part one epidemiology and risk...
TRANSCRIPT

BLBK041-Kim September 22, 2008 17:30
PART ONE
Epidemiology and riskfactors
COPYRIG
HTED M
ATERIAL

BLBK041-Kim September 22, 2008 17:30

BLBK041-Kim September 22, 2008 17:30
1 Anatomy of intracranial arteriesDavid S Liebeskind and Louis R Caplan
Comprehensive knowledge of intracranial arterialanatomy forms the basis for consideration ofintracranial atherosclerosis. Anatomy defines thelocation of such neurovascular lesions, delineatesthe extent and involvement of branching perfora-tors, and consequently determines the effects ondownstream perfusion. Anatomy is also intertwinedwith pathophysiology, as vessel morphology influ-ences hemodynamic variables that promote plaquegrowth and vessel wall constituents may predisposeto atherosclerotic involvement. Once an atheroscle-rotic plaque has formed, the arterial territories withinthe brain may shift, reflecting diminished perfu-sion beyond a stenosis and compensatory collat-eral flow via anastomoses from adjacent arterialsources.1 Simply stated, the anatomical features ofthese arteries or pipes and their perforators de-termine perfusion, penumbra, and the parenchymalconsequences of brain ischemia. These intracranialvessels differ in anatomy from other circulatory bedsin the heart or periphery, with only limited corre-lates noted in the comparative anatomy of intracra-nial arteries across species. Arterial anatomy adds tothe complexity of neurological localization, provid-ing a unique classification of neurovascular disorders.Consideration of intracranial arterial anatomy is mostgermane to clinical management where recognitionof particular stroke syndromes influences treatmentdecisions. Identification of a culprit atheroscleroticlesion also hinges on anatomical details of thecase.
The historical perspective on characterizinganatomy of the intracranial arteries includes an ironictwist where only marginal advances regarding pathol-
ogy of these arterial segments have been made sincethe autopsy series performed hundreds of years ago,and angiography reigns as the definitive modalityfor defining these structures almost a century afterits introduction. Pathology related to atheroscleroticinvolvement of the major intracranial arteries haslargely eluded modern imaging techniques becauseof the small size of these vessels and the orientationof these segments that defy conventional imagingplanes. Numerous noninvasive methods have beendeveloped to image intracranial arterial anatomy,2
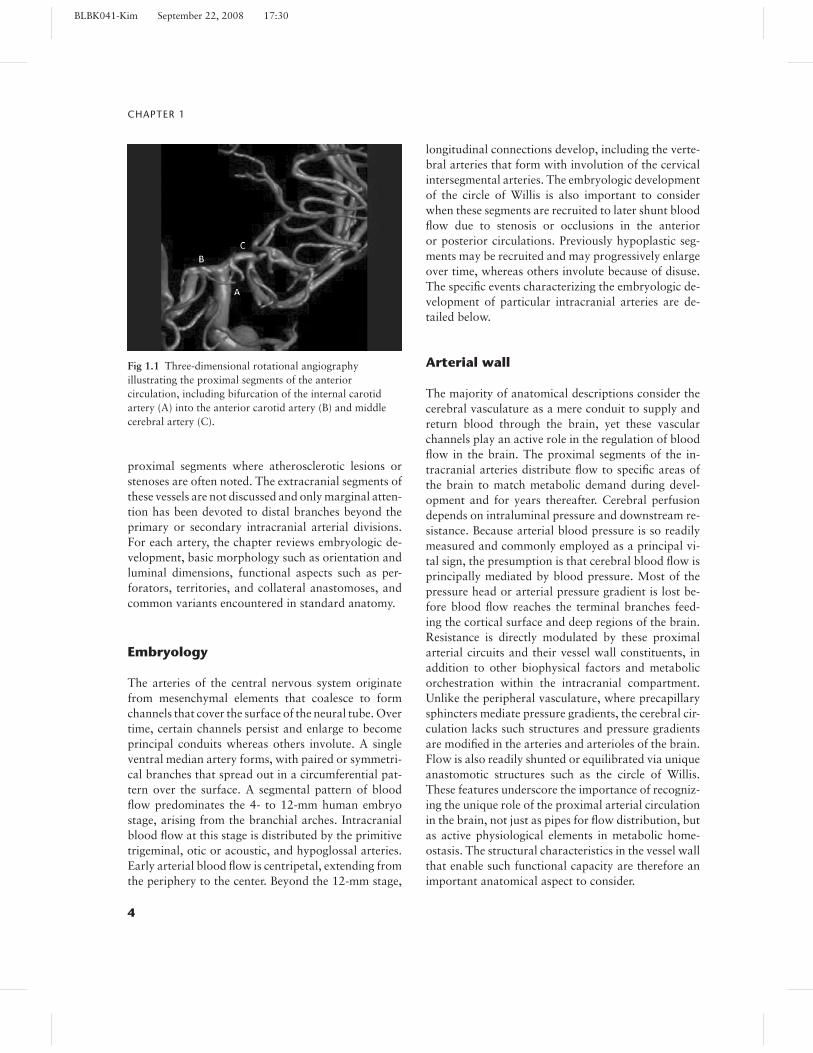
yet these modern vascular imaging techniques in-cluding transcranial Doppler ultrasound (TCD),computed tomographic angiography (CTA), andmagnetic resonance angiography (MRA) are not asaccurate as the gold standard of conventional ordigital subtraction angiography (DSA).3 Recently, theadvent of angioplasty and stenting for intracranialatherosclerotic disease has reinforced the importanceof DSA, since arterial access is needed for treatment.Noninvasive imaging modalities such as TCD, CTA,and MRA each provide differing information re-garding a balance of anatomical information, suchas measures of the arterial lumen versus physiolog-ical data reflecting flow through a specific arterialsegment and distal perfusion. DSA remains theprevailing method for evaluating vascular anatomyin the brain, although modifications such as three-dimensional rotational angiography (Fig. 1.1) haveallowed for expansion across numerous frames ofreference.
This introductory chapter considers the vascu-lar anatomy of the major intracranial arterial seg-ments supplying blood flow to the brain, emphasizing
3
BLBK041-Kim September 22, 2008 17:30
CHAPTER 1
Fig 1.1 Three-dimensional rotational angiographyillustrating the proximal segments of the anteriorcirculation, including bifurcation of the internal carotidartery (A) into the anterior carotid artery (B) and middlecerebral artery (C).
proximal segments where atherosclerotic lesions orstenoses are often noted. The extracranial segments ofthese vessels are not discussed and only marginal atten-tion has been devoted to distal branches beyond theprimary or secondary intracranial arterial divisions.For each artery, the chapter reviews embryologic de-velopment, basic morphology such as orientation andluminal dimensions, functional aspects such as per-forators, territories, and collateral anastomoses, andcommon variants encountered in standard anatomy.
Embryology
The arteries of the central nervous system originatefrom mesenchymal elements that coalesce to formchannels that cover the surface of the neural tube. Overtime, certain channels persist and enlarge to becomeprincipal conduits whereas others involute. A singleventral median artery forms, with paired or symmetri-cal branches that spread out in a circumferential pat-tern over the surface. A segmental pattern of bloodflow predominates the 4- to 12-mm human embryostage, arising from the branchial arches. Intracranialblood flow at this stage is distributed by the primitivetrigeminal, otic or acoustic, and hypoglossal arteries.Early arterial blood flow is centripetal, extending fromthe periphery to the center. Beyond the 12-mm stage,
longitudinal connections develop, including the verte-bral arteries that form with involution of the cervicalintersegmental arteries. The embryologic developmentof the circle of Willis is also important to considerwhen these segments are recruited to later shunt bloodflow due to stenosis or occlusions in the anterioror posterior circulations. Previously hypoplastic seg-ments may be recruited and may progressively enlargeover time, whereas others involute because of disuse.The specific events characterizing the embryologic de-velopment of particular intracranial arteries are de-tailed below.
Arterial wall
The majority of anatomical descriptions consider thecerebral vasculature as a mere conduit to supply andreturn blood through the brain, yet these vascularchannels play an active role in the regulation of bloodflow in the brain. The proximal segments of the in-tracranial arteries distribute flow to specific areas ofthe brain to match metabolic demand during devel-opment and for years thereafter. Cerebral perfusiondepends on intraluminal pressure and downstream re-sistance. Because arterial blood pressure is so readilymeasured and commonly employed as a principal vi-tal sign, the presumption is that cerebral blood flow isprincipally mediated by blood pressure. Most of thepressure head or arterial pressure gradient is lost be-fore blood flow reaches the terminal branches feed-ing the cortical surface and deep regions of the brain.Resistance is directly modulated by these proximalarterial circuits and their vessel wall constituents, inaddition to other biophysical factors and metabolicorchestration within the intracranial compartment.Unlike the peripheral vasculature, where precapillarysphincters mediate pressure gradients, the cerebral cir-culation lacks such structures and pressure gradientsare modified in the arteries and arterioles of the brain.Flow is also readily shunted or equilibrated via uniqueanastomotic structures such as the circle of Willis.These features underscore the importance of recogniz-ing the unique role of the proximal arterial circulationin the brain, not just as pipes for flow distribution, butas active physiological elements in metabolic home-ostasis. The structural characteristics in the vessel wallthat enable such functional capacity are therefore animportant anatomical aspect to consider.
4

