on the reproductive processe ofs the brandlin g worm ... · various pabula were tried, but the most...
TRANSCRIPT

On the Reproductive Processes of the BrandlingWorm, Eisenia foetida. (Sav.)
By
A. J. Grove, M.A. (Cantab.), D.Sc. (Birm.),
Lecturer in Zoology,
and
L. F. Cowley, M.Sc. (Bristol),A s s i s t a n t C u r a t o r o f t h e Z o o l o g i c a l M u s e u m , U n i v e r s i t y o f S h e f f i e l d .
W i t h P l a t e 3 0 a n d 4 T e x t - f i g u r e s .
C O N T E N T S .
P A G E
I N T R O D U C T I O N . . . . . . . . . . 5 5 9H I S T O R I C A L 5 6 0
M E T H O D S 5 6 1
C O I T I O N 5 6 4
D i r e c t O b s e r v a t i o n s . . . . . . . . 5 6 4T h e C o i t i o n S l i m e - t u b e 5 6 7T h e S e m i n a l G r o o v e . . . . . . . . 5 6 9T h e A t t a c h m e n t o f t h e W o r m s 5 6 9
C O C O O N F O R M A T I O N A N D D E P O S I T I O N . . . . . . 5 7 1
T h e C o c o o n 5 7 2' C o c o o n F o r m a t i o n . . . . . . . . 5 7 2
C o c o o n D e p o s i t i o n . . . . . . . . 5 7 3T h e P a s s a g e o f t h e E g g s i n t o t h e C o c o o n . . . . 5 7 5T h e P a s s a g e o f t h e S p e r m i n t o t h e C o c o o n . . . . 5 7 7
S U M M A R Y . . . . . . . . . . 5 7 9REFERENCES . . . . . . . . . . 580
EXPLANATION OF PLATE . . . . . . , . 581
INTRODUCTION.
IN a previous paper one of us (Grove, 1925) described someobservations on the sexual congress of L. t e r r e s t r i s . Theseobservations led naturally to the consideration of other impor-
NO. 280 p p

560 A. J . GROVE AND L. F. COWLBY
tant reproductive processes, e. g. cocoon formation and deposi-tion, regarding which our knowledge is scanty in respect ofmany essential details. Perhaps the most pressing point uponwhich light is desired is the exact relationship between coitionand cocoon formation. In this connexion Foot (1898) statesthat in the Brandling (E. f o e t i d a) during coition both wormsform cocoons whilst they are still united, and that each depositsits cocoon, enclosed in a moiety of the slime-tube, after separa-tion. This statement, and the fact that these worms providedmore easily manipulated material than L . t e r r e s t r i s , ledto the selection of the Brandling for these present observations.
In B . f o e t i d a the oviducts open on segment 14, the vasadeferentia on 15, and the spermathecae between 9-10 and10-11. The clitellum extends usually from circa 26-32. Theonly important differences between E . f o e t i d a and L. t e r -r e s t r i s are therefore the position of the clitellum, and thatthe apertures of the spermathecae are dorsal and not lateral inposition. Although the general arrangement of the reproduc-tive organs is similar to that which obtains in L. t e r r e s t r i s ,yet it seemed probable that the dorsal position of the aperturesof the spermathecae in E . f o e t i d a would result in some diver-gences from the account of the process of coition which hasbeen given for L. t e r r e s t r i s . This probability necessitatedfresh observations on this process in the Brandling, and asseveral new points have been recorded, an account of thecoition in addition to that of cocoon formation and depositionseemed desirable.
Our thanks are due to the Committee of the Endowmentof Research Fund of the Birmingham Natural History andPhilosophical Society for a grant to assist these investigations,and also to Professor A. Denny for his continued encourage-ment.
HISTORICAL.
Among the earliest accounts of the coition of the Brandlingis that of Perrier (1875), who calls attention to the marked con-strictions which occur in the region of the girdle. Andrews

