p1,6gknac-branched n-glycans regulate ......figure 8 of chapter 4 and the northern blot shown in...
TRANSCRIPT

P1,6GkNAc-BRANCHED N-GLYCANS REGULATE
INFLAMMATION and CANCER
Maria Granovsky
A thesis submitted in confonnity with the requirements for the degree of Doctor of Philosophy,
Graduate Department of Medical and Molecular Genetics, University of Toronto
O Copyright by MariaGranovsky, 1999

NationaI Library 1+1 of Canada Bibliothèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services services bibliographiques 395 Wellington Street 395. rw Wellington Ottawa ON K i A ON4 Octawa ON K 1 A W Canada Canada
The author has granted a non- exclusive licence allowing the National Library of Canada to reproduce, loan, distribute or sel1 copies of this thesis in microform, paper or electronic formats.
L'auteur a accordé une licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distribuer ou vendre des copies de cette thèse sous la forme de microfiche/film, de reproduction sur papier ou sur format électronique.
The author retains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial extracts fiom it Ni la thèse ni des extraits substantiels may be printed or othewise de celle-ci ne doivent être imprimés reproduced without the author's ou autrement reproduits sans son permission. autorisation.

Maria Granovsky 1,oGlcNAc-branched N-glycans regulate inflammation and cancer progression
1999, Doctor of Philosophy Graduate Department of Medical and Molecular Genetics University of Toronto
Glycosylation of cell-surface and secreted glycoproteins is the most complex type of post-
translational modification, consuming a large portion of cellular resources, and may have as
broad an impact on metazoan biology as phosphorylation. $1,6GlcNAc-branched asparagine
0- l inked oiigosaccharkies, the biosynthesis of which requires the activity of B 1,6N- Acetylglucosarninyltransferase V (GlcNAc-TV), constitute a subset of structuraily
heterogeneous gl ycans found on ceIl surface and secreted gl ycoproteins. Increased B 1,dGlcNAc-
branching of N-glycans has been linked to cancer progression and metastatic potential. In
addition, the expression of GlcNAc-TV is induced by activation of SK: and Ras.
We first examined the expression pattern of Mgat.5, the gene encoding GlcNAc-TV, during murine
developmen t. GIcNAc-TV transcripts were absent in embryonic tissues at E7, but becarne
expressed throughout the embryo at E9.5, and then progressively restricted to regions of the
developing central nervous system and to the epithelia of skin, intestine, kidney, endocrine tissues
and respiratory tract. A comrnon feature of ceils in basal epithelia and in the cortical neural
epithelium is the capacity to migrate, a cellular function, which may require GlcNAc-TV - dependent glycoconjugates.
To examine the function of GlcNAc-TV dependent glycosylation in murine biology, we
generated mice deficient in this enzyme by targeted mutation of the Mgat5 locus. Homozygous
~ ~ a t . 5 ~ rnice were viable, fertile. These mice lacked pl,6GlcNAc-branched N-glycans,
indicating that only one gene encodes GlcNAc-TV activity. GlcNAc-TV deficient mice
displayed enhanced hypersensitivity to skin imtants. However, the onset of the inflamrnatory
response was slower, likely due to a reduced rate of leukoçyte migration in vivo, which was
associated with increased adhesion to substratum, measured in vitro. GlcNAc-TV deficient T
cells, but not B cells, were hypersensitive to mitogenic stimulation. Our results suggest that
B 1.6GlcNAc-branched N-glycans on the T cell surface have a direct role as a negative regulator
of T ce11 response to antigen, thus contributing to shutting down immune responses.
In order to study the role of GlcNAc-TV in metastatic breast cancer, we crossed GlcNAc-TV
deficient rnice with mice transgenic for the polyoma middle T antigen (PyMT) under the . . 11

regulation of a marnrnary speci fic promoter. PyMT is a potent oncogene that interaçts and
activates S hc, Src and PI3 kinase. Signaling through these pathways increases Mgat.5
expression, making this a relevant mode1 for studying the function of GlcNAc-TV in cancer.
GlcNAc-TV deficiency caused a delay in the development of palpable mammary tumors, the
area of which grew at a rate 6.7-fold slower than did tumors of wild type littermates, resulting in
4.5-fold reduction in tumor weight. GlcNAc-TV deficiency also resulted in >95% reduction in
the incidence of lung metastases. Our results suggest that GlcNAc-TV activity may enhance the
formation and turnover of focal adhesions, thereby increasing signaling via the integridPI3
kinase/PKB pathway, and influencing ceIl migration and metastatic potential.

1 would like to thank my supervisor, Jim Dennis, who has given me a great project to work on. While giving rnany helpful suggestions, Jim also allowed me enough freedom to explore different avenues and to make mistakes of my own devising.
1 am grateful to my codttee, Drs. Andras Nagy and Harry Schachter for their attention and comments. They were always available for discussion, and provided a different slant on some of the problems that were facing me during this project.
Past and present mexrtbers of the Dennis lab have left an enduring impression. In particular, 1 would like to thank Charles Warren, who has been my role mode1 throughout m y graduate school years. His - shall we Say - original views on appropriate experimental procedures and controls are unforgettable. Chris Binkert was a great sounding board for both procedural concerns and persona1 issues. Judy Pawling is worth her weight in gold for her incredible technical, organizational and baking skills. Al1 that dong with a cheerful and even personality too! 1 started in the lab when Carol Fode and Mike Demetriou were finishing their degrees. Carol set me up and got me started on m y project. Mike8s friendly picking enabled me to develop a tougher skin, which I'm told is a prerequisite for a career in science. Thanks must also go to Melanie, Tania, John, Aldis, Pam, Jemifer and Carol Swallow, who, collectively and individually, have contributed to my ongoing education as a person. 1 am indebted to several people in the Samuel Lunenfeld institue. Alec Cheng's contribution to this work is considerable, as he helped me to formulate the necessary questions about the phenotype of my mice. I've had many helpful discussions with Jeff Henderson who was very patient in illuminating the mysteries of the murine brain and in vivo perfusions to me. Deana Church was instrumental in hooking me up with my post-doctoral supervisor, and has thus provided the push 1 needed to complete m y degree. She has also threatened me with physical violence and thus kept my motivation at a high level during the writing of the thesis. Thanks must also go to fellow graduate students Vivette Brown and Jackie Koehler without whom my graduate school experience would have been considerably more drab. Finally, although 1 promised m y parents to leave them out of the acknowledgements, this list would be glaringly incomplete without them. So that's it. Just thank you.

Attribution of Data
1 am responsible for al1 the data that is presented in this thesis with the following exceptions.
The lectin histochemistry shown in Figure 2 of Chapter 2 was done by Dr. Robert Campbell.
Immunofluorescense images of fibroblasts and tumor cells show in Figure 6 of Chapter 3 and
Figure 8 of Chapter 4 and the Northern blot shown in Figure 1B of Chapter 4 were generated by
Judy Pawling. Western blots, shown in Figure 9 of Chapter 3, and in Figure 9 of Chapter 4 were
done by Dr. James Dennis. Jamie Fata stained mammary pads for histology, ceIl proliferation
and apoptosis (Figure SB, SA-D and table 1 of Chapter 4).

Table of Contents
Abstract
Acknowledgements
Attribution of Data
Table of Contents
List of Tables and Figures
List of Abbreviations
CWAPTER 1 : General Introduction
N ï R O D UCTION
GLYCOSYLATION AND CANCER Cancer initiation and progression Tumor Cell Proliferation Cell Migration and metastasis Sialylation and Metastasis Endogenous Lectins and T u m r Cell Adhesion Host anti-tumor immunity Carbohydrate processing inhibitors as anti-cancer agents
HERITABLE HUMAN GLYCOSYLATION DISORDERS AND ENGINEERED MUTATIONS IN MICE
Murations ogecring initiation and branching of N-glycans Mutations aflecting O-glycosylation Mutations aflecting Glycan elongation Mutations affecting glycan capping
CHAPTER 2: GlcNAc-TV and Core 2 GlcNAc-T Expression in the Developing Mouse Embryo
ABSTRACT
INTRODUCTION
RESULTS GlcNAc-TV expression in post-implantation mouse embryo Core 2 GlcNAc- T expression in the post- implantation mouse embryo
DISCUSSION
MATERIALS AND METHODS Animals

In situ RNA analysis Lectin histochemistry
CHAETER 3: Generation and Characterization of GlcNAc-TV Deficient Mice 72
INTRODUCTION 73
RESLJLTS Generation of ~ ~ a t 5 ' - mice GlcNAc-TV deficiency causes hyperinflammatory response Ce11 migration is slowed and adhesion to jibronectin is enchanced in ~ ~ a t f l - mice
GlcNAc-TV activity may regulate integrin signaling ~ ~ a t 5 - l - T-splenocytes are hypersensitive to T ce11 mitogens GlcNAc-TV deficiency is synergistic with CD28 CO- receptor stinrulation GlcNAc-TV defciency directly affects antigen-induced TCR intemalizaîion Mgat5 expression is regulated by T ce11 activation
DISCUSSION GlcNAc-TV deficient mice GlcNAc- TV deficiency alters cellular adhesion and migration TCR signaling and flf.6GIcNAc-branched N-glycans Immune regulation by MgatS
MATERIALS AND METHODS Mutation of the Mgat5 gene L-PHA lectin and Western blotting Skin Inflammation Induction of penioneal exudates and ce11 adhesion Stimulation of T cells in vitro FACS analysis and TCR internalization
CHAPTER 4: GlcNAc-TV Regulates Tumor Growth and Metastasis
INTRODUCTION 120
RESULTS Generation of P y ~ ~ ~ ~ a t 5 ' - mice Mammary tumors form with greoier lotency in ~ ~ a t 5 ' - mice Tumor growth is slowed in ~ ~ a t 5 ' - mice EXpressionji=om the Mgat5 locus is enhanced with PyMT transformation and tumor progression GlcNAc-TV deflciency supresses metastasis ~ ~ a t 5 - I - PyMT tumor cells display aberrant intracellular architecture Wild-iype ES cells f o m tumors in M g d mice
DISCUSSION 146 GlcNAc-TVdejkiency delays but dues not inhibit tumongenesis 146
vii

GlcNAc-TV defciency slows tumor growth Mgat5 expression is a murker of tumor progression GlcNAc-TV activiîy is required for efficient metasrmis and might be necessary for proper PZ3 kinase signaling Fast-growing Mgat5'- tumors may be the result of a somatic mutation Autonomous vs host-dependent factors in tumor progression
MATERIALS AND METHODS Generation of ~~at - ' -PyMr ' mice Northern analysis of PyMT expression Detection and measurement of tumors and metmatic foci Histological examination of turnors Immunofluorescent microscopy ES-cell teratocarcinomus
CHAPTER 5 : Conclusions and Future Directions GlcNAc-TV deftciency aflects maternai behavior GlcNAc-TV in immunity GlcNAc- TV deficiency causes deluyed tumor progression and inh ibits merastasis Mgat.5 expression may require signaling through both the Ras and the PZ3 kinase pathways The biological significance of GlcNAc-TV activity may be investigated in worm
List of References
viii

List of Tables and Figures
Chapter 3 Table 1: Sumrnary of the phenotypic aberrations observed in ~ ~ a t 5 - ' - mice. 82
Chapter 4 Table 1: Number of apoptotic cells per 10X magnification view
Figures
Chapter 1 Figure 1: Schematic of N-linked glycan biosynthesis showing the Golgi
compartments.
Figure 2: Schematic of O-linked glycan biosynthesis.
Chapter 2 Figure 1: In situ localization of GlcNAc-TV (TV) and core 2 GlcNAc-T (C2)
expression during murine em bryogenesis.
Figure 2: Lectin histochernistry of the E7 embryo.
Figure 3: GlcNAc-TV and core 2 GlcNAc-T expression in adult and embryonic mouse intestine.
Figure 4: GIcNAc-TV and core 2 GlcNAc-T expression in kidney at E17.5.
Figure 5: GlcNAc-TV expression in murine skin.
Figure 6: GlcNAc-TV and Core 2GlcNAc-T are not expressed in the same ceIl populations.
Figure 7: GlcNAc-TV and core 2 GlcNAc-T expression in female reproductive organs.
Figure 8: Summary of the expression patterns.
Chapter 3
Figure 1: Generation and charactenzation of GlcNAc-TV deficient mice.
Figure 2: LacZ expression from the mutant MgarS- allele is identical to the MgatS expression pattern.
Figure 3: Aberrant histology in ~ ~ a r 5 - I - mouse tissues.
Figure 4: Inflammation in ~ ~ a t . 5 ' - and littermate controls.

Figure 5: Ce11 migration is slowed and adhesion to fibronectin is enhanced in ~ ~ a r 5 ' - rnice.
Figure 6: Focal adhesions in ernbryonic fibroblasts plated on fibronectin.
Figure 7: ~ ~ a z 5 ' - T splenocytes are hypersensitive to T ce11 mitogens.
Figure 8: GlcNAc-TV deficiency Enhances Coopertivity of TCR-rnediated celiular activation.
Figure 9: GlcNAc-TV deficiency directly affects antigen-induced TCR intemalization.
Figure 10: Mgut5 expression is regulated by T cell activation.
Figure 11: Signaling initiated by TCR engagement.
Chapter 4
Figure 1: Generation of M~U~F/-P~W mice and P y m transgene expression.
Figure 2: PyMT tumor development is delayed in ~ ~ a t 5 ~ rnice.
Figure 3: GlcNAc-TV deficiency causes slower tumor growth.
Figure 4: Histopathology of mammary fat pads and tumors in 26 week old P ~ M T M ~ U ~ Y ~ and P ~ M T M ~ ~ ~ S ~ mice.
Figure 5: Proliferative index in mammary fat pads and tumors in P~MTM~U~S/ - and P ~ M T M ~ ~ ~ ~ + ' - mice.
Figure 6: Lac Z expression in ~ ~ a t 5 + ~ and ~ ~ a t . 5 ~ tumors.
Figure 7: GlcNAc-TV activity is required for metastasis.
Figure 8: Effects of GlcNAc-TV deficiency on intrrtcellular cytoskeletal architecture.
Figure 9: GlcNAc-TV deficiency in host animais affects growth characteristics of wild type ES cell-derived teratocarcinornas.
Figure 10: The effect of GlcNAc-TV deficiency on MAP kinase and PKB phosphory Iation.
Chapter 5
Figure 1: Schematic of hypohesis for GlcNAc-TV function in ce11 adhesion.

List of Abbreviations
aa a-MEM APC CD CHO DTH E ECM ELISA ES FACS FITC Ga1 GalNAc GlcNAc GlcNAc-TV i.p. i.v. IT AM LAT LFA- 1 LPS MHC MMP PH S.C.
SA Sm TCR T W a VEGF X-gal ZAP-70
m i n o acid alpha-minimal essential medium antigen presenting ce11 cluster designation Chinese hamster ovary delayed-t ype h ypersensi tivity embryonic day extracellular matrix enzyme-linked immunosorbent assay embryonic stem fluorescence aiâed ce11 sorting flourescien isothioc yanate galactose N-acetylgalactosamine N-acetylglucosamine p 1,6N-Acetyi glucosamin y ltrans ferase V intraperi toneal intravenous immunoreceptor tyrosine -based activation sequence motif Leukoçyte lymphocyte function-associated antigen lipopol ysaccharide major histocornpatibility complex matrix metaloproteinase plekstrin homology subcutaneous sialic acid Src homology 2 T ce11 receptor-CD3 complex tumor necrosis factor alpha vascular endothelid growth factor 4-chloro-5-bromo-3-indolyl-~-galactoside zeta-chain associated protein

CHAPTER 1 : General Introduction

INTRODUCTION
Glycosylation of cell-surface and secreted glycoproteins is the most complex
post-translational modification. As such, it requires a large portion of cellular resources.
For example, an estimated 0.5 to 1% of the vertebrate translated genome is devoted to
glycosylation-related genes (Varki and Marth, 1995), a commitment sirnilar in magnitude
to that of protein phosphorylation. Anal ysis of somatic gl ycos ylation mutants and
transgenic mice suggests that the functionai impact of glycosyiation on metazoan biology
might be as broad as that of phosphorylation as well. Glycosylation modulates and
regulates a variety of protein-protein interactions relevant to cellular proliferation,
adhesion and migration. The structural complexity of glycans hm impeded functional
analysis of specific structurd subgroups, but targeted mutations in mice affecting specific
steps in the biosynthetic pathways of glycosylation offer a powerful tool for studying
such structure-function relationships. This thesis describes the results of ablating a
specific subset of N-glycan structures by targeting Mgat5 the gene encoding P1,6N-
Acetylglucosaminyltransferase V (GlcNAc-TV), which catalyzes the addition of the
B 1,oGlcNAc-branch on N-glycans.
Oligosaccharides on glycoproteins are classified by the nature of their Iinkage to
the protein as either N-linked or O-Iinked (reviewed in (Komfeld and Komfeld, 1985;
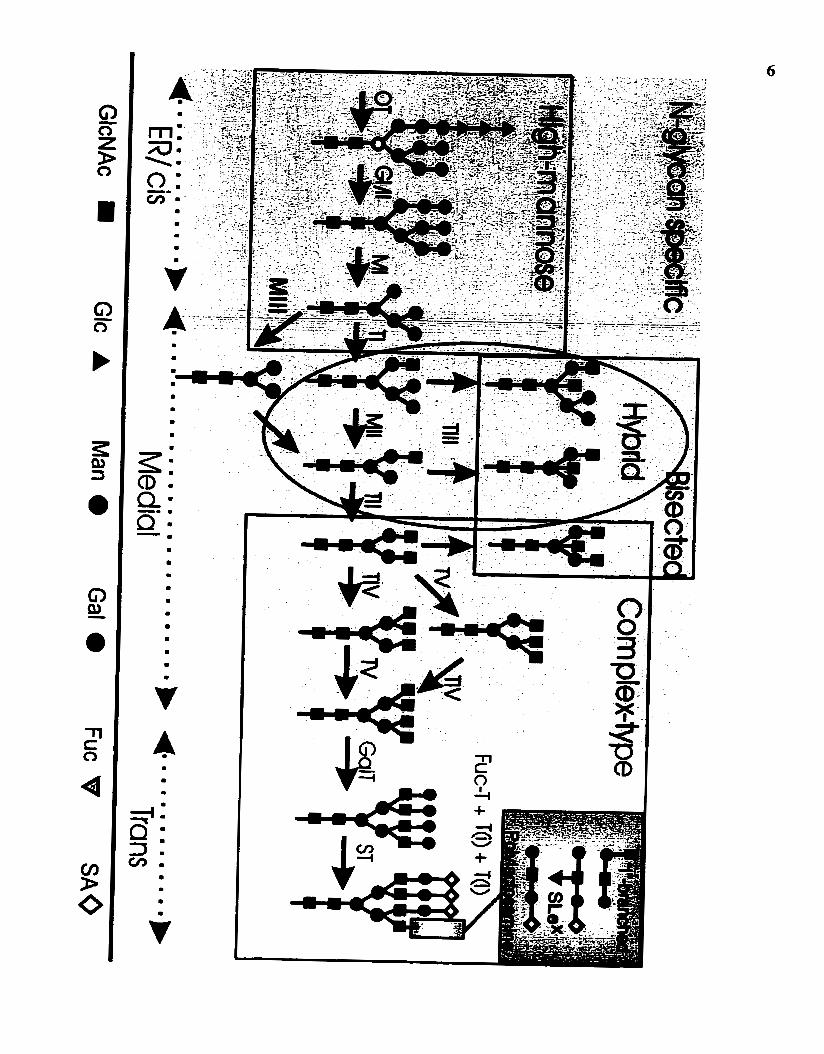
Schachter, 1986)). N-glycans are the focus of this work (for biosynthesis, see Figure 1).
Dolichol-mediated N-gl ycosylation is absent in bacteria, but present in al1 eukaryotic
species, including yeast. Glycosylation aids the proper folding of glycoproteins and the
mediation of chaperone-protein recognition in the endoplasrnic reticulum (ER) of
eukaryotes (Velders and Kast, 1999; Ware et al., 1995). A glucosyltransferase recognizes

3 misfolded glycoproteins and reglucosylates the polypeptide so it is recirculated for
degradation (Velders and Kast, 1999).
The activity of GlcNAc-TI, which is essential for the synthesis of hybrid- and
complex-type N-glycans (Schachter, 1986), is absent in yeast but present in al1
multicellular organisms studied to date. Thus, GlcNAc-TI demarcates a fundamental
evolutionary boundary between uni- and multicellular organisms. This divergence
suggests a role for complex and hybrid N-glycans in intercellular communication and in
the interactions of the ce11 with its environment. Consistently, comptex- and hybrid-type
glycosylation occurs in the Golgi network, an organelle devoted to secretory and cell-
surface proteins.
N-glycosylation can be viewed as consisting of four phases (Figure 1). Transfer
of a pre-assembled precursor oligosaccharide from a dolichol pyrophosphate to the
nascent polypeptide in the rough ER is followed by tnmming by glycosidases in the
rough ER and cis-Golgi. The trimrned intermediate passes into the medial-Golgi, where
branching by GlcNAc-Ts is added to the growing structure that is then elongated and
capped in the trans-Golgi. The structural varïability of glycans is dictated by tissue-
specific regulation of glycosyltransferase genes, acceptor and sugar nucleotide
availability in the Golgi, compartmentalization, and by competition between enzymes for
acceptor intermediates dunng glycan elongation. Several genes encoding
glycosyltransferases appear to be regulated by multiple promoters, and
glycosyltransferase activity levels measured in vitro do not necessarily reflect message
levels, suggesting pst-transcnptional andor pst-translational regulatory mechanisms
(Lo and Lau, 1996; Perng et al., 1994; Saito et ai., 1995; Yang et al., 1994). Given the
possible combinations of available monosaccharides, linkages, branching and variable

4
lengths of glycan chains, the potential for stnictural diversity of glycans in metazoan ceIls
is very large.
Glycosyltransferases catalyze the transfer of a monosaccharide from specific
sugar nucleotide donors ont0 a particular hydroxyl position of a monosaccharide in a
growing glycan chain in one of two possible anomeric linkages (either a or B). The
majori ty of gl ycosyltransferases characterized to date show exquisi te specificity for the
donor and acceptor they recognize and the Iinkages they cataiyze. The protein rnicro-
environment in which the immature glycan chah is synthesized dso affects
gI ycos yltransferase catalytic efficiency (Do et al., 1994; Granovsky et ai., 1994) and
leads to structural heterogeneity of gl ycans between gl ycoproteins, and even between
different glycosylation sites on individual glycoproteins produced by the sarne ceIl
(Rademacher et al., 1988; Sheares, 1988). A range of biosynthetically related glycan
structures may be present at any particular glycosylation site of a mature glycoprotein,
creating a population of "glycoforms". The prevalence of a particular glycoform can
affect the intrinsic biological activity of an individual gl ycoprotein. For example, the
peptide hormones lutropin and erythropoietin are produced with stnicturally diverse
EI ycans and the discrete glycoforms of these hormones have significantly different serum - half-life and potency in vivo (Fiete et al., 1991; Takeuchi and Kobata, 1991). Cytokine
and adhesion receptors are similariy subject to glycoform heterogeneity with the potential
to modify the kinetics of physical association of these receptors, and thereby intracellular
signaling.
The Golgi biosynthetic pathways appear to have evolved to generate structural
diversity on glycoproteins, but the selective pressure favoring such resourceexpensive
complexity of glycosylation remains unclear. One possible explanation is parasite

5 Figure 1: Schematic of N-liaked glycan biosynthesis showing the Golgi
compartments.
A high-mannose pre-assembled precursor oligosaccharide is transferred from a
dolichol pyrophosphate to the nascent polypeptide in the rough ER. It is tnrnmed by
glucosidases and by a-mannosidase 1 in the rough ER and cis-Golgi. The action of
GlcNAc-TI is required for the production of hybrid- and cornplex-type N-glycans. The
action of a-mannosidase II or a-mannosidase III, followed by GlcNAc-TI1 is required to
synthesize the precursor to ail cornplex-type N-glycans. Note that GlcNAc-TIII
substitutes the core $-mannose residue and redirects the pathway into "bisected N-
gl ycans", blocking subsequent action of GlcNAc-branching transferases (Schachter,
1986). The terminal sequences are added to both N- and O-linked glycans by B1,4Gal-T,
B 1,3GlcNAc-T(i), f3 1,6GlcNAc -Tm, a2,3ST, a2.6ST and al ,3Fuc-T. GlcNAc-TV is
one of several medial-Golgi enzymes that initiate a specific branch (the P1.6GIcNAc
branch) that is variably elongated in the trans-Golgi generating structural diversity in the
mature N-glycans. The GlcNAc-TV product is the preferred intermediate for extension
with polyIactosarnine chains (Le. Gal$l,4GlcNAc$1,3 repeating units of 2 to >10 in
length). Polylactosamine synthesis is also regulated by pl,3GlcNAc-T(i) activity,
glycoprotein transit time in the trans-Golgi, and cornpetition by chain-terminating
enzymes including a l ,2Fuc-T and a2,6SA-T. The pol ylactosamine and Lewis antigens
that preferentially elongate the $1,6GlcNAc-branch are expanded in the grey box.
Abbreviations used are oligosaccharyltransferase, OT; the a-glucosidases, GI, GII; the p- N-acetylglucosaminyltransferases, TI, TII, Tm, TIV, TV, T(i) and T(ï); the
al ,2mannosidases, MI, al,3/6mannosidases MII, Mm; f31,4-galactosyltransferases (Gal-
T), a- fucos y l tram ferases @uc-T), a-sial y1 transferases (ST).

7
Figure 2: Schematic of O-iinked glycan biosynthesis.
O-glycan specific enzymes are shown in the shaded area. Unlike N-glycosylation, O-
glycosylation is initiated directly on the nascent poIypeptide, without an intermediate.
The terminal sequences shown in the non-shaded area are added to both N- and O-linked
glycans by a common set of enzyme as noted in Figure 1. The numbers in front of the
sialyltransferases (STs) indicate the linkage of the transferred SA. Note that core 2
GlcNAc-T initiates an antenna required for addition of polylactosamine and polyvalent
Lewis antigens. GaiNAc in mucins c m also be substituted with $l,3GlcNAc and in
several other linkages creating "other cores" disaccharides, which mature differently.
Abbreviations are polypeptide O-a-N-acetylgalactosaminyltransferase, (ppGT);
B 1,3galactosyitransferase, (3Gal-T); the core 2, p-N-acetylglycosaminyltransferases, (C2-
Tl-

Other terminal sequences - polylactosamine
w "1"- branc hed
GlcNAc . GalNAc . Gal 0 FUC I SAO

9
evasion. Glycan binding to lectin is a common means of adhesion between multicellular
organisms and parasitic organisms. For example, adhesion of Helicobacter pylori to the
mucous and gastric epithelial cells is mediated by fucosylated blood-group antigens of
the O blood-group, and has been associated with a greater incidence of ulcerative disease
in O blood group individuals (Boren et al., 1994). Infection by Entamoeba histolytica,
which can result in colitis and dysentery, is mediated by recognition of Ga1 and GaiNAc
residues by a parasite-expressed lectin (Dodson et al., 1999)- and Tritrichmonas foetus,
an infectious bovine parasite associated with inflammation, abortion and infertility,
adheres to its host by a sialic-acid specific lectin (Baba1 and Russell, 1999). However,
different terminal sequences on glycans c m also mask lectin-binding sites for parasite
infection. Thus, continuous generation of molecular diversity in metazoans may enhance
population fitness in the ongoing battle to evade parasitic organisms.
Newly arising glycan structures are CO-opted, on occasion, into functions
unrelated to pathogen evasion. The conserved structures become the açceptors for newly
evolved gl ycos yltransferases, and this generates additional diversi ty. In this scenarîo, the
oIder the capacity for a particular carbohydrate structure, the more likely it is that the
structure has acquired biological functions intrinsic to the organism, and unrelated to
pathogen evasion. Consistently, mutations in ancient parts of the glycosylation pathway
will affect many downstream oligosaccharide structures, some of which now
performdevelopmental functions. Indeed, the phenotypes of glycosyltsansferase-deficient
mice are progressively rnilder for enzymes operating later in the biosynthetic pathway.
GlcNAc-TI deficient ( ~ g a t l - " ) mice lack al1 hybrid- and complex-type N-glycans, and
they die between embryonic day (E) 8.5 and E9.5 due to multiple organ failure.
Mutations in downstream genes, affecting only subsets of complex-type N-glycans are
iess severe. ~ g a t 2 ' ~ mice survive until birth, and ~ ~ o t 3 - ' * . ~ ~ a t 5 ' - .

10
G ~ ~ T I - ~ . and FMCTVII' mice are viable (Asano e t al., 1997; Chui et al., 1997; Granovsky
et al., 1997; Ioffe and Stanley, 1994; Maly et al., 1996; Metzler et al., 1994; Priatel e t al.,
1997).
The effects of glycosylation on cellular proliferation and migration have k e n
studied rnostly in the context of tumor progression and metastasis. Previously, over 40
somatic mutants of N-glycosylation enzymes have been isolated and characterized
(Stanley, 1983). Glycosylation mutant tumor cells grow autonomously in culture, but in
vivo, solid tumor p w t h andor expeimentd metastasis are compromised by certain
mutations. Cancer growth and metastasis involve many cell-ce11 interactions including
ce11 migration, invasion, differentiation, angiogenesis and immune system evasion. As
such, one or more of these processes would appear to require precise glycosylation of
macrornolecules for efficient tumor growth and metastasis.
GLYCOSYLATION AND CANCER
Malignant transformation is often associated with altered glycosylation of
glycoproteins and glycolipids, and expression of certain glycan structures in tumors
correlates with clinical prognosis (reviewed in (Hakomori, 1996; Kobata, 1998)).
GIycans are structural components of many ce11 surface glycoprotein receptors, such as
ce11 adhesion receptors and cytokine receptors, the signaling of which controls
fundamental cellular processes that are of prime interest when studying cancer
progression and metastasis. The role of carbohydntes in cancer initiation, progression
and metastasis is an area of intense study using somatic tumor ce11 mutants, tumor cells
transfected with glycosyltransferase genes and transgenic mice.

11
Cancer initiation and progression: The study of cancer genetics is key to Our
understanding of the disease, and provides a context in which to consider glycosylation
effects. Multiple genetic changes, combined with selection pressure in the host
environment, provide conditions for tumor formation and progression. Mutations that
impart growth advantages are maintained and selected for in the cellular population,
allowing mutant cells to preferentiaily expand during tumor progression and acquisition
of metastatic potential (Cairns, 1981). For example, the pre-neoplastic cells of aberrant
crypt foci in colon display increased proliferation; a cancer-predisposing condition
because these rapidly cycling cells, with increased DNA replication, are more likely to
sustain subsequent rare mutations. These rare mutations allow aberrant crypt foci cells to
form pre-malignant polyps, which can progress to adenornas and finally to metastatic
colon carcinomas (Lengauer et al., 1997).
Cancer mutations result in either loss-of-function in a gene product,
designatecf "tumor suppressor proteins" (e.g. p53, APC, WTl), or missense mutations
which activate a "proto-oncogene" (e.g. h-ras). Tumor suppressor genes have also k e n
described as "caretaker" and "gatekeeper" genes, controlling genomic integrity and
aspects of cellular proliferation, respectively (reviewed in (ICinzler and Vogelstein,
1997)). The retinoblastoma gene, Rb-1, is a gatekeeper gene, responsible for maintaining
cells in Gi phase (Weinberg, 1999). Loss-of-function mutations in Rb-1 promote entry
into the S phase of the ce11 cycle, and subsequent tumorigenesis in retinai epitheliai cells.
The mismatch DNA repair enzymes MSH2 and MLHl are caretaker genes and their
inactivation in colon cancers leads to mutations in other genes that can enhance growth,
such as the TGF-f! receptor type II gene (Grady et al., 1999). P53 and other genes that
ensure faithful segregation of chromosomes during mitosis also serve a caretaker function
(Cross et al., 1995). Heritable mutations in turnor suppressor genes such as P53, Rb-1,

12
and APC greatly increase the risk of cancer, as only one wild-type allele is present and
therefore only one somatic inactivating mutation is required for complete Ioss of
function. Some of the heritable mutations common in human cancers were engineered in
mice to facilitate the study of the pathways leading to oncogenesis (reviewed in (Jacks,
1996; Webster and Muller, 1994)). Indeed, many of these transgenic mice show
increased incidence of pre-rnalignant lesions and tumors, and thereby are useful in
establishing cause and effect relationships in the cascade of events leading to metastatic
cancer.
Currently, genes encoding glycosylation-related enzymes are not included in
the catalogue of somatic and heritable mutations responsible for cancer initiation or
progression. However, changes in glycosylation are commonly observed in human
carcinomas, and in ce11 Iines transformed with activated Ras and v-Src (Easton et al.,
1991; Lu and Chaney, 1993; Pierce and Arango, 1986; Yarnashita et al., 1985; Yarnashita
et al., 1985; Yarnashita et al., 1985). In earlier studies of tumor ce11 glycosylation
mutants, a2,3sialylation, increased polyIactosamine content and B1,6GlcNAc-branching
of N-glycans were suggested to enhance tumor growth and metastasis in mice (Dennis,
1986; Dennis et al., 1987; Finne et al., 1982; Lu et al., 1994). More recently, over-
expression of glycosyltransferase genes in tumor ceIl lines, and studies in
glycosyltransferase deficient mice confirm that cancer-associated changes in
glycosylation play a causal role in cancer progression and metastasis.
Turnor Ce11 Proli$erafion: The ras proto-oncogene sustains activating mutations in
approximately 20% of al1 human tumors (Velders and Kast, 1999). In addition, Ras
signaling is induced by other common mutations such as amplification of NedErbB-2 in
breast cancer (Denhardt, 1996). Signaling by the Ras GTPase activates Raf-1 kinase,
which leads to the activation of AP1 (i.e. c-Fos/c-Jun dimers) and Ets transcription

13
factors (Wasylyk et al., 1998). These transcription factors regulate expression of multiple
genes involved in ce11 cycle progression and ce11 motihty, as well as metalloproteases,
growth factors and gl ycos y ltrans ferases.
The relationship between Ras and c-Fos was dernonstrated genetically.
Application of skin carcinogens results in invasive skin tumors in mice transgenic for
activated Ras. However, c-Fos deficient mice carrying the activated v-H-ras transgene
develop benign tumors only, in which matrix metalloproteases (MMPs) and vascular
endothelial growth factor (VEGF) transcripts are suppressed (Saez et al., 1995). VEGF
induces host endothelial cells to develop microvasculature, thereby providing the
necessary oxygen and nutnents to the expanding tumor. MMPs secreted by tumor cells
digest extracellular matrix and facilitate tumor ce11 invasion through extracellular matrix
t hat separates tissue compartments. Thus, according to this experiment, Ras acts upstream
of c-fos, which is an affector of VEGF and MMP transcription. This study is an exarnple
of how genetic analysis facilitates the ordering of specific molecules in a signaling
cascade.
The Ras signaling pathway is activated by growth factors including members of
the EGF, PDGF and NGF farnilies (Chao, 1992). In Mgar3 transfected U373 glioma
cells, EGF receptor levels and signaling were observed to decrease, and both the EGF
receptor and the NGF receptor, Trk, were substrates of GlcNAc-Tïïï-dependent
glycosylation (Rebbaa et al., 1997). Dimerization and phosphorylation of Trk were also
reduced in Mgut3 transfected PC 12 cells (Ihara et al., 1997). Converse1 y, overexpression
of the O-glycan $1,6GIcNAc-branching enzyme, core 2 GlcNAc-T in PC12 cells
enhanced Trk receptor O-glycosylation, signaling via MAPK, and PC 12 differentiation
(Persona1 communication, C.E. Warren). Trk receptor levels on the ce11 surface, as well
as receptor activity are enhanced by B1,6GIcNAc-branching of either N- or 0- glycans.

14
Therefore, it is likely that a comrnon feature, such as polylactosamine extensions rnay be
involved in stabilizing the receptor in some manner.
NGFmrk and EGFEGFR both activate RasIMAPK in PC12 cells but with
different cellular outcornes. NGF/Trk activation results in differentiation, whereas
EGFIEGFR activation results in proliferation (Tan and Kim, 1999). The apparent
dichotomy cm be resolved by considering the kinetics of activation through these
receptors, which are different. The studies with Mgar3 and core 2 GlcNAc-T transfected
cells suggest that glycosylation may modify intracellutar signding and that GlcNAc-Tm-
mediated glycosylation and B1,dGlcNAc-branching of either N- or O-glycans exert
opposite effects on Trk signaling. GlcNAc-Tm and GlcNAc-TV, which catalyzes the
addition of the f31,6GlcNAc-branch on N-glycans, compete for glycan acceptor
intermediates and the action of GlcNAc-TIII renders glycan intermediates poor substrate
for GlcNAc-TV in in vitro biochemical studies (Schachter, 1986). Indeed, Bl6 melanoma
cells transfected with a GlcNAc-Tm expression vector showed an increase in hybrid-type
gl ycans and a reduction in GlcNAc-TV products (Sheng et al., 1997). Thus, the relative
levels of GlcNAc-Tm and GlcNAc-TV in a particular ce11 may provide kinetic fine-
tuning to the all-or-none phenomenon of Trk dimerization and signaling.
In many cancer cells, where Ras is constitutively activated, the observed effects of
glycosylation are probably due to their modulation of signaling through receptors
utilizing other pathways. Activating mutations in ras, of the type found in human
tumors, bypass the need for growth factors and ce11 surface glycoprotein receptors.
Therefore, the glycosylation status of glycoprotein receptors and cytokines upstrearn of
Ras may be less significant in cells with a mutated ras gene, as the cells acquire a large
measure of growth-factor independence. A sirni lar phenomenon was recentl y
demonstrated for the cytokine pathway initiated by Wnt 1 Frz signaling in Drosophila.

