of diabetes insipidus in brattleboro rats
TRANSCRIPT

Although the ability ofMetRS to repress theswitched mRNA leader is clearly demonstrat-ed, we believe that the repression can still beamplified. The possibility remains that nucle-otides not present in the mRNA leader par-ticipate in tRNAMer identity. It is also possi-ble that nucleotides in the mRNA leaderspecifically prevent recognition by MetRS.The present genetic system permits an easyselection for mutants acting in cis that aremore sensitive to repression byMetRS. Theseselections could yield nudeotide changes thatwould give us information about howMetRSrecognizes its tRNA isoacceptors.Our results indicate that an excess ofMetRS
somewhat represses wild-tpe thrS expression.This repression is seen in the decrease ofexpres-sion of thrS-lacZ fusions (Table 1) and onprotein immunoblots (Fig. 2B), where theband corresponding to the endogenous ThrRSis weaker in the presenceofpNAV7 than in thepresence of pUC12. Because this effect is lostwith changes other than the CGU -- CAUchange, it appears that the wild-type thirSmRNA leader is weakly but specifically recog-nized by MetRS. Thus, the thrS mRNA leadermay be recognized with lower affinity byMetRS and possibly other synthetases. Thebiological relevance ofthis cross talk remains tobe determined.The present data, coupled with genetic
(10) and biochemical (5) analyses of the thrSmRNA leader-ThrRS complex, show thatthe synthetase recognizes both RNAs by asimilar mechanism. Because the three-dimen-sional structure ofthe synthetase-tRNA inter-action is under analysis, this system shouldprovide a mechanism for studying the three-dimensional structure of an mRNA leadercomplexed with a translational repressor. Theability to change the regulatory specificity ofthrS by changing the mRNA leader maymake three-dimensional analysis more facile.The specificity of the mRNA leader could bechanged to an E. coli synthetase for which thethree-dimensional interaction with its cog-nate tRNA is available (17). Also, the abilityto change the regulation specificity of thrSprovides the best indication of mimicry be-tween a regulatory site on an mRNA andanother site on a stable RNA, to which asingle control protein binds to both in orderto perform a specific cellular activity.
REFERENCES AND NOTES
1. P. Lestienne, J. A. Plumbridge, M. Grunberg-Man-ago, S. Blanquet, J. Biol. Chem. 259, 5232 (1984).
2. M. Springer et al., J. Mol. Biol. 185, 93 (1985).3. M. Springer, M. Graffe, J. S. Butler, M. Grunberg-
Manago, Proc. Nad. Acad. Sci. U.SA. 83, 4384(1986).
4. H. Moine et al., ibid. 85, 7892 (1988).5. H. Moine et al., J. Mol. Biol. 216, 299 (1990).6. L. Lindahl and J. M. Zengel, Annu. Rev. Genet. 20,
297 (1986).7. J. Normanly and J. Abelson, Annu. Rev. Biochem.
58, 1029 (1989).
8. M. Springer et al., unpublished data.9. Y. Komnine, T. Adachi, H. Inokuchi, H. Ozeki, J.
Mol. Biol. 212, 579 (1990).10. M. Springer, M. Graffe, J. Dondon, M. Grunberg-
Manago, EMBOJ. 8, 2417 (1989).11. L. H. Schulman and H. Pelka, Nuclekic Acids Res. 18,
285 (1990).12. W. H. McClain and H. B. Nicholas, Jr., J. Mol.
