opposing effects of swi/snf and mi-2/nurd chromatin remodeling
TRANSCRIPT

Opposing effects of SWI/SNF and Mi-2/NuRDchromatin remodeling complexes on epigeneticreprogramming by EBF and Pax5Hua Gaoa, Kara Lukinb, Julita Ramírezb, Scott Fieldsb, Desiree Lopezb, and James Hagmana,b,1
aProgram in Molecular Biology, University of Colorado at Denver Health Sciences Center, Aurora, CO 80045; and bIntegrated Department of Immunology,National Jewish Health, Denver, CO 80206
Edited by Anjana Rao, Harvard Medical School, Boston, MA, and approved May 12, 2009 (received for review September 24, 2008)
Transcriptionally silent genes are maintained in inaccessible chro-matin. Accessibility of these genes requires their modification bychromatin remodeling complexes (CRCs), which are recruited to pro-moters by sequence-specific DNA-binding proteins. Early B-cell factor(EBF), which is crucial for B-cell lineage specification, reprograms mb-1(Ig-�) promoters by increasing chromatin accessibility and initiatingthe loss of DNA methylation. In turn, this facilitates promoter activa-tion by Pax5. Here, we investigated the roles of ATP-dependent CRCsin these mechanisms. Fusion of EBF and Pax5 with the ligand-bindingdomain of ER� allowed for 4-hydroxytamoxifen-dependent, syner-gistic activation of mb-1 transcription in plasmacytoma cells. Knock-down of the SWI/SNF ATPases Brg1 and Brm inhibited transcriptionalactivation by EBF:ER and Pax5:ER. In contrast, knock-down of theMi-2/NuRD complex subunit Mi-2� greatly enhanced chromatin ac-cessibility and mb-1 transcription in response to the activators. Thereduction of Mi-2� also propagated DNA demethylation in re-sponse to EBF:ER and Pax5:ER, resulting in fully unmethylated mb-1promoters. In EBF- or EBF/Pax5-deficient fetal liver cells, both EBFand Pax5 were required for efficient demethylation of mb-1 pro-moters. Together, our data suggest that Mi-2/NuRD is importantfor the maintenance of hypermethylated chromatin in B cells. Weconclude that SWI/SNF and Mi-2/NuRD function in opposition toenable or limit the reprogramming of genes by EBF and Pax5 duringB-cell development.
DNA methylation � mb-1 promoter � Cd19 promoter � chromatin accessibility
The development of B cells from progenitor cells in the bonemarrow is controlled by a network of transcriptional regu-
lators (reviewed in 1). Early B-cell Factor (EBF; also known asEBF1/O/E-1/COE1) plays an integral role in this network andhas been implicated as a major determinant of the B-cell fate (2,3). In the absence of EBF, B-cell development is arrested at anearly progenitor stage (3, 4). EBF is essential for the rearrange-ment and expression of Ig (Ig) genes in B cells and is requiredfor expression of the B-cell commitment factor Pax5. EBF andPax5 synergistically activate transcription of B cell-specific genesincluding mb-1 (Cd79a), which encodes the Ig-� subunit of thepre-B and B-cell receptors (5). We have proposed that EBFfunctions as a ‘pioneer’ factor by controlling the epigenetic statesof its target genes (6). In response to EBF, the accessibility ofmb-1 promoter chromatin is increased, while DNA methylationof the promoter is decreased.
Specific biochemical interactions necessary for the pioneer func-tions of EBF have not been identified. However, transitions be-tween active and inactive states of chromatin can be mediated by therecruitment of chromatin-remodeling complexes (CRCs) by tran-scription factors (7). CRCs serve as ‘molecular motors’ that mediatechanges in the relative positions of nucleosomes. In this regard,CRCs of the mammalian SWI/SNF (related to yeast switch/sucrosenon-Fermenter) subfamily are important for the functions of manytissue-specific transcription factors. SWI/SNF components includethe Brahma (Brm/Smarca2) or Brahma-related gene 1 (Brg1/Smarca4) ATPases, which provide the energy to slide or evict
nucleosomes. SWI/SNF CRCs are important for controlling dif-ferentiation and proliferation in many cell types. In lymphocytes,CRCs have been implicated in activating transcription and antigenreceptor rearrangements (8–10).
Mi-2/nucleosome remodeling deacetylase (Mi-2/NuRD) com-plexes constitute a second class of heterogeneous CRCs that areprimarily associated with gene silencing (11–13). Mi-2/NuRD com-plexes include the SNF2-related ATPase Mi-2� (Chd4) togetherwith histone deacetylases (HDAC1 and HDAC2), histone bind-ing proteins (RbAp46 and RbAp48), metastasis-associated(MTA) proteins 1, 2, or 3 and methyl(CpG)-binding domain 3(MBD3) proteins (14, 15). Mi-2/NuRD induces chromatin com-paction via the coupling of nucleosome sliding and histone deacety-lase activities. Moreover, Mi-2/NuRD associates with methylatedDNA (16). In these capacities, Mi-2/NuRD has been implicated inthe control of both B- and T-cell differentiation (15, 17, 18).
We investigated roles of SWI/SNF and Mi-2/NuRD complexesin the activation of the mb-1 promoter by EBF and Pax5. Here,we show that the activation of mb-1 promoters by EBF and Pax5is SWI/SNF-dependent. Activation of mb-1 promoters by EBFand Pax5 is restrained by Mi-2/NuRD, which limits the extent ofchromatin remodeling and DNA demethylation.
Results4-OHT-Dependent EBF and Pax5. We produced retroviral vectors forexpression of FLAG-tagged EBF (18–591) or Pax5 (1–381) asfusion proteins with the ligand-binding domain (residues 282–595) of human estrogen receptor � (ER-LBD). The mutatedER-LBD used here is activated by the synthetic estrogen 4-hydroxytamoxifen (4-OHT) (19), but the domain does notinclude sequences that are important for recruitment of SWI/SNF or Mi-2/NuRD by ER� (20).
