of b1omxc.a chemi~y vol. 269; no. 2, of 14, pp. 1294-1300 ... · the baculovirus transplacement...
TRANSCRIPT

’lb JOURNAL OF B1omxc.a C H E M I ~ Y 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269; No. 2, Issue of January 14, pp. 1294-1300, 1994 Printed in U S A .
Purple Acid Phosphatase of the Human Macrophage and Osteoclast CHARACTERIZATION, MOLECULAR PROPERTIES, AND CRYSTALLIZATION OF THE RECOMBINANT DI-IRON-OXO PROTEIN SECRETED BY BACULOVIRUS-INFECTED INSECT CELLS*
(Received for publication, July 21, 1993)
Alison R. HaymanS and Timothy M. Cox5 From the DeDartment of Medicine, Universitv of Cambridge, Level 5, Addenbrooke’s Hospital, Cambridge dB2 2QQ, United Kingdom
“ .
The purple phosphatases catalyze hydrolysis of phos- phate esters (optimum pH - 5 ) and are resistant to in- hibition by dextro-rotatory tartrate; their distinctive color is due to Fe(II1)-phenolate charge-transfer transi- tions at their active site. Expression of human purple phosphatase, designated type 5 acid phosphatase, is re- stricted to osteoclasts and other activated cells of mono- histiocytic lineage, but its biological Ale in relation to bone resorption and phagocytosis is unknown. To char- acterize this enzyme further, we have engineered the human type 5 acid phosphatase into a baculovirus vec- tor expression system that enabled milligram quantities of purple protein to be purified from medium containing Sf9 host cells. The phosphatase cDNAwas transcribed as a single RNA species of 1.6 kilobases as in human tissues. Tartrate-resistant acid phosphatase activity reacting with uteroferrin antisera appeared in the culture me- dium, from which up to 8 mglliter was purified by two- step cation-exchange chromatography at pH 8.0. Two isoforms of -36 kDa were identified by SDS-polyacryl- amide electrophoresis and were converted to a single species of apparent molecular size 34 kDa upon treat- ment with N-glycosidase F, indicating secreted glyco- forms of a single polypeptide. Mass spectroscopy showed that the mean molecular mass of the active, se- creted glycoprotein was 35849 Da. The recombinant en- zyme (specific activity, 190 p o l p-nitrophenoYmidmg at 37 “C) contained 2 iron atomdmolecule and formed purple, monoclinic crystals. Exposure to the ferric che- lator, 1,2-dimethyl-3-hydroxypyrid-4-one, rapidly inacti- vated the enzyme, which was not inhibited by a,a’-bi- pyridyl, a ferrous chelator. That ferric iron is essential for enzymatic catalysis, was further indicated by the synergistic effects of the reductant, dithiothreitol, and bipyridyl on phosphatase activity. The recombinant purple phosphatase catalyzed the peroxidation of 5-aminophthalhydrazide (luminol), as evidenced by the induction of chemiluminescence; this reaction was in- hibited by a,a‘-bipyridyl at concentrations that did not inhibit phosphatase activity. The divalent iron moiety of human type 6 phosphatase may therefore participate in the generation of free radical species by fluid-phase re- actions involving Fenton chemistry that are dissociated from its phosphatase function.
* This work was supported in part by the Cancer Research Campaign and the Wellcome Trust. The costs of publication of this article were defrayed in part by the payment of page charges. This article must
U.S.C. Section 1734 solely to indicate this fact. therefore be hereby marked “advertisement” in accordance with 18
$ Supported by a grant from SmithKline Beecham. 9 To whom correspondence should be addressed. Tel.: 011-44-223-
336864; Fax: 011-44-223-336846.
Human type 5 acid phosphatase (orthophosphoric monoester phosphohydrolase (acid optimum); EC 3.1.3.2) is a lysosomal protein expressed principally in differentiated cells of monohis- tiocytic lineage. The isozyme is identified by its unique catho- dal electrophoretic mobility at pH 4 and is further defined by its resistance to inhibition by I,(+)-tartrate (1). Type 5 acid phos- phatase, which promotes the hydrolysis of phosphoproteins, nucleotides (including ATP), and arylphosphates, belongs to a class of purple proteins that contain a catalytically active di- iron-oxo cluster (2). Although the porcine homologue, uterofer- rin, may serve to deliver iron secreted by the endometrium to the developing piglet in utero (31, the biological function of, and natural substrate for, human type 5 acid phosphatase is un- known (4).
Strong histochemical reactions for tartrate-resistant acid phosphatase are observed in normal osteoclasts (5) and are characteristic of Gaucher’s cells (6) and the phagocytic B lym- phocytes of hairy cell leukemia (7). Moreover, serum tartrate- resistant acid phosphatase activity is pathologically increased in conditions including Paget’s disease and hyperparathyroid- ism where bone resorption is enhanced (8-10). Northern blot analysis using a human cDNA hybridization probe has recently confirmed that expression of type 5 acid phosphatase is re- stricted to cells originating from granulocyte-monocyte pro- genitors. A single 1.5-kilobase transcript was identified in os- teoclasts, alveolar, and monocyte-derived macrophages but was not detected in human blood monocytes or normal splenic tis- sue (11). Similar studies indicate that expression of the phos- phatase accompanies phorbol ester-induced hairy cell transfor- mation of B-lymphocytic leukemia cells (12). Thus expression of type 5 acid phosphatase is correlated with activation and dif- ferentiation of osteoclasts, macrophages, and other cells of monohistiocytic phenotype.
Recently, it has become clear that di-iron-oxo clusters are found at the metal-binding sites of proteins that are widely dis- tributed and associated with diverse functions. Low tempera- ture electron paramagnetic resonance studies have detected an intense lowg value signal (g’ = 1.74) in the purple phosphatases with integrated intensities accounting for a maximum of half of the specifically bound iron, suggesting that the iron pairs are sequestered in anti-parallel spin-coupled binuclear clusters (2, 13-17). This proposal has been confirmed by magnetic suscep- tibility studies (18). Apart from the purple phosphatases, di- iron-oxo clusters have been identified in hemerythrin and ri- bonucelotide reductase (19); evidence of these iron centers has been found also in toluene-4-hydroxylase, phenol hydroxylase, stearoyl-acyl carrier protein Ag-desaturase, and methane monooxygenase hydroxylase (20-26). The involvement of sev- eral of these proteins in the catalysis of oxygen-dependent re- actions and the occurrence of inter-convertible redox forms of the purple phosphatases, suggests that type 5 acid phosphatase may also participate in electrochemical processes.
