original effects of mir-223 on expression of il-1β and il
TRANSCRIPT

101
Abstract: MicroRNAs (miRNAs) are small non-coding RNAs that regulate post-transcriptional expression by translational inhibition or mRNA degradation. miRNAs bind to target mRNAs through partial complementarity, and can regulate many genes. In the present study, we investigated the effects of miR-223 on the expression of inflammatory cytokines in human gingival fibroblasts (HGF). To determine the effects of miR-223 on the expressions of interleukin-1β (IL-1β) and IL-6, HGF were stimu-lated by IL-1β (1 ng/mL) or tumor necrosis factor-α (TNF-α; 10 ng/mL) and transfected with a miR-223 expression plasmid. Levels of mRNA for IL-1β, IL-6, inhibitor of kappa-B kinase α (IKKα) and mitogen-activated protein kinase phosphatase-5 (MKP-5) were measured by real-time PCR, and levels IL-1β, IL-6 and IKKα protein were determined by enzyme-linked immunosorbent assay and Western blotting. Expression of IL-1β and IL-6 mRNAs was induced by IL-1β and TNF-α and further increased by miR-223 overexpression. IL-1β and TNF-α induced the expres-sion of IL-1β and IL-6 mRNAs, and this was reduced by miR-223 inhibitor. Overexpression of miR-223 decreased the levels of IKKα protein and MKP-5 mRNA in HGF. These findings indicate that miR-223 might control the inflammatory response via IKKα and MKP-5 in periodontal tissue.(J Oral Sci 58, 101-108, 2016)
Keywords: cytokine; gene expression; gingival fibroblast; inflammation; microRNA.
IntroductionPeriodontitis is a chronic inflammatory disease caused by periodontopathic bacteria and viruses. It affects the peri-odontal tissues, and when advanced can result in tissue destruction, occasional pain, alveolar bone resorption, and eventual tooth loss (1-3). The response to periodontal pathogens is determined by the nature and control of both adaptive and innate immune responses (4). Cyto-kines are central regulators of the immune responses that are produced by various cell types including fibroblasts, epithelial cells, macrophages, dendritic cells and T-helper cells in response to microbial contact (5). Inflammatory cytokines are regulated by activation of nuclear factor-kappa-light-chain-enhancer in activated B cells (NF-κB), interferon-regulatory factor transcription factors, and mitogen-activated protein kinases (MAPKs) (6,7). The NF-κB family is composed of five protein members; RelA (p65), RelB, c-Rel, NFκB1 (p105/p50), and NFκB2 ((p100/p52), which are synthesized as large precursors. NF-κB exists as a heterodimer consisting of p65 and p50 subunits that are associated with inhibitor of kappa-B (IκB) in the cytoplasm as an inactive form. IκB kinase (IKK) induces phosphorylation of IκB, and the phosphorylated IκB activates NF-κB to regulate the transcription of a variety of target genes, including interleukin-1β (IL-1β) and IL-6 (8-10). The IKK complex consists of three subunits each encoded by a separate gene: IKKα (also known as conserved helix-loop-helix ubiquitous kinase (CHUK)), IKKβ (also known as inhibitor of kappa light polypeptide gene enhancer in B-cells, kinase beta (IKBKB)), and IKKγ (also known
Journal of Oral Science, Vol. 58, No. 1, 101-108, 2016
Original
Effects of miR-223 on expression of IL-1β and IL-6 in human gingival fibroblasts
Sari Matsui1) and Yorimasa Ogata1,2)
1)Department of Periodontology, Nihon University School of Dentistry at Matsudo, Matsudo, Japan2)Research Institute of Oral Science, Nihon University School of Dentistry at Matsudo, Matsudo, Japan
(Received October 27, 2015; Accepted December 17, 2015)
Correspondence to Dr. Yorimasa Ogata, Department of Periodontology, Nihon University School of Dentistry at Matsudo, 2-870-1 Sakaecho-nishi, Matsudo, Chiba 271-8587, JapanFax: +81-47-360-9362 E-mail: [email protected]/10.2334/josnusd.58.101DN/JST.JSTAGE/josnusd/58.101

102
as IKBKG) (11). MAPKs include the p38, extracellular signal-regulated kinases (ERKs) and c-Jun N-terminal kinase (JNK) subfamilies, which control various cellular processes such as apoptosis, cell proliferation, differen-tiation, survival, inflammation and innate immunity (7). Activation of MAPKs is negatively regulated by MAPK phosphatases (MKPs; also known as dual specificity phosphatases (DUSPs)). MKPs regulate the inflam-matory response and are involved in the development of certain cancer types. There are 11 MKPs, which are divided into three distinct groups. The first group are the nuclear phosphatases, MKP-1/DUSP1, PAC-1/DUSP2, MKP-2/DUSP4 and DUSP5. The second group are the ERK-specific cytosolic phosphatases, MKP-3/DUSP6, MKP-X/DUSP7, and MKP-4/DUSP9. And the third group are the p38 MAPK/JUNK specific nuclear and cytosolic phosphatases, DUSP8, MKP-5/DUSP10, and MKP-7/DUSP16 (7,12,13).