EBPRODUOTIVB PROCESSES OF BISENIA FOETIDA 561
(1895) gives a more extended account derived from the examina-tion of preserved material, stating that direct observation isprecluded by the fact that the worms do not come to the surfaceto conjugate. He describes the dorsal position of the aperturesof the spermathecae, and emphasizes the extent to which theclitellum embraces the adposed segments. From his observa-tions he concludes that coition in E . f o e t i d a follows closelythat of L. t e r r e s t r i s as described by Hering (1857).
Foot (1898) does not describe the details of coition, butfrom her observations deduced certain relationships betweencoition and cocoon formation. On page 483 this authorstates : ' Both cocoons are formed while, the worms are united,and when they separate each deposits a cocoon encasedby a moiety of the slime-tube. . . . Each cocoon appearsto be formed around both worms, encircling the clitellum ofone worm and three or more of the anterior segments of theother worm. As the worms withdraw backwards out of thecocoons, the eight free anterior segments of each must be with-drawn first, leaving a cocoon around the clitellum of eachworm ; the worm finally leaving this cocoon at the end repre-sented in Text-fig. 3 by the thread-like extension.' It will be shownlater that this author has mistaken the constricting bands seenduring coition for the ends of the cocoons, and further, hasfailed to distinguish between the slime-tube formed duringcoition and the cocoon slime-tube. Further, her statementpresents a quite different aspect of the process of cocoon forma-tion from that given in the only other record of direct observa-tions, viz. that of Vedjovsky (1892) on E h y n c h e l m i s .Foot (p. 497) was also induced to suggest that the usual functionascribed to the spermathecae is not the correct one, and thattwo acts of coition are necessary before cocoon deposition.
METHODS.
The common habitat of E . f o e t i d a is in heaps of well-rotted manure where, when conditions are favourable, theymay be found in considerable numbers. The worms alwaysremain below the surface of the compost, and on exposure to
pp 2
562 A. J . GROVE AND L. F . COWLEY
light exhibit a definite negative heliotropic reaction. It wasalso found that even under conditions of total darkness suchas those which were used for the observations on L . t e r r e s -t r i s , the worms did not come to the surface for coition. Thisis confirmed by the observation that in examining manureheaps, pairs of worms may be found copulating below thesurface. This habit necessitated the development of specialapparatus to render observations possible. This apparatus
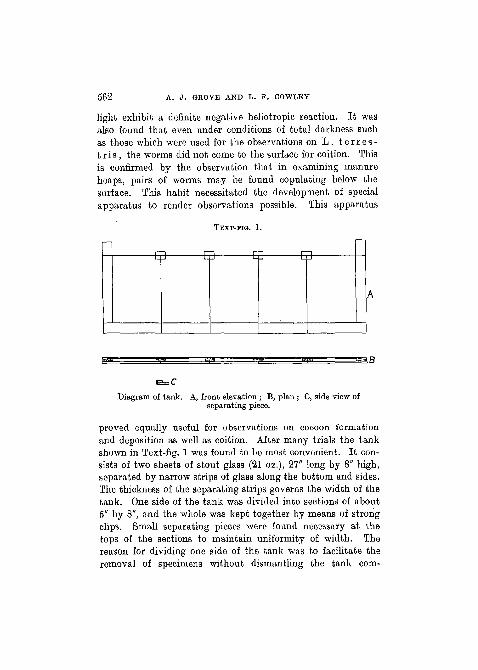
TEXT-FIG. 1.
Diagram of tank. A, front elevation ; B, plan ; C, side view ofseparating piece.
proved equally useful for observations on cocoon formationand deposition as well as coition. After many trials the tankshown in Text-fig. 1 was found to be most convenient. It con-sists of two sheets of stout glass (21 oz.), 27" long by 8" high,separated by narrow strips of glass along the bottom and sides.The thickness of the separating strips governs the width of thetank. One side of the tank was divided into sections of about5" by 8", and the whole was kept together by means of strongclips. Small separating pieces were found necessary at thetops of the sections to maintain uniformity of width. Thereason for dividing one side of the tank was to facilitate theremoval of specimens without dismantling the tank com-

REPRODUCTIVE PROCESSES OF EISENIA FOETIDA 568
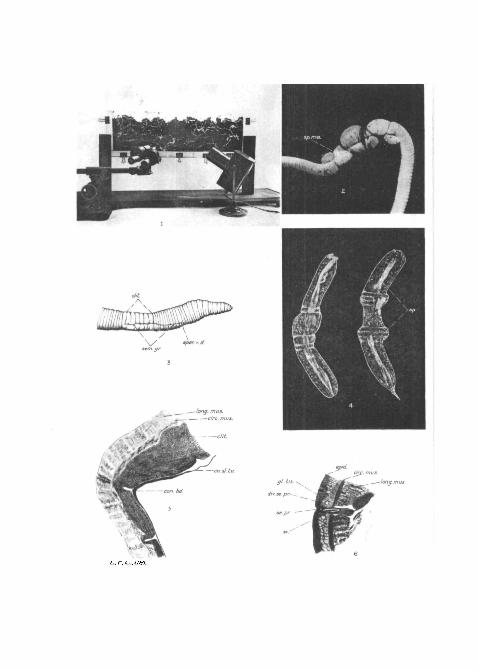
pletely. Each tank was supported-by a wooden stand (PI. 30,fig. 1), which was constructed in such a way that the tank,resting in two grooves in the wooden uprights, enabled it tobe removed with ease and rapidity. The tanks were filled witha convenient medium and the width was adjusted so that,although the worms could move about freely, one or both ofthe sides of their bodies was in contact with the glass permittingobservations to be made. In practice it was found that if thetanks were too narrow, coition was interfered with, the wormsbeing apparently unable to complete the process.
Various pabula were tried, but the most suitable was foundto be a mixture of rotten leaves and well-rotted horse manurein the proportion of one part of the former to two of the latter.For convenience in filling the narrow tanks, the mixed leavesand horse manure, whilst still wet, were finely divided bypassing them through a mincing machine. The material wasthen partially dried and then again passed through the mincerand finally sifted. [The drying process should only be allowedto proceed sufficiently to enable the material to be sifted, other-wise difficulty will be experienced in remoistening.] In thispowdery condition the material could be shaken down aninclined plane into the tank. Water was then added until thepabulum was suitably moist, when the worms (12-18 per tank)were introduced. The pronounced negative heliotropic reactionof the worms necessitated keeping the tanks in a dark room sothat natural conditions were closely simulated.
Mast (1917) showed that in L u m b r i c u s the maximumnegative heliotropic reaction was with light of a wave length of483-4 fi/x, i. e. where the blue portion of the spectrum merges intothe green. Experiments with E . f o e t i d a showed that theseanimals were relatively insensitive to weak orange light, andthis was used for general illumination of the room duringobservations. It was also found that during the ' intense 'periods of coition or cocoon formation, condensed red light ofsufficient brilliancy to permit of the use of a binocular microscopecould be employed.
Specimens for examination were killed by allowing them to

