nonribosomal peptide synthesis and toxigenicity of...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY,0021-9193/99/$04.0010
July 1999, p. 4089–4097 Vol. 181, No. 13
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Nonribosomal Peptide Synthesis and Toxigenicityof Cyanobacteria
BRETT A. NEILAN,1,2* ELKE DITTMANN,2 LEO ROUHIAINEN,3 R. AMANDA BASS,1
VERENA SCHAUB,2 KAARINA SIVONEN,3 AND THOMAS BORNER2
School of Microbiology and Immunology, The University of New South Wales, Sydney 2052, New South Wales,Australia1; Institute for Biology (Genetics), Humboldt University of Berlin, D-10115 Berlin, Germany2; andDepartment of Applied Chemistry and Microbiology, University of Helsinki, FIN-00014 Helsinki, Finland3
Received 14 December 1998/Accepted 21 April 1999
Nonribosomal peptide synthesis is achieved in prokaryotes and lower eukaryotes by the thiotemplatefunction of large, modular enzyme complexes known collectively as peptide synthetases. These and other multi-functional enzyme complexes, such as polyketide synthases, are of interest due to their use in unnatural-prod-uct or combinatorial biosynthesis (R. McDaniel, S. Ebert-Khosla, D. A. Hopwood, and C. Khosla, Science 262:1546–1557, 1993; T. Stachelhaus, A. Schneider, and M. A. Marahiel, Science 269:69–72, 1995). Most nonribo-somal peptides from microorganisms are classified as secondary metabolites; that is, they rarely have a rolein primary metabolism, growth, or reproduction but have evolved to somehow benefit the producing organisms.Cyanobacteria produce a myriad array of secondary metabolites, including alkaloids, polyketides, and non-ribosomal peptides, some of which are potent toxins. This paper addresses the molecular genetic basis of non-ribosomal peptide synthesis in diverse species of cyanobacteria. Amplification of peptide synthetase genes wasachieved by use of degenerate primers directed to conserved functional motifs of these modular enzyme com-plexes. Specific detection of the gene cluster encoding the biosynthetic pathway of the cyanobacterial toxinmicrocystin was shown for both cultured and uncultured samples. Blot hybridizations, DNA amplifications,sequencing, and evolutionary analysis revealed a broad distribution of peptide synthetase gene orthologues incyanobacteria. The results demonstrate a molecular approach to assessing preexpression microbial functionaldiversity in uncultured cyanobacteria. The nonribosomal peptide biosynthetic pathways detected may lead tothe discovery and engineering of novel antibiotics, immunosuppressants, or antiviral agents.
Not all proteins are synthesized on the ribosome. Smallpolypeptides, with fewer than about 50 amino acids, can beassembled by peptide synthetases just as other compounds,such as fatty acids, are linked by other synthases. The productsof microbial nonribosomal peptide synthesis include the im-munosuppressant cyclosporine and antibiotics such as grami-cidin S, tyrocidin A, and surfactins (for a review, see reference13). A modular structure of peptide synthetases has beenshown to be responsible for the sequential and amino-acid-specific elongation of peptide chains (17). The specific combi-nation of modules and various functional domains within thepeptide synthetase determines the structure and hence theactivity of the peptide product.
The Cyanobacteria, as determined on the basis of severalmolecular phylogenies, comprise a single and coherent groupof prokaryotes (35). Commonly, these bacteria proliferate ineutrophic marine and freshwater habitats, resulting in the for-mation of water blooms. Cyanobacteria represent a relativelyunexplored and potentially rich source of bioactive secondarymetabolites (6, 20, 21). Of these bioactive compounds, the toxinsproduced by certain planktonic species of cyanobacteria havebeen particularly well studied. These oxygenic phototrophs arethe only known producers of the hepatotoxic microcystins, andseveral morphologically and physiologically diverse generahave been shown to synthesize these compounds (11, 14, 30).
Microcystins are cyclic heptapeptides (Fig. 1A). Sixty-fiveisoforms of these compounds which vary by degree of methyl-
ation, hydroxylation, epimerization, peptide sequence, and tox-icity have been identified (26, 29). These peptides are potentinhibitors of eukaryotic protein phosphatases type 1 and 2A,with inhibition being dependent on particular structural vari-ations (1), including the substitution of two variable L-aminoacids and the methylation of aspartate (b-iso-Asp) and dehy-droalanine (Fig. 1A). The modified b-amino acid (Adda in Fig.1A), which is also found in the hepatotoxic pentapeptidesnodularin and motuporin, is conserved in all known toxic mi-crocystins. Microcystins and related cyclic peptides are carriedinto hepatocytes via the bile acid transport system, where hy-perphosphorylation of microfilaments, including cytokeratins,is the primary toxic effect (33). Microcystins may also activatephospholipase A2 and cyclooxygenase in hepatocytes, while inmacrophages they induce tumor necrosis factor alpha and in-terleukin 1. These functions, together with hyperphosphoryla-tion of DNA, have implicated microcystins as agents promot-ing hepatocellular carcinoma (8).
Like other small peptides which contain unusual amino ac-ids, microcystins are synthesized nonribosomally (2, 4). Wehave recently identified peptide synthetase genes in cyanobac-teria of the genera Microcystis and Anabaena (3, 19). Thecharacteristic modular structure of the peptide synthetasegenes and particular conserved sequence motifs seen in otherbacteria and fungi were also found in these cyanobacterialgenes (Fig. 1B) (3). The insertional inactivation of a peptidesynthetase gene from the hepatotoxic strain Microcystis aerugi-nosa PCC7806 resulted in transformation to the nontoxic stateand a loss of microcystins, demonstrating that this gene, calledmcyB, encodes a microcystin synthetase (4).
To date, peptide synthetase genes have been isolated fromtwo cyanobacterial species, while a microcystin synthetase gene
* Corresponding author. Mailing address: School of Microbiologyand Immunology, The University of New South Wales, Sydney 2052,NSW, Australia. Phone: 612 9385 3235. Fax: 612 9385 1591. E-mail: [email protected].