MicroRNAs (miRNAs) are short, non-coding RNA molecules. They function as post-transcriptional regula-tors that bind to the 3'-untranslated regions (3'-UTRs) of target mRNAs, thereby inducing gene silencing. miRNAs play important roles in biological processes such as development, differentiation, cell proliferation and apoptosis. Several studies have demonstrated that a range of miRNAs are closely associated with inflammatory responses (14-16). We have previously reported that the three most overexpressed miRNAs in inflamed gingiva from Japanese patients with chronic periodontitis (CP) were hsa-miR-150, hsa-miR-223 and hsa-miR-200b, and that IL-1β and tumor necrosis factor-α (TNF-α) induced the expression of hsa-miR-150 in human gingival fibroblasts (HGF) (17). To clarify the mechanism whereby miRNA regulates the expression of inflammatory cytokines, we overexpressed hsa-miR-223 in HGF and examined the association between hsa-miR-223 and inflammatory cytokines.
Materials and MethodsReagentsDulbecco’s modified Eagle medium (DMEM), ISOGEN II, and recombinant human IL-1β were obtained from Wako (Tokyo, Japan). Recombinant human TNF-α was purchased from R&D Systems (Minneapolis, MN, USA). Fetal calf serum (FCS), Lipofectamine 2000, penicillin and streptomycin, and TrypLE Express were purchased from Invitrogen (Carlsbad, CA, USA). The PrimeScript RT reagent kit and SYBR Premix Ex Taq II were obtained from Takara (Tokyo, Japan). The miRNeasy Mini Kit was from Qiagen (Valencia, CA, USA). The Mir-X miRNA First-Strand Synthesis Kit and SYBR
Advantage qPCR Premix were purchased from Clontech (Mountain View, CA, USA). An expression plasmid for miRNA (pEZX-MR04) was obtained from GeneCopoeia (Rockville, MD, USA). miRCURY LNA inhibitor and miRCURY LNA inhibitor control were purchased from Exiqon (Woburn, MA, USA). Anti-rabbit IgG (whole molecule)-horseradish peroxidase antibody goat serum was from Sigma-Aldrich Japan (Tokyo, Japan). ELC plus Western Blotting Detection Reagent was purchased from GE Healthcare UK Ltd. (Buckinghamshire, UK).
Cell cultureHGF were cultured in 5% CO2 and 95% air at 37°C in DMEM medium containing 10% FCS as described previously (17-19). The Ethics Committee of Nihon University School of Dentistry at Matsudo approved the study (EC03-041, EC10-040, EC14-023), and a consent form was obtained from each patient after all the protocol had been explained. The cells were grown to confluence in 100-mm cell culture dishes. Twenty-four hours after plating, cells at 50-70% confluence were transfected with the control plasmid (pEZX-MR04; 3 μg), miRExpress precursor miRNA expression plasmid for miR-223 (3 μg), miRCURY LNA Inhibitor Control (10 nM) or miRCURY LNA Inhibitor for miR-223 (10 nM) using Lipofectamine 2000, then cultured for 12 h in DMEM without FCS, and stimulated with IL-1β (1 ng/mL) and TNF-α (10 ng/mL). Total RNA was purified from triplicate cultures after stimulation for 24 h.