564 A. J . GEOVE AND L. F. COWLEY
fall into hot water (about 50° C.)> and for histological purposesBouin's fixative was used. Ehrlich's acid haematoxylin wasused for general staining.
COITION.
D i r e c t O b s e r v a t i o n s .The following account of the coition of E . f o e t i d a is a
summary of the details obtained from the observation underthe binocular microscope of many pairs, supplemented in someinstances by the macro- and microscopic examination of thepair after killing.
Among the first external indications in the worms that coitionis about to take place, is an apparent increased tumescence inthe clitellum. In the narrow tanks used this tumescence wasthe more marked because of the pressure against the glass sides.The preliminaries to coition appear to be a mutual caressing ofthe anterior segments of one worm by the ' head ' of the other.During this process the worms usually approach one anotherfrom opposite directions, and the caressing movements aregenerally confined to the ventral surface of the first 8-10 seg-ments. The movements are relatively rapid, probably indica-tive of some condition of nervous tension, and they may con-tinue for an extended period (in some cases hours) before con-nexion is established. An increased secretion of mucus is alsoevident at this time. No constancy of position assumed by theworms in the tanks during coition was observed, and all possiblevariations between the horizontal and vertical positions werenoticed. During coition the worms assume the typical head-to-tail position with their ventral surfaces applied to one another,the clitellum of one being adposed to segments 8-11 of theother. The relation of the clitellum to the adposed segmentsis an intimate one. During the most intense periods the clitel-lum practically encloses the adposed segments, so that, in someinstances, the free edges of the clitellum have been observedalmost to meet over their dorsal surfaces. Also, as will beshown later, the ventral surface of the adposed segments is
REPRODUCTIVE PROCESSES OF EISENIA FOETIDA 565
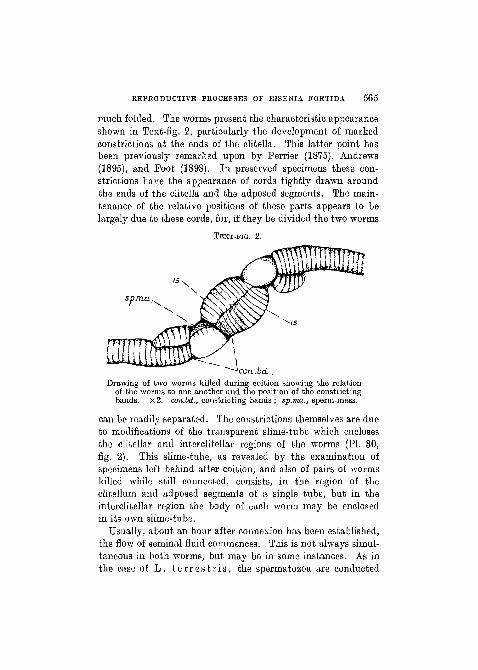
much folded. The worms present the characteristic appearanceshown in Text-fig. 2, particularly the development of markedconstrictions at the ends of the elitella. This latter point hasbeen previously remarked upon by Perrier (1875), Andrews(1895), and Foot (1898). In preserved specimens these con-strictions have the appearance of cords tightly drawn aroundthe ends of the elitella and the adposed segments. The main-tenance of the relative positions of these parts appears to belargely due to these cords, for, if they be divided the two worms
TEXT-FIG. 2.
Drawing of two worms killed during coition showing the relationof tho worms to one another and the position of the constrictingbands. x2. con.bd., constricting bands ; sp.ma., sperm-mass.
can be readily separated. The constrictions themselves are dueto modifications of the transparent slime-tube which enclosesthe clitellar and interclitellar regions of the worms (PL 30,fig. 2). This slime-tube, as revealed by the examination ofspecimens left behind after coition, and also of pairs of wormskilled while still connected, consists, in the region of theclitellum and adposed segments of a single tube, but in theinterclitellar region the body of each worm may be enclosedin its own slime-tube.
Usually, about an hour after connexion has been established,the flow of seminal fluid commences. This is not always simul-taneous in both worms, but may be in some instances. As inthe case of L . t e r r e s t r i s , the spermatozoa are conducted

566 A. J. GROVE AND L. F. COWLEY
from the apertures of the vasa deferentia to the clitellum alongthe lines of the seminal grooves. Each groove is distinctlymarked in spirit specimens, and extends backwards (PI. 30,fig. 3) from the outer margin of the tumid lips about the aper-ture of the vas deferens almost to the posterior margin of theclitellum. In the pairing worms, however, the groove as awhole is not obvious, but its position is indicated by the lineof backwardly moving pit-like depressions in the epidermis inwhich the seminal fluid is conveyed. The tanks used permittedthe examination of both sides of the worms, and it was possibleto determine by direct observation that the flow of seminalfluid took place on both sides of each of the co-operating worms.This afforded additional evidence in support of this view putforward in the case of L . t e r r e s t r i s , in which, from theposition taken up by the worms, examination of both sideswas not possible whilst the process was in progress. The pit-like depressions appear at intervals of about three segments,and each sperm-mass covers about one segment; so that asone depression appears in the vicinity of the aperture of thevas deferens, a second is mid-way between it and a third whichis situated at the anterior end of the clitellum. During themost active period the depressions appear and travel backwardsat the rate of about ten per minute. The passage of spermcontinues usually for about 30—45 minutes, but may in someinstances be of longer duration.
On reaching the clitellum the depressions with their containedsperm travel along the lateral surface to the end of the seminalgroove, whence (Text-fig. 3) they pass ventrally round theposterior margin of the free edge of the clitellum and the spermis deposited between it and the adposed segments. Along thefree edge, waves of a much shorter amplitude are visible runningin a postero-anterior direction which serve to aggregate thesperm into masses over the dorsally placed apertures of thespermathecae. This result is also assisted by the embracingand releasing movements of the clitellum which become pro-nounced at this time, for, at the moment of closest embrace thecircumstance that the clitellum practically encloses the adposed