BLBK041-Kim September 22, 2008 17:30
ANATOMY OF INTRACRANIAL ARTERIES
Several features distinguish intracranial arteriesfrom arteries of similar caliber elsewhere in the body.Arteries in the brain have a well-developed internalelastic lamina with only a minimal degree of elasticfibers scattered in the media.4 Unlike arteries elsewherethroughout the body, the intracranial arteries do nothave an external elastic lamina. Other distinctive fea-tures of the intracranial arteries include the presenceof tight endothelial junctions with a relative paucityof pinocytic vesicles, and differing distribution of en-zymes within the vessel wall. The adventitial layer istypically thin compared with the systemic arteries. Ingeneral, the cerebral arteries have a smaller wall-to-lumen ratio than arteries elsewhere in the body.5 Over-all, the intimal layer accounts for about 17% of totalvessel wall thickness, with the media constituting 52%and adventitia only 31%.6,7
The arterial lumen is defined by the adjacent ar-chitecture of the vessel wall. Cerebral endothelialcells with tight junctions form a critical element ofthe blood–brain barrier.8 These endothelial cells arenot fenestrated and the tight junctions bestow onlyselective permeability to this boundary, preventingexchange of numerous substances. This boundary isoften referred to as the “blood–brain barrier.” Underpathophysiologic conditions, this selective permeabil-ity boundary may be deranged.9 The number of en-docytotic vesicles is also limited compared with theendothelial lining of other vascular beds. Cerebralendothelial cells have a high concentration of mito-chondria, denoting their active metabolic role and,possibly, their vulnerability to ischemia.10 Endothe-lial cells in cerebral arteries and arterioles play anactive role in the regulation of hemodynamics. This ca-pacity is partially related to the expression of a widearray of vasoactive substances, including endothelinand nitric oxide.11 The internal elastic lamina of in-tracranial arteries is fenestrated, with holes that varyin size according to the arterial segment.12 Beyond theendothelial layer, the cerebral arteries have protuber-ances at distal branching sites that also modulate flow.These structures have been variably defined as intimalcushions, bifurcation pads, or subendothelial protu-berances. Underneath the luminal surface, these struc-tures contain groups of smooth muscle cells arrangedin irregular fashion, with intertwined collagenous fib-rils and are encompassed by the split internal elasticmembrane.13 Although the exact role of these struc-tures in titration of arterial pressure has not been fully
elucidated, it appears that these structures help alterflow via fluid shear stress mechanisms. Fluid shearstress is a critical physiological variable both in thedevelopment of atherosclerosis and in compensatoryarteriogenesis.14,15 A circumferential orientation of thesmooth muscle cells at branching sites may be relatedto titration of arterial inflow resistance by acting via asphincter-like mechanism.
In normal intracranial arteries, smooth muscle cellsmake up 72% of the media, whereas this composi-tion is radically altered under pathophysiologic con-ditions such as intracranial atherosclerosis or chronichypertension.4 Age-related changes are found in thecomposition of the media. Autonomic nerves locatedin the tunica adventitia have connections with thesesubendothelial structures via intercellular smoothmuscle cell contacts. Within the media, smooth mus-cle cells are generally oriented in a pattern circumfer-ential to the lumen except at bifurcations.6 Adjacentcollagen and elastin fibers run perpendicular to thesmooth muscle layer or in parallel with the long axisof the vessel. The thin medial layer of intracranial ar-teries compared with systemic vessels is thought to berelated to compliance differences associated with sur-rounding cerebrospinal fluid. The number of smoothmuscle cell layers within the media diminishes distally.A basement membrane associated with the adjacentsmooth muscle cells forms the framework for adjoin-ing layers of the intima and adventitia. Nerve fibersapproach the media from the adventitial layer. Withinthe adventitia, loose connective tissue surrounds auto-nomic nerve fibers and all vessel wall structures are en-closed by spindle-shaped fibrocytes. Once beyond thedura mater, the intracranial arteries have no vasa va-sorum. The external surface of the intracranial arteriesin these regions is in direct contact with surroundingcerebrospinal fluid. A rete vasorum in the adventitiais permeable to large proteins, allowing ingress or ex-change with cerebrospinal fluid in the subarachnoidspace.16
Characteristics of the intracranial arterial wall inhumans typically consider the proximal intracranialarteries such as the middle cerebral artery (MCA) sep-arately from much smaller intracerebral or pial ar-terioles. As the internal carotid artery (ICA) coursesdistally, there is progressive disappearance of the ex-ternal elastic lamina. The MCA is a terminal continu-ation of the ICA with a gradual change in blood vesselwall characteristics of histopathology. The relative
5
BLBK041-Kim September 22, 2008 17:30
CHAPTER 1
amounts of intima, media, and adventitia in the MCAare less than the equivalent amount per vessel size inthe more proximal ICA. The MCA internal elastic lam-ina is thicker and partially fenestrated. Compared withsimilar sized extracranial arteries, the MCAs have lessadventitia with less elastic tissue and few perivascularsupporting structures, including an absence of vasavasorum.17
Internal carotid artery
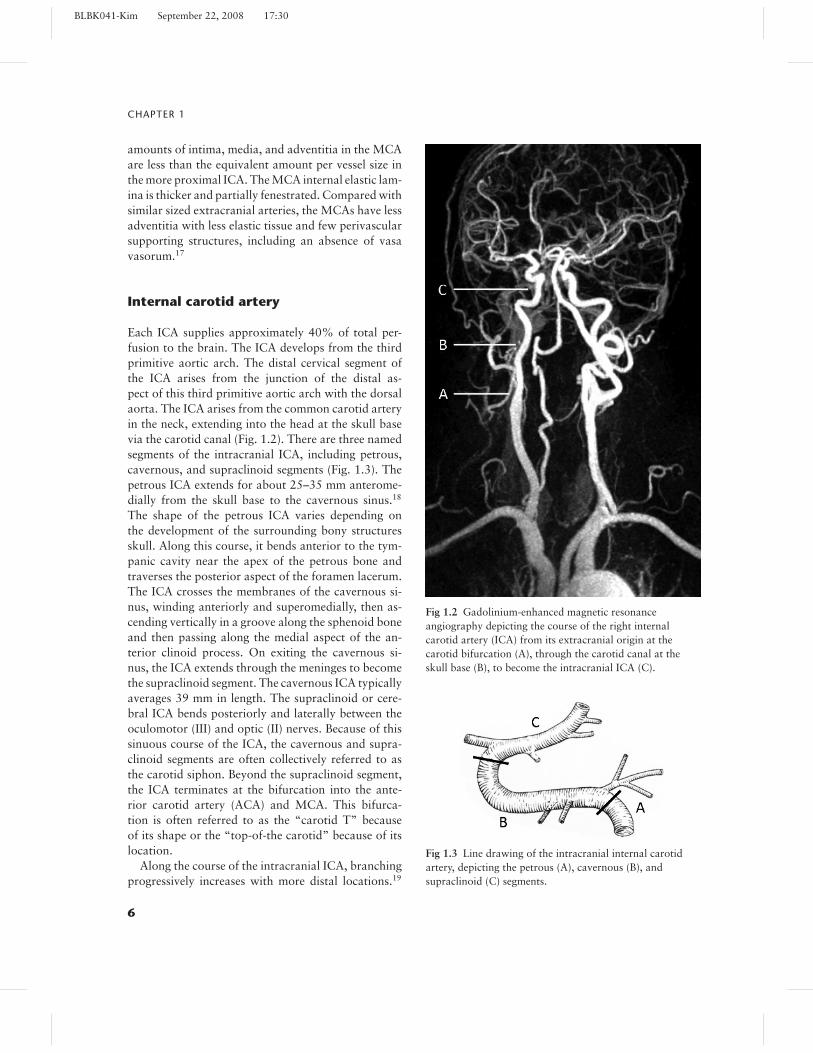
Each ICA supplies approximately 40% of total per-fusion to the brain. The ICA develops from the thirdprimitive aortic arch. The distal cervical segment ofthe ICA arises from the junction of the distal as-pect of this third primitive aortic arch with the dorsalaorta. The ICA arises from the common carotid arteryin the neck, extending into the head at the skull basevia the carotid canal (Fig. 1.2). There are three namedsegments of the intracranial ICA, including petrous,cavernous, and supraclinoid segments (Fig. 1.3). Thepetrous ICA extends for about 25–35 mm anterome-dially from the skull base to the cavernous sinus.18
The shape of the petrous ICA varies depending onthe development of the surrounding bony structuresskull. Along this course, it bends anterior to the tym-panic cavity near the apex of the petrous bone andtraverses the posterior aspect of the foramen lacerum.The ICA crosses the membranes of the cavernous si-nus, winding anteriorly and superomedially, then as-cending vertically in a groove along the sphenoid boneand then passing along the medial aspect of the an-terior clinoid process. On exiting the cavernous si-nus, the ICA extends through the meninges to becomethe supraclinoid segment. The cavernous ICA typicallyaverages 39 mm in length. The supraclinoid or cere-bral ICA bends posteriorly and laterally between theoculomotor (III) and optic (II) nerves. Because of thissinuous course of the ICA, the cavernous and supra-clinoid segments are often collectively referred to asthe carotid siphon. Beyond the supraclinoid segment,the ICA terminates at the bifurcation into the ante-rior carotid artery (ACA) and MCA. This bifurca-tion is often referred to as the “carotid T” becauseof its shape or the “top-of-the carotid” because of itslocation.
Along the course of the intracranial ICA, branchingprogressively increases with more distal locations.19
Fig 1.2 Gadolinium-enhanced magnetic resonanceangiography depicting the course of the right internalcarotid artery (ICA) from its extracranial origin at thecarotid bifurcation (A), through the carotid canal at theskull base (B), to become the intracranial ICA (C).
Fig 1.3 Line drawing of the intracranial internal carotidartery, depicting the petrous (A), cavernous (B), andsupraclinoid (C) segments.
6

