new species of slender salamander, genus … 2002 by the american society of ichthyologists and...
TRANSCRIPT

q 2002 by the American Society of Ichthyologists and Herpetologists
Copeia, 2002(4), pp. 1016–1028
New Species of Slender Salamander, Genus Batrachoseps, from theSouthern Sierra Nevada of California
DAVID B. WAKE, KAY P. YANEV, AND ROBERT W. HANSEN
Populations of robust salamanders belonging to the plethodontid salamander ge-nus Batrachoseps (subgenus Plethopsis) from the southern Sierra Nevada and adjacentregions represent a previously unknown species here described as Batrachoseps ro-bustus. The new species is robust, with a short trunk (17–18 trunk vertebrae) andwell-developed limbs. It differs from its close geographic neighbor, Batrachosepscampi, in lacking patches of dorsal silvery iridophores and in having (typically) alightly pigmented dorsal stripe, and from Batrachoseps wrighti in being more robust,having more trunk vertebrae, and in lacking conspicuous white spots ventrally. Thisspecies is widely distributed on the semiarid Kern Plateau of the southeastern SierraNevada and extends along the east slopes of the mountains into the lower OwensValley; it also is found to the south in the isolated Scodie Mountains. It occurs athigh elevations, from 1615–2800 m, in areas of low rainfall and high summer tem-peratures.
THE slender salamanders, Batrachoseps, rangefrom the Columbia River in northern
Oregon (458339N) to the vicinity of El Rosario,Baja California Norte (308009N). These sala-manders were considered to display mainly in-traspecific morphological variation, and formany years only two (Hendrickson, 1954) orthree (Stebbins, 1951) species were recognized.Brame and Murray (1968) and Yanev (1980)demonstrated that several to many additionalspecies should be recognized. Some of theserepresent subdivision of existing taxa based onnew information, mainly allozyme and mito-chondrial DNA (mtDNA) characters (e.g., Jock-usch et al., 1998, 2001), but new discoveries ofmorphologically distinct forms, such as Batra-choseps aridus, Batrachoseps campi, and Batracho-seps gabrieli, have also led to increases in thenumber of recognized species. Here we de-scribe a morphologically and genetically distinctspecies from the southern Sierra Nevada thatwas discovered many years ago but misidentifiedas Batrachoseps stebbinsi (Richman, 1973). Untilrecently, we were unsure of the geographic lim-its of the taxa represented, but now sufficientinformation is available to justify description ofa strikingly distinct and previously unknownspecies. The new species initially was thought tobe restricted to the Kern Plateau region of thesoutheastern Sierra Nevada, east of the KernRiver (as reported and illustrated in Stebbins,1985). Later, populations were found in the Sco-die Mountains, to the southeast, and in OwensValley. We now think that all of these popula-tions, which display substantial morphological(especially coloration) variation and range in el-evation from about 1600 to 2800 m, are conspe-
cific. They occur in relatively extreme environ-ments for terrestrial salamanders, in areas thatare exceptionally hot and dry in the summer,and that have a short, unpredictable wet seasonwith little rainfall. Some of them occur at highelevations that are under snow cover for severalmonths each year.
This new species is a member of the genusBatrachoseps because it has four toes on the hindlimb, a large cranial fontanelle, and an elongat-ed genioglossal muscle. Furthermore, it is amember of the subgenus Plethopsis and a relativeof both Batrachoseps (Plethopsis) wrighti and Ba-trachoseps (Plethopsis) campi on the basis of a phy-logenetic analysis of sequences of the mitochon-drial gene cytochrome b ( Jackman et al., 1997;Jockusch and Wake, 2002).
MATERIALS AND METHODS
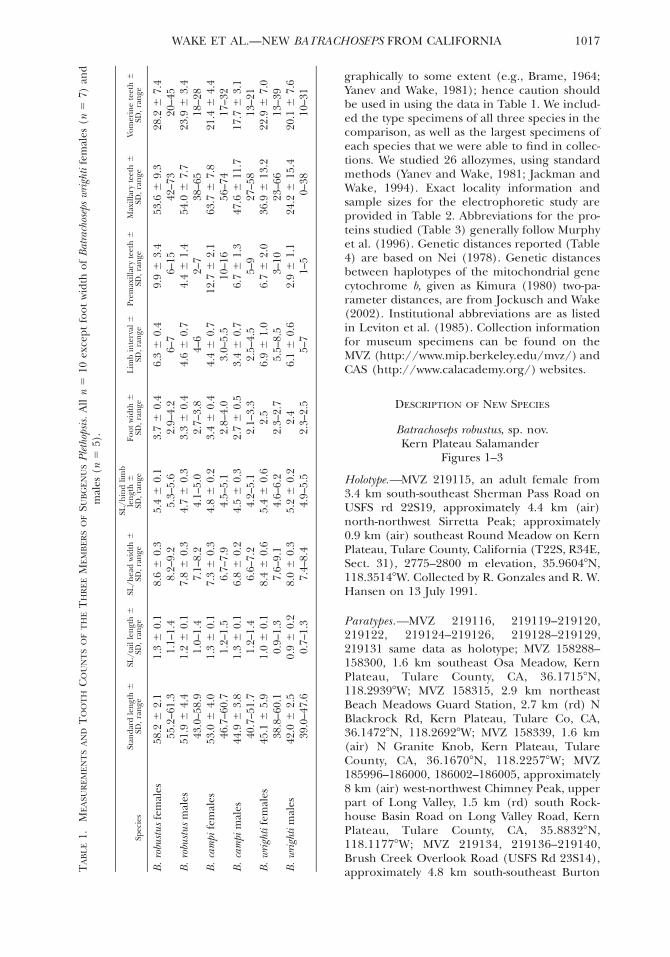
All specimens used in this study, with the ex-ception of a single specimen mentioned in thetext, were collected from the field for this study.Morphological data were taken as in Wake(1996). All measurements are in millimeters,only clearly visible teeth are counted, and stan-dard length (SL) is the distance from the tip ofthe snout to the posterior angle of the vent. Per-tinent morphological measurements are pre-sented in Table 1. We measured 10 adult malesand 10 adult females of the new species and ofits closest relatives, B. campi and B. wrighti. Sam-ple sizes for the new species and B. campi weretoo small for us to obtain 20 adults from a singlepopulation; thus, we were forced to combinesamples from two or more populations. Weknow that all of these species are variable geo-
1017WAKE ET AL.—NEW BATRACHOSEPS FROM CALIFORNIA
TA
BL
E1.