We demonstrated previously that enforced expression of Pax5is sufficient for activation of endogenous mb-1 gene transcriptionin the �M.10 subclone of 558L�M plasmacytoma cells (21). Onaverage, each of these cells possesses 1 hypomethylated mb-1promoter that is accessible to Pax5 in the absence of EBF.Membrane-bound IgM (mIgM) is displayed on �M.10 cells indirect proportion to mb-1 gene transcripts. We expressedPax5:ER in �M.10 cells using MSCV-Pax5:ER-YFP virus. YFP�
cells were purified using fluorescence activated cell sorting(FACS) and incubated for 48 h without or with 0.5 �M 4-OHT.In the absence of 4-OHT (Fig. 1A), mIgM� cells were detectedat a low frequency (2.6%) and with a low mean fluorescence
Author contributions: H.G., K.L., J.R., S.F., and J.H. designed research; H.G., K.L., J.R., S.F.,and D.L. performed research; H.G., K.L., J.R., S.F., and J.H. analyzed data; and H.G., K.L., J.R.,and J.H. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0809485106/DCSupplemental.
11258–11263 � PNAS � July 7, 2009 � vol. 106 � no. 27 www.pnas.org�cgi�doi�10.1073�pnas.0809485106

intensity (MFI) of mIgM staining (MFI � 3.1). Both reflect lowlevels of mb-1 gene transcripts. In contrast, 4-OHT stimulatedmIgM expression on 28.8% of the cells (MFI � 17.9), indicatinga significant increase of mb-1 transcripts in response to Pax5:ER.
To generate cells expressing EBF:ER, we expressed the fusionprotein stably in the 558L�M subclone �M.2 (21). Unlike �M.10cells, �M.2 cells are refractory to activation by Pax5 becausetheir mb-1 promoters are uniformly hypermethylated in rela-tively inaccessible chromatin (21, 22). Following incubation with4-OHT (Fig. 1B), �M.2 cells expressing EBF:ER exhibitedminimal mIgM expression (1.1% mIgM� cells; MFI � 3.9). Inthe absence of 4-OHT, expression of Pax5 in these cells resultedin few mIgM� cells (3.5% mIgM� cells; MFI � 4.7). However,in the presence of both EBF:ER and Pax5, incubation with4-OHT increased mIgM� cells (37.8%) and the levels of mIgMthat they express (MFI � 121). In other experiments, EBF orEBF:ER exhibited nearly identical abilities to increase mIgMsynergistically with Pax5 (Fig. S1). Therefore, synergistic acti-vation of mb-1 gene expression by the EBF:ER fusion protein isdependent on 4-OHT and Pax5.
Roles of CRCs in Transcriptional Activation of the mb-1 Gene. Wetested whether transcriptional activation of mb-1 promoters byEBF and Pax5 requires CRCs. For these experiments we used�M.2 cells that stably expressed EBF:ER (�M.2�EBF:ERcells). The cells were infected with control YFP or Pax5:ER-YFPretroviruses. YFP� cells were collected 48 h after infection. Thepurified cells were infected to express previously validatedshRNAs specific for a sequence that is conserved between Brg1and Brm transcripts, or specific for mRNA encoding Mi-2�(Chd4; referred to hereafter as Mi-2�) (23). All cells wereincubated with 4-OHT for the final 48 h of culture. Afterexpression of shRNAs for 3 or 4 days, Western analysis dem-onstrated that Brg1 and Brm (Fig. 2A), or Mi-2� (Fig. 2B),proteins were depleted efficiently in the presence of shRNAs.Western blotting also confirmed the expression of EBF:ER and
Pax5:ER. Co-expression with Pax5:ER consistently reducedexpression of EBF:ER in the presence of 4-OHT.
We next examined the induction of mIgM on cells expressingEBF:ER, Pax5:ER, and/or shRNAs in the presence of 4-OHT(Fig. 2C). Expression of Pax5:ER alone in �M.2 cells resulted in9.4% mIgM� (MFI � 12.6) cells (increased from 3.3% obtainedwith control retroviruses). In the absence of Pax5:ER, only asmall fraction (5.7%; MFI � 4.8) of EBF:ER-expressing cellsdisplayed mIgM in the presence of 4-OHT. In contrast, co-expression of EBF:ER and Pax5:ER in the presence of 4-OHTresulted in 60.7% mIgM� cells (MFI � 76.2), which, althoughreduced somewhat in intensity, is similar to results obtainedusing unmodified EBF and Pax5 (Fig. S1). Next, we examinedwhether transcriptional activation in response to EBF:ER and/orPax5:ER is dependent on SWI/SNF. The Brg1/Brm-specificshRNA reduced the percentage of mIgM� cells induced byeither factor, separately or together. In the presence of Pax5:ER,EBF:ER, or both factors together, the shRNA reduced thefraction of mIgM� cells to 1.9% (MFI � 2.7), 2.8% (MFI � 3.8)or 11.7% (MFI � 7.6), respectively (Fig. 2C). Also, the back-ground expression of mIgM� was reduced by the shRNA alone(from 3.3% to 0.5%). We conclude that EBF and Pax5 bothrequire SWI/SNF for mb-1 promoter activation.
In contrast to the inhibitory effects of knocking down Brg1 andBrm, the knock-down of Mi-2� enhanced the frequency of mIgM�
cells under all conditions (Fig. 2D). In �M.2 cells alone, theMi-2�-specific shRNA had only a small effect on the frequencyof mIgM� cells (increased from 3.3% to 7.8%). In the presence ofMi-2� shRNA, Pax5:ER and 4-OHT increased the frequency ofmIgM� cells from 7.8% to 27.4% and the MFI from 5.2 to 38.Therefore, Mi-2� knock-down greatly enhanced induction of mIgMexpression mediated by Pax5:ER. The knock-down of Mi-2� alsoincreased the percentage of mIgM� cells in the presence ofEBF:ER (from 5.7% to 21.1%; MFI � 9.3)(Fig. 2 C and D).Strikingly, when �M.2 cells expressed EBF:ER, Pax5:ER andMi-2� shRNA, mIgM was detected on nearly all infected cells(82.2%; MFI � 484). The MFI of these cells was much greater thanthat obtained in the presence of EBF:ER, Pax5:ER and normallevels of Mi-2� (MFI � 76.2). These results indicate that Mi-2/NuRD complexes attenuate the activation of mb-1 promoters byPax5:ER, EBF:ER or by both factors together.
