penicillins and activity of nitrogen metabolism enzymes in plant tissue cultures
TRANSCRIPT

Plant Science, 59 {1989) 119-- 125 119 Elsevier Scientific Publishers Ireland Ltd.
PENICILLINS AND ACTIVITY OF NITROGEN METABOLISM ENZYMES IN PLANT TISSUE CULTURES
ISABEL SANTOS and R. SALEMA
Institute of Botany and Centre of Experimental Cytology (INIC), University of Porto, Rua do Campo Alegre, 823, P-~IO0 Porto ~PortugaI)
{Received April 11th, 1988) {Revision received July 11th, 1988) (Accepted August 25th, 1988~
Since plant tissues grown in vitro are not indifferent to antibiotics added to the culture medium, effects of penicillin G (PG) and V (PV) on the uptake and metabolism of nitrogen were studied by following levels of activity of nitrate reductase (NR), glu- tamate dehydrogenase (GDH), glutamine synthetase (GS), glutamate syntbase (GOGAT) and the fate of NO~ in the culture media, using callus tissue cultured during 14 days. In comparison to control both NR and GDH showed some degree of stimula- tion: higher levels of GS than in control were found up to day 10, with a reverse situation from that point till the end of treat- ment; activity of GOGAT was always lower in treated tissues, throughout the experiments; soluble protein contents were always higher in treated tissues. Uptake of N0~ was generally more intense in treated material, discounting an initial two days period during which the inverse was observed. Data obtained show that penicillins enhance certain metabolic pathways of cul- tured plant cells.
Key words: nitrogen metabolism enzymes; callus cultures; penicillins
Introduction
Cultures of plant cells or callus tissue are presently valuable tools in a wide variety of research fields, from applied biotechnology to many areas of basic physiology and metabolic behaviour. When working with delicate plant material that do not endure the currently used harsh disinfection procedures, contamination with microorganisms is likely to occur.
Bacterial contamination is frequently a hazardous situation in cultures of woody plants, mainly in large scale micropropagation labora- tories [1], since repeated treatments with chem- ical disinfectants very often produce damage of the plant tissues. Also infection of valuable cul- tures are sometimes very damaging to the work under way. To prevent, or control, these undesirable situations antibiotics have been used [1--3], and also suggested as disinfection media, substituting for aggressive chemicals currently used.
However, culture media with added antibiot- ics were mostly used for short-term cultures, generally for 5 days or even shorter periods [1]. The behaviour of cultures for longer periods have been less frequently studied [3] and work previously carried out in our laboratory showed that cultured plant cells were not indif- ferent to such type of treatments and it was even found that some antibiotic treated callus showed stimulation of metabolic pathways as well as growth ratios [4--6]. Beneficial effects of penieillins were reported for higher plant cells [7,8] and also for short-term plant tissue cultures [1,2].
The observed stimulatory effects of some penicillins prompted further studies to better know the response of cultured callus tissue. On the other hand it seemed of interest to have some account of how basic metabolic mecha- nism were affected, notably those concerned to nitrogen assimilation in which the inorganic nitrogen is incorporated into different biologi-
0168-9452/89/$03.50 © 1989 Elsevier Scientific Publishers Ireland Ltd. Printed and Published in Ireland
120
cal nitrogenous compounds. The effects of peni- cillins on the uptake and metabolism of nitrogen in tissue cultures were appraised through the activity levels of enzymes related to those processes, results which are reported in the present paper.
Material and methods
Established callus cultures derived from leaf mesophyl] of Sedum telephium L. were used as plant material for this study. Innocula (approx. 5 g) were t ransferred to Gamborg's B-5 medium [9] containing 10 mg/l benzylamino purine plus 0.1 mg/l naphthalene acetic acid, solidified with 0.6% agar, containing penicillin G (PG) or peni- cillin V (PV) at 100 ~g/ml, or without them as control.
Callus tissue was grown at 27°C in a 12-h light period (10 W m -2) for 14 days. Samples were collected after 2, 4, 6, 8, 10, 12 and 14 days of growth and for each material studied 4 repli- cates were prepared and used for enzyme activ- ity evaluation. This schedule of experimen- tation was run thrice.