1294

Human Purple Phosphatase 1295
Macrophages and osteoclasts possess a common lineage (27) and generate oxygen-derived free radicals that have been as- sociated with microbial killing and bone resorption (28-31). Given that human monocyte-derived macrophages and osteo- clasts synthesize the purple phosphatase at a high level (5 , 10, 11,32), we have considered that this unusual protein may play a central role in the formation of reactive intermediates that bring about these effector-cell functions. Accordingly, to char- acterize further the catalytic, structural, and biochemical prop- erties of the purple phosphatases, we have engineered the hu- man type 5 phosphatase into a baculovirus expression vector system that allowed large quantities of functional protein to be secreted by insect cells into culture medium, from which it was easily purified for investigation.
EXPERIMENTAL PROCEDURES Materials-The full-length complementary DNAfor type 5 acid phos-
phatase had been isolated from a human placental expression library in hgtll (32) using a probe derived by mixed oligonucleotide-primed am- plification of human splenic cDNA (33) in the polymerase chain reac- tion. The degenerate primers used in the polymerase chain reaction were derived from amino acid sequences that are conserved between porcine uterofemn and bovine spleen acid phosphatase.
The baculovirus transplacement vector PAC CL29 and wild type Au- tograph californica nuclear polyhedrosis virus (AcNPV)' were ob- tained from Dr. Vivienne Murphy, SmithKline Beecham laboratories, Great Burgh, Surrey, United Kingdom; insect host cells from the ovar- ian streak of the fall armyworm Spodoptera frugiperda (Sf9) were ob- tained from Dr. J . Sinclair, Department of Medicine, University of Cam- bridge.
Uteroferrin Antisem-Antisera to porcine uterofemn were obtained by immunizing rabbits with the purified protein that had been kindly donated previously by Dr. Philip Aisen, Albert Einstein College of Medi- cine. Antisera were validated by two-dimensional immunodiffusion and immunoenzymatic capture assay of tartrate-resistant acid phosphatase activity in human tissue extracts as described previously (32, 34, 35). Immunospecific antibody was purified by affinity absorption to utero- femn immobilized on CNBr-agarose (2 mg/ml swollen gel; Pharmacia Fine Biochemicals) and eluted with 2% (w/v) aqueous acetic acid (34). An additional sample of pure porcine uterofemn was generously sup- plied by Dr. R. M. Roberts, Animal Sciences Center, Columbia, MO.
Cell Cultures-Sf9 cells were propagated at 28 "C either as suspen- sion cultures in TClOO medium (Flow Laboratories) supplemented with 10% (w/v) fetal calf serum, or as monolayer cultures in TC100/10% (w/v) fetal calf serum or Sf 900 medium (Life Technologies, Inc.).
Purification of Viral DNA-Wild type AcNPV DNA was purified by the proteinase K digestion procedure (36).
Recombinant Vector Construct-The human type 5 acid phosphatase cDNA was ligated into the BamHI site of the transfer vector pAcCL29 according to Maniatis et al. (37).
Recombinant Virus Production-A baculovirus containing the acid phosphatase coding sequence driven by the polyhedrin promoter was produced by co-transfection of recombinant plasmid DNAwith wild type AcNPV DNA into Sf9 cells. 96 h after co-transfection, virions extruded into the culture medium were analyzed using the plaque assay (38) and subsequent staining with neutral red. Plaques resulting from recombi- nant virus were identified by the presence of a bluish halo following overnight incubation at 28 "C in the presence of 5 mg/ml 5-bromo-4- chloro-3-indolyl phosphate, 5 mg/ml nitro blue tetrazolium, 200 m sodium acetate (pH 5.6), and 100 m disodium tartrate. Positive plaques were purified to homogeneity by two more rounds of plaque assay. High titer virus stocks (lo7 to lo8 plaque-forming units/ml) were raised from the purified virus (38).
Large Scale Production ofRecombinant Protein-Sf9 cells (1.8 x lo7) were seeded into T75 tissue culture flasks containing 15 ml of Sf900 medium and inoculated with 0.5 ml of high titer virus. After incubating for 5-6 days at 28 "C, the medium was decanted, centrifuged to remove cell debris, and stored at 4 "C.
Purification of Expressed Recombinant Protein-Serum-free insect medium (220 ml) containing recombinant protein, was alkalinized to pH 8 by addition of NaOH and incubated at 4 "C for 16 h. After centrifu-
The abbreviations used are: AcNPV, A. californica nuclear polyhe-
resistant) acid phosphatase. drosis virus; Sf9, S. frugiperdu cell line; AP5, type (band) 5 (tartrate-
gation at 2000 x g for 15 min, the supernatant was applied to a 0.9 cm x 2.0-cm S-Sepharose column (Pharmacia) equilibrated with 50 II~M HEPES (pH 8). A 60-ml linear gradient of 0-0.6 M NaCl in the same buffer was applied to elute the protein. Acid phosphatase activity was determined, and active fractions were pooled, diluted 10 times with buffer, and applied to a 0.5 x 5.0-cm Mono S column (Pharmacia) and eluted under the same conditions as for S-Sepharose chromatography.
Electrophoresis-SDS-polyacrylamide gel electrophoresis was car- ried out according to the method of Laemmli (39). Gels were stained with Coomassie Brilliant Blue or subjected to analysis by immunoblot- ting.