To assess effects of the early B cell-specific factors and CRCson mb-1 transcription, we isolated RNA from 4-OHT-treatedcell cultures and quantitated steady state levels of mb-1 mRNAtranscripts using quantitative PCR (qPCR). Relative to unin-fected cells (Fig. 2E), mb-1 transcript levels were unchanged in�M.2 cells in the absence of EBF:ER and Pax5:ER, regardlessof the status of Brg1/Brm or Mi-2� (1.7- or 1.1-fold withknock-downs, respectively). Expression of Pax5:ER alone re-sulted in increased mb-1 transcripts (5.7-fold relative to control;P � 0.005) that were dependent on Brg1/Brm (reduced to2.4-fold in the presence of Brg1/Brm shRNA). As expected fromthe flow cytometry data, expression of Pax5:ER together withthe knock-down of Mi-2� increased transcript levels to 18.4-fold(P � 0.004). In the presence of EBF:ER alone, mb-1 transcrip-tion was stimulated weakly (2.3-fold) relative to that in control�M.2 cells. The weak activation by EBF:ER was relativelyunaffected by the knock-down of Brg1/Brm (2.2-fold); however,the knock-down of Mi-2� increased activation to 9.8-fold (P �0.0003). Synergistic effects were observed in the presence ofEBF:ER and Pax5:ER, which strongly increased mb-1 transcripts(70.4-fold; P � 0.002). This was greatly reduced by the knock-down of Brg1/Brm (to only 8.0-fold; P � 0.002). Significantly, inthe presence of EBF:ER and Pax5:ER, the knock-down of Mi-2�increased mb-1 transcripts 1,727-fold (P � 0.002).
Previously, we concluded that EBF increases the accessibilityof mb-1 promoter DNA (22). In turn, this facilitates DNAbinding and activation of the promoter by Pax5. To determine
Fig. 1. Tamoxifen-dependent activation of mb-1 gene expression. (A)Pax5:ER increases the display of mIgM on �M.10 cells. Cells were infected withPax5:ER-YFP retroviruses. YFP� cells were sorted and 0.5 �M 4-OHT was addedat 48 h posttransduction. 48 h after the addition of 4-OHT, cell surfaceexpression of mIgM (which directly reflects mb-1 expression) was analyzed bylabeling cells with biotinylated anti-IgM and streptavidin-conjugated APC.mIgM was detected using a FACScalibur™ flow cytometer. (B) Synergisticactivation of mb-1 transcription by EBF:ER and Pax5 in �M.2 cells. Cells werestably transfected to express EBF:ER (�M.2�EBF:ER cells), then infected withMSCV-Pax5-GFP retroviruses. Cells are shown following incubation with orwithout 4-OHT and mIgM staining as in A.
Gao et al. PNAS � July 7, 2009 � vol. 106 � no. 27 � 11259
IMM
UN
OLO
GY

whether changes in DNA accessibility are mediated by CRCs, wemeasured the relative accessibility of a Sau96I restriction site inmb-1 promoters in intact nuclei (the location of the Sau96I siteis indicated in Fig. 3A). Nuclei were obtained from 4-OHT-treated cells and incubated with a limiting concentration ofSau96I enzyme (Test). Signals obtained from completely cleavedDNA are shown (Control). As demonstrated in Fig. 2F, mb-1promoters in un-manipulated �M.2 cells were relatively inac-cessible to Sau96I digestion (arbitrarily assessed as 1.0). Knock-downs of Brg1/Brm or Mi-2� affected mb-1 promoter accessi-bility only slightly in the absence of EBF:ER or Pax5:ER (1.0- or1.5-fold, respectively). Activation of EBF:ER increased accessi-bility by 2.9-fold. This activity was decreased by the knock-downof Brg1/Brm (reduced to 1.1-fold). We conclude that EBF-dependent chromatin remodeling is dependent on SWI/SNF. Incontrast, the knock-down of Mi-2� increased accessibility in-duced by EBF:ER (to 5.9-fold). Accessibility was not affected byPax5:ER alone (1.4-fold). In the presence of Pax5:ER, accessi-bility was enhanced 2.4-fold by the knock-down of Mi-2�. We
Fig. 2. mb-1 gene activation in the context of EBF, Pax5 and knock-down ofchromatin remodeling complexes (CRCs). (A) Western detection of Brg1, Brm,EBF:ER, and Pax5:ER in �M.2 or �M.2�EBF:ER cells. Cells were infected se-quentially with Pax5:ER-YFP or YFP control retroviruses and Brg1/Brm-specificshRNA-GFP or control GFP retroviruses. GFP�YFP� cells were purified on day 2postinfection with GFP/shRNA-GFP viruses. 0.5 �M 4-OHT was added for thefinal 48 h. Western blotting was performed on the sorted cells at day 3 or 4 ofshRNA expression. EBF:ER and Pax5:ER were detected using anti-ER� serum.USF1 served as a loading control. (B) Western detection of Mi-2�, EBF:ER andPax5:ER in �M.2 or �M.2�EBF:ER cells. Other aspects of this experiment aresimilar to (A). (C) mIgM expression in response to EBF:ER, Pax:ER without orwith Brg1/Brm shRNA. In this and all subsequent experiments, cells werelabeled with Cy5-conjugated anti-IgM and detected using a CyAn™ flowcytometer. (D) mIgM expression in response to Mi-2� shRNA without or withEBF:ER, Pax:ER or both factors. (E) Quantitative RT-PCR analysis of mb-1transcripts for the cells in (C and D). All data were obtained from sortedYFP�GFP� cells and normalized relative to �-actin transcripts (n � 3) (mean �SEM.). (F) Accessibility of mb-1 promoter chromatin in response to EBF:ER,Pax5:ER and shRNA. Purified YFP�GFP� cells were incubated 48 h with 4-OHT.(C and D). Relative cleavage by Sau96I (test; see Fig. 3A for location) wasdetected using LM-PCR. Total cleavable DNA was measured following diges-tion of samples to completion (control). Exposure of control gel was one-fifththat of test gel exposure.