Enzyme extraction Tissue was homogenized in 100 mM phos-
phate buffer (pH 7.5) containing I mM disodium ethylenediaminetetra-acetate, 1% of polyvinyl- polypyrrolidone, 1 mM dithiothreitol and 1o/0 bovine serum albumin, and centrifuged at 16 000 × g for 10 min. The supernatant was col- lected and used as enzyme source for measure- ment of nitrate reductase (NR) activity. For glutamate synthase (GOGAT), glutamine syn- thetase (GS) and glutamate dehydrogenase (GDH) Tris--HCl buffer was used instead of phosphate buffer and cysteine (5 raM) added. All operations were performed at 4 °C.
Enzyme assay Nitrate reductase (NR, EC 1.6.6.1) activity
was assayed using the procedure described by Hageman and Reed [10] with some alterations. The assay mixture at pH 7.5 contained, potas- sium phosphate buffer 50 pmol; KNO 8 20/~mol; NADH 0.5 pmo] and enzyme extract 0.2 ml, in a
final volume of 2.0 ml. After 15 min of incuba- tion at 30°C the reaction was terminated by adding 1 ml of sulfanilamide (1% in 1.0 M HC1) plus 1 ml of the N-(1-naphtyl) ethylenediamine (0.020/0). The color was allowed to develop for 20 rain, the contents of the incubation tube were centrifuged at 16 000 × g for 5 min and the supernatant used for reading at 540 nm against a blank containing all the above referred ingre- dients except NADH.
Glutamine synthetase (GS, EC 6.3.1.2) was assayed through Boyer et al. [11] method; gluta- mate synthase (GOGAT, EC 1.4.7.1) activity was measured by the method of Sodek and Da Silva [12]. Glutamate dehydrogenase (GDH, EC 1.4.1.3) activity was assayed in accordance to the method of Pahlich and Joy [13]. Soluble pro- teins were determined by the Lowry method [14]. The fate of nitrate in the culture media was followed by polarography using a selective ion electrode.
Results
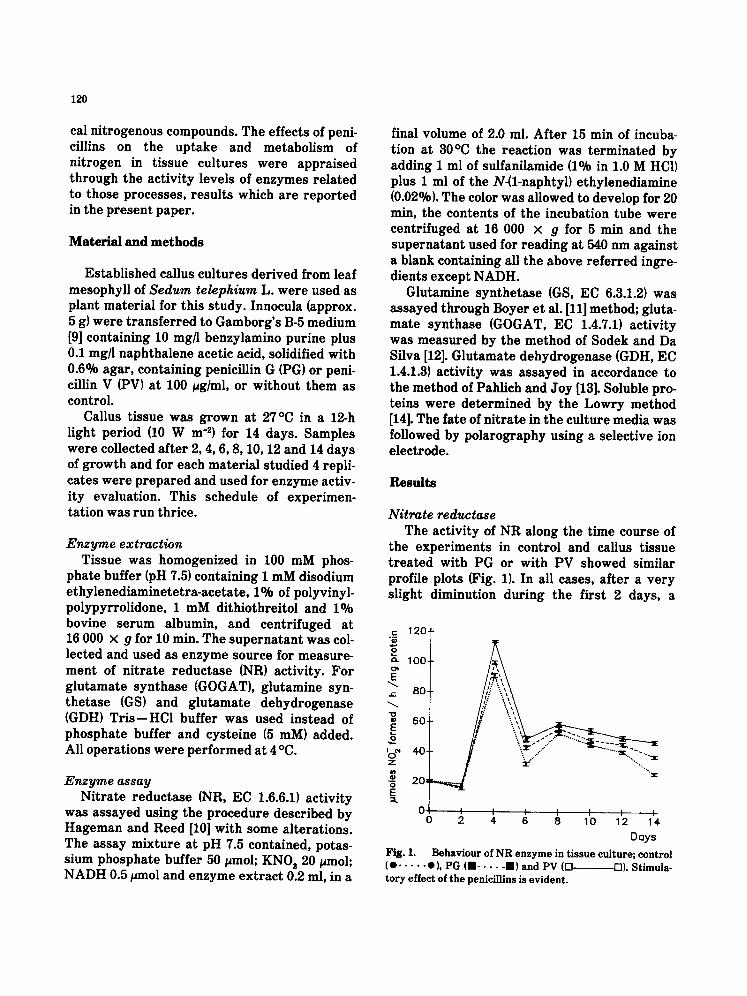
Nitrate reductase The activity of NR along the time course of
the experiments in control and callus tissue t reated with PG or with PV showed similar profile plots (Fig. 1). In all cases, after a very slight diminution during the first 2 days, a
c 120-
¢z 100- o t
E .~ 80-
60-
'o ~ 40- Z
o E
0.~, 0
_ . . . . . . . . . . . . . . . ~...'..-- -.