Immunoblotting-Proteins separated by SDS-polyacrylamide gel electrophoresis were transferred to polyvinylidene difluoride mem- branes (36) (Immobilon, Millipore Ltd.) using an LKB semi-drive elec- troblotting apparatus. The membrane was incubated for 2 h in 3% dried milk ("Marvel"), phosphate-buffered saline (pH 7.4). 0.2% Tween 20 and then incubated sequentially with rabbit uterofemn antiserum diluted 1:100, biotin-conjugated anti-rabbit IgG (1:20,000, Sigma), and avidin- alkaline phosphatase conjugate (1:20,000, Sigma). These incubations were camed out for 18,2, and 2 h respectively, and the membrane was washed three times with phosphate-buffered saline, 0.2% Tween 20 between each change of reagent. The membrane was incubated with 0.1 M ethanolamine-HCl (pH 9.6) for 5 min and then with developing sub- strate solution (100 pg/ml 5-bromo-4-chloro-3-indolyl phosphate, 100 pg/ml nitro blue tetrazolium, 1 m MgC12, 0.1 M ethanolamine-HC1 (pH 9.6)). The reaction was terminated by rinsing the membrane with dis- tilled water.
Human Acid Phosphatase cDNA Probe-The acid phosphatase cDNA was labelled with [CX-~~PI~CTP using the Multiprime DNA-labeling sys- tem (Amersham Corp.).
Southem Blot Analysis-DNA was extracted from insect cells by alkaline lysis (37), digested, electrophoresed in a 1% agarose gel, and than transferred to a Hybond membrane (Amersham Corp.) as de- scribed by Southern (37). Hybridization with the phosphatase probe was carried out for 16 h a t 65 "C in Blotto (1.5 nm NaP04 (pH 7.7), 0.27 M NaCl, 1.5 nm EDTA, 0.5% (w/v) dried milk, 1% SDS, 6% polyethylene glycol 6000). Filters were washed in 1 x SSC (SSC = 0.15 M NaCl, 15 nm trisodium citrate (pH 7.5)), 0.1% SDS, and then in 0.2 x SSC, 0.1% SDS at 65 "C for 1 h.
Northern Blot Analysis-Total RNA was prepared from insect cells by the guanidinium isothiocyanate method (40) and quantified by ultra- violet spectroscopy (37). Samples were electrophoresed on 1% formaldehyddagarose gels transferred to Hybond N membranes and processed as for Southern blotting.
Assay Procedures-Phosphatase activity was assayed in the presence of 100 lll~ disodium tartrate using 10 nm 4-nitrophenyl phosphate as substrate (32, 41), which was buffered to pH 5.6 by 200 nm sodium acetate. The release of nitrophenol was determined spectrophotometri- cally by measuring absorbance at 405 m after stopping the reaction by alkalinization with NaOH (41).
Protein Determinations-Protein concentrations were quantified by the dye binding method of Bradford (42) and by the fluorescamine procedure (43).
Protein Deglycosylation-Proteins were incubated at 95 "C for 15 min and sonicated for 1 min in the presence of 1% (w/v) SDS. The samples were diluted with 5 volumes of 50 nm sodium phosphate (pH 8), 20 II~M EDTA, 10 nm p-mercaptoethanol, 1% n-octyl glucoside, and digested with N-glycosidase F (Boehringer Mannheim) for 16 h at 37 "C.
Iron Content-Iron analysis was carried out using the bathophenan- throline (Sigma) method of Landers and Zak (44). a,d-Bipyridyl was purchased from Sigma; 1,2-dimethyl-3-hydroxypyrid-4-one was gener- ously provided by Dr. G. Kontiorghes, Royal Free Hospital, London.
Protein Crystallization-Crystalline human acid phosphatase was obtained from solutions containing 5-10 mg/ml purified human type 5 acid phosphatase protein by vapor diffusion using the hanging-drop method with 12% (w/v) polyethylene glycol 6000 as the precipitant (45). Crystals were examined by transmission light microscopy using a Ni- kon type 115 instrument (Nikon, Tokyo, Japan).
Mass Spectroscopy-The molecular weight of purified recombinant human purple phosphatase was determined in the VG Bio-Q mass spectrometer using electrospray ionization. The mobile phase was 50% (w/v) aqueous methanol, containing 1% (w/v) acetic acid, and 2 mg/liter gramicidin-S.
Chemiluminescence-Chemiluminescence was recorded in a Hamil- ton Lumicon luminometer. The reaction was started by addition of hydrogen peroxide ( h a 1 concentration 4.4 nm) to a solution of 5-amin- ophthalhydrazide (luminol, Sigma) in 1 ml of 0.2 M Tris-HC1 (pH 9.0), containing S 5 0 1 1 ~ purple phosphatase. Photon emission was meas-

1296 Human Purple Phosphatase
A B
Kb Kb
11.5 -
5.07 -
2.8 -
1.1
0.51 -
9.5 - 7.5 -
4.4 -
2.4
1.2 -
0.2 -
1 2 3 1 2 3
FIG. 1. A, Southern blot hybridization analysis of transfected insect cells. Five pg of genomic DNA obtained from Sf9 infected with recom- binant AcNPV vector was digested with the restriction endonucleases EcoRV and BglII, whose recognition sequences occur uniquely in the plasmid transplacement vector and the human cDNA insert, respec- tively. After electrophoretic separation of the digestion products and transfer to a nylon filter, as described under “Experimental Proce- dures,” the filter was hybridized to 32P-labeled full-length human cDNA for type 5 acid phosphatase (-10’ cpdpg). After stringent washing, the dried filter was exposed to Kodak X-Omat film. The panel is an radio- autograph in which the position of DNA markers of known size is also shown (in kilobases). Lane 1, DNA from uninfected cells; lune 2, DNA from Sf9 cells infected with recombinant AcNPV; lune 3, Sf9 cells in- fected with wild type AcNPV. B, Northern blot hybridization analysis of human type 5 acid phosphatase expression in insect cells. Total RNA (-10 pg) was extracted from Sf9 cells infected with recombinant bacu- lovirus as described under “Experimental Procedures” and separated by electrophoresis in formaldehyde-agarose gel before capillary transfer to a nylon filter. f i r washing, the filter was hybridized and radioauto- graphed as described inpunel A. The position of RNAmarkers of known size (Boehringer Mannheim) is also depicted in this panel. Lune 1, RNA from uninfected Sf9 cells; lune 2, RNA from Sf9 cells infected with recombinant AcNPV; lune 3, RNA from Sf9 cells infected with wild type AcNPV.
ured continuously for 10 s. In control experiments, bovine serum albu- min (Type V, Sigma) or saturated diferric human transferrin (Boeh- ringer Mannheim) replaced the recombinant human enzyme. The effects of ferrous and ferric iron chelators were determined by addition to the protein solution a t 4 “C 30 min before the peroxidation reaction was initiated. a,a’-bipyridyl was dissolved in ethanol; identical concen- trations of ethanol were added to control incubations in which the effects of ferrous iron chelation were examined.