ME
ASU
RE
ME
NT
SA
ND
TO
OT
HC
OU
NT
SO
FT
HE
TH
RE
EM
EM
BE
RS
OF
SUB
GE
NU
SPl
etho
psis
.A
lln
510
exce
ptfo
otw
idth
ofBa
trac
hose
psw
righ
tife
mal
es(n
57)
and
mal
es(n
55)
.
Spec
ies
Stan
dard
leng
th6
SD,r
ange
SL/t
aill
engt
h6
SD,r
ange
SL/h
ead
wid
th6
SD,r
ange
SL/h
ind
limb
leng
th6
SD,r
ange
Foot
wid
th6
SD,r
ange
Lim
bin
terv
al6
SD,r
ange
Prem
axill
ary
teet
h6
SD,r
ange
Max
illar
yte
eth
6SD
,ran
geVo
mer
ine
teet
h6
SD,r
ange
B.ro
bust
usfe
mal
es58
.26
2.1
55.2
–61.
31.
36
0.1
1.1–
1.4
8.6
60.
38.
2–9.
25.
46
0.1
5.3–
5.6
3.7
60.
42.
9–4.
26.
36
0.4
6–7
9.9
63.
46–
1553
.66
9.3
42–7
328
.26
7.4
20–4
5B.
robu
stus
mal
es51
.96
4.4
43.0
–58.
91.
26
0.1
1.0–
1.4
7.8
60.
37.
1–8.
24.
76
0.3
4.1–
5.0
3.3
60.
42.
7–3.
84.
66
0.7
4–6
4.4
61.
42–
754
.06
7.7
38–6
523
.96
3.4
18–2
8B.
cam
pife
mal
es53
.06
4.0
46.7
–60.
71.
36
0.1
1.2–
1.5
7.3
60.
36.
7–7.
94.
86
0.2
4.5–
5.1
3.4
60.
42.
8–4.
04.
46
0.7
3.0–
5.5
12.7
62.
110
–16
63.7
67.
856
–74
21.4
64.
417
–32
B.ca
mpi
mal
es44
.96
3.8
40.7
–51.
71.
36
0.1
1.2–
1.4
6.8
60.
26.
6–7.
24.
56
0.3
4.2–
5.1
2.7
60.
52.
1–3.
33.
46
0.7
2.5–
4.5
6.7
61.
35–
947
.66
11.7
27–5
817
.76
3.1
13–2
1B.
wri
ghti
fem
ales
45.1
65.
938
.8–6
0.1
1.0
60.
10.
9–1.
38.
46
0.6
7.6–
9.1
5.4
60.
64.
6–6.
22.
52.
3–2.
76.
96
1.0
5.5–
8.5
6.7
62.
03–
1036
.96
13.2
23–6
622
.96
7.0
13–3
9B.
wri
ghti
mal
es42
.06
2.5
39.0
–47.
60.
96
0.2
0.7–
1.3
8.0
60.
37.
4–8.
45.
26
0.2
4.9–
5.5
2.4
2.3–
2.5
6.1
60.
65–
72.
96
1.1
1–5
24.2
615
.40–
3820
.16
7.6
10–3
1
graphically to some extent (e.g., Brame, 1964;Yanev and Wake, 1981); hence caution shouldbe used in using the data in Table 1. We includ-ed the type specimens of all three species in thecomparison, as well as the largest specimens ofeach species that we were able to find in collec-tions. We studied 26 allozymes, using standardmethods (Yanev and Wake, 1981; Jackman andWake, 1994). Exact locality information andsample sizes for the electrophoretic study areprovided in Table 2. Abbreviations for the pro-teins studied (Table 3) generally follow Murphyet al. (1996). Genetic distances reported (Table4) are based on Nei (1978). Genetic distancesbetween haplotypes of the mitochondrial genecytochrome b, given as Kimura (1980) two-pa-rameter distances, are from Jockusch and Wake(2002). Institutional abbreviations are as listedin Leviton et al. (1985). Collection informationfor museum specimens can be found on theMVZ (http://www.mip.berkeley.edu/mvz/) andCAS (http://www.calacademy.org/) websites.
DESCRIPTION OF NEW SPECIES
Batrachoseps robustus, sp. nov.Kern Plateau Salamander
Figures 1–3
Holotype.—MVZ 219115, an adult female from3.4 km south-southeast Sherman Pass Road onUSFS rd 22S19, approximately 4.4 km (air)north-northwest Sirretta Peak; approximately0.9 km (air) southeast Round Meadow on KernPlateau, Tulare County, California (T22S, R34E,Sect. 31), 2775–2800 m elevation, 35.96048N,118.35148W. Collected by R. Gonzales and R. W.Hansen on 13 July 1991.
Paratypes.—MVZ 219116, 219119–219120,219122, 219124–219126, 219128–219129,219131 same data as holotype; MVZ 158288–158300, 1.6 km southeast Osa Meadow, KernPlateau, Tulare County, CA, 36.17158N,118.29398W; MVZ 158315, 2.9 km northeastBeach Meadows Guard Station, 2.7 km (rd) NBlackrock Rd, Kern Plateau, Tulare Co, CA,36.14728N, 118.26928W; MVZ 158339, 1.6 km(air) N Granite Knob, Kern Plateau, TulareCounty, CA, 36.16708N, 118.22578W; MVZ185996–186000, 186002–186005, approximately8 km (air) west-northwest Chimney Peak, upperpart of Long Valley, 1.5 km (rd) south Rock-house Basin Road on Long Valley Road, KernPlateau, Tulare County, CA, 35.88328N,118.11778W; MVZ 219134, 219136–219140,Brush Creek Overlook Road (USFS Rd 23S14),approximately 4.8 km south-southeast Burton

1018 COPEIA, 2002, NO. 4
TABLE 2. SAMPLES USED FOR ALLOZYME STUDY.
