
153
2005 299
1260
TOTAL 3564
1210 1273 L _1840 636
TOTAL a 3366 TOTAL a 3587
RADIOACTIVITY (.cpm /6 sections)
www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
z0
U)
U.
4
0
049U)
z
I-
4c2
I.-z
C.)
80
70
601_
501.
401.
301
20 .
10
0
CONCENTRATION OF IAA IN SOURCE (M)CONCENTRATION OF IAA IN SOURCE (M)
:,4000 430
S~ ~2106 z4 420
3000 +10
z
> 2000 O -lO
0~ ~ ~ ~ ~ ~~ 2
0
C1000 OFU
z
0 5 10 20 25 0 5 10 20 25
CONCENTRATION OF IAA IN SOURCE (10 M) CONCENTRATION OF [AA IN SOURCE (IO"M)
FIG. 4. The total radioactivity and that recovered opposite the source in vertical (X's) and horizontal sections(circles) as a function of concentration of IAA in the asymmetric source. Shading indicates the part of the sectionconsidered. The source was present for 2 hours. Points are averages of duplicate samples. Both axes are loga-tithmic, but a straight line relationship is also obtained with linear axes.
FIG. 5. The fraction of the radioactivity in the section which was recovered in the half of the section opposite thesource, or in the case of symmetric sources in tshe lower half (shaded half of sections). For the sections with sym-metric souces, solid symbols give the fraction actually observed on the lower side, while open symbols, give the fractionpredicted from results with asymmetric sources. The 2 different symbols (squares, triangles) used for symmetricsources designate results from 2 independent experiments. The curvature of these same sections is given in figure 7.
1540
.9 4
U
S
023
z
0
I-
a
02
a.2004Oz
I-
020
4
-I I I- --
4
-|g/-I-/
* x
I I aI
I I.
5
aa~~~
X 00
xx~~~~~
I0 -1 a
1-7 10 z
www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.

GOLDSMITH AND WILKINS-AUXIN MOVEMENT DURING GEOTROPISM1
of radioactivity was determined at the end of 2 hours.Responses of Sections to Different Concentrations
of IAA in the Asymmletric Source. Sections sup-
ported in troughs of agar were given asymmetricsources from 5 X 10-7 to 2.5 x 10-5 M IAA, andafter 2 hours, both the curvature and distribution ofactivity were determined. The curvature of thesesections reached a maximum value at this time,whereas the curvature of horizontal sections suppliedwith asymmetric agar blocks without auxin was stillonly 2 to 30.
The distribution of radioactivity in the sections issummarized in figures 4, 5, 6 for 1 of these experi-ments. Over a 50-fold increase in the concentrationof the source, the radioactivity both in entire sectionsand in each half of the sections increased linearly(fig 4); therefore, the ratio of radioactivity betweenthe 2 sides of the section was independent of concen-
tration (fig 5). Although the total radioactivity in
horizontal and vertical sections was the same at eachconcentration, more activity was found in the lowerhalf of horizontal sections with upper sources thanin the half opposite the source in vertical sections(lower 2 curves, fig 4). Approximately 10% of theactivity of vertical sections and 25% of that in hori-zontal sections with upper sources was found in thehalf opposite the source (fig 5). It also follows fronmthe linear increase in the radioactivity of each halfof the section that the difference between the radio-activities of the 2 sides increased linearly with con-
centration (fig 6).About 4% of the sections' activity was recovered
in the half opposite lower sources. This was abouta sixth the activity recovered in the half oppositeupper sources, and it was consistently less than theactivity opposite the source in vertical sections (fig5).
Figure 7 shows the curvature of the sections fromwhich the data in figures 4, 5, and 6 were obtained.The results of 3 other experiments, including 2 withunlabeled IAA, were similar to those in figure 7.Both vertical sections and horizontal ones with lowersources curved away from the asymmetric source,and as the concentration of IAA in the source in-creased, this curvature approached a maximum value.The curvature of horizontal sections with a lowersource slightly exceeded that of vertical ones; how-ever, the continuing increase in curvature of thesehorizontal sections with concentrations greater than10-5 M (fig 7) was not observed in 3 other experi-ments. On the other hand, at low concentrationshorizontal sections with upper sources curved towardsthe source, but as the concentration increased, the
upward curvature decreased to zero and then slightdownward curvature occurred (fig 7, 8). At thehighest concentrations, these sections became dis-tinctly S-shaped; the persistent upward curvaturewas localized near the base of the section and a down-ward curvature appeared immediately under thesource. For S-shaped sections only the resultantcurvature was measured.
In these experiments, the curvature of the hori-zontal sections with upper sources was usually notsignificantly different from that of decapitated coleop-tiles without IAA. In 3 out of 4 experiments, how-ever, there was an upward curvature towards a
source of 10-6 M IAA that was significant at the 5%level (fig 8). A more important point, however, isthat at 10-6 3& and higher concentrations, the curva-
ture of horizontal sections away from the source was
always significantly less (at well below the 1% level)than that of vertical sections. The geotropic re-
sponse-that is, the difference between the curvatureof vertical and horizontal sections with equivalentsources (fig 9)-rose sharply with concentration ofIAA and reached a maximum at about 5 X 10-6 MIAA.