RESULTS Production of Recombinant Human Purple Phosphatase-A
recombinant baculovirus was generated by replacement of the nonessential wild type polyhedrin gene with the full-length cDNA encoding human type 5 acid phosphatase. Homologous recombination with the wild type AcNPV baculovirus was me- diated by use of the plasmid transplacement vector PAC CL29. Homologous recombination between the wild type polyhedrin gene and the acid phosphatase cDNA together with the 5’- and 3”flanking regions of the polyhedrin gene in the transplace- ment vector was confirmed by digestion with two restriction endonucleases followed by Southern blot analysis of Sf9 ge- nomic DNA (Fig. lA). The vector contains a unique EcoRV restriction site and there is a unique recognition site for BgZII in the phosphatase cDNA sequence. The acid phosphatase cDNA hybridized to genomic fragments of the expected size derived from AcNPV-infected Sf9 cells. The foreign gene insert
was transcribed in the Sf9 insect host cells as a single RNA species of 1.5 kilobases as found in human tissues (11,321 (Fig. lB ).
Expression of Acid Phosphatase Activity-Culture medium obtained from Sf9 cells infected with recombinant AcNPV bacu- lovirus contained enzymatically active acid phosphatase that could be removed by binding to antibodies raised against por- cine uterofemn. Tartrate-resistant acid phosphatase was gen- erally detectable in the medium from 48 h after infection; the levels of activity, however, increased for several days.
Purification of Secreted Protein-Human type 5 acid phos- phatase was purified from serum-free medium by sequential chromatography using columns of S-Sepharose and Mono S at pH 8 (see Table I). The yield of recombinant protein in the culture medium could be enhanced by increasing the cell den- sity up to 1.2 x 106/ml, and subsequent purifications yielded up to 8 mg of purple phosphatase proteiditer of medium. The specific activity of the purified recombinant protein (100-200 pmol of p-nitrophenol releasedlmidmg at 37 “C) resembles that determined in enzyme samples purified independently from human osteoclastoma (41) and splenic tissue (46).
When the purified protein was analyzed by SDS-polyacryl- amide gel electrophoresis and staining with Coomassie Blue, two proteins of approximately 36 kDa were identified (Fig. 2). These polypeptides were converted to a single species of -34 kDa upon digestion with N-glycosidase F, indicating that they represented two glycoforms of a single polypeptide. Both gly- coproteins reacted specifically with uterofemn antibody as shown by immunoblotting analysis, but the expressed human phosphatase had a slightly lower molecular weight than utero- ferrin. This suggests that uteroferrin may be more highly gly- cosylated. The intracellular form of the human recombinant acid phosphatase was identified as a single protein species of approximately 38 kDa. The intracellular form of the recombi- nant protein was found not to react with uterofenin antibody and did not possess acid phosphatase activity (data not shown). Mass spectroscopy of the purified protein provided four esti- mates of the molecular weight (36276.20, 35580.71, 35415.80, and 36121.61), with a mean molecular weight determination of 35848.58.
Analysis of the Iron Center-Recombinant human type 5 acid phosphatase, purified in its active form from insect cell me- dium, contained 2 atoms of irodmolecule. The protein pos- sessed a striking pink-purple color in 0.1 M sodium acetate buffer at pH 5.5, giving rise to a spectroscopic absorbance maxi- mum at 537 nm. Reduction with ascorbic acid by addition to 0.015 M resulted in a slight change of color to dark purple in conjunction with a shift of maximum absorbance to 531 nm. The pH of the solution was unchanged.
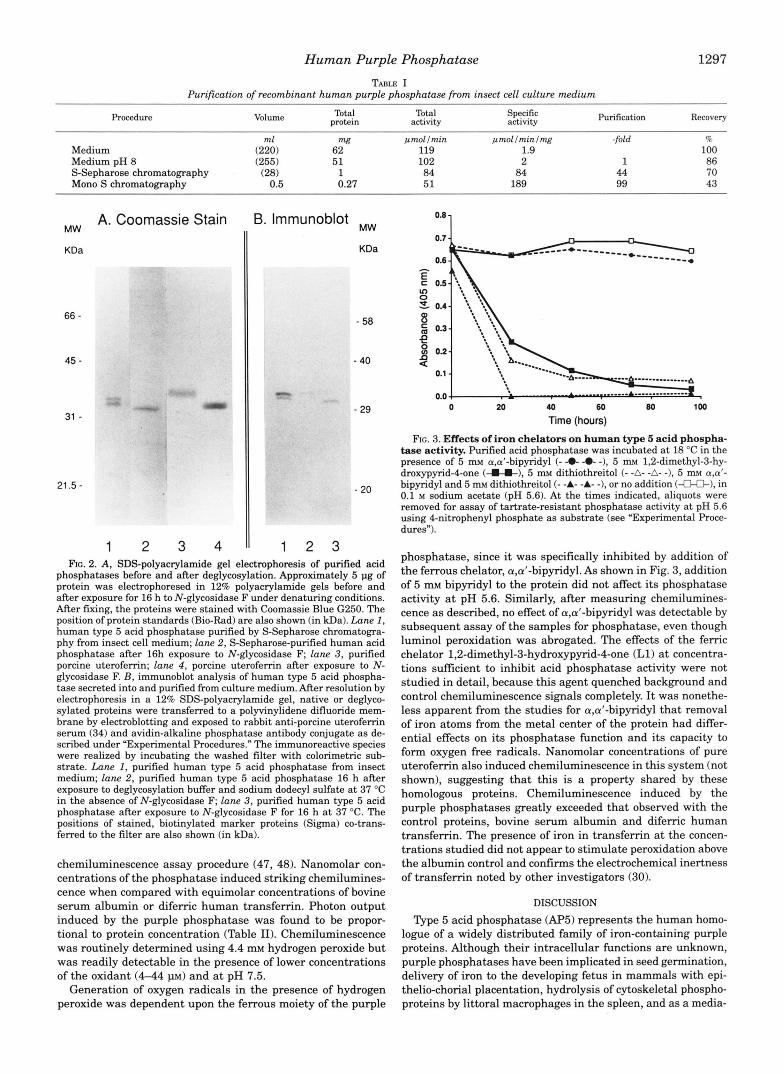
When human acid phosphatase was exposed to the ferrous chelator, a,a‘-bipyridyl(5 m ~ ) at room temperature for 5 days, phosphatase activity remained similar to that of a control sample maintained for the same period. Exposure to the ferric chelator, 1,2-dimethyl-3-hydroxypyrid-4-one inhibited acid phosphatase activity by more than 70% after 2 days’ exposure, as did reduction in the presence of dithiothreitol (Fig. 3). Incu- bations in the presence of dithiothreitol and a,a’-bipyridyl re- sulted in complete inhibition of acid phosphatase activity within a few hours.