4089
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

has been isolated from only one of several microcystin-produc-ing genera. The present study describes the detection andcharacterization of microcystin and peptide synthetase genes ina genetically diverse range of microcystin-producing and non-toxic cyanobacterial species. Oligonucleotide primers whichamplify DNA from both the conserved sequence motifs ofpeptide synthetase genes and specific sequences of microcystinsynthetase genes in cyanobacteria were designed (Table 1).These PCR products were used as hybridization probes and/ordirectly sequenced. Correlations were made between the pres-
ence of peptide synthetase genes and the production ofmicrocystins by hepatotoxic cyanobacteria. The data present-ed provide initial indications of the distribution of microcystinsynthetase and other peptide synthetase genes in the phylumCyanobacteria and the possible mechanism underlying theirtransmission. In addition, these gene-targeting procedures en-able the isolation and characterization of sequences from novelpeptide synthetase modules with potentially diverse biosynthe-tic activities. Isolation and culturing of the many microorganismswhich may produce nonribosomal peptides are not required.
FIG. 1. (A) General structure of microcystins, cyanobacterial heptapeptide hepatotoxins, showing the most frequently found variations. X and Z, variable L-aminoacids L-leucine and L-arginine, respectively; R1 and R2, H (demethylmicrocystins) and CH3, respectively; D-MeAsp, D-erythro-b-methylaspartic acid; Adda,(2S,3S,8S,9S)-3-amino-9-methoxy-2,6,8-trimethyl-10-phenyldeca-4,6-dienoic acid; Mdha, N-methyldehydroalanine (Dha, dehydroalanine). (B) Diagram depicting thegene encoding a single peptide synthetase module. The functional domains of a type I peptide synthetase module include condensation (shaded box), amino-acid-specific acyladenylation (open box), thioesterification (hatching), and possibly epimerization (cross-hatching). A type II module would contain an N-methyltransferasedomain between the acyladenylation and thioesterification regions. Peptide synthase conserved motifs (13) are shown as roman numerals (I to VI) (for a more detaileddescription of functional domains in peptide synthetases, see reference 17). Arrows indicate the relative positions of degenerate cyanobacterial PS-PCR primers (atmotifs I and V; MTR2) and MS-PCR primers.
TABLE 1. Peptide synthetase gene consensus and specific PCR primers
Primer (annealing)temp Degeneracy (fold) Tm (°C) Product (bp) Oligonucleotide primer sequence, from 59339 (conserved peptide motif)
PS-PCR (52°C)MTF2 512 53 ;1,000 GCNGG(C/T)GG(C/T)GCNTA(C/T)GTNCC(AGGAYVP, core motif I)MTR 192 51 ;1,000 CCNCG(AGT)AT(TC)TTNAC(T/C)TG(QVKIRG, core motif V)
MS-PCR (50°C)FAA 1 50 758 CTATGTTATTTATACATCAGGRAA 1 50 758 CTCAGCTTAACTTGATTATC
4090 NEILAN ET AL. J. BACTERIOL.
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

MATERIALS AND METHODS
Cyanobacterial strains and culturing. Cyanobacterial strains with the desig-nations AWT, HUB, NIES, PCC, and UNSW were obtained from the culturecollections of Australian Water Technologies (AWT; Sydney, Australia), Hum-boldt University (HUB; Berlin, Germany), the National Institute for Environ-mental Studies (NIES; Tsukuba, Japan), the Institut Pasteur (PCC; Paris,France), and The University of New South Wales (UNSW; Sydney, Australia),respectively (Table 2). The remaining strains were from a culture collection ofone of the authors (K. Sivonen, University of Helsinki). Strains were grown inJM, BG-11, or Z8 (Helsinki strains only) medium at 20 6 2°C and undercontinuous illumination of 25 mmol/m2/s as detailed earlier (16, 25). Nitrogen-fixing species were grown in nitrogen-free medium (25), and Nodularia specieswere grown in salt-containing medium (16). Strains obtained from culture col-lections were axenic.
Measurement of strain toxicity and chemical analyses of microcystins. De-tection of microcystins by mouse bioassay, high-pressure liquid chromatography(HPLC), commercially available enzyme-linked immunosorbent assay (ELISA)(EnviroGard Microcystins Plate Kit; Strategic Diagnostics), and protein phos-phatase inhibition was performed according to previously published procedures.Briefly, the mouse bioassay was performed by intraperitoneal injection of aque-ous cell suspensions, measurement of the 50% lethal dose, and histologicalobservation of hepatic hemorrhage (5). For HPLC analyses, microcystin wasextracted from a log-phase culture with methanol. Unquantified measurement ofmicrocystin content was performed with a LiChroCART RP18 column (15).Protein phosphatase type 2A inhibition activity was determined with frozen-thawed cell extracts and standardized for total protein and dry cell mass (1, 4).For many of the strains listed in Table 2, at least one microcystin has beenisolated and the structure has been identified (26, 29).
DNA extraction, amplification, sequencing, and probe hybridization. Totalgenomic DNA was extracted by standard methods commonly used for cyanobac-teria and plants (10, 22). Alternatively, a PCR template was prepared by rapidcell lysis in the presence of a cation-exchange resin and nonionic detergents (24).PCR annealing step temperatures are shown in Table 1 along with peptidesynthetase gene-directed oligonucleotide primer sequences. In capillary or200-ml tubes, the PCR thermal cycling protocol included an initial denaturationat 94°C for 2 min, followed by 35 cycles at 93°C for 10 s, at the annealingtemperature (Table 1) for 20 s, and at 72°C for 1 min. Amplification reactioncomponents were as previously described (23), and incubations were performedwith an FTS-1S capillary thermocycler (Corbett Research, Sydney, Australia) ora PE2400 apparatus (Perkin-Elmer Cetus Corporation, Emeryville, Calif.).
Amplified DNA was purified from surplus reaction components and se-quenced directly by standard automated fluorescence techniques (19, 23).Larger, contiguous fragments of the M. aeruginosa PCC7806 and M. aeruginosaHUB524 microcystin synthetase genes (accession no. U97078 and Z28338, re-spectively) were also isolated from phage and plasmid gene libraries by PCRprobe hybridization. A method of semidegenerate, long PCR was developed toextend sequence information flanking that obtained from the gene libraries. Thisprocedure used the highly redundant primers MTF2 and MTR (Table 1) di-rected to the conserved motifs of known peptide synthetase genes combined withprimers specific for the Microcystis genome (19). By modifying and using long-PCR protocols, we found it possible to amplify regions of DNA encoding entiremodules of the synthetase. In this way, sequences flanking the known microcystinsynthetase gene could be extended to unknown conserved regions of the peptidesynthetase gene. This method is termed module jumping. Sequencing of thesePCR fragments was performed with 100 pmol of the degenerate primer andautomated protocols. DNA sequences of the peptide and microcystin synthetasegenes were aligned with the program Pileup (Genetics Computer Group) and themultiple-sequence-alignment tool Clustal W. Manual confirmation of the se-quence alignment was also performed. Phylogenetic analyses are described in thecorresponding figure legends.