Distributtion of Radioactivity and Cutrvatutre ofHorizontal Sections with Synmmetric Sources. After2 hours, sections with symmetric sources containedtwice as much activity as those with asymmetricsources of equal concentration (fig 4). Over a range
of concentration from 2.5 X 10-7 to 2.5 X 10-5 MIAA, equal amounts of C14 were recovered in eachhalf of vertical sections while 54 to 59% of theradioactivity was found in the lower half of horizontalsections. This asymmetry was essentially independ-ent of concentration (fig 5). Gillespie and Thimann(6) found that the asymmetry in receivers was in-dependent of the source concentration between 2.7and 5.7 X 10-6 M. At approximately 10-6 M, how-ever, they reported a slight but statistically significantincrease in asymmetry. At present this discrepancybetween our results and those of Gillespie and Thi-mann can not be resolved.We undertook to see how the distribution of radio-
activity in sections with symmetric sources agreeswith the average distribution of the 2 differenthorizontal sections with asymmetric sources. Withlower sources the ratio of the radioactivity in thelower to that in the upper halves of sections was
approximately 95: 5; with upper sources, approxi-mately 25: 75 (fig 5). If lateral redistributionproceeds independently of the concentration on eitherside of the section, the expected distribution in sec-
tions with symmetric sources would be 60: 40.
<(JFIG. 6. The difference in radioactivity of the 2 halves of sections supplied with radioactive IAA for 2 hours. Points
are averages of duplicate samples.FIG. 7. The curvature after 2 hours as a function of concentration of IAA-C14 in the agar source. The mean
curvature of 10 sections and standard error of the means are given. By convention, curvatures towards an asymmetricsource are plotted as positive; curvatures away as negative. Upward curvatures with symmetric sources are assignedpositive values. The amount and distribution of radioactivity in these sections is shown as a function of source con-centration in figures 4, 5, and 6.
155
www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.
PLANT PHYSIOLOGY
0 2 4 6 8 10CONCENTRATION OF IAA IN THE ASYMMETRIC
SOURCE ( x 16M )
r.I I ..
0~~~~~I"\ 10
\\\\
\\v0--- A-- 0~- H,
- ~ ~~~ ~~~~b_¢E
I I II
O 1 2 3
TIME SECTIONS HAVE DRAINED ( hours)
20L
4
4
Z
n
I-
zwC')
w
wa
30
wU
0Ut)
w
-Jo
44r
P0a Jn
U. I
Z
Z_Z
O
zz
wo
W. c)
201
0
0 5 10 15 20 25
CONCENTRATION OF IAA IN THE ASYMMETRICSOURCE ( x 10 6 M )
0 2 3 4TIME AFTER SJPPLYING SOURCE ( hours)
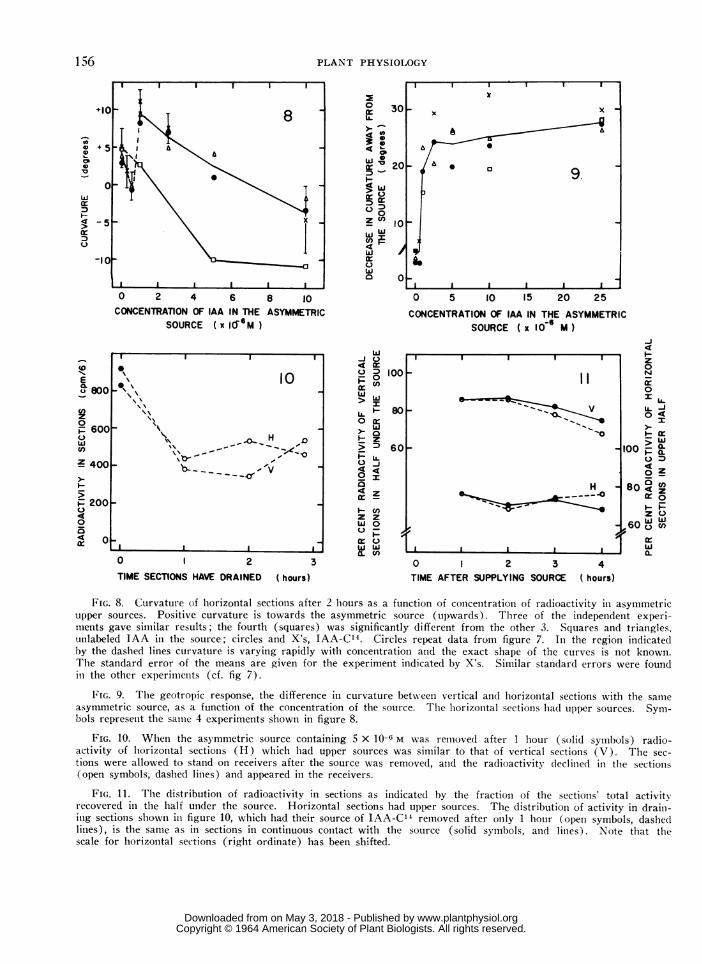
FIG. 8. Curvature of horizontal sections after 2 hours as a function of concentrationi of radioactivity in asymmetricupper sources. Positive curvature is towards the asymmetric source (upwards). Three of the independenit experi-nients gave similar results; the fourth (squares) was significantly different from the other 3. Squares and triangles,unlabeled IAA in the source; circles and X's, IAA-C14. Circles repeat data from figure 7. In the region indicatedby the dashed lines curvature is varying rapidly with concentration and the exact shape of the curves is not known.The standard error of the means are given for the experiment indicated by X's. Similar standard errors were foundin the other experimenits (cf. fig 7).