15
Wnt receptor (Frz) signaiing requires UDP-glucose dehydrogenase, which produces
glycosarninoglycans, required as CO-recepton. Inactivating mutations in p-catenin and
APC, which are negative regulators of the growth stimulation by the Wnt pathway,
circumvent the requirement for Wnt / Frz, and for UDP-glucose dehydrogenase
(Cumberledge and Reichsman, 1997).
Ce11 Migrdon and metastusis: Metastatic carcinoma cells are not restricted by the tight
junctions c haracteristic of nomal epithelial cells. With the transition h m normal to
malignant status, cell-cell adhesion molecules are switched off, and substratum adhesion is
modified to optimize focal adhesion turnover and cell motility. In normal epithelial cells,
tight junctions and desmosomes associate with cytoskeleton intermediate filaments to
maintain ce11 shape and polarity. Criticai to the maintenance of these contacts is ca2*-
dependent homophilic binding of the cell-adhesion molecule E-cadherin in the zones of
adhesion (Christofori and Semb, 1999). Gemiline mutations in the E-cadherin gene have
k e n found in cases of familial gastric cancer, and either Ecadherin expression or its
function is suppressed in sporadic carcinomas of rnost tissues (Birchmeier, 1995; Fnxen et
al., 1991). Forced over-expression of E-cadherin in tumor cells suppresses tumor growth in
rnice (Birchmeier, 1995). E-cadherin and CD44 were subject to GlcNAc-TIII-dependent N-
glycosylation in transfected B l 6 cells (Sheng et al., 1997; Yoshimura et al., 1996), and
Ievels of E-cadherin increased in GlcNAc-TIII transfectants. Therefore, inactivating
mutations, reduced gene expression or aberrant glycosylation of the gene product can cause
suppression of Ecadherin activity, and the subsequent enhancement of tumor ce11 growth
autonomy. The overexpression of the hyaluronic acid-binding ceil adhesion receptor CD44
suppresses tumor progression (Schmits et al., 1997). CD44 activity is glycosylation-
dependent, as enzymatic removal of sialic acid and galactose from complex-type N-glycans
of CD44 enhances binding to hyaluronate (Skelton et al., 1998). GlcNAc-Tm transfection

16
of B 16 melanoma cells enhanced CD44-mediated ce11 adhesion, reduced invasion through
extracellutar matrix in vitro, and reduced lung colonization in vivo when transfected cells
were injected intravenously into mice (Sheng et al., 1997; Yoshimura et al., 1995). These
studies demonstrate that GlcNAc-TIII-mediated glycosylation may enhance tumor cell
adhesion, thereby reducing tumor ce11 growth autonomy. This outcome may be achieved
partly by competing for substrate with GlcNAc-TV and reducing 81,6-branching of N-
glycans, which have been implicated in reducing adhesion and promoting ce11 migration.
Metastatic tumor cells rnust migrate over extraceilular matrix (ECM) to make
their escape from the primary tumor, then gain access to the blood Stream, attach to the
distal vascular bed, migrate over ECM and grow in the secondary organ (Nicolson,
1988). Tumors of various origins differ greatly in phenotypes that affect metastasis.
Various steps in the metastatic process, such as ce11 migration on ECM, required for both
escape from the primary tumor and seeding at the secondary site, can be rate lirniting for
metastasis, depending on the tumor phenotype. Ce11 migration on ECM is mediated
primarily by signaling through the integrin receptors. These receptors mediate
attachment to substratum, aggregating into focal adhesions when bound to the ECM
glycoproteins fibronectin, laminin and collagen (Ruoslahti, 1996). In growth-arrested
non-transformed cells, ligand-engaged integrins forrn stable adhesion plaques with links
to actin stress fibers inside the cell. In either growth factor stimulated cells or
transformed cells, focal adhesions induce continuous recruitment of signaling complexes
on the cytosolic side of the plasma membrane. This results in turnover of the actin
microfilarnents, and activation of the RadMAPK and PI3K/PKB signaling pathways
(Jaln et ai., 3653; Schlaepfer et al., 1995). Cell migration rates depend upon optimal
turnover of focal adhesion complexes. Integrin receptor levels and activity are balanced
against ECM adhesion domains to regulate focal adhesion turnover and thereby migration

17
rates. Substratum density changes can Vary reciprocally to integrin levels or their affinities
(Palecek et al., 1997). As such, rates of focai-adhesion turnover and ce11 motility exhibit
bell-shaped responses to changing ligand or reçeptor levels. Focal adhesion turnover also
stimulates intracellular signaling and ce11 proliferation via PI3K and c-Src kinases, which
creates a positive feedback loop.
Studies in MvlLu cells, S 115 marnmary carcinoma cells and T cells from patients
with Sezary syndrome indicate that B1,6GlcNAc-branched N-glycans are present on as.
a, and f31 integrin subunits (Braut-Boucher et ai.. 1998; Demetriou et al.. 1995; Leppa et
al., 1995). Cellular adhesion to ECM inverse1 y comlates with f3 1 ,GGlcNAc-branching,
while the motility of the cells correlates with f3 1,dGlcNAc-branching directly (Demetriou
et al., 1995). Therefore, it is possible that integrin-mediated signaling is subject to
regulation by $1,6GlcNAc-branched N-glycans.
Sialylation and Metasfasis: Terminal N- and 0- glycan sequences added in the trans-
Golgi cornpartment contribute to rnalignancy, as suggested by studies on tumor cell
glycosylation mutations. MDAY-D2 mutants, overexpressing a2,6SA-T due to a retroviral
insertion into the gene promoter, showed 3-10 fold fewer metastases and 60% slower turnor
p w t h (J. Liu, personal cornmunication). The mutant cells had predominantly a2,GSA
rather than the wild-type cQ,3SA on the ce11 surface (Takano et al., 1994). Sirnilarly,
transfection of a glioma ce11 line, U373 MG cells with a2,6SA-T reduced invasion
(Yamamoto et al ., 1997). The h yposial ylated CMP-NeuNAc-h ydrox ylase-expressing
MDAY-D2 mutants gave rise to slower growing solid tumors than did the parent cells. In
addition, loss of sialylation in B 16 melanoma mutants due to over-expression of al,3Fuc-T,
a1,ZFuc-T or al3Gd-T, resulted in loss of metastatic potential (Dennis, 1986; Finne et
al., 1982; Gorelik et ai., 1997; Yamamoto et al., 1997). These results imply that capping

18
N-acetyllactosamine with N-acetylneurarninic acid, specifically in the a2,3 linkage, may
be conducive to efficient metastasis.
Endogenous Lectins and Tumor CeU Adhesion: The GlcNAc-TV enzyme product in the
N-glycan pathway and core 2 GlcNAc-T product of the O-glycan pathway are preferred
intermediates for extension with polylactosamine chains, which add heterogeneity in the
form of polymer length and various capping sequences (Cumrnings and Komfeld, 1984;
Schachter, 1986; Yousefi et al., 1991). Galectins binding to N-acetyllactosamine, are
widely expressed, and have been implicated in tumor ce11 adhesion during metastatic
spread (Perillo et al., 1998). Gaiectins 1 and 3 are expressed on the surface of B 16
melanoma, the UV-2237 fibrosarcoma and the K-1735 melanoma cells, and have
previously k e n shown to facilitate organ colonization and metastasis by blood-borne
turnor ce1 1s (Ohannesian et al., 1995). Intravenous infusion of Ga1 or arabinogalactan
inhibited Iiver colonization by murine tumor cells, presumably by blocking their retention
to the microvasculature (Beuth et ai., 1987). LJDP-Ga1 transporter mutants lack
polylactosarnine in both 0- and N-glycans, and show the most severe attenuation of
tumor growth and metastasis. Restonng Ga1 to the surface of UDP-Ga1 transporter
mutants, using bovine B1,4Gal-T, increased tumor ce11 adhesion to non-activated
endothelial cells and enhanced metastasis of the mutants in mice (Comil et al., 1990).
Genetic revertants of the UDP-Gal transporter mutation also regain the malignant
phenotype (Dennis and Laferte, 1986).
The Lewis carbohydrate antigens Lex, Sialyl-Lex, L,ey and SLe' are ofien over-
expressed in human carcinomas and have been shown to mediate attachment of colon tumor
ce1 1s to selectins in vitro (Mannon et al., 1995). Pol ylactosamine forms the backbne of the
dimeric Lex and Lt? antigens, and dimenc Le sequences comlate with poor prognosis in
colon cancers (Hoff et al., 1989). E- and P- selectins on endothelial cells bind SLeX and

19
related sequences found on core 2 GlcNAc-branched O-glycans (Li et ai., 19%). Core 2
GlcNAc-T transcripts increase in human colon carcinomas, as do Fuc-TIV and ST3Gal II,
both of which are required for the biosynthesis of Slex (Ito et al., 1999; Shimodaira et al.,
1995). Indeed, inhibition of core 2 GlcNAc-T activity and GalNAc-Serm substitution in
vivo, by treating twnor cells with benzyl-a-GalNAc reduces organ colonization (Bresalier et
ai., 1991).
The normal function of selectin ligands, including Slex, is the mediation of
Ieukocyte attachment to blood vesse1 endotheliai ceils, an important step for Ieukocyte
extravasation (reviewed in (Vestweber and Blanks, 1999)). Leukocytes express selectin
ligands on ce11 surface glycoproteins like PSGL-1, which may not be expressed by cancer
cells. However, on cancer cells, selectin ligands may be overexpressed on other
glycoproteins, and thus may still be able to contribute to metastasis in vivo. Additionally,
by expressing selectin ligands, blood-borne tumor cells can aggregate with platelets and
leukocytes, which may express L-selectins and c m mediate attachment indirectly. Forced
expression of E-selectin in the liver of transpnic mice enhanced metastasis of Slex-
expressing B16F10 melanoma ceIl to the liver, rather than to their usual destination of the
lung (Biancone et al., 1996). However, it is unclear that attachment of blood-borne tumor
cells to endothelium is a rate-lirniting step in clinical metastasis for most tumors. Patients
with ascites turnors treated with peritoneal shunts to maintain their sait balance push
millions of peritoneal tumor cells into the circulation, but this does not significantly increase
the number of metastases observed at autopsy (Jamjoom et al., 1993).
Host anti-tumor Unmuni#y: Tumor-specific antigens are widely present but whether they
elicit meaningful anti-tumor immune response is still open for debate. For example, in
normal tissues, The O-glycan Tn and T antigens (GalNAc-polypeptide and
Galpl,3GalNAc-polypeptide, respectively) are shielded by subsequent branching or

20
capping, but -90% of these epitopes are uncovered in tumors. A twenty-year study
suggests that immunization with purified T n n antigen in adjuvant and a trace of the
h yperantigen phosphogl ycolipid A resul ts in signi ficantl y improved 5 and IO-year
survival (Springer, 1999). However, the study was conducted on a limited nurnber of
patients with tumor grades ranging from II to IV, al1 of whom were receiving additional
treatment, such as chemotherapy or radiation therapy. Thus, the benefit of Tmn
vaccination remains controversial.
Tumors produce cytokines such as TGF-f! and IL-10 that act to suppress cell-
mediated immunity (Le. Th 1 responses) (Botti et al., 1998), and prevalent carbohydrate
epitopes, such as Lex and polylactosamine may also suppress Th 1 responses. LeX and
polylactosamine sequences, found in Schistusoma mansoni eggs, have been shown to
suppress Th 1 response required to ciear the infection (Palanivel et al., 1996). Susceptible
mice mount the ineffective humoral Th2 response, which leads to tissue damage and
continued infection (Palanivel et al., 1996). Thus, it is suggested that Lewis sequences
and poiylactosamine on tumor cells may also suppress Th1 cellular immunity and T ce11
production of INF-y and IL-2 cytokines that can reduce the growth of some tumors.
Carbohydrate processing in hibitors as anti-cancer agents: The al kaloids sw ainsonine
and castanosperrnine block tumor ceil metastasis and invasion through extracellular
matnx in vitro. Swainsonine is a cornpetitive inhibitor of Golgi a-mannosidase II, which
bloc ks the N-gl ycan biosynthetic path way prior to g 1,6GlcNA~-branching, and resu!ts in
production of hybrid-type glycans (Figurel). Swainsonine-treated cells showed increased
transcription rates for tissue inhibitor of metalloproteinases (TïMP-1). Swainsonine also
suppresses MMP-2 expression in human tumor cells, a metalloproteinase assoçiated with
cancer progression in humans.

21 Swainsonine has k e n particularly useful for anti-cancer studies due to its
apparent lack of toxicity (reviewed in(Goss et al., 1995)). Swainsonine has been tested in
two phase I clinical trials with encouraging results that show both low toxicity and
evidence of clinical responses. Phase II trials designed to rneasure efficacy of oral
sw ainsonine treatment in rend ce11 carcinoma are currentl y being done. S wainsonine
s hould be considered a first generation compound, with room for i mprovement.
Swainsonine inhibits lysosomal a-mannosidases with potency equal to that of the Golgi
a-mannosidase il, and therefore induces lysosomal storage, and complex-type N- glycan
biosynthesis can proceed via altemate pathways that circumvent a-mannosidase II and
the swainsonine block (Romero and Herscovics, 1986; Chui et aI., 1997).
Somatic tumor ce11 mutants with a deficiency in UDP-Ga1 transport activity
show the most severe attenuation of malignancy, suggesting that a bloçker of lactosarnine
extension in N- and O-glycans rnay be an effective anti-cancer agent. Alternatively,
administering a combination of inhibitors of polylactosarnine extension and B1,6GlcNAc-
branching in the N- and 0- glycosylation pathways may prove to be a potent anti-cancer
strategy. These strategies will also circumvent side effects associated with carbohydrate
stomge in lysosomes. Further cancer studies with mutant rnice lacking specific
gl ycos yltransferase genes wi 11 increase our understanding of the relative importance of
these glycan structures to cancer growth and metastasis, and should provide useful
information for h g development efforts.
HERITABLE HUlMAN GLYCOSYLATION DISORDERS AND
ENGINEERED MUTATIONS IN MICE Somatic glycosylation tumor ceIl mutants do not have obvious cell-autonomous
phenotypes in tissue culture. This is perhaps not surprising, given the predicted role for

22 glycosylation in cell-ce11 and cell-ECM interactions. Therefore, analyses of whole
organisms are instructive when investigating the role of glycosylation in marnrnalian
bioIogy. The insights into the functions of specific glycosylated structures, provided by
rare human heritable disorders and engineered mutations in rnice are surnrnarized below.
Mutatons affectirzg initiafion and branching of N-glycum: The rare human hentable
disorder CDGSI (carbohydrate deficient gl ycoprotein syndrome type 1) is characterized
b y h ypo-N-gl ycos ylation of available si tes, and clinicall y presents with severe neurologk
dysfunction, rnultisystemic abnormalities and developmental delay observed in infancy
(Keir et al., 1999). The majority of CDGS type 1 patients are deficient in
phosphomannomutase activity, and are therefore depleted of GDP-mannose, a required
substrate for synthesis of dolichol-phosphate-mannose, dolichol pyrophosphate
oligosaccharide and glycosyl-phosphatidyl-inositol anchors (Hansen et al., 1997; Keir et
al., 1999). Consequently, the defect disrupts oligosaccharide transfer to Asn residues and
GPI anchor sites of nascent glycoproteins. CDGS type 1 patients show subnormal
mannose Ievels in serum, and the addition of mannose to cultured cells from CDGS type I
patients corrects the hypoglycosylation phenotype (Alton et al., 1999; Panneerselvarn et
al., 1999). Thus, norrnalizing blood mannose levels might correct protein
hypogIycosylation in some patients, an intervention now k i n g explored (Alton et al.,
1999).
GlcNAc-TI action is a prerequisite for the biosynthesis of the entire hybrid and
cornplex-type glycan series (Figure l), and in mouse appears to be encoded by a single
gene, Mgatl (Ioffe and Stanley, 1994; Metzler et al., 1994). Mgatl message is
ubiquitously expressed, as are the complex-type glycan products of the GIcNAc-Ti
enzyme (Campbell et al., 1995). GlcNAc-TI deficient mouse embryos die at around E9.5
due to a failure of multiple organ systems, including deformation of the neural tube (Ioffe

23 and Stanley, 1994; Metzler et ai., 1994). Complex-type glycans are obsewed in pre-E6
day ~ ~ a t l - ' - embryos, but are absent in E9.5 embryos, suggesting that materna1
compensation rnay prevent even earlier lethality (Ioffe et al., 1997). Lecl, a mutmt CHO
ceII line deficient in GlcNAc-TI, has no observable phenotype in tissue culture (Stanley,
1984), suggesting that GlcNAc-TI activity is not essential for cell-autonomous functions.
Consistent1 y, chimeric anal ysis of embryos has shown that GlcNAc-TI defîcient ES ceils
were able to contribute to every tissue with the exception of bronchial epithelium (Ioffe et
al., 1996). This finding also implies that GlcNAc-TI mediated gtycosyiation does not
have an essentiai ce11 autonomous role in the majority of tissues, but is required for the
differentiation and/or maintenance of bronchial epithelium. Cunously, ~ ~ a t l ~ - ES cells
aIso contributed significantly less to bronchial epithelium, suggesting that a full dose of
GlcNAc-TI activity is required for the proper formation of this ceIl layer. The degree to
which the nuIl cells in other tissues are functionally normal by vinue of their contact with
wild-type cells remains to be d e t e d n e d .
Following the action of GlcNAc-Ti, the majority of complex N-glycan structures
are processed by a-mannosidase II. However, an alternate route to complex N-glycans
was discovered by analyzing residual glycan structures and activities in the a-
mannosidase iI deficient mice (Chui et d., 1997). A new enzyme, a-mannosidase m,
which uses Man5GIcNAcr as substrate (Figure 1), was detected in most tissues of the a-
Mannosidase 11 deficient mouse, resulting in a partial loss of complex-type N-glycans
only. In erythroid cells, a-mannosidase III activity is not evident, and therefore complex
N-glycans are not synthesized. The a-mannosidase II deficient mice are viable and
fertile, but develop a condition similar to human congenital dyserythropoietic anemia
(CDA type II), also known as HEMPAS, a genetic disorder characterized by anemia,

24
splenomegaly and marrow erythroplasia, but without neurological involvement. CDA II
lymphocytes and red cells exhibit decreased complex-type glycan content, and in some
patients, reduced a-mannosidase II levels were observed- However, the presence of
normal a-mannosidase II levels in some patients suggests that the syndrome may have
multiple biochemical causes. In patients with a-mannosidase II deficiency, the exact
genetic defect remains unidentified, but may involve aberrant transcriptional regulation
(Iolascon et al., 1999).
GlcNAc-TII uses the a-mannosidase II-trirnmed core as substrate, and its activity
is required for the biosynthesis of complex N-glycans (Schachter, 1986). In its absence,
hybrid-type structures are forrned and only a portion of sialyllactosamine antennae can be
added (Figure 1). Like GIcNAc-TI, its activity appears to be ubiquitous. Mice lacking
GlcNAc-TII are runted, and die at or shortly after birth with multiple organ defects
(Priatel et al., 1997). As is the case with GlcNAc-TI, murine GlcNAc-TLI activity
appears to be encoded by a single gene. GlcNAc-'III deficiency causes the rare human
autosomal recessive disease, carboh ydrate-deficient gl ycoprotein syndrome type II
(CDGS type II) (Jaeken et al., 1994). CDGS type II is characterized by multisystemic
involvement and severe impairment of the nervous system. Inactivating point mutations
in the catalytic domain of the GlcNAc-TI1 gene (Ser290Phe and His 262Arg) have k e n
identified in two unrelated patients (Tan et al., 1996).
Substitution by GlcNAc-TIII adds a bisecting GlcNAc-residue in a $1,4 linkage
to the underlying mannose of the N-glycan core, and renders the glycan chains poor
substrates for a-mannosidase II, GlcNAc-TII, GlcNAc-TIV and GlcNAc-TV, thus
divening the pathway towards hybrid glycans as shown in Figure 1 (Schachter, 1986).
GlcNAc-TIIi activity is normally detected in brain, kidney and B cells (Miyoshi et al.,

25
1993). Mice deficient in GlcNAc-Tm are viable and fertile with no obvious
abnonnalities (Priatel et al., 1997; Bhaurnik et al., 1998). Although the Celegans
genome has the coding potential for most known glycosyltransferse genes, GlcNAc-TIII
appears to have no homologue in the worm (C.E.Warren, personal communication). It is,
therefore, likely that GlcNAc-Tm function is specific to vertebrates or marnmals, and
therefore GlcNAc-Tm deficient mice might respond aberrantly to stimuli of higher
organisms, such as stressors of the immune system or carcinogens. Indeed, GlcNAc-TIII
deficient mice respond differently to diethylnitrosamine than do wild type mice.
Exposure to diethylnitrosamine results in hepatoçarcinogenesis, which is inhibited by
GlcNAc-Tm deficiency. However, Mgat3, the gene encoding GlcNAc-Tm, is not
expressed in normal or transformed hepatocytes, suggesting that a host factor is
dependent on GlcNAc-Tm-dependent glycosylation to promote cancer progression
(Bhaumik et al., 1998).
Mutations aflecting O-glycosylation: To date, few somatic or naturall y occumng
mutations have k e n identified that specifically affect O-glycan biosynthesis. Tn
polyagglutination is a notable exception. This is a rare, acquired hematological
abnormality affecting a subset of bone marrow stem cells, where a loss of 1.3Gal-T
activity due to transcnptionai repression is obsewed (Thurnher et al., 1999) (Figure 2).
Core 2 GlcNAc-T initiates f!1,6GlcNAc-branching of O-glycans, and its expression is
subject to tissue-specific regulation (Granovsky et al., 1995). The antenna initiated by
core 2 GIcNAc-T forms the primary scaffold for polylactosamine and SLex on O-glycans
of PSGL-1, and is required for P- and E- selectin binding (Ellies et ai., 1998; Li et ai.,
1996). Pol ylactosamine on f! 1,GGlcNAc-branched O-gl ycans of CD43 and the CD45 on
thymocytes bind to the mammalian lectin, galectin-1, found on thymic epithelial cells and
this interaction causes enhanced apoptosis (Perillo et al., 1995).

26 CD43 acts as an anti-adhesive at the leading edge of migrating T cells and
faci l i tates extravasation from the blood into secondary 1 yrnphoid tissues (Manjunath et
al., 1995; McEvoy et al., 1997). Core 2 GlcNAc-T activity and branched O-glycans on
CD43 are upregulated in peripheral T and B cells following activation by antigens,
possibly enhancing the anti-adhesive properties of CD43 (Piller et al., 1988). The high
density of O-linked glycan chains on CD43 results in an extended protein structure
protruding from the ce11 surface on T lymphocytes, activated B cells, NK cells and
immature dendritic cells (Cyster et al., 1991). Transgenic mice expressing Core 2
GlcNAc-T under the regulation of the T-ce11 specific Lck promoter, display reduced
delayed type hypersensitivity reactions in vivo. T-cells from these mice adhere less to
fi bronectin and ICAM- 1 (Tsuboi and Fukuda, 1997).
The gene encoding Core 2 GIcNAc-T has been knocked out in rnice. Deficient
mice are viable and fertile, with no detectable core 2 GlcNAc-TV enzymatic activity or
fi 1.6GlcNAc-branched O-glycans. The rnice develop neutrophilia and partial deficiency
of selectin ligands (Ellies et al., 1998). Loss of core 2 oligosaccharides reduces
neutrophil rolling in vitro and neutrophil recruitrnent to sites of inflammation in vivo.
However, although core 2 GlcNAc-T deficiency significantly reduces E-, L- and P-
selectin ligands on leukocytes, it does not ablate the presence of selectin ligands entirely,
suggesting that ~Iigosaccha~îdes other than $1,6GlcNAc-branched O-glycans can be
substrates for selectin ligand biosynthesis. It is surprising that B1,6GlcNAc-branching of
O-glycans is not detected in the core 2 GlcNAc-T deficient mouse because a second gene
encoding an enzyme with core 2, core 4 and 'Tl catalytic activity has been isolated and
characterized (Yeh et al., 1999). Therefore, it is possible that the relatively mild
phenotype observed in this mouse is due to compensation by this second gene.

M e n s affecfing Glycun elongritron: Branched N- and O-gl ycans, as well as gl ycolipids
are elongated by polylactosamine c h a h (2 to >10 repeating uni& of Ga@ 1,4GlcNAc$1,3).
p l PGal-Ts substitute GIcNAcS- antennae, and their action is essential for polylactosttmine
production (Figure 1). The predominant marnrnalian enzyme. $1 ,4Gai-T 1 has two distinct
functions. In the crans-Golgi network of most cells, the enzyme participates in
pol ylactosamine production, but in lactating mammary gland, $1 ,&al-Tl complexes with
a-lactalbumin, altering the enzyme's catalytic specificity to favor glucose over GlcNAc
(Bell et al., 1976). Under these circurnstances, the enzyme preferentialIy participates in
lactose biosynthesis. $1 ,&alTl deficient mice show a marked decrease in galactosylation
of semm glycoproteins (Asano et al., 1997). These mice are viable and fertile, but exhibit
g o w t retardation and eady lethaiity. This gmss phenotype might be related to the
observed abnormalities in epithelial ce11 proliferation and differentiation of skin and
intestine, which suggest that p l ,4GalT 1 is required for the maintenance of the epithelial ce11
layer after birth.
fi I ,4GalT expressed on the surface of sperm may bind the egg coat glycoprotein
ZP3, thus mediating garnete recognition. Indeed, p 1,4GalT 1 deficient spem bound less
ZP3 than wild type spenn, and were unable to undergo acrosome reaction in response to
either ZP3 or anti-galactosyltransferase antibody, which rendered them physiologically
inferior to wild-type sperm. However, $1,4GalTl nul1 males are fertile, indicating that
efficient ZP3 binding and the subsequent induction of the acrosome reaction are not
essential for fertilization (Lu and Shur, 1997).
The viability and fertility of $1,4GalT1 deficient mice niight be attributable to
compensation by other enzymes possessing similar activity. To date, 5 additional human
p l ,4Gal-Ts have been identified and cloned with the aid of ESTs. These enzymes,

28
designated $1,4Gal-TII through fi1.4Gai-TV1, have 33% to 55% sequence identity to
B1,4Gal-Ti (Lo et al., 1998).
Mutdons affecting glycan capping: The terminal sequences of N-and O-glycans
usually contain fucose andor sialic acid residues. There are 8 marnrnalian genes
encoding fucosyltransferases and at least 13 genes encoding sialyltransferases (reviewed
in (Oriol et al., 1999) and (Tsuji, 1996)). Fucose residues are present on N- and 0-
glycans in one of four possible linkages (a1,2, a1.3, a1,4 and al$). Sialyltransferases
transfer sialic acids, of which there are 20 isotypes, in four possible linkages with
remarkable specificity for the c h h y d r a t e acceptor (aî ,3Gd, or2,6Gal, at2,6GalNAc
and a2,8Sia) (Tsuji, 19%). As has been previously discussed, genetic and epigenetic
diversity of terminal oligosaccharide sequences in the human population may have
evolved as a defense against bacteriai and viral pathogens, including influenza A, E. coli
0 1 5 7 , and NeissenQ gononheu, al1 of which exploit glycoconjugates in their host
recognition and subsequent pathogenicity. For example. influenza A hemagglutinin is
known to bind sialic acid in an isotype and linkage-specific manner, which affects the
tissue tropism and the infectivity of the virus (Pritchett et al., 1987).
Fucosyltransferases are ancient glycosyltransferases, conserved from prokaryotes
to mammals (Oriol et al., 1999). The bacterium Helicobacter pylon is known to
spthesize Lex epitopes. for which al,3Fuc-T activity is required. The al3Fuc-T gene
found in another bacterium, Schisrosoma mansoni appears to be munne in origin (Onol et
al., 1999). This gene encodes a fucosyltransferase with siaiyllactosamine acceptor
specificity, an acceptor that is not synthesized by Schistosoma mansoni. It is suggested
that the parasite has incorporated a fragment of mouse DNA into its genome. It would be

29
interesting to know what selective advantage, if any, the bacterium derives from
incorponting this mammalian fucosyltransferase into its genome.
In mamrnals, al.3Fuc-Ts synthesise the Lewis epitope, SLex, on E- and P-
selectin ligands. The selectin ligands are a subset of fucosylated O-glycan structures on
PSGL- 1, ESL-1, GlyCAM- 1, CD43 and MadCam (Springer, 1994). Selectin-Selectin
ligand interactions enable leukocytes to roll on endothelial cells, a prerequisite for
extravasation into sites of infiamation (Vestweber and Blanks, 1999). Five human genes
encode al,3Fuc-Ts (m, N, V, VI, W. Genes encoding only 2 of these enzymes,
a l ,3Fuc-TV11 and al,3Fuc -TW, are expressed in leukocytes, and al,3Fuc-TVII appears
to be the most active in the synthesis of SLex (Knibbs et al., 1996). Mice lacking
al ,3Fuc-TVII develop blood leukocytosis due to a deficiency in leukocyte homing and
recirculation (Maly et al., 1996). These mice also have poor leukocyte extravasation into
areas of inflammation. al3~uc-TVE'- mice are phenotypicaily similar to P- and E-
selectin deficient mice and to individuais with LADII. Leukocyte adhesion deficiency II
syndrome (LADII) is a rare human congenitai disease caused by a defect in GDP-fucose
biosynthesis, and is characterized by immunodeficiency and a severe delay in wound
healing (Karsan et al., 1998). As discussed above, fucose is essential for the biosynthesis
of selectin ligands and is, therefore, important in neutrophil and lymphocyte homing.
In contrast to the severe consequences of al,3Fuc-TVn deficiency, inactivating
mutations in al3/4Fuc-TIII are common in the population and impart no obvious
phenotype (Orntoft et al., 1996). Sirnilarly, lack of functional al,2Fuc-TI and TII genes,
required for the biosynthesis of A B 0 and H blood group oligosaccharide antigens, occurs
with no evidence of pathology, and inactivating mutations in al,3FucTVI also cause no
apparent harm (Kelly et ai., 1994).

30
Fucos y ltransferases and sial yltrans ferases often compte for the same acceptors.
Sialylation is absent in C. elegans, which cap its glycans with fucose (Oriol et al., 1999),
suggesting that sialylation may be required for functions restricted to vertebrates. The
roles ascribed to siaiylation include enhanced cellular migration during
neurodevelopment and regulating leukocyte extravasation from blood to sites of
inflammation and lymphoid organs (Sjoberg et al., 1999). The regulated expression of
sialic acids is due in large part to the spatial and temporal pattern of expression of
sialyltransfenses. Tbese enzymes are grouped into four families according to the
carbohydrate linkages they produce; ST3Ga1, ST6Gd, ST6GalNAc, and STSSia, and are
sensitive to the oligosaccharide composition of the acceptor beyond the specific sugar
residue to be substituted (Sjoberg et al., 1999). In mammals, ST6Gal sialyltransferase
appears to be solel y responsi ble for producing the Siaa2,6Gal$l,4GlcNAc terminus on
N-glycans and possibly on some O-glycans (Hennet et al., 1998)- The transcription of
thi s gene is regulated by multiple promoters and altered by glucocorticoids and cytokines.
ST6Gal is unique in producing the ligand for the CD22 lectin molecule expressed on B
cells, a member of the 1-type lectin farnily, which also includes myelin-associated
glycoprotein (Sgroi et al., 1993). As CD22 itself carries Sia6LacNAc, homotypic binding
interactions have been shown to occcur and may play a regulatory role in immune
function. FolIowing antigen stimulation of B cells, the cytosolic domain of CD22
recruits the SHPl tyrosine phosphatase into the Ig receptor-signaling cornplex, and
dampens intracellular signaling. Predictably, therefore, B ceIls in mice lacking CD22 are
hypersensitive to antigens, while ST6Gal deficient mice show impaired B ce11 maturation
and IgM production. The phenotype of ST6Gal deficient rnice appears to be more severe
than the phenotype resulting from CD22 deficiency, suggesting that there might be other,

31
as yet unidentified, Sia6LacNAc lectins with regulatory functions in B cells (Hennet et
al., 1998).
As described above, whole animal models of aberrant glycosylation provide new
insights into the function of specific carbohydrate structures. In particular, immunity and
cancer progression and metastasis are complex phenornena, which involve many cellular
responses to the cell's environment, such as signal-induced proliferation andor
differentiation, and ECM-rnediated cellular adhesion and migration. The complexity of
these interactions cannot be fully mimicked in tissue culture, and therefore in vitro
models have limited use for studying the role of glycosylation in these processes. In this
regard, the developrnent of gene knockout technology has been a timely blessing for the
study of biological functions of glycosylation.
GlcNAc- TV
GlcNAc-TV is a branching enzyme in the complex-type N-glycosylation
biosynthetic pathway (Cumrnings and Komfeld, 1984). As such, it adds complexity to
the N-gtycans it acts upon, by providing the B1,aGlcNAc-branch, an enzymatically
favored substrate for the addition of p l ylactosamine c hains. Indeed, GlcNAc-TV
activity appears to be rate lirniting for polylactosamine addition to N-glycans in tumor
cells (Easton et al., 1991). For example, GicNAc-TV deficiency in MDAY-D2 cells
results in significant loss of polylactosamine on N-glycans, but not on O-glycans
(Yousefi et al., 1991).
The rat gene for GlcNAc-TV eiicodes a 740 amino acid (aa) type II
transmembrane protein (Shoreibah et al., 1993), a structure shared with dl
glycosyltransferases cloned to date. Type II transmembrane proteins comprise a shon

cytoplasrnic N-terminus tail, a single hydrophobie transmembrane domain followed by a
proline-rich stem region, and a large cataiytic domain at the C terminus. The C-terminal
portion of rat GIcNAc-TV (aa 2 13-740) is essentid for catalytic activity (Korczak et al.,
1997), and the transmembrane domain and at les t one residue in the stem region (aa 188)
are required For Golgi Iocalization. Two CHO mutant ce11 lines, Lec4 and Lec 4A, Iack
fi 1,6GlcNAc-branched N-glycans on their ceIl surface (Weinstein et ai., 1996). Lec4
cells do not have GlcNAc-TV activity, due to the dîsruption of the gene encoding the
enzyme, but Lec4A cells have GlcNAc-TV enzymatic activity. The gene e n c d n g
GlcNAc-TV in Lec4A cells has a missense mutation, converting leucine to arginine at
position 188 (L188R), causing loss of Golgi Iocalization. Therefore, the LI 88R mutation
causes a GlcNAc-TV deficient cellular phenotype by mislocalizing the enzyme without
affecting catal ytic activity.
GlcNAc-W is conserved in metazoans. The C,elegans gene, Gly-2, encodes a
protein possessing GlcNAc-TV enzymatic activity (Warren and Dennis, 1999,
subrnitted). The gene is able to rescue the ce11 surface phenotype of Lec4 cells, but not
when a L116R mutation, analogous to the L188R mutation in Lec4A, is introduced.
Thus, Gly-2 is orthologous to the mairnalian genes encoding GlcNAc-TV. It is
structurally conserved at both the genornic and peptide levels, and it is functionally
interchangeable with mammalian GlcNAc-TV. Such "deep homology" has been
demonstrated for only a few genes, the products of which are important within ancient
and conserved pathways.
Gly-2 and mammalian genes encoding GlcNAc-TV are multi-exonic, and the
structure of the gene (i.e. intron-exon boundries) is conserved between wonn and human.
The human gene is comprised of 17 exons spanning 155kb and is located on chromosome
2q2 1 (Saito et ai.. 1995). Database searches and the characterization of the ~ ~ a t . 5 ' -

33
mouse (described in Chapter 3 of this thesis) suggest that mammalian and worm
GlcNAc-TV activity is encoded by a single gene. This is an unusual feature, as many
gl ycosyltransferases are encoded by fami lies of genes in C-elegans ancilor in mammalian
species.
GlcNAc-TV gene expression and enzyme activity appear to be regulated in a
complex fashion. Mgat.5 gene transcription is regulated by Ras activation (Chen et al.,
1998; Dennis et al., 1987). The promoter region of the human gene contains M l , AP2,
HNFl, HP1 and PEA3/ets binding sites (Saito et ai., 1995). The v-src oncogene has been
shown to stimulate transcription from the MgatS promoter by activating Raf-1 kinase and
the Ets-2 transcription factor (Buckhaults et al., 1997). GlcNAc-TV message is
detectable in most rat and mouse tissues, dbeit in greatly varying amounts (Granovsky et
al., 1995; Perng et al., 1994). In addition, levels of GlcNAc-TV mRNA do not
necessarily reflect the levels of enzymatic activity in a particular tissue. For example, rat
brain has the highest level of GlcNAc-TV transcripts but low levels of enzymatic activity
and product, suggesting additional pst-translational andor pst-transcriptional
regulatory mechanisms (Perng et al., 1994). Intriguingly, transfectants with high
expression of GlcNAc-TV do not survive in tissue culture and Mgat5 transfection
enhanced apoptosis, suggesting that there is selection against overexpression of GlcNAc-
TV (Demetriou et al., 1995). Thus, proper regulation of GlcNAc-TV activity may be
important for cellular viability as well as specific glycoprotein functions.
B 1,oGlcNAc-branched N-glycans, as detected by L-PHA lectin
immunohistochemistry were detected in the majority of epithelial cell populations, with
the exception of breast and colon epithelia in rat and human tissues (Li and Roth, 1997).
L-PHA is a plant lectin with specific binding to Galf3f AGlcNAcf31,6-
(Ga@ 1,4GlcNAcS 1,2)Mana of tri' and tetraantennary N-glycans (Cummings and

34
Kornfeld, 1982). High L-PHA reactivity and GlcNAc-TV activity were detected in the
small intestine and in the kidney (Dennis and Laferte, 1989). In addition, activation of
human CD4+ and CDS+ thpocytes, which induces high cellular proliferation and
migration, results in increased GlcNAc-TV activity and f31,oGlcNAc-branching of N-
glycans (Lemaire et al., 1994).
GlcNAc-TV activity and B1,oGlcNAc-branching of N-glycans have been
correlated with cancer progression and metastasis. $1,6GlcNAc-branching of N-glycans
increases in BHK cells transformed with poIyomavirus or RSV virus (Pierce and Arango,
1986; Yarnashita et al., 1984). GlcNAc-TV activity increased in NIH3T3 cells
transfomed with activated Ras, and in rat2 fibroblasts and SPl epithelial cells transfected
with activated Ras or the v-Fps oncogene (Dennis et al., 1989; Dennis et al., 1987;
Heffeman et al., 1993; Lu and Chaney, 1993). This enhancement correlated with
phenotypic changes consistent with morphological transformation. increased L-PHA
reactivity was reported in a variety of malignant and invasive human tumors, including
melanomas and tumors in breast and colon. For example, L-PHA staining was
significantly increased in 50% of malignant human breast biopsies and intensity of L-
PHA staining significantly correlated with the progression of these malignant tumors
(Fernandes et al., 1991). In addition, L-PHA staining in human colorectal carcinoma
sections provides an independent prognostic indicator for tumor recurrence and patient
outcome, and is associated with the presence of lymph node metastases (Seelentag et al.,
1998).
There is evidence that the relationship between increased p1,6 branching of N-
glycans and aspects of the malignant phenotype is causal. Somatic mutant tumor cells
selected for reduced L-PHA reactivity and GlcNAc-TV activity are comprornised in their
ability to form lung colonies p s t i.v, injection, while ce11 lines with increased sensitivity

35
to L-PHA show enhanced metastatic potential (Dennis et al., 1986; Dennis et al., 1987).
In addition, N-glycosylation inhibitors, such as swainsonine, are able to slow solid tumor
gowth in mice and affect proliferation of munne lymphoma and human sarcoma lines in
serum free medium (Goss et al., 1995). Two genetic studies looked at the specific d e of
GlcNAc-TV in transformation and metastasis. Demetriou et al. transfeçted rat GlcNAc-
TV cDNA into MvlLu cells, a premalignant irnmortalized rnink lung epithelial ce11 Iine
(Dernetriou e t al., 1995). The forced expression of GlcNAc-TV and the subsequent
increase in B i ,6-branched N-glycans resulted in toss of contact-inhibition of ce11 growth.
GlcNAc-TV transfectants formed foci in serum-deprived and high density monolayer
cultures, and maintained microfi larnent structures consistent with a proli ferative state.
These phenotypic traits correlated with the degree of GlcNAc-TV expression. In vivo,
transfectants with the highest Ievels of GlcNAc-TV expression and activity were able to
form solid tumors in nude mice (Demetriou e t al., 1995).
More recently, Seberger and Chaney studied metastatic and transformed ce11 lines
transfected with GIcNAc-TV cDNA (Seberger and Chaney, 1999). The transfection
resulted in an increase in the metastatic potential of these cells by 4-40 fold. The
increased metastatic potential was not associated with a gross change in cellular growth
rate as measured in tissue culture, nor with aberrant adhesion to extracellular matnx
proteins, such as fibronectin or collagen type TV.
In order to understand the role of GlcNAc-TV in marnmalian development and in
cancer progression and metastasis, we undertook to study its expression pattern during
murine embryogenesis and to generate GlcNAc-TV deficient mice. We have confimed
that GlcNAc-TV is not ubiquitously expressed, and that its expression is highest in cells
with stem-cell-like properties, i.e. possessing regenerative and migratory capacity. The
knockout mouse has provided us with a possible explanation of how Bl,6GlcNAc-

36 branched N-glycans participate in such processes as T ce11 and tumor cell proliferation,
and leukocyte adhesion and migration. The data suggest a direct regulatory role for
GlcNAc-TV in T ce11 receptor and inteHn trammembrane receptor aggregation and
subsequent signaiing.