Real-time polymerase chain reaction (PCR)Total RNA (1 μg) was used as a template for cDNA synthesis. cDNA was prepared using the PrimeScript RT reagent kit. Quantitative real-time PCR was per-formed using the following primer sets: IL-1β forward; 5'-CCAGGGACAGGTATGGAGCA-3', IL-1β reverse; 5'-TTCAACACGCAGGACAGGTACAG-3', IL-6 for-ward; 5'-AAGCCAGAGCTGTGCAGATGAGTA-3', IL-6 reverse; 5'-TGTCCTGCAGCCACTGGTTC-3', IKKα forward; 5'-TGCCTTGGCCATTTAAGCACTA-3' IKKα reverse; 5'-GGGACAGTGAACAAGTGACAACTC-3', MKP-5 forward; 5'-AGCGGCTGAACATCGGCTA-3', MKP-5 reverse; 5'-AGCCTGGCAGTGGATGAGAA-3', GAPDH forward; 5'-GCACCGTCAAGGCTGAGAAC-3', GAPDH reverse; 5'-ATGGTGGTGAAGACGC-CAGT-3', using the SYBR Premix Ex Taq II in a TP800 thermal cycler dice real-time system (Takara, Tokyo, Japan). The PCR reactions were performed in a total volume of 25 µL 2 × SYBR Premix Ex Taq II (12.5 μL), 10 μM forward and reverse primers and 50 ng cDNA for IL-1β and IL-6 and 10 ng cDNA for GAPDH. For

Fig. 1 Inflammatory cytokines induce miR-223 expression in HGF. Expression of miR-223 and U6 snRNA in HGF after stimulation with IL-1β (1 ng/mL), IL-6 (1 ng/mL) and TNF-α (10 ng/mL) for 24 h was measured by real-time PCR. Quantitative analyses of the datasets are shown with standard errors. Signifi-cant differences versus control. ***P < 0.001.
103
miRNA analysis, cDNA was synthesized using the Mir-X miRNA First-Strand Synthesis kit. Quantitative real-time PCR was performed using SYBR Advantage qPCR Pre-mix and the following primer sets: hsa-mir-223 forward primer; 5'-TGTCAGTTTGTCAAATACCCCA-3', mRQ 3' reverse primer, U6 forward primer and U6 reverse primer. The PCR reactions were run in 25 μL of the reaction mixture containing 2 × SYBR Advantage qPCR Premix (12.5 μL), 10 μM forward and reverse primers (final concentration, 0.2 μM), and 25 ng (2.0 μL) cDNA. To reduce variability between replicates, PCR premixes containing all reagents except for cDNA were prepared and aliquoted into 0.2 mL PCR tubes. The thermal cycling conditions were 10 s at 95°C, and 40 cycles of 5 s at 95°C and 30 s at 60°C. The expressions of IL-1β, IL-6, IKKα and MKP-5 relative to GAPDH, and miR-223 relative to U6 were determined in triplicate.
Western blottingProtein samples from HGF were separated in 10% SDS-PAGE and transferred onto Hybond-P membranes. The membranes were incubated for 2 h with anti-IKKα (Y463; Abcam, Cambridge, UK), anti-phospho-p38 MAPK (#9211; Cell Signaling, Technology Inc., Beverly, MA, USA), anti-Cdk2 (sc-163; Santa Cruz, Paso Robles, CA, USA) and anti-α tubulin (sc-5286; Santa Cruz) anti-bodies. Anti-rabbit or anti-mouse IgG conjugated with horseradish peroxidase was used as the second antibody. Immunoreactivities were determined using ELC plus Western Blotting Detection Reagent.
Cytokine measurementsIL-1β and IL-6 in the culture supernatants were measured using human enzyme-linked immunosorbent assay (ELISA) kits (Diaclone, Besançon Cedex, France) in accordance with the manufacturer’s protocols.
Statistical analysisTriplicate samples were analyzed for each experiment. The significance of differences between control and miR-223 and inflammatory cytokine treatments was analyzed using one-way ANOVA.
ResultsInflammatory cytokines induce miR-223 expression in HGFWe have previously reported that miR-223 expression is significantly upregulated in inflamed gingiva in compar-ison with non-inflamed gingiva (17). To determine whether inflammatory cytokines can induce miR-223 expression in HGF, we stimulated HGF with IL-1β (1 ng/
mL), IL-6 (1 ng/mL), and TNF-α (10 ng/mL) for 24 h and confirmed that miR-223 expression was induced by these cytokines (Fig. 1).
Regulation of L-1β and IL-6 mRNA levels by miR-223 To determine the effects of miR-223 on the expression of inflammatory cytokines (L-1β and IL-6), HGF were transfected with the miR-223 expression plasmid or control plasmid and stimulated with IL-1β (1 ng/mL) or TNF-α (10 ng/mL). IL-1β and TNF-α induced the expression of IL-1β and IL-6 mRNA in HGF, and this was further increased by overexpression of miR-223. Also, the levels of IL-1β and IL-6 mRNA were increased by miR-223 overexpression in the absence of stimulation with IL-1β or TNF-α (Fig. 2A, B).