Cyanobacterial genomic DNA (about 100 ng) was dot blotted onto nylonmembranes (Hybond N1; Amersham, Little Chalfont, United Kingdom) by apreviously described protocol (27). A 2.0-kb clone and a 2.4-kb clone corre-sponding to different modules of a peptide synthetase gene from Anabaena sp.strain 90, as well as a 1.15-kb amplified clone of the mcyB gene of strain HUB524(4), were used as probes. Probe labeling, DNA hybridization, which was per-formed at 60°C, and membrane washing were done by standard procedures (27).
RESULTS AND DISCUSSION
Detection of peptide synthetase genes by dot blot hybridiza-tion. In order to compare the distributions of two peptidesynthetase genes derived from M. aeruginosa HUB524 (mcyB)(4) and Anabaena sp. strain 90, DNAs from hepatotoxic, neu-rotoxic, and nontoxic strains belonging to various species ofAnabaena, Nostoc, Microcystis, Nodularia, Oscillatoria, andAphanizomenon were simultaneously probed in dot blot exper-iments (Fig. 2A). The mcyB fragment recently identified as a
microcystin synthetase gene was shown to hybridize, with vari-able signal intensity, to DNAs from all hepatotoxic strains andto DNAs from three nontoxic strains (Microcystis sp. strain 130,Aphanizomenon sp. strain 202, and Oscillatoria sp. strain 2)(Table 2 and Fig. 2B). This probe did not hybridize to DNAsfrom neurotoxic strains and other nontoxic strains (Table 2).The two Anabaena sp. strain 90 peptide synthetase gene frag-ments gave strong signals with all hepatotoxic Anabaena strainsbut showed insignificant hybridization to most other microcys-tin-producing species (Fig. 2C). These probes did, however,cross-hybridize to Nostoc sp. strain 152, Nodularia sp. strainHEM, nontoxic Anabaena sp. strain 299 (data not shown), and(weakly) Nodularia sp. strain NSOR12. Therefore, we haveevidence for two distinct peptide synthetase genes showingdifferent distributions among the cyanobacterial strains tested.These results reflect the observation that nonribosomal pep-tide and microcystin contents vary among cyanobacterial spe-cies (26). The mcyB hybridization data revealed the presenceof similar genes across all genera investigated. It remains to bedetermined whether all positive hybridization signals corre-sponded to microcystin synthetase genes. To obtain more de-tailed information on the distribution of peptide synthetasegenes among toxic and bloom-forming cyanobacteria, severalselected strains were further investigated by DNA amplifica-tion and sequencing.
Design of amplification primers for peptide synthetasegenes in cyanobacteria. A comparison of peptide synthetasesand other adenylate-forming enzymes from various pro-karyotes and eukaryotes revealed the presence of highly con-served functional domains (13, 17) (Fig. 1B). Degenerate PCRprimers were directed to these conserved sequence motifs (Ta-ble 1). The design of these primers was based on conservedpeptide motifs in other bacteria and fungi. Back-translation(reverse translation from protein to DNA) of these consensusmotifs was sensitive to the codon bias of cyanobacterial genesin general (7). Specifically, sequence motifs I and V of theadenylate-forming domain of known peptide synthetase mod-ules were used as target sequences (Fig. 1B). These functionaldomains are only weakly conserved in non-peptide synthetaseadenylate-forming enzymes (34). Degenerate primers wereused to detect genes encoding peptide synthetases (degeneratepeptide synthetase PCR [PS-PCR]).
The mcyB gene of M. aeruginosa PCC7608 (4) and its ortho-logue in M. aeruginosa HUB524 (19) were aligned in order todesign PCR primers specific for peptide synthetase genes in-volved in microcystin biosynthesis (microcystin synthetasePCR [MS-PCR]). These primers were directed to regionswithin the peptide synthetase module which were shown not tobe part of the series of conserved functional motifs (17). Theresulting PCR product did, however, contain a conserved coresequence (core motif II) which was used to align DNA se-quences for further phylogenetic analyses (Fig. 1B). Specificamplification primers (for MS-PCR) based on the character-ized peptide synthetase gene sequence of M. aeruginosa (4)were designed and directed to the region between conservedpeptide synthetase motifs I and III (Fig. 1B and Table 1). Thepriming sites for MS-PCR did not have a sequence identical inthe two Microcystis strains, PCC7806 and HUB524, and wereselected to enable amplification of a microcystin synthetasefragment from a broad range of microcystin-producing cya-nobacteria.