Biol. 194, 635 (1987).13. P. H. Hirel et al., Biochimie 70, 773 (1988).14. We made the diffirent bacteriophages (with the excep-
tion of XMA20-10-XII-25-ML2) used in Table 1 bycloning the Hind lf-Eco RI insert of M13mp8A20-10-XII-25 (or its derivatives) between the left arm ofXSKS107 (to its Hind Ill site) and the right arm ofXgt4 from its Eco RI site, as described (2). This cloningreconstitutes the dS-IacZ hybrid in its integrity. Thephage XSKS107 is irmn21 and nin5, and we made it bycloning the Hind IEl-SstIfragmentofpSKS107 [S. K.Shapiro, J. Chou, F. V. Richaud, M. J. Casadaban,Gene 25,71 (1983)] that carries the 5' tenminal part oflacZ without a translation initiation site between the leftarm of Xnav8-5 (to its Sst I site) and the right arm ofANM540 (from its Hind m site) as described (2).M13mp8A20-10-XII-25 is an Ml3mp8 derivative car-rying an insert ~edxting from the Pst I site 1194 bp infront of thrS to a recontited Hindm site 53 bp onthe 3' side oftheATG ofthrS. The insert also carries theXII-25 mutation (Fig. 1) that was introduced witholigonulotd sitedirected mutagenesis, as was donefor all the other mutations of the thrS mRNA leader.The phage XMA20-10-Xf-25-ML2 was selected froma XMA20-10-XII-25 lysogen of E. coli IBPC5421 asconferring a Lacphenotype (3, 8). Strain IBPC5421was lysog d with a single copy ofthe recombinantphages. The F'lacPTnlO episome was introduced inthe single lysogens by conjugation. The male lysogenswere then transformed with pUC12, pUB4 (2), orpNAV7 (13). The strains were grown in MOPS-glucose medium, supplemented with all amino acids
(18), tetracycline at 10 ~g/ml, and isopropyl-1-thio-- galactoside (IPTG) at 10-3 M. We added ampi-
cillin at a concentration of 100 [Lg/ml at 1-hourintervals during growth to select for the presence ofthe plasmids.
15. High expression from the fusions mutated in themRNA leader equivalent of the anticodon is due tothe inability of the endogenous ThrRS to repressp-galactosidase synthesis from the thrS-lacZ fusions.
16. Low production ofThrRS by pUB4 is probably dueto the repression of thrS expression from the lacpromoter in the presence of glucose or the presenceof the F'lacPTnlO episome. The episome somewhatrepresses thrS expression even in the presence ofIPTG. The F'lacJPTnWO is necessary because pNAV7is lethal to the cells when MetRS is expressed fromboth the lac and its own promoter but not whenexpressed only from the latter.
17. M. A. Rould, J. J. Perona, D. S611, T. A. Steitz,Science 246, 1135 (1989).
18. F. C. Neidhardt, P. L. Bloch, S. Pedersen, S. Reeh,J. Bacteriol. 129, 378 (1977).
19. J. H. Miller, Experiments in Molecular Genetics (ColdSpring Harbor Laboratory, Cold Spring Harbor,NY, 1972).
20. J. S. Butler, M. Springer, J. Dondon, M. Grunberg-Manago, J. Bacteriol. 165, 198 (1986).
21. This work is dedicated to the memory ofG. Fayat. Wethank G. Fayat and C. L. Olsson for discussions; Y.Mechulam for the pNAV7; C. L. Olsson, E. N.Brody, M. Nomura, and P. Shimmel for reading themanuscript; andM. Grunberg-Manago and J. P. Ebelfor their support and interest. Supported by grantsfrom the CNRS (UA 1139 and LP6201), INSERM(891017 to M.S.), the European Economic Commu-nity [SC1*/0194-C(AM) to M. Grunberg-Manago],and the Fondation pour la Recherche Medicale (toM.Grunberg-Manago).
8 July 1991; accepted 10 October 1991
Reversal of Diabetes Insipidus in Brattleboro Rats:Intrahypothalamic Injection of Vasopressin mRNAGUSTAV F. JIRIKOWSKI,* PIETRO PAOLO SANNA,DOMINIQUE MACIEJEWSKI-LENOIR, FLOYD E. BLOOM
Messenger RNAs occur within the axons of magnocellular hypothalamic neuronsknown to secrete oxytocin and vasopressin. In Brattleboro rats, which have a geneticmutation that renders them incapable ofvasopressin expression and secretion and thuscauses diabetes insipidus, injection into the hypothalamus of purified mRNAs fromnormal rat hypothalami or of synthetic copies of the vasopressin mRNA leads toselective uptake, retrograde transport, and expression of vasopressin exclusively in themagnocellular neurons. Temporary reversal of their diabetes insipidus (for up to 5days) can be observed within hours of the injection. Intra-axonal mRNAs mayrepresent an additional category of chemical signals for neurons.