Effects of miR-223 inhibitor on IL-1β and IL-6 mRNA levelsNext we stimulated HGF with IL-1β (1 ng/mL) or TNF-α (10 ng/mL) and transfected the cells with miR-223 inhibitor or a negative inhibitor control (miRCURY LNA Inhibitor Control; 10 nM). IL-1β increased the expression of both IL-1β and IL-6 mRNA in HGF, and the mRNA levels were significantly decreased by miR-223 inhibitor.

Fig. 2 Effects of miR-223 on levels of mRNA for inflammatory cytokines in HGF. The expression of IL-1β (A) and IL-6 (B) mRNAs in HGF was measured by real-time PCR. Cells transfected with the miR-223 expression plasmid or a control plasmid were treated with IL-1β (1 ng/mL) or TNF-α (10 ng/mL) for 24 h. Quantitative analyses of the data sets are shown with standard errors. Significant differences from control. *P < 0.05, **P < 0.01.
Fig. 3 Effects of miR-223 inhibitor on levels of IL-1β and IL-6 mRNA. The expression of mRNAs for IL-1β (A) and IL-6 (B) in HGF was measured by real-time PCR. Cells were transfected with the miR-223 inhibitor (10 nM) or control, and then treated with IL-1β (1 ng/mL) or TNF-α (10 ng/mL) for 24 h. Quantitative analyses of the data sets are shown with standard errors. Significant differences from control. *P < 0.05, **P < 0.01.
104
Also, TNF-α induced the expression of both IL-1β and IL-6 mRNA, and the mRNA levels were significantly decreased by miR-223 inhibitor (Fig. 3A, B).
Regulation of L-1β and IL-6 protein levels by miR-223To determine the effects of miR-223 on expression of IL-1β and IL-6 proteins, HGF were stimulated with IL-1β (1 ng/mL) or TNF-α (10 ng/mL), and transfected with the miR-223 expression plasmid or control plasmid. IL-1β induced the expressions of IL-1β and IL-6 proteins, and the expressions were further increased by overexpression of miR-223 (Fig. 4A, B). TNF-α induced the expressions of IL-1β and IL-6 proteins, but miR-223 overexpression
caused no further increase in these expressions (Fig. 4A, B).
Identification of miR-223 targets associated with cytokine productionTo clarify how miR-223 regulates the production of inflammatory cytokines, we used miRanda and TargetScanHuman (6.2) to predict the target genes of miR-223. We found two targets associated with inflam-mation: IKKα, a subunit of IKK that plays a crucial role in the non-canonical NF-κB signaling pathway, and MKP-5/DUSP10, a member of the MKPs, which control p38 MAPK phosphorylation. The IKKα 3'-UTR contains a putative miR-223 seed sequence and the

Fig. 4 Regulation of IL-1β and IL-6 protein levels by miR-223. Cells were transfected with the miR-223 expression plasmid or a control plasmid. IL-1β (A) and IL-6 (B) production was measured by ELISA after stimulation with IL-1β (1 ng/mL) or TNF-α (10 ng/mL) Significant differences from control. *P < 0.05, **P < 0.01.
Fig. 5 5 miR-223 targeting of IKKα and MKP-5. The expression of IKKα, MKP-5 and GAPDH mRNAs was measures by real-time PCR. HGF were transfected with the miR-223 expression plasmid or a control plasmid, and then treated with IL-1β (1 ng/mL) or TNF-α (10 ng/mL) for 24 h. Quantitative analyses of the data sets are shown with standard errors. Significant differences from control. *P < 0.05, **P < 0.01, NS: not significant.