Population Species n Locality MVZ#
1234567
B. robustusB. robustusB. robustusB. robustusB. robustusB. robustusB. robustus
810133672
Walker CreekHogback CreekOsa MeadowsBlackrock RoadKennedy MeadowBurton CampHorse Meadow
202331–36202362–71158288–300158315–17158318–21158307–13158338–40
89
1011
B. robustusB. robustusB. campiB. wrighti
101055
Long ValleyMcIver’s SpringFrench SpringHidden Lake
185996–6005158274–77, 79–83, 184900167183–87177846–50
Camp (northeast 1/4 Sect. 11, T23S, R33E),2375 m, Kern Plateau, Tulare County, CA,35.94678N, 118.38508W; LACM 99469–70, 4 kmE Beach Meadows Guard Station (36889300N,1188159300W; Richman, 1973), 2530 m, [KernPlateau], Tulare County, CA, 36.14178N,1188259830W; MVZ 202331–202336, WalkerCreek, 8.1 km soouth Olancha, Owens Valley,1740 m, Inyo County, CA, 36.23638N,118.06858W; MVZ 202362–202364, HogbackCreek, 11.3 km south-southwest Olancha(northwest 1/4 Sect 13, T20S, R36E), 1770 m,Inyo County, CA, 36.18998N, 118.03478W; MVZ158275–158277, 169168–169169, vicinity ofMcIvers (Melvers, on some maps) Spring, Sco-die Mtns, 1980–2010 m, Kern County, CA,35.62158N, 118.07248W; MVZ 158279–158281,3.2 km (air) W McIvers Spring, Scodie Mtns,2010 m, Kern County, CA, 35.62248N,118.10568W.
Referred material.—MVZ 158304, 158307, 158329,158331, 158338, 202365–202371, 219114,219117–18, 219127, 219130, 219132–219133,219135, 219141–43 219158, 219990, 220813,222939–222949, 222962, 224862–224863, and225713, CAS 213163, and CAS 213154. The fol-lowing cleared-and-stained skeletal preparationsare also assigned to the species: MVZ 158274,158301–158303, 158316–158317, 169171,172646.
Diagnosis.—A large, robust species of Batracho-seps (SL of adults averages 51.9 in males, 58.2 infemales, Table 1) with a short trunk (17–19trunk vertebrae), broad head, a relatively short,stout tail, well-developed limbs, and relativelylarge hands and feet with robust digits. Distin-guished from B. campi by its narrower head,shorter limbs, and distinctive coloration (oftenwith a dorsal stripe, lacking patches of silvery
iridophores but with a complicated dorsal mot-tling of rusty and gray); from B. wrighti by hav-ing more trunk vertebrae (mode of 18 ratherthan 17), a more robust habitus with largerhands and feet, and lacking prominent ventro-lateral white spots; from B. stebbinsi and B. sim-atus by having generally fewer trunk vertebraeand paired premaxillary bones; from all otherspecies of Batrachoseps by its combination oflarge size, robust habitus, relatively few trunkvertebrae, color pattern and paired premaxil-lary bones.
Description.—Batrachoseps robustus, one of thelargest species in the genus (Table 1), has abroad head that is well demarcated from theneck. The facial region of the head is relativelylarge, and the snout is broad and rather trun-cated. The head is relatively robust and deepand is not flattened. Nostrils are small, andthere are modest nasolabial protuberances as-sociated with the prominent nasolabial groove.Sexually mature males have more pronouncedswelling associated with the terminus of the na-solabial grooves that extend beyond the jawline. No mental glands are found in males.Grooving patterns of the head, throat and neckare typical of the genus. Eyes are relatively largebut are not prominent, nor do they protrudenoticeably from the side of the head; eyes areeither not visible, or barely visible, in ventralview. Vomerine teeth are in moderately long se-ries, or occasionally in patches. Small maxillaryteeth are borne in a series of moderate lengththat ends under the eye. Premaxillary teeth aresmall and more numerous in females than inmales. In some large males, the teeth are some-what enlarged and are located far forward,where they pierce the upper lip. Of all speci-mens of known sex that were radiographed orcleared and stained, all females (13) have 18

1019WAKE ET AL.—NEW BATRACHOSEPS FROM CALIFORNIA
Fig. 1. Holotype. Museum of Vertebrate Zoology (MVZ) 219115, an adult female from the Kern Plateau,Tulare County, California.
trunk vertebrae, but of the eight males studied,two have 19 trunk vertebrae, three 18, two 17,and one was asymmetric (recorded as 17 1/2).There are either 16 or 17 costal grooves be-tween the limbs (counting one each in the ax-illa and the groin). The tail is short and rela-tively stout, not tapering much until near thetip. The tail has no discernible basal constric-tion and is broader than deep basally, becominground posteriorly. No postiliac gland is evident.Limbs are relatively long and well developed.The hands and feet are relatively large and welldeveloped, with well-demarcated, stoutly round-ed digits and expanded digital tips that bearwell-developed subterminal pads. There arefour fingers and four toes. The third toe ismarkedly longer than the other digits. Webbingis insignificant. Fingers and toes in order of de-creasing length are 3-2-4-1.
Measurements of the holotype (in millimeters).—Max-imum head width 7.2 (7.8 in life); snout to gularfold (head length) 11.5; head depth at posteriorangle of jaw 3.6; eyelid length 3.2, eyelid width2.1; anterior rim of orbit to snout 2.5; horizon-tal orbital diameter 2.1; interorbital distance1.8; snout to forelimb 15.0; distance separatingexternal nares 2.1; snout projection beyondmandible 0.5; snout to posterior angle of vent(standard length) 53.7 (55.2 in life); snout toanterior angle of vent 50.1; axilla to groinlength 29.4; tail length 43.5 (45.5 in life); tailwidth at base 3.5; tail depth at base 3.1; forelimblength 10.5; hind-limb length 11.7; limb intervalfour costal interspaces; width of right hand 3.0;width of right foot 3.8; length of third toe 1.9;body width behind forelimbs 4.4. There are fivepremaxillary, 65 maxillary, and 20 vomerineteeth and 19 trunk, two caudosacral, and 30caudal vertebrae.
Coloration (in life).—Color was recorded in theanesthetized holotype by DBW (25 July 1991).‘‘Ground color dark brownish black. Dorsal sur-faces heavily covered with iridophores that giveappearance of a broad dorsal band with irreg-ular and indistinct borders from the eyes ontothe anterior part of the tail. Iridophoresgrouped together and overlain with second lev-el so that white has bronze- to reddish-brown ontop of it. Whole band has lively tan-rusty brownappearance, with many irregular gaps whereground color shows. Band becomes very brokenup on head, which is a complex mottling—verylively, attractive. Bronze color limited to dorsumwith flanks having a heavy guanophore washthat ends rather abruptly at ventrolateral mar-gin. Venter midline unmarked gray-black, butgular region heavily mottled with white iridop-hores. Dorsal surfaces of limbs heavily markedwith iridophores. Iris dark with bronze high-lights. Tail irregularly but heavily marked dor-sally with iridophores, including bronze color.Large recent scar on tail. Venter of tail un-marked gray-black.’’