T HE MRNAS ENCODING THE HOR-
mones arginine vasopressin (AVP)and oxytocin (OT) are present in
axons of the hypothalamoneurohypophysialtract (1-3). Oxytocin mRNA has been local-ized by in situ hybridization and electronmicroscopy in large granular vesicles of ax-onal varicosities in the lateral hypothalamus,the median eminence, and the posteriorpituitary (2). Furthermore, the amounts of
Department of Neuropharmacology, The Scripps Re-search Institute, La Jolla, CA 92037.
*To whom correspondence should be addressed.
996
OT mRNA in the hypothalamoneuro-hypophysial tract change with fimctionaldemand on the neurons (4).The presence of a neuropeptide mRNA in
vesicles and evidence for its axonal transport(2) suggest that this mRNA could providean intraneuronal or intercellular signal. Be-cause of the low activity of brain ribonucle-ase (RNase) (5), we were able to investigatethe metabolic fate of exogenous AVPmRNA injected into the axonal fields ofthese magnocellular neurons. To do so, weused the Brattleboro rat, which has a singlebase deletion in exon B of the propresso-
SCIENCE, VOL. 255
on January 31, 2021
http://science.sciencemag.org/
Dow
nloaded from

Fig. 1. (A) Vasopressin immunostaining of the.PVN of a Brattleboro rat, 2 hours after unilateralinjection of 50 ng of synthetic sense RNA coding ,for AVP. Rats were anesthetized with chloral r -Ihydrate (5 mg per 100 g of body weight) andkilled by cardiac perfusion with 4% paraformal--;dehyde in phosphate-buffered saline (PBS) for 20 2;min at 40C. Hypothalami were removed and te7sectioned on a Vibratome (Oxford Instruments) Jyinto serial frontal sections (50 pAm thick). Sectionswere washed in PBS and immunostained by the .. ,peroxidase-antiperoxidase method (16) with a rab-bit anti-AVP-serum (Chemicon), which had been -,preabsorbed with synthetic OT (Sigma). Immuno-precipitates were made visible with diaminobenzi- j '4 k .dine and hydrogen peroxide. For control purposes, Asome sections were stained with antisera to ACTH and to P-endorphin (both antisera obtained fromChemicon). Scale bar = 100 pum. (B) Vibratome section of the PVN of a Brattleboro rat injected withisotonic saline, devoid of vasopressin immunostaining. Scale bar = 100 pum. (C) The posterior lobe of thepituitary of a Brattleboro rat injected with total hypothalamic RNA. Scale bar = 10 pgm.
physin gene (6). This mutation prevents thetranslation of AVP in homozygotes andleads to chronic diabetes insipidus.
In preliminary experiments, Brattlebororats received injections into the lateral hypo-thalamus (7) of 5 Kug of total cytoplasmicRNA prepared from hypothalami of normalSprague-Dawley rats. Northern (RNA) blotanalysis of this total mRNA pool confirmedthe presence ofAVP mRNA as expected (8).We found that 18 hours after injection withthis preparation Brattleboro rats exhibitedcytoplasmic AVP immunoreactivitv withinneurons of the magnocellular hypothalamicnuclei (Fig. 1 and Table 1).
In subsequent experiments, Brattlebororats were injected with 50 ng of a syntheticsense-strand 35S-labeled AVP RNA, with a35S-labeled AVP antisense RNA (9), or withunlabeled AVP RNA. In confirmation of theimmunoreactivity changes observed after thetotal mRNA injections, magnocellular peri-karya of the supraoptic (SON) and paraven-tricular nuclei (PVN) selectively showed in-corporation of radioactivity within 2 hours ofinjection with sense-strand RNA. In fourrepetitions of this experiment (each groupcontaining three rats) about 25% (±+7%) ofthe magnocellular neurons in the SONshowed 35S-labeling, as did approximately10% (+4%) of the neurons in the PVN (Fig.2A). The density of silver grains per cellappeared to be higher in the SON than in the
PVN. After these sense-strand injections, ac-cumulation of radioactivity was also observedin the median eminence and the posteriorlobe (Fig. 2B), regions known to contain theaxons of the SON and PVN neurons. Ratsinjected with comparable amounts of thelabeled antisense AVP RNA showed radioac-tivity only in the ventricles and blood vessels,whereas hypothalamic perikarya did not accu-mulate radioactivity. The median eminenceand the posterior lobe were also unlabeled inthese controls (Fig. 2C).