Crystal Structure-Concentrated solutions of purified hu- man type 5 acid phosphatase (Fig. 4) formed dark monoclinic crystals that could be distinguished from the pale mother liq- uor after 6 weeks at room temperature (Fig. 5). The average length of the crystals was 50 pm.
Generation of Reactive Oxygen Species-The ability of human purple phosphatase to catalyze the peroxidation of luminol was examined in three independent experiments by means of the
Human Purple Phosphatase 1297 TABLE I
Purification of recombinant human purple phosphatase fiom insect cell culture medium
Procedure Volume protem ””I
actlwty Total Specific activity Purification Reeovery
ml w pmollmin prnollrninlmg -fold Medium (220) 62 119 1.9
9% 100
Medium pH 8 (255) 51 102 S-Sepharose chromatography (28) 1 84 Mono S chromatography 0.5 0.27 51
2 84
189
1 86 44 70 99 43
MW A. Coomassie Stain
KDa
66 -
45 -
31 -
21.5 -
B. lmmunoblot MW O.* 1
KDa
.58
40
29
20
1 2 3 4 1 1 1 2 3 FIG. 2. A, SDS-polyacrylamide gel electrophoresis of purified acid
phosphatases before and after deglycosylation. Approximately 5 pg of protein was electrophoresed in 12% polyacrylamide gels before and after exposure for 16 h to N-glycosidase F under denaturing conditions. After fixing, the proteins were stained with Coomassie Blue G250. The position of protein standards (Bio-Rad) are also shown (in m a ) . Lane 1, human type 5 acid phosphatase purified by S-Sepharose chromatogra- phy from insect cell medium; lane 2, S-Sepharose-purified human acid phosphatase after 16h exposure to N-glycosidase F; lune 3, purified porcine uterofemn; lune 4, porcine uterofemn after exposure to N- glycosidase F. B, immunoblot analysis of human type 5 acid phospha- tase secreted into and purified from culture medium. After resolution by electrophoresis in a 12% SDS-polyacrylamide gel, native or deglyco- sylated proteins were transferred to a polyvinylidene difluoride mem- brane by electroblotting and exposed to rabbit anti-porcine uterofemn serum (34) and avidin-alkaline phosphatase antibody conjugate as de- scribed under “Experimental Procedures.” The immunoreactive species were realized by incubating the washed filter with colorimetric sub- strate. Lane 1, purified human type 5 acid phosphatase from insect medium; lane 2, purified human type 5 acid phosphatase 16 h after exposure to deglycosylation buffer and sodium dodecyl sulfate a t 37 “C in the absence of N-glycosidase F; lune 3, purified human type 5 acid phosphatase after exposure to N-glycosidase F for 16 h a t 37 “C. The positions of stained, biotinylated marker proteins (Sigma) co-trans- ferred to the filter are also shown (in kDa).
chemiluminescence assay procedure (47, 48). Nanomolar con- centrations of the phosphatase induced striking chemilumines- cence when compared with equimolar concentrations of bovine serum albumin or difemc human transfemn. Photon output induced by the purple phosphatase was found to be propor- tional to protein concentration (Table 11). Chemiluminescence was routinely determined using 4.4 m~ hydrogen peroxide but was readily detectable in the presence of lower concentrations of the oxidant (444 p ~ ) and at pH 7.5.
Generation of oxygen radicals in the presence of hydrogen peroxide was dependent upon the ferrous moiety of the purple
0.7
0.6
0 20 40 60 80 100
Time (hours)
FIG. 3. Effects of iron chelators on human type 5 acid phospha- tase activity. Purified acid phosphatase was incubated a t 18 “C in the presence of 5 m~ a,d-bipyridyl (- -0- -0- -), 5 m~ 1,2-dimethyl-3-hy- droxypyrid-4-one ( I C ) , 5 m~ dithiothreitol (- -A- -A- -1, 5 rn a,a‘- bipyridyl and 5 m~ dithiothreitol (- -A- -A- -), or no addition (€LO-), in 0.1 M sodium acetate (pH 5.6). At the times indicated, aliquots were removed for assay of tartrate-resistant phosphatase activity a t pH 5.6 using 4-nitrophenyl phosphate as substrate (see “Experimental Proce- dures”).
phosphatase, since it was specifically inhibited by addition of the ferrous chelator, apt-bipyridyl. As shown in Fig. 3, addition of 5 mM bipyridyl to the protein did not affect its phosphatase activity a t pH 5.6. Similarly, after measuring chemilumines- cence as described, no effect of a,a’-bipyridyl was detectable by subsequent assay of the samples for phosphatase, even though luminol peroxidation was abrogated. The effects of the femc chelator 1,2-dimethyl-3-hydroxypyrid-4-one (Ll) at concentra- tions sufficient to inhibit acid phosphatase activity were not studied in detail, because this agent quenched background and control chemiluminescence signals completely. It was nonethe- less apparent from the studies for a,d-bipyridyl that removal of iron atoms from the metal center of the protein had differ- ential effects on its phosphatase function and its capacity to form oxygen free radicals. Nanomolar concentrations of pure uterofemn also induced chemiluminescence in this system (not shown), suggesting that this is a property shared by these homologous proteins. Chemiluminescence induced by the purple phosphatases greatly exceeded that observed with the control proteins, bovine serum albumin and difemc human transferrin. The presence of iron in transferrin at the concen- trations studied did not appear to stimulate peroxidation above the albumin control and confirms the electrochemical inertness of transferrin noted by other investigators (30).