Coloration of an anesthetized paratype (MVZ219140) was recorded the same day. ‘‘Like ho-lotype, but band on dorsum more distinct andbordered by a broken black dorsolateral lineproduced by gaps in the superficial pigment al-lowing the very dark black ground color to showthrough. Black particularly evident on costalgrooves. Band much less reddish than holotypeand is essentially tan to cream, with only a littlebronze. Some cream areas so concentrated thatband appears to be spotted or blotched. Sides,venter and limbs and iris like holotype.’’
Variation in coloration.—Batrachoseps robustus dis-plays considerable variation in coloration overits range, both within populations and amonggeographically separate populations. A general

1020 COPEIA, 2002, NO. 4
TA
BL
E3.
AL
LE
LE
FRE
QU
EN
CIE
SFO
R24
PRO
TE
INS
USE
DIN
AL
LO
ZYM
EST
UD
Y.Po
pula
tions
asin
Tabl
e2.
Inad
ditio
n,al
lpo
pula
tions
fixed
for
the
sam
eal
lozy
me
for
SOD
and
AD
A-2
.
Locu
s
Popu
latio
n
12
34
56
78
910
11
AC
OH
-1F
EE
EE
C0.
143
E0.
857
EE
EE
C
AC
OH
-2A
AA
AA
AA
AA
BE
PGD
HD
0.75
G0.
25G
D0.
192
G0.
269
I0.
462
J0.
077
G0.
667
I0.
333
D0.
667
F0.
083
G0.
167
H0.
083
G0.
786
I0.
214
D0.
25G
0.75
G0.
55I
0.45
DF
H
IDH
-1B
BA
0.07
7B
0.92
3B
BA
0.28
6B
0.71
4B
BA
0.3
B0.
7B
0.9
C0.
1B
IDH
-2A
AA
AA
AA
AA
CA
G6P
DH
B0.
875
C0.
125
A0.
05B
0.95
BB
BB
BB
BB
B
AA
T-1
G0.
187
J0.
813
G0.
9J
0.1
E0.
115
G0.
808
J0.
077
GE
0.41
7G
0.08
3J
0.5
G0.
5J
0.5
E0.
25J
0.75
GG
0.25
J0.
75F
I
AA
T-2
BB
BB
BB
BB
BA
BL
DH
-1A
AA
AA
AA
AA
AD
LD
H-2
DD
DD
DD
DD
DC
0.2
D0.
8D
PEP
B(I
gg)
EE
EE
EE
EB
0.15
E0.
85E
G0.
5H
0.5
E
CK
BB
A0.
038
B0.
962
BA
0.08
3B
0.91
7B
BB
BB
0.9
C0.
1B
G3P
DH
GG
GG
GG
GG
F0.
55G
0.45
ED
MPI
CC
CC
CC
CC
CC
0.9
E0.
1A
0.2
C0.
8PG
MC
B0.
6C
0.4
CC
C0.
833
D0.
167
CC
B0.
333
C0.
667
CC
C
MD
HP
CC
0.5
E0.
5A
0.15
4C
0.84
6C
CC
CC
B0.
15C
0.85
CC
PEP
D(I
-a)
A0.
063
E0.
938
A0.
05E
0.95
A0.
038
B0.
462
E0.
462
F0.
038
B0.
5E
0.5
E0.
917
F0.
083
EE
EB
0.05
E0.
95B
0.1
E0.
9B
0.6
E0.
4

1021WAKE ET AL.—NEW BATRACHOSEPS FROM CALIFORNIA
TA
BL
E3.
CO
NT
INU
ED
Locu
s
Popu
latio
n
12
34
56
78
910
11
AD
A-1
DD
DD
DD
DD
DE
CM
DH
-IB
BB
BB
A0.
143
B0.
857
BB
BD
B
MD
H-2
FF
D0.
192
F0.
808
FF
FF
C0.
1F
0.9
FE
B
GPI
BB
B0.
654
C0.
346
B0.
5C
0.5
B0.
917
D0.
083
BA
0.25
B0.
75A
0.35
B0.
45D
0.2
B0.
85D
0.18
BB
GL
UD
BB
BB
BB
BB
B0.
9C
0.1
BB
IDD
HE
EC
0.15
4E
0.84
6C
0.5
E0.
5E
EC
0.5
E0.
5E
E0.
75G
0.25
EE
AD
HA
AA
A0.
833
B0.
167
A0.
917
B0.
083
AA
AA
AA
trend, however, is that animals with light dorsalcoloration (e.g., gray or silver) are restricted tothe driest portions of the range, such as the Sco-die Mountains or the southeastern part of theKern Plateau. The light-colored dorsal surfacesof these animals may contribute to crypsis onpale-colored substrates. In contrast, salaman-ders occurring in the most mesic portions of therange, such as the Red Fir forests on the north-ern portion of the Kern Plateau, are invariablydark-colored. These salamanders closely ap-proximate the reds and browns of downed anddecaying Red Fir and Lodgepole Pine logs andassociated bark rubble. Color variation was re-corded at time of capture for samples from tworelatively mesic localities, as follows. (1) northof Poison Meadow, Kern Plateau, 11 July 1991:20 (SL of living animal), black with brownishiridophores; 20, brownish mottling; 29, 36, redplus brown; 31, few rust-red patches but distinctdorsal band present that overall is brownish-red;32, prominent rust-red patches; 36, reddish; 40,46, overall very reddish (‘‘red fir bark red’’); 42,silvery gray with faint rust blotching but littlerust on tail, dorsal ‘‘band’’ indistinct; 44, rustpatches all over w/black edging; 45, reddishwith black lateral edge line with distinct dorsalband; sides mottled with gray stippling; 50, red-dish overall, venter dark chocolate, light spot-ting ventrally, mostly in gular region; 54, olivegray with few coppery rust-brown patches, moreon tail, dorsal ‘‘band’’ not distinct. (2) BrushCreek Overlook, Kern Plateau, 12 July 1991: 18,18, black dorsally; 18, 28, fine rust dorsal fleck-ing, 19, dark with brownish-rust flecking; 42, sil-very gray dorsal color; 43, 46, reddish-rust dor-sal color.