I I I I I I 2 4 6 8 10 12 14.
Days
Fig. 1. Behav iour of N R e n z y m e in t i s sue cul ture ; control (O . . . . . O), PG (urn . . . . . ran) and PV (D D). St imula- to ry effect of t he peniciLlins is ev ident .
rapid increase in activity was observed up to day 4, followed by a sharp decline measured two days later; at day 8 a high activity was noted, followed by a slow and steady lower- ing of activity till the end of the experiment. However differences among the three situa- tions were noted since in comparison to control significantly higher levels of enzyme activity were measured throughout the growth period in antibiotic t reated callus and more so in the presence of PV. This stimulation of enzyme activity during almost all the experiment shows out clearly by the end of the growth period and is easily perceived in (Fig. 1), where the activity of NR at day 14 was 44.9% higher in PG-treated material and 83% higher in PV- t reated tissue than in control.
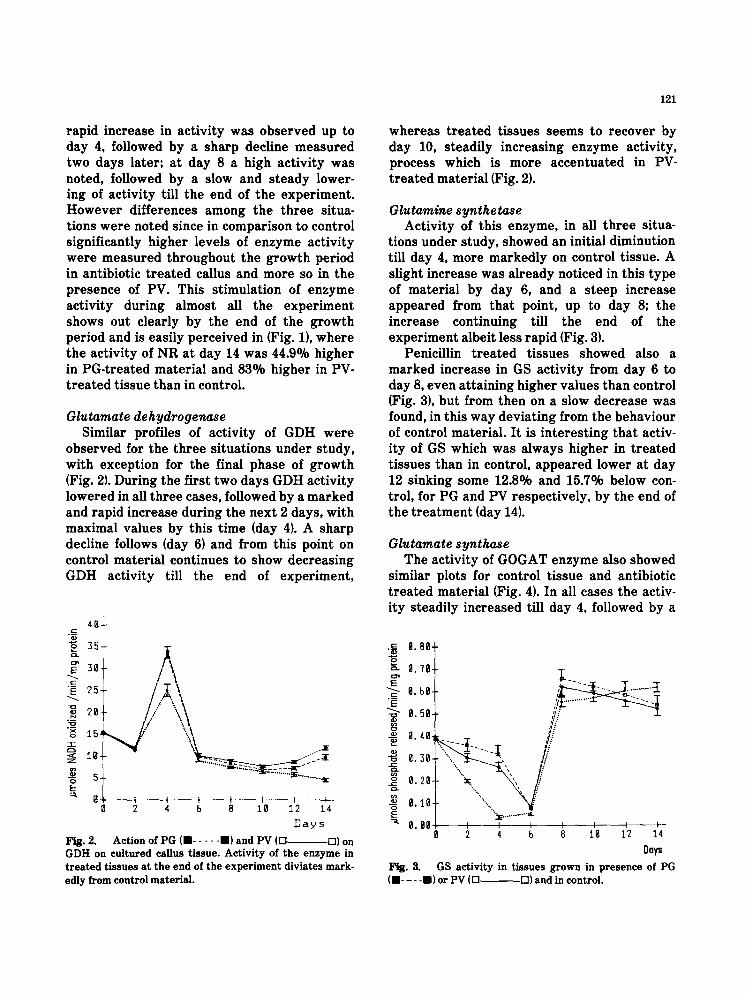
Glutamate dehydrogenase Similar profiles of activity of GDH were
observed for the three situations under study, with exception for the final phase of growth (Fig. 2). During the first two days GDH activity lowered in all three cases, followed by a marked and rapid increase during the next 2 days, with maximal values by this time (day 4). A sharp decline follows (day 6) and from this point on control material continues to show decreasing GDH activity till the end of experiment,
4° i
I 30
• ~ 2~-
26-
< I g -
o E
0 0
i I i I I I 2 4 b 8 10 12 14
Days
Fig. 2. Action of PG (" . . . . . ") and PV (D D) on GDH on cultured callus tissue. Activity of the enzyme in treated tissues at the end of the experiment diviates mark- edly from control material.