DISCUSSION
Type 5 acid phosphatase (AP5) represents the human homo- logue of a widely distributed family of iron-containing purple proteins. Although their intracellular functions are unknown, purple phosphatases have been implicated in seed germination, delivery of iron to the developing fetus in mammals with epi- thelio-chorial placentation, hydrolysis of cytoskeletal phospho- proteins by littoral macrophages in the spleen, and as a media-

Human Purple Phosphatase
I
tor of bone resorption by oeteoclasta (3,10,19,49-61). In man, a single A P S gene has been localized to chromosome 19p (32); this cOrregpOnds to the mapping position of the murine gene on a syntenic region of mouBe chromosome 9 (52). Expression of
IIY X 10~110 8 Bovineeenunalbumin 20 32 134
10 28 122 6 24 102
€inman difarric taaneferrin 20 26 123 10 27 111 6 28 108
Human purple phoaphataae 20 403 197 10 214 1-40 6 89 122
theAP6 gene in human h u e s is highly selective and accounts for tartrate-resistant acid phosphatase activity characteristic of certain differentiated cells of monocyte-macrophage lineage, including the dast (11). Although several functiom have been pro@ for the human purple phphatase in speciabd cells hitherto, owing to its low abundance, insuflicient amounta have been purified tivm whole tissues to permit a detailed examination of ita molecular structure and catalytic pmperties. We describe here the use of a baculovirw system to induca high level expression of human A P S secreted by Sf9 insect cells.
Auified human A P S protein expressed by Sf9 cells possesd t a r t r ab -d tan t acid phosphatase activity cloeely similar to that measured using enzyme isolated from human aourtxs (6, 41,461. The secreted protein has a molecular maas of 36 lrDa and contains 2 iron atoms/molecule as shown previously with the enzyme purified by Ketcham et al. (46) from hairy cell leukemic spleen. k similar glycofonns of the expresd re- combinant protein were converted to a single protein species of -34 kDa after treatment with N-glycosidase P; this corm- sponds to the predicted molecular mass of the native polypep- tide after proteolytic processing of the 2.4-kDa lysoeomal signal sequence (32,631 and of mature polypeptide isolated from hu- man spleen by Roberts and colleagues (46). In this respect, when expressed by insect cells, human AP6 behaves as a se- creted lysosomal protein like its porcine homologue, uterofer- M . Biosynthetic labeling of the carbohydrate eubetituents of porcine u t e r o f d eecreted by cultured endometrial explants shows that it contains a masked mannose &phosphate lyso- somal recognition ligand (64) .
The purple phosphatases are members of a m u l t i h c t i d claee ofdi-iron-oxo proteins that are of diverse origin (19). This class of metalloproteins posseesea a eommoll structural feature at ita active site, characterized by strong anti-ferromagnetic coupling between the iron centers in their bi-femc form and unique EPR signals in the mixed valence forms. However, with the exception of the residues that serve as metal-binding li- gands, they show marked divergence in their amino acid 88- quences (17,251. Human purple phosphatase, expremed in ita active form by insect cells, contains catalytically BB1)811tiB1 ferric iron and was rapidly inhibited by treatment with the reducing agent dithiothreitol. Exposure to the ferrous chelator, a,a'-bi- pyridyl had little effect on phosphatase activity but treatment wi& the femc chelator, 1,2-dimethyl-3-hydroxypyrid4one, rapidly inhibited enzymatic activity. The effecte of combined treatment with dithiobitol and a,a'-bipyridyl likewise mark- edly reduced phosphatase activity, presumably as a result of the reduction of an essential ferric atam that was removed tivm the catalytic center by the chelator. We believe that the ferric iron is also essential for protein stability, since it did not prove

Human Purple Phosphatase 1299
possible to restore enzymatic activity on re-addition of ferric iron (not shown). Recently, the analysis of the murine AP5 promoter has demonstrated the presence of steroid hormone and 1,25-dihydroxy vitamin D response elements similar to those found in the human osteocalcin gene. The 5"untrans- lated region of the murine AP5 gene is active in mediating transcription in macrophage lines (55). Preliminary evidence indicates also that transcription of the acid phosphatase is also subject to control by cellular iron; expression of acid phospha- tase-constructs is regulated by iron-responsive promoter se- quences in the 5"untranslated region of the human AP5 gene (56).
A unifying property of di-iron-oxo proteins is their participa- tion in reactions involving the transfer of single electrons; in deoxyhemerythrin, electrons from ferrous irons are transferred to molecular di-oxygen in a reversible process that allows the protein to function as an oxygen carrier (17); in methane monooxygenase hydroxylase, ribonucleotide reductase, stearo- yl-acyl carrier protein Ag-desaturase, and other oxygenases, 02-dependent cleavage of unactivated C-H bonds appears to depend on high valent iron-oxo catalytic intermediates (25). Although the amino acid sequence of the human purple phos- phatase does not contain the paired motif, -(DE)-E-X-R-H-, analogous to the known iron-binding sites of ribonucleotide reductase and methane monooxygenase hydrolase that are ca- pable of reaction with molecular di-oxygen, nonetheless we speculate that the human phosphatase participates in electron transfer reactions involving oxygen species.
Addition of hydrogen peroxide to dilute solutions of the pro- tein in the presence of luminol induced striking chemilumines- cence, indicating the formation of electronically excited mol- ecules. This chemiluminescence was inhibited by chelating ferrous iron from the purple phosphatase.