Fig. 3. Knockdown of Mi-2� increases and propagates EBF- and Pax5-dependent DNA demethylation. (A) Positions of EBF, Pax5/Ets and proximalpromoter factor binding sites are indicated relative to CpGs1–8. Binding sitesof other factors present in early B cells (Runx1/CBF�, E2A), but not plasmacy-toma cells, are not shown. The location of the Sau96I site used to measurechromatin accessibility is indicated. (B) DNA methylation of mb-1 promoters isdecreased synergistically by EBF:ER, Pax5:ER, and Mi-2� shRNA in �M.2 cells.All cells were cultured 48 h in the presence of 4-OHT. Genomic DNA wasrecovered from �M.2 cells without EBF:ER (�EBF:ER) or �M.2 �EBF:ER cells asshown and converted with sodium bisulfite before amplification and subclon-ing of mb-1 promoters. The frequency of methylated CpGs at each of the eightCpGs is shown for 20 sequenced clones (black bars). (C) The lack of DNAmethylation correlates with high levels of mIgM. �M.2 cells expressing EBF:ER,Pax:ER, and Mi-2� shRNA were sorted to recover the highest and lowest 5% ofmIgM� cells. Bisulfite converted mb-1 promoters were amplified and twentyindividual clones were sequenced as in B. Methylated (black circles) andunmethylated (open circles) CpGs are indicated in each individual sequence.
11260 � www.pnas.org�cgi�doi�10.1073�pnas.0809485106 Gao et al.

conclude that Pax5, by itself, is less able to modulate accessibilitythan EBF. However, EBF:ER and Pax5:ER increased accessi-bility in concert (3.8-fold relative to control �M.2 cells). Thisincrease was blocked by the knock-down of Brg1/Brm (reducedto 1.3-fold), but the knock-down of Mi-2� increased accessibilitypotently when both EBF:ER and Pax5:ER were active (10.4-fold).
We performed chromatin immunoprecipitation (ChIP) anal-yses to measure changes in factor occupancy of mb-1 promotersin response to EBF:ER, Pax5:ER and Mi-2� shRNA. As ex-pected from the increase in mb-1 transcripts, ChIP assaysdetected increased occupancy of mb-1 promoters in �M.2 cellsby RNA polymerase II (pol II) in the presence of these inducers(Fig. S2 A). We also detected the association of Brg1 with mb-1promoters, confirming that it is recruited in the course of mb-1promoter activation. Notably, the knock-down of Mi-2� in-creased detection of associated Brg1. Increased detection of polII and Brg1 correlated with reduced levels of histone H3 at mb-1promoters in cells expressing EBF:ER, Pax5:ER and Mi-2�shRNA (Fig. S2B). This suggests that nucleosomes are reposi-tioned or removed during mb-1 transcriptional activation.
The question arose as to whether other genes are repro-grammed similarly in �M.2 cells by EBF, Pax5 and CRCs.Previous reports demonstrated that activation of murine andhuman promoters of Cd19 genes, which encode a costimulatoryreceptor on B cells, is dependent on EBF and Pax5 (24–26).Therefore, we measured Cd19 transcripts in �M.2 cells express-ing EBF:ER, Pax5:ER, or both factors in the presence ofBrg1/Brm- or Mi-2�-specific shRNAs and 4-OHT (Fig. S3).Similar to the control of mb-1 expression, EBF:ER and Pax5:ERincreased endogenous Cd19 transcripts synergistically (23.5-fold;P � 0.03). The increase was ablated completely in the presenceof Brg1/Brm shRNA, indicating a requirement for SWI/SNF.Inclusion of Mi-2� shRNA significantly increased Cd19 tran-scripts (to 52.2-fold; P � 0.07). However, the expression ofEBF:ER is increased similarly in cells expressing Mi-2� shRNA.The reduced enhancement of Cd19 versus mb-1 transcripts byMi-2� shRNA in the presence of EBF and Pax5 may reflectdifferences in chromatin structure and/or requirements foractivation of their respective promoters in �M.2 cells.
Demethylation of mb-1 Promoters in Response to EBF and Pax5 IsRestrained by Mi-2/NuRD. Our previous studies demonstrated thatmb-1 promoter activation is blocked, in part, by DNA methyl-ation because 5-methylcytosine (5-meC) interferes with Pax5’srecruitment of Ets transcription factors to bind mb-1 promotersin �M.2 cells (21, 22). Our observation that mb-1 transcripts areincreased efficiently by EBF:ER, Pax5:ER and the knock-downof Mi-2� suggested that these conditions result in demethylationof mb-1 promoters. Therefore, we measured the frequency of5-meC at 8 positions in mb-1 promoters under various conditionsin the presence of 4-OHT (Fig. 3 A and B). Bisulfite-treated mb-1promoter antisense DNA strands were amplified, cloned, andsequenced. Activation of EBF:ER (with or without Pax5:ER)increased demethylation of CpG-1 and CpG-2. Significantly,with expression of EBF:ER, Pax5:ER and Mi-2� shRNA, CpG-1and CpG-2 were almost completely demethylated, while demeth-ylation of CpG-3 and CpG-4 was substantially increased. Overall,these conditions resulted in the graded loss of methylation from5� to 3� ends of mb-1 promoters.
We next examined patterns of methylation of individual mb-1promoters in cells exhibiting different levels of mIgM expression.�M.2 cells expressing EBF:ER, Pax5:ER and Mi-2� shRNAwere incubated with 4-OHT and stained to detect mIgM. UsingFACS, we purified cell populations that expressed maximal (top5%) or minimal (bottom 5%) mIgM. We sequenced 20 individ-ual bisulfite-treated clones from each of these populations (Fig.3C). The data confirmed that mb-1 transcription is increased in
proportion to the overall demethylation of mb-1 promoters. Onlycells expressing high levels of mIgM exhibited extensive demeth-ylation of the Pax5-dependent Ets binding site (overlapping withCpG-3) and other downstream CpGs. CpG-5, which is nestedwithin a site known to bind Sp1 (27), was demethylated with thelowest efficiency. Four of 20 sequences derived from cellsexpressing high levels of mIgM were demethylated completely.We conclude that EBF:ER and Pax5:ER induce mb-1 promoterdemethylation, but they do so inefficiently in the presence ofMi-2/NuRD.