BLBK041-Kim September 22, 2008 17:30
ANATOMY OF INTRACRANIAL ARTERIES
The petrous segment gives rise to the caroticotym-panic artery, supplying the tympanic cavity, and thepterygoid or vidian branch passing through the ptery-goid canal.18 This vidian artery anastomoses with theinternal maxillary artery. On occasion, the persistentstapedial branch of the petrous segment traverses abony canal and continues as the middle meningealartery.18 The cavernous portion, however, has farmore tributaries including the meningohypophysealtrunk, the anterior meningeal artery, the artery tothe inferior portion of the cavernous sinus, and theophthalmic artery. The meningohypophyseal trunkfurther subdivides into diminutive branches that in-clude the basal and marginal (artery of Bernasconiand Cassinari20) tentorial arteries, the inferior hy-pophyseal artery, and the dorsal meningeal artery.The inferolateral trunk arises from the inferolateralaspect of the cavernous ICA, supplying many smallbranches to the tentorium and trigeminal (V) nerve di-visions. Collateral anastomoses between the ICA andthe external carotid artery (ECA) are formed by theinferolateral trunk extending to the internal maxil-lary artery. The supraclinoid ICA also has numerousbranches including the superior hypophyseal perfo-rators to the anterior pituitary and stalk, posteriorcommunicating artery (PCoA), and anterior choroidalartery (AChA) before bifurcating into the ACA andMCA.21
The two ACAs connect through the anterior com-municating artery (ACoA) thus joining the left andright carotid circulations. The PCoA extends poste-riorly to connect with the primary segment of theposterior cerebral artery, allowing collateral flow topass between the anterior and posterior circulations.1
This vascular network, referred to as the circle ofWillis (Fig. 1.4), plays a critical role in shunting bloodflow between adjacent territories in the brain. At itsorigin, the PCoA often has a widened segment re-ferred to as the infundibulum. The PCoA passes ven-tral to the optic tract, with perforators that supplythe optic tract, posterior aspect of the chiasm, pos-terior hypothalamus, and anterior and ventral nucleiof the thalamus. In 15% of individuals, this vesselcontinues distally as the posterior cerebral artery.22,23
Great variability may be noted in the caliber of thePCoA, ranging from less than 1 mm to greater than2 mm. The anatomy of the PCoA differs in variouspopulations and in clinical conditions associated withischemia.23,24 Hypoplasia or absence of the PCoA is
Fig 1.4 Line drawing of anastomotic connections at thecircle of Willis, including the anterior communicating artery(A), the proximal or A1 segment of the anterior carotidartery (B), the posterior communicating artery (C), and theproximal or P1 segment of the posterior cerebral artery (D).
found in a minority of cases at autopsy, with bilateralhypoplasia in only 0.25% of individuals.22 The con-figuration and size of the PCoA also differs betweenrates gleaned from autopsy studies and angiographyseries.
There are numerous variant configurations of theICA, including its rare absence or hypoplasia. Theamount of blood volume supplied to distal structurescan vary depending on the caliber of the terminalICA. The course of the ICA sometimes varies, cours-ing through the middle ear or bending towards themidline in a configuration termed kissing ICAs at thecavernous segments. Anomalous origins of the poste-rior fossa arteries from the ICA, including the superiorcerebellar artery (SCA), anterior inferior cerebellarartery (AICA), or posterior inferior cerebellar artery(PICA), may also occur. Persistent fetal connectionsto the posterior circulation may involve the PCoA,trigeminal, otic or acoustic, hypoglossal, and proat-lantal intersegmental arteries. The persistent trigemi-nal artery is the most common persistent embryonicconnection (85%), arising from the cavernous ICAand joining the upper basilar artery.25 The persistentotic artery is very rare, connecting the petrous ICAwith the basilar artery inferior to AICA. The persis-tent hypoglossal connects the distal cervical ICA with
7