In contrast, 2 hours after injection of syn-thetic unlabeled AVP RNA (the earliest timeexamined), immunoreactive AVP was ob-served in a fraction of the magnocellularneurons of the PVN (Fig. 1) and the SON;immunostaining was more pronounced in thePVN on the ipsilateral side of the injection,and AVP-immunoreactive penikarya werecompletely absent in the SON contralateral tothe injection site. Furthermore, AVP-immu-nostained nerve endings in the posterior lobeappeared to be less densely distributed thanstructures visible in autoradiograms obtainedfrom this organ after injection of [35S]AVPRNA into the hypothalamoneurohypophys-ial tract. This may indicate that the fractionof nerve terminals in the posterior lobe thatcontained radioactive RNA was greater thanthe number of nerve terminals containingimmunoreactive, translated AVP.To determine whether the injection of syn-
thetic AVP RNA or total hypothalamic RNAresulted in functional signals, we took urinesamples from Brattleboro rats before andafter the intracerebral injections. In normalrats urine osmolarity is well above 1200 mos-mol. The urinary osmolarity of untreatedhomozygous Brattleboro rats, however, wasin all cases below 200 mosmol throughoutthe time observed. Brattleboro rats treatedwith total cytoplasmic mRNA as well as withsynthetic RNA showed increased urine os-molarity within 2 hours after the injection,declining to control levels after 5 days (Fig.3), whereas ones injected with isotonic saline,antisense AVP RNA, or cytoplasmic mRNAtaken from normal rat hindbrain did notshow this effect. Moreover, the ability ofmicroinjected hypothalamic mRNA or syn-thetic AVP RNA to raise urine osmolaritywas lost when these materials had been di-gested with RNase (10).
Radioimmunoassays (RIAs) of homoge-nized hypothalamic nuclei (11) showed clearincreases in AVP levels upon injections ofsynthetic AVP RNA (Table 1). Animals in-jected with 50 ng of synthetic frill-lengthsense AVP RNA (9) had a distribution of
Table 1. Amounts of immunoreactive vasopressin measured by RIA (9) in homogenates of hypo-thalamic regions from Brattleboro rats 18 hours after unilateral injection with total hypothalamicRNA. Vasopressin concentration was below detection limit in animals that had been injected withisotonic saline.
AVP per milligram of protein (pg)
LocationPVN SON Median Posterioreminence lobe
Injected brain side 410 (+ 75) 173 (+ 54) 133 (-+- 26) 340 (-+- 87)Contralateral side 120 (+ 25) 67 (+ 12)
Fig. 2. Autoradiograms ofPVN and posterior lobeof a Brattleboro rat injected with [35S]AVP RNA.Hypothalami and pituitaries of animals, perfusedwith 4% paraformaldehyde 2 hours after injectionswith the labeled probes, were rapidly frozen andsectioned into 10-pxm cryostat sections. These sec-tions were processed for autoradiography withIlford G5 emulsion. (A) Magnocellular perikarya inthe PVN. (B) Dark-field micrograph of the poste-rior lobe. (C) Dark-field micrograph of the poste-rior lobe of a rat injected with the 35S-labeledantisense probe. Scale bar = 10 ptm.