FIG. 9. The geotropic response, the difference in curvature between vertical and horizonital sections with the sameasymmetric source, as a function of the concentration of the source. The horizonital sections hadl upper sources. Sym-bols represent the samie 4 experimenits shown in figure 8.
FIG. 10. Wheni the asymmetric source containing 5 X 10-t6 M was removed after 1 hour (solid symbols) radio-activity of horizonital sections (H) wvhich had upper sources was similar to that of vertical sections (V). The sec-tions were allowed to stand on receivers after the source was removed, and the radioactivity declined in the sectioIns(open symbols, dashed lines) and appeared in the receivers.
FIG. 11. The distribution of radioactivity in sections as indicatedl by the fraction of the sectionis' total activityrecovered in the half under the source. Horizontal sections had upper sources. The distribution of activity in drain-ing sections sl1ownI iii figure 10, which had their source of IAA-C14 removed after only 1 hour (open symbols, dashe(dlines), is the same as in sections in continuous contact with the source (solid symbols, and lines). Note that thescale for horizointal sections (right ordinate) has been shifted.
156
I I I I I I
8 x
A _
.
0 9.
i0I
I I I I I I
+10
co5
V
0
IdD
5 -5
E0
-0
P 600
cn
84000
s 600
ir
O-
10 _
10I~~~~~~~~~~~~ _
H
I I I I
I-z0N
0
Il
100FC 4
5; z
80
60 i icr0p
_ L)
l I I I
I I I I I
www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.

GOLDSMlITH AND WN'ILKINS-AUXIN MOVEMENT DURING GEOTROPISM
Above a concenitration of 5 X 10-6 M[, the distributionof activity in horizontal sections with symmetricsources agreed closely with that predicted from sec-tions with asyimmetric sources (fig 5). Even belowthis concentration, the asymmetry in sections withsymmetric sources was not significantly less thanpredicted.
Horizontal sections with symmetric sourcescu,rved upwards (fig 7). This curvature reached amaximum about 10-6 M, remained constant as theconcentration of the source rose tenfold, and then fellalmost to zero above 10-5 M.
Distributtioni. of Mobile and Imm1zobile Radioactivityin Sections. A major fraction of the C14 in sectionsof Avena coleoptiles is immobile and is not exportedto a receiver even if the source is removed (8). Bothhorizontal ancd vertical sections of Zea coleoptileswere compared to determine if geotropic stimulationinfluences either the total amount or the distributionof immobilized activity. The initial source concen-tration was 5 X 10-6 m, and horizontal sections hadupper sources. At the end of an hour, the asym-metric sources were removed but the original receiv-ers were left at the basal ends. During the firsthour after the asymmetric source was removed, theactivity of the sections declined 50 to 60%, but withlonger collection periods it did not decrease further(fig 10). Accordingly, after 1 hour of export, theradioactivity of sections was largely immobile.Horizontal and vertical sections contained the sameamount of immiiiobile activity (fig 10).
After the source was removed, the radioactivityof the sections was compared at intervals with thatof sections supplied with IAA-C14 throughout theexperiment. During export without a source, thedistribution of radioactivity across vertical sectionsdid not differ significantly from that across verticalsections kept in constant contact with the donor.The same result was obtained for horizontal sections(fig 11). This shows that the amount of activityimmobilizedl and the amount transported longituclin-ally in each half of the sections are proportional tothe total activity present in that half. Geotropicstimulation thus changes the distribution of immobileradioactivity but not the proportion of the total radio-activity immobilized.
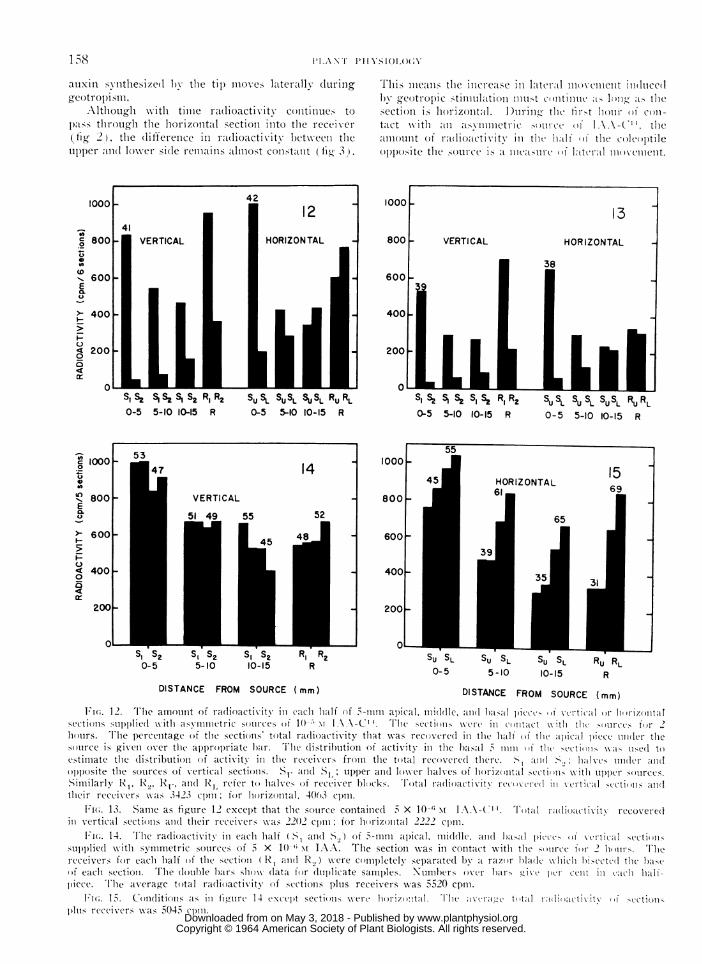
The Distributiont of Radioactizity Across theSection at Different Distances fromn the Source.In these experiments, vertical sections with asym-metric sources and horizontal sections with uppersources of IAA-C14 were halved longitudinally andeach half wvas further subdivided transversly into3 pieces each 5 mm long. In both horizontal andvertical sections the radioactivity in the half beneaththe asymmetric source decreased from the apical tothe basal end of the sections, while in the oppositehalf of the sections, radioactivity increased with dis-tance from the source (fig 12, 13). The gradientacross the apical 5-mm portion of both vertical andhorizontal sections was approximately 10: 1. Bycontrast, in the basal 5 mm the gradient across hori-zontal sections almost disappeared while across ver-
tical sections it was still approximately 3: 1. Theseresults were independent of the concentration in thesource; that is, the gradient across each 5-mm pieceof the sections in figure 12 (source equal to 10-5 MIAA) was not significantly different from the cor-responding gradient in figure 13 (source equal to5 X 10-6 M IAA). Likewise, the fraction of thetotal radioactivity recovered in each longitu(dinalhalf of 15-mm sections was also independent of thesource concentration (fig 5). Finally, in all sections,independent of both orientation and source concentra-tion, about 40% of the sections' total activity wasrecovered in the first 5 mm of the half directly underthe source.