105
MKP-5 3'-UTR has two putative miR-223 target sites. To confirm the contribution of miR-223 to IKKα and MKP-5 mRNA levels, HGF were stimulated with IL-1β (1 ng/mL) or TNF-α (10 ng/mL) and transfected with the miR-223 expression plasmid or control plasmid. IL-1β did not change the level of IKKα mRNA, whereas TNF-α suppressed it significantly relative to the control. When we transfected the HGF with the miR-223 expression plasmid, the level of IKKα mRNA did not change (Fig. 5A). IL-1β and TNF-α induced the expression of MKP-5 mRNA, and the mRNA levels were decreased by overex-pression of miR-223 (Fig. 5B). miRNAs down-regulate the expression of target genes by either of two post-transcriptional mechanisms: mRNA destabilization and
translational inhibition. It has been shown that miRNAs can repress gene expression without decreasing the levels of mRNA. The results of Western blotting suggested that miR-223 altered the level of IKKα by translational repression rather than by mRNA destabilization. The levels of IKKα protein in HGF were decreased by IL-1β and TNF-α, and further decreased by overexpression of miR-223 (Fig. 6A). In addition, we investigated whether miR-223 might regulate p38 MAPK phosphorylation via inhibition of MKP-5 production. It was found that miR-223 increased p38 MAPK phosphorylation relative to the control in response to stimulation with IL-1β or TNF-α (Fig. 6B).
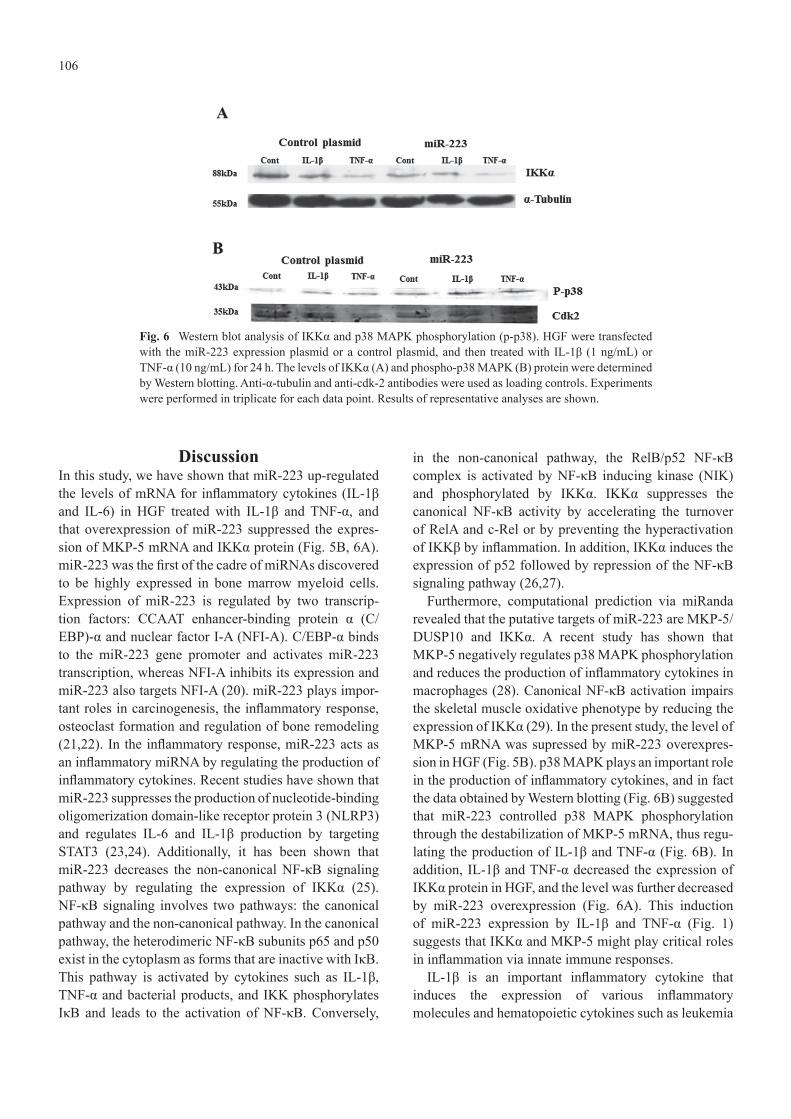
Fig. 6 Western blot analysis of IKKα and p38 MAPK phosphorylation (p-p38). HGF were transfected with the miR-223 expression plasmid or a control plasmid, and then treated with IL-1β (1 ng/mL) or TNF-α (10 ng/mL) for 24 h. The levels of IKKα (A) and phospho-p38 MAPK (B) protein were determined by Western blotting. Anti-α-tubulin and anti-cdk-2 antibodies were used as loading controls. Experiments were performed in triplicate for each data point. Results of representative analyses are shown.