BLBK041-Kim September 22, 2008 17:30
CHAPTER 1
the distal vertebral artery. Intercavernous ICA collater-als may also allow for blood to flow laterally to eitherhemisphere.
Anterior choroidal artery
The AChA arises from the posterior aspect of theICA, about 2–4 mm distal to the origin of the PCoAand about 5 mm proximal to the carotid terminus.26
The AChA is relatively small, yet serves as an im-portant landmark in delineating important structuresat angiography.27 There are two segments of theAChA, including the cisternal and plexal segments.The AChA may have a single origin or may consistof several smaller vessels (4% of individuals).26,28 TheAChA arises from the MCA or PCoA in 2–11% ofindividuals.28 Complete absence of the AChA has alsobeen reported.27 The external diameter of this vesselis often only 0.5–1 mm, although a reciprocal rela-tionship has been noted in the caliber of this vesselwith the ipsilateral PCoA. The cisternal segment passesposteriorly from the lateral to medial aspect of the op-tic tract in close proximity to the posterior cerebralartery (PCA), extending for about 12 mm, extendingto a total length of about 26 mm. The AChA givesoff penetrating branches to the optic tract in this seg-ment. As the AChA courses posteriorly it gives offpenetrating branches to the globus pallidus and thegenu and posterior limb of the internal capsule. Sub-sequent branches extend laterally to supply the medialtemporal lobe cortex, hippocampal and dentate gyri,caudate, and amygdala. Medial branches supply thecerebral peduncle, substantia nigra, red nucleus, sub-thalamus, and ventral anterior and lateral nuclei of thethalamus. The AChA is the only branch of the ICA thatsupplies a portion of both the anterior and posteriorcirculation, although the midbrain and thalamic sup-ply is very variable. More distally, the AChA extendsthrough the choroidal fissure to become the plexal seg-ment. The juncture of the AChA at the choroidal fis-sure is often referred to as the plexal point. The plexalsegment then enters the choroid plexus near the pos-terior aspect of the temporal horn. Arterial supply ofthis segment includes the lateral geniculate body, opticradiations, and posterior limb of the internal capsule.The AChA anastomoses with lateral branches of theposterior choroidal artery, PCoA, PCA, and MCA.27,29
Variants include AChA origin from the PCoA or MCA.
Middle cerebral artery
The MCA provides arterial blood flow to the largestextent of the intracranial circulation. The MCA is typ-ically 75% of the caliber of the parent ICA.30 Af-ter diverging from the terminal ICA below the ante-rior perforated substance, it courses horizontally andslightly anteriorly to reach the sylvian fissure, wherebranches perfuse the frontal, parietal, and some ex-tent of the temporal and occipital cortices (Fig. 1.5).The proximal or horizontal segment of the MCA av-erages around 15 mm in length yet may be as longas 30 mm.30 At younger ages, the M1 segment risesobliquely but this segment tends to course more infe-riorly or anteriorly with increasing age later in life.31
Between the 7 and 12 mm (7 weeks) embryonic stage,small perforators that are precursors of the MCA maybe seen arising from the ICA. The MCA is smaller thanthe AChA at these early stages and then grows larger.During the second month of fetal life, the sylvian
Fig 1.5 Schematic of the middle cerebral artery, illustratingthe proximal segment or M1, lenticulostriate arteries (LSA),and bifurcation into M2, with downstream territoriesdelineated between adjacent anterior carotid artery andposterior cerebral artery regions.
8

BLBK041-Kim September 22, 2008 17:30
ANATOMY OF INTRACRANIAL ARTERIES
fissure develops as a groove over the cerebral hemi-sphere and the MCA grows within this depression.The MCA becomes enveloped in the sulcation of thecerebral cortex, following the growth of each specificbrain region.
The proximal or M1 division of the MCA provideslenticulostriate arteries that feed the globus pallidus,putamen, internal capsule, corona radiata, and cau-date nucleus. This segment is typically around 2.5 mmin internal diameter.30 These end arteries originatefrom the M1 segment in almost perpendicular fash-ion to penetrate the brain parenchyma. The laterallenticulostriates ascend for 2–5 mm posteromediallyfrom the M1 and then course laterally and superi-orly for an additional 9–30 mm to penetrate the in-ternal capsule. The medial lenticulostriates generallyarise from more proximal segments of the MCA orfrom distal reaches of the terminal ICA and proxi-mal ACA.32–35 There is considerable variation in therelative distribution and origins of medial versus lat-eral lenticulostriate perforators. The arterial diame-ter of lateral lenticulostriates is typically greater thanthe medial lenticulostriate perforators. Overall, thereare typically 5–17 lenticulostriate arteries, althoughall are barely identifiable at angiography.30 There arethree principal patterns that have been described forthe anatomy of the lenticulostriates.32–35 Grand et al.33
described a pattern where either one or more of thelarger lenticulostriates arise just beyond the MCA bi-furcation (49%), all arise proximal to the major bi-furcation (39%), and a minority of cases where someof the larger perforators arise from the medial por-tion of the stem. According to Jain,34 54.1% orig-inate from the MCA trunk, 25.6% from the divi-sion point, and 20.3% from one of the branches ofthe MCA. The lateral lenticulostriates supply the lat-eral portion of the anterior commissure, the putamen,the lateral segment of the globus pallidus, the supe-rior half of the internal capsule, the adjacent coronaradiate, and the body and head of the caudate nu-cleus. The medial lenticulostriates arise in perpendic-ular fashion to the parent MCA or ACA, yet bendin mesial fashion. The areas supplied by the me-dial lenticulostriates, including the prominent recur-rent artery of Heubner, and the AChA are adjacentto the territories of the lateral lenticulostriates. Therelative territorial extents are reciprocal in size anddepend on the development of each of these arterialgroups.32–35
The largest branch of the proximal MCA is the an-terior temporal artery, extending from the middle ofthe proximal MCA and winding anteriorly and in-feriorly. Although the configuration of the proximalMCA often varies, the vessel most often splits intotwo or more principal divisions near the sylvian fis-sure. Although prior studies have suggested symmetryin the morphology of bilateral MCAs, no clear corre-lations exist. The anterior and posterior divisions ofthe MCA extend into the sylvian fissure and spreadout over the hemisphere. These cortical branches in-clude the temporopolar, frontobasal, operculofrontal,precentral, postcentral, posterior parietal, angular, an-terior temporal, middle temporal, and posterior tem-poral arteries. As the MCA branches loop over theinsula in the sylvian fissure, they form the sylvian tri-angle, a landmark classically used to identify mass le-sions on angiography. Terminal branches of the MCAform collateral anastomoses with the ACA and PCA.1
Variation in MCA anatomy is less common thanvariants in other intracranial arteries. Fenestrationof the M1 segment occurs, and duplicated M1 seg-ments may also arise from the ICA.36 Angiographicdemonstration of MCA fenestration may be evidentin approximately 0.26% of individuals.37 Yamamotoet al.38 described 14 accessory MCAs and seven du-plicated MCAs in a series of 455 bilateral carotidangiographies. The M1–M2 junction is characterizedby a bifurcation in 64–90% of individuals, trifurca-tion in 12–29%, and complex branching in isolatedindividuals.30 Some controversy has surrounded spe-cific landmarks and associated classification of theMCA segments. Whereas many identify the segmentsof the MCA based on each successive branch point,others use a nomenclature that relates each of theseMCA segments with a specific adjacent anatomicalstructure. For instance, some refer to the M2 originat the initial bifurcation of the proximal or M1 MCA,whereas others identify the M2 segment as the arterialsegment that overlies the insula.
Anterior cerebral artery
The ACA develops from residual elements of the prim-itive olfactory artery at the terminus of the ICA. Thepaired primitive olfactory arteries from each side forma plexus in the midline that gives rise to the ACoA.During development, the ACA extends superiorly and
9

BLBK041-Kim September 22, 2008 17:30
CHAPTER 1
Fig 1.6 CT angiography illustration of two differentconfigurations in the anterior carotid artery (ACA) complexand anterior communicating artery (ACoA) anastomosis.
(A) A patent ACoA provides interhemispheric flow betweenleft and right ACAs. (B) ACoA is absent, yet a prominentrecurrent artery of Heubner (RAH) is demonstrated.
then posteriorly over the hemispheres in the midlinewhereas the remainder of the primitive olfactory arteryregresses to become a small perforating vessel. TheACA is typically 50% of the caliber of the parentICA.39 The internal diameter of the A1 is usually 0.9–4 mm, with hypoplasia defined as a diameter less than1 mm. The A1 segment measures 7–18 mm, with anaverage span of 12.7 mm.39 The ACA extends antero-medially between the optic chiasm (70% of individu-als) or optic nerve (30% of individuals) and the an-terior perforated substance to join the contralateralACA through an anastomosis via the ACoA.
The ACoA forms the anterior aspect of the circle ofWillis, a critical route for collateral flow between thecerebral hemispheres. The ACoA is the shortest cere-bral artery, measuring only 0.1–3 mm in length.39 Theanatomy of the ACA–ACoA is variable (Fig. 1.6) withhypoplasia of different segments, including absence ofthe ACoA. Accessory routes, fenestrations, and othercomplex azygous connections between the proximalACAs are also described.
The proximal ACA or A1 segment gives off numer-ous perforating arteries that supply the adjacent opticnerves and chiasm inferiorly, and the hypothalamus,septum pellucidum, anterior commissure, fornix, andcorpus striatum. These mesial lenticulostriate vesselsoften include a prominent recurrent artery of Heubner
that supplies the caudate head, putamen, and anteriorlimb of the internal capsule.40 The A2 segment be-gins at the juncture of the ACA with the ACoA andextends to the genu of the corpus callosum. The re-current artery of Heubner arises from the A2 segmentin 49–78% of individuals.39 The recurrent artery ofHeubner may be a single vessel or can be representedby a number of parallel arteries. Beyond the proximalsegment of the ACA, azygous connections allow forshunting of flow between the cerebral hemispheres.The ACAs course over the cerebral hemispheres in theinterhemispheric fissure as paired vessels, with theirdistal extent typically determined by the correspond-ing anatomy of the PCAs. Subsequent divisions in-cluding the pericallosal and callosomarginal arteriesdivide to provide arterial supply to the corpus callo-sum and anteromesial cortices. Several variations indistal ACA anatomy have been described, includingthe observation that the left pericallosal artery is lo-cated more posteriorly than the corresponding right-sided vessel in 72%.41 Similarly, absence of the callo-somarginal artery has been noted in 18–60% of casesstudied.41
Cortical branches of the ACA include the or-bitofrontal, frontopolar, callosomarginal, and perical-losal arteries. As the terminal portion of the ACA trav-els along the corpus callosum, its anterior pericallosal
10