With symmetric sources, the radioactivity in bothhalves of vertical and horizontal sections decreasedfrom the apical to the basal end. As in previousexperiments, equal radioactivity was recovered inthe 2 halves of a vertical section (fig 14), while thelower half of an horizontal section contained moreactivity than the upper half (fig 15). In the apical5-mm pieces of horizontal sections, the differencein the radioactivity of the 2 halves was barely signifi-cant; 55% of the activity was recovered on the lowerside. In the middle and basal pieces of the section,the proportion of the radioactivity in the lower halfincreased to 61 and 65% respectively. The actualdifference in radioactivity between the upper andlower halves was approximately twice as great inboth the middle and basal pieces as in the apical one.In this experiment, the receivers for the 2 halvesof the sections were completely separated b) a razorblade which penetrated 2 mm into the basal piece ofthe sections. With vertical sections, the radlioactivityin the 2 receivers was approximately equal. Bycontrast, at the base of horizontal sections 15 mm inlength, 70% of the total activity was in the lowerreceiver. In all approximately 1.6 times as muchactivity remained in or had passed through the lowerthan the upper half of horizontal sections with sym-metric sources.
Discussion
The Existence and Amount of Lateral Movemiient.The lower halves of horizontal sections with uppersources have more activity than the halves of verticalsections opposite the source. Since the total radio-activity recovered in sections and receivers is thesame for both orientations (fig 1) the altered distri-bution is not attributable to a differential loss ofradioactivity. Furthermore, since immobile activityincreases in the lower halves in direct proportion tothe increase in total activity of these halves, immobili-zation of radioactivity is not directly responsible forthe increased activity in the lower halves. Radio-activity recovered in the half of a section oppositean asymmetric source must have moved laterallywithin the tissue. Our results thus support the viewthat radioactivity taken up from an asymmetric sourceof IAA-C14 moves laterally in vertical sections, andthat this movement is increased by geotropic stimula-tion. This suggests that in intact coleoptiles the
157
www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.
PL N 1 P11 XSlOlA Y
auxini svnthesize(d lbv tlle til) illoves laterally (luringgceotro)i sin.
A1tlhoughi xN it11 timie radioacttiv itv continues top)as throtu l the horizontal sectioin iilto the receiverhig 2 ), tlle ilifterelnce in ra(lioactivity hetwxeen the
ul)p)er anlt lower si(le remliainis almost constant ( ti 3
''lhii means the increase in lateral nIovemnent iin(lItCcel)v g,Cotlrol)iC stimtiulati on1i1clit ilitilltne a ing sa theoection ihozorizntal. D)nrintz the fir't hlonei- 0o con0-
t1act xxwith an asvnmmetric. in c l \ \(I'A thle(In1lotinlt of radliotctiv ity in the llt ol the c( oItt)tileopposite the soUl-rce ia me n ()I Ilatttrdl n]ox ciieilt.
42
1241
VERTICAL HORIZONTAL
LL hII1 S2 S S2 SI S2 R R20-5 5-10 10-15 R
SUSL SUSL SUSL RURL0-5 5-10 10-15 R
S0 S2 S5 S2 S0 SI RI R20-5 5-10 10-15 R
SU SL SU SL SUSL RU RL0-5 5-10 10-15 R
VERTICALC. I A e 55
15L 69
65 I
52 1 1 20-5 5- 10 10-15
'i K2R
DISTANCE FROM SOURCE (mm)
SU SL SU SL SU SL Ru RL0-5 5-10 10-15 R
DISTANCE FROM SOURCE (mm)
It(.. 1 2. TIhe amountlt of radioactixitv in each half of 5-mm apical, middle, and haslal pieee> '0 xcrtil il oli li1iiizontalIsectionIs sul)iplied w ith asymmetric so r(i ces of 1() I1\\1-t seCti()ins x (ere in (mta ct \\xtx l the ( mIr , f( I 2hours. Tlle per-cenItage of tlle sections' total radjioacttixity that was recovtre(l ill tlle hali of thle apical piece io(ler the
ssource is givenl over thle ap)proprl~iate} hair. Ti'le distrihution of activitx in the hasal nin (if tihi secti' 11nscd to
estimiate the (listributioni of activity in thle receivers fromii the total recovere(l thlere. >a >I halxves mlei(-r and
olplposite tlle sources of vertical sectiolis. SU. anid( SI,; uIper anld loxer halves of lhorizo ntal secti(ms wxith utipper si mnrces.SimillaI-Ir R1p R, Rj-. and 1 refer to halves of receiver b1)1cks. Total radioacti xittv re( )xc-ic(I inI xvtit ical sect D'i( and1(thieir reccivers as 3423ct p ; for hlorizontal, 4003 cpm.