106
DiscussionIn this study, we have shown that miR-223 up-regulated the levels of mRNA for inflammatory cytokines (IL-1β and IL-6) in HGF treated with IL-1β and TNF-α, and that overexpression of miR-223 suppressed the expres-sion of MKP-5 mRNA and IKKα protein (Fig. 5B, 6A). miR-223 was the first of the cadre of miRNAs discovered to be highly expressed in bone marrow myeloid cells. Expression of miR-223 is regulated by two transcrip-tion factors: CCAAT enhancer-binding protein α (C/EBP)-α and nuclear factor I-A (NFI-A). C/EBP-α binds to the miR-223 gene promoter and activates miR-223 transcription, whereas NFI-A inhibits its expression and miR-223 also targets NFI-A (20). miR-223 plays impor-tant roles in carcinogenesis, the inflammatory response, osteoclast formation and regulation of bone remodeling (21,22). In the inflammatory response, miR-223 acts as an inflammatory miRNA by regulating the production of inflammatory cytokines. Recent studies have shown that miR-223 suppresses the production of nucleotide-binding oligomerization domain-like receptor protein 3 (NLRP3) and regulates IL-6 and IL-1β production by targeting STAT3 (23,24). Additionally, it has been shown that miR-223 decreases the non-canonical NF-κB signaling pathway by regulating the expression of IKKα (25). NF-κB signaling involves two pathways: the canonical pathway and the non-canonical pathway. In the canonical pathway, the heterodimeric NF-κB subunits p65 and p50 exist in the cytoplasm as forms that are inactive with IκB. This pathway is activated by cytokines such as IL-1β, TNF-α and bacterial products, and IKK phosphorylates IκB and leads to the activation of NF-κB. Conversely,
in the non-canonical pathway, the RelB/p52 NF-κB complex is activated by NF-κB inducing kinase (NIK) and phosphorylated by IKKα. IKKα suppresses the canonical NF-κB activity by accelerating the turnover of RelA and c-Rel or by preventing the hyperactivation of IKKβ by inflammation. In addition, IKKα induces the expression of p52 followed by repression of the NF-κB signaling pathway (26,27).
Furthermore, computational prediction via miRanda revealed that the putative targets of miR-223 are MKP-5/DUSP10 and IKKα. A recent study has shown that MKP-5 negatively regulates p38 MAPK phosphorylation and reduces the production of inflammatory cytokines in macrophages (28). Canonical NF-κB activation impairs the skeletal muscle oxidative phenotype by reducing the expression of IKKα (29). In the present study, the level of MKP-5 mRNA was supressed by miR-223 overexpres-sion in HGF (Fig. 5B). p38 MAPK plays an important role in the production of inflammatory cytokines, and in fact the data obtained by Western blotting (Fig. 6B) suggested that miR-223 controlled p38 MAPK phosphorylation through the destabilization of MKP-5 mRNA, thus regu-lating the production of IL-1β and TNF-α (Fig. 6B). In addition, IL-1β and TNF-α decreased the expression of IKKα protein in HGF, and the level was further decreased by miR-223 overexpression (Fig. 6A). This induction of miR-223 expression by IL-1β and TNF-α (Fig. 1) suggests that IKKα and MKP-5 might play critical roles in inflammation via innate immune responses.
IL-1β is an important inflammatory cytokine that induces the expression of various inflammatory molecules and hematopoietic cytokines such as leukemia

107
inhibitory factor (LIF) and IL-6. IL-1β initiates a signaling cascade that culminates in activation of NF-κB and JNK (30). IL-6 is also known to play central roles in the pathogenesis of rheumatoid arthritis and peri-odontitis. Fibroblasts constitutively express a low level of IL-6. Furthermore, IL-1 and TNF-α synergistically enhance the production of IL-6 by fibroblasts (31-33). IL-1β and TNF-α induce NF-κB expression, and then NF-κB subsequently controls the expression of IL-1β, IL-6 and TNF-α (34-36). We found that IL-1β increased the expression of IL-1β and IL-6 protein, and that this was further increased upon overexpression of miR-223 (Fig. 4A, B). However, the expression of IL-1β and IL-6 protein induced by TNF-α was unchanged by miR-223 (Fig. 4A, B). Further studies will be necessary to clarify the reasons for these discrepancies.