REPRODUCTIVE PROCESSES OF EISENIA FOETIDA 567
segments ensures that the sperm lies on the dorsal surface of thesesegments, between the free edges of the clitellum. With thesecontinued movements the sperm-masses gradually diminish insize having passed into the spermathecae. When these pro-cesses are completed each worm backs out of the coition slime-tube, which is left behind.
TEXT-PIG. 3.
a.per. v.d...
Diagram of the two worms together to show the course of the seminalfluid from the aperture of the vas deferens to the clitellum. x 2.aper.v.d., aperture of vas deferens; sp.ma., sperm-mass.
The Co i t i on S l i m e - t u b e .This structure is, in the Brandling, sufficiently stable in com-
position to maintain its integrity after the worms have with-drawn from it. It is extremely delicate, however, and can onlybe satisfactorily examined in water, for, composed as it is, ofa transparent mucilaginous material, it is very susceptible todrying, and under dry conditions shrivels up and becomesindistinguishable in a short time after the withdrawal of theworms. Its appearance is conditional upon the way in whichthe worms have separated ; in some instances it may bepartially rolled up, but usually, in the tanks, it was left attachedto th& glass, and more or less extended. In water or 5 per cent,formalin it rapidly assumes its normal contour, and the positionsof the constricting bands, although they are not so much inevidence, can be distinguished (PI. 30, fig. 4). Foot (1898,
568 A. J . GKOVE AND L. P. COWLEY
p. 481) gives the average extent of this slime-tube to be fromthe eighth to the thirty-third segments in each worm, and thishas been confirmed during the present observations. In sectionsthrough the pairing worms the slime-tube is. seen to consist ofseveral layers or lamellae (PI. 30, fig. 5), suggesting that it isproduced by a series of periods of secretive activity. The tubeitself gives the mucin reaction, so that it may be inferred thatit is a product of the mucin secreting cells of the epidermis.
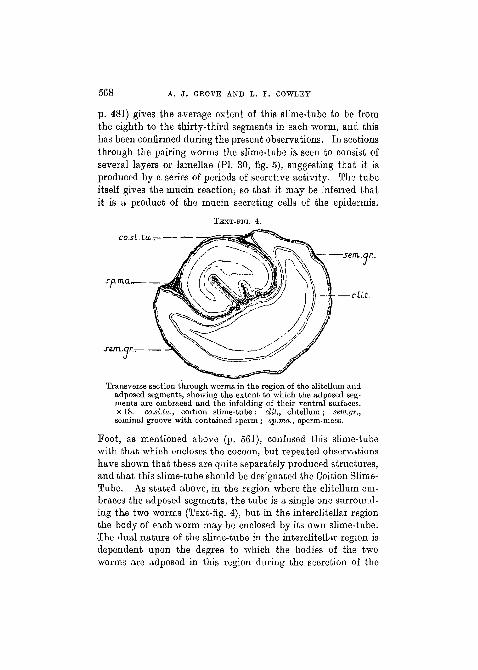
TEXT-FIG. 4.
co.si.tu,.-, '—
sp.m.an
c sem.qr..
—cat.
Transverse section through worms in the region of the clitellum andadposed segments, showing the extent to which the adposed seg-ments are embraced and the infolding of their ventral surfaces,x 18. co.sl.la., coition slime-tube; clit., clitellum; sem.gr.,seminal groove with contained sperm ; sp.ma., sperm-mass.
Foot, as mentioned above (p. 561), confused this slime-tubewith that which encloses the cocoon, but repeated observationshave shown that these are quite separately produced structures,and that this slime-tube should be designated the Coition Slime-Tube. As stated above, in the region where the clitellum em-braces the adposed segments, the tube is a single one surround-ing the two worms (Text-fig. 4), but in the interclitellar regionthe body of each worm may be enclosed by its own slime-tube.The dual nature of the slime-tube in the interclitellar region isdependent upon the degree to which the bodies of the twoworms are adposed in this region during the secretion of the

REPRODUCTIVE PROCESSES OF EISBNIA FOETIDA 569
mucus ; for, if there is a small space between the bodies of theworms, then each worm will be completely surrounded by itsown secretion, whereas, if the bodies are closely adposed through-out, then the fluid mucus will coalesce along the lateral marginsand form a common tube enclosing both worms. In either casethe interclitellar portion of the tube merges insensibly into thesingle tubes surrounding the clitella and adposed segments.
From the examination of sections through the pairing worms(PI. 30, fig. 5), it is seen that the characteristic constrictingbands, so evident in a surface view, consist merely of thickeningsand to some extent foldings of the slime-tube. Exactly howthese slight thickenings and foldings produce such markedconstrictions in the bodies of the worms is difficult to conjecture,and so far no further light has been thrown upon this pointduring these observations.
The Seminal Groove.In E . foe t ida the seminal groove never functions as a
continuous furrow extending from the aperture of the vasdeferens to the clitellum, the condition in which it is seen inspirit specimens. In effect, the so-called seminal groove merelyindicates the line along which, during coition, the pit-likedepressions with their contained sperm pass. The coursetraversed by these pits has been described above. The pitsare brought into existence by a similar system of arciformmuscles to that which has been previously described forL. t e r r e s t r i s .
The A t t a c h m e n t of the Wo rms .The Cl i t e l lum and Adposed Segments.—The rela-
tion between the worms in this region is, in some respects, moreintimate than is the case with L. t e r r e s t r i s . A transversesection (Text-fig. 4) through the worms whilst they are stillconnected shows that the ventral surface of the clitellum ismarkedly hollowed out so as to form a trough in which theadposed segments lie. This effect is also enhanced by a con-siderable folding of the ventral surface of the adposed segments,

570 A. J . GROVE AND L. F. COWLBY
which effectively reduces the circumference to be enclosed bythe clitellum. This folding of the ventral surface is probablybrought about largely by the pressure exerted in the embracingmovements of the clitellum, assisted by reciprocal contractionsof the musculature of the ventral part of these segments. Theopposed surfaces of the clitellum and the adposed segmentsalso show an interdigitation. Between these surfaces traces ofmucus and the secretions from the glands associated with thesetal pores are to be seen. This characteristic relation betweenthe clitellum and the adposed segments has been previouslyfigured by Andrews, and the similarity between this author'sfigure and Text-fig. 4 is so striking that it cannot be mere coin-cidence, but represents the normal relation of these parts duringcoition.
This rather remarkable relation between the worms in thisregion becomes intelligible when it is remembered that theapertures of the spermathecae are dorsal in position. It follows,therefore, that in order to render the entrance of the sperma-tozoa possible, it is essential that they must be brought on tothe dorsal surface of the adposed segments by the action ofthe free edges of the clitellum. The folding of the ventral sur-face of the adposed segments with the consequent reductionof the circumference enables the embracing process to be morecomplete and the requisite end attained. The embracing andreleasing movements will also probably assist in the opening ofthe apertures of the spermathecae, further facilitating theentrance of the sperm.
The A c t i o n of t h e Setae.—No evidence has been forth-coming that the setae participate to any extent in the attach-ment of the worms as is the case in L. t e r r e s t r i s . A fewinstances have been observed in sections where rupture of theepidermis by the setae was in- evidence, but this was quite super-ficial.
The G l a n d s A s s o c i a t e d w i t h t h e S e t a l Po re s .—From the examination of a number of specimens killed in hotwater the occurrence of these glands showed considerablevariation, which has also been confirmed by the examination