BLBK041-Kim September 22, 2008 17:30
ANATOMY OF INTRACRANIAL ARTERIES
branches form anastomoses with the posterior perical-losal branches of the PCA.41
Variant anatomy of the ACA most commonly in-cludes hypoplasia or absence of the A1 segment(10% of individuals).39 Other variations may includeanomalous origin of the ACA from the ICA, agenesisor accessory branches, direct connection of bilateralA1 segments, or other combinations that involve azy-gous orientation of distal ACA segments.
Vertebral artery
The vertebral artery enters the skull at the level of C1through the foramen magnum. The intracranial or in-tradural (V4) segment of the vertebral artery ascendsanteriorly to the medulla, approaching the midline atthe pontomedullary junction, where it meets the con-tralateral vertebral artery to form the basilar artery.The paired longitudinal arteries that form the arterialsupply to the posterior circulation during early fetaldevelopment retain their proximal course as the ver-tebral arteries. The left vertebral artery is larger thanthe right 42% of the time, whereas the right is largerthan the left 32% of the time. In the remainder of indi-viduals, the vertebral arteries are equivalent in caliber.Vertebral artery hypoplasia is fairly common, ofteninvolving the right side. The frequency of this find-ing depends largely on the modality used to image thevessel, the size threshold used to define hypoplasia,and the study population, including healthy subjectsor patients with ischemic stroke. Defining hypoplasiaas ≤2 mm by ultrasonography, one group reported afrequency of 1.9% in 451 subjects.42 Among healthysubjects with a threshold of <3 mm, another groupnoted a frequency of 6% in 50 healthy subjects.43 Oth-ers have recently noted a frequency as high as 35.2%in 529 patients with ischemic stroke.44 Utilizing a lu-minal diameter threshold of 2.2 mm, prominent asym-metry in vertebral artery hypoplasia has also been de-scribed (7.8% on the right and 3.8% on the left) in447 subjects.45 Differentiation of hypoplasia from adiseased arterial segment may be difficult to define onthe basis of luminal dimensions alone. The configu-ration or compensatory enlargement of neighboringsegments may provide clues to this distinction. Fur-thermore, definitions may vary considerably depend-ing on the imaging technique employed. The vertebralartery may also terminate in the PICA rather than ex-
tend to the junction with the basilar artery. In suchcases, the vertebral artery is generally smaller than thecontralateral vertebral artery.
The terminal vertebral artery yields several branchesthat supply the rostral end of the spinal cord andposterior inferior aspect of the cerebellum. Anteriorand posterior spinal arteries extend from this segment.Each anterior spinal artery fuses with its counterpart,supplying the ventral medulla and rostral spinal cord.The posterior spinal arteries do not pair in the midline,but descend the spinal cord at the level of the dorsalroots. The posterior inferior cerebellar artery branchesfrom the vertebral artery to supply the inferior aspectof the cerebellum.
Posterior inferior cerebellar artery
The largest tributary of the vertebral artery is thePICA, arising 10–20 mm before the vertebrobasilarjunction (Fig. 1.7).46 In 20% of individuals, the PICAarises from below the foramen magnum.47 Duringembryogenesis, the PICA may be evident as a largerbranch of numerous arteries that extend posteriorlyfrom the hindbrain in the 20–24 mm embryo stage.
Fig 1.7 Digital subtraction angiography of the posteriorinferior cerebellar artery (PICA), showing proximalmedullary segments and hemispheric tributaries.
11

BLBK041-Kim September 22, 2008 17:30
CHAPTER 1
At later stages, this vessel continues to predominate,growing into the largest arterial offshoot. There arefour segments of this vessel, including the anterior,lateral, posterior medullary, and supratonsillar PICA.The anterior medullary segment travels laterally nearthe inferior aspect of the olive of the medulla oblan-gata, continuing in a loop that courses between thecerebellum and medulla. Numerous perforating arter-ies extend from the first three segments of the PICAto supply anterior, lateral, and posterior aspects of themedulla. The PICA then extends posteriorly in the ton-sillomedullary fissure adjacent to the glossopharyn-geal (IX) and vagus (X) nerves. Beyond this point, thePICA curves over the cerebellar tonsil to becomethe supratonsillar segment extending further acrossthe cerebellum as the medial and lateral terminal PICAbranches. At the juncture of the posterior medullaryand supratonsillar segments of the PICA, perforatingvessels arise to feed the choroid plexus of the fourthventricle. This choroidal point is used as a landmarkto identify masses within the posterior fossa.
Variations in PICA anatomy include hypoplasia orabsence of this branch (10–20% of individuals), typ-ically accompanied by a prominent ipsilateral ante-rior inferior cerebellar artery. Absence of the PICA isalso accompanied by numerous medullary perforatorsthat arise directly from the vertebral artery. Duplica-tion of the PICA and double origin with distal arterialconvergence of the PICA48 may occur. PICA–AICAconnections and other anomalies may be seen, yet thespecific frequencies of such anatomic configurationsremain unclear. On occasion, both inferior cerebellarterritories may be supplied by a bihemispheric PICAoriginating from one vertebral artery.49
Basilar artery
The basilar artery extends from the confluence of thevertebrals near the pontomedullary junction to the ter-minal bifurcation as the PCAs at the level of the mid-brain. During embryologic development, paired ves-sels on the ventral surface of the hindbrain fuse to formthe basilar artery. Distal segments of these paired basi-lar arteries have connections with the ipsilateral ICA.Over time, the plexus formed by the paired basilararteries fuses with progressive disappearance of fen-estrations. The basilar artery is often tortuous or ser-pentine, with a straight course noted about 25% of the
Fig 1.8 Schematic of posterior circulation, demonstratingcerebellar arteries including superior cerebellar artery (SPA),anterior inferior cerebellar artery (AICA), and posteriorinferior cerebellar artery (PICA).
time. Curvature of the basilar as it ascends has been as-sociated with the presence of a wider contralateral ver-tebral artery, suggesting hemodynamic modeling. Thelength of the basilar artery is consistently 25–35 mm,irrespective of body size.50 The diameter is about 2.7–4.3 mm at the proximal portion. The luminal diameterof the basilar artery tends to taper towards the distalend.
The largest branches of the basilar artery include theanterior inferior cerebellar artery (AICA) and superiorcerebellar artery (SCA) (Fig. 1.8). Numerous smallerperforators embrace the brainstem, coursing from themidline ventral aspect around the surface to the lat-eral dorsal surface, and diving deep into the substanceof the brainstem between fiber tracts. These pontineperforators are grouped into medial and lateral subdi-visions, often referred to as paramedian and circum-ferential arteries. Lateral pontine perforators extendto also supply the ventrolateral surface of the cerebel-lum, whereas the medial perforators perfuse midlinestructures of the midbrain. There is a somewhat down-ward trajectory of brainstem perforating arteries sothat the most rostral portion of the basilar artery sup-plies penetrators to the pontine tegmentum. The inter-nal auditory or labyrinthine artery may arise from thebasilar to provide arterial blood flow to the cochlea,labyrinth, and facial (VII) nerve.51 Alternatively, thisartery may arise as a branch of the AICA. Owing tothe paired structure of posterior circulation arteries,
12