In the present study, we have shown that, in HGF, miR-223 increased the levels of IL-1β and IL-6 mRNA induced by treatment with IL-1β and TNF-α, and that these mRNAs were down-regulated by miR-223 inhibitor. We also found that miR-223 increased the phosphorylation of p38 MAPK and suppressed the level of IKKα protein in HGF, whereas miR-223 did not decrease the level of IKKα mRNA. These results suggest that miR-223 may regulate the process of IKKα transla-tion rather than mRNA degradation. Taken together, our data indicate that miR-223 regulates the production of inflammatory cytokines through IKKα and MKP-5 in periodontal tissue.
AcknowledgmentsThis work was supported in part by a Grant-in-Aid for Scientific Research (C; No. 25463229), a Nihon University President’s Grant for Specified Multidisciplinary Research, and a grant from the Strategic Research Base Development Program for Private Universities from the Ministry of Education, Culture, Sports, Science, and Technology, Japan (MEXT), 2010-2014 (S1001024).
Conflict of interestThe authors have no conflicts of interest to declare.
References1. Pihlstrom BL, Michalowicz BS, Johnson NW (2005) Peri-
odontal diseases. Lancet 366, 1809-1820.2. Kato A, Imai K, Ochiai K, Ogata Y (2013) Higher prevalence
of Epstein-Barr virus DNA in deeper periodontal pockets of chronic periodontitis in Japanese patients. PLoS One 8, e71990.
3. Kato A, Imai K, Ochiai K, Ogata Y (2015) Prevalence and quantitative analysis of Epstein-Barr virus DNA and Porphyromonas gingivalis associated with Japanese chronic
periodontitis patients. Clin Oral Investig 19, 1605-1610.4. Seymour GJ, Gemmell E, Reinhardt RA, Eastcott J, Taubman
MA (1993) Immunopathogenesis of chronic inflammatory periodontal disease: cellular and molecular mechanisms. J Periodontal Res 28, 478-486.
5. Wang PL, Shirasu S, Shinohara M, Azuma Y, Daito M, Yasuda H et al. (1999) IL-10 inhibits Porphyromonas gingi-valis LPS-stimulated human gingival fibroblasts production of IL-6. Biochem Biophys Res Commun 263, 372-377.
6. Kida Y, Kobayashi M, Suzuki T, Takeshita A, Okamatsu Y, Hanazawa S et al. (2005) Interleukin-1 stimulates cytokines, prostaglandin E2 and matrix metalloproteinase-1 production via activation of MAPK/AP-1 and NF-κB in human gingival fibroblasts. Cytokine 29, 159-168.
7. Arthur JS, Ley SC (2013) Mitogen-activated protein kinases in innate immunity. Nat Rev Immunol 13, 679-692.
8. Li Q, Verma IM (2002) NF-κB regulation in the immune system. Nat Rev Immunol 2, 725-734.
9. Bonizzi G, Karin M (2004) The two NF-κB activation path-ways and their role in innate and adaptive immunity. Trends Immunol 25, 280-288.
10. Hayden MS, Ghosh S (2004) Signaling to NF-κB. Genes Dev 18, 2195-2224.
11. Li X, Massa PE, Hanidu A, Peet GW, Aro P, Savitt A et al. (2002) IKKα, IKKβ, and NEMO/IKKγ are each required for the NF-κ B-mediated inflammatory response program. J Biol Chem 277, 45129-45140.
12. Lang R, Hammer M, Mages J (2006) DUSP meet immu-nology: dual specificity MAPK phosphatases in control of the inflammatory response. J Immunol 177, 7497-7504.
13. Boutros T, Chevet E, Metrakos P (2008) Mitogen-activated protein (MAP) kinase/MAP kinase phosphatase regulation: roles in cell growth, death, and cancer. Pharmacol Rev 60, 261-310.
14. He L, Hannon GJ (2004) MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet 5, 522-531.
15. Klein ME1, Impey S, Goodman RH (2005) Role reversal: the regulation of neuronal gene expression by microRNAs. Curr Opin Neurobiol 15, 507-513.
16. Kozomara A, Griffiths-Jones S (2011) miRBase: integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res 39, D152-157.
17. Ogata Y, Matsui S, Kato A, Zhou L, Nakayama Y, Takai H (2014) MicroRNA expression in inflamed and noninflamed gingival tissues from Japanese patients. J Oral Sci 56, 253-260.
18. Ogata Y, Niisato N, Sakurai T, Furuyama S, Sugiya H (1995) Comparison of the characteristics of human gingival fibroblasts and periodontal ligament cells. J Periodontol 66, 1025-1031.