37
TKESIS OUTLINE
The cDNA encoding rat GlcNAc-TV was cloned shortly before 1 started my studies.
We were, therefore, in a position to take a genetic approach to study the function of
1.6GicNAc-branched N-glycans.
1 fitst detennined the temporal and spatial expression pattern of Mg&, the gene
encoding GlcNAc-TV, during murine embryogenesis. High Mgut5 expression was
observed in the @nt cells of the trophoblast, basal epithelia in skin and intestine and in the
cortical neural epithelium. A cornmon feature of these cells is their capacity to migrate, a
cellular function that may require GlcNAc-TVdependent glycosylation. The detailed
expression pattern is described in Chapter 2.
To study the rote of GlcNAc-TV in mammalian biology, we generated GlcNAc-TV
deficient rnice by targeting the Mgal5 locus by homologous recombination in embryonic
stem (ES) cells. Chapter 3 describes these mice, with particular emphasis on the function of
leukocytes and T-lymphocytes. We hypothesized that if 1,GGlcNAc-branched N-glycans
regulate cellular migration, leukoc yte migration in response to in flammatory stimuli would
be altered by GlcNAc-TV deficiency. Two independent models of acute inflammation were
used to test this hypothesis. ln addition, a delayed type hypersensitivity mode1 was used to
study the effect of GlcNAc-TV deficiency on T cell function in vivo. ~gat.5'- mice
responded more strongly to the delayed type hypersensitivity antigen than did wild type
mice. This result prompted an in vitro study of Mgut.? T ce11 response to rnitogens.
We hypothesized that tumor progression and metastasis may be impeded in
GlcNAc-TV deficient rnice. This hypothesis was based on work with GIcNAc-TV deficient
somatic tumor cell mutants done previously in the Dennis lab and in several other labs,
consistently showing slower tumor progression and reduced metastasis. In addition, studies

38
with MvLul cells and mammary carcinoma ceil lines transfected to overexpress GIcNAc-
TV, suggest that GlcNAc-TV activity may be required for transformation and metastasis.
Since al1 previous studies have been done with transplanted tumor cells, which were already
fully transformed, the role of GlcNAc-TV in tumor initiation, or the effect of GlcNAc-TV
deficiency in the host animal on Nmor progression could not be studied. Chapter 4
describes the effect of GlcNAc-TV on tumor formation and development in a whole animal
mode1 in which GlcNAc-TV deficient mice were crossed with transgenic mice expressing
the polyorna middle T oncogene under the control of a marnmary-specific promotet
(MMTV-F'yMT). PyMT is a potent oncogene, and its transfoming activity is dependent on
its ability to associate and activate c-Src, PI3K and Shc. Expression of the PyMT transgene
in mammary epi thelium results in global hyperplasia, rnulti focal mammasy
adenocarcinornas and frequent metastases to the lung (Guy et al., 1992). Therefore, this
cross gave us the opporîunity to study the effect of GlcNAc-TV on tumor formation, tumor
growth, metastatic potential and oncogenic pathways dependent on GlcNAc-TV mediated
glycosylation.

CHAPTER 2: GlcNAc-TV and Core 2 GlcNAc-T
Expression in the Developing Mouse Embryo.
This chapter was pubtished as follows:
Granovsky, M., Fode, C., Warren, C.E., Campbeü, R.M., Martb, J.D, Pierce, M., Fregien, N., and Demis, J.W. ( 1995). GlcNAc-transferase V and core 2 GlcNAc-transferase expression in the developing mouse embryo. Glycobiology 5,797-806.
Reproduced by premission o f Oxford University Press

40
ABSTRACT
UDP-G1cNAc:Mana l.6Man P-R P 1,6-N-acetylglucosamin yltransferase V (GlcNAc-
TV) and UDP-GlcNAc:Gal p 1,3GaWAc-R 1,6-N-acetylglucosamin yltransfense (core 2
GlcNAc-T) are Golgi enzymes that catalyze the biosynthesis of P1,ffilcNAc-branched
intermediates in the N- and O-linked biosynthesis pathways, respectively. The activities of
these enzymes change dunng haematopoiesis, embryocarcinorna ce11 di fferentiation and
following maiignant transformation, but little is known about their expression in normal
adul t tissues and during embryogenesis. We have examineci the expression of GlcNAc-TV
and core 2 GlcNAc-T in sections of pst-implantation mouse embryos by in situ RNA
hybridization. The two enzymes showed distinct temporal and spatial patterns of
expression. Core 2 GlcNAc-T mRNA was widely expressed at embryonic day (E) 7 and
becarne restricted to a subset of mucin-producing and cartilagenous tissues at El 1.5 through
to El75 GlcNAc-TV transcripts were absent at E7, becarne expressed throughout E9.5
embryos, and then pnigressively restricted to regions of the developing central nervous
system (CNS) and to speciaiized epithelia of skin, intestine, kidney, endocrine tissues and
respiratory tract. In the adult gonads, GlcNAc-TV trancnpts were excluded from germ cells
but were detected in the follicular and testicular cells. Leukoagglutinin (L-PHA)- reactive
oligosaccharides CO-localized with GlcNAc-TV transcripts in skin, kidney and intestine, but
brain showed unexpectedly low overail staining punctated by bright staining of the vascular
endotheliurn. A common feature of cells in basal epithelia and in the cortical neural
epithelium is the capacity to migrate, a cellular function which may require GtcNAc-TV - dependent gl ycoconjugates.

41
INTRODUCTION
Of igosacc harides, such as the stage-speci fic embryonic an tigens (eg. Lewis
an tigens), show precise spatial and temporal patterns of expression during mouse
embryogenesis (Childs et ai., 1983; Feizi, 1985; Hakomori and Kannagi, 1983; Pennington
et al., 1985; Solter and Knowles, 1978). These c h h y d r a t e sequences fom the termini of
N- and O-Iinked oligosaccharides and can serve as ligands for lectins, or may directly
mediate cell-cell interactions by homotypic binding (Eggens et al., 1989; Fenderson et al.,
1990). Multivalent conjugates of Le' have been s h o w to inhibit mouse embryo compaction
in vitro, suggesting that they may participate in cell-cell interactions at this early stage mird
and Kimber, 1984; Fenderson et al., 1984). In adult tissues, sialylle' sequences are
expressed on neutrophil ce11 surface glycoproteins which can mediate ceIl adhesion by
binding to selectins found on activated or darnaged vascular endothelia (Spinger, 1994).
Sirnilar interactions may be involved in cancer ce11 invasion and metastasis, as these
embryonic carbohydrate sequences are cornrnonly re-expressed in cancer cells (Dennis,
1991).
G d (3 l,3GalNAca-Sermr and Mana l,3 Mana l,6]Man p 1 -4GlcNAc p 1 SGlcNAc-
Asn, comrnon core stmctures for 0- and N- linked glycans respectively, are substituted to
varying degrees by branching GlcNAc-Ts in the medial Golgi. This ailows for considerable
structural diversity as the GlcNAc residues can be extended by the many
glycosyltransferases of the tram-Golgi to produce different tenninal sequences. Acceptor
specificity and cornpetition between glycosyltransferases for common acceptors (Schachter,
1986), coupled with tissue-specific expression of the enzymes, appear to be major factors
regulating the distribution of oligosaccharide stmctures (Paulson and Colley, 1989a; Paulson
et al., 1989b). GlcNAc-TV substitutes GlcNAc p 1,2Mancil,6Manp- of the core N-linked
structure at the 6 position of the a1,6 linked mannose, creating a preferred acceptor for

42
polylactosamine extension and associated Lewis antigens (van den Eijnden et al., 1988;
Yousefi et al., 1991). In the O-linked oligosaccharide pathway, core 2 GlcNAc-T plays a
similar role, converting core 1 (ie. GalQ1,3GlcNAccr-O) to core 2 (ie.
Ga1 p 1,3(GlcNAc p 1,6)Gal.NAcar-O), thereby creating a branch for the addition of
polylactosamine (Carlsson et al., 1986; Yousefi et al., 1991). Oncogene-induced
transformation of fibroblasts (Dennis et al., 1987; Lu and Chaney, 1993; Pierce and Arango,
1986; Yarnasfuta et al., 1985), differentiation of teratocarcinoma cells (Heffernan et al.,
1993), and activation of lymphocytes (Lemaire et al-, L994) have been show to induce
GlcNAc-TV activity and N-glycan-associated pl ylactosamine. Core 2 GlcNAc-T activi ty
and O-glycan associated polylactosamine is also elevated following lymphocyte activation
(Higgins et al., 1991; Piller et al., 1988) and in some malignant cells (Higgins et al., 1991).
Therefore, GlcNAc-TV and core 2 GlcNAc-T activities are developmentally-regulated, and
these enzymes control expression of polylactosamine and associated terminal carbohydrate
sequences in N- and 0- linked glycan biosynthesis pathways.
To initiate a genetic study of the d e of GlcNAc-transferases in mouse development,
we have examined the tissue distribution of core 2 GlcNAc-T and GlcNAc-TV transcripts
dunng mouse embryogenesis by in situ RNA hybridization. GlcNAc-TV and core 2
GlcNAc-T showed distinctive temporal and spatial patterns of expression in post-
implantation embryos. Core 2 GlcNAc-T was expressed at E7 prior to organogenesis and
becarne restricted to mucin-producing and cartilaginous tissues in later-stage embryos.
GIcNAc-TV was upregulated in a widespread manner in E9.5 embryos, and later became
confined to several specialized epithelial ce11 layers, as well as cells found in the
neuroepithelium of the developing CNS.

43
RESULTS
GlcNAc- T V expression in post-Unplanrotion mouse embryo: Implantation of blastoc y st
stage embryos into the uterine epithelium takes place between E4.5 and E6, and by E7 in
primitive streak embryos, the three embryonic ce11 layers can be readiiy be identified in
sagittal sections. GlcNAc-TV transcripts were highly expressed in the giant cells of the
trophoblast and in the decidua basalis at E7 but absent in the embryo (Figure 1). To
compare expression of GlcNAc-TV and its products, lectin histochemistry was also
preformed on embryo sections. 1,6GlcNAc-branched oligosaccharides were detected
with L-PHA (Curnmings and Komfeld, 1982), which showed very low reactivity on al1
three ce11 layers of the E7 embryo when compared to the deciduum which stained heavily
(Figure 2). However, ConA and E - P m stained the E7 embryo intensely, suggesting that
bianntennary andor high mannose -type as well as complex-type N-linked
oligosaccharides were present, while structures with f3 1,aGlcNAc-branching were absent.
GlcNAc-TV transcript levels were greatly increased and widely distributed in the E9.5
embryo, with particularly high signals observed in the developing neuroepithelium of the
CNS (Figure 1). In E9.5 and E10.5 embryos, transcripts were distributed throughout the
ventricular zones of the CNS, the region where the actively dividing neuroblasts are
located. By El 1.5, the accumulation of GlcNAc-TV message shifted to the intermediate
and marginal zones of the spinal cord, the cranid flexure, the lamina terminalis and the
fourth ven tricle. Later stage embryos consistent1 y expressed high levels of message in
the marginal and intermediate zones of the CNS which are the more differentiated areas
of the nervous system compared to the underlying ventricular zone.

44
Figure 1: In situ localkation of GlcNAc-TV (TV) and core 2 GlcNAc-T (C2)
expression during mutine embryogenesis.
Bright- and darkf~eld photornicrographs of sagittal sections through embryos at E7, E9.5,
E 13.5 and E17.5 as indicated at the left, Tissue sections were prepared and hybridized with
antisense RNA probes as described in Materials and Methods. Labels are: aç, amniotic
cavity; ex, extraembryonic region; et ectoplacental cone; fb, forebrain; gu, gut; h, hem; hb,
hindbrain; k,kidney; li, liver; lu, lung; mb, midbrain; ne, neuroepitheliurn; ot, otic vesicle; rp
Rathke's pouch; SC, spinal cord; t, trophoblast; v, vertebrae; wf, whisker follicles; iii 3rd
ventricle; iv, 4th ventricle; ba branchial arches; tr, thyroid; th, thymus; p, pancreas; O,
01 factory epithelium.


Figure 2: Lectin histmhemistry of the E7 embryo.
E7 ernbryos were stained with (A) E-PHA, (B) L-PHA and (C) ConA. Labels are: am,
mesoderm and ectoderm component of amnion; Pe, parietal endoderm; ee extra-embryonic
endoderm; m, mesoderm; ne, neural ectodem.


48
Expression was also observed in the auditory semicircular canal, in the lens epithelium of
the eye and in the olfactory epitheiium. Although GlcNAc-TV transcripts appeared to be
widely expressed in the central nervous system, L-PH. staining of E15.5 and E17.5
embryos was restricted largely to the vascular endothelium of brain (data not shown).
GlcNAc-TV transcripts were observed in several endocrine organs at di fferent
stages of development (Figure 1). At EL 1.5, expression was observed in the Rathke's
pocket, the primordial structure for the pituitary gland, a pattern that persisted through
E17.5, when highest expression was &tected in the pars distalis of the pituitary gland In
contnst, GlcNAc-TV message was detected in the parathyroid, the adrenal gland and pineal
gland only later in organ development as observed in E17.5 sections. The most striking
example was the genesis of the parathyroid, which detaches h m the thyroid gland at E12,
with continued morphological development until E l5 when GlcNAc-TV message becomes
highly expressed. However, expression was not observed in the thyroid at any of the stages
examined. Embryonic thymus showed low GlcNAc-TV expression, although message has
previousl y been detected in adult thymus (Pemg et al., 1994). It is possible that GIcNAc-TV
expression is upregulated and required in thymocytes afier birth, as GlcNAc-TV enzyme
activi ty has k e n observed to increase following thymocyte activation (Lemaire et al., 1994).
In the gastrointestinal tract, GlcNAc-TV transcripts were observed in embryos at
E11.5, the earliest stages of intestinal development, persisting through to E17.5.
Furthemore, this pattern of expression was similar in E17.5 embryos and adult intestine
(Figure 3B,F). GlcNAc-TV was present in epithelial and Paneth cells at the base of the
crypts, and in the columnar epithelial cells in the lower half of the villi.

49
Figure 3: GlcNAc-TV and core 2 GlcNAc-T expression in adult and embryonic
mouse intestine.
Panels A-D are photomimgraphs of sections from E17.5, and panels E,F show adult
intestine. Light-fields (A) is paired with the dark field in situ (B), GlcNAc-TV; and @),
core 2 GlcNAc-T. Panel C shows L-PHA staining for GlcNAc-TV product in E17.5
intestine. Panel E is the light-field of a section through adult intestine and it is paired with
the dark-field of the sarne section shown in panel F. Labels are: vi. villi; cr, crypt cells; il,
intestinal lumen; sm, smooth muscle layer.


51
Transcripts were absent in lamina propria and in the smooth muscle layer located
immediately below the crypts- L-PHA lectin histochemistry showed reactivity in the
epithelium lining the villi, coincident with the location of GlcNAc-TV transcripts (Figure
3C). L-PHA staining extended to the top of the villi, presumably reflecting a longer half-life
of the N-linked oligosaccharides than the GlcNAc-TV transcripts in the epithelial cells
moving up the villi from the crypts.
GlcNAc-TV was highly expressed in the developing kidney beginning at E13.5, and
appeared to be limiteci to the epitheiial ceIls of Bowman's capsuies, a ceil layer which also
stained intensely with L-PHA (Figure 4).
In skin, GlcNAc-TV was expressed in the basal ce11 layer of the epidermis beginning
at €15.5 (Figure 5). The basal or pendemal layer of the epidermis is the outermost layer at
E14, the earliest stage of skin differentiation, and stratification begins on E l 6 5 At E15.5,
prior to stratification of the epidermis, GlcNAc-TV t randp t s and L-PHA reactivity were
observed in the outermost ceIl layer. Following stratification at E17.5, transcripts and L-
PHA reactivity were confined to the basal ce11 layer and absent in the comîfied layer.
GlcNAc-TV expression was also observed in the whisker follicles at E13.5 and was
maintained through E17.5 (Figure 6). Expression was restricted to cells of the external rmt
sheath, an involuted basal epithelium of the skin which also stained strongly with L-PHA
(data not shown).
Low levels of GlcNAc-TV message were observed in the nasal epithelium, in the
lining of the oral cavity and in the oesophagus and trachea at El 1 -5. By E15.5, prominent
expression was seen in the respiratory epithelium of the nasal cavity, and in the similarly
stmctured epithelium of the pharynx, the oesophagus, the trachea and the bmnchi (data not
shown). This pattern of expression persisted to E17.5 (Figure 1).

52 Senoli and granulosa cells are support cells in the adult testis and ovaries,
respectively, both of which expressed GlcNAc-TV message. Transcripts were observed in
pnulosa cells of superovulated ovaries at al1 stages of follicle maturation, and in the
epithelium of the oviduct (Figure 7). However, message was not detected in spermatozoa or
ova.
Core 2 GlcNAc-T expression in the post-hphntation mouse embryo: Core 2 GlcNAc-
T transcripts were highly expressed in embryonic and extraembryonic regions of the E7
embryo, as well as in the giant cells of the trophoblast and the materna1 deciduum (Figure
1). The pattern of expression became more restricted by E9.5. Between E9.5 and E11.5,
message was detected primari!y in tissues of mesodemal origin, such as the liver bud, the
visceral arches, the heart primordium and the somites. The expression pattern of core 2
GlcNAc-T in later stage embryos was associated with mucin- and cartilage- producing
tissues.
Core 2 GlcNAc-T was not expressed in the oral and nasal epithelium prior to El 1.5,
but were detected by E13.5 and persisted through E l 7 5 At E15.5 and E17.5, core 2
GlcNAc-T expression was also detected in the respiratory epithelium of the nasal cavity, the
trachea and the pleural sac (Figure 1).
At E9.5, core 2 GIcNAc-T transcripts were highly expressed in the mesenchyme of
the sornites and in the otic vesicle, regions which will later form cartilage. By E13.5, high
expression of core 2 GlcNAc-T message was also observed in the cartilage of the front and
hind paws, where cartilage synthesis is a preliminary step to bone fornation (data not
shown). At E17.5, transcripts were reduced in the paws correlating with reduced cartilage

53
Figure 4: GlcNAc-TV and core 2 GlcNAc-T expression in kidney at E17.5.
Light-field photornicrograph (A) is paired with dark-field of in situ for core 2 GlcNAc-T
(B), and GlcNAc-TV (C). Panel D shows L-PHA staining for GlcNAc-TV product which is
localized to Bowmann's capsule, (bc). Other labels are: dt, distal tubule; pt, proximal tubule;
gl, glomerulus.

Figure 5: GlcNAc- TV expression in murine skin.
(A), Light-field photomimgraph at E17.5; and corresponding dark-fields of (B), in sihr for
GlcNAc-TV transcript and (C), L-PHA staining. O), shows in situ for GlcNAc-TV
transcript at E15.5. The peridermal layer, which expresses GIcNAc-TV at E15.5 becomes
the basal ce11 layer by day 17.5. Labels are: bc, basal cells; cl, cornifieci layer, pd,
peridermal layer, dm, derrnis.


57
Figure 6: GlcNAc-TV and Core 2GlcNAc-T are not expresseci in the same cell
populations.
Bright- and corresponding darkfield photornicrographs showing GlcNAc-TV, (AB); and
core 2 GlcNAc-T, (CD) transçript expression in whisker follicles, and GIcNAc-TV in lung
(EJ;) at E17.5. Labels are: irs, inner mot sheath; ers, extemal mot sheath; cts, connective
tissue sheath; rb, respiratory bronchioles.


59 Figure 7: GlcNAc-TV and core 2 GlcNAc-T expression in fernale reproductive
organs.
In sitri locdization of GlcNAc-TV (BE) and core 2 GlcNAc-T (CS;) transcripts in
superovulated ovary (A-C) and oviduct @-F). Labels are: fo, follicles; am, amptulla; is,
isthmus.


61
production and higher bone formation at this developmental stage. Other cartilage
producing tissues also expressed core 2 GIcNAc-T transcripts at E17.5, including the nasal
passages, trachea, invertebrate disks and Meckel's cartilage of the lower jaw (Figure 1). Core
2 GlcNAc-T transcripts were observed in the proximal tubules, the mucin producing
compartrnents of the kidney (Figure 4B). The distal tubules and Bowman's capsule produce
less mucin and these regions showed very little message. In the intestine, expression was
observed in the epithelial cells of the villi at El1.5, and appeared to increased by E l 3 5
Expression persisted through E17.5 and in the adult intestine @igue 3D).
In whisker follicles, core 2 GlcNAc-T message was observed in the inner root
sheath, a ce11 layer close to the lumen which becomes occupied by a haïr at E13.5 through
E17.5 Figure 6CD). This contrasts with GlcNAc-TV transcripts which were restncted to
cells of the extemai mot sheath, a ce11 iayer contiguous with the basai ce11 layer of skin.
Core 2 GlcNAc-T message was not detected in the ovaxîes of superovulated females.
However, the isthmus of the oviducts showed high levels of expression, again correlating
with a region of high mucin production (Figure 7). In the adult testis, core 2 GlcNAc-T
transcripts were detected in the male germ cells at al1 stages of sperrnatogenesis (data not
shown).

62
DISCUSSION
In this study, we have examined the distribution of GlcNAc-TV and core 2 GlcNAc-
T transcripts in p s t implantation mouse embryos by RNA in situ hybridization. Core 2
GlcNAc-T message was expressed widely at E7, considerably earlier than GlcNAc-TV
transcripts, which were observed at E9.5 in al1 embryonic and extraembryo~c ce11 layers.
Retinoic acid-induced differentiation of F9 cells into primitive endoderm is believed to
reflect a similar developmental period and is accompanied by the induction of several
glycosyltransferases (Heffeman et al., 1993). In this system, core 2 GlcNAc-T shows a
larger and more rapid induction following retinoic acid treatrnent than does GlcNAc-TV
(Heffeman et al., 1993), a developmental sequence which may be analogous to the earlier
appearance of core 2 GlcNAc-T as compared to GlcNAc-TV during embryogenesis. These
observations may reflect a general temporai ciifference in the induction of enzymes in the 0-
linked and N-linked pathways. Core 2 GlcNAc-T expression becornes restncted to specific
tissues by E9.5, and similarly, a restricted expression pattern was observeci for GlcNAc-TV
by El 1.5. However, patterns of tissue-specific expression were distinctly different for the
two genes (Figure 8).
Core 2 GlcNAc-T expression was observed in epithelial cells of most, but not dl,
rnucin producing organs, where O-glycosylation is expected to be high. Core 2 GlcNAc-T
expression overlapped with a subset of tissues that express the MUCs, with highest levels in
the proximal tubules of the kidney, the pancreas, respiratory tract and small intestine (Braga
et al., 1992). Core 2 GlcNAc-T transcripts were also observed in the pericardium, the
visceral pleura and around the vertebrae, where lubricative and synovial fiuid secretions
occur. In the adult oviduct, core 2 GlcNAc-T transcripts were highest in the isthrnus where
mucins are also highly expressed. Core 2-GlcNAc-T transcripts were also found in regions

Figure 8: Summary of the expression patterns.
The temporal and spatial expression pattern of GlcNAc-TV (open box) and core 2 GlcNAc-
T (closed box) in the p s t implantation mouse embryo.

Brain ventricular
medial, marginal
bndocnne p i t u i t q
adrenal
pineal
Intestine villi
PtS
Kidney Proximal tubules
Bowman's capsule
Skin
Pancreas
Respiratory epithelium
Cartilage

65 of cartilage production, suggesting a possible role in bone formation. Core 2 GlcNAc-T
expression was not detected in embryonic thymus but the message is present in the cortex of
postnatal adult thymus (Baum et al., 1995). Core 2 GLcNAc-T activity has also been shown
to increase following activation of human T cells in tissue culture (Higgins et al., 1991;
Piller et al., 1988), and may therefore be associated with later stages of T ce11 maturation.
In the early pst-implantation embryo, GlcNAc-TV transcripts show similar
temporal regulation as that reported for GlcNAc-Ti. with very low levels at E7, followed by
upregulation at E9.5. However, GlcNAc-TI remains widely expressed in al1 tissues pst
E9.5 (unpublished data), while GlcNAc-TV becomes progressively resuicted to speci fic cell
layen (Campbell et al., 1995). Lectin histochemistry showed that E-PHA reactive complex-
type gl ycans were present in the E7 day embryo, but L - P H . reacti ve P 1,6GlcNAc-branched
structures were relatively low. These observations suggest that the oligosaccharide
structures are either not necessary, or matemal sources of GIcNAc-TV and GlcNAc-TI
and/or glycoconjugates are sufficient for gastrulation and implantation. In the sea urchin
embryo, gastmlation occurs in the presence of N-linked processing inhibitors 1-
deox ynojirimycin and 1 -deoxymannojirimycin but subsequentl y, spicule formation is
blocked suggesting a need for cornplex-type oligosaccharides at this latter stage of
development (Kabakoff and Lennarz, 1990).
E9.5 marks the beginning of rapid embryo growth with a 330x increase in weight by
El75 Maternal sources of message and proteins are expected to be depleted by dilution
during this phase, and therefore autonomous expression of genes becomes essential for rapid
growth and continued organogenesis. The induction of GlcNAc-TI, as well as GlcNAc-TV,
core 2 GlcNAc-T, P 1,3Gal-T, p 1-4Gat-T and GlcNAc-T(i) activities observed in retinoic
acid treated F9 cells suggests that a general increase in glycoprotein gIycosy1ation occurs
following E7, pssibty to accomodate the requirements for increased extracellular matrix

66 and connecti ve tissue gl ycoproteins. In this regard, GlcNAc-TI nul l mutant embryos, which
are defective in neural tube closure and vascularization, show evidence of abnorrnal
development only afier E85E9.5, coincident with the advent of GlcNAc-TI and GlcNAc-
TV expression in wild-type embryos (Joffe and Stanley, 1994; Metzler et al., 1994).
Because the product of GIcNAc-TI is required in the synthesis of acceptors for GlcNAc-TII,
Tl[V and TV enzymes (Schachter, 1986), its absence results in a complete loss of complex-
type N-linked oligosaccharides. Thus, it cannot yet be detemiined which defects are
attributable to the failure to form subsets of structures downstrearn of GlcNAc-TI. Patients
with carbohydrate deficiency syndrome type II, a rare human congenital defect in GlcNAc-
TII, show severe multi-organ defects (Jaeken et al., 1994). A point mutation in the coding
sequence of the GlcNAc-TII gene from two patients has recently been identified (Schachter
et al. personal communication). Further studies with GlcNAc-T gene "knockout" mice will
be required to dissect the nul1 phenotypes of GlcNAc-TII and TV, speçificaily.
As organogenesis proceeds at E9.5 and thereafter, GlcNAc-TV transcript in non-
neural tissues becomes progressively restricted to specialized epithetia. The cornpiex
pattern of expression of GlcNAc-TV, and its potential to act on many target glycoproteins,
suggests that the gene product rnight be required in a variety of biological functions. A
subset of cells which express GIcNAc-TV share the common features of basal ce11 layers
such as their stem ce11 properties, and attachment to basal lamina. These include basal
epithelia of the intestine, skin, lung, and Bowman's capsules in kidney, as well as the Sertoli
cells of the testis and the granulosa cells of the ovary. The skin, and gastrointestinal tract
rapidly repair denuded epithelium by migration and proliferation of surrounding epithelial
cells to cover exposed basement membrane. In the small intestine, this involves
proliferation and upward migration of epithetial cells from the lower half of the villus
followed by terminal differentiation as the cells continue to ascend to the top portion of the

67 villus. Although GIcNAc-TV transcripts were absent in the upper portion of the villus, L-
PHA staining indicated that the cells retain the oligosaccharide structures forrned by the
enzyme until the ce11 are shed.
In normal basal epithelium, ce11 proliferation and migration is suppressed, but can be
rapidly induced in response to cues such as tissue damage. However, the growth and
motility of transformeci cells are not well controlled by their tissue environment.
Transformed fibroblasts and epithelial ce11 have k n shown to have elevated GlcNAc-TV
activity (Dennis et al., 1987; Lu and Chaney, 1993; Pierce and Arango, 1986; Yarnashita et
al., 1985) which may contribute to ce11 growth and motility. indeai, transfection of MvlLu
epithelial cells with a GlcNAc-TV expression vector resulted in relaxation of growth
controls and increased ce11 motility (Demetriou et al., 1995). Trophoblasts, an invasive ce11
in the E7 embryo also express high levels of both GlcNAc-TV transcripts and L-PHA
reactive oIigosaccharides~ Furthemore, swainsonine, an a-mannosidase II inhibitor that
blocks N-linked processing pathway pnor to GlcNAc-TV, has been shown to inhibit both
tumor ce11 and first trimester human trophoblast cells invasion in vitro (Yagel et al., 1989;
Yagel et d., 1990).
A more recent survey of nomal human and rat tissues detected L-PHA reactive
glycans in most non-mitotic and non-migrating epiuielial ce11 types, with the notable
exception of colonic and mammary gland epithelia (Li and Roth, 1997). Therefore, it is
possible that in addition to the suggested role for B1.6GlcNAc-branched N-glycans in aiding
ce1 1 rnigrati on, these gl ycans have additional functions, not readil y apparent from their
location. For example, our RNA in siîu results indicate that GlcNAc-TV activity may also
be necessary for ei ther the formation of secretory epithelia of some endocrine glands, or the
proper glycosylation of the hormones that these glands produce. These include the secretory
epithelia of the pituitary gland, the parathyroid, the pineai gland and the adrenai gland.

68 GlcNAc-TV transcript levels appeared to be highest in embryonic neuroepithelium
of the bnin and spinal cord from E13.5 to El75 Expression was first observed at E9.5 to
E10.5 in the ventricular zone. In Iater stage embryos (ie. E11.5-17.5), transcripts were
observed in the intennediate and marginal zones, where p s t mitotic neurons and glial cells
are ac ti vel y migrating outw ard to populace the cortical layers. Therefore, GlcNAc-TV
niTU\IA expression correlates with regions of active ce11 migration and neurite outgrowth in
the developing CNS. Cerebellar organization also involves selective cell death by apoptosis
which may determine the correct size of sensory neural areas and that of its targets.
Interestingly, MvlLu epitheiiai cells transfected with a GlcNAc-N expression vector also
show accelerated rates of cell death in serurn-free medium (Demetriou et al., 1995), sirnilar
to pnmary mouse f i broblasts partial1 y-trans forrned by a single oncogene (eg. H-ras) (Tanaka
et al., 1994).
Our RNA in sim hybridization and L-PHA histochemistry, as well as previously
reported Northern and enzyme activity analysis (Dennis and Laferte, 1989; Pemg et al.,
1994) suggest that the ratio of GlcNAc-TV transaipts to enzyme activity in brain is
considerably greater than in other GlcNAc-TV expressing tissues such as the intestine and
kidney. It is possible that GlcNAc-TV enzyme activity is regulated by pst-transcriptional
or post-translational mechanism(s) that suppress activity in the centrai nervous system. For
example, p l4Gal-T activity during testicular development appears to be regulated by
altemate splicing of 5' untranslated sequences that affect translation efficiency. In addition,
p 1-4Gal-T (Bunneil et ai., 1990) and core 2 GlcNAc-T @atti and Dennis, 1993) may also
be regulated by protein phosphorylation. The brain GlcNAc-TV transcript is 7.5 kb, similar
in size to that of other tissues. However, the GlcNAc-TV gene has multiple exons @ennis
et al. unpublished &ta), raising the possibility that transcripts might undergo tissue-specific
alternative splicing to produce different protein products, as has b e n observai for a2-6SA-

69 T (Svensson et al., 1990). Secondly, the predicted protein sequence is 740 amino acids,
almost mice the length of the other GlcNAc-Ts suggesting that the enzyme may have
additional functional domains not involved in glycosyltransfer.

MATERIALS AND METHODS
Animals: CD1 female mice wefe mated overnight, checked for vaginal plugs the following
moming and gestation time counted from the preceding night. Super-ovulation was
achieved by intraperitoneal injection of 5 IU of pregnant mare semm gonadotropin,
followed 48h later by an injection of 5 IU human chorionic gonadotropin (hCG).
In situ RNA Analysk cDNA encoding mouse GlcNAc-TV (Shoreibah et al.. 1993) and
mouse core 2 GlcNAc-T (Warren et al. in preparation, accession #U19265), which is
highIy homologous to the human gene (Bierhuizen and Fukuda, I992), were used to
prepare the anti-sense RNA probes. The 2384bp HinDm: Bglil fragment of pcDlnGnT-
V3 was subcloned into HinDIiI: BamHI cut pBluescript KS(+). This plasmid was
linearized by HinDm digestion and ami-sense probes generated by in vitro transcription
with T7 RNA polyrnerase incorporating [ y - 3 5 ~ ] ~ ~ E ? The 1715bp XhoI fngrnent of
pmC2-25 1 was subcloned into Sa11 cut pGEM-52f(+). This construct was linearized by
NotI digestion and antisense probe synthesized by in vitro transcription with SP6 RNA
polymerase incorporating [ y - 3 5 ~ ] ~ ~ ~ .
Ir1 sirzi RNA hybridization was carried out on 7 Pm cryostat sections, as previously
descri bed (Peter et al., 1990). Adjacent sections were probed with antisense GIcNAc-TV
and core 2 GlcNAc-T, and as a control with sense probes. Post-hybridization washes
included treatment with M pglml N a s e A at 370~ or 4 2 ' ~ for 30 minutes, and 2 stringent
washes of 20 minutes each at 600C in 0.1X standard saline citrate (15 mM NaCI, 1.5 rnM
sodium citrate pH 7). The slides were dipped into Kodak NTB-2 emulsion, exposed for 7
days, developed and stained with toluidine blue.
Lectin hiktochemisîry: Cryostat sections of 7 pn in thickness were blocked with 1 %
BSA in PBS at 20°C for 30 min. FITC-conjugate L-PHA at 20 &ml was overlayed ont0

71
the section, and incubated for 30 min at 37°C in a humidified chamber. The slides were
then washed twice with PBS, 1% BSA for 10 min each at room temperature.
Fluorescence was observed and photographed using a Leica Leitz DM RXE microscope.
In Figure 2, cryostat sections were stained with biotinylated E - P H . and L - P H .
(Vector), postfixed for 30 min in 3.7% formddehyde. The slides were washed in PBS,
rinsed in methanol and incubated with 0.3% WO, in methanol for 20 min. The slides were
blocked with 1% BSA in PBS and then incubated with the lectins at 10 uglml in the same
buffer ovemight at room temp. M e r three washes in PBS, the color was developed with a
solution of 1 m g / d of diaminobenzidine, 0 .03% --O2. for 10 min, after which slides were
was hed in water, mounted under Gel-mount (Biomeda) and photographed (Campbell et al.,
1995).