121
whereas t rea ted tissues seems to recover by day 10, steadily increasing enzyme activity, process which is more accentuated in PV- t reated material (Fig. 2).
Glutamine synthetase Activity of this enzyme, in all three situa-
tions under study, showed an initial diminution till day 4, more markedly on control tissue. A slight increase was already noticed in this type of material by day 6, and a steep increase appeared from that point, up to day 8; the increase continuing till the end of the experiment albeit less rapid (Fig. 3).
Penicillin t rea ted tissues showed also a marked increase in GS activity from day 6 to day 8, even attaining higher values than control (Fig. 3), but from then on a slow decrease was found, in this way deviating from the behaviour of control material. It is interesting that activ- ity of GS which was always higher in t reated tissues than in control, appeared lower at day 12 sinking some 12.8O/o and 15.7% below con- trol, for PG and PV respectively, by the end of the treatment (day 14).
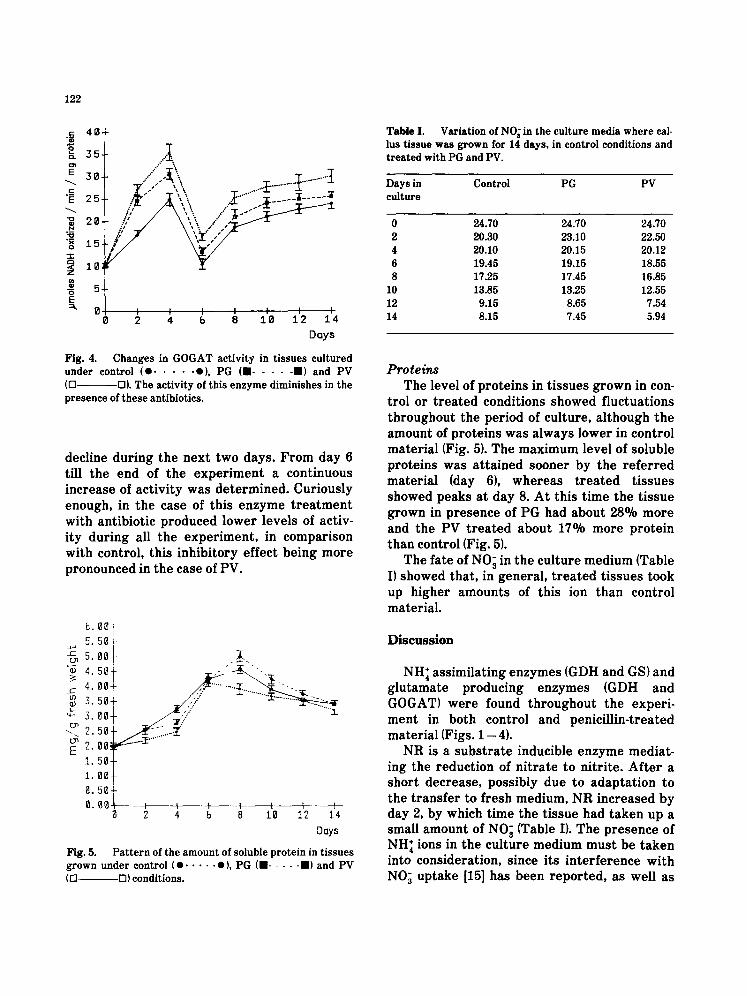
Glutamate synthase The activity of GOGAT enzyme also showed
similar plots for control t issue and antibiotic t reated material (Fig. 4). In all cases the activ- ity steadily increased till day 4, followed by a
Fig, 3, (m .... i) or PV (D
9.86~-
8.48#
8.39-
g,2g-
g . l g -
0.00 I I L I i i I 2 4 b B Ig 12 14
Days GS activity in tissues grown in presence of PG
D) and in control.