BLBK041-Kim September 22, 2008 17:30
ANATOMY OF INTRACRANIAL ARTERIES
asymmetries or relative dominance of one artery suchas the PICA, AICA, or SCA may occur. Contralateralcerebellar infarction may therefore result from occlu-sion or disease of one cerebellar artery. The frequen-cies of such patterns vary and are difficult to estimate,although the advent of MRI and MRA allows for sys-tematic evaluation of such relatively subtle features.Terminal branches of the PICA, AICA, and SCA formanastomoses that allow for collateral flow to easilyshift between their arterial territories.1,52
Anterior inferior cerebellar artery
The AICA extends off of the basilar artery approxi-mately one-third to halfway through its course.52 TheAICA arises from the caudal third of the basilar 74%of the time.52 This artery is the smallest of the principalcerebellar arteries. During development, the AICA ap-pears as one of the larger perforatoring vessels extend-ing to the posterior aspect of the hindbrain. The AICAextends laterally and inferiorly, in close proximity tothe abducens (VI) nerve. Similar to the lateral pontineperforators, it courses around the brainstem and thenenters the cerebellopontine angle cistern along with thefacial (VII) and vestibulocochlear (VIII) nerves. TheAICA crosses the anteroinferior aspect of the cerebel-lum to supply the middle cerebellar peduncle, floc-culus, and adjacent cerebellum. The AICAs supplya rather small variable portion of the anterior infe-rior cerebellum.53 The supply to the brachium pontisand flocculus is consistent. Numerous pontine perfora-tors may arise from its proximal segment. The lateralbranch runs across the cerebellum in the horizontalfissure. The medial branch of the AICA courses inferi-orly to supply the biventral lobule. Similar to absenceof the PICA, the AICA may be absent or hypoplasticand is typically accompanied by a prominent PICA.Variable infarct patterns may be noted in these re-gions of the posterior circulation, likely reflecting arte-rial configurations of the principal cerebellar arteriesthat may involve hypoplastic segments or anomalousanastomoses.53,54 Combined or multiple territorial in-farcts such as the PICA and AICA or AICA and SCAmay reflect dominant patterns of arterial supply orig-inating from the vertebral and basilar arteries. Simi-larly, collateral anastomoses between these territoriesmay provide sufficient arterial inflow to spare distalsegments of a particular arterial territory. As individ-
ual anatomy is often studied only after the onset ofstroke, the original arterial configuration and mech-anistic events may only be surmised. Similarly, thefrequency of specific arterial supply patterns may bedifficult to ascertain in the healthy population as indi-viduals are most often studied after presentation withpotential neurovascular disorders.
Superior cerebellar artery
The SCA also extends from the basilar in symmetricfashion, just proximal to the terminal bifurcation ofthe basilar into the proximal PCAs. SCA morphologyis more consistent across individuals than other cere-bellar branches.55 The SCA courses laterally belowthe oculomotor (III) nerve, passing around the cere-bral peduncles and below the trochlear (IV) nerve.28
Numerous perforators extend from the proximal orambient SCA to supply the adjacent pons and mid-brain, whereas distal segments split into the lateralmarginal and superior vermian branches. These divi-sions may also arise independently from the basilarartery or even the PCA. Duplication of the SCA isnoted in 28% of individuals, with bilateral duplicationin 10%.56 The SCA variably divides into medial SCAand lateral SCA branches. The lateral marginal SCAsupplies the anterosuperior cerebellum, superior cere-bellar peduncle, middle cerebral peduncle, and dentatenuclei. The superior vermian SCA supplies the supe-rior cerebellar peduncle, tentorium, inferior colliculi,cerebellar hemispheres, and dentate nuclei. Anasto-moses between this branch and the inferior vermianbranch of the PICA allow for robust collateral perfu-sion across the cerebellar hemispheres.57
Posterior cerebral artery
The PCA develops from fusion of several vessels thatsupply the mesencephalon, diencephalon, and choroidplexus in the fetus.58 These vessels stem from the termi-nal aspect of the PCoA at the distal end of the carotidcirculation. The PCA most often then extends posteri-orly to spread over the ipsilateral cortex, whereas theproximal connection with the PCoA regresses. In thiscommon scenario, the primary arterial supply shiftsto a source from the terminal basilar artery. In theremainder of individuals, the PCA supply continues
13

BLBK041-Kim September 22, 2008 17:30
CHAPTER 1
Fig 1.9 Digital subtraction angiography illustratingposterior cerebral artery anastomosis with posteriorcommunicating artery, thalamoperforators, and distalcortical branches.
from what has been termed a fetal PCoA. Variants ofPCoA anatomy include a diverse range of caliber inthis segment, complete agenesis, and anomalous ori-gins of other vessels from this arterial segment.59
The PCA extends from the terminal portion of thebasilar artery in the interpeduncular cistern, passingabove the oculomotor (III) nerve to circle the midbrainabove the tentorium (Fig. 1.9). As it passes through thepeduncular, ambient, and quadrigeminal cisterns, nu-merous perforators supply adjacent structures.60 Thispattern of arterial limbs includes paramedian perfo-rators, short circumferential and long circumferentialbranches that typify the general structure of the ma-jor arterial territories in the posterior circulation. Theperforating arteries from these segments range from200 μm to 800 μm in diameter.60 The artery of David-off and Schechter extends from the P1 segment tosupply part of the inferior surface of the tentorium.The midbrain receives arterial blood from the pedun-cular or P1 segment before posterior thalamoperfo-rators arise. In the successive ambient segment, thethalamogeniculate arteries diverge to supply the lat-eral geniculate and pulvinar nuclei. Medial and lateral
branches of the posterior choroidal arteries extendfrom this portion of the PCA to supply the pinealgland, third ventricle, dorsomedial thalamus, pulvinar,lateral geniculate body, and choroid plexus.61 Thesedistal posterior choroidal arteries form anastomoseswith the AChA.
These deep arterial territories composed of perfora-toring arterioles that encompass the thalamus are of-ten difficult to comprehend because of their complexconfiguration (Fig. 1.10).62 The P1 or proximal PCAserves as an important arterial segment in this config-uration, with variable contributions from the basilarartery, PCoA, and AChA.63 These vessels have perfora-tors that supply these critical structures at the juncturebetween the anterior and posterior circulations. Theanterior thalamoperforating arteries consist of about7–10 branches that arise from the superior and lateralsurfaces of the PCoA.63 A larger branch, the premamil-lary artery, is often noted.64 This vessel courses fromthe posterior aspect of the PCoA, penetrates the hy-pothalamus and subsequently terminates in branchesthat supply the anterior and ventroanterior nucleiof the thalamus. Further posteriorly, the thalamus is
Fig 1.10 Schematic of the arterial supply to the thalamus,illustrating premamillary arteries (A), interpeduncularperforators (B), thalamogeniculate arteries (C), posteriorchoroidal arteries (PCoA) (D), and anterior choroidal artery(AChA) (E). PCA, posterior cerebral artery.
14