CHAPTER 3: Generation and Characterization of
GlcNAc-TV Deficient Mice

INTRODUCTION
Tissue-specific patterns of glycopmtein glycosylation (Childs et ai., 1983; Feizi,
1985; Hakomori and Kannagi, 1983; Pennington et ai., 1985; Solter and Knowles, 1978),
and more specifically GlcNAc-TV-dependent glycosylation (Chapter 2) are documented,
but the functions of N-glycans in development and disease are poorly understood
(Hakomori, 1989). Studies on somatic tumor ce11 mutants have causally linked the
GlcNAc-TV activity and its products, B1,oGlcN~c-branched N-glycans with increased
metastatic potential (Yagel et aI., 1989; Yousefi et al., 1991; Myc et al., 1989). The
1,oGlcNAc branch of N-glycans is the enzyrnatically preferred acceptor site for
polylactosarnine chah addition and enhanced expression of polylactosamine sequences
on the ce11 surface is suggested to reduce ce11 adhesion to substratum and enhance organ
colonization by tumor ceils.
Lymphocyte adhesion may be similarl y affected by f3 1,oGlcNAc-branched N-
glycans, with consequences for immune ceIl maturation, activation and migration.
Indeed, lymphoid ce11 functions are known to be sensitive to changes in glycosylation
(Baum et al., 1995; Jiang et al., 1995; Lasky et al., 1989). A rare human congenital
syndrome, LADIl (Leukocyte adhesion deficiency U) caused by a defect in the synthesis
of GDP-fucose, is clinically characterized by immunodeficiency and impaired wound
healing (Phillips et ai., 1995). Mice deficient in al,3Fuc-TVII, which is required for the
spthesis of Slex antigen in neutrophils and lymphocytes (Maly et al., 1996), have
elevated penpheral leukocyte counts and are deficient in leukocyte extravasation into
areas of inflammation (Maly et al., 1996). Gal:a2,6 sialyttransferase (ST6Gal)
synthesizes SAa2,6GaIp1,4 sequences recognized by CD22 on the surface of B cells

74
(Powell et al., 1993). Mice lacking this enzyme are severely immunosuppressed and
display reduced serum IgM levels, impaired B ce11 proliferation and attenuated antibody
production (Hennet et al., 1998). Transgenic mice expressing the O-glycan core 2
GIcNAc-T under the regulation of the lymphocyte-specific Lck promoter, show reduced
T ce11 activation (Tsuboi and Fukuda, 1997).
To examine the function of GlcNAc-TV dependent glycosylation in murine
biology, mice deficient in this enzyme were generated by targeted mutation of the Mgat.5
locus. This c hapter sumarizes the phenotypic anal ysis of GlcNAc-TV &ficient mice.
Homozygous ~ ~ a t . Y ' - mice were viable, fertile and lacked pl,6GlcNAc-branched N-
glycans. Although these mice showed no gross abnonnalities, detailed histological
examination revealed lymphocytic infiltrates in liver and kidney of ~ ~ a t 5 ' - mice, which
were not present in wild type littermate and cage-mate animais.
GlcNAc-TV deficient mice dispiayed enhanced hypersensitivity to skin irritants,
but the onset of the inflamrnatory response was slower. This was likely due to a reduced
rate of leukocyte migration in vivo, which was associated with increased adhesion to
substratum, measured in vitro. Sensi tivity and cooperati vi ty of T ce11 receptor (TCR)-
dependent activation, as well as the rate of receptor intemalization and actin
reorganization, were increased in GlcNAc-TV deficient T cells. We also demonstrated
that MgarS gene expression and P1,6GlcNAc-branched N-glycans increase with T cell
activation. Therefore, levels of p1,6GlcNAc-branched N-glycans are regulated by the
status of T cell activation. Consequently, our results suggest that B 16GlcN~c-branched
N-glycans have a direct role as a negative regulator of T ceIl response to antigen.

75
RESULTS
Generufïion of Mg& mice: The Mgat5 targeting vector was designed to replace the
coding portion of the first coding exon of Mgat5 with the lac2 reporter gene (Figure
lA,B). GlcNAc-TV enzyme activity was approximately 50% in heterozygous mice, and
below the level of detection in ~ ~ a t s / - mice (Figure lC,D). The products of GlcNAc-
TV, f! 1,oGlcNAc-branched N-gl ycans, were also absent in the organs of ~ ~ a t 5 - I - mice, as
determined b y L-PHA lectin biotting (Figure 1 E). L-PHA binds to tri' and tetraantennary
N-glycans, the synthesis of which is dependent on a substitution by GIcNAc-TV.
As expected, the Lac2 expression pattern closely matched Mgut5 expression, which
we have previously characterized by RNA in situ hybridization (Figure 2, Figure
3A)(Granovsky et al., 1995). Lac2 expression was observed in the giant cells of the
trophoblast at E7.5 (Figure 2A). At E13.5, lac2 expression was observed in the crypts of
the developing gut (Figure 2B), the media1 and marginal zones of the developing braîn
(Figure 2C), the spinal chord and the skin (Figure 2D). Lac2 staining was specific to
neuraI cells in newborn and adult brain, and corresponded with L-PHA staining of neural
axonal trees in ~ ~ a t 9 ' ' CNS (Figure 2E,F). In adult kidney, staining was observed in
the podocytes of the glomerulli (Bowman's capsuie), corresponding to the previously
observed Mgar5 transcripts in this tissue. As expected, LacZ expression was significantly
suonger in ~ ~ a t 5 - I - embryos than in ~ ~ a t S ' - embryos.
~ ~ a t 5 ' - mice were fertile and showed no oven abnonnalities. However,
histological and biochemical examination of tissues revealed several subtle abnormalities.

76
Figure 1: Generation and characterization of GlcNAc-TV deficient mice.
(A) Strategy for the inactivation of the MgatS gene. The structure of the wild type Mgai5
locus, the targeting vector, and the resulting targeted locus are illustrated.
Nucleotides -22 to 241 of the coding portion of the first cating exon were replaced with
lacZ and a neomycin-resistance gene; filled region represents the exon, 5' extemal probe
is indicated by a solid bar above the Mgut5 genomic locus. P , Pst1 restriction site. (B)
Southem blot analysis of genomic DNA of F2 offsping derived from heterozygous
crosses. PstI-digested DNA was hybridized with the 5' extemal probe. Lanes 2 and 4
show the ~ ~ a t 5 ' - genotype of F2 mice generated from two independently targeted ES
clones. (C) GlcNAc-TV enzyme activity measured in ~ ~ a t 5 + / + and ~ ~ a t S / - tissue
homogenates. Sarnples were assayed in duplicate and the assays were repeated on tissues
from mice of both lines. @) Time course of GIcNAc-TV enzyme activity in small
intestine from ad+/+, ~gar .5+ / - and ~ g a t 5 ' - mice. Results represent duplicate assays.
(E) L-PHA reactive glycoproteins in homogenates of 1, intestine; K, kidney; H, heart; B,
brain; S, spleen; Lu, lung and Li, Iiver separated by SDS-PAGE.

A Probe -
Mg& Genomic locus
Targeting construct
P * I - , m i - [ neo P )P %.6 w
d kidney brain spleen liver O 50 100 150 200 250 300
Time (min)

78
Figure 2: Lac2 expression from the mutant M g 6 allele is identical to the
Mgai5 expression pattern.
Lac2 expression pattern in murine embryos at E7.5 and at E13.5, and lac2 expression
and L-PHA reactivity in adult cerebellum. Lac2 expression was visualized by $-
galactosidase cleavage of x-gai, which ~tains cells blue. (A) E7.5 embryo. The embryo
proper does not express l a d , but the giant cells of the trophoblast show strong
expression. E 13.5 ~ ~ a t . 5 ' - embryos were stained for b-galactosidase activity (B-D). (B)
E13.5 gut, staining in the crypts. (C) E13.5 rnid-brain, where lac2 expression is seen in
the media1 and marginal zones. @) E. 13.5 spinal cord and dermis express lacZ. (E) Lac2
expression is observed in the Purkinje cells, a neural ce11 type in the adult cerebelum. (F)
The lac2 expression in the hrkinje cells is matched by F[TC-L-PHA reactivity of the
dendritic trees of these cells, staining green. e, embryo; t, trophoblast; v, ventricle; m,
medial and marginal zone of the 3" vemtricle; d, dermis; s, spinal chord; p, Purkinjee
cells; dp, dendritic trees of Purkinjee cells.


Figure 3: Aberrant histology in M ~ ~ S / - mouse tissues.
(A,B,C,) Photomicrographs of kidney from 9 months old mice. (A) Lac2 staining of
M ~ U ~ S " kidney section, showing intense staining in podocytes, the filtering cells of the
glomerulus. (B) ~ ~ a t y - glomerulli. (C) ~ ~ a t S ' + glomerulli. ~ ~ a t 5 " glomerulli show
a reduction of 47% in nuclei when normalized to the area of the glomerulus. (D)
Lymphocytic infiltrates, indicated by the arrow, in the liver of a 3 months old ~ ~ a r S ' '
mouse. (E) Lymphocytic infiltrates. indicated by the arrow. in the kidney of a ~ ~ a t S ' -
mouse.


Table 1: Summnry of the phenotypic aberrations observed in ~ ~ a r t s ' ~ mice.
Aff ected system
Neurologicai
Kidney
Lymphoid
Phenotype
15 1 18 ~ ~ o t 5 ' - females fail to nurse their
young and are aggressive towards them
(4 1 3 1 ~ ~ a t 9 ' - and 1 1 5 ~ ~ a t F " + females
exhibited similar behavior).
Glomeruli have 28*7% fewer celldarea in
3 months old mice, and 47115% fewer
cellslarea in 9 months old rnice
- - -
Lymphocytic infiltrates in solid organs.

83
The cellularïty of kidney glomerulli was reduced by 28% in 3 months old ~ g a t S / -
animals, and by 47% in 9 monihs old ~ ~ a r . 5 ' ' animais, as compared to wild type
Iittermates (Figure 3A-C). In addition, a strain dependent behavioral abnormality was
observed in ~ ~ o t 5 - I - nursing fernales. 129/sv ~ ~ a r y - females (15 / 18) behaved in a
negligent or agressive manner towards their pups, fading to nurse and cannibalizing
them. Only 4 1 3 1 ~ g a t 5 " - and 1 1 5 ~ g a t 5 * ' + 129/sv females behaved similarly (results
summarized in Table 1). Laçk of nurturing behavior was not apparent on rnixed 1291s~-
CD 1 or 1 W s v - C57BU6 strain bac kground.
Lyrnphocytic foci were detected by histology in the liver and kidney at 3 months
of age, but wasting o r mortality due to autoimmune disease was not observed (Figure
3D,E). PeripheraI white ce11 and erythrocyte counts were normal, and populations of T
and B celis in spleen, thymus, and lymph nodes were also in the normal range by FACS
anal ysis.
GIcNAc- TV defciency causes hyperinftammatory response: The 1 ymp hoc yte in fi 1 trates
in Mgat5'- mice suggest they may respond aberrantly to immune stimulants. To examine
T ce11 dependent responses in vivo, a type N (T ce11 dependent) delayed-type
hypersensitivity @TH) reaction was induced in mutant and ~ ~ a t F ' + mice. The chemical
antigen oxazolone was applied topically to the backs of the mice, then again 4 days later
to the right ear (Correll e t al., 1997). Ear swelling in ~ ~ a t Y " + rnice peaked 24 h after
application of oxazolone to the ear, and returned to normal by day 5. Ear swelling in
Mgar5" mice attained a higher maximum between 48 and 72 h, and persisted until day 5
(Figure 4A).

Figure 4: Inflammation in ~ ~ a f s / * and Uttermate coatrols.
(A) DTH response in ~ ~ a t 5 ' - and Mgat5+'+ mice exposed to oxazolone fint on their
back, then 4 days later on the nght ear. The results are plotted as mean change + S.E. in
ear thickness relative to the vehicle-treated lefi ear for 7 ~ g a t 5 ' - rnice and 6 Mgat5+/+
littermate controls (Irdenotes p 4 . 0 1 ) . (B) Ear swelling induced by topicai application
of arachidonic acid in ~ g o t 5 ' - and Mgat5+'+ mice. The results are the average of five
mice per genotype. One month later, when swelling had subsided completely, the
experiment was repeated with the same rnice, reversing the control and experimental ears
on each mouse. The results were identical (*denotes p4.001) .

Time (hrs)
Time (hrs)

86 A single application of arachidonic acid, which is converted to leukotriene B4, a
potent chernotactic agent for infiltrating neutrophils, macrophages and T lymphocytes,
was used as a mode1 of immediate skin hypersensitivity (Jiang e t al., 1998). As shown in
Figure 4B, ear swelling induced by arachidonic acid was also significantly greater and
more persistent in ~ ~ a t . 5 ' mice. Maximum swelling was observed 2 h post application in
the ~ ~ a t 9 ' ' mice but was delayed until7 h in the Mgats/- mice.
Cell migrdion is slowed and dhesion tu fibromeetin is enhanced in ~~at5 ' - mieor The
delayed onset of inflammation observed in ~ g a t s / - mice rnay be due to a glycosylation-
dependent change in leukocyte adhesion. To examine this possibility, the adjuvant
thioglycollate was injected i.p. and elicited peritoneal leukocytes were counted 3. 7 and
30 hrs later (Maly et al., 1996). Leukocyte infiltration was reduced at al1 time points in
the ~ ~ a r 5 ' - mice (Figure SA). Indeed, at 30 hm, the inflammatory response was
dampened to basal levels in MgatS'' mice but was still rising in ~ ~ a t p + and ~ ~ a t 9 ' -
mice. Fuc-TVII deficient rnice lack selectin ligand and show a similar impairment in
leukocyte extravasation (Maly e t al., 1996). Leukocytes accumulate in blood of Fuc-
TVII nul1 mice due to impaired selectin-mediated attachment to endothelium, while
leukocyte accumulation in ~ ~ a t 5 ' - mice occurs in solid tissues and may be due to altered
cell-substratum adhesion. Indeed, the thioglycollate-elicited leukocytes from M@F"
mice attached more avidly to fibronectin coated plastic, as did activated T cells (Figure
5B,C).
GlcNAc-TV activity may regulafe integrin signaiingr Leukocyte adhesion to fibronectin
is mediated primarily by integrins VLA4 ( a 4 g l ) and VLAS (or5$l), and the latter is
known to modified with ~1,6GlcNAc-branched glycans (Demetriou e t al., 1995).

87
Figure 5: Cell migration is slowed and adhesion to fibronectin is enhanced in
(A) Leukocyte motility into the peritoneal cavity of mice injected i.p. with
thioglycollate. Ce11 counts represent the average I ME of results for three mice per
genotype. (B) Thioglycollate-activated leukocytes from ~ g a & * and ~ g a 2 . Y ' mice were
seeded on various concentrations of fibronectin. The results represent the average + SD
of duplicate wells, and the experiment was repeated twice. (C) Adhesion of leukocytes
and activated T cells to fibronectin-coated plastic. Neutrophils were activated by
thioglycollate, and purified splenic T cells were activated with 40 ng/ml of PMA. Cells
from 3 rnice per genotype were assayed for adhesion, and the expenments were done
twice.
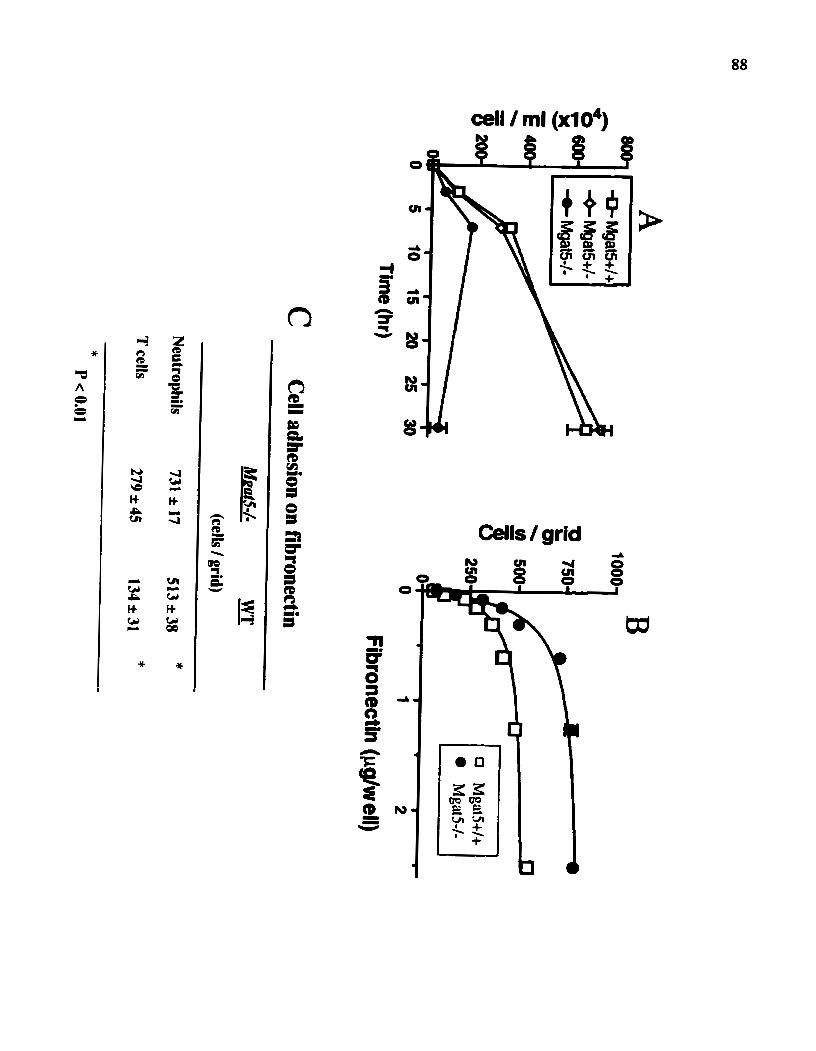

89
Ce1 1 moti lity is dependent upon formation and turnover of integrin-mediated focal
adhesions (Paiecek et al., f 997). These structures can be readily exarnined in embryonic
fi broblasts plated on fibronectin. ~ ~ a t ? ' - fi broblasts spread more and pseudopodia
showed fine actin microfilament, while ~ g a t 5 ' - cells had coarse actin stress fibers
characteristic of non-motile cells (Figure 6A.B). The distribution of the cytosolic protein
paxillin, which binds pl integin and the signaling molecules focal adhesion kinase
(FAK), Csk and ppoOSrc at discrete sites of ce11 attachment in spreading and motile cells
(Schaller et al.. 1995), was also different in ~ ~ ~ t . 9 ' - and ~ ~ a t Z / - cells. In ~ ~ u t p - cells.
paxillin localized to the distal ends of microfilaments characteristic of focal adhesions,
but the protein was dispersed in the cytoplasm of ~ ~ a t 3 ' - fibroblasts. Thus, GlcNAc-TV-
dependent glycosylation may enhance focal adhesion formation in vitro, and may account
for the observed motility of Mgat5+" leukocytes in vivo.
~ ~ a & ~ T-splenocytes are hypersensin've to T cell rnitogenc The exaggerated and
persistent DTH response in Mgat.5" mice was reflected in the behavior of splenocytes in
vitro. Spleen cells were stimulated with mitogens applied in solution. Mïtogens used
were the lectins leukoagglutinin (L-PHA) and Concanavalin A (ConA), and soluble
monoclonal antibodies reactive to CD3, clone 2Cl l (anti-CD3), and to TCRdP, clone
H59.72 (anti-TCR). Following stimulation with ConA, anti-CD3 or anti-TCR, T ce11
proliferation was 3-5 fold greater for spleen cells from ~ ~ a t ~ ' - compared to ~ ~ a t F ' +
li ttermates (Figure 7A-C). In contrast, ~ ~ a t . 5 ' - splenoc ytes were completely
unresponsive to L-PHA (Figure 7D). Thus, these results demonstrate that T ce11
activation by L-PHA requires the presence of surface Pl$GlcNAc-branched N-glycans.

90
Figure 6: Focai adhesions in embryonic fibroblasts plated on fibronectin.
~ p d " (A,C) and ~ ~ a t 5 ' (B,D) fibroblasts were stained with rhodamine-phalloidin,
FITC conjugated anti-paxillin anti bodies, and Hoec hst 33258 stain. Fluorescence images of
the celIs were obtained using a deconvolution microscope and digital data capture. Paxillin
(green) i s locdized to ends of microfilaments ( r d ) in focal adhesions of ~ ~ a t ' i + ' - , but not
in ~ ~ a t 5 ' - fibroblasts. ~ ~ a r 9 ' - fibroblasts spread to cover a greater surface area than
M ~ Q Z Z ~ cells, and the nucleus in panel A is out of view to the left. Panels A and B are at
1ûûX magnification, and panels C and D are at 40X magnification.


92 Figure 7: ~ ~ a t S / - T spienocytes are hypersensitive to T ceU mitogem.
~ ~ a t 5 ' - (a) and ~ ~ a t 5 " ' (a) spleen cells were cultured at 2 x 1 6 cells/ml and
stimulated with increasing concentrations of (A) anti-TC&p (B) anti-CD3 (C) Con A
@) L-PHA (E) PMA (10 ng/ml) plus ionomycin (OSpg/ml) and (F) LPS. Data in al1
panels represent the average k SD of triplicate measurements and experiments were
repeated 3 times. The experiments shown in panel C and D were repeated with rnke
generated from the second targeted ES ce11 line, and with the MgarS mutation on the
mixed 129/sv-CD 1 and 129/sv-C57black6 background with identical results.
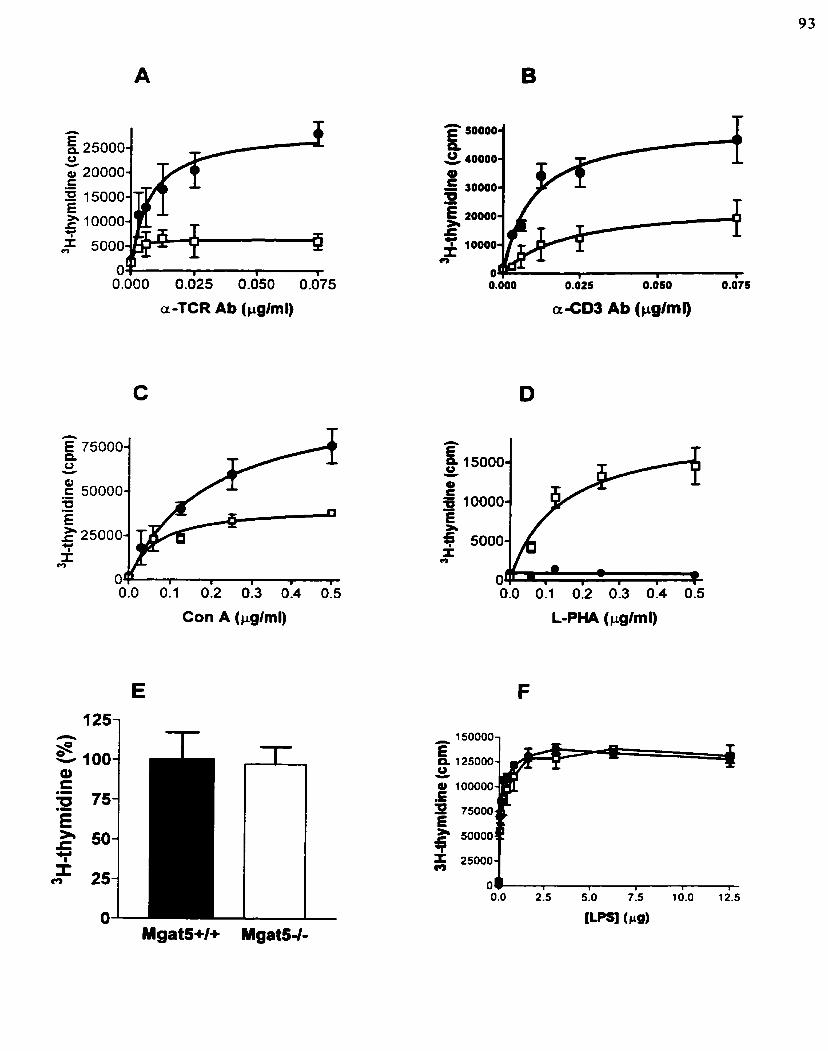
Con A (pglml)
0.0 0.1 0.2 0.3 0.4 0.5
L-PHA (pglml)

94
~ ~ a t y * and ~ ~ a t ~ ' - spleen cultures treated with phorbol 12-myristate 13-acetate
(PU4) and the Ca* ionophore, ionomycin, showed similar levels of proliferation
indicating that intracellular signaling downstrearn of these mediators is normal in ~ ~ a t Z ' -
cells (Figure 7E). The ~ ~ a t ~ ' - phenotype was cell-type specific, as B cell responses to
lipopolysaccharide LPS (Figure 7F) and to anti-IgM or anti-CD40 antibodies, in the
presence or absence of interleukin-4 (data not shown), were similar in mutant and wild
type littermates.
GlcNAc-TV deficiency is synergistic wi#h CD28 cu-receptor stimulrrtinn: CD28 co-
receptors on T cells stabilize TCR-mediated signaling aggregates and reduce the number
of engaged TCR required to propagate intracellular signals and initiate cell proliferation
(Bachmann et al., 1997; Wulfing and Davis, 1998a; Wulfing and Davis, 1998b). Mgat.5
dependent glycosylation of T ce11 surface glycoproteins may aiso affect the kinetics of
agonist-induced TCR aggregation and signaling. To examine this possibility, purified T
celis were cultured at low density and stimulated with increrising concentrations of
soluble anti-CD3 in the presence or absence of anti-CD28 antibodies (anti-CD28). Both
the GlcNAc-TV deficiency and anti-CD28 reduced the requirements for TCR agonist as
indicated by Dso values (Figure 8A,B). Furthemore, the apparent Hill coefficient (nH), a
measure of cooperativity, was increased both by the GlcNAc-TV deficiency and by the
addition of anti-CD28. The Hi11 coefficient for the TCR dependent activation of wild-
type T cells in the absence of CO-receptor stimulation was 6. GlcNAc-TV deficiency
increased the apparent cooperativity by 5.5 fold, and this effect was synergistic with that
of CD28 CO-receptor engagement (Figure SB). Cooperativity or ultrasensitivity in
signaling pathways adds precision to "switch-like" events (measured here as entry into S
phase).

95
Figure 8: GlcNAc-TV deficiency Enhanas Cooperativity of TCR-mediateci
cellular activation.
(A) T ce11 activation by anti-CD3 antibodies in the presence or absence of anti-CD28
anti bodies (0.5 pglml) for ~ ~ a t ~ ' ' and ~ ~ a t 5 + ' + cells. Cultures were seeded at 5 x
10' cells/ml. (B) Sumrnary of the kinetics of responses in panel A. Hill coefficients
were calculated using log(y/l-y) = log Ka + n log A, where Ka is the Hill coefficient
and A is substrate concentration.

Dso and n, values for TCR response

97
(B) A sirnilar mechanism was recently described for progesterone triggenng of meiosis
in Xenopus oocytes (Ferre11 and Machleder, 1998). Thus, GlcNAc-TV-dependent
gl ycos ylation has a significant influence on agonist threshold and cooperativi ty of
TCR signaling.
GlcNAc-TV deficiency ùüectly affects antîgen-induceci TCR i n t e ~ n : We
reasoned that formation of TCR receptor aggregates in the plane of the plasma membrane
is a source of ultrasensitivity that may be regulated by glycosylation. A minimal or
threshold number of TCR must be engaged to initiate entry into S phase machman et
al., 1997; Wulfing and Davis, 1998a; Wulfing and Davis, 1998b), followed by rapid
cytoskeletal reorganization and TCR intemalization (Fischer et ai., 1998; Viola and
Lanzavecc hi a, 19%). To determine whether the ~ g a t ? mutation affects TCR
movement in response to antigen, we measured TCR intemaiization induced by anti-CD3
or by PMA plus ionomycin. Ce11 surface TCR levels in unstimulated Mgat5+'+ and
~ ~ a t 5 " T ce11 populations were comparable. TCR intemalization induced by anti-CD3
proceeded more rapidly in GlcNAc-TV deficient cells compared to ~ ~ a t 5 + / * cells with a
half maximal internalization observed at 1.5 and 2.2 min, respectively (Figure 9A).
Maximum TCR internalization was approximately 2.4 fold greater in Mgat5'- cells
compared to ~ ~ a t 5 + / + T cells. Consistently, TCR ligation induced a more rapid
depol yrnerization of actin filaments, an event that precedes the first step in actin polymer
reorganization, in ~ g a f 5 ' - T cells compared to wild type cells. MAPK phosphorylation,
a downstrearn effector of TCR signaling, aiso peaked more quickly and reached higher
levels in GlcNAc-TV deficient T cells (Figure 9B). Phosphorylated PKB, a kinase
downstream of both TCR and integrin signaling, was abnormdly low in resting ~ ~ a t 5 ' -
T cells, but increased afier stimulation to levels observed in ~ ~ a t 5 + " cells (Figure 9B).

98
In addition, cytoskeletal reorganization, as visuaiized by the breakdown of actin
polymers, was faster in ~ ~ a t 5 ' - T cells than in wild type cells (Figure 9C).
PMA stimulation results in CD3 recycling to the ce11 surface, and endocytic
degradation of CD4 (Ruegg et al., L992). To examine endocytosis in ~ ~ a t 5 ' - cells, we
monitored CD3 and CD4 levels in PMA-activated T cells. Although PMA induced
optimal ceIl proliferation, CD3 intemalization was only weakly induced, and
signi ficantl y, was simi lar for both Mgat.Sc/* and ~ ~ a t . F " T cells (Figure 9D). In contrast,
80% of CD4 was intemalized and d e p d e d in lysozornes cornpanxl to unstimulated cell-
surface levels in al1 three genotypes (Ruegg et al., 1992). Thus, endocytosis in ~ ~ a t . 5 ' ' T
cells does not differ from endocytosis in Mgat9'+ T cells. Rather, ~ ~ a t 5 + ' + and Mgat5'-
T cells differ at the level of extracellular TCR ligation, the pnmary force directing
receptor aggregation and internahzation.
Mg& expression is regulated by T cell activaîion: GlcNAc-TV enzyme activity
increases with activation of CD4+ and CDS' T cells, possibly through the activation of
Mgat5 transcription by the Ras/MAPK/Ets pathway (Buckhaults et al., 1997; Lemaire et
al., 1994; Lu and Chaney, 1993). Consistent with this possibility, B-galactosidase
activity produced by the gene product of the inserted lac2 reporter gene in ~ ~ a t . 5 ' - T-
cells increased 5 fold foilowing T ce11 activation for 48 hours. Furthemore, this resulted
in altered glycosylation of the ceIl surface, as L-PHA reactivity also increased in
~ ~ a t p " ' T cells following activation (Figure LOA,B).

99
Figure 9: GlcNAc-TV deficiency directly &ts antigen-induced TCR
(A) T cells from ~ ~ a t y - or ~ ~ a t 5 + / + mice were stimulated with 0.1 @ml anti-CDS, and
ceIl surface levels of TCR (relative mean flourescent intensity, MFI) were measured by
FACS analysis using FïïC-anti-TCRdg. The same result was obtained when the roles of
anti-TCRdp and anti-CD3 were reversed. (B) Phosphorylated MAPK and Akt/PKB in T
ceils fottowing addition of mti-CD3 antibdes. The numbers represent the ratio of
phosphorylated MAPK and PKB norrnalized to time O for each series. (C) Breakdown
and reorganization of actin was measured by rhodamine-phalloidin binding to actin
bundles following the addition of 0.1 Wrnl anti-CD3. The results represent duplicate
rneasurernents. (D) Splenocytes from ~ ~ a t 5 ' - rnice and from ~ ~ a t ? controls.
stimulated with 10 nglml PMA, were stained with FïT'C-anti-CD3 and PE-anti-CD4 No
significant difference in staining with either antibody was observed between the
genotypes.


Figure 10: Mgai5 expression is m t e d by T ceii activation,
(A) L-PHA binding to ~ ~ a t y ' + T lymphocytes either untreated (white) or stimulated
wi th O. 1 pg/ml of anti-CD3 and 0.5 pg/ml anti-CD28 for 48h (grey). (B) Lac2 activity in
untreated (white) and anti-CD3 and anti-CD28 stimulated (grey) T cells from M~U~Z' '

cell number
O s 8 8 8 8
cell number

DISCUSSION
GlcNAc-TV d e m n t mice: We g e n e d mice deficient in GlcNAc-TV activity to study
the biological role of B 1,oGlcNAc-branched N-glycans. M g a S message, GlcNAc-TV
activity and B1,oGlcNAc-branched N-glycans were not detected in Mce homozygous for
the MgarS targeted mutant allele. Therefore, these mice are functionally nul1 indicating that
only one gene encodes GlcNAc-TV activity.
GlcNAc-TV deficient mice are viabIe and fertile, unlike GlcNAc-TI deficient
mice, which do not survive past E9.5 (Ioffe and Stanley, 1994; Metzler et al., 1994).
Ioffe et al. have demonstrated chat matemal message of Mgat l is present in post-
implantation embryos and, therefore, possibly rescues the embryos from an even earlier
death (Ioffe et al., 1997). Although Mgal5 is highly expressed in the embryo from the
age of E9.5 (Granovsky et al., 1995), lack of GlcNAc-TV activity does not result in
significant developmental retardation or abnormalities. GlcNAc-TV deficient females
produce normal-sized litters when mated with males of al1 Mgut5 genotypes, suggesting
that. indeed, Mgat5 message is not necessary for normal development or fertility. At
E7.5, MgatS transcripts are expressed in the giant cells of the trophoblast, an extra-
embryonic invasive ce11 type, phenotypically similar to transfonned cells (Granovsky et
al., 1995; Hoffman and Wooding, 1993). Since the O-glycan core 2 GlcNAc-T is also
expressed in these cells at the same developmental stage (Granovsky et al., 1995), it is
possible that $1,6GlcNAc-branched O-glycans are able to functionally compensate for
the lac k of p l ,6GlcNAc-branched N-gl ycans. Even though N-gl ycans and O-gi ycans are
present on different glycoproteins and fulfill distinct biologicd roles, the cornrnon
extension of the $1,6GlcNAc-branch by polylactosarnine chahs may be the functionally
important structural feature of glycans in the giant cells of the trophoblast, enabling O-

104
glycans to rescue the ~ g a t s / - cells. A cross between core2 GlcNAc-T deficient mice
and ~ ~ a t . 5 ' - mice may address this question.
Although GlcNAc-TV deficient 129/sv female mice produced nomai litters, the
majority of these rnice failed to nurse their young. This deficiency could have been due
to either a lactation defect or to a behavioral abnonnality. Mamrnary development
appeared normal in M ~ ~ z z ' - females (see example in Figure 3, chapter 4), and milk was
evident in the marnmary glands of severai ~ g a t 5 ' - females that were examined within 24
hours of giving birth. These data, and the observed aggressiveness towards their pups,
suggest that the failure of ~ ~ î z t 5 ' - 129/sv females to nurse their young is due to a
neurological defect. Nurturing behavior defects have been ascribed to neural
abnormalities in previousiy characterized mutant mice. Mice deficient in FosB exhibit a
nurturing defect, while showing no other cognitive deficiency (Brown et al., 1996). The
aberrant behavior is IikeIy due to the absence of FosB in the preoptic area, a region of the
hypothalamus that is criticd for nurturing. A more severe cognitive phenotype, which
includes failure to nurture, is observed in Fkh-5-deficient rnice (Wehr et al., 1997). Fldi-
5 is a winged helix transcription factor and in adult mice it is expressed in the
marnmiliary body region of the caudal hypothalamus and in the midbrain. Mgar.5 is
highl y expressed in neural epithelium from E 9.5 and into adulthood (Granovsky et al.,
1995) and expression is detected in the hypothalamus. In the adult mouse brain, its
expression is restricted to neural cells and is absent from the supporting glial cells (Figure
2E). However, histological examination of ~ ~ a t 5 ' - central nervous system reveals no
obvious anatornical abnormaiities, suggesting that the defect in behavior may be a
biochemical defect in some aspects of potentiation (petsonal communication, J.
Henderson).