To further investigate the mechanism of mb-1 promoterdemethylation in response to EBF and Pax5, we asked whetherscheduled DNA replication (i.e., S phase of the cell cycle) isrequired. We performed experiments similar to Fig. 2D, but inthe presence or absence of the DNA cross-linker mitomycin C(MitC). After 48 h incubation with 4-OHT, the generation ofmIgM� cells was unaffected by MitC (Fig. S4A), which reducedDNA synthesis by more than 76% (Fig. S4B). Moreover, mb-1promoter demethylation in response to EBF:ER, Pax5:ER andMi-2� shRNA was similar in the absence or presence of MitC(Fig. S4C). Together, these data suggest that mb-1 promoterdemethylation is an active process that does not require sched-uled DNA replication.
Initiation of mb-1 Promoter Demethylation by EBF in ex Vivo B CellProgenitors. We previously demonstrated that mb-1 promoters arecompletely unmethylated in pro-B cells and subsequent stages ofB-cell development (22), whereas, only hypermethylated mb-1promoters were detected in B cell progenitors from EBF-deficientmice. Therefore, to study DNA demethylation in cells ex vivo weused cytokine-dependent B cell progenitors from E � 14.5 Ebf1�/�
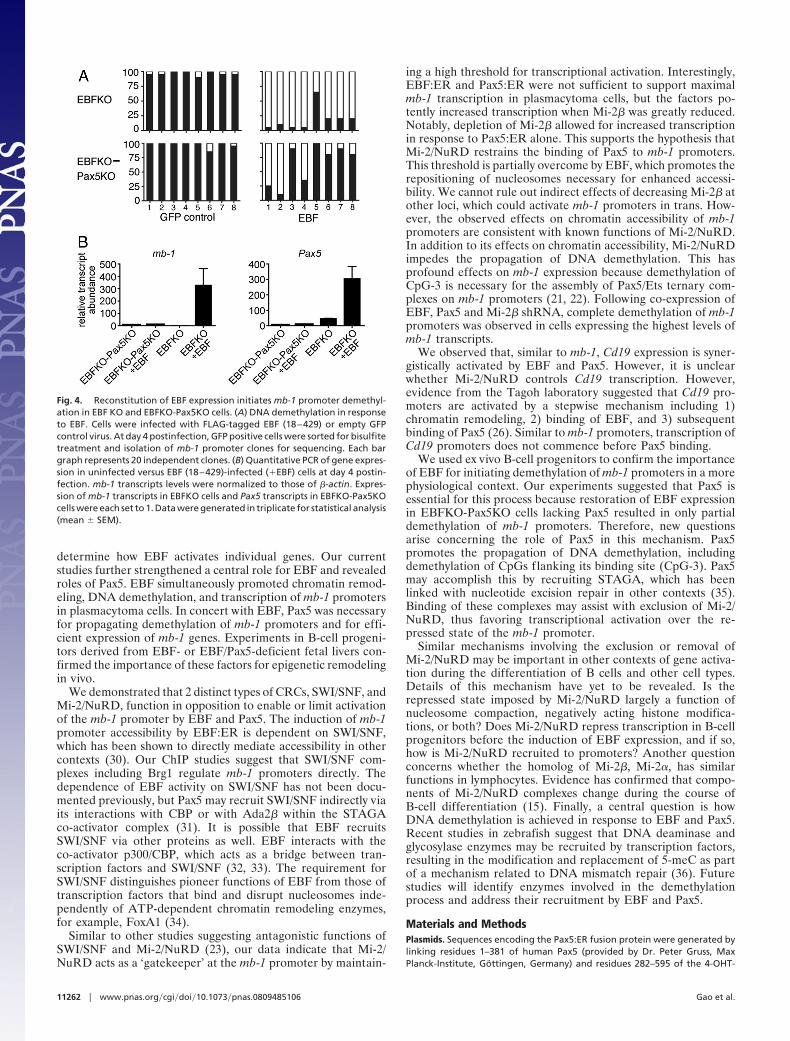
(EBFKO) or Ebf1�/�Pax5�/� (EBFKO-Pax5KO) fetal livers. Wegenerated B220�CD43�CD117�CD28loCD19�CD24�BP-1�IgM� cells, which represent EBFKO or EBFKO-Pax5KO ‘B-biased progenitor’ cell populations. As expected for cells lackingEBF, we detected only hypermethylated mb-1 promotersin EBFKO or EBFKO-Pax5KO cells (Fig. 4A). Next, we infectedEBFKO or EBFKO-Pax5KO cells with retroviruses for expressionof EBF (18–429) (28). After 4 days, infected cells were purified foranalysis of mb-1 promoter DNA methylation. Clones obtained fromGFP� control cells were uniformly hypermethylated. However,expression of EBF (18–429) resulted in nearly complete demeth-ylation of mb-1 promoters in EBFKO cells (except for CpG-5).Although expression of EBF (18–429) in EBFKO-Pax5KO cellsresulted in significant demethylation of CpG-1 -2, and -4, demeth-ylation was not propagated to 3� ends of mb-1 promoters. Notably,EBFKO-Pax5KO cells failed to demethylate CpG-3, which coin-cides with the Pax5-dependent Ets binding site. Analysis of mb-1transcripts in EBFKO-Pax5KO cells demonstrated a correlationbetween inefficient mb-1 promoter demethylation and the absenceof mb-1 transcripts (Fig. 4B). However, EBF increased chromatinaccessibility in these cells by 3.5-fold (Fig. S5). In contrast, the morecomplete demethylation of mb-1 promoters in EBFKO cells re-constituted with EBF correlated with greatly increased (327-fold;P � 0.07) mb-1 gene transcription in these cells. EBF (18–429) alsoincreased endogenous Pax5 transcripts (6.7-fold; P � 0.04) inEBFKO cells. Taken together, our data suggest that EBF and Pax5drive demethylation and transcription of mb-1 promoters synergis-tically during early B-cell development.
DiscussionThe specification of developmental programs by transcriptionfactors requires epigenetic changes necessary for the activationof silent genes. Our studies suggest that EBF initiates epigeneticreprogramming in the course of B-cell lineage specification. EBFis expressed at low levels in common lymphoid progenitors(CLPs) and drives their differentiation to become B cells (29).To address how EBF accomplishes this, we have sought to
Gao et al. PNAS � July 7, 2009 � vol. 106 � no. 27 � 11261
IMM
UN
OLO
GY
determine how EBF activates individual genes. Our currentstudies further strengthened a central role for EBF and revealedroles of Pax5. EBF simultaneously promoted chromatin remod-eling, DNA demethylation, and transcription of mb-1 promotersin plasmacytoma cells. In concert with EBF, Pax5 was necessaryfor propagating demethylation of mb-1 promoters and for effi-cient expression of mb-1 genes. Experiments in B-cell progeni-tors derived from EBF- or EBF/Pax5-deficient fetal livers con-firmed the importance of these factors for epigenetic remodelingin vivo.