122
= 40
~. 3 5
~ 30.
"~ 25.
~ 20
"~ 1 5 - r -
10:
o E ~ 0
0
Fig. 4.
Y W I I 1 I I I I 2 4 b 8 1 0 12 14
Days
Changes in GOGAT activity in tissues cultured under control (e . . . . . o), PG (11 . . . . . II) and PV (Q D). The activity of this enzyme diminishes in the presence of these antibiotics.
decline dur ing the nex t two days . F r o m day 6 till the end of the e x p e r i m e n t a cont inuous increase of ac t iv i ty was de t e rmined . Cur iously enough, in the case of th is e n z y m e t r e a t m e n t wi th ant ibiot ic p roduced lower levels of activ- i ty dur ing all the e x p e r i m e n t , in compar i son with control , this inh ib i to ry ef fec t be ing m o r e p ronounced in the case of PV.
[3,
5. 5. ED
"~. 4. 4. 3.
E 2. 1. 1.
0 0 + I
504-
.:.!. i. 50+
/ / "'"1 g 0- _:;If'/" 50-~;;/ Og.~ " 50- 00- 50-
2 4 b 8 10 12 14 Days
Fig. 5. Pattern of the amount of soluble protein in tissues grown under control (0 . . . . . 0), PG (11 . . . . . II) and PV (12] D) conditions.
Table I. Variation of NO~ in the culture media where cal- lus tissue was grown for 14 days, in control conditions and treated with PG and PV.
Days in Control PG PV culture
0 24.70 24.70 24.70 2 20.30 23.10 22.50 4 20.10 20.15 20.12 6 19.45 19.15 18.55 8 17.25 17.45 16.85
10 13.85 13.25 12.55 12 9.15 8.65 7.54 14 8.15 7.45 5.94
Proteins The level of p ro te ins in t i s sues g rown in con-
t rol or t r e a t e d condit ions showed f luc tua t ions th roughou t the per iod of cul ture , a l though the amount of p ro te ins was a lways lower in control ma te r i a l (Fig. 5). The m a x i m u m level of soluble pro te ins was a t t a ined sooner by the r e f e r r e d mate r i a l (day 6), whe rea s t r e a t e d t i s sues showed peaks a t day 8. A t this t ime the t i s sue grown in p r e sence of PG had abou t 28% more and the PV t r e a t e d abou t 17O/o more p ro te in than control (Fig. 5).
The fa te of NO 3 in the cu l ture m e d i u m (Table I) showed tha t , in genera l , t r e a t e d t i s sues took up higher amoun t s of this ion than control mater ia l .
D i s c u s s i o n
N H 4 ass imi la t ing enzymes (GDH and GS) and g l u t a m a t e p roduc ing enzymes (GDH and GOGAT) w e r e found t h r o u g h o u t the exper i - m e n t in both control and penic i l l in- t reated mate r ia l (Figs. 1 - 4).