BLBK041-Kim September 22, 2008 17:30
ANATOMY OF INTRACRANIAL ARTERIES
supplied by a combination of vessels arising from theposterior circulation. Interpeduncular branches fromthe basilar artery and P1 segment ascend superiorly toperfuse mesial aspects of the thalamus. More lateralaspects of the thalamus, including the ventropostero-medial and ventroposterolateral nuclei are supplied bythe thalamogeniculate arteries.65 These typically con-sist of 3–5 small branches that penetrate the inferioraspect of the thalamus and also supply blood to thegeniculate bodies.60 The most posterior reaches of thethalamus are supplied via the posterior choroidal ar-teries, arising from more distal aspects of the subcor-tical PCA. These posterior choroidal branches supplyportions of the medial nuclei, habenula, and rostro-medial pulvinar, and choroid plexus.66 The posteriorchoroidal arteries adjoin and overlap to some extentwith distal reaches of the AChA, extending from theICA to supply lateral and posterior regions of thethalamus.
The PCA passes along the free edge of the tentoriumto eventually reach the medial aspect of the occipitallobe. Further branching within the hippocampal fis-sure produces cortical divisions that course over theinferior and mesial aspects of the hemisphere. The cor-tical territories of the PCA are supplied via the anteriorand posterior divisions, fanning out to follow the ar-chitecture of the cortical surface in these regions. Cor-tical branches of the PCA include the hippocampal, an-terior temporal, middle temporal, posterior temporal,parieto-occipital, calcarine, and posterior pericallosalor perisplenial arteries.67 Anastomoses of the PCA al-low for collateral flow into the MCA via the anteriorand posterior temporal arteries and into the ACA viapericallosal divisions that arise from the quadrigemi-nal PCA.1
Conclusions
Anatomy and physiology, or pathophysiology such asintracranial atherosclerosis, are inextricably linked.Knowledge of abnormal or pathological conditionssuch as intracranial atherosclerosis stems from de-tailed recognition of the normal or baseline patternof vascular anatomy. The vascular anatomy of the in-tracranial arteries, at the level of the vessel wall and asa larger structure or conduit, is a reflection of physiol-ogy over time, from in utero stages through adult life.The embryologic development of these structures is
therefore pivotal, for understanding the flux of carotidsupply to the hindbrain that ultimately determinesPCoA morphology or for deciphering the complex pat-tern of evolving cortical architecture that shapes distalarterial territories. Variant anatomy may also be un-derstood from this perspective, as persistence of em-bryologic variants typically reflects ongoing flow re-quirements. The unique characteristics of arteries atthe base of the brain may help our understanding ofatherosclerotic lesions that tend to afflict specific arte-rial segments.
Replacement of smooth muscle cells in the arterialmedia with other cellular components or focal in-creases in fluid shear stress because of vessel mor-phology may hasten disease. Similarly, medical andendovascular therapeutic strategies may be refinedthrough consideration of this unique anatomicalsystem. Although much of the knowledge regardingintracranial arteries originates from pathology andangiography series over several centuries, evolvingnoninvasive techniques have rapidly expanded ourperspective on these key arterial segments. As eachmodality provides a depiction that combines anatomyand flow physiology, it is important to interpret eachimage with a solid understanding of typical arterialanatomy. Ongoing studies that illustrate the anatomyand pathophysiology of these proximal arterial seg-ments across modalities will help refine our knowl-edge of the interplay between vascular anatomy andcerebral blood flow. Such descriptions will likely ex-tend far beyond the current standard of characterizinga vascular lesion solely based on the degree of luminalstenosis.
Although distal arterial segments have not been em-phasized in this chapter, collateral anastomoses at suchsites may greatly influence disease management of in-tracranial atherosclerosis in the proximal intracranialarteries. Collateral flow should theoretically be incon-sequential or nonexistent if an atherosclerotic lesion isnot hemodynamically significant, determined by a crit-ical threshold around 60–70%. Nevertheless, anecdo-tal descriptions relate findings of collateral flow witheven slight or moderate stenoses. Future studies mayhelp elucidate pivotal arterial factors far beyond thedegree of stenosis, examining downstream influenceson cerebral perfusion, artery-to-artery thromboem-bolic potential, amenability to endovascular therapiesand stent conformation, or propensity for restenosisdue to biophysical factors.
15

BLBK041-Kim September 22, 2008 17:30
CHAPTER 1
References
1 Liebeskind DS. Collateral circulation. Stroke 2003; 34:2279–2284.
2 Parmar H, Sitoh YY, Hui F. Normal variants of the in-tracranial circulation demonstrated by MR angiographyat 3T. Eur J Radiol 2005; 56: 220–228.
3 Feldmann E, Wilterdink JL, Kosinski A, et al. TheStroke Outcomes and Neuroimaging of IntracranialAtherosclerosis (SONIA) trial. Neurology 2007; 68:2099–2106.
4 Stehbens WE. Pathology of the cerebral blood vessels. StLouis, MO: CV Mosby 1972.
5 Walmsley JG, Canham PB. Orientation of nuclei as in-dicators of smooth muscle cell alignment in the cerebralartery. Blood Vessels 1979; 16: 43–51.
6 Walmsley JG. Vascular smooth muscle orientation incurved branches and bifurcations of human cerebral ar-teries. J Microsc 1983; 131: 377–389.
7 Walmsley JG. Vascular smooth muscle orientation instraight portions of human cerebral arteries. J Microsc1983; 131: 361–375.
8 Farrall AJ, Wardlaw JM. Blood–brain barrier: ageingand microvascular disease – systematic review and meta-analysis. Neurobiol Aging (in press).
9 Bang OY, Buck BH, Saver JL, et al. Prediction of hemor-rhagic transformation after recanalization therapy usingT2∗-permeability magnetic resonance imaging. Ann Neu-rol 2007; 62: 170–176.
10 Oldendorf WH, Cornford ME, Brown WJ. The large ap-parent work capability of the blood–brain barrier: a studyof the mitochondrial content of capillary endothelial cellsin brain and other tissues of the rat. Ann Neurol 1977; 1:409–417.
11 McCarron RM, Chen Y, Tomori, T, et al. Endothelial-mediated regulation of cerebral microcirculation. J Phys-iol Pharmacol 2006; 57 (Suppl 11): 133–144.
12 Hassler O. The windows of the internal elastic lamella ofthe cerebral arteries. Virchows Arch Pathol Anat PhysiolKlin Med 1962; 335: 127–132.
13 Takayanagi T, Rennels ML, Nelson E. An electron micro-scopic study of intimal cushions in intracranial arteries ofthe cat. Am J Anat 1972; 133: 415–429.
14 Heil M, Eitenmuller I, Schmitz-Rixen T, Schaper W.Arteriogenesis versus angiogenesis: similarities and dif-ferences. J Cell Mol Med 2006; 10: 45–55.
15 Schaper W, Scholz D. Factors regulating arteriogenesis.Arterioscler Thromb Vasc Biol 2003; 23: 1143–1151.
16 Zervas NT, Liszczak TM, Mayberg MR, Black PM. Cere-brospinal fluid may nourish cerebral vessels through path-ways in the adventitia that may be analogous to systemicvasa vasorum. J Neurosurg 1982; 56: 475–481.
17 Clower BR, Sullivan DM, Smith RR. Intracranial vesselslack vasa vasorum. J Neurosurg 1984; 61: 44–48.
18 Quisling RG, Rhoton AL, Jr. Intrapetrous carotid arterybranches: radioanatomic analysis. Radiology 1979; 131:133–136.
19 Allen JW, Alastra AJ, Nelson PK. Proximal intracranialinternal carotid artery branches: prevalence and impor-tance for balloon occlusion test. J Neurosurg 2005; 102:45–52.
20 Frugoni P, Nori A, Galligioni F, Giammusso V. Furtherconsiderations on the Bernasconi and Cassinari’s arteryand other meningeal rami of the internal carotid artery.Neurochirurgia (Stuttg) 1964; 108: 18–23.
21 Gibo H, Lenkey C, Rhoton AL, Jr. Microsurgicalanatomy of the supraclinoid portion of the internalcarotid artery. 1981; J Neurosurg 55: 560–574.
22 Alpers BJ, Berry RG, Paddison RM. Anatomical studiesof the circle of Willis in normal brain. AMA Arch NeurolPsychiatry 1959; 81: 409–418.
23 Schomer DF, Marks MP, Steinberg GK, et al. The anatomyof the posterior communicating artery as a risk factor forischemic cerebral infarction. N Engl J Med 1994; 330:1565–1570.
24 Eftekhar B, Dadmehr M, Ansari S, Ghodsi M, NazparvarB, Ketabchi E. Are the distributions of variations of circleof Willis different in different populations? Results of ananatomical study and review of literature. BMC Neurol2006; 6: 22.
25 Uchino A, Kato A, Takase Y, Kudo S. Persistent trigeminalartery variants detected by MR angiography. Eur Radiol2000; 10: 1801–1804.
26 Rhoton AL, Jr, Fujii K, Fradd B. Microsurgical anatomyof the anterior choroidal artery. Surg Neurol 1979; 12:171–187.
27 Takahashi S, Suga T, Kawata Y, Sakamoto K. Anteriorchoroidal artery: angiographic analysis of variations andanomalies. AJNR Am J Neuroradiol 1990; 11: 719–729.
28 Saeki N, Rhoton AL, Jr. Microsurgical anatomy of theupper basilar artery and the posterior circle of Willis. JNeurosurg 1977; 46: 563–578.
29 Morandi X, Brassier G, Darnault P, Mercier P, Scara-bin JM, Duval JM. Microsurgical anatomy of the ante-rior choroidal artery. Surg Radiol Anat 1996; 18: 275–280.
30 Gibo H, Carver CC, Rhoton AL, Jr, Lenkey C, MitchellRJ. Microsurgical anatomy of the middle cerebral artery.J Neurosurg 1981; 54: 151–169.
31 Bouissou H, Emery MC, Sorbara R. Age related changesof the middle cerebral artery and a comparison with theradial and coronary artery. Angiology 1975; 26: 257–268.
16