Osteology.—Information concerning osteology isderived from nine cleared-and-stained speci-mens (MVZ 158301–158303, 158316–158317,158274, 169171, 172646, 184900) and from ra-diographs of many of the type series. The well-developed skull (Fig. 2) is similar to that of B.campi. Premaxillary bones of adults are paired,and prefrontal bones are typically present (ab-sent on one side in two individuals). In a juve-nile (MVZ 158317, 25.3 SL), there is still a sin-gle premaxillary, but tiny paired prefrontalshave already developed. The prefrontals varygreatly in size, from tiny spots of bone in some,to bones that are rather large and exceed theseptomaxillary in size in others. The bone isnever more than about one-quarter the area ofthe nasal, which is always large and well devel-oped. In two individuals, the nasal is divided onone side (Fig. 2). The nasals overlap the well-developed septomaxillaries and frontals and

1022 COPEIA, 2002, NO. 4
TABLE 4. NEI (1978) GENETIC DISTANCE FOR NINE POPULATIONS (SEE TABLE 1) OF Batrachoseps robustus AND ONE
POPULATION EACH OF Batrachoseps campi AND Batrachoseps wrighti.
Popula-tion 1 2 3 4 5 6 7 8 9 10 11
123456789
1011
00.1120.0910.110.0830.0610.0560.1030.0630.5480.402
00.0550.050.1020.0350.0560.030.1030.5420.459
00.0010.0650.0320.0430.0260.0670.5050.392
00.0570.0350.0350.0240.0910.540.423
00.0640.0090.090.0520.5320.447
00.0140.0290.0530.4790.394
00.0470.0410.4960.417
00.0870.5060.435
00.5020.435
00.675 0
Fig. 2. Dorsal (left) and ventral (right) views ofthe skull of an adult male Batrachoseps robustus (MVZ158303). Only bones and teeth are shown, apart fromthe dorsal fontanelle, which is stippled. The nasalbone on the right side is fragmented, and the rightvomer also has a small anterior fragment.
Fig. 3. Dorsal view of the right hind limb of anadult Batrachoseps robustus (MVZ 158303). Cartilage isstippled.
contact both the prefrontals and facial process-es of the maxillaries. The nasolacrimal ductpasses over the prefrontal, and it may be partlyincised into the anterodorsal surface of thatbone. The duct enters the nasal capsule at theanterior end of the prefrontal, between the na-sal and the maxillary. The maxillaries are longand slender and bear small teeth in a single se-ries that extends well posterior to the center ofthe eyeball. Each maxillary extends about two-thirds to three-quarters of the way through theeye, and the last 20% is typically edentulous andattenuate. There is a sizable dorsal fontanellelying between the frontals and the parietals, andpaired bones do not touch their counterparts.The parietals are well separated from each oth-er; they are moderately broad for this genus.The parietals narrowly fall short of the synotic
tectum. There are no crests on the otic capsules,but there is a small ridgelike ledge above thearticulation zone of the elongate and slenderdorsal portion of the squamosals. The quadrateis a relatively well-developed bone. The oper-culum bears a well-developed columellar rod.The vomers are well articulated to each other,and each bears a small to large preorbital pro-cess that varies among individuals; in some, it

1023WAKE ET AL.—NEW BATRACHOSEPS FROM CALIFORNIA
Fig. 4. Distribution of Batrachoseps robustus on theKern Plateau and western margins of the Owens Val-ley, Inyo County, CA, and in the Scodie Mountains,Kern County, CA.
reaches nearly to the lateral margin of the bodyof the vomer and is as well developed as seenin any members of the genus. The process sup-ports a row of vomerine teeth. Paired, relativelynarrow patches of teeth underlie the large par-asphenoid bone. Each patch contains fromabout 80 to 120 tiny teeth. The patches arewidely separated from each other. The lower jawis relatively stout and consists of the elongatedentaries and prearticulars, the latter having abroadly expanded, moderately high coronoidprocess.
The hyolingual skeleton is typical of the ge-nus in being entirely cartilaginous, lacking theurohyal, having elongated radii, and havinglong epibranchials that extend under the skinof the neck and shoulder region as far posteri-orly as the forelimb insertion.
There are 18 or 19 trunk vertebrae, the lastlacking ribs or having very short ribs. There arethree caudosacral vertebrae. Tails of most indi-viduals are regenerated, but there are 34 verte-brae in one apparently complete tail and 30 and31 in two others.
The limbs are long, and the tibial spur is welldeveloped, long and free. Phalangeal formulasare 1-2-3-2 for both hands and feet. The handsand feet are broad and have well-developed dig-its made up of relatively stout phalanges (Fig.3). The terminal phalanges are expanded andflattened, often with an irregular surface finish.The mesopodials have the standard arrange-ment for the genus (illustrated in Marlow et al.,1979). There are eight carpals and eight tarsals,but in one individual, there are only seven car-pals as a result of the fusion of the intermediumand centrale.
Habitat and distribution.—Batrachoseps robustus,now known from over 30 sites, occurs in a sur-prisingly broad range of habitats in a regionthat appears to be generally unfavorable for am-phibians. The elevational range is from 1615 min Portuguese Canyon on the east slope of theSierra Nevada to 2804 m at Sherman Pass onthe Kern Plateau (Fig. 4). In general, popula-tions are more numerous on the northern andwestern parts of the Kern Plateau, correspond-ing to more mesic fir and Jeffrey Pine forests,than in the more isolated and xeric regions tothe east and south. With decreasing elevationand precipitation, the occurrence of these sal-amanders is increasingly restricted to seasonalsprings and seeps in otherwise harsh, arid sur-roundings.
Descriptions of sites at which this species hasbeen collected are based on field notes of RWH,and Figure 5 illustrates habitat conditions.