NR is a s u b s t r a t e inducible e n z y m e media t - ing the reduct ion of n i t r a t e to ni t r i te . A f t e r a shor t decrease , poss ib ly due to adap ta t ion to the t r a n s f e r to f resh medium, N R increased by day 2, by which t ime the t i s sue had t a k e n up a small amoun t of NO 3 (Table I). The p r e sence of N H 4 ions in the cu l ture med ium m u s t be t a k e n into considerat ion, since i ts i n t e r f e r ence wi th NO 3 up take [15] has been r e p o r t e d , as well as

lowering extractable NR [16], preventing inducible NR activity in roots [15,16], in culture cells [17] and in Chlorella [18], not to mention the 5 h delay in substrate induction of mem- brane transporters for the ion itself and some 6
- 10 h for NO~ transporters [19]. NR activity declined during the two initial
days but afterwards increased sharply, reach- ing maximum value by day 4 (Fig. 1), possibly due to progressive uptake of N0~ from the medium {Table I) and consequent stimulation of NR synthesis and also very likely due to stimu- latory effect of BA present in the medium, as reported in other cases [20,21].
Since the inoculum of tissue showed the highest growth ratio from day 4 to 6 {data not shown) the cellular concentration of NO~ correspondingly decreases, which might explain the decrease of NR activity by day 6, in agreement to references that both induction and steady-state level of NR activity are affected by nitrate concentration [22,23]. The decrease might also be due to NR-inactivating factors since they were detected in other cell cultures [24]. The slow and progressive decline of NR activity during the last 6 days of the experiment is similar to the diminution of activ- ity found in other cases [25].
The presence of PG or PV promoted higher levels of NR activity than in control, with the latter more effective, producing 83o/0 more activity than in control, in comparison with some 44o/0 for the former.
Various metabolic pathways in plants [7,8,26] and in tissue cultures [4--6] are known to be affected by penicillins. Mukerji's group reported that the antibiotic led to increased levels of cytokinins and that it could emulate cytokinin itself by increasing chloroplast pig- ment contents [26,27]. This fits well in our study because cytokinins are capable of inducing NR synthesis [20,21]; it seems reasonable to admit that in our cultures penicillins act like cytoki- nins {promoting NR synthesis) and would also raise the levels of the phytohormone, thus fur- ther promoting synthesis of NR.
GDH activity, in control and treated condi- tions, increased markedly from day 2 to day 4
123
(Fig. 2), interestingly in accordance with NR activity. This latter enzyme would produce a high intraceUular concentration of ammonium, making available one of the reagents for the reaction mediated by GDH. The high values found for the activity of GDH suggest that dur- ing the initial phase of culture NH 4 assimilation proceeded mostly through intervention of this enzyme, similarly as described for Bouvardia ternifolia cultures [28]; high enzyme activity has been reported as a consequence of high ammonium concentration [29,30]. GDH activity declined sharply from day 4 to day 6 very likely due to lowering of intracellular NH~. From this point on until the end of the experiment control tissue progressively looses enzyme activity, suggesting that this route of NH~ assimilation progressively fades away in favour of the other known pathway, an assumption supported by steady increase of GS and GOGAT activities (Figs. 3 and 4). Treated tissues showed similar behaviour up to day 10, at which time GDH activity started rising in them. Here again, this behaviour seems linked to GS levels, since peni- cillin treatment led to a lowering of activity of this enzyme in the last days of the experiment (Fig. 3).
GS activity in control material declined dur- ing the first four days (Fig. 3), behaviour that could be the result of the cell dedifferentiation, which is known to occur when this callus tissue is transferred to fresh medium [31], with asso- ciated revertion of chloroplasts to proplastids and diminished supply of ATP.
As mentioned above the activity of GDH reached very low levels by day 6 and never again increased significantly, suggesting that from this point on assimilation of ammonium was taken over by the enzyme tandem system of GS-GOGAT. In support of this reasoning those two enzymes began to increase also at day 6, in both control and treated tissue (Figs. 3 and 4).
The data obtained seem to indicate that dur- ing the initial 6 days of control culture, gluta- mine was not produced through GS mediation, since the activity of this enzyme falls off rap- idly, approaching zero. Most certainly GOGAT

124
was using glutamine already present in the cells, as this molecule is known to be one of the major nitrogen storage compounds [32]. This pool of glutamine possibly approached deplec- tion by day 4, when GOGAT activity started decreasing sharply. This enzyme markedly gained activity from day 6 on, in accordance with GS, this latter enzyme certainly producing large amounts of glutamine, the substrate for GOGAT.