19. Nakayama Y, Takai H, Matsui S, Matsumura H, Zhou L, Kato A et al. (2014) Proinflammatory cytokines induce amelotin transcription in human gingival fibroblasts. J Oral Sci 56, 261-268.
20. Fazi F, Rosa A, Fatica A, Gelmetti V, De Marchis ML, Nervi

108
C et al. (2005) A minicircuitry comprised of microRNA-223 and transcription factors NFI-A and C/EBPα regulates human granulopoiesis. Cell 123, 819-831.
21. Haneklaus M, Gerlic M, O'Neill LA, Masters SL (2013) miR-223: infection, inflammation and cancer. J Intern Med 274, 215-226.
22. Taïbi F, Metzinger-Le Meuth V, Massy ZA, Metzinger L (2014) miR-223: an inflammatory oncomiR enters the cardio-vascular field. Biochim Biophys Acta 1842, 1001-1009.
23. Bauernfeind F, Rieger A, Schildberg FA, Knolle PA, Schmid-Burgk JL, Hornung V (2012) NLRP3 inflammasome activity is negatively controlled by miR-223. J Immunol 189, 4175-4181.
24. Chen Q, Wang H, Liu Y, Song Y, Lai L, Han Q et al. (2012) Inducible microRNA-223 down-regulation promotes TLR-triggered IL-6 and IL-1β production in macrophages by targeting STAT3. PLoS One 7, e42971.
25. Li T, Morgan MJ, Choksi S, Zhang Y, Kim YS, Liu ZG (2010) MicroRNAs modulate the noncanonical transcription factor NF-κB pathway by regulating expression of the kinase IKKα during macrophage differentiation. Nat Immunol 11, 799-805.
26. Lawrence T, Bebien M, Liu GY, Nizet V, Karin M (2005) IKKα limits macrophage NF-κB activation and contributes to the resolution of inflammation. Nature 434, 1138-1143.
27. Li Q, Lu Q, Bottero V, Estepa G, Morrison L, Mercurio F et al. (2005) Enhanced NF-κB activation and cellular function in macrophages lacking IκB kinase 1 (IKK1). Proc Natl Acad Sci U S A 102, 12425-12430.
28. Hömmö T, Pesu M, Moilanen E, Korhonen R (2015) Regu-lation of inflammatory cytokine production by MKP-5 in macrophages. Basic Clin Pharmacol Toxicol 117, 96-104.
29. Remels AH, Gosker HR, Langen RC, Polkey M, Sliwinski P, Galdiz J et al. (2014) Classical NF-κB activation impairs skeletal muscle oxidative phenotype by reducing IKK-α expression. Biochim Biophys Acta 1842, 175-185.
30. Kwan Tat S, Padrines M, Théoleyre S, Heymann D, Fortun Y (2004) IL-6, RANKL, TNF-α/IL-1: interrelations in bone resorption pathophysiology. Cytokine Growth Factor Rev 15, 49-60.
31. Takahashi K, Takigawa M, Takashiba S, Nagai A, Miyamoto M, Kurihara H et al. (1994) Role of cytokine in the induction of adhesion molecules on cultured human gingival fibroblasts. J Periodontol 65, 230-235.
32. Kent LW, Rahemtulla F, Michalek SM (1999) Interleukin (IL)-1 and Porphyromonas gingivalis lipopolysaccharide stimulation of IL-6 production by fibroblasts derived from healthy or periodontally diseased human gingival tissue. J Periodontol 70, 274-282.
33. Sawada S, Chosa N, Ishisaki A, Naruishi K (2013) Enhance-ment of gingival inflammation induced by synergism of IL-1β and IL-6. Biomed Res 34, 31-40.
34. Zhang YH, Lin JX, Vilcek J (1990) Interleukin-6 induction by tumor necrosis factor and interleukin-1 in human fibroblasts involves activation of a nuclear factor binding to a κB-like sequence. Mol Cell Biol 10, 3818-3823.
35. Grassl C, Luckow B, Schlöndorff D, Dendorfer U (1999) Transcriptional regulation of the interleukin-6 gene in mesan-gial cells. J Am Soc Nephrol 10, 1466-1477.
36. Xing L, Bushnell TP, Carlson L, Tai Z, Tondravi M, Siebenlist U et al. (2002) NF-κB p50 and p52 expression is not required for RANK-expressing osteoclast progenitor formation but is essential for RANK- and cytokine-mediated osteoclastogen-esis. J Bone Miner Res 17, 1200-1210.