Fi(;. 13. Sami1e as figure 12 except that the sotirce containie(d X 10- II.\A-( ". T(1tal radio.ttixvitv recovered
in vertical sectiols and(I their receixers xxas 2202 cpmi for rizontal 2222 cpm.
UFn;. 14. The radioactivity in eachi lhalf S1 ad(I S.,) of 5-mm ta)ical. middle, and hasal pictt ()I' xtctitcal se(ttiiSnsupipiliedl jith symmetric soutrces of 5 X 10 x; IA A. Tlle section xwas in contact xvith the Si nrece fir- 2 ]Imrs. Tliereceivers for each lhalf (of th1c section (IN I aIn(I R.,) xere commpletelv sep)aratetl 1y a razor haldeltx\vhlih hi sectil tll s)ct(f eachi sectioi. The douhle h)airs shoxx data for duplicate samples. Numbers over hars Jt t tnIt iIi eat hall-piece. Th'lle average total radlioactixity of sectioins plus receivers xxas 5520 cpmli.
Fi'( 15. Conditions as in fig,ure 14 textcept secti(nis vert Ii '1talIIhr1ax-t tilt1il hiCtti cetioii1hlIs receiverswx as 5045 (lIiip.
158
1000
800 _
6001_
400 _
200 _
c
(0co'._
EC)
00CL
cr:
)k
14c 10000
S
U) 800E
>- 600F-5:
4 4000
5r
1000
800
)-
1-
HORIZONTAL61 _
2001
6001-
400k
200-
mU _6
OL
www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.

GOLDSMITH AND WILKINS-AUXIN MOVEMENT DURING GEOTROPISAI
After the first hour, however, activity which hasmoved laterally starts to pass into the receiver blockand the total amount that has moved laterally is theactivity opposite the source in sections plus receiver.After 2 hours with a source of 5 X 10-6 M, a totalof 402 cpm has moved laterally in vertical sectionsand 716 cpm in horizontal ones (fig 13). This isequivalent to 3.5 X 10-12 mole of IAA moved later-ally in a single vertical section and 6.3 X 10-12 molein a horizontal one.
Dolk (3) reported that receivers in contact withthe lower half of coleoptile tips contained approxi-mately 60% of the total auxin activity, whereasupper receivers accounted for 40%. He suggestedthat the asymmetry in the distribution of auxin inthe receivers increased with the length of the tips.The present experiments confirm Dolk's suggestionand show that this occurs because radioactivity moves
laterally all along the length of 15-mm sections (fig12-15). The asymmetry in either sections or re-
ceivers changes with distance from the source becauseat any point the activity on the lower side is the sum
of activity that moved laterally in the immediatevicinity plus that which moved laterally in the more
apical regions of the coleoptile and subsequentlyunderwent longitudinal transport.
The net amount of radioactivity which moves
across each successive 5-mm piece of longer sections(table I) was calculated from the following relation-ship:
C14L = (C14A C14B) C14S,where C14L is the amount moving laterally from theupper half-piece of horizontal sections (or the half-piece under an asymmetric source of vertical sec-
tions); C14A, the amount entering the half at itsapical end; C14B, the amount leaving its basal end;C14s the amount remaining in the half-piece. Thenet amount of lateral movement as a fraction of theradioactivity which enters each successive 5-mmhalf-piece of the 15-mm sections is constant or may
even slightly increase from the apical to basal end ofthe section (table I). Since only 10 to 30% of thetotal radioactivity which enters the upper half of any
5-mm piece moves laterally (table I) and since con-
siderable radioactivity is recovered in the upper halfof receivers (fig 15), it is clear that only part of themobile activity in the upper half of a section is di-verted to the lower side during geotropic stimulation.Assuming the cells to be as long as 0.5 mm, each celldiverts laterally only 1 to 3% of the radioactivity itreceives. The suggestion that a large part of theauxin in the upper half of a horizontal coleoptilecontinues to move in the longitudinal direction was
first advanced by Dolk (3). Koch (12) supported
Table IAimount of C14 That Moves Laterally in Successive 5-mm Pieces of 15-mm Sections Supplied with IAA-CI4
Amount Amountentering mount entering Amounteacpice moving %7 moving ofc halfe moving %movingunder laterally laterally under laterally laterallysource (cpm) source (cpm)(cpm) (cpm)
Horizontal sections withsymmetric sources
Total cpm in 5 sections + 5 receivers(Source = 5 X1O-6 M) 5045*
0- 5mm piece 2522* 274* 10.85-10 mm piece 1440 196 13.610-15 mm piece 770 121 15.7
Vertical sections with Horizontal sections withasymmetric sources upper sources
Total cpm in 6 sections + 6 receivers(Source = 5X1O-6 M) 2202** 2222
0- S mmpiece 2202** 133** 6.0 2222** 196** 8.85-10 mm piece 1540 165 10.7 1372 328 23.810-15 mm piece 1080 104 9.6 765 192 25.1Total .... 402 18.2 .... 716 32.2
Total cpm in 6 sections + 6 receivers(Source = 10-5 M) 3423*** 4063
0- 5mmpiece 3423*** 179*** 5.2 4063*** 658*** 16.25-10 mm piece 2410 173 7.2 2400 682 28.410-15 mm piece 1690 267 15.8 1290 344 26.6Total .... 619 18.8 .... 1684 41.4
* Calculated from data of figure 15.** Calculated from data in figure 13.