BLBK041-Kim September 22, 2008 17:30
ANATOMY OF INTRACRANIAL ARTERIES
32 Donzelli R, Marinkovic S, Brigante L, et al. Territoriesof the perforating (lenticulostriate) branches of the mid-dle cerebral artery. Surg Radiol Anat 1998; 20: 393–398.
33 Grand W. Microsurgical anatomy of the proximal middlecerebral artery and the internal carotid artery bifurcation.Neurosurgery 1980; 7: 215–218.
34 Jain KK. Some observations on the anatomy of the middlecerebral artery. Can J Surg 1964; 7: 134–139.
35 Marinkovic SV, Milisavljevic MM, Kovacevic MS, StevicZD. Perforating branches of the middle cerebral artery.Microanatomy and clinical significance of their intracere-bral segments. Stroke 1985; 16: 1022–1029.
36 Komiyama M, Nakajima H, Nishikawa M, Yasui T.Middle cerebral artery variations: duplicated and ac-cessory arteries. AJNR Am J Neuroradiol 1998; 19:45–49.
37 Lazar ML, Bland JE, North RR, Bringewald PR. Middlecerebral artery fenestration. Neurosurgery 1980; 6: 297–300.
38 Yamamoto H, Marubayashi T, Soejima T, MatsuokaS, Matsukado Y, Ushio Y. Accessory middle cerebralartery and duplication of middle cerebral artery –terminology, incidence, vascular etiology, and develop-mental significance. Neurol Med Chir (Tokyo) 1992; 32:262–267.
39 Perlmutter D, Rhoton AL, Jr. Microsurgical anatomy ofthe anterior cerebral-anterior communicating-recurrentartery complex. J Neurosurg 1976; 45: 259–272.
40 Pearce JM. Heubner’s artery. Eur Neurol 2005; 54: 112–114.
41 Perlmutter D, Rhoton AL, Jr. Microsurgical anatomy ofthe distal anterior cerebral artery. J Neurosurg 1978; 49:204–228.
42 Delcker A, Diener HC. [Various ultrasound methods forstudying the vertebral artery – comparative evaluation].Ultraschall Med 1992; 13: 213–220.
43 Touboul PJ, Bousser MG, LaPlane D, Castaigne P. Duplexscanning of normal vertebral arteries. Stroke 1986; 17:921–923.
44 Park JH, Kim JM, Roh JK. Hypoplastic vertebral artery:frequency and associations with ischaemic stroke ter-ritory. J Neurol Neurosurg Psychiatry 2007; 78: 954–958.
45 Jeng JS, Yip PK. Evaluation of vertebral artery hypoplasiaand asymmetry by color-coded duplex ultrasonography.Ultrasound Med Biol 2004; 30: 605–609.
46 Lister JR, Rhoton AL, Jr, Matsushima T, Peace DA. Mi-crosurgical anatomy of the posterior inferior cerebellarartery. Neurosurgery 1982; 10: 170–199.
47 Fine AD, Cardoso A, Rhoton AL, Jr. Microsurgicalanatomy of the extracranial-extradural origin of the
posterior inferior cerebellar artery. J Neurosurg 1999; 91:645–652.
48 Lesley WS. and Dalsania HJ. Double origin of the pos-terior inferior cerebellar artery. AJNR Am J Neuroradiol2004; 25: 425–427.
49 Cullen SP, Ozanne A, Alvarez H, Lasjaunias P. The bi-hemispheric posterior inferior cerebellar artery. Neurora-diology 2005; 47: 809–812.
50 Brassier G, Morandi X, Riffaud L and Mercier P. Basilarartery anatomy. J Neurosurg 2000; 93: 368–369.
51 Kim JS, Lopez I, DiPatre PL, Liu F, Ishiyama A, BalohRW. Internal auditory artery infarction: clinicopathologiccorrelation. Neurology 1999; 52: 40–44.
52 Rhoton AL, Jr. The cerebellar arteries. Neurosurgery2000; 47: S29–68.
53 Kumral E, Kisabay A, Atac C. Lesion patterns and etiol-ogy of ischemia in the anterior inferior cerebellar arteryterritory involvement: a clinical – diffusion weighted –MRI study. Eur J Neurol 2006; 13: 395–401.
54 Terao S, Miura N, Osano Y, et al. Multiple cerebellarinfarcts: clinical and pathophysiologic features. J StrokeCerebrovasc Dis 2005; 14: 193–198.
55 Hardy DG, Peace DA, Rhoton AL, Jr. Microsurgicalanatomy of the superior cerebellar artery. Neurosurgery1980; 6: 10–28.
56 Icardo JM, Ojeda JL, Garcia-Porrero JA, Hurle JM. Thecerebellar arteries: cortical patterns and vascularizationof the cerebellar nuclei. Acta Anat (Basle) 1982; 113:108–116.
57 Brandt T, von Kummer R, Muller-Kuppers M, Hacke W.Thrombolytic therapy of acute basilar artery occlusion.Variables affecting recanalization and outcome. Stroke1996; 27: 875–881.
58 Zeal AA. and Rhoton AL, Jr. Microsurgical anatomy ofthe posterior cerebral artery. J Neurosurg 1978; 48: 534–559.
59 Bisaria KK. Anomalies of the posterior communicatingartery and their potential clinical significance. J Neuro-surg 1984; 60: 572–576.
60 Milisavljevic MM, Marinkovic SV, Gibo H, Puskas LF.The thalamogeniculate perforators of the posterior cere-bral artery: the microsurgical anatomy. Neurosurgery1991; 28: 523–529; Discussion 9–30.
61 Fujii K, Lenkey C, Rhoton AL, Jr. Microsurgical anatomyof the choroidal arteries: lateral and third ventricles. JNeurosurg 1980; 52: 165–188.
62 Carrera E, Michel P, Bogousslavsky J. Anteromedian, cen-tral, and posterolateral infarcts of the thalamus: threevariant types. Stroke 2004; 35: 2826–2831.
63 Percheron G. [Arteries of the thalamus in man. Choroidalarteries. III. Absence of the constituted thalamic territoryof the anterior choroidal artery. IV. Arteries and thalamic
17

BLBK041-Kim September 22, 2008 17:30
CHAPTER 1
territories of the choroidal and postero-median thalamicarterial system. V. Arteries and thalamic territories of thechoroidal and postero-lateral thalamic arterial system].Rev Neurol (Paris) 1977; 133: 547–558.
64 Gibo H, Marinkovic S, Brigante L. The microsurgicalanatomy of the premamillary artery. J Clin Neurosci2001; 8: 256–260.
65 Georgiadis AL, Yamamoto Y, Kwan ES, Pessin MS, Ca-plan LR. Anatomy of sensory findings in patients with
posterior cerebral artery territory infarction. Arch Neurol1999; 56: 835–838.
66 Liebeskind DS, Hurst RW. Infarction of the choroidplexus. AJNR Am J Neuroradiol 2004; 25: 289–290.
67 Brandt T, Steinke W, Thie A, Pessin MS, Caplan LR. Pos-terior cerebral artery territory infarcts: clinical features,infarct topography, causes and outcome. Multicenter re-sults and a review of the literature. Cerebrovasc Dis 2000;10: 170–182.
18