Detection of peptide synthetase genes in cyanobacteria byPCR. All DNA samples were checked for integrity in a cya-nobacterium-specific 16S ribosomal DNA amplification reac-tion prior to use in PS-PCR and MS-PCR (23). The degeneratePCR (PS-PCR) was used to identify cyanobacterial strains
VOL. 181, 1999 NONRIBOSOMAL PEPTIDE SYNTHESIS OF CYANOBACTERIA 4091
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
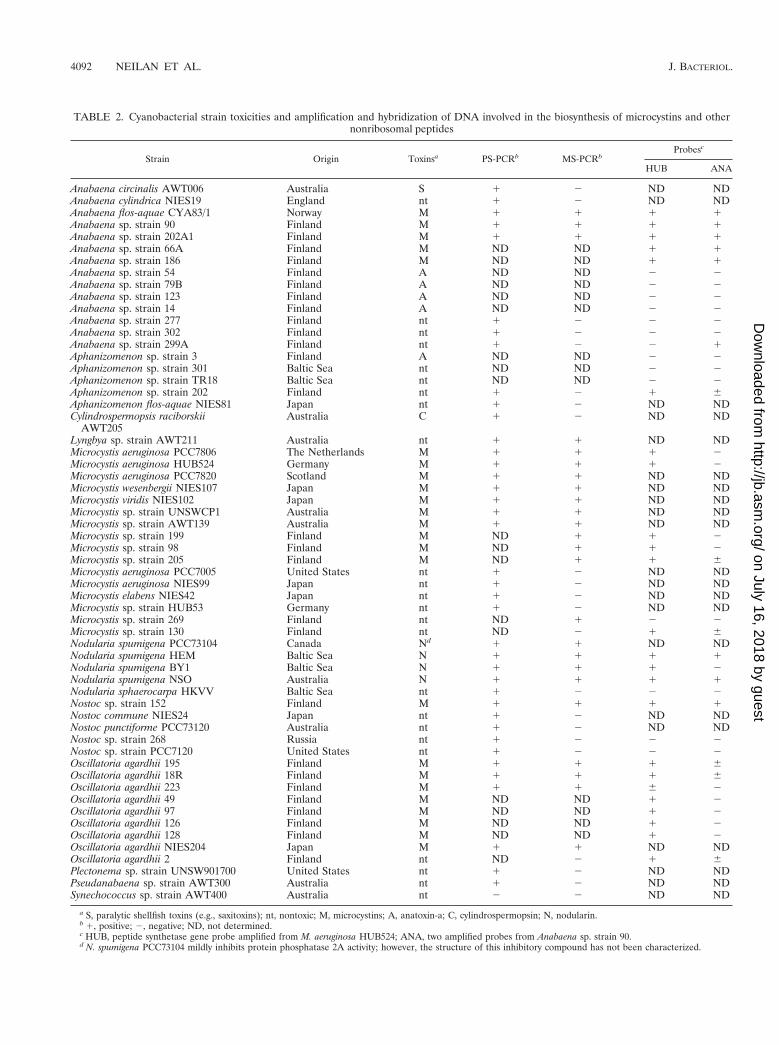
TABLE 2. Cyanobacterial strain toxicities and amplification and hybridization of DNA involved in the biosynthesis of microcystins and othernonribosomal peptides
Strain Origin Toxinsa PS-PCRb MS-PCRbProbesc
HUB ANA
Anabaena circinalis AWT006 Australia S 1 2 ND NDAnabaena cylindrica NIES19 England nt 1 2 ND NDAnabaena flos-aquae CYA83/1 Norway M 1 1 1 1Anabaena sp. strain 90 Finland M 1 1 1 1Anabaena sp. strain 202A1 Finland M 1 1 1 1Anabaena sp. strain 66A Finland M ND ND 1 1Anabaena sp. strain 186 Finland M ND ND 1 1Anabaena sp. strain 54 Finland A ND ND 2 2Anabaena sp. strain 79B Finland A ND ND 2 2Anabaena sp. strain 123 Finland A ND ND 2 2Anabaena sp. strain 14 Finland A ND ND 2 2Anabaena sp. strain 277 Finland nt 1 2 2 2Anabaena sp. strain 302 Finland nt 1 2 2 2Anabaena sp. strain 299A Finland nt 1 2 2 1Aphanizomenon sp. strain 3 Finland A ND ND 2 2Aphanizomenon sp. strain 301 Baltic Sea nt ND ND 2 2Aphanizomenon sp. strain TR18 Baltic Sea nt ND ND 2 2Aphanizomenon sp. strain 202 Finland nt 1 2 1 6Aphanizomenon flos-aquae NIES81 Japan nt 1 2 ND NDCylindrospermopsis raciborskii
AWT205Australia C 1 2 ND ND
Lyngbya sp. strain AWT211 Australia nt 1 1 ND NDMicrocystis aeruginosa PCC7806 The Netherlands M 1 1 1 2Microcystis aeruginosa HUB524 Germany M 1 1 1 2Microcystis aeruginosa PCC7820 Scotland M 1 1 ND NDMicrocystis wesenbergii NIES107 Japan M 1 1 ND NDMicrocystis viridis NIES102 Japan M 1 1 ND NDMicrocystis sp. strain UNSWCP1 Australia M 1 1 ND NDMicrocystis sp. strain AWT139 Australia M 1 1 ND NDMicrocystis sp. strain 199 Finland M ND 1 1 2Microcystis sp. strain 98 Finland M ND 1 1 2Microcystis sp. strain 205 Finland M ND 1 1 6Microcystis aeruginosa PCC7005 United States nt 1 2 ND NDMicrocystis aeruginosa NIES99 Japan nt 1 2 ND NDMicrocystis elabens NIES42 Japan nt 1 2 ND NDMicrocystis sp. strain HUB53 Germany nt 1 2 ND NDMicrocystis sp. strain 269 Finland nt ND 1 2 2Microcystis sp. strain 130 Finland nt ND 2 1 6Nodularia spumigena PCC73104 Canada Nd 1 1 ND NDNodularia spumigena HEM Baltic Sea N 1 1 1 1Nodularia spumigena BY1 Baltic Sea N 1 1 1 2Nodularia spumigena NSO Australia N 1 1 1 1Nodularia sphaerocarpa HKVV Baltic Sea nt 1 2 2 2Nostoc sp. strain 152 Finland M 1 1 1 1Nostoc commune NIES24 Japan nt 1 2 ND NDNostoc punctiforme PCC73120 Australia nt 1 2 ND NDNostoc sp. strain 268 Russia nt 1 2 2 2Nostoc sp. strain PCC7120 United States nt 1 2 2 2Oscillatoria agardhii 195 Finland M 1 1 1 6Oscillatoria agardhii 18R Finland M 1 1 1 6Oscillatoria agardhii 223 Finland M 1 1 6 2Oscillatoria agardhii 49 Finland M ND ND 1 2Oscillatoria agardhii 97 Finland M ND ND 1 2Oscillatoria agardhii 126 Finland M ND ND 1 2Oscillatoria agardhii 128 Finland M ND ND 1 2Oscillatoria agardhii NIES204 Japan M 1 1 ND NDOscillatoria agardhii 2 Finland nt ND 2 1 6Plectonema sp. strain UNSW901700 United States nt 1 2 ND NDPseudanabaena sp. strain AWT300 Australia nt 1 2 ND NDSynechococcus sp. strain AWT400 Australia nt 2 2 ND ND
a S, paralytic shellfish toxins (e.g., saxitoxins); nt, nontoxic; M, microcystins; A, anatoxin-a; C, cylindrospermopsin; N, nodularin.b 1, positive; 2, negative; ND, not determined.c HUB, peptide synthetase gene probe amplified from M. aeruginosa HUB524; ANA, two amplified probes from Anabaena sp. strain 90.d N. spumigena PCC73104 mildly inhibits protein phosphatase 2A activity; however, the structure of this inhibitory compound has not been characterized.