*** Calculated from data of figure 12.
159
www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
Dolk's view with the finding that decapitated hori-zontal coleoptiles with an upper source remain nearlystraight. Our results, like Koch's fail to confirmNuernbergk's report (13) that curvatures of verticalcoleoptiles with asymmetric sources are the same
as those of hor-izontal ones with either upper or lowersources.
Gillespie and Thimann (5, 6) found an unequaldistribution of radioactivity in receivers at the baseof horizontal 7-mm sections of Avena coleoptilessupplied with symmetric sources of IAA-C14 but didnot detect an asymmetrical distribution in the sectionsthemselves. They suggested that the immobileradioactivity in the oat sections masks the asym-metry of mobile radioactivity (6). This may be so,
but it is also possible that 7-mm sectionis of Avenawith symmetric sources were not long enough for a
detectable asymmetry of radioactivity to developwithin the tissue. Since an asynmmetry can be de-tected with,in a 7-mm horizontal section of Zea (fig15 and reference 6), each cell of the coleoptile of Zeamust divert laterally a larger fraction of its radio-activity than does a cell of Avena. Furthermore, theasymmetry in a receiver at the base of a section isalways greater than in the entire sectioni because thedistribution in the receiver is that of the most basalcells of the sections.
Other workers (2, 14, 15) failed to detect unequaldistribution of radioactivity in sections a centimeteror longer because they removed the source of IAA-C14 before stimulation. Under these conditions, mostof the radioactivity in the sections is probably im-mobile (4, 8).
The Mechanismi, of Lateral Mlovemnent. Threekinds of evidenice indicate that diffusion cannotaccount for the lateral movement in either vertical or
horizontal sections. A) Where movement across
successive 5-imm pieces of the section is down a
gradient of radioactivity (fig 12, 13), the amount(table I) is not proportional to the gradient. B)Under conclitions where the radioactiv-ity of the lowerhalf exceeds that of the upper (fig 15), net movement
of radioactivity (table I) apparently proceeds againsta concentration gradient. C) In horizontal sections,lateral movenment is polarized; it occurs muore readilyfrom the upper to the lower side (fig 5).
The upper half of a horizontal secti on with an
asymmetric lower source has significantly less activitythan the opposite half of a vertical section (fig 5).This could result fromi an actual decrease in move-
nment from lower to upper side, or an acceleratedtransport from upper to lower side which immediatelyreturns any activity that moves upwvard from thelower side. There is no evidence to distinguish be-tween these possibilities; wvhichever is correct, theresult is a preferential direction of auxin transport.In vertical sections with asymmetric sources as muchas 18% of the radioactivity moves laterally (table I);therefore, completely restricting upward movenmentin horizontal sections with symmetric sources couldlead to a maximunm ratio betweein the upper and lowerhalf of 41 to 59.
Lateral movement of radioactivity in sections ofZea coleoptiles exhibits several properties in commonwith the longitudinal movement of auxin. A) It ispolarized from the upper to the lower side of thesectionis just as the movement of radioactivity froma source of IAA-Cl4 through Zea coleoptiles is polar-ize(I from apex to base (10, and unpublished observa-tions of authors). B) Lateral movement proceedsindependently of the gradient of radioactivity be-tween the upper and lower halves of the sections justas movement of auxin occurs independently of thegradient between apex and base of sections of Avenacoleoptiles (7, 17, 18). C) Lateral movement occursthroughout the section, as does basipetal movement.D) Lateral movement must occur in parenchymacells, which in Zea coleoptiles are also capable ofbasipetal transport of radioactivity from a source ofIAA-C14 (10, an(d unpublished observations of theauthors).
On the other haand, in longitudinal transport theorientation of the coleoptiles does not influence polar-ity (17, 11), anid either does not affect the totalamount of transport, as in Avenia (17), or affects itonly slightly, as in Zea (11). In the case of lateraltransport in Zea coleoptiles, however, both directionand amount are greatly dependent upoon orientation.The rapid increase in lateral movement on geotropicstimulation is not necessarily accompanied by a de-crease in the total amount of basipetal transport ina section (fig 1); lateral movement increases in lessthan an hour (fig 3), wvhereas the total amount oflongitudinal movement is affected more slowly if atall (fig 2). Furthermore, the geotropic curvatureis fully developed before any decrease in total basi-petal transport of radioactivity is detectable. Ap-parently lateral movement makes more auxin avail-able for basipetal transport on the lover than theupper side of a horizontal coleoptile. At present.the simplest assumption is that orientation (leterminesthe distribution between lateral and basipetal trans-port of auxin in an individual cell anid that lateralmovement occurs by the samiie nmechanism as basi-petal transport of IAA.