(1) McIvers Spring, Scodie Mountains, KernCounty (elevation 2025 m; Fig. 5A), visited 12May 1979 and 17 May 1980. This is the south-eastern-most locality and appears to be margin-al for salamanders. The McIvers Spring drain-age is dry Singleleaf Pinyon Pine (Pinus mono-phylla) forest in an area of gentle relief—a pla-teau of sorts before dropping off into SageCanyon and down into the Mojave Desert. Thesoil is very porous and dry. A few Jeffrey Pines(P. jeffreyi) grow here, but this species is not ex-tensive, and the trees all appear to be fairly old.Precipitation is estimated to be less than 300mm per year, with most occurring as wintersnowfall. Outflow from McIvers Spring is pres-ent for several hundred meters before the sur-face dries. We found 36 salamanders along thecreek during two visits: 24 adults or subadults,and 12 juveniles. All but one of the salamanderswere found beneath rocks in sandy soil at theedge of the creek. The soil was moist for 1–2 mon either side of the creek; all salamanders werefound within this band of moisture. A singlelarge adult was found under a small log at thebase of a large Jeffrey Pine; the ground was cov-ered with pine needles and fallen bark. Othervegetation here includes Artemisia tridentata,Chrysothamnus nauseosus, Carex sp., Taraxacumsp., Eriogonum sp., Opuntia basilaris, Salix sp.(patchy), along with a few scattered Quercus wis-lizenii and Q. kelloggii. Rock outcrops borderingthe creek consist of weathered granite. The total

1024 COPEIA, 2002, NO. 4
Fig. 5. Habitat of Batrachoseps robustus. (A) McIvers Spring, Scodie Mountains, 2025 m, Kern County, CA.Salamanders are apparently restricted to the band of moisture associated with spring runoff, occurring beneathscattered rocks on moist soil. Bordering woodland consists principally of Singleleaf Pinyon Pine (Pinus mono-phylla). 12 May 1979. (B) Vicinity of Brush Creek Overlook, approximately 3 map miles south-southeast BurtonCamp, north of Poison Meadow, 2380 m elevation, Tulare County, CA. This site lies on the western edge ofthe Kern Plateau. Salamanders were found beneath flat rocks at the margins of a seepage. Bordering forest iscomprised of Jeffrey Pines (Pinus jeffreyi) and White Fir (Abies concolor). 10 July 1980. (C) Type locality for B.robustus, north-northwest Sirretta Peak, southeast Round Meadow on Kern Plateau, Tulare County, CA. Eleva-tion 2775–2800 m. 13 July 1991. (D) Sage Flat Creek, east slope Sierra Nevada, Inyo County, CA. This site istypical of the eastern slope localities, dominated by Singleleaf Pinyon and Canyon Oaks (Quercus chrysolepis),with Bigleaf Sagebrush (Artemisia tridentata) common in open areas. Salamanders were found only on or atthe base of the shaded, north-facing slope. The arid Owens Valley and the Inyo Mountains are visible in thedistance. 25 March 1986.

1025WAKE ET AL.—NEW BATRACHOSEPS FROM CALIFORNIA
corridor length over which salamanders werefound is no more than 200 m. Bufo boreas, Hylaregilla, Sceloporus graciosus, and Thamnophis cou-chii also were found here.
(2) Brush Creek Overlook, Kern Plateau, Tu-lare County (elevation 2377 m; Fig. 5B), visited10 July 1980 and 12 July 1991. Outflow from asmall spring courses over a metamorphic rockoutcropping with talus before entering a denseWhite Fir (Abies concolor) and Jeffrey Pine forest.Salamanders (n 5 21 in two visits) were foundbeneath mostly flat rocks (talus) bordering, butnot away from, the spring runoff. Borderingslopes are moderately steep (approximately358), with the stream flowing along a gradientof 10–358. There is little streamside vegetation,except for monkey flowers (Mimulus sp.) andcurrents (Ribes sp.).
(3) South of Sherman Pass, Kern Plateau, attype locality, Tulare County (elevation 2774–2804 m; Fig. 5C), visited 13 July 1991. Averageannual precipitation is estimated to be . 750mm, most as winter snowfall, with some summerthundershowers. The habitat here consists of alarge seep on a steep (approximately 308) west-facing slope. Surface flow is present in severalareas on the slope, as are scattered rocks anddowned logs. The surrounding area is heavilyforested and is dominated by Red Fir (Abies mag-nifica) and Lodgepole Pine (Pinus contorta). Veg-etation within the seep area consists of Veratrumcalifornicum and dwarf willows (Salix sp.; plantscharacteristic of wet meadows), and Dodecatheonsp. Salamanders (19 adults, 1 subadult) werefound both singly and in small groups, underrocks or under or within wet logs. There waslittle canopy cover at most of the collectionsites.
(4) Sage Flat Creek, east slope Sierra Nevada,Inyo County (elevation 1800–1950 m; Fig. 5D),visited 25 March 1986, 15 June 1991, and 6 April1993, is an example of a relatively open site onthe east slope of the Sierra Nevada. It is domi-nated by Singleleaf Pinyon Pine and CanyonOaks (Quercus chrysolepis), with Bigleaf Sage-brush common in open areas. Salamanderswere found under single rocks, within rock rub-ble and crevices, and under wet logs, exclusivelyon or at the base of a moderately steep north-facing slope.
Based upon our now considerable field ex-perience with B. robustus (. 350 specimens),several patterns emerge concerning its generalecology. Timing and duration of seasonal activ-ity vary according to elevation and available sur-face moisture. Lower elevation populations (,2000 m) are probably restricted to a few monthsof activity in late winter and spring, before sur-
face conditions become warm and dry. Popula-tions occurring at higher elevations are limitedin their activity by cold temperatures and snow.We suspect that at higher elevations surface ac-tivity is confined to a few months, perhaps Mayor June to October. Thus, the timing of court-ship and egg laying is expected to vary accord-ing to local conditions.
Substrate temperatures were recorded for 217salamanders at point of discovery (under cover)from a Schultheis or Miller and Weber quick-reading thermometer. The mean substrate tem-perature was 13.5 C, with a range of 5.2–25.0 C.This is well within the range of temperature val-ues for other species of Batrachoseps (Feder etal. 1982; RWH, unpubl.).
Near the end of the seasonal activity periodat some sites, especially those at lower elevationsand xeric environments, we have found only ju-venile salamanders (e.g., hatchlings or year-lings). These same sites have produced adults atother times. Although risk of desiccation be-cause of unfavorable surface area-volume rela-tionships is greater for juveniles, we attributelate-season juvenile activity under marginal sur-face moisture conditions to the need to accu-mulate fat reserves prior to summer aestivation.Adults, by contrast, do not face the same pres-sures and, thus, are not required to maintainsurface activity during suboptimal periods. Allmembers of Batrachoseps are relatively elongateand suited for subterranean existence. However,B. robustus has a relatively robust body and ashort tail, features that should result in a sur-face:volume relationship that is beneficial in re-sisting water loss.