4092 NEILAN ET AL. J. BACTERIOL.
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

which contained significant DNA sequence similarity to theadenylate-forming domains of known peptide synthetase genes(13, 28, 34). With the described amplification reaction, putativepeptide synthetase genes were detected in strains of the cya-nobacterial genera Anabaena, Aphanizomenon, Cylindrosper-
mopsis, Microcystis, Nodularia, Nostoc, Oscillatoria, Plecto-nema, and Pseudanabaena. However, similar degenerate PCRproducts were not observed for the Synechococcus strain ana-lyzed (Table 2). The results presented in Table 2 were consis-tent with a larger body of data generated for several otherstrains of the species listed (data not shown).
Several members of the genera Anabaena, Aphanizomenon,and Nostoc were tested for the presence of cyclic peptide toxinsand sequences homologous to those of peptide synthetasegenes (Fig. 3A). No strains of nonhepatotoxic Anabaena rep-resenting the species Anabaena circinalis, A. cylindrica, and A.flos-aquae were shown to possess a microcystin synthetase geneorthologue by the described MS-PCR or probe hybridization.However, these species were shown to contain peptide syn-thetase gene orthologues. Of the 15 Anabaena strains exam-ined and listed in Table 2, 7 were microcystin producers; theother strains were either nontoxic or produced alkaloid neu-rotoxins (anatoxins or saxitoxins). These seven strains were
FIG. 2. Schematic representation of cyanobacterial DNAs on a template dotblot filter and results of hybridization and autoradiography. (A) Abbreviationsfor cyanobacterial genera are as follows: Anab, Anabaena; Nos, Nostoc; Mic,Microcystis; Osc, Oscillatoria; Nod, Nodularia; and Aph, Aphanizomenon. Hepa-totoxic (H), neurotoxic (N), and nontoxic (—) strains were investigated. (B)Hybridization pattern of the cyanobacterial dot blot filter with a 1.157-kb frag-ment of the mcyB gene of M. aeruginosa HUB524 as the probe. (C) Hybridizationpattern of the dot blot filter with a 2.0-kb fragment of a peptide synthetase genefrom Anabaena sp. strain 90 as the probe.
FIG. 3. Ethidium bromide-stained 1.5% agarose–Tris-acetate-EDTA elec-trophoresis gels showing peptide synthetase gene amplification products fromvarious toxic and nontoxic cyanobacteria (Table 2). (A) Amplification productsobtained with the degenerate PS-PCR. Lanes 1 to 13, PCR fragments from thecultured cyanobacteria Anabaena sp. strain 90, A. circinalis AWT006, Apha-nizomenon sp. strain 202, C. raciborskii AWT205, Lyngbya sp. strain AWT211, M.aeruginosa PCC7806, M. aeruginosa PCC7005, M. elabens NIES42, N. spumigenaPCC73104, Nostoc sp. strain 152, Nostoc sp. strain 203, Oscillatoria sp. strain 195,and O. agardhii NIES204, respectively. The marker lane (M) contains fX174digested with HaeIII, the top four bands being 1,358, 1,078, 872, and 603 bp. (B)Specific amplification of the microcystin synthetase gene by use of the MS-PCRdescribed in the text. Lanes 1 to 13 are the same as in panel A. (C) Results of theMS-PCR for uncultured cyanobacterial bloom samples containing the generaOscillatoria (lane 1), Nodularia (lane 2), and Microcystis (lanes 3 to 6). Themarker lane (M) contains SPP-1 digested with EcoRI, the six bottom bands being1,390, 1,160, 980, 720, 480, and 360 bp.
VOL. 181, 1999 NONRIBOSOMAL PEPTIDE SYNTHESIS OF CYANOBACTERIA 4093
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

identified by amplification of the MS-PCR product and bypositive probe hybridization (Fig. 2B and 3B).
The closely related genus Nostoc, which was represented byboth microcystin-producing and nontoxic strains possessedpeptide synthetase gene homologues in toxic and nontoxictypes and microcystin synthetase gene homologues selectivelyin only the hepatotoxic strain. Neurotoxic and nontoxic strainsof Aphanizomenon possessed peptide synthetase but not mi-crocystin synthetase gene orthologues. This result correlateswith the lack of microcystin-based toxicity observed to date forthe genus Aphanizomenon (Fig. 2 and 3). MS-PCR of non-microcystin-producing strains of cyanobacteria from thesethree genera did not reveal an amplification product.
The other filamentous and heterocyst-forming cyanobacte-ria used in this study, Nodularia and Cylindrospermopsis, alsoshowed gene detection experiment results which were congru-ent with toxin analyses. Members of the genus Nodularia areresponsible for the synthesis of nodularin, a cyclic pentapep-tide, similar in structure to microcystin and likewise hepato-toxic due to its potent inhibition of eukaryotic protein phos-phatases. PS-PCR and MS-PCR products were amplified fromall strains of Nodularia spumigena. These strains were alsoshown to produce nodularin, as detected by HPLC, or to in-hibit protein phosphatase 2A, as in the case of N. spumigenaPCC73104. The Nodularia strains used in this study were iso-lated from distinct geographic regions, implying a high degreeof gene conservation or a cosmopolitan distribution of hepa-totoxic strains of this brackish-water cyanobacterium (31). Noamplification of the MS-PCR fragment correlated with the lackof microcystin-based toxicity in Cylindrospermopsis raciborskii.This cyanobacterium is capable of synthesizing a hepatotoxicalkaloid known as cylindrospermopsin. The presence of PS-PCR products for the strains studied may be related to thepresence of peptide synthetase genes of unknown function.