21 FEBRUARY 1992 R-EPORTS 997
on January 31, 2021
http://science.sciencemag.org/
Dow
nloaded from
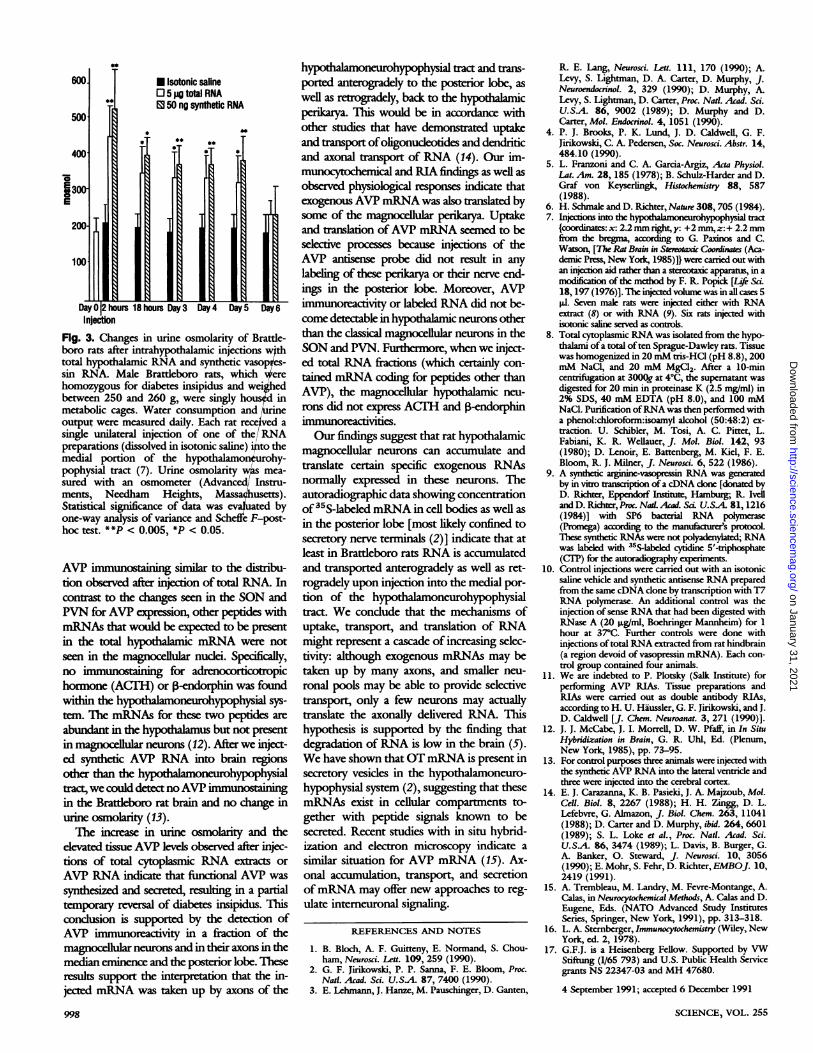
hours 18 hours Day 3 Day 4 Day 5 Day6
Fig. 3. Changes in urine osmolarity of Brattle-boro rats after intrahypothalamic injections wthtotal hypothalamic RNA and synthetic vasop es-
sin RNA. Male Brattleboro rats, which erehomozygous for diabetes insipidus and weigedbetween 250 and 260 g, were singly ho in
metabolic cages. Water consumption and e
output were measured daily. Each rat received a
single unilateral injection of one of the/RNApreparations (dissolved in isotonic saline) to themedial portion of the hypothalamon urohy-pophysial tract (7). Urine osmolaritywls mea-
sured with an osmometer (Advanced/ Instru-ments, Needham Heights, Massachusetts).Statistical significance of data was evauated byone-way analysis of variance and Scheffe F-post-
hoc test. **P < 0.005, *P < 0.05.
AVP immunostaining similar to the distribu-tion observed after injection of total RNA. Incontrast to the changes seen in the SON andPVN for AVP expression, other peptides withmRNAs that would be expected to be presentin the total hypothalamic mRNA were notseen in the magnocellular nudei. Specifically,no immunostaining for adrenocorticotropichormone (ACITH) or I3-endorphin was foundwithin the hypothalamoneurohypophysial sys-tem. The mRNAs for these two peptides are
abundant in the hypothalamus but not presentin magnocellular neurons (12). After we inject-ed synthetic AVP RNA into brain regionsother than the h la phal
tract, we could detect noAVP immunostainingin the Brattleboro rat brain and no change inurine osmolarity (13).The increase in urine osmolarity and the
elevated tissue AVP levels observed after injec-tions of total cytoplasmic RNA extracts or
AVP RNA indicate that fiuctional AVP was
synthesized and secreted, resulting in a partialtemporary reversal of diabetes insipidus. Thisconcusion is supported by the detection ofAVP immunoreactivity in a fraction of themagnocellular neurons and in their axons in themedian eminence and the posterior lobe. Theseresults support the interpretation that the in-jected mRNA was taken up by axons of the
998
hypothalamoneurohypophysial tract and trans-ported anterogradely to the posterior lobe, aswell as retrogradely, back to the hypothalamicperikarya. This would be in accordance withother studies that have demonstrated uptakeand transport ofoligonudeotides and dendriticand axonal transport of RNA (14). Our im-munocytochemical and RIA findings as well asobserved physiological responses indicate thatexogenous AVP mRNA was also translated bysome of the magnocellular perikarya. Uptakeand translation ofAVP mRNA seemed to beselective processes because injections of theAVP antisense probe did not result in anylabeling of these perikarya or their nerve end-ings in the posterior lobe. Moreover, AVPimmunoreactivity or labeled RNA did not be-come detectable in hypothalamic neurons otherthan the classical magnocellular neurons in theSON and PVN. Furthermore, when we inject-ed total RNA fractions (which certainly con-tained mRNA coding for peptides other thanAVP), the magnocellular hypouthalamic neu-rons did not express ACTH and P-endorphinimmunoreactivities.Our findings suggest that rat hypothalamic