We demonstrated that 2 distinct types of CRCs, SWI/SNF, andMi-2/NuRD, function in opposition to enable or limit activationof the mb-1 promoter by EBF and Pax5. The induction of mb-1promoter accessibility by EBF:ER is dependent on SWI/SNF,which has been shown to directly mediate accessibility in othercontexts (30). Our ChIP studies suggest that SWI/SNF com-plexes including Brg1 regulate mb-1 promoters directly. Thedependence of EBF activity on SWI/SNF has not been docu-mented previously, but Pax5 may recruit SWI/SNF indirectly viaits interactions with CBP or with Ada2� within the STAGAco-activator complex (31). It is possible that EBF recruitsSWI/SNF via other proteins as well. EBF interacts with theco-activator p300/CBP, which acts as a bridge between tran-scription factors and SWI/SNF (32, 33). The requirement forSWI/SNF distinguishes pioneer functions of EBF from those oftranscription factors that bind and disrupt nucleosomes inde-pendently of ATP-dependent chromatin remodeling enzymes,for example, FoxA1 (34).
Similar to other studies suggesting antagonistic functions ofSWI/SNF and Mi-2/NuRD (23), our data indicate that Mi-2/NuRD acts as a ‘gatekeeper’ at the mb-1 promoter by maintain-
ing a high threshold for transcriptional activation. Interestingly,EBF:ER and Pax5:ER were not sufficient to support maximalmb-1 transcription in plasmacytoma cells, but the factors po-tently increased transcription when Mi-2� was greatly reduced.Notably, depletion of Mi-2� allowed for increased transcriptionin response to Pax5:ER alone. This supports the hypothesis thatMi-2/NuRD restrains the binding of Pax5 to mb-1 promoters.This threshold is partially overcome by EBF, which promotes therepositioning of nucleosomes necessary for enhanced accessi-bility. We cannot rule out indirect effects of decreasing Mi-2� atother loci, which could activate mb-1 promoters in trans. How-ever, the observed effects on chromatin accessibility of mb-1promoters are consistent with known functions of Mi-2/NuRD.In addition to its effects on chromatin accessibility, Mi-2/NuRDimpedes the propagation of DNA demethylation. This hasprofound effects on mb-1 expression because demethylation ofCpG-3 is necessary for the assembly of Pax5/Ets ternary com-plexes on mb-1 promoters (21, 22). Following co-expression ofEBF, Pax5 and Mi-2� shRNA, complete demethylation of mb-1promoters was observed in cells expressing the highest levels ofmb-1 transcripts.
We observed that, similar to mb-1, Cd19 expression is syner-gistically activated by EBF and Pax5. However, it is unclearwhether Mi-2/NuRD controls Cd19 transcription. However,evidence from the Tagoh laboratory suggested that Cd19 pro-moters are activated by a stepwise mechanism including 1)chromatin remodeling, 2) binding of EBF, and 3) subsequentbinding of Pax5 (26). Similar to mb-1 promoters, transcription ofCd19 promoters does not commence before Pax5 binding.
We used ex vivo B-cell progenitors to confirm the importanceof EBF for initiating demethylation of mb-1 promoters in a morephysiological context. Our experiments suggested that Pax5 isessential for this process because restoration of EBF expressionin EBFKO-Pax5KO cells lacking Pax5 resulted in only partialdemethylation of mb-1 promoters. Therefore, new questionsarise concerning the role of Pax5 in this mechanism. Pax5promotes the propagation of DNA demethylation, includingdemethylation of CpGs flanking its binding site (CpG-3). Pax5may accomplish this by recruiting STAGA, which has beenlinked with nucleotide excision repair in other contexts (35).Binding of these complexes may assist with exclusion of Mi-2/NuRD, thus favoring transcriptional activation over the re-pressed state of the mb-1 promoter.
Similar mechanisms involving the exclusion or removal ofMi-2/NuRD may be important in other contexts of gene activa-tion during the differentiation of B cells and other cell types.Details of this mechanism have yet to be revealed. Is therepressed state imposed by Mi-2/NuRD largely a function ofnucleosome compaction, negatively acting histone modifica-tions, or both? Does Mi-2/NuRD repress transcription in B-cellprogenitors before the induction of EBF expression, and if so,how is Mi-2/NuRD recruited to promoters? Another questionconcerns whether the homolog of Mi-2�, Mi-2�, has similarfunctions in lymphocytes. Evidence has confirmed that compo-nents of Mi-2/NuRD complexes change during the course ofB-cell differentiation (15). Finally, a central question is howDNA demethylation is achieved in response to EBF and Pax5.Recent studies in zebrafish suggest that DNA deaminase andglycosylase enzymes may be recruited by transcription factors,resulting in the modification and replacement of 5-meC as partof a mechanism related to DNA mismatch repair (36). Futurestudies will identify enzymes involved in the demethylationprocess and address their recruitment by EBF and Pax5.
Materials and MethodsPlasmids. Sequences encoding the Pax5:ER fusion protein were generated bylinking residues 1–381 of human Pax5 (provided by Dr. Peter Gruss, MaxPlanck-Institute, Gottingen, Germany) and residues 282–595 of the 4-OHT-
Fig. 4. Reconstitution of EBF expression initiates mb-1 promoter demethyl-ation in EBF KO and EBFKO-Pax5KO cells. (A) DNA demethylation in responseto EBF. Cells were infected with FLAG-tagged EBF (18–429) or empty GFPcontrol virus. At day 4 postinfection, GFP positive cells were sorted for bisulfitetreatment and isolation of mb-1 promoter clones for sequencing. Each bargraph represents 20 independent clones. (B) Quantitative PCR of gene expres-sion in uninfected versus EBF (18–429)-infected (�EBF) cells at day 4 postin-fection. mb-1 transcripts levels were normalized to those of �-actin. Expres-sion of mb-1 transcripts in EBFKO cells and Pax5 transcripts in EBFKO-Pax5KOcells were each set to 1. Data were generated in triplicate for statistical analysis(mean � SEM).