PS-PCR revealed that all strains of Microcystis studied pos-sess one or more peptide synthetase loci. It has been shownthat strains of Microcystis, irrespective of their ability to pro-duce microcystin, as determined by chemical analyses and bio-assays, contain DNA sequences with significant identity toknown peptide synthetase genes (3). The data presented heresupport these findings and describe a rapid method for thedetermination of potentially hepatotoxic Microcystis based onthe amplification of the MS-PCR template. Samples of uncul-tured cyanobacterial blooms containing hepatotoxic Micro-cystis, Nodularia, or Oscillatoria strains were all detected byMS-PCR (Fig. 3C). Both the degenerate and the specific am-plification products were generated coordinately only forstrains which were shown to produce microcystin or otherwiseinhibit eukaryotic protein phosphatases (Fig. 3 and Table 2).Moreover, we sequenced the products obtained by MS-PCRfrom M. aeruginosa strains and consistently found significantidentity to the mcyB gene of strain PCC7806 (see below). It hasrecently been shown that a single gene cluster inclusive ofmcyB is responsible for the synthesis of all microcystin isoformsin M. aeruginosa PCC7806 and that all protein phosphataseinhibition activity is due to cellular microcystin content (4).
The amplification of an MS-PCR product for nonhepato-toxic Microcystis sp. strain 269 was not supported by theHUB524 microcystin synthetase gene probe hybridization (Fig.2A). All other nontoxic strains of this genus did not showamplification of the microcystin-specific DNA sequence (Table2). Strain 269 may contain a genome sequence compatible withthe MS-PCR primers but not the dot blot probe. In addition, itis also possible that toxin levels in this strain are below detec-tion limits or that the altered expression of peptide synthetasegenes in laboratory cultures has inhibited microcystin biosyn-
thesis by this strain (36). It is also possible that mutationalinactivation of the microcystin synthetase gene in regions otherthan the fragments assayed resulted in the observed differencesamong MS-PCR, probe hybridizations, and toxicity tests.
All the Oscillatoria agardhii strains produced detectable lev-els of microcystin, except for one, Oscillatoria sp. strain 2. Thisstrain also cross-hybridized with the HUB524 peptide syn-thetase probe but was found to be nontoxigenic by MS-PCR.MS-PCR experiments and/or probe hybridizations indicatedthat these strains, including strain 2, contained orthologoustoxin biosynthesis genes (Fig. 2B and 3B).
Of the remaining cyanobacterial groups to be tested with thedescribed molecular methods, the filamentous and non-hetero-cyst-forming genera Lyngbya, Pseudanabaena, and Plectonema(Table 2) and several stromatolite-associated (benthic) cya-nobacteria (data not shown) showed genomic orthology to theconserved peptide synthetase loci but not to the microcystinsynthetase gene. The sole Synechococcus isolate examined didnot possess peptide synthetase or microcystin synthetase (Ta-ble 2). Similarly, there was a lack of peptide synthetase ortho-logues in the genome of Synechocystis sp. strain PCC6803 (12).These results indicate that a broad range of cyanobacteria arecapable of nonribosomal peptide synthesis.
Sequence analysis and evolutionary relationships betweenthe putative microcystin synthetase gene and other peptidesynthetase genes in cyanobacteria. The specific MS-PCR prod-ucts from 12 hepatotoxic cyanobacterial strains (representingfive genera and seven species) and four PS-PCR products(representing peptide synthetase modules of unknown func-tion) from nontoxic M. aeruginosa HUB53 (three modules)and NIES99 (one module) were purified and sequenced. Ap-proximately 700 bp of the putative microcystin synthetase genewas determined for each strain. Furthermore, we included therespective most similar sequence from another module of themicrocystin synthetase gene, mcyC, that had been determinedduring a study aimed at identifying the entire microcystin syn-thetase gene cluster of M. aeruginosa PCC7806 (33a). DNAsequences of the microcystin synthetase gene and other pep-tide synthetase gene orthologues were aligned with each other,and the pairwise (Jukes-Cantor) genetic distances were calcu-lated. These distances were represented in a phylogenetic anal-ysis (Fig. 4A). Differences between the peptide synthetase genesequences were surprisingly large and reflected relatively lowsequence similarity between the highly conserved and func-tional domain motifs of each peptide synthetase module (Fig.4B). The unicellular cyanobacteria of the genus Microcystis hada second cluster of MS-PCR sequences which was delineatedfrom the PS-PCR sequences of the nontoxic Microcystis strains.We concluded from this observation that, for the Microcystisspecies studied, MS-PCR specifically amplified a sequence ofthe mcyB gene and thus may be used to identify potentialmicrocystin producers in these species.
However, the situation appears more complicated for strainsof Anabaena, Nodularia, Nostoc, and Oscillatoria. For thesestrains, MS-PCR amplified sequences were clearly differentfrom the sequences amplified for Microcystis strains. Moreover,some of these sequences (N. spumigena BY1, Nostoc sp. strain152, and O. agardhii 195) clustered more closely with the MS-PCR products from Microcystis strains, whereas other se-quences (Anabaena sp. strain 90 and N. spumigena HEM andPCC73104) were more related to the PS-PCR products fromnontoxic M. aeruginosa strains. Since the sequences betweendifferent modules (having different functions) and of differentpeptide synthetase genes (again, having different functions)were compared (Fig. 4B), no phylogenetic relationships can beconcluded from the PS-PCR sequences. The above-mentioned
4094 NEILAN ET AL. J. BACTERIOL.
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

FIG. 4. (A) Phylogenetic affiliations in a defined region of peptide synthetase genes from strains of the cyanobacterial genera Anabaena, Microcystis, Oscillatoria,Nodularia, and Nostoc. M. aeruginosa HUB53 is nonhepatotoxic and contains a peptide synthetase with paralogous modules designated M, S, and U (27a). Comparisonswere based on partial sequence data from MS-PCR products (hepatotoxic strains in Table 2) and PS-PCR products (nontoxic strains). The phenogram wasreconstructed from a pairwise distance matrix by use of the neighbor-joining method. Hepatotoxic and nontoxic clusters of Microcystis strains are enclosed by squarebrackets. (B) Evolutionary relationships among peptide synthetases of microcystin-producing cyanobacteria, nontoxic cyanobacteria, and other bacteria and fungi.Translated MS-PCR and PS-PCR DNA sequences of cyanobacteria were compared to sequences of similar database peptides. Divergence between amino acidsequences was calculated by use of PAM-Dayhoff matrix, and the tree was constructed by use of the neighbor-joining method obtained from the PHYLIP suite ofprograms. Peptide synthetases, including various modules of the same multienzyme complex, are as follows: GrsA and GrsB, gramicidin A and B synthetases (withmodules A to C), respectively, from Bacillus brevis; SrfA, SrfB, and SrfC, surfactin A, B, and C synthetases, respectively, from B. subtilis; TycA, tyrocidin A synthetasefrom Brevibacillus brevis; EntF, enterobactin synthetase component F from Escherichia coli; Hts1, HC toxin synthetase 1 (modules A to D) from Cochliobolus carbonum;and SimA, cyclosporin A synthetase from Tolypocladium niveum. McyB and McyC are involved in microcystin biosynthesis in the M. aeruginosa strains indicated (Table2). M. aeruginosa strains HUB53 and NIES99 are nontoxic and possess putative peptide synthetases of unknown function. The scale for each tree indicates inferredevolutionary distances. Bootstrap values greater than 50% which were derived from 100 resampling events of the aligned sequence data and which support the treetopologies are also shown.