Chemiluminescence has been used as direct measure of the formation of free radicals by cells undergoing phagocytosis dur- ing microbial killing (47). In this report we show that purified human purple phosphatase serves as a catalyst for the peroxi- dation of aminophthalate leading to the formation of superox- ide ions, hydroxyl ions, or the HO-Fe2+-OOH moiety (30). The peroxidation of aminophthalate probably involves the forma- tion of organic radicals that can produce superoxide in the presence of oxygen. Further free radicals can react with super- oxides, leading to the generation of excited aminophthalate dianions that cause photon emission on decay to the ground state (47). That the purple phosphatase is active in the cataly- sis of luminol peroxidation, a process inhibited by ferrous ion chelators, indicates that its iron center is accessible for ferroxi- dation and may participate in biological reactions that generate destructive free radicals. The constraints imposed by the re- quirements of the luminol reaction prevented the determina- tion of chemiluminescence under acid conditions, but in the pH range 7-9 the peroxidative activity of purple phosphatase in inducing chemiluminescence exceeded by more than an order of magnitude that of human transferrin, even though the latter also contains 2 iron atoms/molecule. Although it is likely that the induction of chemiluminescence results from formation of hydroxyl or peroxy radicals as products of the Fenton reaction catalyzed by ferrous iron at the active center (30), we have not obtained formal proof of decomposition to these free radical species.
We propose that induction of iron-dependent chemilumines- cence by the human purple phosphatase is significant for our understanding of microbial killing by mature activated macro- phages and osteoclast function. As mononuclear phagocytes mature, myeloperoxidase activity is lost so that oxygen-depend- ent killing is accomplished by reactive species such as the hy- droxyl radical that can be generated directly from superoxide
and hydrogen peroxide (57). Recent studies have shown that low concentrations of hydrogen peroxide stimulate osteoclast activity (31, 58, 59). Osteoclasts produce free radical interme- diates that reduce nitro blue tetrazolium (58), and altered bone resorptive activity in response to calcitonin or interleukin-1 is accompanied by parallel changes in free radical-generating power (31). Since human type 5 acid phosphatase expression is restricted to activated macrophages, and is secreted at the mar- gin of the ruffled border in the resorptive vacuole of the osteo- clast (60), we believe that it has the appropriate location and properties for the generation of oxygen-derived free radicals that participate in microbial killing and bone resorption. That this may occur in the appropriate cellular microenvironment, remains to be demonstrated.
The possibility that the human purple phosphatase is a mul- tifunctional protein also remains. The enzyme possesses re- gions homologous to mammalian phosphoprotein phosphatases (61) and, as in the pig uterus, may serve as a labile source of iron. We report here the purification of substantial quantities of this unusual metalloprotein in an advance on its systematic characterization. Expression of the human enzyme by means of the baculovirus vector in insect cells renders possible the pro- duction of sufficient purple phosphatase for large scale crystal- lization and determination of its three-dimensional structure at atomic resolution.
Acknowledgments-Joan Grantham provided expert secretarial as- sistance. We thank Drs. Janet Dewdney, Robert Imrie, and Sherin Ab- del-Meguid for encouragement and generous discussion. Dr. R. Michael Roberts kindly supplied pure porcine uteroferrin for investigation. Mass spectroscopy was carried out by Paul Skelton, Department of Chemis- try, University of Cambridge. We are grateful to Drs. Nicholas Cross and Ian Cassady for sharing information about the murine acid phos- phatase gene in advance of publication.
1
2,
4 3
5 6
8. 7.
10. 9.
11.
12.
13.
14.
15.
16.
17. 18.
19.
20.
21.
22.
23.
24.
25.
REFERENCES Li, C. Y., Yam, L. T., and Lam, K. W. (1970) J. Histochem. Cytochem. 18,
Davis, J. C., and Averill, B. A. (1982) Proc. Natl. Acud. Sci. U. S. A. 79,4623-
Roberts, R. M., Raub, T. J., and Bazer, F. W. (1986) Fed. Proc. 45, 2513-2518 Roberts, R. M., and Bazer, F. W. (1984) Bioessays 1, %I1 Burstone, M. S. (1959) J. Histochem. Cytochem. 7 , 3 9 4 1
Yam, L. T., Li, C. Y., and Finkel, H. E. (1972)Arch. Intern. Med. 130,24&256 Robinson, D. B., and Glew, R. H. (1980) Clin. Chem. 26,371-382
Choy, F. Y. M. (1985)Am. J. Med. Genet. 21,519-528 Yam, L. T. (1974)Am. J. Med. 56,604-616 Lau, K.-H. W., Onishi, T., Wergedal, J. E., Singer, F. R., and Baylink, D. J.
(1987) Clin. Chem. 33,458462 Bevilacqua, M., Lord, D. K., Cross, N. C. P., Whitaker, K. B., Moss, D. W., and
Cox, T. M. (1991) Mol. Biol. Med. 8, 135-140 Gignac, S. M., Buschle, M., Roberts, R. M., Pettit, G. R., Hornrand, A. V., and
Drexler, H. G. (1990) Iauk. Lymphoma 3, 19-29 Schlosnagle, D. C., Sander, E. G . Bazer, F. W., and Roberts, R. M. (1976) J. Biol.
Chem. 251,468&4685 Antanaitis, B. C., Aisen, P., Lilienthal, H. R., Roberts, R. M., and Bazer, F. M.
(1980) J. Biol. Chem. 266, 11204-11209 Antanaitis, B. C., Aisen, P., and Lilienthal, H. R. (1983) J. Biol. Chem. 268,
316&3172 Antanaitis, B., and Aisen, P. (1983) in Structure and Function of Iron Storage
and Dunsport Proteins (Urushizaki, I., Listowsky, I., and Aisen, P., eds) pp. 503-511, Elsevier Science Publishers B. V., Amsterdam
Antanaitis, B. C., and Aisen, P. (1983b) in Advances in Znorganic Biochemistry Que, L., and Scarrow, R. C. (1988) Am. Chem. Soc. Symp. Ser: 372, 152-178
(Theil, E. C., Eichhorn, G. L., and Marzilli, L., eds) Vol. 7, pp. 111-136,
Que, L., and True, A. E. (1990) in Progress in Znorganic Chemistry: Bioinor- Elsevier Science Publishing Co., h e . , New York
ganic Chemistry (Lippard, S. J., ed) Vol. 38, pp. 97-200, John Wiley & Sons, New York
Fox, B. G., Borneman, J. G., Wackett, L. P., and Lipscomb, J. D. (1990) Bio- chemistry 2 9 , 6 4 1 9 4 2 7
Stenkamp, R. E., Sieker. L. C.. Jensen. L. H.. Sanders-Loehr. J. (1981) Nature
901-910
4627
291,263-264
96-102
. .