magnocellular neurons can accumulate andtranslate certain specific exogenous RNAsnormally expressed in these neurons. Theautoradiographic data showing concentrationof35S-labeled mRNA in cell bodies as well asin the posterior lobe [most likely confined tosecretory nerve terminals (2)] indicate that atleast in Brattleboro rats RNA is accumulatedand transported anterogradely as well as ret-rogradely upon injection into the medial por-tion of the hypota e physialtract. We condude that the mechanisms ofuptake, transport, and translation of RNAmight represent a cascade of increasing selec-tivity: although exogenous mRNAs may betaken up by many axons, and smaller neu-ronal pools may be able to provide selectivetransport, only a few neurons may actuallytranslate the axonally delivered RNA. Thishypothesis is supported by the finding thatdegradation ofRNA is low in the brain (5).We have shown thatOTmRNA is present insecretory vesicles in the hypothalamoneuro-hypophysial system (2), suggesting that thesemRNAs exist in cellular compartments to-gether with peptide signals known to besecreted. Recent studies with in situ hybrid-ization and electron microscopy indicate asimilar situation for AVP mRNA (15). Ax-onal accumulation, transport, and secretionofmRNA may offer new approaches to reg-ulate interneuronal signaling.
REFERENCES AND NOTES
1. B. Bloch, A. F. Guitteny, E. Normand, S. Chou-ham, Neurosci. Lett. 109, 259 (1990).
2. G. F. Jirikowski, P. P. Sanna, F. E. Bloom, Proc.Nat!. Acad. Sci. U.SA. 87, 7400 (1990).
3. E. Lchmann, J. Hanze, M. Pauschinger, D. Ganten,
R. E. Lang, Neurosci. Lett. 111, 170 (1990); A.Levy, S. Lightman, D. A. Carter, D. Murphy, J.Neuroendocrinol. 2, 329 (1990); D. Murphy, A.Levy, S. Lightman, D. Carter, Proc. Nad. Acad. Sci.U.SA. 86, 9002 (1989); D. Murphy and D.Carter, Mol. Endocrinol. 4, 1051 (1990).
4. P. J. Brooks, P. K. Lund, J. D. Caldwell, G. F.Jirikowski, C. A. Pedersen, Soc. Neurosci. Abstr. 14,484.10 (1990).
5. L. Fran7oni and C. A. Garcia-Argiz, Acta Physiol.Lat. Am. 28, 185 (1978); B. Schulz-Harder and D.Graf von Keyserlingk, Histochemnistry 88, 587(1988).
6. H. Schmale and D. Richter, Nature 308,705 (1984).7. Injections into the hypothanlm rohypophysial tract
{coordinates: x: 2.2 mm right,y: +2 mm,z:+ 2.2 mmfrom the bregma, according to G. Paxinos and C.Watson, [The Rat Brain in Streotaxic Coordinates (Aca-demic Press, New York, 1985)]} were carried out withan injection aid rather than a stereotaxic apparatus, in amodification of the method by F. R. Popick [Lie Sci.18, 197 (1976)]. The injected volume was in all cases 5pl. Seven male rats were injected either with RNAextract (8) or with RNA (9). Six rats injected withisotonic saline served as controls.