4095
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

cluster of sequences from different cyanobacterial genera thatpossibly represents the mcyB gene (Fig. 4A), however, reflectsorganismic phylogenetic relationships, as inferred by 16SrRNA gene sequences (9, 23, 35). More information on thesequence and organization of peptide synthetase genes in cya-nobacteria and of the microcystin synthetase gene in particularis required before final conclusions can be made regarding theevolution and phylogeny of microcystin biosynthesis.
The present study has shown, for the first time, that micro-cystin synthetase gene orthologues are present not only in alltoxic strains of the genus Microcystis but also in microcystin-producing strains of the genera Anabaena, Oscillatoria, andNostoc. Nodularin-producing cyanobacteria of the genusNodularia also appear to possess a microcystin synthetase geneorthologue and therefore a similar biosynthetic pathway fortoxin production. We have also shown that strains of othertoxic and nontoxic cyanobacterial genera, such as Anabaena,Aphanizomenon, Cylindrospermopsis, Lyngbya, Nodularia, Nos-toc, Oscillatoria, Plectonema, and Pseudanabaena contain genesfor similar peptide synthetase complexes of unknown function.Due to this broad intergeneric distribution of integrated en-zyme systems, therefore, cyanobacteria provide a rich andnovel source of many uncharacterized amino-acid-activatingand -modifying peptide synthetase modules (18, 28, 32). Thisstudy reveals a molecular approach to the discovery of novelbioactive compound synthetic pathways in uncultured cya-nobacteria and probably other microorganisms. The specificPCR (MS-PCR) was applicable to the rapid and sensitivedetection of toxigenic strains of Microcystis. This methodcould also be used to identify microcystin-producing strains ofother cyanobacterial genera. Further characterization (includ-ing insertional inactivation) of various microcystin synthetasegene orthologues from Nodularia, Anabaena, Nostoc, and Os-cillatoria will enable the design of specific PCRs for the detec-tion of potential hepatoxin producers in each of these genera.This strategy will provide a procedure for detecting toxic ge-notypes prior to the production of toxins by relevant cyanobac-terial species.
ACKNOWLEDGMENTS
Additional strains used in this study were kindly supplied by SueBlackburn (CSIRO) and Boris Gromov (University of St. Petersburg).The anonymous reviewers are thanked for their contributions to themanuscript.
This work was funded in Australia by the Australian ResearchCouncil, Australian Water Technologies, and the Co-operative Re-search Center for Water Quality and Treatment; in Germany by theGerman Research Foundation, DFG (grant Bo 1045/13-3), and theEuropean Commission (grant BIO 4 CT96-0256); and in Finland bythe University of Helsinki and the Academy of Finland. B.A.N. wassupported by fellowships from the Alexander von Humboldt Founda-tion and the Australian Research Council. K.S. is a senior researchscientist of the Academy of Finland.
REFERENCES
1. An, J., and W. W. Carmichael. 1994. Use of a colorimetric protein phospha-tase inhibition assay and enzyme linked immunosorbent assay for the studyof microcystins and nodularins. Toxicon 32:1495–1507.
2. Arment, A. R., and W. W. Carmichael. 1995. Evidence that microcystin is athiotemplate product. J. Phycol. 32:591–597.
3. Dittmann, E., K. Meißner, and T. Borner. 1996. Conserved sequences ofpeptide synthetase genes in the cyanobacterium Microcystis aeruginosa. Phy-cologia 35:62–67.
4. Dittmann, E., B. A. Neilan, M. Erhard, H. von Dohren, and T. Borner. 1997.Insertional mutagenesis of a peptide synthetase gene which is responsible forhepatotoxin production in the cyanobacterium Microcystis PCC7806. Mol.Microbiol. 26:779–787.
5. Eloff, J. N. 1981. Autecological studies on Microcystis, p. 71–96. In W. W.Carmichael (ed.), The water environment—algal toxins and health. PlenumPress, New York, N.Y.
6. Erhard, M., H. von Dohren, and P. Junblut. 1997. Rapid typing and eluci-dation of new secondary metabolites of intact cyanobacteria using MALDI-TOF mass spectrometry. Nat. Biotechnol. 15:906–909.
7. Fay, P., and C. van Baalen. 1987. The cyanobacteria. Elsevier/North-HollandPublishing Co., Amsterdam, The Netherlands.
8. Fujiki, H. 1992. Is the inhibition of protein phosphatase 1 and 2A activitiesa general mechanism of tumor promotion in human cancer development?Mol. Carcinog. 5:91–94.
9. Giovanonni, S. J., S. Turner, G. J. Olsen, S. Barns, D. J. Lane, and N. R.Pace. 1988. Evolutionary relationships among cyanobacteria and green chlo-roplasts. J. Bacteriol. 170:3584–3592.
10. Golden, J. W., C. D. Carrasco, M. E. Mulligan, G. J. Schneider, and R.Haselkorn. 1988. Deletion of a 55-kilobase-pair DNA element from thechromosome during heterocyst differentiation of Anabaena sp. strain PCC7120. J. Bacteriol. 170:5034–5041.