Sjoberg, B.-M., Loehr, T. M., and Sanders-Loehr, J. (1982) Biochemistry 21,
Yen, K.-M., Karl, M. R., Blatt, L. M. Simon, M. J., Winter, R. B., Fausset, P. R., Lu, H. S., Harcourt, A. A., and Chen, K. K. (1991) J. Bacteriol. 173, 5315-
Nordlund, I., Powlowski, J., and Shingler, V. (1990) J. Bacteriol. 172, 6826- 5327
Fox, B. G., Shanklin, J., Somerville, C., and Miinck, E. (1993) Proc. Nad. Acad. 6833

1300 Human Purple Phosphatase
26. Cardy, D. L. N., Laidler, V., Salmond, G. P. C., and Murrell, J. C. (1991) Mol.
27. Osdoby, P., Krukowski, M., Oursler, M. J., and Salino-Hugg, T. (1987) Bioes-
28. Murray, H. W., and Cohn, 2. A. (1980) J. Inuest. Dermatol. 74, 285287 29. Nathan, C. F. (1987) J. Clin. Inuest. 70, 319-326 30. Halliwell, B., and Gutteridge, J. M. C. (1989) Free Radicals in Biology and
31. Garrett, I. R., Boyce, B. F., Oreffo, R. 0. C., Bonewald, L., Poser, J., and Mundy, Medicine, 2nd Ed., Oxford University Press, Oxford
32. Lord, D. K, Cross, N. C. P., Bevilacqua, M. A., Rider, S . H., Gorman, P. A,, G. R. (1990) J. Clin. Inuest. 85,632-639
Groves, A. V., Moss, D. W., Sheer, D., and Cox, T. M. (1990) Eur J. Biochem. 189,287-293
33. Lee, C. C., Wu, X., Gibbs, R. A,, Cook, R. G., Muzny, D. M., and Caskey, C . T.
34. Echetebu, Z. O., Cox, T. M., and Moss, D. W. (1987) Clin. Chem. 33.18321836 35. Whitaker, K B., Cox, T. M., and Moss, D. W. (1989) Clin. Chem. 35,86439 36. Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. D., Smith,
J. A., and Struhl, K (eds) (1990) Current Protocols in Molecular Biology,
37. Maniatis, T., Fritach, E. F., and Sambrook, J. (1989) Molecular Cloning: A John Wiley & Sons, New York
Labomtory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
Sci. U. S. A 90,2486-2490
Microbiol. 5, 33-42
says 7, 30-34
(1988) Science 239,1288-1291
38. Summers, M. D., and Smith, G. E. (1987) %. Agric. Exp. Stn. Bull. 1656
40. Chomczynski, P., and Sacchi, N. (1987) Anal. Biochem. 162, 156-159 39. Laemmli, U. K (1970) Nature 227,680-685
41. Hayman, A. R., Warburton, M. J., Pringle, J. A. S. , Coles, B., and Chambers, T.
42. Bradford, M. M. (1976) Anal. Biochem. 72,248-254 43. Udenfriend, S., Stein, S. , Bohlen, P., Dairman, W., Leingruhei, W., and Weigel,
44. Landers, J. W., and Zak, B. (1958)Am. J. Clin. Pathol. 29, 590-592
J. (1989) Biochern. J . 261,601-609
M. (1972) Science 178,871-872
45. McPherson, A. (1982)Prepamtion and Analysis ofProtein Crystals, John Wiley
46. Ketcham, C. M., Baumbach, G. A., Bazer, F. W., and Roberts, R. M. (1985) J.
47. Henley, R., and Worwood, M. (1991) Arch. Biochem. Biophys. 286.238-243 48. McPhail, L. C., DeChatelet, L. R., and Johnston, R. B., Jr. (1979) J. Clin.
49. Beck, J. L., McConachie, L. A,, Summors, A. C., Arnold, W. N., De Jersey, J., Inuest. 63,648-655
50. Moonga, B. S. , Moss, D. W., Patchell, A,, and Zaidi, M. (1990) J. Physiol. 429, and Zerner, B. (1986) Biochim. Biophys. Acta 869,6148
29-45 51. Schindelmeiser, J., Munstermann, D., and Witzel, H. (1987) Histochemistry
52. Grimes, R., Reddy, S . V., Leach, R. J., Scareez, T., Rmdman, G. D., Sakaguchi, 87.13-19
53. Ketcham, C. M., Roberts, R. M., Simmen, R. C. M., and Nick, H. S . (1989) J. A. Y., Lalley, P. A., and Windle, J. J. (1993) Gemmics 15, 421422
54. Baumbach, G . A,, Saunders, P. T. K, Bazer, E W., and Roberts, R. M. (1984) Biol. Chem. 264,557563
55. Cassady, A. I., King, A. G., Cross, N. C. P., and H u e , D. A. (1993) Gene Proc. Natl. Acad. Sci. U. 5'. A. 81, 298L2989
56. Boldt, D. H., Reddy, S . V. J., Craig, F., Alcantara, O., and Roodman, G. D. (1992) (Amst. ), in press
57. Johnston, R. B. Jr., Gcdzik, C. A., and Cohn, Z. A. (1978) J. Exp. Med. 148, Blood 80, Suppl. 1,182a
58. Key, L. L. Jr., Reis, W. L., Taylor, R. G., Hays, B. D., and Pitzer, B. R. (1990) 115-127
59. Bax, B. E., Alam, A. S . M. T., Baneji, B., Bax, C. M. R., Bevis, P. J. R., Stevens, Bone 11, 115-119
C. R., Moonga, B. S. , Blake, D. R., and Zaidi, M. (1992) Biochem. Biophys. Res. Commun. 183, 1153-1158
& Sons, New York
Biol. Chem. 260,5768-5776
60. Miller, S . C. (1985) Calcif lIlsslre Int. 37, 526529 61. Vincent, J. B., and Averill, B. A. (1990) FEBS Lett. 263, 265-268