8. Total cytoplasmic RNA was isolated from the hypo-thalami ofa total often Sprague-Dawley rats. Tissuewas homogenized in 20 mM tris-HCl (pH 8.8), 200mM NaCI, and 20 mM MgCI2. After a 10-mincentrifligation at 3000g at 4'C, the supematant wasdigested for 20 min in proteinase K (2.5 mg/ml) in2% SDS, 40 mM EDTA (pH 8.0), and 100 mMNaCl. Purification ofRNA was then performed witha phenol:chloroform:isoamnyl alcohol (50:48:2) ex-traction. U. Schibler, M. Tosi, A. C. Pittet, L.Fabiani, K. R. Wellauer, J. Mol. Biol. 142, 93(1980); D. Lenoir, E. Battenberg, M. Kiel, F. E.Bloom, R. J. Milner, J. Neurosci. 6, 522 (1986).
9. A synthetic arginine-vasopressin RNA was geneatdby in viro transcription of a cDNA clone [donated byD. Richter, Eppendorf Institute, Hamburg; R. Ivelland D. Richter, Proc. Nad. Aad. So. U.SA 81, 1216(1984)] with SP6 bacterial RNA polymrase(Promega) according to the manufactns protocol.These synthetic RNAs were not polyadenylaed, RNAwas labeled with 35S-hbal cytidine 5'-tmphosphate(CIP) for the autoradiography experiments.
10. Control injections were carried out with an isotonicsaline vehicle and synthetic antisense RNA preparedfrom the same cDNA clone by transcription with T7RNA polymerase. An additional control was theinjection of sense RNA that had been digested withRNase A (20 jig/ml, Boehringer Mannheim) for 1hour at 37C. Further controls were done withinjections oftotal RNA extracted from rat hindbrain(a region devoid of vasopressin mRNA). Each con-trol group contained four animals.
11. We are indebted to P. Plotsky (Salk Institute) forperforming AVP RIAs. Tissue preparations andRIAs were carried out as double antibody RIAs,according to H. U. Haussler, G. F. Jirikowski, and J.D. Caldwell [J. Chem. Neuroanat. 3, 271 (1990)].
12. J. J. McCabe, J. I. Morrell, D. W. Pfaff, in In SituHybridization in Brain, G. R. Uhl, Ed. (Plenum,New York, 1985), pp. 73-95.
13. For control purposes three animals were injected withthe synthetic AVP RNA into the lateral ventricle andthree were injected into the cerebral cortex.
14. E. J. Carazanna, K. B. Pasieki, J. A. Majzoub, Mol.Cell. Biol. 8, 2267 (1988); H. H. Zingg, D. L.Lefebvre, G. Almazon, J. Biol. Chem. 263, 11041(1988); D. Carter and D. Murphy, ibid. 264, 6601(1989); S. L. Loke et al., Proc. Nat!. Acad. Sci.U.SA. 86, 3474 (1989); L. Davis, B. Burger, G.A. Banker, 0. Steward, J. Neurosci. 10, 3056(1990); E. Mohr, S. Fehr, D. Richter, EMBOJ. 10,2419 (1991).
15. A. Trembleau, M. Landry, M. Fevre-Montange, A.Calas, in Neurocytochemical Methods, A. Calas and D.Eugene, Eds. (NATO Advanced Study InstitutesSeries, Springer, New York, 1991), pp. 313-318.
16. L. A. Stemberger, Immunocytochemistry (Wiley, NewYork, ed. 2, 1978).
17. G.F.J. is a Heisenberg Fellow. Supported by VWStiftung (1/65 793) and U.S. Public Health Servicegrants NS 22347-03 and MH 47680.
4 September 1991; accepted 6 December 1991
SCIENCE, VOL. 255
on January 31, 2021
http://science.sciencemag.org/
Dow
nloaded from

mRNAReversal of diabetes insipidus in Brattleboro rats: intrahypothalamic injection of vasopressin
GF Jirikowski, PP Sanna, D Maciejewski-Lenoir and FE Bloom
DOI: 10.1126/science.1546298 (5047), 996-998.255Science
ARTICLE TOOLS http://science.sciencemag.org/content/255/5047/996
REFERENCES
http://science.sciencemag.org/content/255/5047/996#BIBLThis article cites 20 articles, 9 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.ScienceScience, 1200 New York Avenue NW, Washington, DC 20005. The title (print ISSN 0036-8075; online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
No claim to original U.S. Government Works.Copyright © 1992 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science.
on January 31, 2021
http://science.sciencemag.org/
Dow
nloaded from