Relations/iip Between DistribIttioi of Radioacti.-ity and Cuirvatufre. An important question is wlhetherthe movement of radioactivity in corn coleoptilesreflects the movement of IAA. Hertel (10) suppliedsections of corn coleoptiles witlh donors containingcarboxyl-labeled IAA for up to 90 miniutes andrecovered all the radioactivity in extracts of the sec-tions ancI their receiving blocks as IAA. Goldsmithand Thimann (8) gave IAA-C'4 to Avena coleoptilesand presented evidence that IAA is the only radio-active substance transported to receivers. Qualita-tively, the present experiments also support the ideathat movement of radibactivitv reflects the nmovementof IAA. For example, not only is lateral move-ment of radioactivity greater in horizontal sectionswith an upper source than in vertical sections withan asymmetric source, but the curvature of thesehorizontal sections away- froml the source is mlarkedlyless than vertical sections (fig 7,9).
160
www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.

GOLDSMITH AND WILKINS-AUXIN MOVEMENT DURING GEOTROPISM 6
We must, however, account for the observationthat uptake and lateral movement of radioactivity(fig 4, 6) but not curvature (fig 7) increase linearlywith external concentration. The curvature of asection depends on the difference in growth of the 2halves. Since the amount of longitudinal growthexponentially approaches a maximum with increasingauxin concentration, the amount of curvature causedby a difference in concentration of auxin in the 2halves will depend on both the magnitude of thisdifference and on the absolute amount of auxin pres-ent in each half. If the radioactivity in the sectionreflects the distribution of auxin, the growth willsaturate first in the half with the highest radioactivityand curvature will approach a maximumii. Furtherthe curvature of horizontal sections with symmetricsources will decline at high concentration because theconcentration of radioactivity on each side of thesection becomes sufficient for maximum growth.
The fact that the curvature of vertical sectionswith asymmetric sources and horizontal ones witheither lower or symmetric sources can be meaning-fully related to the average radioactivity in eachhalf of the section may, however, be fortuitous.Nothing eliminates the possibilities that 1) onlypart of the radioactivity in the coleoptile has auxinactivity and only this radioactivity moves laterallyand that 2) sensitivity to auxin varies along thelength of the section (19,3). That such considera-tions must also be taken into account is made clearby the observation that horizontal sections withupper sources always contain about 75% of theirradioactivity in the upper half and about 25% in thelower, and yet these sections do not curve signifi-cantly downward and at low concentration may evencurve upward (fig 8). If the growth of the basalregions of the section is more sensitive to auxin,curvature would be determined mainly by the amountand difference in radioactivity between the 2 halvesin this region (fig 12-15). In the basal portion ofhorizontal sections with asymmetric sources, thedifference in radioactivity is small or else the lowerside has only slightly greater activity (fig 12, 13).This could account for these sections remainingnearly straight or bending upward only slightly. TheS-shaped curvature of horizontal sections with uppersources also provides support for the view that thebasal end of these sections is more sensitive to auxinthan the apical end; although the difference in radio-activity is much greater at the apical than the basalend of the section, the downward curvature just underthe source only appears at high concentration andthen about equals the upward geotropic curvatureat the base. Furthermore, the curvature of hori-zontal sections with symmetric sources may be aboutthe same magnitude as vertical ones with a twofoldgreater concentration in asymmetric sources, (fig 7),and for both sections the difference between theradioactivity of the 2 halves at their basal end issimilar (fig 6, 12,15).
The question whether the asymmetric distributionof auxin caused by lateral movement is sufficient by
itself to account for geotropic curvature of Zeacoleoptiles will be pursued further in future work.
Summary
Sources containing C14 carboxyl-labeled indole-acetic acid (IAA) were placed either svnmmetricallyor asymmetrically on the apical cut surface of 15-mmsections of Zea w1lavs L. coleoptiles wlhiclh wereoriented either horizontally or vertically. The asym-metric source was placed on either the upper or thelower half of the apical surface of horizontal sections.The amount of radioactivity in each longitudinal halfof the sections and the curvature of the sections wasdetermined as a function of time and concentration(2.5 x 10-7 to 2.5 x 10-5 -I) of IAA in the donor.
During the first 2 hours, uptake and longitudinaltransport of radioactivity are the same in horizontalas in vertical sections; hos-ever, w-ithin the first houran increased lateral movement of radioactivity isevident in the horizontal sections. This enhancedlateral movement persists for at least 4 hours.
Vertical sections with svmminetric sources haveequal radioactivity in each half. In vertical orhorizontal sections with asymmetric sources, thetotal radioactivity as well as the difference betweenthe radioactivities of the 2 halves increase linearlywith the concentration of IAA in the source, and sothe asymmetry in the sections is independent of con-centration. Approximately 10% of the radioactivityin vertical sections is recovered in the half oppositean asymmetric source. wvhereas in horizontal sectionsabout 25% is found opposite an upper source butonly 4% opposite a lower source. Geotropic stimu-lation polarizes the lateral transport: lateral move-ment across vertical sections is less than towardsthe lower side of horizontal sections but more thantowards the upper side of horizontal sections.
Radioactivity can move laterally anywhere alongthe length of a 15-mm section after which it canagain be transported longitudinally. Consequentlythe asymmetry in both sections and receivers varieswith the length of the sections. At any (listance fromthe apical end of the section, only about 3% (or less)of the radioactivity arriving at a given cell is divertedlaterally.