During the initial days penicillin treated tis- sue had much higher levels of GS activity than control material, indicating that some gluta- mine was being produced. However, the same tissues showed less GOGAT activity, a differ- ence which might be explained by the fact that the treated tissues had also higher amounts of proteins (Fig. 5).
PeniciUins can enhance some aspects of metabolism of cells from higher plants [6], and the activity of enzymes involved in nitrogen metabolism were found to be no exception. Indeed, PG and PV stimulated activities of NR, GDH, and, except at the end of the experiment, also GS; the lowering of GOGAT activity should be viewed in conjunction with the levels of protein synthesis, compounds which appeared at significantly higher levels than in untreated tissues, a situation which might be responsible for the divergence of some gluta- mine, since this metabolite is essential for nucleic acid and protein synthesis [33].
Acknowledgements
Grants from INIC (Lisbon, Portugal) and from Stiftung Volkswagenwerk (Hannover, F.R.G.) are gratefully acknowledged.
References
P.M. Young, A.S. Hutchins and M.L. Canfield, Use of antibiotic to control bacteria in shoot cultures of woody plants. Plant Sci. Lett., 34 (1984) 203--209. S.M. Phillips, S.M. Arnott and S.E. Kaplan, Antibiotics in plant tissue culture: rifampicin effectively controls bacterial contaminants without affecting the growth of short-term explant cultures of Helianthus tuberosus. Plant Sci. Lett., 21 (1981) 235--240.
3 K. Pollock, D.G. Barfield and R. Shields, The toxicity of antibiotics to plant cell culture. Plant Cell Pep., 2 (1983) 36-- 39.
4 R. Salema, Ultrastructural and biochemical aspects of callus tissue grown in antibiotic containing media. Electron Microsc., 3 (1984) 2099--2100.
5 R. Salema and I. Santos, Some effects of penicillin on callus cultures of Sedum telephium L. Cien. Biol., 9 (1984) 257.
6 I. Santos and R. Salema, Effects of penicillins on plant tissue cultures: ultrastructural and biochemical aspects. Bol. Soc. Brot., 58 (1985) 269- 285.
7 S. Mukherji and A.K. Biswas, Penicillin action on chiw roplast pigment composition. Hill reaction and chloro- phyllase activity in Euphorbia pulcherrima L. leaf discs. Ind. J. Exp. Biol., 19 (1981) 70--72.
8 S. Mukherji and A.K. Biswas, Penicillin action stimu- lating growth in seedling of rice (Oeyza sutiva). Can. J. Bot., 63 (1984) 1150-1156.
9 O.L. Gamborg, R.A. Miller and K. Ojima, Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res., 50 (1968) 151-158.
10 R.H. Hageman and A.J. Reed, Nitrate reductaso from higher plants. Methods Enzymol., 69 (1980) 270--280.
11 P.D. Boyer, R.C. Mills and H.J. Fromm, Hypothesis for and some kinetic studies with glutamine synthetase and acetate thiokinase. Arch. Biochem. Biophys., 81 (1959) 249 - 263.
12 K. Sodek and W.J. Da Silva, Glutamate synthase a possible role in nitrogen metabolism of the developing maize endosperm. Plant Physiol., 60 (1977) 602-- 605.
13 E. Pahllch and K.W. Joy, Glutamate dehydrogenase from pea roots: purification and properties of the enzyme. Can. J. Biochem., 49 (1971) 127-138.
14 O.H. Lowry, N. Pesenbrough, A.L. Farr and R.J. Randall, Protein measurement with folinphenol reagent. J. Biol. Chem., 193 (1951) 265- 275.
15 G.J.T. Frith, Effect of ammonium nutrition on the activity of nitrate reductase in the roots of apple seed- lings. Plant Cell Physiol., 13 (1972) 1085-- 1090.