105
Mgut5 deficiency affects several ce11 types and tissues that have been previously
identified as high in GlcNAc-TV activity (Perng et al., 1994). In addition to the
behavioral defect, the ~ g a t f - mutation affects the kidney and the lpphoid system.
These findings suggest that GlcNAc-TV activity might be necessary for cellular
processes common to many ce11 types.
The lymphocytic infiltrates, obsewed in the histological examination of ~ ~ a t S ' -
mouse tissues, prompted us to investigate the inflammatory responses of these mice.
Arachidonic acid is the metabolic precursor molecule for the production of
prostaglandins and leukotrienes, whic h are vasodiaiators and ieukocyte-attracting
c hemokines, respective1 y. Topical application of arachidonic acid induces a rapid
migration of leukocytes to the site of application, and the subsequent attraction and
activation of T cells several hours later. Inflammation in response to arachidonic acid
was delayed but stronger in ~ g a t y - mice compared to wild type littermates. These
results suggest that the initial rate of leukocyte migration to the site of application may be
slowed, but that once leukocytes and lymphocytes are attracted, they are stimulated to a
greater degree and their stimulation lasts longer. The specific hypmeactivity of T cells
was confinned by the increased response in ~ g a t 5 ' - rnice to oxazolone. which induces a
T cell-mediated DTH reaction (type IV). Therefore, the results of these two
hypersensitivity models suggest that leukocyte migration and T ce11 activation may be
aberrant in ~ ~ a t . F ' ' mice. Interestingly, a full inflammatory response was not mounted
when ~ ~ o t 5 ' - rnice were injected with thioglycollate. Indeed, the infiammatory response
appeared to be over at 30 hrs, while the in flammatory response in wild type mice was still
rising. Thioglycollate stimulation results in the inhibition of arachidonic acid release,
with subsequent reduction in the production of arachidonic acid metabolites, but with
e n hanced production of TNFa (Carrick et ai., 1995). Thus, it is possible that the ~ ~ a t 5 ' -

106
mice may be hypersensitive to arachidonic acid metabolites, but hyposensitive to
thioglicollate and TNFa,
GlcNAc-TV deficiency &ers cellular aùhesion and migration: In this study, we
observed that fewer leukocytes migrated into the peritoneum when induced with
thioglycollate. Both peritoneal leukocytes and PMA-activated T cells were more
adherent to fibronectin in vitro. These results suggest that the observed delay in the
inflarnmatory response to arachidonic acid may be due to slower leukocyte migration
resul ting fmm stronger cellular adhesion to fibronectin.
GlcNAc-TV deficient fibroblasts exhibited intracellular architecture consistent
with stationary cells, including heavy actin stress fibers, lack of loçalization of focal
adhesion proteins and reduced spreading. ~ ~ a t 5 ' - fibroblasts in serum-free conditions,
as well as freshly harvested resting T cells, had aberrantly low levels of phosphorylated
PKB. PKB is a kinase downstream of PI3K, the activation of which is important for
integrin-mediated cytoskeletal reorganization that allows focal adhesion turnover,
membrane ruffling and subsequent migration (Clark and Brugge, 1995; Parent and
Devreotes, 1999; Stephens et al., 1998). The aspi integrin heterodimer (VLA-5) is the
prirnary cellular receptor for fibronectin, and both chains are subject to GlcNAc-TV-
dependent glycosylation (Demetriou et al., 1995). Overexpression of GlcNAc-TV in
MvLul cells resulted in enhanced cellular migration and decreased adhesion to ECM
proteins in vitro (Demetriou et al,, 1995)- Therefore, the enhanced adhesion of GlcNAc-
TV deficient leukocytes and T cells to fibronectin is consistent with the hypothesis that
GlcNAc-TV activity may modulate VLA5 binding to fibronectin and its subsequent
signaling.
TCR signalhg and flI,6GleNAc-&ronched N-glycans: We have shown that T cells from
GlcNAc-TV deficient mice are hypersensitive to T ce11 mitogens. In contrast, B cells

107
from GlcNAc-TV deficient and wild type mice were stimulated similarly by a variety of
B ce11 mitogens, indicating that the hypersensitivty is T ceil specific. Therefore,
f31.6GlcNAc-branched N-glycans may be present on and affect the biological activity of
glycoproteins required exclusively for T ce11 function.
The majority of the ceIl surface molecules comprising the TCRKD3 complex
(TCR), and the CO-receptor molecules CD4 and CD8, are glycosylated (Huppa and
Ploegh, 1997) (for detailed description of KR-mediated signaling see Figure 1 l)(Hanis
et al., 1990; Wu et al., 1996). CD45, a phosphatase important in the initiation of TCR
signaling has L-PHA reactive structures, indicating it is subject to GIcNAc-TV mediated
glycosylation. In addition, many of the reported costirnulatory molecules, such as CD28
and LFA- I (lymphocyte function-associated antigen- 1) are also N-glycosylated (Bajorath
et al., 1997; Miller and Spnnger, 1999; Monostori et al., 1997), although detailed
structural analysis of the glycans is yet to be done. The crystal structure of the C@ chains
of the TCR shows that 4 N-glycosylation sites are occupied on the B chain, and 3 are
occupied on the a chain(Wang et al., 1998). Of these seven chains, 5 are present on the
invariant regions (Ca and C$) (Wang et al., 1998). The carbohydrates of the a chain
have been previously characterized as complex-type, a likely substrate for GlcNAc-TV
(Hubbard et al., 1986).

f O8
Figure 11: Signaling initiated by TCR engagement.
The TCR is an oligomeric complex containing both antigen recognition subunits and
signal transduction subunits. The af3 subunits of the T ce11 receptor recognize peptide in
the context of a major histocompatibility complex (MHC) on the antigen presenting ce11
(APC), but they do not possess intracellular signaling domains (Huppa and Ploegh,
1997). The a p chahs are associated by salt bridges with CD3y, CD36 and C D ~ E , known
as the CD3 complex, and with a fourth protein, CD3c7 essential for signaiing(Janeway,
1992)). The cytoplasmic domains of d l four CD3 proteins contain sequences called
immunoreceptor tyrosine-based activation sequence motifs (ITAMs) (Janeway, 1992). In
addition to the stable TCR complex, a CO-receptor molecule (CD4 or CDS) transiently
associates with the complex dunng antigen recognition and enhances the sensitivity of T
cells to antigen.
Antigen induced signaling through the TCR is initiated by bringing together the TCR,
CD4 or CDS, and CD45, each of which is N-glycosylated (Zenner et al., 1995). CD45 is
a tyrosine phosphatase which removes inhibitory phosphate groups and thereby activates
the Lck and Fyn tyrosine kinases associated with the CO-receptor and the TCR,
respectively (Figure 10) (Clements et al., 1999). Lck phosphorylates iTAMs on CD36,
and the phosphorylated CD36 is then able to recruit ZAP-70 (zeta chah associated
protein-70) (clements et al., 1999)- The activation of ZAP-70 triggers intracellular
signaling cascades including the Ras pathway (Clements et ai., 1999), and signaling
through Ras culminates in the activation of Fos and the AP-1 transcription factors,
initiating new gene transcription necessary for T ceIl differentiation, activation and
proliferation.
TCR signaling is enhanced by the engagement of costimulatory molecules on the
T ce11 by their ligands on the APC. In particuIar, the costimulatory effects of CD28 when
engaged by the B7 lignads and of LFA-1 bound to 1-CAM1 are well doçumented
(Bachmann et ai., 1997; Lenschow et al., 1996; Shahinian et al., 1993; Yashiro et ai.,
1999).

a@ TCR chains CD4ICD8 CO-rcceptor A 6 1 y/& CD3 chüins
CD45 phosphatüse * ( CD3 chain
- ~ n t i ~ e n - ~ ~ ~ o n APC + PI3 kinase
87 ligand on APC
Fyn kinase
Lck kinase
ZAP 70
Phosphate

1 IO
X-ray crystdlography studies indicate that the N-glycan chains are not Iikely to obstmct
peptide presentation to the TCR, but by projecting outward, away from the protein, they
have the the potential to influence TCR aggregation (Garcia et al., 1996; Wang et al.,
1998).
Antigen induced signaling through the TCR is initiated by bringing together or
clustering the TCR, CD4 or CDS, and CD45, each of which is N-glycosylated (Zenner et
al., 1995). CD45 is a tyrosine phosphatase which removes inhibitory phosphate groups
and thereby activates the Lck and Fyn tyrosine kinases associated with the CO-receptor
and the TCR, respectively (Clements et al., 1999). Clustering of receptor and CO-receptor
glycoproteins brings together the various phosphatases and kinases and their substrates
and may be influenced by glycosylation. The bulk and charge of the glycan chains
provides steric hindrance to protein-protein interactions, impeding receptor clustering.
Alternatively, the mobility of the various molecules in the plane of the membrane might
be influenced by the kinetics of sugar-lectin interactions in cis or in trans (i.e. single ce11
or between cells, respectively). Indeed, the addition of exogenous gaiectin 1 to cultured
T cells antagonizes TCR signaling (Vespa et al., 1999). On B cells, the binding in cis of
oc2,6SA on polylactosamine to CD22 is essential for proper B ce11 maturation and IgM
production (Hennet et al., 1998).
There are three possible mechanisms by which glycan-glycan or glycan-lectin
interactions could modulate the physical interactions required for T cell activation. The
formation of the initial TCR-CD41CD8-CD45 complex, or subsequent TCR clustering
could be impeded by glycosylation, thereby slowing the rate with which activated TCR
aggregates are formed. Altematively, carbohydrate chains may destabilize the activated
cornplex, thereby reducing the time available for signaling, and, in effect, acting to
counterbalance CD4 and CDS. A third possibility is chat glycosylation alters the rate of

11 1
migration of the activated complex into kinase-rich "rafts", w hich are cholesterol-rich
membrane microdomains where many signaling molecules important for TCR activation,
such as ZAP-70, tçk and LAT are sequestered (Xavier et al., 1998). Slowed migration
into the "rafts" lengthens the period between TCR engagement and its interaction with its
signaling partners, thus reducing the probability of initiating T cell activation. The
mechanisms mentioned above could apply equally well to the binding and intramembrane
movernents of N-glycosylated costirnulatory molecules, leading to the modulation of
their biological activity. A combination of these mechanisms may operate in a complex
manner to produce the observed phenotypes.
TCR intemalization in response to mitogen stimulation is increased in GlcNAc-
TV deficient T cells (Figure 9A). TCR intemalization has k e n previously used as a
measure of TCR aggregation, a necessary step for TCR mediated signaling. TCR
aggregation and internalization require cytoskeletal rearrangement, which occurs faster in
~ ~ a t 5 ' - T cells compared to wild type T cells (Figure 9C). Cytoskeletal reorganization
in T cells, which allows glycoprotein receptors to move in the plane of the membrane,
plays a key role in TCR signaling (Lanzavecchia et al., 1999; Xavier et al., 1998).
Specifically, antigen presentation by APCs results in the formation of ordered protein-
rich domains at the interface between the APC and the T-cell, where TCR complexes and
other signaling molecules, such as LFA-1 aggregate in distict spatial distributions (Monks
et al., 1998). In addition, efficient TCR signaling is dependent on the formation of kinase-
rich "rafts" into which triggered TCRs are recruited (Xavier et al., 1998).
TCR agonists, such as anti-CD3 antibody and peptide-MHC on APCs induce
TCR clustering, and the threshold of T cell activation, as measured by calcium influx, is 3
TCRs per cluster, or 1 TCR cross-iinked to a CO-receptor molecule (Boniface et al., 1999;
Delon et al., 1999). When polyrners of haptens or polypeptide oligomers of peptide

112
antigens were used to stimulate T ce11 activation, an increased potency was observed with
increased number of antigenic units, reaching a plateau at about 16 units (Boniface et al.,
1999). Thus, T ce11 activation is dependent on the cooperativity generated by receptor
clustering, and Our results indicate that glycosylation may regulate TCR ciustering.
Cooperativity of T ceIl activation is also enhanced by the engagement of
costimulatory molecules, such as CD28. CD28 engagement by B7 ligands resuits in the
phosphorylation of CD28, and the activation of the tyrosine kinases Lck and EMT, thus
acting to amplify TCR signaling by enhancing early phosphorylation events (Gibson et
al., 1998). CD28 stimulation results in cytoskeletal rearrangement and raFt aggregation
(Viola et al., 1999), which appears to physicaily aid in sustaining TCR signaling. CD28
is heavil y N-gl ycosyiated, and lack of f3 1,6GlcNAc-branching may enhance this
receptor's engagement by the B7 ligands. Indeed, the effects of GlcNAc-TV deficiency
and CD38 engagement on T ce11 activation were synergistic, suggesting that signaling
mediated by both TCR and CD28 may be regulated by glycosylation.
The binding kinetics of effective APC-presented antigens are optimal when on-
rates allow recruitment of TCR clustering, but dissociation occurs at a rate optimal for
serial binding and recruitement of additional TCR molecules into the clusters. In this
fashion, a few agonistic peptide-MHC complexes are able, with time, to engage and
trigger a much larger number of TCRs by seriaily binding, triggering and disengaging
from TCRs (Valitutti et al., 1995). Serial triggering is an important regulatory element in
T ceIl activation, since a threshold number of triggered TCR must be achieved for
activation. The threshold number is not absolute, and is subject to modification by
costimulatory signals. For example, in order to activate human T-ceIl clones (-30,000
TCR/cell), 8,000 tnggered TCRs were required in the absence of CD28 costimulation,
but only 1,000 triggered TCRs were needed in its presence (Viola and Lanzavecchia,

113 1996). The engagement of TCR by peptide-MHC does not result in the immediate
triggering of TCR signaling. Rather, TCR signaling requires a series of phosphorylation
steps, ieading to the recruitment and activation of ZAP-70, which activates TCR-
mediated signaiing (See Figure 11). The multiple phosphorylation steps act as a
proofreading mechanism, since weakly bound peptides with fast off-rates do not remain
bound to the TCR long enough to complete the multiple phosphorylation events, and
therefore fail to trigger T ce11 activation. In this context, CD4 and CD8 coreceptor
molecules may act to enhance activation by binding the MHC and stabilizing TCR-
peptide-MHC interactions to allow weak agonists to engage TCRs long enough (-10
seconds) to reach the triggering threshold. Our study suggests that B 1,6GlcNAc-
branching of N-glycans may destabilize the TCR cornplex, thus exerting the opposite
effect to CD4 and CD8 engagement.
GlcNAc-TV deficient T-splenocytes of ~ ~ a t 5 ' - mice are quiescent and do not
spontaneously proliferate in tissue culture in the absence of T ce11 rnitogens. Thus,
f31.6GlcNAc-branched N-glycans do not appear to be essential for maintaining
quiescence, but are important in regulating T ce11 activation through altering the kinetics
of TCR aggregation in response to agonist. In the absence of ce11 surface 0 1.6GlcNAc-
branched N-glycans, TCR oligomerize more efficiently, thereby allowing activation to
occur at lower agonist levels and with greater cooperativity. In contrast, PMA-induced
activation, which bypasses the need for ce11 surface receptor ligation (Ruegg et al., 1992),
is similar in mutant and wild type cells, suggesting that downstream signaling in ~ ~ a t 5 ' '
T cells is intact. Therefore, the P1,6GlcNAc-branched N-glycans on the surface of T
cells appear to attenuate receptor aggregation, and thereby may prevent or reduce
spurious activation in the absence of high-affmity ligands in vivo.

114
Our results indicate that p l ,6GlcNAc-branched N-gl ycans suppress T ce11
activation and that Mgut5 gene expression is induced by T£ell activation. Therefore,
GlcNAc-TV upregulation rnay provide negative feedback, suppressing TCR responses in
activated T cells. Negative feedback rnay be required to suppress hyperactivation of T
ceIIs as they migrate into sites of infection where antigen concentration is higher.
Hyperactivation would be detrimental, as it rnay cause premature apoptosis, leading to an
unsustained and therefore ineffective T ceIl response. In addition, negative feedback is
necessary for shutting down immune responses and inflammation once infection is
cleared, and our results suggest that regulated expression of Mgat.5 in T cells is a potent
means used to regulate this process.
Immune regulation by Mg&: The results presented in this chapter rnay have
implications for hypersensitivities, allergies and the ability to clear infections. GlcNAc-
TV deficiency resulted in slower onset of inflammation in the two models of
hypersensitivities tested. Therefore, innate immunity rnay be compromised in these rnice
by the inability of leukocytes to migrate to the site of infection as rapidly as in wild type
mice.
The profile of the immediate-type hypersensitivity response to arachidonic acid in
~ ~ a t 5 ' - mice was markedly different than the response in P P A R ~ - mice, where the
oxidative degradation of leukotriene B4 is disrupted (Devchand et al., 1996). The
PPARQ" mice did not differ from the wild type mice in the amplitude of the
inflarnmatory response nor in the time during which maximal response was reached.
However. due to the slower clearence of leukotriene B4 in the PPAR~-'- mice,
inflammation persisted longer. In the ~ ~ o t 5 - I - rnice, the magnitude. the timing and the
duration of the response were al1 significantly different from the response in wild type
mice, suggesting that several processes rnay be penurbed by GlcNAc-TV deficiency.

115
The phenotype of the ~ g a t ~ ' - mouse is suggestive of the ~ ~ 4 3 " mouse, both by
the hypersensitivity of T cells and by the enhanced adhesion of T lymphocytes to
fi bronectin (Manjunath et al., 1995). In the ~ ~ 4 3 " mouse, vaccinia virus was cleared
less efficiently than in wild type mice, and the authors speculate that slowed migration,
tissue localization or the pst-lysis viral clearance by phagocytes may be impaired
(Manjunath et al., 1995).
The hyperactivation of T cells in M ~ U ~ S ' - mice suggests that these mice may be
susceptible to contact hypersensitivities and dlergies. T ce11 mediated responses were
not only higher in these mice, but were also slower to subside, suggesting that GlcNAc-
TV activity is required for the efficient shutting down of an immune response. This
defect may have implications for induced autoimmuni ty.
There is no data available on the variances in basal levels of GlcNAc-TV activity
in the human population. There is also no information about the differences in maximal
induction of the Mgut5 gene expression following T ce11 activation. The results presented
above suggest that both factors rnay contribute to the observed variability in human
responses to immune challenges. In addition to changing T ce11 sensitivity to antigens,
GlcNAc-TV activity changes the rate of ce11 migration, and therefore could potentially
have implications for the ability to clear infections. Our results suggest that patients
suffenng from hypersensitivities and allergies would be of particular interest for studying
their GlcNAc-TV activity profiles.

116
MATERIALS and METHODS
Mutation of the Mg& gene: A genornic library from strain 129/sv mice was screened
with a Mgat5 cDNA probe. A 13.5 kb genomic clone containing the 205 nt 5'
untranslated region and 241 nt spanning the first coding exon was used to constnict the
Mgat5 targeting vector. The Mgat.5 targeting vector was constmcted with LacZ replacing
the coding region of the first exon. The targeting vector was linearized with NotI,
electroporated into R1 ES cells, and transfected cells were selected in the presence of
G418 and gancyclovir as previously described. DNA from dnig resistant colonies was
digested with PstI and screened for homologous recombination by Southem blot analysis
using a 1.7 kb PstVXbaI genomic fragment extemai to the targeting vector. Two ~ g a t y -
ES ce11 Iines were aggregated with blastocysts from CD-1 mice, and implanted into
pseudo-pregnant CD-1 females. The resultant chimeras were mated with 129/sv females.
Heterozygous progeny were intercrossed to generate ~ g a t 5 ' - mice, and experiments were
performed on the 129/sv background- For histologicd examination, mice were perfused
with 10% buffered formalin in vivo, tissues were paraffin embedded, sectioned and
stained with hematoxylin and eosin. GIcNAc-TV enzyme activity in tissue homogenates
was determined as transfer of [ 3 ~ - ~ l c ~ ~ c from UDP-[~H]-GICNAC to the synthetic
acceptor GlcNAcS 1,2Manal,6Glc$l-octyl per mg of protein as described.
C P H A lectin and Western bloning: Proteins (50 pg per sample), separated on 8-16%
gradient SDS-PAGE under reducing conditions, were transferred electrophoretically ont0
PVDF membrane. Western blots were performed with antibodies to phospho-MAPK
(Thr202lTyr204) and anti-Akt/PKB and phospho-Akt/PKB (New Engiand Biolabs). For
lectin blots, membranes were probed with 0.05 pg/ml L-PHA, foliowed by incubation
with a rabbit anti-L-PHA antibody (1: 1000) and with HRP-donkey anti-rabbit antibody
(Arnersham).

117
Skïn Z n ~ n t a n o n : DTH response was primed by applying LOO pl of 5% (w/v) 4-
ethoxymethyiene-2-phenyl-2-0xazolin-5-o (oxazolone) in ethanoYacetone (3:1, vlv)
epicutaneousiy to the shaved backs of ~ g d mice and littemate ~ g a t y controls. Four
days after sensitization, the mice were challenged by applying 2 5 ~ 1 of 1% (wlv) oxazolone
in olive oiYacetone (3: 1, v/v) on each side of the right ear. As control, the left ear received
25 p1 of olive oiVacetone on each side. Ear swelling was measured with a micrometer at 24
hr intervals for the next 5 &YS, and swelling was reporteci as the difference between the
thickness of the right ear minus the thickness of the left ear. Results are reported for 7
~ ~ a t 5 " rnice and 6 ~ ~ a t S " mice. To investigate skin hypersensitivity. arachadonic acid (2
mg in 25 pl ethanol) was applied to the right ear of ~ g d - and ~ ~ a t T * littermate conml
rnice. The left ear received 25 pl of ethanol alone. Ear swelling was measured with a
micrometer at 2 hr intervals and was reported as above. Five mice of each genotype were
used per experiment and the experiment was repeated twice.
Induction of peritoneai exudafes and ceil adheswn: To induce leukoc yte extravasation,
mice received an i.p. injection of thioglycollate broth (2 ml of 3% w/v), and 3 h later
ieukocytes were recovered fmm the peritoneal cavity by lavage and counted. The cells were
irnrnediately resuspended in alpha-MEM at ld cells/mi and applied in 100 pl alaliquots to %
well plates coated with serial dilutions of fibronectin. Neutrophil and T ce11 adhesion
proceeded for 30 min, plates were washed with PBS and attached cells counted as described.
To examine focal adhesions, embryonic fibroblasts were allowed to attach ovemight to glas
slides coated with 1 pg/ml of fibronectin in semm free alpha-MEM. The cells were stained
with hodamine-phalloidin, anti-paxillin antibodies (Transduction Laboratories), and
Hoechst 33258 stain according to manufacturer's instructions. Fluorescence images of the
cells were obtained using a deconvolution microscope and digital capture.

I I 8
Stimulraion of T c e k in vitro: Spleen cells from 8-12 wk old mice were harvested and
stimulated for 48 h as previously described (Kniisbeek, 1991). High seeding density was
2 . 5 ~ 1 0 ~ cells/well, and low seeding density was 5x10~ cells/well. Two pCi of 3 ~ -
thymidine were added for the 1s t 20 h of incubation, and cells were harvested on
fiberglass filters and radioactivity was measured in a p-counter. T celis were purified
(>go% pure) by panning on plates pre-coated with anti-CD19 antibody (Pharrningen).
Stimulatory lectins and antibodies used were L-PHA and Con A (Sigma), anti-CD3
antibody, cione 2Cll (Cedadane) and anti-TCR antibody, clone H59.72 (generous gif t of
Dr. K. Sirninovitch).
FACS analysis and TCR intemalizdon: Single cell suspension of spleens, thymi and
lymph nodes were stained with FITC- or PE-conjugated antibodies (Pharmingen) reactive to
CD3c TCR-WB, CD4 CDS, CD69, B220, CD45 and CTLA4. FïïC-L-PH. (Sigma) was
also used for analysis. Propidium iodide (5 @ml) was added to the cells to facilitate dead
ceIl exclusion. Analysis was carried out on a FACScalibur machine using FACScan
software. To measure TCR internaiization, purified splenic T cells, stimulated with either
0.2 pg/ml anti-CD3 or 10 ng/ml PMA for varying lengths of time were harvested and
stained with FITC-anti-TCR. PMA concentrations were not lirniting as 10, 50 and 100
nglml produced similar intemalization and ce11 activation results. Lac2 activity in ~ ~ a r S " T
cells was detected by loading cells with fluorescin-di-PD-galactopyranoside (FDG)
(Molecular Probes) at l m , and ailowing the reaction to proceed for 30 min. The reaction
was stopped by the addition of 1 mM phenyl-$-thiogalactoside, and analysis was carried out
on a FACScalibur machine.

CHAPTER 4: GlcNAc-TV Regulates Tumor Growth and
Metas tasis

120
INTRODUCTION
GlcNAc-TV is transcriptionally regulated by the RadMAPKIEts pathway (Kang
et al., 1996; Lu and Chaney, 1993) and its activity is commonly increased in human and
rodent cancers. Studies on mutant tumor ce11 lines deficient in the enzyme, as well as
overexpression studies of Mgat.5. support the h ypothesis ihat f3 1,6GlcNAc-branched N-
glycans are positive effectors of tumor growth and metastatic potentiai (Dennis et al.,
1987; Lu et al., 1994; Seberger and Chaney, 1999). Although experiments with somatic
mutant tumor lines, and with revertant ce11 Iines, have provided convincing evidence that
1,oGlcNAc-branched N-glycans are required for efficient tumor development and
metastasis, some aspects of oncogenesis could not be explored using this approach. Al1
previous studies on the role of GlcNAc-TV in tumor progression and metastasis were
done with transplanted tumor cells, which were already fully transforrned. Therefore,
these experiments could not explore the role of GlcNAc-TV in tumor initiation, nor look
at the effect of GlcNAc-TV deficiency in the host animal versus tumor.
In order to examine the effect of GlcNAc-TV on tumor formation and metastasis
in a whole animal model, we crossed GlcNAc-TV deficient mice ( ~ ~ a r ? rnice) with
mice transgenic for the polyoma middle T antigen under the regulation of the mammary
specific MMTV promoter (PyMT). The PyMT antigen is a potent oncogene and its
transforming activity is dependent on its ability to associate and activate c-Src, PI3K and
Shc, which collaborate via intersecting signaling pathways to alter gene expression and
ce11 behavior (Guy et al., 1998; Guy et al., 1994; Webster et al., 1998). Expression of the
MMTV-PyMT transgene results in the widespread transformation of the mamrnary
epithelium leading to multifocal marnrnary adenocarcinornas, and the majority of mice
develop multiple metastases in the lung (Guy et al., 1992). The short latency p e n d

121
between the expression of PyMT, which starts with sex steroid production dunng the
maturation of the mammary gland, and widespread transformation, and the multifocal
nature of the arising tumors suggest that the expression of P y m alone is sufficient for
tumongenesis (Guy et al., 1992). Therefore, this mode1 is attractive because the influence
of heritable mutations on PyMT oncogenesis can be studied in vivo in a system where no
additional mutations are necessary to produce tumors.
In this study, we show that GlcNAc-TV deficient mice bearing the PyMT
transgene have a 1.5 fold extended latency p e n d in forming palpable rnammary tumors.
These tumors aiso grow at a 6.7-fotd slower rate than do tumors of wild type littermates,
resulting in 4.5-fold reduction in tumor weight in late stages of the disease. GlcNAc-TV
deficiency also resulted in >95% reduction in the incidence of lung metastases. By
activating c-SrcIFAK, Shc/Ras and PI3K, the P y m protein contributes to the
reorganization of cytoskeletal structures and turnover of focal adhesions. GlcNAc-TV
deficient embryonic fibroblasts and primary Pym-transformed tumor cells show
reduced spreading and increased cortical stress fiber content when plated on fibronectin
in serum free media. The results presented below suggest that GlcNAc-TV activity may
enhance the formation and turnover of focai adhesions and thereby increase signaling via
the integrifl13WPKB pathway, thus enhancing ce11 proliferation, migration and
metastatic potential.

122
RESULTS
Ceneration of P y ~ ~ ~ ~ a t ~ ' - r n i c e : ~ g a t 5 + ~ femaies were crossed with a PyMT founder
male to generate P ~ M T M ~ U ~ ~ + / - mice. P ~ M T M ~ ~ ~ S + ~ males were crossed with Mgat5+'-
fernales, and 42 PyMT fernale progeny were used in the expenments. A PCR strategy
was employed to identify PyMT mice and to determine their Mgat5 genotype (Figure
1A). PyMT expression was similar in tumors from mice of al1 three genotypes, as
determined by Northern analysis (Figure 1B).
Mammary tumors fom wilh greater Iolency in ~ ~ a t 5 ' - mire: ~ ~ a t 9 " and Mgat.5+'-
rnice developed palpable tumors by the age of 8 weeks, but no palpable tumors appeared
on ~ga2.F'- mice until 12 weeks of age. Tumors were detected in al1 10 marnmary pads
of ~ g ~ t 5 + / + and ~ g a t S * / Mce by 20 weeks of age, but not until week 27 in Mgat5'-
mice (results summarized in Figure 2a). The time (Tso) to 50% of the marnmary glands
developing palpable tumors was 14 weeks for Mgat9'+ and MgatS'- mice, and 20.7
weeks for ~ g a t 5 ' - rnice (a 1.5-fold delay). Marnrnary pad from ~ ~ a t s / - mice developed
normally, indicating that the effect of GlcNAc-TV deficiency on PyMT-mediated
tumorigenesis is not due to altered mammary pad development. In addition, microscopic
tumor nodules were observed in mammary pads of 19 week old rnice of al1 Mgat5
genotypes. The number of tumor foci ranged from O to >LOO, and did not correlate with
the Mgat5 genotypes. Thus, the multifocal nature of PyMT-dependent tumors is
main tained on a GlcNAc-TV deficient background (Figure 2b).

123
FIGURE 1: Generation of ~ ~ o t 5 ~ ~ ~ h l T mice and Pym transgene expression.
(A) PCR was used to identify PyMT positive rnice and to determine their Mgat5
genotype. PCR with PyMT specific primers amplifies a 530 bp fragment. The wild type
allele of MgarS was detected b y PCR with primers specific for the first coding exon of
Mgar.5, which generated a 446 bp fragment. The mutant allele was detected with primers
specific for LacZ, which generated a 320 bp fragment. (B) Northem andysis of P y m
transgene expression. RNA was extracted from PyMT marnmary tumon h m ~~at .5+'+,
~ ~ a r . 5 " - and ~ ~ a t 5 ' - rnice and 10 pg were loaded per lane. The RNA was probed with
a1.5 kb cDNA fragment comprising the entire coding sequence of PyMT and the
membrane was stained with methylene blue to visualize the 28s RNA band to ensure
equivalent loading and uansfer.


FIGURE 2: PyMT tumor development is delayed in ~ ~ u f 5 ' - mice.
(A) Li ttermates transgenic for P y m on ~ g a t 5 + ' + , ~ g a t 5 + ~ or ~ ~ a t 5 ~ bac kground were
palpated for the development of mammary tumors starting at 6 weeks of age. In total, 9
~pt .S"' , 17 ~ ~ a t ~ + / - , and 14 ~ g a t 5 ' - P y m female rnice were examined. (B)
wholemount preparations of the 4th mammary pad from 19 week old ~ y h 4 T ~ ~ a t y ' mice
were stained with Cannine alum stain to visualize tumor microfoci prior to the formation
of palpable tumors. Multiple foci were observed in these mammary pads, and no apparent
correlation was observed between the number of foci per mammary paa and Mgut5
genotype, when 4 ~ ~ a t 5 " + , 6 ~ ~ a t s ' ~ ' and 4 ~ g a t 5 ' - mammary fat pads were compared.

% Tumour free

127
Tumor growth k slowed Ui ~ ~ o t 5 ~ mice: In order to compare the rate of tumor growth
in ~ ~ a t 5 - l - mice to ~ g a t 5 + ' - and ~ p t s ' / + rnice, palpable tumors were measured weekly
and the tumor area was plotted (Figure 3A). Tumors in tu ad- mice grew at a 6.7 fold
slower rate than did tumors in ~ g a t s ' ~ or ~ ~ a t 5 " + mice. The slower growth rate was
reflected in a 4.5 fold decrease in tumor weight at 26 week old rnice (Figure 3B). MgatS-
A rnice had 3.4 f 0.8 g of tumor as compared to the ~ g a t 5 " - and ~ g a t 5 + ' + mice which
averaged 15.1 k 1.8 g and 13 f 2.8 g, respectively. A small subset of GlcNAc-TV
deficient tumors (9140, 3.7%) grew slowly initidly, but had a sharp increase in their
growth rate to resernble ~ ~ a t F ' + and ~ g a t 5 " - tumor growth 3-4 weeks after tumors
became palpable (Figure 3C). Marnmary pads from 26 week old GlcNAc-TV deficient
mice displayed a range of histopathology, including hyperplasia, non invasive dysplasia
and invasive carcinoma with necrosis (Figure 4A-B). In contrast, Al1 mammary pads
from 26 week old ~ g a t 5 + ' + and ~ ~ a r 5 " ~ mice had tumors that were without exception
invasive, undifferentiated and with central areas of necrosis (Figure 4C). Hyperplasia is
defined as intraluminal proliferation of cells, resulting in a duct wall that is more than one
ce11 thick. Dysplasia is defined as the loss of the uniforrnity of individual cells, as well as
their architectural orientation. Malignant tumors show increased nuclear content and an
irregular, invasive growth through basement membrane.
Adjacent sections to the hematoxylin and eosin stained sections were
imrnunostained to quantify apoptosis and proliferation. Klenow was used to label broken
DNA fragments in apoptotic cells, and a monoclonal antibody was used to detect
proliferating cell nuclear antigen (PCNA) in cycling cells. The percentage of apoptotic
cells was not significantly different between genotypes (Table 1). However, proliferation
was significantly reduced in Mga& tumors (Figure 5A-D) in al1 stages of tumorigenesis.

FIGURE 3: GlcNAc-TV deficiency causes slower tumor growth.
(A) Palpable rnammary tumors on rnice of al1 Mgat5 genetic backgrounds were measured
weekly with calipers, and the average of two measurements / tumor / week was plotted.
In total 50 ~ g a z 5 + ~ + , 5 0 ~ g a t Y ' - and 5 0 ~ g a t 5 ' tumon were measured. Fast growing
tumors were not included in the ~ g a t 5 ' - category. (B) Pym-transgenic mice were
weighed at 26 weeks and the weight of a littermate femaie not bearing the PyMT
transgene was subtracted. 11 ~ g a t 5 + ' + , 15 ~ g a t 5 + / and 14 ~ ~ a t 5 ~ were weighed. (C)
A small subset of GlcNAc-W deficient tumors (51140, 3.6%) displayed an altered
growth rate 3-4 weeks after they were first detected by palpation. The growth of three
such tumors are plotted-

2 4 6 8
Time (weeks)
2 4 6
Time (weeks)

130
FIGURE 4: Histopathology of mammary fat pads and turnors in 26 week old
P~MTM~&- and P ~ M T M ~ U ~ ~ mice.
Tumon from ~ ~ a t 5 ' - (A,B) and ~ ~ a t 5 + ' - (C) were paraffin-embedded and 8 micron
sections were stained with hematoxylin and eosin. h, hyperplasia; d, dysplasia; t, tumor;
n, necrotic area.


132
FIGURE 5: Proliferntive index in mammary fat pads and hunors in F ~ M T M ~ &
and P ~ M T M ~ U ~ ~ + ' - mice.
D ysplastic epithelia (A.B) and invasive carcinomas (CD) from ~ ~ a r 5 + ' - (AC) and
~ ~ a t 5 ' - (B,D) were subjected to immunohistochernistry with an antibody against the
proliferating cellular nuclear antigen (PCNA) to visualize proliferating cells. (E) A
significant difference (p4 .05) was observed in the proliferative index in al1 stages of
transformation between the two genotypes.

PCNA positive cells (%)

Table 1: Number of apoptotic celis per IOX marncation view.
Dysplasia
Invasive tumor
Tumon from ~ ~ a t p - and ~ ~ a t . ' ' rnice were parafin embedded and sections were
Iabeied with Klenow to visualize apoptotic cells. Ten random fields / mouse were scored,
and tumors from two mice genotype were exarnined.

135
Expression Jiom the MgatS locus is enhanced with PyMT îmnsfonnation and tumor
progression: The mutant allele of MgatS was generated by replacing the coding portion
of the first coding exon with the bacterial reporter gene LacZ, encoding p-galactosidase.
Thus, expression of L a d is under the control of the MgatS promoter, and reflects the
tissue distribution observed for Mgut5 transcripts by RNA in situ hybridization
(Granovsky et al., 1995), (Chapter 2). Marnmary fat pads from ~ ~ a t y ~ mice were
stained with x-gal to visualize p-galactosidase expression in tumors of different sizes and
apparent stages of progression. f3-galactosidase activity was not evident in normal
mammary epithelium, but activity was detected in tumors (Figure 6A). Uniformly intense
p-galactosidase activity was observed in Mga&- tumors. However, in ~ ~ a t . F ' - tumors,
intense staining was observed focal1 y, while the majonty of the tumor area was weakl y
stained (Figure 6B). Lac2 expression was uniform and high in 3/3 fast growing ~gat.5'
tumors, comparable to that of Mgat5+'- tumon (Figure 6C).
GlclVAc-TV deficiency supresses metastasis: Mice transgenic for Pym develop
metastatic disease in lung, and GlcNAc-TV activity and the B 1,oGlcNAc-branched N-
glycans has been causally implicated in increased metastatic potential. The numben of
surface lung metastases were compared across the M g d genotypes (Figure 7A). A 20-fold
reduction was observed in the number of metastases in Mg& mice compared to Mg&
and MgatS" controls. When the number of metastatic nodules was nomalized to Nmor
weight pet- animal. an Il-fold reduction was observed in ~~a2 .Y' ' rnice compared to M~UZY
and ~ , g a t S ' - controls (Figure 7B). In addition, the average size of the metastatic nodules
was smaller in the Mg& mice. Microscopie examination of lungs fiom 6 ~ ~ a t 9 mice
revealed no rnicroscopic tumor foçi.