The same amount of radioactivity is immobilizedin horizontal and vertical sections. Since the amountimmobilized is proportional to the total radioactivity,immobilization is not the cause of the increasedradioactivity in the lower half of horizontal sections.
The curvature of vertical sections away fronm anasymmetric source increases to a miaximunm withconcentration, and at concentrations of 10-6 Mt andhigher, the curvature of horizontal sections awayfrom an upper source is always highly significantlyless than that of the vertical sections. This differenceindicates that lateral moivement of radioactivity inhorizontal corn coleoptiles reflects the movement ofIAA.
Lateral movement does not occur by diffusion.Since lateral movement has properties in common
161
www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
with the longitudinal movement of auxin, the samemechanism may be responsible for both. In thiscase, the orientation of the tissue may simply affectthe ratio between lateral and basipetal transport ofauxiI1.
Acknowledgments
We are grateful to the Yale University Committee onAtypical Growth for a grant that provided the IAA-C14used in this study and to Dr. Bruce B. Stowe who per-formed the synthesis, to Professor Arthur W. Galston forthe use of counting and other equipment, and to Drs.Timothy H. Goldsmith and Beatrice AI. Sweeney forhelpful discussions.
Literature Cited
1. ANKER, L. 1962. Ortho-geotropism in shoots andcoleoptiles. p 102-52. In: Encyclopedia of PlantPhysiology, XVII pt 2, W. Ruhland, ed. Springer-Verlag, Berlin.
2. CHING, T. M. AND S. C. FANG. 1958. The redis-tribution of radioactivity in geotropically stimulatedplants pretreated with radioactive IAA. Physiol.Plantarum 11: 722-27.
3. DOL K, H. E. 1930. Geotropie en Groeistof. Dis-sertation, Utrecht; English transl. by F. Dolk-Hoekand K. V. Thimann, 1936. Rec. Trav. Botan.N6erl. 33: 509-85.
4. GILLESPIE, B. AND W. R. BRIGGS. 1961. Mediationof geotropic response by lateral transport of auxin.Plant Physiol. 36: 364-68.
5. GILLESPIE, B. AND K. V. THINMANN. 1961. The la-teral transport of IAA-C14 in geotropism. Expe-rientia 17: 126-29.
6. GILLESPIE, B. AND K. V. THIMANN. 1963. Trans-port and distribution of auxin during tropistic re-sponse. I. The lateral migration of auxin ingeotropism. Plant Physiol. 38: 214-25.
7. GOLDSMITH, M. H. M. 1959. Characteristics ofTranslocation of IAA in the Coleoptile of Avena.Ph.D. Thesis. Radcliffe College, Cambridge,Massachusetts.
8. GOLDSMITH, MI. H. M. AND K. V. THIMIAN-N. 1962.Some characteristics of moveement of IAA in coleop-tiles of Avena. I. Uptake, destruction, immobil-ization, and distribution of IAA during basipetaltranslocation. Plant Physiol. 37: 492-505.
9. GOLDSMITH, M. H. NM. AND M1. B. W ILKINS. 1962.New evidence for lateral molvement of auxin insections of maize coleoptiles stimulated by gravity.Plant Physiol. 37: xvii.
10. HERTEL, R. 1962. Der Auxiiitrainsport in derKoleoptile von Zea wiaYs L. Ph.D. Thesis. Lud-wig-Maximilians Uliiversitdit, Miiunchen.
11. HERTEL, R. AND A. C. LEOPOLD. 1962. Auxilntrains-port und Schwerkraft. Naturwisseiischaftei49: 377.
12. KOCH, K. 1934. Untersuchunigeni uiber den Quer-und Langstransport des \Vuchsstoffes in Pflanzein-organen. Planta 22: 190-220.
13. NUERNBERGK, E. 1933. tVber dein Auxin-Quer-transport und den Geotropismus der Aveena-Koleop-tile: Einfluss der Dekapitation. Flora 28: 99-110.
14. REISENER, H. J. 1957. Versuche zurI1 Geotropismusmit radioaktiver (-Indolylessigs'aure. Naturwiss-enschaften 44: 120.
15. REISENER, H. J. AND H. SIMiON. 1960. \NVeitereGeotropismus-Versuche mit radioaktiver f-Indoly-lessigsaure. Z. Botan. 48: 66-70.
16. STOWE, B. B. 1963. Synthesis of high specific ac-tivity C14-carboxyl IAA and of C14-nitrile ind(ole-acetonitrile. Anal. Biochem. 5: 107-15.
17. WEIJ, H. G. VAN DER. 1932. Der 'Mechanisnitus desWuchsstofftransportes. Rec. Trav. Botan. N6erl.29: 380-496.
18. WEIJ, H. G. VAN DER. 1934. Der 'Mechanismus desWuchsstofftransportes II. Rec. Trav. Botan.Neerl. 31: 810-57.
19. WENT, F. W. 1928. WN-uchstoff und \Vachstum.Rec. Trav. Botan. N6erl. 25: 1-116.
20. WENT, F. W. AND K. V. TIII'IANN. 1937. Plhyto-hormonies. Macmillan, New York.
21. WILKINS, .4. B. AND 1M. H. M4. GOLDSrMITH. 1964.The effects of red, far-red, and blue light on thegeotropic response of coleoptiles of Zca miavs. J.Exptl. Botany 14 (in press).
162
www.plantphysiol.orgon May 3, 2018 - Published by Downloaded from Copyright © 1964 American Society of Plant Biologists. All rights reserved.