11. Honkanen, R. E., F. R. Caplan, K. K. Baker, C. L. Baldwin, S. C. Bobzin,C. M. Bolis, G. M. Cabrera, L. A. Johnson, J. H. Jung, L. K. Larsen, I. A.Levine, R. E. Moore, C. S. Nelson, G. M. L. Patterson, K. D. Tschappat,G. D. Tuang, A. L. Boynton, A. R. Arment, J. An, W. W. Carmichael, K. D.Rodland, B. E. Magun, and R. A. Lewin. 1995. Protein phosphatase inhibi-tory activity in extracts of cultured blue-green algae (Cyanophyta). J. Phycol.31:478–486.
12. Kaneko, T., A. Tanaka, S. Sato, H. Kotani, T. Sazuka, N. Miyajima, M.Sugiura, and S. Tabata. 1995. Sequence analysis of the genome of theunicellular cyanobacterium Synechocystis sp. PCC6803. I. Sequence featuresin the 1 Mb region from map position 64% to 92% of the genome. DNA Res.2:153–166.
13. Kleinkauf, H., and H. von Dohren. 1996. A non-ribosomal system of peptidebiosynthesis. Eur. J. Biochem. 236:335–351.
14. Krishnamurthy, T., W. W. Carmichael, and E. W. Sarver. 1986. Toxic pep-tides from freshwater cyanobacteria (blue-green algae). I. Isolation, purifi-cation and characterisation of peptides from Microcystis aeruginosa andAnabaena flos-aquae. Toxicon 24:865–873.
15. Lawton, L. A., C. E. Edward, and G. A. Codd. 1994. Extraction and highperformance liquid chromatographic method for the determination of mi-crocystins in raw and treated waters. Analyst 119:1525.
16. Lehtimaki, J., P. Moisander, K. Sivonen, and K. Kononen. 1997. Growth,nitrogen fixation, and nodularin production by two Baltic Sea cyanobacteria.Appl. Environ. Microbiol. 63:1647–1656.
17. Marahiel, M. A., T. Stachelhaus, and H. D. Mootz. 1997. Modular peptidesynthetases involved in non-ribosomal peptide synthesis. Mol. Gen. Genet.97:2651–2673.
18. McDaniel, R., S. Ebert-Khosla, D. A. Hopwood, and C. Khosla. 1993. Engi-neered biosynthesis of novel polyketides. Science 262:1546–1557.
19. Meißner, K., E. Dittmann, and T. Borner. 1996. Toxic and non-toxic strainsof the cyanobacterium Microcystis aeruginosa contain sequences homologousto peptide synthetase genes. FEMS Microbiol. Lett. 135:295–303.
20. Moore, R. E. 1996. Cyclic peptides and depsipeptides from cyanobacteria: areview. J. Ind. Microbiol. 16:134–143.
21. Namikoshi, M., and K. L. Rinehart. 1996. Bioactive compounds produced bycyanobacteria. J. Ind. Microbiol. 17:373–384.
22. Neilan, B. A. 1995. Identification and phylogenetic analysis of toxigeniccyanobacteria by multiplex randomly amplified polymorphic DNA PCR.Appl. Environ. Microbiol. 61:2286–2291.
23. Neilan, B. A., D. Jacobs, T. Del Dot, L. Blackall, P. R. Hawkins, P. T. Cox,and A. E. Goodman. 1997. Ribosomal RNA sequences and evolutionaryrelationships among the toxigenic cyanobacteria of genus Microcystis. Int. J.Syst. Bacteriol. 47:693–697.
24. Neilan, B. A., D. Jacobs, and A. E. Goodman. 1995. Genetic diversity andphylogeny of toxic cyanobacteria determined by DNA polymorphisms withinthe phycocyanin locus. Appl. Environ. Microbiol. 61:3875–3883.
25. Rapala, J., K. Sivonen, C. Lyra, and S. I. Niemela. 1997. Variation ofmicrocystins, cyanobacterial hepatotoxins, in Anabaena spp. as a function ofgrowth stimuli. Appl. Environ. Microbiol. 64:2206–2212.
26. Rinehart, K. L., M. Namikoshi, and B. W. Choi. 1994. Structure and bio-synthesis of toxins from blue-green algae (cyanobacteria). J. Appl. Phycol.6:159–176.
27. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, N.Y.
27a.Schaub, V. Unpublished results.28. Schneider, A., T. Stachelhaus, and M. A. Marahiel. 1998. Targeted alter-
ation of the substrate specificity of peptide synthetases by rational moduleswapping. Mol. Gen. Genet. 257:308–318.
29. Sivonen, K. 1996. Cyanobacterial toxins and toxin production. Phycologia35:12–24.
30. Sivonen, K., W. W. Carmichael, M. Namikoshi, K. L. Rinehart, A. M.Dahlem, and S. I. Niemela. 1990. Isolation and characterization of hepato-
4096 NEILAN ET AL. J. BACTERIOL.
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

toxic microcystin homologs from Nostoc sp. strain 152. Appl. Environ. Mi-crobiol. 56:2650–2657.
31. Sivonen, K., K. Kononen, W. W. Carmichael, A. M. Dahlem, K. L. Rinehart,J. Kiviranta, and S. I. Niemela. 1989. Occurrence of the hepatotoxic cya-nobacterium Nodularia spumigena in the Baltic Sea and structure of thetoxin. Appl. Environ. Microbiol. 55:1990–1995.
32. Stachelhaus, T., A. Schneider, and M. A. Marahiel. 1995. Rational design ofpeptide antibiotics by targeted replacement of bacterial and fungal domains.Science 269:69–72.
33. Theiss, W. C., W. W. Carmichael, J. Wyman, and R. Bruner. 1988. Bloodpressure and hepatocellular effects of the cyclic heptapeptide toxin produced
by the freshwater cyanobacterium (blue-green alga) Microcystis aeruginosastrain PCC7820. Toxicon 26:603–613.
33a.Tillett, D. Unpublished data.34. Turgay, K., and M. A. Marahiel. 1994. A general approach for identifying
and cloning peptide synthetase genes. Peptide Res. 7:238–241.35. Turner, S. 1998. Molecular systematics of oxygenic photosynthetic bacteria.
Plant Syst. Evol. 11:13–52.36. Watanabe, M. F., and S. Oishi. 1985. Effects of environmental factors on
toxicity of a cyanobacterium (Microcystis aeruginosa) under culture condi-tions. Appl. Environ. Microbiol. 49:1342–1344.
VOL. 181, 1999 NONRIBOSOMAL PEPTIDE SYNTHESIS OF CYANOBACTERIA 4097
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from