136
FIGURE 6: Lac Z expression in hfgotS/' and ~ ~ a t ~ tumors.
(A) Whole marnmary pads from 14 week old ~ ~ a r 5 + ' - mice were stained for $-
galactosidase activity. Staining was not observed in premalignant marnmary fat pads, but
intense staining was observed in tumor foci (arrows). (B,C) Tumors were resected from
26 week old ~ ~ a t 5 ' - mice and stained for p-galactosidase activity. The majority of
~ ~ a r ~ ~ tumors exhibited punctate x-gal staining (B), but fast growing tumors (C) stained
intensiy and uniforrnly. lac Z, foci of x-gal staining; n, necrosis.

FIGURE 7: GlcNAc-TV activity is required for metastasis.
(A) ~ ~ a t s ' ' + and ~ p t 9 ' - rnice were sacrificed when tumor burden reached !4 body
weight, or when bleeding from ulcerated tumors was observed, and their lungs were
resected and visually exarnined for surface metastatic foci. ~~a t .5 - ' - were sacrificed at the
sarne time as their ~ ~ a t 5 + ' ' + and ~ ~ a r 9 ' ' littemates (26-28 weeks, denoted by black
circles). White circles indicate the number of metastatic foci on lungs from mice
sacrificed at 19 weeks of age with low tumor burden and with no evidence of ulcerated
turnors. White squares indicate the number of metastatic foci on lungs from ~ ~ a r 5 - I -
mice with fast-growing tumors. (B) The number of metastatic foci was norrnalized to the
tumor weight of the animai.

1 O Mce sacrificed at 19 weeks I 1 O Mgat5-1- mice with fast growing tumors 1

140
The number of metastatic foci was normalized to the tumor weight of the animal.
The average number of metastatic foci / g tumor was 0.37M.14 in ~ ~ a t . 5 ' - mice,
compared to 5 . 0 . 9 and 6.lf 1.9 foci / g tumor in ~ g a t 9 ' - and ~gat.?'+ mice,
respectively. Tumor burden on the lungs often results in cardiac hypertrophy, a
physioiogical response to poor lung capacity. Cardiac hypertrophy was scored in 13/27
~ ~ a t . 5 ' ' ~ and ~ g a t 5 + / mice, but in 0/15 ~gat.5' mice.
~ ~ ~ r 5 ' - PyMT tumor cells display aberrant cytoskelelal architecture: In order to
investigate the effect of GIcNAc-TV deficiency on ce11 adhesion and cytoskeletd
rnorphology, PyMT tumor cells were plated on fibronectin-coated cover slips in serum
free media, and subsequentl y stained with the nuclear stain Hoechst322, actin-specific
rhodamine-phalloidin and FïïC-anti-paxillin antibody (Figure 8A-D). The PyMTMgatS
'- (Figure 8B) tumor cells showed impaired membrane ruffling and a dispersed and
punctate distribution of focal adhesions compared to ~ ~ M T ~ g a t ~ + / - t u r n o r cells (Figure
8.4). The P ~ M T M ~ ~ Z ~ ' - tumor cells had denser actin fibers with very little paxillin
localized to putative anachment sites at the edges of the cell. Paxillin binds 01 integrin,
as well as signaling molecules FAK, Csk and c-Src at discrete sites of ceil attachment in
spreading and motile cells. Membrane ruffling and filopodia formation require PUK
activation. The pattern of actin stress fibers in M@+/- tumor cells treated with
wortmannin, a P13K inhibitor, was sirnilar to that of Mgat5/ tumor cells, with heavy
cortical actin fibers replacing the fine actin filaments of ~ g a t 5 + ~ cells cultured in the
absence of wortmannin (Figure 8A and C). Cell spreading and membrane ruffling
induced

141
FIGURE 8. Effects of GlcNAc-TV deficiency on intracellular cytoskeletal
architecture.
Mgat5+'- (A) and ~ ~ a t 5 ' - (B) P y m tumor cells were plated on fibronectin and stained
with the nuclear stain Hoechst322, actin-specific rhodarnine-phalloidin and FïîC-anti-
paxillin antibody. Fine actin filaments and ruffling were obsewed in ~ g a t 5 + ~ tumor
cells. characteristic of motile cells. In contrast, ~ ~ a t 9 - tumor cells exhibited
morphology characteristic of stationary cells, with cortical actin stress fibers establishing
the perimeter of the cell.
(C) The staining pattern of ~ g a t . ? ~ tumor cells incubated with wortmannin was sirni lar
to that of ~ ~ a t 5 ~ cells, with heavy actin stress fibers replacing the fine actin filaments of
~ ~ a t 5 ' ~ cells. @) Cells from a fast-growing ~ ~ a t 5 ' - tumor were stained for actin and
paxi 1 lin. The ce1 1 population showed considerable heterogeneity.


143
by PyMT in serum free conditions is impaired in ~ ~ M T ' ~ g a t 5 ~ tumor cells. suggesting
that PyMT-induced activation of focal adhesion signaling is reduced in ~ ~ M T ~ g a t f -
tumor cells.
Cells from a fast-growing ~ ~ a t 5 ~ tumor were also plated on fibronectin and
stai ned for actin and paxillin (Figure 8D). Greater morphological heterogenei ty was
observed in these cells than in slow-growing ~gar5'- cells. Cells with filapocha, focal
adhesions and actin fibers, as well as cells with focal adhesions but little actin filaments
were observed, This mixed ce11 population may reflect tumor progression and evolution
of the population towards a phenotype similar to that of the ~gat5+'- phenotype.
Wild-type ES cells fonn tumots in ~ g d mke: Subcutaneously injected ES cells form
teratocarcinornas in syngeneic mice. GlcNAc-TV-dependent glycosylation affects both
host and tumor glycoproteins, and therefore a host effect on turnor growth could not be
ruied out. In order to investigate the contribution of host GlcNAc-TV on tumor
formation and growth, we injected wild-type ES cells into ~gat5* /+ , ~ ~ a t 5 + ~ and Mgar5
'- rnice. Tumors forrned in al1 ~gat5+'+ and ~ ~ a t 5 ~ animals, demonstrating that ~gat5 ' -
animals do not mount a significant anti-tumor immune response against cells with
B1,6GlcNAc-branched glycans. However, the kinetics of tumor formation and growth
were significantly different in ~gar5+/+ and ~ ~ a t 5 ~ mice (Figure 9). Formation of
tumors was delayed in ~ g a t . 5 ~ mice until week 6 compared to week 3 for wild type
mice, but once detected, tumors grew significantly faster than tumors in ~ ~ a r 5 + / + mice.
Surprisingly, tumors did not grow in 414 ~ ~ a t . S + ~ , a result requiring further investigation.
These findings suggest that glycosylation of glycoproteins in the host microenvironment
of the tumor plays a significant role in tumor progression.

144
FIGURE 9: GIcNAc-TV deficiency in host animrils acfixts growth characteristics
of wild type ES cell-derived teratocarchomas.
2x10~ wild-type ES cells were injected S.C. into the left hind leg of ~ ~ a t ~ ' - & f g a t f l - and
~ ~ a r 5 " + mice. Mice were palpated weekly for the formation of tumors, and the
diameter of palpable tumon was measured weekly. Tumon formed on 515 ~ g a t 5 ' - , 014
~ ~ a t 9 ' ' and 4/4 ~ ~ a t p + mice. The graph represents growth measurements of 415
~ ~ a t 5 ' - , 4/4 ~ ~ a t f " and 3/4 ~ ~ a t S / + rnice. One ~ ~ a t 5 ' - mouse and 1 ~ g a t P + rnouse
developed ascites tumors and were not încluded in the growth measurements.

Time (weeks)

146
DISCUSSION
GlcNAc- T V deficàèncy dekys but does not inhibit tumorigenesis: GlcNAc-TV acti vity
has b e n causally implicated in tumor growth and metastasis (Dennis et al., 1987; Lu et al.,
1994; Seberger and Chaney, 1999). To elutidate the role of GlcNAc-TV in these processes,
we generated a GlcNAc-TV deficient mouse and mssed it with a transgenic mouse
expressing the PyMT antigen under the conml of a mammary-specific promoter. Pym is
a potent oncogene, which becomes tyrosine phosphorylated and serves as a docking protein
for membrane localization and activation of Src kinases, PI3K and the Shc adaptor protein,
al1 of which contribute to transformation (Blaikie et al., 1997; Webster et al., 1998; Yi and
Freund, 1998). P y m is sufficient to transform immortalized cells in culture, and the
MMTV-controlled transgene induces multifocal tumorigenesis in breast epithelium
@ilworth et al., 1994; Guy et al., 1998).
Mice of al1 Mgat.5 genotypes developed invasive mammary tumors in 10/10
marnmary pads by 26 weeks of age. Multiple tumor foci were detected in 19 week old
rnice, even in mammary fat pads that lacked palpable tumors. The number of foci was
variable with no significant difference amongst the genotypes in the small number of
sarnples examined. The high incidence of in situ carcinomas and the development of
multiple tumors per mouse and per fat pad indicate that Pym-induced multifocal
transformation occurs in ~ ~ o r . 5 ' - rnice, but growth of the tumon is delayed markedly.
The PyMT oncogene activates two main pathways that collaborate to allow the
formation of tumors. The Ras/MAPK pathway is activated by the association of PyMT
with Shc and Src, and the PI3K/PKB pathway is activated by engagement of PI3K (Guy
et al., 1994; Meili et al., 1998; Webster et al., 1998). In contrast to wild type mice
carrying the MMTV-Pym transgene, c-Src deficient mice rareiy developed mammary

147
tumors, suggesting that c-Src is essential for PyMT mediated transformation (Guy et ai.,
1994). Src activation is cornmonly increased in human breast tumors making the study of
Src as relevant a mode1 to the human disease (Verbeek BS et al., 1999). as the study of
mutations affecting the Ras and the PI3WPKB pathways, also common in human tumors
(Velders and Kast, 1999). However, Src is not sufficient for transformation, as mice
transgenic for activated c-Src develop hyperplasia but no tumors (Webster et ai., 1995).
Muller's group has generated two transgenic mouse lines carrying mutant QMT antigens
that are unable to associate with either PI3K or Shc (Webster et al., 1998). Both
mutations attenuated the delay and reduced tumorigenesis. The presence of either mutant
was sufficient for the formation of global marnrnary epithelial hyperplasia, which
progressed to the formation of focal mammary tumors with a longer latency than
observed in mice transgenic for wild type P y m . Consistently, both mutations in PyMT
caused a 2.3-2.7 fold delay before palpable tumors were formed. In mice transgenic for
either PyMT mutant, the transition from hyperplasia to tumor was commonly associated
with a secondary mutation. A second mutation in the PyMT gene, restoring the ability of
PyMT to bind Shc, was observed in a subset of tumors arising in mice canying the Shc
binding mutant of PyMT. Tumors in mice transgenic for either PyMT mutant showed
increased expression of ErbB-2 and ErbB-3 receptor tyrosine kinases, which recruit and
activate P13K and Shc in a ligand dependent manner (Webster et al., 1998). The
phenotype of the ~ ~ a t 5 ' - mice was different from that of the mice transgenic for either
mutant in several respects. Palpable tumors arose earlier in the ~ ~ a t 5 ' - mice than in
either of the PyMT mutants, which were delayed significantly longer. As well, the
rnultifocal nature of the tumors in ~ ~ a t S / ' rnice was similar to the phenotype of the
tumors arising in mice transgenic for wild type PyMT. Thus, we concluded that GlcNAc-

148
TV activity is not critical to PyMT-dependent tumorigenesis, but rather, significantly
retards tumor growth.
GlcNAe-TV dejirency slows amor growth: Although MgafT" mice developed tumors in
al1 marnrnary pads examined, there was a significant lag in the formation of palpable
tumors. The delay observed on the MgatS" genetic background is comparable to that
observed for Pym-mediated tumors in Grbî* and ~ t s - 2 4 rnice (Cheng et al., 1999)
(personal communication, R. Oshima). Grb2 is an adaptor pmtein, which mediates Ras
activation by Shc and by receptor tyrosine kinases (Cheng et al., 1999). Ets-2 is a
transcription factor positioned downstream of Ras, which is required for expression of
MMPs and VEGF (Wasylyk et al., 1998). The promoter of MgarS contains APL, AP2 and
PEA3/ets sites (Saito et al., 1995), and Mgai5 is activated by oncogenes in the Ras pathway
(Kang et al., 19%; Chen et al.. 1998). The slower development of tumors in MgatF- mice is
consistent with the observed phenotypes of mice bearïng other mutations in the Ras
pathw ay . There fore, GlcNAc-TV may be a downstream effector of PyMT-mediated
transformation and metastasis, in a manner similar to downstream effectors of Ras signaling.
VanderElst and Dennis have previously shown that reduced branching of N-
glycans on the surface of MDAY-D2 mutant tumor cells resulted in slower cellular
growth rates in vitro and in differences in tumor weight in vivo (VanderElst and Dennis,
1991). Interestingly, relatively srnall differences in the measured doubling time of the
cells in vitro ( 12%) resulted in more than doubling of the tumor size in vivo over 3 weeks.
~ ~ a r . 5 ' - cells proliferate significantly less than ~ ~ a t . 5 ' " cells in al1 stages of
transformation (see Figure 6E), as measured by PCNA immunohistochemistry. ~ g a t ~
mice have a reduced proliferative index by 2-fold in hyperplastic epithelium, by 1.5-fold
in dysplastic epithelium, and by 1.1 fold in invasive tumors, compared to ~ ~ a t 5 " - mice.
Al1 differences were statisticaily significant. These diffetences in proliferation are

149
sufficient to account for the 6-fold difference observed in the growth rate of tumor area
and the 4.5 difference in tumor weight. Indeed, if al1 other conditions were the sarne, a
4.4% difference in cellular doubling times would be sufficient to account for a 4.5 fold
difference in tumor weight over the period of 7 weeks, which is the period used in this
experiment*.
Mg& expression is a marker of tumor progression: Analysis of Lac2 reporter gefie
expression in M g a t y and M g m y mammary fat pads showed that there are 3 relative levels
of f3-galactosidase activity in these tissues. Noxmai and hyperplastic mammary epithelium
of ~ g a t S " mice did not express L a c 2 but intense staining was observed in Nmors from
these mice. The majority of tumon from ~ g a t . 5 ' - mice (96%) grew more slowly than tumors
from M~L& or MgaiS" rnice, and Lac2 expression was low. with only sporadic foci of
intense staining. However, in the fast-growing ~ ~ a t S " turnors lac2 expression was
comparable to that of Mga? tumors. This suggests a secondary mutation or epigenetic
event has oçcured in these PyMT tumors and provided both a growth advantage and restores
Mgal5 promoter activity to that of the ~ g a t T * or wild type F'yMT tumors. The staining
pattern indicates that GlcNAc-TV activity and its products provide positive feedback for
optimal signaling leading to expression from the Mg& locus.
* formula for calculating the relationship between tumor weight and cellular proliferation
ratios: [ ~ p d t u r n o r weight] / [ ~ ~ a t p + tumor weight] = [e 4'n2>49] 1 [e(1n2v9] , w here A
is the fold difference in cellular proliferation (VanderElst and Demnis, 1991).

150
Optimal expression is restored in the fast-growing M ~ ~ Z P - tumors, suggesting
that the restoration of fast growth and MgatS expression require signaling through the
same pathway. Lac Z expression is not observed in normai or hyperplastic mammary
tissues, indicating that transcription from the Mgut5 locus is turned on after PyMT
induced transformation. This staining pattern further supports the mode1 that GlcNAc-TV
activity functions p s t transformation to influence the rate of tumor growth, but is not
required for tumor formation.
GlcNAc-TV activity is requkd for e m n f metustasis and might be necessuq for
optimal fyMT-induced PI3K signaling: Tmsfection of Mg& into metastatic mamrnary
tumor cells enhances their metastatic potential 4 - 4 ) fold (Lu et ai., 1994). In the current
study, GlcNAc-TV deficiency resulted in >95% reduction in lung metastases. This
observation held true when the number of metastatic foci was nonnalized to the weight of
the primary tumors per animai and in Mgat5'- rnice with fast-growing tumors. Therefore, it
is possible that metastatic potential and tumor growth rates are separate and distinct
phenotypic features of the Mga& mouse. The distinction between tumor jpwth and
metastatic potential has been previously observed in plasminogen deficient rnice (Bugge et
al., 1999). Plasrninogen is a protease that efficiently degrades fibrin, thus participating in
ECM remodeling. Expression of the P y m transgene in plasminogen deficient mice
resul ted in the developmen t of mamrnary tumors indistinguis hable from those developed by
wild type littermates. However, a 6-fold reduction in lung metastases was observed in
plasminogendeficient mice (Bugge et al., 1999).
The abrogation of metastasis in ~ ~ a t . ! f mice might be due to changes in the
ability of ~ ~ a t ~ ' / ' tumor cells to migrate on ECM. ECM-dependent proliferation and
migration is mediated primarily by the integins, a family of ceIl surface heterodimeric
receptors. Integrins are comprised of extracellular domains that recognize specific ECM

151
proteins, and cytoplasrnic domains, which associate with bundles of actin microfilaments
and affiliated structural proteins, wch as vinculin. Integrin subunits as, a. and pi have
been previously shown to be substrates for GlcNAc-TVdependent glycosylation
(Demetnou et al., 1995). These subunits fom VLA-5, the cellular fibronectin receptor
(Ruoslahti, 1996). MgarS-transfected MvLul cells are less adherent to fibronectin and
are more migratory than the parental ce11 Iine (Demetriou et ai,, 1995), supporting the
hypothesis that 1,oGlcNAc-branching of N-glycans may replate cellular adhesion and
migration.
As was shown in Chapter 3 (Figure 6) and in this chapter, the cytoskeletal
organization of ~ ~ a t 5 " - fibroblasts and Pym tumor cells differs from wild type and
~ ~ a t Y ' - cells. In addition, resting M ~ U Z ? T cells have higher content of actin stress
fiben than do wild type cells. These observations suggest that ~ ~ a t r - cells are more
stationary, and that adhesion-dependent signaiing requires f31,ffilcNAc-branched N-
glycans. In ~ ~ a r 5 ' - embryonic fibroblasts, paxillin, a downstrearn effector of Src
activation by integrin signaling, was not localized to focal adhesions, in contrast to
~,garT'- cells. The failure of ~ ~ u t y - fi broblasts to properl y localize paxillin indicates
that GlcNAc-TV deficiency inhibits focal adhesion formation and turnover required for
ce11 spreading and motility. Paxillin is a multi-domain molecule with binding sites for
both structural proteins and signaling proteins involved in cell motility and cell
proliferation (Turner, 1998). Upon cellular adhesion to ECM, paxillin is targeted to focal
adhesions by an as yet undefined recruiting molecule, where it interacts with the focal
adhesion proteins vinculin and talin, and where it is phosphorylated by FAK and Src. The
phosphorylation of paxillin results in the formation of S H 2 domains which serve as
docking sites for signaiing proteins such as Crk, thus contributing to signaling through
the Ras pathway. The physical features of paxillin suggest an important role for this

152 protein in recruiting common signai cascade components into close proximity with one
another, thereby allowing efficient signal transduction through both ceIl proliferation and
ceIl motility cascades (Turner, 1998). Thus, the lack of paxillin localization to putative
focal adhesions in GlcNAc-TV deficient fibroblasts may result in inefficient signaling
through RadMAPK and PI3K/PKB pathways.
Semm starved GlcNAc-TV deficient fibroblasts and resting T cells had similar levels of
MAPK phosphorylation to wild type cells, but had reduced PKB phosphorylation. This
finding suggests that signaling downstream of Ras was intact in GlcNAc-TV deficient
cells, but that signaling through PI3K may have been inhibited. P I ' K catalyzes the
phosphoryIation of inositol lipids at the 3' position on the inositol ring, resulting in the
formation of 3-phophorylated phosphoinositides (PtdIns) (Leevers et al., 1999).
Phosphorylated PtdIns are second messenger molecules that participate in the
transduction of mitogenic signals from activated tyrosine kinase receptors and of signals
mediated by integrin engagement. Phosphorylated PtdIns are necessary for the activation
of PKB. The binding of PtdIns(3,4)Pz to the plextrin homology (PH) domain of PKB
results in the tramIocation of PKB to the plasma membrane where it is phosphorylated by
phosphoinositide-dependent ThrISer kinases. Therefore, PKB phosphorylation is a
measure of PI3K activity (Leevers et al., 1999; Wyrnann and PiroIa, 1998).
PI3K activity transduces signais downstream of receptor tyrosine kinases as well
as engaged integrins. The addition of serum to cultured M ~ U ~ Z " fibroblasts, and the
mitogenic stimulation of ~ ~ a t 5 ' - T cells resulted in nomai PKB phosphorylation,
indicating that stimulation of PI3K activity through receptor tyrosine kinases was intact.
However, phosphorylation was reduced in ~ ~ i z t . ! ? cells in serum free conditions,
consistent with the cytoskeletai morphology of the cells, and confirming than integrin
signaling is cornpromi sed.

153
FIGURE 10: The effect of GlcNAc-TV deficiency on MAPK and PKB
phosphorylation.
A) Levels of phosphorylated and total Akt/PKB, and of phosphorylated MAPK in T cells
following addition of serum. B) Levels of phosphorylated and totd Akt/PKB, and of
p hosphorylated MAPK in tumor cells from P ~ M T M ~ ~ ~ . ? . P ~ M T M ~ u ~ s * " and
P ~ M T M ~ ~ ~ F ' - fast growing tumors.
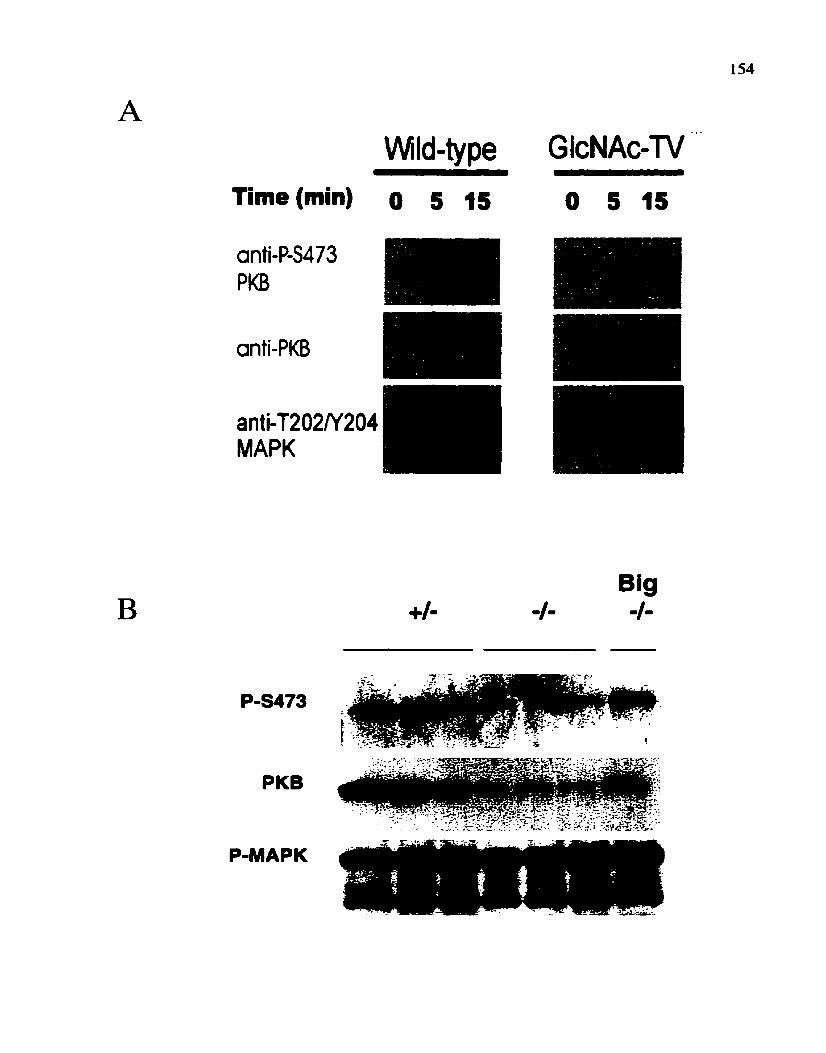
anti-P-S473 PKB
Big +/- -1- -1-
PKB

155
PyMT binds and activates Src and PI3K, and therefore it is possible that tumor
cells are Iess dependent on integrin engagement to initiate signaling cascades leading to
ceIl motility, as indicated by the translocation of paxillin to focal adhesions. However,
our results suggest that PyMT tumor cells remain at least partially dependent on focal
adhesion turnover, requiring GlcNAc-TV activity. In PyMT transformed cells, paxillin
localized to focal adhesions in cells of al1 genotypes (this chapter, Figure 9). However,
~ ~ a r . 5 ' - cees failed to undergo membrane mffling, a necessary step for ce11 motility.
Rather, P~M'T'M~U~S'- cells maintained heavy actin stress fibers suggesting little tumover
of actin and focal adhesions. In contrast, the P~MT'M~~ZY- tumor cells show membrane
ruffling and fine actin filaments. A recent study suggests that paxillin phosphorylation in
response to integrin engagement is partly dependent on PI3K activity. W e have observed
that Mgat5'- tumors have hypophosphorylated PKB, and preliminary
immunoprecipitation experiments show reduced phosphorylation of paxillin as well.
These results indicate that P U K activity is inhibited in ~ ~ a t 5 ' - tumors similarly to the
inhibition obsewed in ~ ~ a t ~ ' - ~ u i e s c e n t fibroblasts and T-cells.
The lack of membrane ruffling and the hypophosphorylation of PKB in
P ~ M T M ~ ~ ~ F " tumor cells suggest that signaling by the Rac and Cdc42 GTPases may be
impaired. Membrane mffling requires the activation of Cdc42 downstream of integrin
engagement. Activated Cdc42 associates with PI3K and stimulates its activity (Reif et
al., 1996), which, in turn, is required for the activation of Rac. PI3K-dependent
activation of Rac results in membrane mffling (Aspenstrom, 1999). Fast growing
P ~ M T M ~ ~ ~ F " tumors have comparable PKB phosphorylation levels t o w iId type P yMT
tumors, and their growth rate is similar to wild type tumors as well. However, cells from
the fast growing P ~ M T M ~ ~ ~ Y - tumon still exhibit considerably Iess membrane ruffling
than wild type PyMT tumor cells do, indicating that the altered growth and cytoskeletal

156
rearrangement are separate and distinct characteristics of the ~ ~ a t . 5 ' - tumor ce11
phenotype. These findings suggest that integrin signding leading to Cdc42 activation
may remain impaired in the fast growing P ~ M T M ~ ~ ~ Y - tumon, even though a secondary
mutation restored PI3K activity to wild type levels, and restores wild type-like growth
rates.
Fast-gmwing ~ ~ a t s / - tumors ntoy be the result of a somattœc rnutnlion: The infrequent
appearance of fast-growing ~~at .5- ' - tumors, which appear to arise from slow growing
Mgat.!? tumors, suggests the occurence of rare mutations. The effect of such mutations
is to release cells from the growth inhibition imposed by GlcNAc-TV deficiency. As in
the case of the mice transgenic for mutant P y m , one such possible secondary mutation
would be the upregulation of the ErbB receptors, which are known to enhance PI3K
activity and Shc phosphorylation. Such a scenario may also enhance Ras signaling, and
therefore result in enhanced expression from the Mgat.5 locus in an Ets-2 and APL
dependent rnanner. The upregulation of Lac2 expression in the fast growing tumors is
consistent with this model,
The fast-growing ~ ~ a t 5 ' - tumors do not give rise to a higher incidence of
metastases. This observation suggests that metastatic potentiai and tumor growth rates
are separate and distinct phenotypic features of the ~ ~ a t ~ ~ mouse. Therefore, it is
possible that the secondary mutation, sustained by the cells giving rise to fast-growing
tumors, is sufficient to compensate for the proliferative deficiency but not for the
metastatic deficiency caused by Mgut5 ablation. An amplification of intracellular
signaling may drive proliferation and survivai, while the deficiency in pl,6GlcNAc-
branched N-glycans continues to restnct focal adhesion turnover and thereby physical
movement of the tumor cells required for invasion and metastasis.
Autonomous vs host-dependent factors in tumor progression: Glycosylation can

157
potentially affect tumor growth in both tumor-autonomous and host-dependent manner.
Wild type ES cells were injected S.C. into rnice of al1 Mgar.5 genotypes to investigate the
role of host GlcNAc-TV activity on tumor formation and growth. ES cells injected into
~ ~ a t 5 " and ~ ~ a t T ' ' + rnice produced tumors in lm% of the mice. However,
surprisingly, none of the ~ ~ a t 5 + ' - rnice developed tumors. Latency was longer in Mgat.5
/- rnice, but once tumors formed, their growth rate was significantly greater, suggesting
that host factors depend on GlcNAc-TV activity for tumor growth. A similar dependence
on proper glycosylation was noted in GlcNAc-ï?D deficient mice. Hepatocarcinomas
resulting from diethylnitrosarnine treatment progress more slowly in ~ ~ a t 3 - ' - mice
compared to wild type littermates. However, Mgat3 is not expressed in mouse
hepatocarcinoma cells, suggesting that a host paracrine effect, such as the full function of
a growth factor, or a growth factor receptor, is dependent upon GlcNAc-TIII
gl ycosylation to promote tumor growth (Bhaumik et al., 1998).
This experiment suggests that GlcNAc-TV deficiency in the host exerts an
opposite effect on tumor growth rates than GlcNAc-TV deficiency of tumor cells. In
addition, the differences in latency and growth rates between tumors in M ~ u ~ S * + and
~ ~ a t 5 ' - mice suggest that GlcNAc-TV rnay regulate more than one aspect of host
perrnissiveness for tumor progression. Obvious candidates for further investigation
incIude host immune responses to the ES cells, and the angiogenic potential of the MgatS-

METHODS
Cenerotion of ~ ~ r r f ' - P y ~ T ' mke: ~ g a t s ~ mice were generated as described in
Chapter 3. ~ ~ a t 9 ' - female mice on 1291Sv background were mated with a male mouse
trangenic for MMTV-PyMT on FVB background. Offspring were screened for the
presence of the PyMT gene and their Mgar5 genotype was identified by PCR. PCR
primers for PyMT were S'GGAAGCAAGTACTTCACAAGGG3' and
5'GGAAAGTCACTAGGAGCAGGG3' and generated a 530 bp fragment. For Mgat5
penotyping, the primer pair S'GTAAGGACTCACAGCTGAGG3' and
S'GCCAAGGGAATGGTACATTGC3' was used to amplify a wild type-specific
fragment and the primer pair S'CCCATCTACACCAACGTAACC3' and
5'CGCCACATATCCTGATCTTCC3' was used to amplify a mutant-specific fragment.
The wild-type primers amplified a 446 bp fragment and the mutant primers amplified a
320 bp fragment.
Northern analysis of PyMT expression: Mice bearing mammary tumors were sacnficed
and the tumors were snap frozen. RNA was isolated using Trizol (Gibco-BRL) according to
manufacturer's instxuctors. RNA was size fractionated on a 1% formamide-agarose gel and
blotted ont0 Genescreen membrane. Radiolabeled PyMT cDNA (1.5 kb) containing the
entire coding region, was used as probe.
Deteetion and measurement of tumors and metastatic foci: Turnors were detected by
palpating each mammary fat pad weekly from the age of 6 weeks. Once tumors became
palpable, two measurements 1 tumor 1 week were done using calipers. Mice were
sacrificed when tumor burden reached 50% of body weight, or when tumors became
ulcerated, and lungs were resected and surface metastatic foci were counted visually.
HistologicaI exanzincdion of tunaors: Tumors were fixed in 10% phosphate-buffered

159
formalin, blocked in parafin, sectioned at 8 pm and stained with hematoxylin and eosin.
Wholemount preparations of marnmary fat pads were done as previously described Briefly,
the fat pads were spread on a glass slide and air-drïed. The slides were fixed ovemight in
Carnoy's solution (75% ethanol, 25% glacial acetic acid) and stained with Carmine alum
stain (02% w/v Carmine dye, 0.5% w/v aluminum). Staining of sections for apoptosis by
Klenow labeling of fragmented DNA and for proliferation by PCNA
irnrnunohistochemistry, were done as previously described (Fata et al., 1999).
Detection of f3-galactosidase achvity :
Marnmary fat pads were fixed in 0.2% glutaraldahyde for 30 minutes, washed in
PBS, and incubated overnight with X-gal staining solution (lmg/ml4-chloro-5-bromo-3-
indolyi-fLgalactoside(X-gal), 4mM We(CN)&HzO, 4mM &Fe(CN)6, 2mM MgCl?,
0.0 1 % deoxycholate, 0.02% Nonidet-P40 in O. lM sodium phosphate pH7.3). After
staining, tumors were further fixed in 10% phosphate-buffered formalin.
ImmunoJIuorescent microscopy : To examine focai adhesions, embryonic fi broblasts and
primary tumor cells were allowed to attach ovemight to glass slides coated with 1 pg/d of
fibronectin in serum free alpha-MEM. The cells were stained with hodamine-phalloidin,
anti-paxillin antibody (Transduction Laboratones), and Hoechst 33258 stain according to
manufacturer's instructions. The anti-paxillin antibody was detected with a FITC-
conjugated anti-rabbit antibody. Fluorescence images of the cells were obtained using a
decovolution microscope and digital capture.
ES-ce11 teraiocarcinornas: 2~ l d wi Id type R 1 ES cells were injected S.C. into the left hind
leg of 4 Mgar5+'+ 4 ~ ~ a t . 5 ' - and 5 Mgatf* mice. Mice weR exarnined for palpable tumors on
a weekly basis, and once tumors were detected, they were measured with cdipers weekly.