REPRODUCTIVE PROCESSES OF EISENIA FOETIDA 571
of sections. They have been detected in segments 9, 10, 11,12, 23, and from thence to about 32. In segments 9-12 theducts from the glands usually open into the l a t e r a l setalpores (PI. 30, fig. 6), and in 9, 10, and 11 their arrangement maybe asymmetrical. It is interesting to note that the associationof the glands with the lateral setae in the anterior segments isa natural concomitant of the greater extent to which thesesegments are embraced by the clitellum and also the infoldingof the ventral surface of these segments during coition.
COCOON FORMATION AND DEPOSITION.
Foot (1898) emphasizes the statement that in E . f o e t i d acocoon formation takes place during coition, and says (p. 483) :' Both cocoons are formed while the worms are united, andwhen they separate each deposits a cocoon, encased by amoiety of the slime-tube.' The direct observations on thesexual congress recorded above, and the examination ofcoition slime-tubes left behind by the worms, failed to revealany trace of cocoons. The accuracy of Foot's observations wasat once called into doubt by these results, and this point wasconclusively determined by a simple experiment. Ten worms, inwhich the condition of their clitella indicated sexual maturity,and which had been kept together in an observation tank forabout a fortnight, were isolated, each being placed in a separateculture. These were examined daily. By the end of forty-sixdays no. 1 had produced 2 cocoons ; no. 2, 4 ; no. 3, 6 ; no. 4,1 ; no. 5, 2 ; no. 6, 6 ; no. 7, 11 ; no. 8, 0 ; no. 9, 3 ; no. 10, 2.Although from later experiments it was discovered that thesefigures do not represent the optimum rate of cocoon deposition,yet the experiment afforded indubitable evidence that theformation and subsequent deposition of the cocoon is a quiteseparate process from that of coition. These observations weresupplemented by the isolation of Avorms which had been observedduring coition, and it was found that as many as six days mayelapse before a cocoon was deposited.

572 A. J. GROVE AND L. F. COWLEY
The Cocoon.
When freshly deposited the cocoon of E . f o e t i d a (PI. 30,fig. 7) is lemon-shaped, of a translucent white colour, and iscompletely enveloped in a slime-tube. The posterior end ofthe cocoon is well rounded and terminates in a short thread-likeprojection which is attached to the posterior end of the slime-tube. Anteriorly the cocoon is drawn out into a funnel-shapedstructure with fimbriated margins. The slime-tube is approxi-mately three to foar times the length of the cocoon which isalways found near the posterior end of the tube. On exposureto the air the cocoon slime-tube dries up and disappears, andthe cocoon attains a straw colour. The cocoon consists of anenclosing wall or membrane which is closely invested by theslime-tube but is different from it in structure. This membraneis, on drying, tough and resistant, resembling somewhat thechitin of insects, and it is evident that the ends of the cocoon aremaintained closed by the elasticity of the membrane, for ifa freshly deposited cocoon is subjected to slight pressure thecontents do not escape.
The contents of the cocoon consist of a gelatinous mass ofalbumen, which is almost transparent in a freshly depositedcocoon but becomes opaque after a few hours. In this albumenfrom eight to sixteen eggs are found.
Cocoon F o r m a t i o n .The very early stages of cocoon formation are difficult to
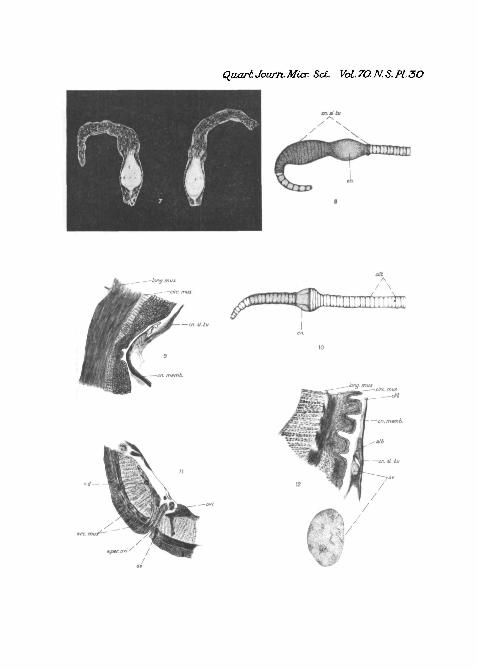
detect, but the first indication has been found to be the forma-tion of a slime-tube extending from about segments 6-8 toabout 2-3 segments posterior to the clitellum. This is practi-cally invisible whilst the worm is in the tank, but it can bedemonstrated after the specimen has been killed, and exampleshave been examined in which the slime-tube was clearlydemonstrable although there was no evidence of the cocoonproper present. When the formation of the cocoon is welladvanced the worm has a characteristic appearance (PI. 30,fig. 8). The most common, though not universal, position isto have the anterior portion curved so that the ' head ' end of