16 T.O. Orebamjo and G.R. Stewart, Ammonium repres- sion of nitrate reductase induction in Lemna minor L. Planta, 122 (1975) 27 - 36.
17 J.M. Bayley, J. King and O.L. Gamborg, The effect of the source of inorganic nitrogen on growth and enzymes of nitrogen assimilation in soybean and wheat cells in suspension cultures. Planta, 105 (1972) 15--24.
18 M. Losada, A. Paneque, P.J. Aparicio, J.M. Vega and J. Cardenas, Inactivation and repression by ammonium of the nitrate reducing system in Chlorella. Biochem. Biophys. Res. Commun., 38 (1970) 1009-1015.
19 S.S. Goyal and C. Huffaker, The uptake of NO~, NO~ and NH~ by intact wheat (Triticum aestivum) seed- lings. Plant Physiol., 82 (1986) 1051 -- 1056.
20 U.N. Dwivedi, B.M. Kahn, S.K. Rawal and A.F. Mascarenhas, Biochemical aspects of shoot differentia- tion in sugarcane callus: 1. Nitrogen assimilating enzymes. J. Plant Physiol., 117 (1984) 7-- 15.

21 H. Kende, H. Hahn and S.E. Kays, Enhancement of nitrate reduetase activity by benzyladenine in Agros- temmum githago. Plant Physiol., 48 (1971) 702--706.
22 J.S. Knypl, Synergistic induction of nitrate reductase activity by nitrate and bonzyaminopurine in detached cucumber cotyledons. Z. Pflanzenphysiol., 70 (1973) 1--11.
28 W. Chantarotwong, R.C. Huffaker, B.L. Miller and R.C. Granstedt, In vivo nitrate reduction in relation to nitrate uptake, nitrate content, and in vitro nitrate reductase activity in intact barley seedlings. Plant Physiol., 57 (1976) 519-- 522.
24 D.L. Shaner and J.S. Boyer, Nitrate reductase activity in maize (Zea mays L.) leaves, II. Regulation by nitrate flux. Plant Physiol., 58 (1976) 499-- 504.
25 T. Yamaya, A. Oaks and I.L. Boesel, Characteristics of nitrate reductase-inactivating proteins obtained from corn roots and rice cell cultures. Plant Physiol., 65 (1980) 141 -- 145.
26 S. Mukherji and A.K. Biswas, Modulation of chloro- phyll carotene and xantophyll formation by penicillin, benzyladenine and embryonic axis in mungbean (Phaseolua aureus L.) cotyledons. Ann. Bot., 43 (1979) 225 - 229.
27 S. Mukherji and P.F. Wareing, The effect of penicillin
125
on the levels of endogenous gibberellins and cytokinins in mungbean (Vigna radiata L.) seedlings. Biol. Plant., 25 (1983) 161 - 165.
28 E. Murillo and E. Sanchez de Jimenez, Alternative pathways for ammonium assimilation in Bouvardia ternifolia cell suspension cultures. J. Plant Physiol., 117 (1984) 57--68.
29 I. Barash, T. Sadon and H. Mor, Evidence of ammon- ium-dependent de novo synthesis of glutamate dehy- drogenase in detached oat leaves. Plant Physiol., 56 (1975) 856-- 858.
30 V.M. Loyola-Vargas and E. Sanchez de Jimenez, Dif- ferential role of glutamate dehydrogenase in nitrogen metabolism of maize tissues. Plant Physiol., 76 (1984) 536--540.
31 I. Brandbo and R. Salema, Callus and plantlets devel- opment from cultured leaf explants of Sedum tele- phium L. Z. Pflanzenphysiol., 85 (1977) 1 --8.
32 R.R.S. Bidwell, Nitrogen Metabolism in Plant Physiol- ogy, Collier MacMillan Int. Editions, London, 1979 pp. 192-- 227.
33 V.M. Loyola-Vargas and E. Sanchez de Jimenez, Effect of nitrate, ammonium and glutamine on nitrogen assimilation enzymes during callus growth of maize. J. Plant Physiol., 125 (1986) 235-- 242.