Conclusions and Future Directions

Structure-function relationships of specific glycan classes can be elucidated by
targeted ablation of key enzymes in the biosynthetic pathways of glycosylation.
Although glycosylation is a pst-translational modification that affects many cell-surface
and secreted polypeptides, and therefore its influence is pleotropic, specific biological
functions have k e n ascribed to carbohydrate structures by studies in knockout mice.
Unlike GlcNAc-TI and GlcNAc-TII, which are ubiquitiously expressed, GlcNAc-TV is
tempomlly and spatially regulated during embryogenesis and in cancer progression. The
pattern of expression of GlcNAc-TV implies a possible functional specificity for
1,6GlcNAc-branched N-glycans. Indeed, the phenotypes observed in the ~ p t . 5 ' '
mouse occur in ce11 populations expressing Mgat.5 in wild type mice. GlcNAc-TV
deficiency leads to a behavioral abnormality, T ce11 hypersensitivity and changes in
leu koc yte migration and adhesion. In addition, as predicted from studies with somatic
tumor ce11 mutants with reduced GlcNAc-TV activity and p1,6GlcNAc-branched N-
glycans. ~ ~ a t 5 ' - mice develop mammary tumors with greater latency, slower growth and
show >20 fold reduction in metastasis. Our resutts suggest that GlcNAc-TV may
regulate processes common to many ce11 types, such as ce11 motility and adhesion to
ECM and receptor aggregation and related signaling.
GlcNAc- TV deficiency affects materna1 behavioc GIcNAc-TV de ficienc y does not
result in obvious developmental defects, as M~u~z' ' mice are viable and fertile.
However, 1 Wsv GlcNAc-TV deficient females fail to nurse their Young. This phenotype
is most likely due to a behavioral defect. The behavior is strain dependent, suggesting
that GlcNAc-TV may be a modifier of some other gene product, such as a signaling
receptor. 129/sv females exhibit poorer matemal skills than CD1 or C57/BL-6 females
and therefore GlcNAc-TV deficiency may enhance an existing phenotype. Since the

162
aberrant materna1 be havior disappears in mixed 1291s~-CD 1 or 1291s~-C571BL-6
females, it may be possible to identify the gene interacting with Mgat.5 to cause the
aberrant nurtunng behavior.
GlcNAc-TV in imrnuniîy: The data presented in Chapter 3 supports a role for GlcNAc-
TV activity in the regdation of T ce11 proliferation and in leukocyte migration. We have
shown that GlcNAc-TV deficiency results in hyperproliferation of T cells and in
decreased migration of leukocytes in response to stimuli such as thioglycollate and
arachidonic acid. Application of imtants causing either a T-ce11 dependent DTH or an
imrnediate-type h ypersensitivity resulted in exaggerated inflammatory responses in
GIcNAc-TV deficient mice. In both models, the recovery time was longer in the
GlcNAc-TV deficient mice, suggesting that the inflammatory responses were not
darnpened as efkiently as in wild type rnice. We have also demonstrated activation-
dependent upregulation of Mgat.5 and L-PHA reactive structures in T cells. Together
these data suggest that pl,6GlcNAc-branched N-glycans may function to dampen the T
ceIl response following antigen-mediated activation of the cells.
Infiammatory responses are tenninated by the apoptosis of the immune cells.
Overexpression of GlcNAc-TV in MvLul cells resulted in increased apoptosis, and
therefore it is possible that the upregulation of GlcNAc-TV activity in activated T cells is
necessary for the efficient apoptosis of these cells. ~ ~ a t S / - T cells may be deficient in
their abiiity to apoptose after antigen-dependent stimulation, which may result in the
increased severity of the inflammatory response obsewed in the GlcNAc-TV deficient
mice. In this manner, the function of GlcNAc-TV activity may be sirnilar to the function
of CTLA-4, which is expressed after the activation of T cells (Lee et al., 1998). CTLA-4
has a higher affinity for the B7 molecules, which are the stimulatory ligands for CD28.
Thus, once CTLA-4 is activated, it downregulates T ceIl responses by outcompeting

163
CD28 for the B7 ligands, while physicaily associating with the TCR and directly
inhibiting TCR signaling. Lack of CD28 signaling results in failure to produce IL-2,
which protects T cells from apoptosis. CTLA-4 deficient mice develop severe
autoimrnunity and die 2-3 weeks after birth (Khattrï et al., 1999). Although the
phenotype of GlcNAc-TV deficient mice is much milder, it is possible that these mice
could develop an autoimmune condition if stressed appropnately. Induced T ce11
dependent multiple sclerosis is a model of autoimmunity now k ing explored in the lab.
The ability of ~ ~ a r 5 " mice to clear viral or parasitic infeaion might also be
different than in wild type mice. We are currently explonng different models of
infection. The Friend leukemia virus is a well-established model for studying genetic
resistance to an immunosuppressive munne virus (Hasenkrug and Chesebro, 1997), and
Schistosomes infection has k e n studied extensively as a model of an immunosuppressive
parasitic infection. Cytotoxic T cells, T helper cells, Th1 and Th2 responses, as well as
antibody responses are required for clearance of infection, varying in importance
depending on the pathogen. Therefore, several models will be used to study these aspects
of immunity in the GIcNAc-TV deficient mouse.
Preliminary studies in the GlcNAc-TV deficient mice suggest that these mice may
produce aberrant cytokine levels when challenged, or that their leukocytes may respond
aberrantly to cytokines. Thioglycollate, an inflamrnatory agent, failed to produce a
significant neutrophil and macrophage infiltration when injected into the peritoneum of
~ ~ a r 5 ' - mice. Thioglycollate stimulates the synthesis of tumor necrosis factor n (TNF-
a) by macrophages and inhibits arachidonic acid production and metabolism. Therefore,
Our experiments suggest that while the GlcNAc-TV deficient mice rnay be hypersensitive
to arachidonic acid metabolites, the initiai delay in leukucyte migration may inhibit the
production of other inflamrnatory cytokines, such as TNF-a, resulting in a mild and

164
aborted response to thioglycollate injection. This hypothesis is supported by a
prelirninary study of hepatic injury induced by injection of carbon tetrachloride (CCh), in
which GlcNAc-TV deficient mice survived better than wild type littermates. Previous
studies implicate interleukin-6 (TL-6), IL-10 and TNF-a in hepatic inflammation induced
by CCIa. IL-IO deficient mice displayed elevated levels of TNF-a, TGF-pl and IL-6,
resulting in an acute inflarnmatory burst. Slowed migration of leukocytes into the liver in
GlcNAc-TV deficient mice would inhibit the acute inflarnmatory burst that causes
rnondity in this model. However, the difference in the response of GlcNAc-TV deficient
mice to CClj could also be due to the levels of one or severai of these cytokines, their
acti vity, or the activity of their receptors k i n g aberrant in these rnice.
ClcNAc-TV deJiciency causes delayed hrmor progression and inhibits metastusis: We
crossed GlcNAc-TV deficient mice with MMTV-PyMT mice to study metastatic breast
cancer. PyMT interacts with Src, Shc and PI3K. Src and Shc both act in the Ras
pathway, which has been previously shown to activate MgalS. Our results show that
GlcNAc-TV activity is necessary for efficient tumor progression and for metastasis.
Tumor growth was retarded in ~ ~ a t . 5 ' - mice, and metastasis was inhibited by s20 fold.
Infrequent ad- tumon exhibited altered growth rates 3-4 weeks after they became
palpable to resemble wild type growth, suggesting that a second mutation occurred in
these tumors. Western analysis of primary T cells and embryonic fibroblasts shows that
PKB phosphorylation, an indicator of PI3K activity, is reduced in Mga& cells.
Preliminary results from Western analysis of PyMT tumors suggest that Mgats/- nul1
tumors aiso have reduced phophorylation of PKB compared to Mgat9'- tumors.
However, PKB phosphorylation levels in fast-growing ~ ~ a t 5 ' - tumors resemble
phophorylation levels of MgatTi- tumors. Thus, it is likely that GlcNAc-TV deficiency

165
results in penurbation of the PI3KIPKB signaling pathway, which may be
counterbalanced by a mutation in the fast-growing tumors.
The fast-growing tumors did not cause an increase in the number of metastases,
suggesting that tumor growth and metastatic potential may be separate and distinct
phenotypic characteristics of the ~ g a t 5 ' - mice. Therefore. it is possible that even though
signaling through both the Ras and the PI3K pathways is sirnilar to wild type in these
tumors, Iack of Bl,6GlcNAc-branched N-glycans still impedes integrin mediated focal
adhesion turnover. The specificity of the defect is underscored by the efficient turnover
of actin in activated ~ ~ a t ~ " T cells, which is directed by TCR and CD28 engagement
and not by integrin engagement. The hypothesis that integrin signaling is impaired in
~ ~ a r 5 ' - fibroblasts and in P ~ M T M ~ U ~ S " tumor cells primarily through the Rho family of
GTPases, could be tested directly by rescue experiments in which the activated Cdc42,
Rac and Rho constructs would be transfected into these cells.
We have shown that bl,6GlcNAc-branched N-glycans have a function essentiai
for efficient metastasis, possibly independent from receptor signaling modification. This
function may relate to physically aiding in focal adhesion turnover, or to extravasation
and organ colonization that may be mediated by interactions with lectins. Candidates for
binding partners of B1,6GlcNAc-branched N-glycans could be the widely expressed
galectins, which bind N-acetyllactosamine and have been implicated in tumor ceIl
adhesion during metastatic spread. If the polylactosamine content of ~ g a t y ' tumor cells
is reduced, it would mean less ligand for the galectins, and therefore less eEcient
colonization by metastatic cells. Structural analysis of N- and O-glycans is required to
determine how the ~ ~ a t 5 ' - mutation affects the distribution of residual structures.
Mgut5 expression may requùe signding through both the Ras and the PI3K pathways
and may be necesscug for proper integrin signaling: The expression of lacZ in nul1

166
tumors suggests that the activation of the Ras pathway alone may not be sufficient to tum
on expression from the MgatS locus. Preliminary results show that the level of MAPK
phosphorylation in P ~ M ' T M ~ ~ Z Y - is sirnilar to that of ~ y M T ' ~ ~ a t p and P ~ M T M ~ U Z ~
tumors. However, expression of lac2 is downregulated in cornparison to expression in
the ~~at .S+'' tumors. In the rare fast growing ~ y M T ' ~ ~ a t 5 ' - tumors, PKB
phosphorylation, as well as lac2 expression is comparable to that observed in
P ~ M T M ~ ~ Y ' - and P ~ M T M ~ u ~ ~ « + tumors. Therefore, these results suggest that Mgat5
expression is regulated by both the Ras and the PUK signaling pathways. This scenario
is similar to the one presented in a recent report showing that the transcription factor
AFX (a member of the forkhead family) requires direct phosphorylation by both PKB, a
PI3K effector, and by the Ra1 GTPase, an effector of Ras s i g n a h g (Kops et al., 1999).
The results presented in this thesis suggest the following working mode1 (Figure
1). Activation of receptor tyrosine kinases initiates signaling through the Ras and PI3K
pat h ways, w hich results in upregulation of Mgar5 gene expression and increased
fi 1,oGlcNAc-branched N-glycan biosynthesis. Thus, the $1,6GlcNAc-branched N-
glycans on adhesion and cytokine receptors increase. Signaling initiated by integrin
engagement is enhanced by $1,6GlcNAc-branching of N-glycans due to their effect of
promoting receptor cluster turnover. Integrin engagement aiso results in the activation of
Ras and P13K pathways, leading to cytoskeletal reorganization, proliferation and
migration. Further signaling through the Ras pathway, and the PUK, amplify MgatS
transcription and f% 1 -6GlcNAc-branched N-glycan synthesis, creating positive feedback.
In the absence of Mg&, integrin signaling is compromised, as indicated by PKB
hypophosphorylation and reduced microfilament and focal adhesion turnover. In
addition, we have shown that the positive feedback is disrupted in ~ y M T ~ ~ a t 5 ' - tumor
cells. P~MTM~~ZY- tumor cells have similar level of MAPK phosphorylation to wild

167
type PyMT tumor cells, indicating that the Ras pathway is intact in these cells. However,
~ ~ h 4 T ~ ~ a t . F ' - tumors have reduced lac2 expression from the Mgat5 locus, a result that is
not consistent with MgatS expression k ing dependent on the Ras pathway alone. The
dependence of Mgat5 expression on PI3K signaiing is further illustrated by the enhanced
lac2 expression in fast growing ~~h4T'~~at.T'- tumors, where PKB phosphorylation are
restored to wild type levels.
Recently, several papers have highlighted the importance of integrin signding in
T ce11 activation. Signaling mediated by LFA-1, the lymphocyte adhesion receptor for
ICAM-1, was shown to be necessary for cytoskeletal rearrangement and TCR
intemalization (Grakoui et al., 1999; Bachmann et al.,1997). In our experiments, aberrant
cytoskeletal remangement was observed in ~ ~ a t 5 ' - T cells, fibroblasts and P y m tumor
cells. It is therefore possible that the glycoproteins affected most cnticaily by
pl ,6GlcNAc-branched N-gl ycosylation are the integins, and that the phenotypes
observed in the ~ ~ a t ~ ' - mouse are due to a common defect, the aberrant hinction of
integins.
The biologrgrcaI significance of GIcNAc-TV activity may be investigated in w o m : As
demonstrated by this work and by work with other glycosyltransferase-deficient mice,
some glycosylation effects are ce11 nonautonomous and therefore must be studied in a
whole organism. Although a great deai of information can be obtained from studies with
mice, these marnrnals are very cornplex. Enhancer/suppressor screens to identify gene
products modified by glycosylation are practically dificult and expensive in mice. C.
elegans provides a genetically tractable and simpler alternative. The components of the
Ras and the PI3K signaling pathways are consewed in C. elegans, as is GlcNAc-TV.
Mutations in the let genes (the worm Ras orthologues) lead to migration defects in the
sex myoblast cells. Mutations in the P13K orthologue (agel) result in altered longitivity.

168 Therefore, screens for enhancer/suppressor mutations on gly-2 (the worm GlcNAc-TV
gene) or other glycosyltransferase deficient Worms would be useful in identifying key
substrates of glycosylation, which may aid in further understanding the moiecular basis
for the mammalian phenotype.

169
Figure 1 Schematic of hypothesis for GlcNAc-TV function in cell adhesion.
A) Signaling through receptor tyrosine kinases initiates signaling through the Ras and
PI3K pathways, which result in upmgulation of Mgut5 expression and increased
B 1,oGlcNAc-bnnched N-glycans. Thus, the B 1,6GlcNAc-branched N-glycans on
adhesion and cytokine receptors are increased. Signaling initiated by a& integrin
engagement is enhanced by p1,6GlcNAc-branching N-glycans and results in the
activation of F M - S r c and of PUK. The activation of these pathways leads to
cytoskeletal reorganization, prolifention, migration and enhancement of signaiing
through the Ras and the PWK pathways, which in turn amplifies Mgut5 transcription and
f3 1,oGlcNAc-branched N-glycan synthesis, creating positive feedback. B) In the absence
of Mgat5, signaling initiated by integrin adhesion to ECM is compromised, possibly by
physicall y inhibiting focal adhesion formation and turnover. Integrin Signaiing through
the PUK is affected in embryonic fibroblasts and in P ~ M T M ~ ~ ~ ~ ' - tumor cells, and the
cytoskeletal organization of P~MTM~~ZS ' - tumor cells indicates that signaling through
Cdc42, positioned downstream of integrins and upstrearn of PI3K is compromised. The
disruption of positive feedback in GlcNAc-TV deficient cells is evident by the reduced
expression of lacZ from the Mgat5 locus in the ~ y M T ~ ~ a t 5 ' - tumors, where PKB is
h ypophosphorylated.

3PPIs ~ R a c i ! ! \
4 I Shc+Sos
reorganization ,
p 1,6GlcNAc-branched N-glycan synt


List of References
Akiyama, S.K., Yamada, S.S., and Yamada, K.M. (1989). Analysis of the role of glycosylation of the human fibronectin receptor. J.Bio1.Chem. 264, 1801 1-180 18.
Alton, G., Kjaergaard, S., Etchison, J.R., Skovby, F., and Freeze, H.H. (1999). Oral ingestion of mannose elevates blood mannose levels: a first step toward a potential therapy for carbohydrate-deficient glycoprotein syndrome type 1. Biochem.Mol.Med. 60, 127-133.
Asano, M., Fumkawa, K., Kido, M., Matsumoto, S., Umesaki, Y., Kochibe, N., and Iwakura, Y. (1997). Growth retardation and early death of beta-1,4- galactosyltransferase knockout rnice with augmented proliferation and abnormal differentiation of epithelial cells. EMBO J. 16, 1850- 1857.
Aspenstrom, P. (1999). The Rho GTPases have multiple effects on the actin cytoskeleton. Exp. Ce11 Res. 246,SO-25.
Babal, P. and Russell, L.C. (1999). Sidic acid-specific lectin-mediated adhesion of Tritrichomonas foetus and Tritrichomonas mobilensis. J-Parasitol. 85, 33-40.
Bachmann, M.F., McKall-Faienza, K., Schmits, R., Bouchard, D., Beach, J., Speiser, D.E., Mak, T.W., and Ohashi, P.S. (1997). Distinct roles for LFA-1 and CD28 during activation of naive T cells: adhesion versus costimulation. Irnmunity 7, 549-557.
Bajorath, J., Metzler, W.J., and insley, P.S. (1997). Molecular modeling of CD28 and three-dimensional analysis of residue conservation in the CD28/CD 152 family. J Mol Graph Mode1 15, 135-139.
Baum, L.G., Pang, M., Perillo, N.L., Wu, T., Delegeane, A., Uittenbogaart, C.H., Fukuda, M., and Seilhamer, J.J. (1995). Human thymic epithelid cells express an endogenous lectin, galectin-1, which binds to core 2 O-glycans on thymocytes and T lymphoblastoic cells. J.Exp.Med. 181, 877-887.
Bell, J.E., Beyer, T.A., and Hill, R.L. (1976). The kinetic mechanism of bovine milk galactosyltransferase. The role of alpha-lactalbumin. J Bi01 Chem 251,3003- 3013.
Beuth, J., Ko, H.L., Oette, K., Pulverer, G., Roszkowski, K., and Uhlenbruck, G. (1987). Inhibition of liver metastasis in mice by blocking hepatocyte lectins with arabinogaiactan infusions and D-galactose. J-Cancer Res.Clin.Oncol. 1 13, 5 1-55.
Bhaumik, M., Harris, T., Sundararn, S., Johnson, L., Guitenplan, J., Rogler, C., and Stanley, P. (1998). Progresson of hapatic neoplasms in severely retarded in mice

lacking the bisecting N-acetylglucosamine on N-glycans: evidence for a glycoprotein factor that facilitates hepatic tumor progression. Cancer Res. 58, 288 1-2887.
Biancone, L., Araki, M., Araki, K., Vassalli, P., and Stamenkovic, 1. (1996). Redirection of turnor metastasis by expression of E-selectin in vivo. J.Exp.Med. 183,58 1- 587.
Bierhuizen, M.F.A. and Fukuda, M. (1992). Molecular coloning of cDNA encoding UPD-GICNAC:G~IS 1->3 GalNAc->R (GlcNAc to GalNAc) 1-6 N- acetylglucosarninyltransferase, and O-glycan branching enzyme, by gene transfer into CHO cells expressing polyoma large T antigen. Proc.Nat1.Acad.Sci.USA 89, 9326-9330.
Birchmeier, W. (1995)- E-cadhenn as a tumor (invasion) suppressor gene. BioEssays 17, 97-99.
Bird, J.M. and Kimber, S.J. (1984). Oligosaccharides containing fucose tinked alpha ( l - 3) and alpha ( 1 4 ) to N-acetylglucosamine cause decompaction of mouse morulae, Dev.Bio1.104,449-460.
Blaikie, P.A., Fournier, E., Dilworth, S.M., Bimbaum, D., Borg, J-P, and Margolis, B. (1997). The role of the Shc phosphotyrosie interaction/phosphotyrosine binding domain and tyrosine pheosphorylation si tes in pol yoma middle T antigen- mediated ceIl transformation. J Bi01 Chem 272,2067 1-20677.
Boniface, J.J., Rabinowitz, J.D., Wulfing, C., Hampl, J., Reich, Z., Altman, J.D., Kantor, R.M., Beeson, C., McConnell, H.M., and Davis, M.M. (1999). Initiation of signal transduction through the T ce11 receptor requires the multivalent engagement of peptide/MHC ligands. Immunity 9,459-466.
Boren, T., Norrnark, S., and Falk, P. (1994). Helicobacter pylori: molecular basis for host recognition and bacterial adherence. Trends Microbiol. 2,221-228.
Botti, C., Seregni, E., Ferari, L., Martinetti, A., and Bombardien, E. (1998). Immunosuppressive factors: d e in cancer development and progression. 1nt.J.Biol.Marker-s 13, 5 1-69.
Braga, V.M., Pemberton, L.F., Duhig, T., and Gendler, S.J. (1992). Spacial and tempera1 expression of an epithelial mucin, Muc- 1 during mouse development. Development 115,427437.
Braut-Boucher, F., Font, J., Pichon, J., Paulin, Y., Boukhelifa, M., Aubery, M., and Derappe, C . (1 998). T lymphocytes from sezary syndrome patients express betal integrins whose beta( 1-6)-branched N-linked oligosaccharides reflect their adhesive capacity. L,euk.Res. 22,947-952.
Bresalier, R.S., Niv, Y., Byrd, J-C., Duh, Q.Y., Toribara, N.W., Rockwell, R.W., Dahiya, R., and Kim, YS. (1991). Mucin production by human colonic carcinoma cells correlates with their metastatic potential in animal models of colon cancer

174
metastasis. J.Clin.invest. 87, 1037-1045.
Brown, J.R., Ye, H., Bronson, R.T., Dikkes, P., and Greenberg, M.E. (1996). A defect in nurtunng in mice lacking the irnmediate early gene fosB. Ce11 86,297-309.
Buckhaults, P., Chen, L., Fregien, N., and Pierce, M. (1997). Transcriptional regulation of N-acetylglucosaminyltransferase V by the src Oncogene. J.Biol.Chem. 272, 19575- 1958 1.
Bugge, T.H., Lund, L.R., Konbrinck, K.K., Nielsen, B.S., Holmback, K., Drew, A.F., Flick, M.J., Witte, D.P., Dano, K., and Degen, J.L. (1999). Reduced metastasis of polyoma virus rniddle t antigen-induced marnmary cancer in plasminogen- deficient mice. Oncogene 16,3097-3 104.
Bunnell, B.A., Adams, D.E., and Kidd, V.J. (1990). Transient expression of p58 protein kinase cDNA enhances mammalian gl ycosyltransferase activi ty. Biochem.Biophys.Res.Cornmun. 171, 196-203.
Cairns, J. (1981). The origin of human cancers. Nature 289, 353-358.
Campbell, R.M., Metzler, M., Granovsky, M., Dennis, J.W., and Marth, J.D. (1995). Complex asparagine-linked oligosaccharides in Mgarl-nul1 embryos. Glycobiology 5, 535-543.
Carrick, J.B., Moore, J.N., Chapkin, R.S., and Schnellmann, R.G. (1995). Increased phospholipid mass with decreased arachidonoyl molecular species is associated with decreased eicosanoid synthesis and increased tumor necrosis factor synthesis by thiogl ycollate-elicited rat peritoneal macrophages. Shock 3,284-29 1.
Carlsson, S.R., Saski, H., and Fukuda, M. (1986). Structural variations of O-linked ~Iigosaccharides present in leukosialin isolated from erythroid, myeloid, and t- Iymphoid ceIl lines. J.Biol.Chem. 261, 12787-12795.
Charnmas, R., Veiga, S.S., Travassos, L.R., and Brentani, R.R. (1993). Functionally distinct roles for glycosylation of a and f3 integrin chains in cell-matrix interactions. Proc.NatI.Acad.Sci.US A 90, 1795- 1799.
Chao, M.V. (1992). Growth factor signaling: where is the specificity? Ce11 68,995-997.
Chen, L., Zhang, W., Fregien, N., and Pierce, M. (1998). The her-Zneu oncogene stimulates the transcription of N-acetyIglucosaminyltransferase V and expression of its cell surface oligosaccharide products. Oncogene 17,2087-2093.
Cheng, A.M., Saxton, T.M., Sakai, R., Kulkarni, S., Mbamalu, G., Vogel, W., Tortonce, C.G., Cardiff, R.D., Cross, J-C., Muller, WJ., and Pawson, T. (1999). Mammalian Grb2 regulates multiple steps in embryonic development and malignant transformation. Cell 95,793-803.
Childs, R.A., Pennington, J., Uemura, K., Scudder, P., Goodfellow, P X , Evans, M.J., and Feizi, T. (1983). High-molecular-weight glycoproteins are the major carriers of the carbohydrate differentiation antigens 1, i and SSEA- 1 oi mouse

175
teratocarcinorna cells. Biochem.J. 215,491-503.
Christofon, G. and Semb, H. (1999). The role of the cell-adhesion molecule E-cadherin as a tumour-suppressor gene. Trends Biochem-Sci. 24,73-76.
Chui, D., Hennet, T., Moremen, K., Freeze, H.H., Paulson, J.C., Schachter, H., Fukuda, M.N., and Marth, J.D. (1999). Cell-type-specific and physiologie roles of oligosacc harides. Gl ycobiology 7, 10 19 Society for Gl ycobiology, Abstract 19.(Abstract)
Chui, D., Oh-Eda, M., Liao, Y.-F., Panneerselvam, K., Lal, A., Marek, K.W., Freeze, H.H., Moremen, K.W., Fukuda, M.N., and Marth, J.D. (1997). Alpha- mannosidase-II deficiency results in dyserythropoiesis and unveils an altemate pathway in oligosaccharide biosynthesis. Ce11 90, 157-167.
Clark, E.A. and Brugge, J.S. (1995). Integrins and signal transduction pathways: the road taken. Science 268,233-239.
Clark, E.A., King, W.G., Brugge, J.S., Symons, M., and Hynes, R.O. (1998). Integrin- mediated signals regulated by members of the rho farnily of GTPases. J Ce11 Bi01 142,573-586.
Clements, J.L., Boerth, N.J., Lee, J.R., and Koretzky, G.A. (1999). Integration of t ce11 receptor-dependent signaling pathways by adapter proteins. Annu Rev Immun01 17, 89-108.
Cornil, I., Kerbel, R.S., and Dennis, J.W. (1990)- Tumor ce11 surface pl-4 linked galactose binds to lectin(s) on microvascular endothelial cells and contributes to organ colonization. J.Cell Biol. 11 1,773-782.
Correll, P.H., Iwama, A., Tondat, S., Mayrhofer, G., Suda, T., and Bernstein, A. (1997). Increased interferon-gamma-mediated nitric oxide production, infiammation and susceptibility to endotoxic shock in mice lacking the STK receptor tyrosine kinase. Genes and Function 1, 1-15.
Cross, S.M., Sanchez, C.A., Morgan, C.A., Schimke, M.K., Ramel, S., Idzerda, R.L., Raskind, W.H., and Reid, B.J. (1995). A p53-dependent mouse spindle checkpoint. Science 267, 1353- 1356.
Cumberledge, S. and Reichsman, F. (1997). Glycosarninoglycans and WNTs: just a spoonful of sugar helps the signal go down. Trends Genet. 13,421-423.
Cummings, RD. and Komfeld, S. (1982). Characterization of the structural determinants required for the high affinity interaction of asparagine-linked oligosaccharides with imrnobilized Phaseolus vulnaris leukoagglutinating and erythroagglutinating lectin. J.Biol.Chem. 257, 1 1230-1 1234.
Cummings, R.D. and Kornfeld, S. (1984). The distribution of repeating Ga1 pl-4GlcNAc fi 1-3 sequences in asparagine-linked oligosaccharides of the mouse 1 ymphoma ce11 line BW5 147 and PHAR 2.1. J.Bio1.Chem. 259,6253-6260.

Cyster, J.G., Shotton, D.M., and Williams, A.F. (1991). The dimensions of the T lymphocyte glycoprotein leukosialin and identification of linear protein epitopes that can be modified by glycosylation. EMBO J. 10,893-902.
Datti, A. and Dennis, J.W. (1993). Regulation of UDP-G1cNAc:Gai~l-3GalNAc-R pl-6- N-acetylglucosaminyltransferase (GlcNAc to GalNAc) in Chinese hamster ovary cells. J.Biol.Chem. 268,5409-5416.
Delon, J., Gregoire, C., Malissen, B., Darche, S., Lemaitre, F., Kourilsky, P., Abastado, J.P., and Trautrnann, A. (1999). CD8 expression allows T ce11 signaling by monomeric peptide-MHC complexes. Lmmunity 9,467473.
Demetriou, M., Nabi, I.R., Coppolino, M., Dedhar, S., and Dennis, J. W. (1995). Reduced contact-inhibition and substratum adhesion in epithelial cells expressing GlcNAc- transferase V I JCell Biol. 130,383-392.
Denhardt, D.T. (1996). Oncogene-initiated aberrant signaling engenders the metastatic phenotype: synergistic transcription factor interactions are targets for cancer therapy. Crit-Rev-Oncog. 7, 261-291.
Dennis, J.W. (1986). Different metastatic phenotypes in two genetic classes of WGA- resistant tumor ce11 mutants. Cancer Res. 46,4594-4600.
Dennis, J.W. (1991). Changes in glycosylation with malignant transformation and tumor progression. In Ce11 Surface Carboh ydrates and Ce11 Development. M. Fukuda, ed. (J3oca Raton: CRC Press), pp. 16 1 - 194.
Dennis, J.W., Kosh, K., Bryce, D.-M., and Breitman, M.L. (1989). Oncogenes confemng metastatic potential induce increased branching of Asn-linked oligosaccharides in rat2 fibroblasts. Oncogene 4,853-860.
Dennis, J.W. and Laferte, S. (1986). Co-reversion of a lectin-resistant mutation and non- metastatic phenotype in murine tumor cells. 1nt.J.Cancer 38,445450.
Dennis, J.W. and Laferte, S. (1989). Oncodevelopmental expression of -GlcNAc pl- 6Man a l -6Man pl-6 branching of Asn-linked oligosaccharides in human breast carcinomas. Cancer Res. 49,945-950.
Dennis, J.W., Laferte, S., Fukuda, M., Dell, A., and Carver, J.P. (1986). Asn-linked oiigosaccharides in lectin resistant tumor ce11 mutants with varying metastatic potential. Eur.J.Biochem. 161, 359-373.
Dennis, J.W., Laferte, S., Waghome, C., Breitman. M.L., and Kerbel, R.S. (1987). 1-6 branching of Asn-linked oligosaccharides is directly associated with metastasis. Science 236,582-585.
Devchand, P.R., Keller, H., Peters, J.M., Vazquez, M., Gonzalez, F.J., Wahti, W. (1996). The PPARalpha-leukotriene B4 pathway to inflammation control. Nature 384,39- 43.

Dilworth, S.M., Brewster, C.E.P., Jones, M.D., Lanfrancone, L., Pelicci, G., and Pelicci, P.G. (1994). Transformation by polyoma virus middle T-antigen involves the binding and tyrosine phosphorylation of Shc. Nature 367,87-90.
Do, K.-Y., Fregien, N., Pierce, M., and Curnmings, RD. (1994). Modification of glycoproteins by N-acetylglucosaminyltransferase V is greatly influenced by accessibility of the enzyme to oligosacharide acceptors. J.Biol.Chem. 269, 23456-23464.
Dodson, J.M., Lenkowski, P-WJr., Eubanks, A.C., Jackson, T.F., Napodano, J., Lyerly, D.M., Loçkhart, L.A., Mann, B.J., and Petri, W.A.Jr. (1999). Infection and immunity mediated by the carbohydrate recognition domain of the Entamoeba histolytica GaUGalNAc Iectin. J.Infect.Dis. 179,46046.
Easton, E.W ., Bolscher, J.G.M., and van den Eijnden, DM. (199 1). Enzymatic amplification involving glycosyltransferases fonns the bais for the increased size of asparagine-linked glycans at the surface of MH 3T3 cells expressing the N-ras proto-oncogene. J.Biol.Chem. 266-2 1674-2 1680.
Eggens, I., Fenderson, B., Toyokuni, T., Dean, B., Stroud, M., and Hakomori, S. (1989). Specific interaction between L+eX and kX determinants. J.Biol.Chem. 264,9476- 9484.
Ellies, L.G., Tsuboi, S., Petryniak, B., Lowe, I.B., Fukuda, M., and Marth, J.D. (1998). Core 2 oligosaccharide biosynthesis distinguishes between selectin lignads essential for leukocyte horning and inflammation. h u n i t y 9,881-890.
Fata, J.E., Lao , K.J., Moorehead, R.A., Martin, D.C., and Khokha, R. (1999). Timp- 1 is important for epithelial proliferation and branching morphogenesis during mouse marnmary development. Dev Bi01 submizted,
Feizi, T. (1985). Demonstration by monoclonal antibodies that carbohydrate structures of glycoproteins and glycolipids are onco-developmental antigens. Nature 314,53- 57.
Fenderson, B.A., Eddy, E.M., and Hakomori, S. (1990). Glycoconjugate expression during embryogenesis and its biological significance. BioEssays 12, 173-179.
Fenderson, B.A., Zehavi, U., and Hakomori, S. (1984). A multivalent lacto-N-fuco penatose HI-lysyllysine conjugate decompacts preimplantation mouse embryos, while the free oligosaccharide is ineffective. J.Exp.Med. 160, 1591-1596.
Fernandes, B., Sagman, U., Auger, M., Demetriou, M., and Dennis, J.W. (1991). B 1-6 branched oligosaccharides as a marker of tumor progression in human breast and colon neoplasia. Cancer Res. 5 l ,7 18-723.
Ferrell, J.E.Jr. and Machleder, E.M. (1998). The biochernical basis of an All-or-none ce11 fate switch in Xenopus oocytes. Science 280,895-898.
Fiete, D., Srivastava, V., Hindsgaul, O., and Baenziger, J.U. (1991). A hepatic reticuloendothelial ce11 receptor specific for S044GalNAcp 1.4GlcNAc~ 1 ,Mana

that mediates rapid clearance of lutropin. Cell67, 1103-11 10.
Finne, J., Burger, M.M., and PrieeIs, J.P. (1982). Enzymatic basis for a lectin resistant phenotype: increase in a fucosyltransferase in mouse melanoma cells. J.Ce1l Biol. 92,277-282.
Fischer, K.-D., Kong, Y.-Y., Nishina, H., Tedford, K., Marengere, L.E.M., Kozieradzki, I., Sasaki, T., Starr, M., Chan, G., Gardener, S., Nghiem, M.P., Bouchard, D., Barbacid, M., Bernstein, A., and Penninger, J.M. (1998). Vav is a regulator of cytoskeletal reorganization mediated by the T-ce11 receptor. Curr-Biol. 8, 554- 562.
Frixen, U.H., Behrens, J., Sachs, M., Eberle, G., Voss, B., Warda, A., Lochner, D., and Birchmeier, W. (1991). E-Cadherin-mediated Cell-Cell Adhesion Prevents Invasiveness of Human Carcinoma Cells. J.Cel1 Biol. 113, 173-185.
Garcia, K.C., Degano, M., Stanfield, R.L.. Brunmark, A., Jackson, M.R., Peterson, P.A., Teyton, L., and Wilson, I.A. (1996). An alphabets T ceIl receptor structure at 2.5 A and its orientation in the TCR-MHC cornplex. Science 274, 209-219.
Gibson, S., Truitt, K., Lu, Y., LaPushin, R., Khan, HI, Imboden, J.B., and Mills, G.B. (1998). Efficient CD28 signalling leads to increases in the kinase activities of the TEC family tyrosine kinase E M T m S K and the SRC family tyrosine kinase LCK. Biochem-J. 330, 1123-1 128.
Gorelik, E., Xu, F., Henion, T., Anaraki, F., and Galili, U. (1997). Reduction of metastatic properties of BL6 melanoma cells expressing terminal fucose(a1pha)l- 2-galactose after alphal,2-fucosyltransferase cDNA transfection. Cancer Res. 57, 332-336.
Goss, P.E., Baker, M.A., Carver, J.P., and Dennis, J. W. (1995). Inhibitors of carbohydrate processing: A new class of anticancer agents. Clin-Cancer Res. I , 935-944.
Grady, W.M., Myeroff, L.L., Swinler, S.E., Rajput, A., Thiagalingam, S., Lutterbaugh, J.D., Neumann, A., Brattain, M.G., Chang, J.k.S.J., Kinzler, K.W., Vogelstein, B., Wilson, J.K., and Markowitz, S. (1 999). Mutational inactivation of transforming growth factor beta receptor type II in microsatellite stable colon cancers. Cancer Res. 59,320-324.
Granovsky, M., Bielfeldt, T., Peters, S., Paulsen, H., Meldal, M., Brockhausen, J., and Brockhausen, 1. (1994). UDPgalactose:glycoprotein-N-acetyl-D-galactone 3- beta-D-galactos yltransferase activi ty synthesizing O-gl ycan core 1 is controlled by the amino acid sequence and glycosylation of glycopeptide substrates. Eur.J.Biochem. 22 1, 1039-1046.
Granovsky, M., Cheng, A., Waterhouse, P., and Dennis, J.W. (1999). Lymphocyters of N-acetylglucosamin yltransferase V nul1 mice show h ypersensi tivi ty to T ce11 mitogens. Glycobiology 7, 1048 Society for Glycobiology, abstract

Granovsky, M., Fode, C., Warren, C.E., Campbell, R.M., Marth, J.D., Pierce, M., Fregien, N., and Dennis, J.W. (1995). GlcNAc-tramferase V and core 2 GlcNAc- transferase expression in the developing mouse embryo. Glycobiology 5,797- 806.
Grakoui, A., Bromley, S.K., Sumen, C., Davis, MM., Shaw, AS., Allen, P.M., Dustin, M.L. (1999) The immunological synapse: a molecular machine controlling T ceil activation. Science 285,221-227
Guy, C.T., Cardiff, R.D., and Muller, W.J. (1992). Induction of mammary tumors by expression of polyomavims middle T oncogene: a transgenic mouse mode1 for metastatic disease. Mol.Cell.Biol. 12,954-96 1.
Guy, C.T., Cardiff, R.D., and Muller, W.J. (1998). Induction of mamrnary tumors by expression of pol yomavirus middle T oncogene: A transgenic mouse model for metastatic disease. Mol.Cell.Biol. 12,954-96 1.
Guy, C.T., Muthuswamy, S.K., Cardiff, R.D., Soriano, P., and Muller, W.J. (1994). Activation of the c-Src tyrosine kinase is required for the induction of mammary tumors in transgenic mice. Genes Dev. 8,23-32.
Hakomori, S .-1. (1996). Tumor malignancy defined by aberrant gl ycos ylation and sphingo(g1 yco)li pid metabolism. Cancer Res. 56, 5309-53 1 8.
Hakomori, S . 4 . (1989). Aberrant glycosylation in tumors and tumor-associated carbohydrate antigens. Adv.Cancer Res. 52,257-33 1.
Hakornori, S. and Kannagi, R. (1983). Glycosphingolipids as tumor associated and di fferentiation markers. JNCI 7 l ,23 1-25 1.
Hansen, W.J., Frank, S.R., and Casanova, J.E. (1997). Cloning and characterization of human phophomannomutase, a mammaiian homologue of yeast SEC53. Glycobiology 7, 829-834.
Hams, R.J., Charnow, S.M., Gregory, T.J., and Spellman, M.W. (1990). Characterization of a soluble form of human CD4 Peptide analyses confirm the expected arnino acid sequence, identify glycosylation sites and demonstrate the presence of three disulfide bonds. Eur.J.Biochem. 188,29 1-300.
Hasenkrug, K.J. and Chesebro, B. (1997). h u n i t y to retroviral infection: the Fnend virus model. Proc Nat1 Acad Sci U S A 94,781 1-7816.
Heffernan, M., Lotan, R., Amos, B., Palcic, M., Takano, Y., and Dennis, J.W. (1993). Branching $1 -6N-Acetylglucosaminyltransferases and pol ylactosarnine expression in mouse F9 teratocarcinoma cells and differentiated counterparts. J.Biol.Chem. 268, 1242- 125 1.
Hennet, T., Chui, D., Paulson, J.C., and Marth, J.D. (1998). Immune regulation by the ST6Gal sial ytransferase. Proc.Nat1. Acad.Sci .USA 95,4504-4509.