REPRODUCTIVE PROCESSES OF EISENIA FOETIDA 573
the worm is quite close to the clitellum, the body anterior toand including the clitellum thus forming a loop with an openend. Also, the clitellum loses its angular ends and presents anoval appearance due, no doubt, to its being enclosed by thecocoon membrane and the latter merging into the slime-tube.The segments immediately posterior to the clitellum haveusually a somewhat attenuated appearance, and there is a con-striction visible which marks the posterior end of the clitellumand naturally of the cocoon also. As fig. 9 (PL 30) shows, thisconstriction is due to the slime-tube being partially telescopedand at the same time thrown into folds, and also, to some extentto the termination of the cocoon membrane. The segmentsof the body anterior to the clitellum exhibit varying conditionsof expansion and contraction. Segments anterior to eleven areusually fully extended longitudinally, while transverse expan-sion is most marked from segment 12 to about 23, where, lyingimmediately in front of the clitellum, a constriction is visiblenot quite so marked in extent as that found posterior to theclitellum.
During this process of cocoon formation the worm remainsrelatively stationary and exhibits the previously mentionedinsensitiveness to red light. Owing to the position of the wormin the tank it was impossible to observe the dorsal or ventralsurfaces. Muscular movements of several kinds were observedlaterally, and although it was not possible to refer these todefinite activities, yet in every instance the same kinds ofmovement were observed. Among the most noticeable wereflowings, both regular and irregular, of the coelomic fluid.Also irregular contractions of the body-wall producing pit-likedepressions moving sometimes anteriorly and sometimesposteriorly. Further, just prior to deposition concertina-likemovements were evident in the segments between 10 and16 (i. e. in the middle of the loop).
Cocoon D e p o s i t i o n .At the commencement of cocoon deposition the segments
immediately posterior to the clitellum are withdrawn from the

574 A. J . GROVE AND L. F. COWLEY
slime-tube, swell up, and force the slime-tube forwards, intensi-fying the constriction previously remarked upon at the posteriorend of the clitellum. Some of the segments of the clitellum arethen withdrawn from within the cocoon and these likewise swellup and, pressing against the posterior margin of the cocoon,thus maintain it in position whilst the segments immediatelyanterior to the swollen ones are withdrawn. It should beemphasized that the processes of withdrawal and swelling arecontinuous and practically simultaneous. This is no doubt dueto the existence of a condition of positive pressure in thecoelomic fluid in the segments immediately behind the cocoon,with the result that immediately a portion of the body becomesfree from the restraint exercised by the cocoon it swells up(PI. 30, fig. 10). This method of withdrawal is continued untilthe worm is free with the exception of the first four or fivesegments. During the time that these are being liberated thereare visible four or five characteristic jerks. Prior to the appear-ance of these jerks the cocoon has remained stationary, whilstthe worm, by the processes indicated above, has moved backwardsout of it. But now, with the onset of these jerks, the move-ments of the anterior portion of the body carry the cocoon withit. The reason for these jerks is not easy to conjecture, but itis possible that they may serve to rectify any invagination ofthe cocoon membrane or slime-tube which may have beencaused during withdrawal, or it may be that they are due to theinability of the anterior segments to follow the same swellingprocess which obtained in the more posterior ones. At thetermination of the jerk-like movements the prostomium isusually either still within the posterior end of the cocoon orquite free from it, though still within the posterior end of theslime-tube. The worm then frees itself completely and movesaway.
The average time taken for deposition, i. e. the time whichelapses from the moment when the clitellum leaves the cocoonuntil the worm is completely free, is about three and a halfminutes. Immediately after cocoon deposition the clitellumof the worm presents a distinctive appearance. It loses its

REPRODUCTIVE PROCESSES OF EISENIA POETIDA 575
normal light coloured, pronounced, smooth appearance ;exhibits definite reduction in bulk, so that now it does notproject from the surface of the body, and approximates moreclosely in colour to that of the other segments.
The P a s s a g e of the Eggs i n t o the Cocoon.In the text-books in common use it is usually stated that,
in L u m b r i c u s , the cocoon receives the ova and spermatozoaduring its passage over the apertures of the oviducts and sperma-theca'e respectively. As the only essential difference betweenL u m b r i c u s and E . foe t ida is the dorsal position of theapertures of the spermathecae in the latter worm, it would bereasonable to expect that the account ascribed to L u m b r i c u swould apply to E i s e n i a also. However, doubts as to theaccuracy of this statement were at once raised when the shorttime taken for cocoon deposition was recorded. Also, the back-ward progress of the worm through the cocoon was so con-tinuous, and the time taken so relatively short that it wasinconceivable that the eggs (or even the spermatozoa) couldbe passed into it in the manner stated. Interesting and con-clusive evidence on this point was obtained in the followingmanner. A worm was kept under observation whilst the pro-cess of cocoon formation was in progress until the momentwhen, from the swelling of the segments immediately posteriorto the clitellum, it was evident that deposition was about totake place. The worm was then allowed to fall, by the removalof one section of the side of the tank, into hot water so that itwas immediately killed. The position of the cocoon, whetherit still remained completely or only partially around theclitellum, depended upon the rapidity with which the wormwas removed from the tank. The cocoon was then dissectedfrom off the worm and its contents examined. According tothe usually accepted view given above, it was evident from itsposition that the cocoon should not have received either eggsor spermatozoa. Yet, when the contents were examined, itwas found that eggs were definitely present varying from eightto sixteen in number. Although in the earlier observations on
NO. 280 Q q