11262 � www.pnas.org�cgi�doi�10.1073�pnas.0809485106 Gao et al.

responsive ER-LBD (provided by D. Metzger, IGBMC Recherche, Strasbourg,France) with the linker GALTGALTGAI. The fusion protein sequences wereinserted into XhoI sites of MSCV-IRES-GFP or MSCV-IRES-YFP vectors (providedby P. Marrack, National Jewish Health, Denver, CO). FLAG-tagged EBF:ER wasmade by linking FLAG-EBF (18–591) and the ER-LBD with DAGALTGALTEAI.The fusion protein sequences were inserted into the PmeI site of the Ig�-basedexpression vector VLPE�2.13 (37) to make V�-FLAG-EBF:ER-E�. Retroviral plas-mids for expression of Brg1/Brm- or Mi-2�-specific shRNAs were provided byS. T. Smale (UCLA, Los Angeles, CA) (23). The MSCV-FLAG-EBF (18–429)-GFPand MSCV-Pax5-GFP plasmids were reported previously (28). All plasmids weresequenced.
Cell lines, Transfection, Retroviral Infection, and Flow Cytometry. �M.2 and�M.10 cells were cultured as described previously (21). To create the �M.2 cellline expressing FLAG-EBF:ER, �M.2 cells were electroporated with 3 �g ofpPUR (Clontech) and 20 �g of V�-FLAG-EBF:ER-E� and cultured as previouslyreported (22). Generation of retroviruses and infection of cells were describedpreviously (21, 22). mIgM was detected using biotinylated anti-IgM (CaltagLaboratories) and streptavidin-conjugated allophycocyanin (APC) and de-tected using a FACScalibur™ flow cytometer (BD). Alternatively, mIgM waslabeled with Cy5-conjugated anti-IgM (Jackson Research Laboratories, WestGrove, PA) and detected using a CyAn™ flow cytometer (Dako). ER fusionproteins were induced using 0.5 �M 4-OHT (Sigma-Aldrich). Cell sorting wasperformed as described (21, 22).
Western Blotting. Whole cell protein extraction and western blotting wereperformed as described (32, 33). Rabbit anti-Mi-2� antibody was kindly pro-vided by S. T. Smale (Univ. Calif., Los Angeles, CA). Rabbit ER�- and USF1-specific antibodies were purchased from Santa Cruz Biotechnology. Rabbit
anti-Brg1 and mouse anti-Brm were purchased from Upstate USA, Inc. and BDBioSciences, respectively.
Analysis of mRNA Transcripts, DNA Methylation, and Chromatin Accessibility.Isolation of total RNA and qPCR were reported previously (28). Data wereanalyzed for significance using Student’s 1-tailed t test. Sodium bisulfitetreatment, PCR amplification, subcloning, and sequencing of mb-1 promoterantisense strands were performed using 2 � 104 cells as previously described(21). Chromatin accessibility assays were performed as described (22), exceptthat 10 U of Sau96I was used in all test digestion reactions.
Derivation and Maintenance of Fetal Liver B-Cell Progenitors. Mice were bredand maintained in the Biological Resource Center at National Jewish Health(NJH). All studies were approved by the NJH Institutional Animal Care and UseCommittee. Ebf1�/� mice were made by R. Grosschedl (Max Planck-Institute ofImmunobiology, Freiburg, Germany) and provided by Y. Zhuang (Duke Uni-versity, Durham, NC). Pax5�/� mice were made and provided by M. Busslinger(Research Institute of Molecular Pathology, Vienna, Austria). Lineage marker-negative (Lin�) cells were isolated from E � 14.5 Ebf1�/� and Ebf1�/�Pax5�/�
fetal livers (4, 38) by negative MACS separation (Miltenyi Biotec) as described(39). Lin� cells were expanded and maintained in the absence of stromal cellsin Iscove’s modified Dulbecco’s medium containing 10% FBS (HyClone), 50�g/mL gentamicin, 1� GlutaMAX-1 (Invitrogen) and recombinant (r) IL-7 (3.3ng/mL), rSCF (3.3 ng/mL), and rFLT3 ligand (3.3 ng/mL) (R&D Systems).
ACKNOWLEDGMENTS. We thank C. Murre and S. T. Smale for helpful discus-sions; R. Grosschedl, Y. Zhuang and M. Busslinger for providing mice; and K.Tuttle for excellent technical assistance. This work was supported by NationalInstitutes of Health Grants R01 AI54661, R01 AI56322, and P01 AI22295 (to J.H.)and National Institute of Allergy and Infectious Diseases Training GrantT32-AI007405 (to S.F. and J. R.).
1. Nutt SL, Kee BL (2007) The transcriptional regulation of B-cell lineage commitment.Immunity 26:715–725.
2. Seet CS, Brumbaugh RL, Kee BL (2004) Early B cell factor promotes B lymphopoiesis withreduced interleukin-7 responsiveness in the absence of E2A. J Exp Med 199:1689–1700.
3. Medina KL, et al. (2004) Defining a regulatory network for specification of the B-cellfate. Dev Cell 7:607–617.
4. Lin H, Grosschedl R (1995) Failure of B-cell differentiation in mice lacking the tran-scription factor EBF. Nature 376:263–267.
5. Sigvardsson M, et al. (2002) Early B-cell Factor, E2A, and Pax-5 cooperate to activate theearly B cell-specific mb-1 promoter. Mol Cell Biol 22:8539–8551.
6. Hagman J, Lukin K (2005) Early B-cell Factor ‘pioneers’ the way to B-cell development.Trends Immunol 26:455–461.
7. Fry CJ, Peterson CL (2001) Chromatin remodeling enzymes: Who’s on first? Curr Biol11:R185–R187.
8. Chi TH, et al. (2002) Reciprocal regulation of CD4/CD8 expression by SWI/SNF-like BAFcomplexes. Nature 418:195–199.
9. Gebuhr TC, et al. (2003) The role of Brg1, a catalytic subunit of mammalian chromatin-remodeling complexes, in T-cell development. J Exp Med 198:1937–1949.