Higgins, L.A., Sirninovitch, KA., Zhuang, D., Broçkhausen, I., and Dennis, J.W. (1991). Aberrant O-linked oligosaccharide biosynthesis in lymphocytes and platelets from patients with the Wiskott-Aldrich syndrome. J-BiolChern. 266,6280-6290.
Hoff, S.D., Matsushita, Y., Ota, D.M., Cleary, KR., Yamori, T., Hakomori, S.-I., and Irimura, T. (1989). Increased expression of siai yl-dimeric Lex antigen in liver metastases of human colorectal carcinoma. Cancer Res. 49,6883-6888.
Hoffrnan, L.H. and Wooding, F.B. (1993). Giant and binucleate trophoblast cells of mammals. J Exp Zoo1 266,559-577.
Hubbard, S C , Kranz, D.M., Longmore, G.D., Sitkovsky, MN., and Eisen, H.N. (1986). Glycosylation of the T-ce11 antigen-specific receptor and its potential role in lectin-mediated cytotoxicity. ProcN.S.Inst.Sci. 83, 1852- 1856.
Huppa, J.B. and Ploegh, H.L. (1997). In Vitro translation and assembly of a complete T ce11 receptor-CD3 complex. J.Exp.Med. 186,393403-
Ihara, Y., Sakamoto, Y., Mihara, M., Shimizu, K., and Taniguchi, N. (1997). Overexpression of N-acetylglucosaminyltransferase iII disrupts the tyrosine phosphorylation of Trk with resultant signaling dysfunction in PC12 cells treated with nerve growth factor. J-Biol-Chem. 272,9629-9634-
Ioffe, E., Liu, Y., and Stanley, P. (1996). Essential role for complex N-glycans in forming an organized layer of branchial epithelium. Proc.Natl.Acad.Sci.USA 93, 11041- llO46.
Ioffe, E., Liu, Y., and Stanley, P. (1997). Complex N-glycans in Mgatl nul1 preimplantation embryos arise from matemal Mgatl RNA. Glycobiology 7,913- 919.
Ioffe, E. and Stanley, P. (1994). Mice lacking N-acetylglucosaminyltransferase 1 activity die at mid-gestation, revealing an essential role for complex or hybrid N-linked carbohydrates. Proc.Nat1.Acad.Sci.USA 91, 728-732.
Iolascon, A., Miraglia del Giudice, E., Perrotta, S., Granatiero, M., Zelante, L., and Gasparini, P. (1999). Exclusion of three candidate genes as determinants of congenital dyserythropoietic anemia type II (CDA-II). Blood 90,4197-4200-
Ito, H., Hiraiwa, N., Sawada-Kasugai, M., Akarnatsu, S., Tachikawa, T., Kasai, Y ., Akiyarna, S., Ito, K., Takagi, H., and Kannagi, R. (1999). Altered mRNA expression of specific molecular species of fucosyl- and sialyl-transferases in human colorectal cancer tissues. 1nt.J.Cancer 71, 556-564.
Jacks, T. ( 1996). Tumor suppressor gene mutations in mice. Annu.Rev.Genet. 30,603- 636.
Jaeken, J., Schachter, H., Carchon, H., De Cock, P., Coddeville, B., and Spik, G. (1994). Carbohydrate deficient glycoprotein syndrome type II: a deficiency in Golgi localised N-acetyl-glucosamin yltransferase II. Archives DisChild. 71, 123- 127.

181
Jaln, J., McCaffrey, P.G., Miner, Z., Kerppola, T.K., Lambert, J.N., Verdine, G.L., Curran, T., and Rao, A. (3653). The T-cell transcription factor NFAT, is a substrate for calcineurin and interacts with Fos and Jun. Nature 352,355
Jarnjoom, Z.A., Jarnjoom, A.B., Sulaiman, A.H., and Naim-Ur-Rahman, a.R.A. (1993). Systemic metastasis of medulloblastoma through vetriculoperitoneal shunt report of a case and critical analysis of the literature. Surg.Neurol.40,403-410.
Janeway, C.A.J. (1992). The T ce11 receptor as a muiticomponent signalling machine: CD4/CD8 coreceptors and CD45 in T ce11 activation. Annu Rev Immun01 IO, 645-674.
Jiang, W., Swiggard, W.J., Heuffer, C., Peng, M., Mirza, A,, Steinman, R.M., and Nussenzweig, M.C. (1995). The receptor DEC-205 expressed by dendritic cells and thymic epithelial cells is involved in antigen processing. Nature 375, 151- 155.
Jones, G.E., Arumugham, R.G., and Tanzer, MIL. (1986). Fibronectin glycosylation modulates fibroblast adhesion and spreading. J.Cell Biol. 103, 1663-1668.
Kabakoff, B. and Lennarz, W.J. (1990). Inhibition of glycoprotein processing blocks assembly of spicules during development of the sea urchin embryo. J.Cell Biol. 111,391-400.
Kang, R., Saito, H., Ihara, Y., Miyoshi, E., Koyama, N., Sheng, Y., and Taniguchi, N. (1996). Transcriptional regulation of the N-acetylglucosaminyltranserase V gene in human bile duct carcinoma cells (HuCC-Tl) is mediated by Ets-1. 1.Biol.Chern. 271,26706-26712.
Karsan, A., Cornejo, C.J., Winn, R.K., Schwartz, B.R., Way, W., Lannir, N., Gershoni- Baruch, R., Etzioni, A., Ochs, H.D., and Harian, J.M. (1998). Leukocyte Adhesion Deficiency Type JI is a generalized defect of de novo GDP-fucose biosynthesis. Endothelial ce11 fucosylation is not required for neutrophil rolling on human non1 ymphoid endothelium. J.Clin.Invest. 101,2438-2445.
Keir, G., Winchester, B.G., and Clayton, P. (1999). Chhydrate-deficient glycoprotein syndromes: inbom errors of protein glycosylation. Ann.Clin.Biochem. 36,20-36.
Kelly, R.J., Ernst, L.K., Larsen, R.D., Bryant, J.G., Robinson, J.S., and Lowe, J.B. (1994). Molecular basis for H blood group deficiency in Bombay (Oh) and para- Bombay individuals. Proc Nat1 Acad Sci U S A 91, 5843-5847.
KerbeI, R.S., Florian, M.S., Man, S., Dennis, J.W., and McKenzie, 1. (1980). Carcinogenicity of tumor cell population: origin of a putative H2 isoantigenic variant. J.Natl.Cancer Inst. 64, 1221- 1230.
Khattri, R., Auger, J.A., Griffin, M.D., Sharpe, A.H., and Bluestone, J.A. (1999). Lymphoproliferative disorder in CTLA-4 knockout rnice is characterized by CD28-regulated

182
activation of Th2 responses. J Immun01 162,5784-579 1.
King, W.G., Mattaliano, M.D., ChanT-O., Tsichlis, P.N., and Brugge, J.S. (1997). Phosphatidylinositol3-kinase is required for integrin-stimulated AKT and Raf- lhitogen-activated protein kinase pathway activation. Mo1.Biol.Cell 17,4406- 4418.
Kinzler, K.W. and Vogelstein, B. (1997). Cancer-susceptibilty genes. Gatekeepers and caretakers. Nature 386,76 1-763.
Knibbs, R N , Craig, R.A., Natsuka, S., Chang, A., Carneron, M., and Lowe, J.B. (19%). The fucos y 1 tram ferase FucT-VII regulates E-selectin ligand s ynthesis in human T cells. J Ce11 Bi01 133,911-920.
Kobata, A. (1998). A retrospective and prospective view of glycopathology. G1ycoconj.J. 15, 323-33 1.
Korczak, B., Le, T., Elowe, S., and Dennis, J.W. (1997). Structure function anaiysis of rat N-acetylglucosarninyltransferase V. Glycoconjugate J. 3123, Abs # 346(Abstract)
Kornfeld, R. and Kornfeld, S. (1985). Assembly of asparagine-linked oligosaccharides. Ann.Rev.Biochem. 54,63 1-664.
Kruisbeek, A.M. (1991). In vitro assays for lymphocyte function. In J.E. Coligam, A.M. Kniisbeek, D.H. Margulies, E.M. Shevach, and W. Strober, eds. (
Lanzavecchia, A., Iezzi, g., and Viola, A. (1999). From TCR engagement to T ce11 activation: a kinetic view of T cell behavior. Ce11 96, 1-4.
Lasky, L.A., Singer, M.S., Yednoçk, T.A., Dowbenko, D., Fennie, C., Rodnguez, H., Nguyen, T., Stachel, S., and Rosen, S.D. (1989). CIoning of a lymphocyte homing receptor reveals a lectin domain. Cell56, 1045- 1055.
Lee, K.-M., Chuang, E., Griffin, M., Khattri, R., Hong, D.K., Zhang, W., Straus, D., Samelson, L.E., Thompson, C.B., and Bluestone, J.A. (1998). Molecular basis of T ce11 inactivation by CTLA4. Science 282,2263-2266.
Leevers, S. J., Vanhaesebroeck, B., and Waterfield, M.D. (1999). Signalling through phosphoinositide 3-kinases: the Iipids take centre stage. Curr Opin Cell Biol II, 2 19-225.
Lemaire, S ., Derappe, C., Michalski, J.C., Aubery, M., and Neel, D. (1994). Expression of $1-6-branched N-linked oligosaccharides is associated with activation in human T4 and Tt3 ceIl populations. J.Biol.Chem. 269,8069-8074.
Lengauer, C., Kinzler, K.W., and Vogelstein, B. (1997). Genetic instability in coiorectal cancers. Nature 386,623-627.
Lenschow, D.J., Walunas, T.L., and Bluestone, J.A. (1996). CD28/B7 system of T cell costimulation. Annu Rev Irnrnunol 1458,233-258.

Leppa, S., Heino, J., and Jalkanen, M. (1995). Increased glycosylation of beta 1 integrins affects the interaction of transformed S 115 mammary epithelial cells with laminin- 1. Ce11 Growth Differ. 6, 853-861.
Li, F., Wilkins, P.P., Crawley, S., Weinstein, J., Cummings, R.D., and McEver, R.P. (1996). Post-translational modifications of recombinant P-selectin glycoprotein ligand- 1 required for binding to P- and E-selectin. J-Biol-Chem. 271, 3255-3264.
Li, W.P. and Roth, J. (1997). Expression of beta 1,6 branched asparagine-linked oligosaccharides in non-rnitotic and non-migratory cells of normal human and rat tissues. Int.J.Cancer 71,834-490.
Lo, N.-W. and Lau, J.T. (1996). Novel heterogeneity exists in the 5'-untranslated region of the beta-galactoside alpha 2,6-siaiytransferase rnRNAs in the human B-lymphoblastoid cell line, louckes. Biochem.Biophys.Res.Cornmun.
Lo, N.-W., Sharper, J.H., Pevsner, J., and Shaper, N.L. (1998). The expanding b4- galactosyltransferase gene family: messages from the databanks. GIycobiology 8, 5 17-526.
Lu, Q. and Shur, B.D. (1997). Sperm from beta 1,4-galactosyltransferase-nul1 mice are refractory to ZP3-induced acrosome teactions and penetrate the zona pellucida poorly. Development 124,412 1-413 1.
Lu, Y. and Chaney, W. (i993). Induction of N-acetylglucosaminyltransferase V by elevated expression of activated or prote Ha-ras oncogenes. Mol.Ce1l.Biochem. 122, 85-92.
Lu, Y., Pelling, J.C., and Chaney, W.G. (1994). Tumor ce11 surface beta 1-6 branched oligosaccharides and lung metastasis. Clin.Exp.Metastasis 12-47-54.
Maly, P., Thall, A.D., Petryniak, B., Rogers, C.E., Smith, P.L., Marks, R.M., Kelly, R.J., Gersten, KM., Cheng, G., Saunders, T.L., Camper, S.A., Camphausen, R.T., Sullivan, F.X., Isogai, Y., Hindsgaul, O., von Andrian, U.H., and Lowe, J.B. (1996). The a(l,3)fucosyltransferase fuc-TVII controls leukocyte traficking through an essential role in L-, E-, and P-selectin ligand biosynthesis. Ce11 86, 643-653.
Manjunath, N., Correa, M., Ardman, M., and Ardman, B. (1995). Negative regulation of T-ce11 adhesion and activation by CD43. Nature 377,535-538.
Mannori, G., Crottet, P., Cecconi, O., Hanasaki, K., Aruffo, A., Nelson, R.M., Varki, A., and Bevilacqua, M.P. (1995). Differential coloncancer cell adhesion to E-, P-, and L-selectin: role of mucin-type glycoproteins. Cancer Res. 55,4425-443 1.
Massague, J. (1996). Crossing receptor boundarîes. Nature 382, 29-30.
McEvoy, L.M., Sun, H., Frelinger, J.G., and Butcher, E.C. (1997). Anti-CD43 inhibition of T ce11 homing. J.Exp.Med. 185, 1493-1498.

184
Meili, R., Cron, P., Hemmings, B.A., and Ballmer-Hofer, K. (1998). protein kinase BIAkt is activated by polyomavirus middle-T antigen via a phosphatidylinositol 3- kinase-dependent mechanism. Oncogene 16,903-907.
Metzler, M., Gertz, A., Sarkar, M., Schachter, H., Schrader, J. W., and Marth, J.D. (1994). Complex asparagine-linked oligosaccharides are required for morphogenic events dunng pst-implantation development. EMBO J. 13,2056-2065.
Miller, L.J. and Springer, T.A. (1999). Biosynthesis and glycosylation of plSO,95 and related leukocyte adhesion proteins. J h u n o l 139,842-847.
Miyoshi, E., Nishikawa, A., Ihara, Y., Gu, J., Sugiyama, T., Hayashi, N., Fusamoto, H., Kamada, T., and Taniguchi, N. (1993). N-acetylglucosaminy1transferase III and V messenger RNA levels in LEC rats during hepatocarcinogenesis. Cancer Res. 53, 3 899-3902.
Monks, C.R., Feiberg, B.A., Kupfer, H., Sciaky, N., and Kupfer, A. (1998). Nature 395, 82-86.
Monoston, E., Hartyani, Z., Hegedus, Z., Oçsovszki, I., Pallinger, E., and Ando, 1. (1997). Microheterogeneity of the cell surface tyrosine phosphatase, CD45RA, on T cells: phytohaemagglutinin binding and non-binding fraction of the 220 kDa isoform. Immun01 Lett 59, 17 1-176.
Myc, A., Kunicka, J.E., Melarned, M.R., and Darzynkiewicz, 2. (1989). Effect of swainsonine on stimulation and ce11 cycle progression of human lymphocytes. Cancer Res. 49,2879-2883.
Nicolson, G. (1988). Organ specificity of tumor metastasis: role of preferential adhesion, invasion and growth of malignant cells at specific secondary sites. Cancer Metastasis Rev. 7, 143-188.
Kops, G.J., de Ruiter, N.D., De Vries-Smits, A.M., Powell, D.R., Bos, J.L., Burgenng, B.M. (1999) Direct control of the Forkhead transcription factor AFX by protein kinase B. Nature 398,630-634.
Ohannesian, D.W., Lotan, D., Thomas, P., Jessup, J.M., Fukuda, M., Gabius, H.-J., and Lotan, R. (1995). Carcioembryonic antigen and other glycoconjugates act as ligands for galectin-3 in human colon carcinoma cells. Cancer Res. 55,2 19 1 - 2199.
Oriol, R., Mollicone, R., Cailleau, A., Balanzino, L., and Breton, C. (1999). Divergent evolution of fucosyltransferase genes from vertebrates, invenebrates, and bacteria. Glycobiology 9,323-334.
Orntoft, T.F., Vestergaard, E.M., Holmes, E., Jakobsen, J.S., Grunnet, N., Mortensen, M., Johnson, P., Bross, P., Gregersen, N., Skorstengaard, K., Jensen, U.B., Bolund, L., and Wolf, H. (1996). Influence of Lewis alphal 1-314-L-fucosyltransferase (FUT3) gene mutations on enzyme activity, erythrocyte phenotyping, and ciruculating tumor marker sialyl-lewis a levels. J.Biol.Chem. 271,32260-32268.

Palanivel, V., Posey, C.H.A.M., Solbach, W., Piessens, W.F., and Ham, D.A. (1996). B- ce1 outgrowth and ligand-specific production of IL-10 correlate with Th2 dominance in certain parasitic diseases. Exp.Parasitol 84, 168- 177.
Palecek, S.P., Loftus, J.C., Ginsberg, M.H., Lauffenburger, DA., and Horwitz, A.F. (1997). Integrin-ligand binding properties govern ce11 migration speed through cell-substratum adhesiveness. Nature 385 , 537-540.
Panneerselvam, K., Skovby, F., and Freeze, H.H. (1999). Abnomal metablism of mannose in fami lies with chhydrate-deficient gi ycoprotein syndrome type 1. Biochem.Mol.Med. 61, 161-167.
Parent, C.A. and Devreotes, P.N. (1999). A cell's sense of direction. Science 284,765- 769.
Paulson, J.C. and Colley, K.J. (1989a). Glycosyltransferases: Structure, loçalization, and control of ce11 type-specific glycosylation. J-BioI-Chem. 264, 176 15-176 18.
Paulson, J.C., Weinstein, J., and Schauer, A. (1989b). Tissue-specific expression of sial yltransferase. J.Bio1 .Chem. 264, 1093 1- 10934.
Pennington, J.E., Rastan, S., Roelcke, D., and Feizi, T. (1985). Saccharide structures of the mouse embryo during the first eight days of development. J-Embryol. 90, 335-36 1.
Perillo, N.L., Marcus, M.E., and Baum, L.G. (1998). Galectins versatile modulators of ce11 adhesion, ceIl proliferation, and ce11 death. J.Mol.Med. 76,402412.
Perillo, N.L., Pace, K.E., Seilhamer, J.J., and Baum, L.G. (1995). Apoptosis of T cell mediated by galectin- 1. Nature 378,736-739.
Perng, G.-S., Shoreibah, M., Margitich, I., Pierce, M-, and Fregien, N. (1994). Expression of N-acetylglucosaminyltransferase V mRNA in rnarnmalian tissues and ce11 lines. Glycobiology 4,867-872.
Peter, M., Nakagawa, J., Doree, M., Labbe, J.C., and Nigg, E.A. (1 990). Identification of major nucleolar proteins as candidate mitotic substrates of cdc2 kinase. Ce11 60, 791-801.
Phillips, M.L., Schwartz, B.R., Etzioni, A., Bayer, R-, Ochs, H.D., Paulson, J.C., and Harlan, J.M. (1995). Neutrophii adhesion in leukoçyte adehsion deficiency syndrome type 2. J.Clin.Invest. 96,2898-2906.
Pierce, M. and Arango, J. (1986). Rous sarcoma virus-transformed baby hamster kidney cells express higher ievels of asparagine-linked tri- and tetraantennary gl ycopeptides containing [GlcNAc-B(l,6)Man-a( l,6)Man ] and poly-N- acetyllactosamine sequences than baby hamster kidney cells. J.Biol.Chem. 261, 10772- 10777.
Piller, F., Piller, V., Fox, R.I., and Fukuda, M. (1988). Human T-lymphocyte activation is associated with changes in O-glycan biosynthesis. J.Biol.Chem. 263, 15 146-

Powell, L.D., Sgroi, D., Sjoberg, E.R., Stamenkovic, I., and Varki, A. (1993). Natural ligands of the B ce11 adhesion molecule CD22$ carry N-linked oligosaccharides with a-2,6-linked sialic acids that are required for recognition. J.Biol.Chem. 268,70 19-7027.
Priatel, J.J., Sarkar, M., Schachter, H., and Marth, J.D. (1997). Isolation, characterization and inactivation of the mouse Mgar3 gene: the bisecting N-acetylglucosamine in asparagine-linked oligosaccharides appears dispensable for viability and reproduction. Glycobiology 7,4556.
Pntchett, T.J., Brossmer, R., Rose, U., and Paulson, J-C. (1987). Recognition of monovalent sialosides by influenza virus H3 hemagglutinin. Virology 160, 502- 506.
Rademacher, T.W., Parekh, B., and Dwek, R.A. (1988). Glycobiology. Ann-Rev-Biochem. 57,785-838.
Rebbaa, A., Yamamoto, H., Saito, T., Meuillet, E., Kim, P., Kersey, D.S., Bremer, E.G., Taniguchi, N., and Moskal, J.R. (1997). Gene transfection-mediated overexpression of beta 1,4-N-acetylglucosamine bisecting oligosaccharides in glioma ce11 line U373 MG inhibits epidermal growth factor receptor function. J-Biol-Chem. 272,9275-9279.
Reif, K., Nobes, C.D., Thomas, G., Hall, A., and Cantrell, D.A. (1996). phosphatidylinosi tol3-kinase signals activate a selective subset of Rac/Rho dependent effector pathways. Curr.Bio1. 6, 1445-1455.
Romero, P.A- and Herscovics, A. (1986). Transfer of non-glucosylated oligosaccharides from lipid to protein in marnmalian cells. J.Biol.Chem. 261, 15936- l~94û.
Ruegg, CL., Rajasekar, S., Stein, B.S., and Engleman, E.G. (1992). Degradation of CD4 fotlowing phorbol-induced intemalization in hyman T lymphocytes. Evidence for distinct endocytic routing of CD4 and CD3. J Bi01 Chem 267, 18837-18843.
Ruoslahti, E. (1996). Integrin signaling and matrix assembly. tumour biol. 17, 1 17- 124.
Saez, E., Rutberg, S*E-, Mueller, E., Oppenheim, H., Smoluk, J., Yuspa, S.H., and Spiegelman, B.M. (1995). c-fos is required for malignant progression of skin tumors. Ce11 82,721-732.
Saito, H., Gu, J., Nishikawa, A., Ihara, Y., Fujii, J., Kohgo, Y., and Taniguchi, N. (1995). Organization of the human N-acetylglucosaminyltransferase V gene. Eur.J.Biochem. 233, 18-26.
Schachter, H. (1986). Biosynthetic controls that determine the branching and microheterogeneity of protein-bound oligosaccharides. Biochem-Ce11 Biol. 64, 163-181.
Schaller, M.D., Otey, C.A., Hildebrand, J.D., and Parsons, J.T. (1995). J Ce11 Biol 130,

Schlaepfer, D.D., Hanks, S.K., Hunter, T., and van der Geer, P. (1995). Integrin-mediated signal transduction linked to Ras pathway by GRB2 binding to focal adhesion kinase. Nature 372,786-791.
Schmits, R., Filmus, J.G.N., Senaldi, G., Kiefer, F., Kundig, T., Wakeharn, A., Shahinian, A., Catzavelos, C., Rak, J., FurIonger, C., Zakarian, A., Simard, J.J.L., Ohashi, P.S., Paige, C.J., Guitemz-Rarnos, J-C., and Mak, T.W. (1997). CD44 regulates hematopoietic progeni tor distri bution, granuloma formation, and tumorigenicit y. Bled 90-22 17-2233.
Seberger. P.J. and Chaney, W.G. (1999). Control of metastasis by açn-linked, $1-6 branched oligosaccharides in mouse mammary cancer cells. Glycobiology 9, 235-24 1.
Seelentag, W.K., Li, W.P., Schmitz, S.F., Metzger, U., Aeberhard, P., Heitz, P.U., and Roth, J. (1998). Prognostic value of beta 1-6-branched ~Iigosaccharides in human colorectal carcinoma. Cancer Res. 58, 5559-5564.
Sgroi, D., Varki, A., Braesch-Andersen, S., and Stamenkovic, 1. (1993). CD22, a B cell- specific immunoglobulin superfamily rnember, is a sialic acid-binding lectin. J.Biol.Chem. 268.701 1-7018.
Shahinian, A., Pfeffer, K., Lee, K.P., Kundig, T.M., Kishihara, K., Wakeharn, A., Kawai, K., Ohashi, P.S., Thompson, C.B., and Mak, T.W. (1993). Differential T ce11 costimulatory requirements in CD28-deficient mice. Science 261,609-6 12.
Sheares, B.T. (1988). Site-specific glycosylation in animal cells. J-Biol-Chem. 263, 12778-12782.
Sheng, Y., Yoshimura, M., houe, S., Oritani, K., Nishiura, T., Yoshida, H., Ogawa, M., Okajima, Y., Matsuzawa, Y., and Taniguchi, N. (1997). Remodeling of glycoconjugates on CD44 enhances ce11 adhesion to hyaluronate, tumor growth and metastasis in B 16 melanoma cells expressing f3 1.4-N- acetylglucosaminyltransferase III. 1nt.J.Cancer 73,850-858.
Shimodaira, K., Nakayama, J., Nakamura, N., Hasebe, O., Katsuyama, T., and Fukuda, M. (1997). Carcinoma-associated expression of core 2 P-1,6-N- Acet ylgIucosarnin yltransferase gene in human colorectal cancer: Role of O- gl ycans in turnor progression. Cancer Res. 57,520 1-5206.
Shoreibah, M., Perng, G.-S., Adler, B., Weinstein, J., Basu, R., Cupples, R., Wen, D., Browne, J.K., Buckhaults, P., Fregien, N., and Pierce, M. (1993). Isolation, c haracterization, and expression of cDNA encoding N- Acetylglucosarninyltransferase V. J-BiolChem. 268, 1538 1-15385.
Sjoberg, E.R., Kitagawa, H., Glushka, J,, van Halbeek, H., and PauIson, J.C. (1999). Molecular cloning of a developmentally regulated N-acetylgalactosamine al p h a2 -6-si al y1 tran s ferase

188
specific for sial ylated gl ycoconjugates. J Bi01 Chem 2 71,7450-7459-
Skelton, T.P., Zeng, C., Nocks, A., and Starnenkovic, 1. (1998). Glycosylation provides both stimulatory and inhibitory effects on ce11 surface and soluble CD44 binding to hyaluronan. J-Ce11 Biol. I40,43 1-446.
Solter, D. and Knowles, B.B. (1978). Monoclonal antibodies defining a stage specific mouse embryonic antigen. Proc.Nat1.Acad.Sci.USA 75,5565-5569.
Springer, G.F. (1999). hunoreactive T and Tn epitopes in cancer diagnosis, prognosis, and immunotherapy. J Mol Med 75,594-602.
Spnnger, T.A. (1994). Traffic signals for lymphocyte recirculation and leukocyte migration: the rnultistep paradigrn. Cell 76,30 1-3 14.
Stanley, P. (1983). Lectin-resistant CHO ceIls: selection of new mutant phentoypes. Somat.Cel1 Genet. 9, 593-608.
Stanley, P. (1984). Glycosylation mutants of animal cells. Ann.Rev.Genet. 18,525-552.
Stephens, L., Anderson, K., Stokoe, D., Erdjument-Bromage, H., Painter, G.F., Holmes, A.B., Gaffney, P.R.J., Reese, C.B., McCormick, F., Tempst, P., Coadwell, J., and Hawkins, P.T. (1 998). Protein kinase B kinases that mediate phosphatidyiinositoI 3,4,5-trisphosphate-dependent activation of protein kinase B. Science 279,7 10- 7 14.
Svensson, E.C., Soreghan. B., and Paulson, J-C. (1990). Organization of the $-galactoside &,6-sialyltransferase gene. J.Biol.Chem. 265,20863-20868.
Takano, R., Muchmore, E.A., and Dennis, J.W. (1994). Sialylation and malignant potential in tumor ce11 glycosylation mutants. Glycobiology 4, 665-674.
Takeuchi, M. and Kobata, A. (1991). Structures and functional roles of the sugar chains of human erythropoietins. Glycobiology 1,337-346.
Tan, J., Dunn, J., Jaeken, J., and Schachter, H. (1996). Mutations in the MGATS gene controlling complex N-glycm synthesis cause carbohydrate-deficient glycoprotein syndrome type II, an autosomal recessive disease with defective brain development. Am J Hum Genet 59,8 10-8 17.
Tan, P.B.O. and Kim, S.K. (1999). Signaling specificity: the RTK/RAS/MAPK pathway in metazoans . Trends Genet. 15, 145-149.
Tanaka, N., Ishihara. M., Kitagawa, M., Harada, H., Kimura, T., Matsuyama, T., Lamphier, M.S., Aizawa, S., and Mak, T.W. (1994). Cellular cornmitment to oncogene-induced transformation or apoptosis is dependent on the transcription factor IRF-1. Ce11 77, 829-839.
Thumher, M., Rusconi, S., and Berger, E.G. (1999). Persistent repression of a functional allele can be responsible for galactosyltransferase deficiency in Tn syndrome.

Tsuboi, S. and Fukuda, M. (1997). Branched O -1inked oligosaccharides ectopically expressed in transgenic mice reduce primary T-cell immune responses. EMBO J. 16,6364-6373.
Tsuji, S. (1996). Molecular cloning and functional analysis of sialyltransferases. J.Biochem.(Tokyo) 120, 1-13.
Turner, C.E. (1998). Molecules in focus, Paxillin. Int. J. Biochem. Ce11 Biol. 30,955-959.
Valitutti, S., Mulle, S., Cella, M., Padovan, E., and Lanzavecchia, A. (1995). Serial triggering of many T-ce11 receptors by a few peptide-MHC complexes. Nature 375, 148-151.
van den Eijnden, D.H., Koenderman, A.H.L., and Schiphorst, W.E.C.M. (1988). Biosynthesis of blood group i-active polylactosaminoglycans. J.Biol.Chem. 263, 12461-12465.
VanderElst, 1. and Dennis, J.W. (1991). N-linked oligosaccharide proçessing and autocrine-stimulation of tumor ce11 proliferation. Exp.Ce11 Res. l92,6 12-613.
Varki, A. and Marth, J. (1995). Oligosaccharides in vertebrate development. Dev-Biol. 6, 127-138.
Velders, M.P. and Kast, W.M. (1999). Modulation of the immune response and tumor growth by activated Ras. leukernia 13,502-5 13.
Verbeek BS, Vroom TM, Adriaansen-Slot SS, Ottenhoff-Kalff AE, Geertzema JG, Henniprnan A, and Rijksen G (1999). c-Src protein expression is increased in human breast cancer. An irnmunohistochernical and biochemical anal ysis. J-Pathol. 180,383-388.
Vespa, G.N., Lewis, L.A., Kozak, KR., Moran, M., Nguyen, J.T., Baum, L.G., and Miceli, M C . (1999). Galectin-1 specifically modulates TCR signals to enhance TCR apoptosis but inhibit IL-2 production and proliferation. J Immunol 162,799-806.
Vestweber, D. and Blanks, J.E. (1999). Mechanisms that regulate the function of the selectins and their ligands. Physiol-Rev. 79, 18 1-2 13.
Viola, A. and Lanzavecchia, A. (1996). T ce11 activation determined by T ceIl receptor number and tunable thresholds. Science 273, 104-106.
Viola, A., Schroeder, S., Sakakibara, Y., and Lanzavecchia, A. (1999). T lymphocyte costimulation mediated by reorganization of membrane microdomains. Science 283,680-682.
Wang, J., Lim, K., Smolyar, A., Teng, M., Liu, J., Tse, A.G., Hussey, R.E., Chishti, Y., Thomson, C.T., Sweet, R.M., Nathenson, S.G., Chang, H.C., Sacchettini, J.C., and Reinherz, E.L. (1998). Atomic structure of an alphabeta T ce11 receptor (TCR)

heterodimer in complex with an anti-TCR fab fragment denved from a mitogenic antibody. EMBO J f 7, 10-26.
Ware, F.E., Vassilakos, A., Peterson, P.A., Jackson, M.R., Lehrman, M-A-, and Williams, D.B. ( 1995). The molecular chaperone calnexin binds Glcl MangGlcNAcz oligosaccharide as an initial step in recognizing unfolded glycoproteins. J.Bio1.Chem. 270,46974704.
Wasylyk, B., Hagman, J., and Gutierrez-Hartmann, A. (1998). Ets transcription factors: nuclear effectors of the Ras-MAP-kinase signaling pathw ays. Trends Biochem.Sci 23,213-216.
Webster, M.A., Cardiff, R.D., and Muller, W.J. (1995). Induction of marnrnary epithelial hyperplasias and rnarnmary tumors in transgenic rnice expressing a murine mammary tumor virudactivated c-src fusion gene. Proc Nat1 Acad Sci U S A 92,7849-7853.
Webster, M.A., Hutchinson, J.N., Rauh, M.J., Muthuswamy, S.K., Anton, M., Tortorice, C.G., Cardiff, R.D., Graham, F.L., Hassell, J.A., and Muller, W.J. (1998). Requirement for both Shc and phosphatidylinositol3' kinase signaling pathways in polyomavirus rniddle T-mediated mammary tumorigenesis. Mol.Cel1.Biol. 18, 2344-3359.
Webster, M.A. and Muller, W.J. (1994). Mammary turnorigenesis and metastasis in transgenic mice. Semin-Cancer Biol. 5,69-76.
Wehr, R., Mansouri, A., de Maeyer, T., and Gruss, P. (1997). Fkh5-deficient mice show dysgenesis in the caudal midbrain and hypothalarnic mammillary body. Development 124,4447-4456.
Weinberg, R.A. (1999). The retinoblastoma protein and cell cycle control. Ce11 81,323- 330.
Weinstein, J., Sundararn, S., Wang, X., Delgado, D., Basu, R., and Stanley, P. (1996). A point mutation causes mistargeting of golgi GlcNAc-TV in the Lec4A Chinese hamster ovary glycosylation mutant. J.Bio1.Chem. 271,27462-27469-
Wu, W., Harley, P.H., Punt, J.A., S h m w , S.O., and Kearse, K.P. (1996). Identification of CD8 as a peanut agglutinin (PNA) receptor molecule on immature th ymocytes. J Exp Med 184,759-764.
Wulfing, C. and Davis, M.M. (1998a). A receptor/cytoskeletal movement triggered by costimulation during T ce11 activation. Science 282,2266-2269.
Wutfing, C . and Davis, M.M. (1998b). A receptor/cytoskeletal movement tnggered by costimulation during T cell activation. Science 282,2266-2269.
Xavier, R., Brennan, T., Li, Q., McConnick, C., and Seed, B. (1998). Membrane compartmentation is required for efficient T ceIl activation. Irnrnunity 8,723- 732.

Yagel, S., Feinmesser, R., Waghorne, C., Lala,P.K., Breitman, M.L., andDennis, J.W. (1989). Evidence that 1-6 branched Asn-linked oligosaccharides on metastatic tumor cells facilitate invasion of basement membranes. 1nt.J.Cancer 44,685690.
Yagel, S., Kerbel, R.S., Eldar-Gera, T., and Dennis, J.W. (1990). Basement membrane invasion by first trimester human trophoblast: requirement for branched comptex- type Asn-linked oligosaccharides. Clin.Exp.Metastasis 8,305-3 18.
Yamamoto, H., Kaneko, Y., Rebbaa, A., Bremer, E.G., and Moskal, J.R. (1997). alpha,6-sialyltransferase gene transfection into a human giioma ce11 line (U373 MG) results in decreased invasivity. J.Neurochem. 68,2566-2576.
Yamashita, K., Ohkura, T., Tachibana, Y., Takasaki, S., and Kobata, A. (1984). Comparative study of the oligosaccharides released from baby hamster kidney cells and their polyoma transformant by hydrazinolysis- JBiol-Chem. 259, 108%- 10840.
Yarnashita, K., Tachibana, Y ., Ohkura, T., and Kobata, A. (1985). Enzymatic basis for the structural changes of asparagine-linked sugar chains of membrane glycoproteins of baby hamster kidney cells induced by polyoma transfonnation. J.BioLChem. 260,3963-3969-
Yang, J., Bhaumik, M., Liu, Y., and Stanley, P. (1994). Regulation of N-linked glycosylation. Neuronal cell-specific expression of a 5' extended transcript from the gene encoding N-acetylglucosaminyltransferase 1. Glycobiology 4,703-7 12.
Yashiro, Y., Tai, X.-G., Toyo-oka, K., Park, C.S., Abe, R., Hamaoka, T., Kobayashi, M., Neben, S., and Fujiwara, H. (1999). A fundamental difference in the capacity to induce proliferation of naive T cells between CD28 and other CO-stimulatory molecules. Eur J Immun01 28,926-935.
Yeh, J.C., Ong, E., and Fukuda, M. (1999). Molecular cloning and expression of a novel beta- 1-6-N-acetylglucosamin yltransferase that foms core 2, core 4, and 1 branches. J Bi01 Chem 274,3215-322 1.
Yi, X. and Freund, R. (1998). Deletion of proline-rich domain in polyornavims T antigens results in virus partially defective in transfonnation and tumorigenesis. Virology 248,420-43 1.
Yoshimura, M., Ihara, Y., Matsuzawa, Y ., and Taniguchi, N. (1996). Aberrant glycosylation of E-cadherin enhances cell-cell binding to suppress metastasis. J.Biol.Chem. 271, 138 11-138 15.
Yoshimura, M., Nishikawa, A., Ihara, Y., Taniguchi, S., andTaniguchi, N. (1995). Suppression of lund metastasis of B 16 mouse melanoma by N- acetylglucosarnin yltransferase II gene transfection. Proc.Natl.Acad.Sci .USA 92, 8753-8758.

Yousefi, S., Higgins, E., Doaling, Z., Hindsgaul, O., Pollex-Kruger, A., and Demis, J.W. (199 1). Increased UDP-G1cNAc:Gal $1 -3GalNAc-R (GlcNAc to GalNAc) B 1-6 N-acetylglucosaminyltransferase activity in transformed and metastatic murine tumor ce11 lines: control of polylactosamine-synthesis. J.Biol.Chem. 266, 1772- 1783.
Zenner, G., Dirk zur Hausen, J., and Mustelin, T. (1995). Towards unraveling the complexity of T ce11 signal transduction. BioEssays 17,967-975.
Zheng, M., Fang, H., and Hakomori, S.-1. (1994). Functional role of N-glycosylation in a5P 1 integrin receptor. I.Biol.Chem. 265, 12325- 1233 1.