576 A. J . GROVE AND L. F. COWLEY
this point, the cocoon was dissected from off the worm tofacilitate the more exact determination of the presence of eggs,later experience showed that if the worm was examined inwater under the binocular microscope immediately after killing,the eggs could be discerned and even counted through the stilltransparent cocoon membrane. In all cases so examined theeggs were found lying between the ventral surface of theclitellum and the cocoon membrane. They were usually con-tained within a small quantity of albumen-like fluid, which insections (PI. 30, fig. 12) shows a different appearance from thegeneral albuminous contents of the cocoon.
This important observation at once raises the question ofhow the ova pass back from the apertures of the oviducts tothe region of the clitellum. As has been previously mentioned,observation of the ventral surface was impossible owing to theposition assumed by the worm during the cocoon formation.Attempts were made to overcome this difficulty by placing thetank in such a way that the worm could only move in the hori-zontal plane, in the hope that by so doing it might be inducedto take up a position during cocoon formation with its ventralsurface against the glass. Up to the present, however, theseattempts have proved fruitless, and as a consequence it hasnot been possible to throw any further light upon this pointby direct observations. There are, however, several theoreticalconsiderations which may be of value in this connexion. Theexamination of transverse sections of the worm throughsegment 14 shows that the lower end of the arciform musclesimpinges upon the epidermis at a point contiguous with theouter margin of the oviduct (PL 30, fig. 11). It will follow,therefore, that the oviduct will open in line with the lowergroove that is visible in spirit specimens as a result of the con-traction of the arciform muscles. It is thus possible that thelower groove functions as an ovarian groove in the same wayas the upper functions as a seminal groove. It is impossibleto say whether, if this surmise is correct, the eggs are conveyedbackwards in pit-like depressions in the epidermis as is thecase with the sperm, but from general considerations this seems

REPRODUCTIVE PROCESSES OF EISENIA FOETIDA 577
probable. It also seems reasonable to suppose that the eggswill be contained in some fluid medium which may be eithercoelomic fluid or a secretion of the receptaculum ovorum. Itwill be of interest to record in this connexion that in one instancea worm was killed during cocoon formation when the slime-tubeand cocoon membrane were fully formed and still surroundingthe clitellum, but which, upon examination by means of serialsections, showed the eggs to be still within the receptaculumovorum.
The P a s s a g e of t h e S p e r m i n t o t h e Cocoon .The determination of the fact that the eggs pass back into
the cocoon whilst the latter is still surrounding the clitellumleads naturally to the consideration of the spermatozoa, andwhether they are passed back into the cocoon also. Conclusiveevidence upon this point proved to be extremely difficult toobtain. In the first place the contents of the cocoon wereexamined after it had been dissected from off the clitellum.This revealed the presence of numerous bodies which conformedin size and appearance to what Foot (1898) termed hyper-trophied spermatozoa. In her description the author states(p. 495) : ' These spermatozoa of abnormal growth vary greatlyin size and are relatively far more numerous in the cocoonscontaining fertilized eggs. In some cases they appear to bedeveloped by an abnormal growth of the head alone, while inothers, though all parts select the same stain, the spine, head,and tail can be clearly identified.' The further the examina-tion of these structures progressed the more it became evidentthat they could not be true spermatozoa. For, apart from thevariation in length—which itself would have been surprisingin spermatozoa that had so recently reached the cocoon—the failure to respond to chromatin stains, and a general lackof differentiation into the typical regions of a spermatozoon,militated strongly against the possibility of their being sperma-tozoa at all. From a comparison with fibres visible in the cutedges of the cocoon, it seemed extremely probable that theywere similar in nature. An attempt was then made to detect
Q q 2

578 A. J . GROVE AND L. F. COWLBY
spermatozoa in sections, both longitudinal and transverse,through the worm and cocoon, whilst the latter was still roundthe clitellum. In these sections many bodies similar to theabove were noticed, but a relatively small number of structures,conforming more closely to the size of true sperm, and respond-ing more definitely to chromatin stains, were observed. Buteven these bodies did not show a sufficiently satisfactorychromatin differentiation from the surrounding albumen.Further, sections through a cocoon after deposition did notreveal a larger number of these bodies. It might be mentionedhere that the preparation of sections through the cocoon andclitellum, and also the cocoon after deposition proved extremelydifficult, owing to the relative impermeability of the cocoonmembrane and albumen to reagents, and also to the shrinkageof the albumen during dehydration. The difficulties of theinvestigation were further increased by the fragmentation of thealbumen during sectioning. Even apart from these difficultiesthe relative scarcity of recognizable sperm-like bodies was bothdisappointing and surprising ; for it seemed reasonable tosuppose that if spermatozoa did pass back into the cocoon theywould be present in larger numbers. This consideration led tothe suggestion that possibly fertilization took place in the slime-tube during the passage of the eggs back to the cocoon. Theobtaining of evidence on this point proved to be even moredifficult, for the shrinkage of the slime-tube during dehydration,and its staining properties, rendered the preparations mostunsatisfactory and no sperm could be definitely detected. Theexamination of eggs in the cocoon whilst the latter was stillsurrounding the clitellum, from the point of view of any indica-tions of fertilization, proved equally abortive, owing to theunsatisfactory condition of the cytoplasm due to imperfectfixation as a result of the difficulties already mentioned in thisconnexion. Evidence was obtained, however, that in theeggs whilst still in the receptaculum ovorum during cocoonformation, and also in those that had been passed back into thecocoon but before cocoon deposition, nuclear division was takingplace (PL 30, fig. 12). The imperfect fixation precluded exact