10. Osipovich O, et al. (2007) Essential function for SWI-SNF chromatin-remodeling com-plexes in the promoter-directed assembly of Tcrb genes. Nat Immunol 8:809–816.
11. Tong JK, et al. (1998) Chromatin deacetylation by an ATP-dependent nucleosomeremodelling complex. Nature 395:917–921.
12. Xue Y, et al. (1998) NURD, a novel complex with both ATP-dependent chromatin-remodeling and histone deacetylase activities. Mol Cell 2:851–861.
13. Zhang Y, LeRoy G, Seelig HP, Lane WS, Reinberg D (1998) The dermatomyositis-specificautoantigen Mi2 is a component of a complex containing histone deacetylase andnucleosome remodeling activities. Cell 95:279–289.
14. Ahringer J (2000) NuRD and SIN3 histone deacetylase complexes in development.Trends Genet 16:351–356.
15. Fujita N, et al. (2004) MTA3 and the Mi-2/NuRD complex regulate cell fate during Blymphocyte differentiation. Cell 119:75–86.
16. Zhang Y, et al. (1999) Analysis of the NuRD subunits reveals a histone deacetylase corecomplex and a connection with DNA methylation. Genes Dev 13:1924–1935.
17. Williams CJ, et al. (2004) The chromatin remodeler Mi-2beta is required for CD4expression and T-cell development. Immunity 20:719–733.
18. Naito T, Gomez-Del Arco P, Williams CJ, Georgopoulos K (2007) Antagonistic interac-tions between Ikaros and the chromatin remodeler Mi-2beta determine silenceractivity and Cd4 gene expression. Immunity 27:723–734.
19. Metzger D, Clifford J, Chiba H, Chambon P (1995) Conditional site-specific recombi-nation in mammalian cells using ligand-dependent chimeric Cre recombinase. ProcNatl Acad Sci USA 92:6991–6995.
20. Belandia B, Orford RL, Hurst HC, Parker M. G (2002) Targeting of SWI/SNF chromatinremodelling complexes to estrogen-responsive genes. EMBO J 21:4094–4103.
21. Maier H, Colbert J, Fitzsimmons D, Clark DR, Hagman J (2003) Activation of the earlyB cell-specific mb-1 (Ig-�) gene by Pax-5 is dependent on an unmethylated Ets bindingsite. Mol Cell Biol 23:1946–1960.
22. Maier H, et al. (2004) Early B-cell Factor cooperates with Runx1 and mediates epige-netic changes associated with mb-1 transcription. Nat Immunol 5:1069–1077.
23. Ramirez-Carrozzi VR, et al. (2006) Selective and antagonistic properties of SWI/SNF andMi-2� nucleosome remodeling complexes during an inflammatory response. GenesDev 20:282–296.
24. Kozmik Z, Wang S, Dorfler P, Adams B, Busslinger M (1992) The promoter of the CD19gene is a target for the B cell-specific transcription factor BSAP. Mol Cell Biol 12:2662–2672.
25. Gisler R, Åkerblad P, Sigvardsson M (1999) A human early B-cell factor-like proteinparticipates in the regulation of the human CD19 promoter. Mol Immunol 36:1067–1077.
26. Walter K, Bonifer C, Tagoh H (2008) Stem cell specific epigenetic priming and Bcell-specific transcriptional activation at the mouse Cd19 locus. Blood 112:1673–1682.
27. Travis A, Hagman J, Grosschedl R (1991) Heterogeneously initiated transcription fromthe pre-B- and B-cell-specific mb-1 promoter: Analysis of the requirement for upstreamfactor-binding sites and initiation site sequences. Mol Cell Biol 11:5756–5755.
28. Fields S, et al. (2008) The ‘zinc knuckle’ motif of Early B-cell Factor is required fortranscriptional activation of B cell-specific genes. Mol Immunol 45:3786–3796.
29. Zandi S, et al. (2008) EBF1 is essential for B-lineage priming and establishment of atranscription factor network in common lymphoid progenitors. J Immunol 181:3364–3372.
30. Cote J, Peterson CL, Workman JL (1998) Perturbation of nucleosome core structure bythe SWI/SNF complex persists after its detachment, enhancing subsequent transcrip-tion factor binding. Proc Natl Acad Sci USA 95:4947–4952.
31. Barlev NA, et al. (2003) A novel human Ada2 homologue functions with Gcn5 or Brg1to coactivate transcription. Mol Cell Biol 23:6944–6957.
32. Zhao F, McCarrick-Walmsley R, Åkerblad P, Sigvardsson M, Kadesch T (2003) Inhibitionof p300/CBP by early B-cell factor. Mol Cell Biol 23:3837–3846.
33. Agalioti T, et al. (2000) Ordered recruitment of chromatin modifying and generaltranscription factors to the IFN-� promoter. Cell 103:667–678.
34. Cirillo LA, et al. (2002) Opening of compacted chromatin by early developmentaltranscription factors HNF3 (FoxA) and GATA-4. Mol Cell 9:279–289.
35. Martinez E, et al. (2001) Human STAGA complex is a chromatin-acetylating transcrip-tion coactivator that interacts with pre-mRNA splicing and DNA damage-bindingfactors in vivo. Mol Cell Biol 21:6782–6795.
36. Rai T, Huggins IJ, James SR, Karpf AR, Jones DA, Cairns BR (2008) DNA demethylationinvolves the coupling of a deaminase, a glycosylase, and Gadd45. Cell 135:1201–1212.
37. Wagner DH Jr, et al. (1996) Thymocytes are rescued from glucocorticoid-mediated celldeath by CD28/CTLA-4 costimulatory interactions with B7–1/B7–2. J Exp Med 184:1631–1638.
38. Urbanek P, Wang Z-Q, Fetka I, Wagner EF, Busslinger M (1994) Complete block of earlyB-cell differentiation and altered patterning of the posterior midbrain in mice lackingPax5/BSAP. Cell 79:901–912.
39. Ikawa T, Kawamoto H, Wright LY, Murre C (2004) Long-term cultured E2A-deficienthematopoietic progenitor cells are pluripotent. Immunity 20:349–360.
Gao et al. PNAS � July 7, 2009 � vol. 106 � no. 27 � 11263
IMM
UN
OLO
GY