Behavior and life history.—We recorded behavior-al responses for 79 individuals as they were dis-covered under surface cover in the field. Themost commonly observed behavior was an at-tempt to crawl away (n 5 58; 73.4%), followedby immobility (16.5%), thrashing (5.1%), andcoiling (3.8%). When handled, a few individualsreleased a sticky skin secretion. A single sala-mander exhibited a rapid coiling and springingresponse (‘‘watch-spring’’ behavior), a traitcommonly observed in some attenuate speciesof Batrachoseps. Coiling as a defensive responseis widespread in Batrachoseps; frequency of de-ployment varies among species. The apparentrarity of coiling behavior in B. robustus is nota-ble.
We have found recently hatched young mostcommonly during May and June (with some inMarch, April, and July), but this likely reflectsour field activity schedules and ability to gainaccess to high-elevation sites following snow

1026 COPEIA, 2002, NO. 4
melt. Hatchlings tend to aggregate in groups of2–5 and occur under smaller pieces of coverthan are used by subadults and adults.
On 3 May 1993, five gravid females were col-lected on the Kern Plateau (Long Valley Roadnear Rockhouse Basin Road). Given the ex-tremely dry conditions, we were surprised tofind any salamanders. None responded imme-diately following hormone injection ( Jockusch,1996). This suggests that these animals were notphysiologically ready for oviposition. Three ofthese individuals responded by laying eggs fol-lowing a second injection about a month later;each laid three eggs while retaining 1–3 eggs.Apparently clutch size is small and egg andhatchling size relatively large. Perhaps oviposi-tion occurs in connection with summer rains,which normally occur in the high mountains inthis area but are unpredictable. Two eggshatched after 96 and 103 days of developmentat 13 C. This is substantially slower developmentthan in members of the subgenus Batrachoseps(range from 65–85 days, with variation bothacross populations and species), but faster thanthe development of B. wrighti (average of 126days) at the same temperature (E. Jockusch,pers. comm.). The smallest female observed tobe gravid (MVZ 219143) is 44 SL.
Etymology.—The species name is derived fromthe Latin word robustus, with reference to its sizeand robust habitus.
DISCUSSION
A single specimen of this species, identifiedas B. stebbinsi, was reported nearly three decadesago (Richman, 1973). This misidentification oc-curred because external dimensions of B. robus-tus and B. stebbinsi are very similar (compare ourTable 1 with Brame and Murray, 1968). We firstrealized that the two were not conspecific whenwe observed plesiomorphic osteology in a singlecleared-and-stained specimen from the KernPlateau. Subsequent biochemical studies ( Jack-man et al., 1997; Jockusch and Wake, 2002; un-publ. allozyme data) demonstrated the distinc-tiveness of the two species, which belong to twodifferent clades. Once we understood that B. ro-bustus is a member of the clade now recognizedas the subgenus Plethopsis, we focused attentionon differentiation within that group, usingstarch-gel electrophoresis to survey allozymes innine populations of B. robustus and one each ofB. campi and B. wrighti (Table 2).
Substantial allozymic differentiation is foundwithin B. robustus but less than that encoun-tered in B. campi (Yanev and Wake, 1981). Fif-
teen of the 26 proteins studied were polymor-phic. Direct count heterozygosity ranges from0.038 (Hogback; n 5 10) to 0.109 (Osa Mead-ow; n 5 13). Genetic distances (D; Nei, 1972)range from 0.016 to 0.124 (mean 0.066). TheScodie Mountains are geographically isolatedfrom the rest of the range. However, the rangeof genetic distances between the populationfrom this region and those in the remainderof the range is small (0.055–0.108, mean0.077), suggesting that isolation is relatively re-cent. The two populations sampled from theOwens Valley (Walker Creek and HogbackCreek) are well differentiated from each other(0.115), with each being most similar to theKern Plateau population located most closelyto them to the west (Walker Creek to BurtonCamp, 0.065; Hogback Creek to Burton Camp,0.039, and to Chimney Peak 0.034). The ninepopulations are well differentiated from bothB. wrighti and B. campi but are more similar tothe former. The value of D 5 0.397–0.463(mean 0.428) to B. wrighti and 0.485–0.556(mean 0.525) to B. campi; between B. wrighti toB. campi D 5 0.681. There are seven fixed dif-ferences between B. robustus and B. wrighti, andanother locus is nearly fixed; eight fixed dif-ferences with another nearly fixed differencedistinguish B. robustus from B. campi; B. wrightiand B. campi differ by 12 fixed differences. Ge-netic distances to members of the subgenus Ba-trachoseps are all in excess of D 5 1.0, withmore than 10 fixed differences (unpubl. data).
The mitochondrial DNA gene cytochrome bof Batrachoseps was studied by Jackman et al.(1997), who sequenced 555 base pairs, and byJockusch and Wake (2002), who studied 278 in-dividuals and sequences up to 784 bp in length.Samples of three populations of B. robustus wereincluded: Scodie Mountains, Kern County, Ni-nemile Canyon, Inyo County, and topotypic ma-terial. Maximum sequence divergence was 3.7%(between Scodie Mountains and the type local-ity), compared with 1.8% within B. wrighti and4.2% within B. campi. Maximum parsimonyanalysis with all characters and types of changeweighted equally suggests a sister-group rela-tionship between B. robustus and B. wrighti(bootstrap support: 65%), but other methodsare unable to distinguish between B. campi andB. wrighti as possible sister taxa ( Jockusch andWake, 2002).
The subgenus Batrachoseps has three osteolog-ical synapomorphies: fused premaxillary bonesthroughout life, prefrontal bones absent, andno preorbital process of the vomer. The newspecies is plesiomorphic with respect to all threecharacters, as are B. campi and B. wrighti. There

1027WAKE ET AL.—NEW BATRACHOSEPS FROM CALIFORNIA
are no osteological synapomorphies of Plethop-sis, but DNA sequences and allozymes supportits status as a clade.
Jockusch and Wake (2002) concluded thatthe subgenus Plethopsis contains three distinctiveand well-differentiated species whose phyloge-netic relationships are unresolved. The diversi-fication of the group is relatively old, and miss-ing taxa would likely have a major influence onresolution of relationships if they were knownand available. Despite the close proximity of B.campi and B. robustus (43 km), these species arediverged from each other, with respect both toallozymes and mtDNA, to roughly similar de-grees as they are diverged from the geographi-cally remote (nearly 900 km) and ecologicallyvery different (mesic, heavily forested habitats)B. wrighti. The three species of Plethopsis are allperipheral in distribution to the more speciesrich subgenus Batrachoseps, which ranges fromnorthwestern Baja California to southwesternOregon, south and west of the range of Pleth-opsis.