BEPBODUCTIVE PROCESSES OF EISENIA FOETIDA 579
counts of the chromosomes, but not more than eleven chromo-somes were detected in any instance. This seemed to indicatethat the eggs were in the first maturation division since no polarbodies were observed. According to Foot (1894, p. 476) thepenetration of the eggs by spermatozoa takes place during thefirst maturation division, so that, if this statement is correct,the eggs as they issue from the oviduct are probably in a condi-tion to receive the spermatozoa.
From all these considerations there appear to be two possi-bilities. First, that the spermatozoa pass backwards, in whichcase fertilization may take place either in the slime-tube or in thecocoon whilst still surrounding ^he clitellum ; second, that thespermatozoa are squeezed out from the spermathecae into thecocoon during its passage over the apertures of the sperma-thecae. With regard to this latter possibility it would be reason-able to expect that there would be present in the freshlydeposited cocoon, free, actively moving spermatozoa, but up tothe present these have not been detected.
SUMMARY.
During coition in E . f o e t i d a the worms come together insuch a way that the clitellum of one embraces segments 8-11of the other. The whole of the clitellar and interclitellar regionsare enclosed in a protective coition slime-tube in which aredeveloped constricting bands at both ends of the clitella. Theline of the seminal groove extends from the fifteenth segment tonear the posterior end of the clitellum. The seminal fluid travelsbackwards, beneath the slime-tube, in pit-like depressions ofthe epidermis, which, on reaching the end of the line of the seminalgroove, travel to the ventral side of the clitellum, depositingthe sperm between its free edges and the adposed segments. Thepit-like depressions appear and travel backwards at the rateof about ten per minute on each side of each worm, makingfour streams of seminal fluid in all. The sperm-masses arebrought over the dorsally placed apertures of the spermathecaeby the embracing and releasing, movements of the clitellum.The almost complete embracing of the adposed segments by the

580 A. J . GROVE AND L. F. COWLEY
clitellum is facilitated by the infolding of the ventral surfacesof these segments.
It has been determined that cocoon deposition is a separateprocess from that of coition. This disposes of Foot's (1898)statement that cocoon formation takes place while the wormsare still united, which probably arose from the failure to distin-guish between the coition slime-tube and the cocoon slime-tube, and the mistaking of the constricting bands evidentduring coition for the ends of the cocoons.
During cocoon formation the slime-tube is developed, extend-ing from about the seventh to about the thirty-fourth segment.The cocoon membrane is then secreted around the clitellum, whichassumes an oval form with a marked constriction posteriorlyand a lesser one anteriorly. Whilst the cocoon still surroundsthe clitellum the eggs are passed back into it. It is still uncer-tain, however, whether the spermatozoa similarly pass back intothe cocoon or are squeezed into it from the spermathecae duringits passage over their apertures. The deposition of the cocoon iseffected by the gradual withdrawal of the worm with the excep-tion of the anterior three or four segments, during the freeingof which there usually occur three or four characteristic jerks.
EEFERBNCBS.
Andrews, E. A. (1895).—" Conjugation of the Brandling ", ' AmericanNaturalist', vol. xxix, pp. 1021-7, 1121-7.
Foot, Kath. (1894).—" Preliminary Note on the Maturation and Fertiliza-tion of the Egg of A. foetida ", ' Journ. of Morph.', vol. ix, pp. 475-85.
(1898).—"The Cocoons and Eggs of Allolobophora foetida", ibid.,vol. xiv, pp. 481-505.
Grove, A. J. (1925).—" On the Reproductive Processes of the Earthworm,Lumbricus terrestris ", ' Quart. Journ. Micr. Sci.', vol. Ixix, pp. 245—90.
Hering, E. (1857).—"Zur Anatomie undPhysiologie der Generations-organedes Regenwurms", 'Zeit. f. wiss. Zool.', Bd. viii, pp. 400-24, PI. xviii.
Mast, S. O. (1917).—" The Relation between Spectral Colour and Stimula-tion in the Lower Organisms", 'Journ. Exper. Zool.', vol. xxii, pp. 471-528.
Perrier, E. (1875).—"Note sur 1'accouplement des lombrics ", 'Arch, deZool. Exper. et Gen.', torn, iv, pp. xiii-xv.
Vejdovsky, F. (1892).—' Entwicklungsgeschichtliche TJntersuehungen'Prag.

REPRODUCTIVE PROCESSES OF EISENIA FOETIDA 581
REFERENCE LETTERING.
alb., albumen; aper.ovi., aperture of oviduct; aper.v.d., aperture ofvas deferens ; arc.mus., arciform muscles ; circ.mus., circular muscles ;clit., clitellum; en., cocoon; cn.memb., cocoon membrane; cn.sl.tu.,cocoon slime-tube; con.bd., constricting band; co.sl.tu., coition slime-tube ; div.se.pr., diverticulum of setal pore; epid., epidermis; gl.tis.,glandular tissue ; long.mus., longitudinal muscles ; wo., egg ; ow., oviduct;se., seta ; se.pr., setal pore ; sem.gr., seminal groove ; sp., sperm ; sp.ma.,sperm-mass ; v.d., vaa deferens.
EXPLANATION OP PLATE 30.Fig. 1.—Photograph of a tank as used during observations.Fig. 2.—Photograph of two worms killed during coition showing the
characteristic attitude and also a sperm-mass between the clitellum andthe adposed segments, x 1-5.
Fig. 3.—The anterior part of a worm after coition to show the courseof the seminal groove. (Drawn from a spirit specimen.) x2.
Fig. 4.—Coition slime-tubes showing sperm in the lumen. x4.Fig. 5.—A portion of a longitudinal section through the clitellum of
one of a pair of worms to show the constricting band, x 30.Fig. 6.—Transverse section through the lateral setal pore of segment 9
to show the glands associated with it. x 38-5.Fig. 7.—Cocoons in their slime-tubes, x 2-5.Fig. 8.—A semi-diagrammatic drawing of a worm during cocoon forma-
tion.Fig. 9.—Longitudinal section through a portion of the clitellum and
constricting band at the anterior end of the cocoon showing the cocoonmembrane merging into the laminate slime-tube. x47.
Fig. 10.—Semi-diagrammatic drawing of a worm during cocoon deposi-tion.
Fig. 11.—Transverse section through the aperture of the oviduct to showthe relation of the arciform muscles to it. x 38-5.
Fig. 12.—Transverse section through the ventral portion of the clitellumwith cocoon still surrounding it, showing an egg in the albumen. X40.Also egg more highly magnified. X180.
.•-. r. c, am.
Quart Journ. Mtcr. Set. Vol. 7O. N. S. PI. 3O
— — long mas.