Some species of the subgenus Batrachosepscome close to the range of B. robustus, but weknow of no sympatry with any other salamanderwith the exception of a small zone of overlapwith a population of Hydromantes sp. on the westside of the Owens Valley. A population of B. re-lictus occurs on the western margin of the KernPlateau, 1.6 km from the nearest B. robustus andwell within the elevational range of the latter,and several species occur in Kern Canyon, theGreenhorn Mountains, and the southern PiuteMountains (this is the closest approach of B.stebbinsi).
ACKNOWLEDGMENTS
Many persons have assisted in the collectionof salamanders, but in particular we wish tothank R. Gonzales, D. Guiliani, D. Holland, E.Jockusch, T. Papenfuss, and B. Macey. We es-pecially thank M. Frelow for able assistance inthe electrophoretic studies. Fieldwork was sup-ported by the California Department of Fishand Game and the U.S. Forest Service (SequoiaNational Forest) under contracts to RWH. Lab-oratory investigations and some fieldwork weresupported by grants from the National ScienceFoundation (DBW, Principal Investigator). Illus-trations were prepared by K. Klitz. We thank E.Jockusch for information and discussion, and E.Jockusch and R. Mueller for comments on themanuscript.
LITERATURE CITED
BRAME JR., A. H. 1964. Distribution of the Oregonslender salamander, Batrachoseps wrighti (Bishop).Bull. So. Calif. Acad. Sci. 63:165–170.
———, AND K. F. MURRAY. 1968. Three new slendersalamanders (Batrachoseps) with a discussion of re-lationships and speciation within the genus. Sci.Bull. Mus. Nat. Hist. Los Angeles Co. 4:1–35.
FEDER, M. E., J. F. LYNCH, H. B. SHAFFER, AND D. B.WAKE. 1982. Field body temperatures of tropicaland temperate zone salamanders. Smiths. Herpe-tol. Inf. Serv. 52:1–23.
HENDRICKSON, J. R. 1954. Ecology and systematics ofsalamanders of the genus Batrachoseps. Univ. Calif.Publ. Zool. 54:1–46.
JACKMAN, T. R., AND D. B. WAKE. 1994. Evolutionaryand historical analysis of protein variation in theblotched forms of salamanders of the Ensatina com-plex (Amphibia: Plethodontidae). Evolution 48:876–897.
———, G. APPLEBAUM, AND D. B. WAKE. 1997. Phylo-genetic relationships of bolitoglossine salamanders:a demonstration of the effects of combining mor-phological and molecular data sets. Mol. Biol. Evol.14:883–891.
JOCKUSCH, E. L. 1996. Techniques for obtaining andraising plethodontid salamander eggs. Int. J. Devel.Biol. 40:911–912.
———, AND D. B. WAKE. 2002. Falling apart andmerging: diversification of Slender Salamanders(Plethodontidae: Batrachoseps) in the AmericanWest. Biol. J. Linn. Soc. 76:361–391.
———, ———, AND K. P. YANEV. 1998. New speciesof slender salamanders, Batrachoseps (Amphibia:Plethodontidae), from the Sierra Nevada of Cali-fornia. Contr. Sci. Nat. Hist. Mus. Los Angeles Co.472:1–17.
———, K. P. YANEV, AND D. B. WAKE. 2001. Molecularphylogenetic analysis of slender salamanders, genusBatrachoseps (Amphibia: Plethodontidae), from cen-tral coastal California with descriptions of four newspecies. Herpetol. Monogr. 15:54–99.
KIMURA, M. 1980. A simple method for estimating evo-lutionary rate of base substitutions through com-parative studies of nucleotide sequences. J. Mol.Evol. 2:87–90.
LEVITON, A. E., R. H. GIBBS JR., E. HEAL, AND C. E.DAWSON. 1985. Standards in herpetology and ich-thyology. Part I. Standard symbolic codes for insti-tutional resource collections in herpetology andichthyology. Copeia 1985:802–832.
MARLOW, W. R., J. M. BRODE, AND D. B. WAKE. 1979.A new salamander, genus Batrachoseps, from theInyo Mountains of California, with a discussion ofrelationships in the genus. Contr. Sci. 308:1–17.
MURPHY, R. W., J. W. SITES JR., D. G. BUTH, AND C. H.HAUFLER. 1996. Proteins: isozyme electrophoresis,p. 51–120 In: Molecular systematics. 2d ed. D. M.Hillis, C. Moritz, and B. K. Mable (eds.). SinauerAssociates, Sunderland, MA.
NEI, M. 1972. Genetic distance estimates betweenpopulations. Am. Nat. 106:283–292.
———. 1978. Genetic variation in natural popula-

1028 COPEIA, 2002, NO. 4
tions: patterns and theory. Theoret. Popul. Biol. 13:121–177.
RICHMAN, J. B. 1973. A range extension for the Te-hachapi Slender Salamander, Batrachoceps[sic] steb-binsi. HISS News-J. 1:97.
STEBBINS, R. C. 1951. Amphibians of western NorthAmerica. Univ. California Press, Berkeley.
———. 1985. A field guide to western reptiles andamphibians. 2d ed. rev. Houghton Mifflin, Boston,MA.
WAKE, D. B. 1996. A new species of Batrachoseps (Am-phibia: Plethodontidae) from the San GabrielMountains, southern California. Contr. Sci. 473:1–12.
YANEV, K. P. 1980. Biogeography and distribution ofthree parapatric salamander species in coastal andborderland California, p. 531–550. In: The Califor-
nia islands: proceedings of a multidisciplinary sym-posium. D. M. Power (ed.). Santa Barbara Museumof Natural History, Santa Barbara, CA.
———, AND D. B. WAKE. 1981. Genic differentiationin a relict desert salamander, Batrachoseps campi.Herpetologica 37:16–28.
(DBW, KPY) MUSEUM OF VERTEBRATE ZOOLO-GY, UNIVERSITY OF CALIFORNIA, 3101 VALLEY
LIFE SCIENCES BUILDING, BERKELEY, CALIFOR-NIA 94720–3160; AND (RWH) 16333 DEER
PATH LANE, CLOVIS, CALIFORNIA 93611–9735. E-mail: (DBW) [email protected]. Send reprint requests toDBW. Submitted: 26 Nov. 2001. Accepted: 8July 2002. Section editor: C. Guyer.