peroxisome proliferators induce mouseliver stearoyl-coa … · scd1 mrna levels. additionally, the...
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 93, pp. 9443-9448, September 1996Biochemistry
Peroxisome proliferators induce mouse liver stearoyl-CoAdesaturase 1 gene expression
(clofibrate/gemfibrozil/peroxisome proliferator-activated receptor/polyunsaturated fatty acid)
CAROLYN WILSON MILLER* AND JAMES M. NTAMBI*tDepartments of *Biochemistry and tNutritional Sciences, University of Wisconsin, Madison, WI 53706
Communicated by M. Daniel Lane, Johns Hopkins University School of Medicine, Baltimore, MD, June 11, 1996 (received for reviewApril 29, 1996)
ABSTRACT Peroxisome proliferators induce stearoyl-CoA desaturase activity (EC 1.14.99.5) in liver [Kawashima,Y., Hanioka, N., Matsumura, M. & Kozuka, H. (1983) Biochim.Biophys. Acta 752, 259-264]. We analyzed the changes instearoyl-CoA desaturase 1 (SCD1) mRNA to further define themolecular mechanism for the induction of stearoyl-CoA de-saturase by peroxisome proliferators. SCD1 mRNA was ana-lyzed from the livers of BALB/c mice that had been fed dietssupplemented with clofibrate or gemfibrozil. Clofibrate wasfound to induce liver SCD1 mRNA levels 3-fold within 6 hr toa maximum of 22-fold in 30 hr. Gemfibrozil administrationresulted in a similar induction pattern. This induction isprimarily due to an increase in transcription of the SCD1gene, as shown by nuclear run-on transcription assays andDNA deletion analysis of transfected SCD1-chloramphenicolacetyltransferase fusion genes. The cis-linked response ele-ment for peroxisome proliferator-activated receptor (PPAR)was localized to an AGGTCA consensus sequence betweenbase pairs -664 to -642 of the SCD1 promoter. Clofibrate-mediated induction of SCD1 mRNA was shown to be inde-pendent of polyunsaturated fatty acids, with peroxisome pro-liferators and arachidonic acid having opposite effects onSCD1 mRNA levels. Additionally, the activation of SCD1mRNA by clofibrate was inhibited 77% by cycloheximideadministration. Levels of liver 13-actin and albumin mRNAswere unchanged by these dietary manipulations. Our datashow that hepatic SCD1 gene expression is regulated byPPARs and suggest that peroxisome proliferators and poly-unsaturated fatty acids act through distinct mechanisms.
Peroxisome proliferators include structurally distinct com-pounds such as amphipathic carboxylates, herbicides, andpesticides. These compounds cause hypolipidemia when ad-ministered to mice, rats, humans, and other mammals (1), and,therefore, have the potential to be used in the treatment ofhyperlipidemia and hypercholesterolemia. Transcription ofperoxisomal (3-oxidation genes and liver cytochrome P450genes is induced up to 20-fold by peroxisome proliferators(2-4), while the transcription of lipogenic genes such as S14and fatty acid synthase is repressed (5). Also, the transcriptionof genes involved in phospholipid biosynthesis is induced,probably as a result of the increase in the size of peroxisomal,mitochondrial, and plasma membranes (6). These transcrip-tional effects are presumed to be mediated by peroxisomeproliferator-activated receptors (PPARs) (7). Three PPARgenes (a, ,B, and ry) have been cloned and sequenced indifferent species: 3 Xenopus (8), 2 mouse (9-11), 1 rat (12),and 1 human (13). PPARs are members of the steroid hor-mone superfamily and have been shown to bind peroxisomeproliferator-activated receptor response elements (PPREs) inassociation with retinoid-X receptors (RXR). PPARs are
activated by polyunsaturated fatty acids (PUFAs), as well asperoxisome proliferators. Peroxisome proliferators have beenshown to repress the activity of certain lipogenic proteins suchas S14 and fatty acid synthase through PPARs (5). In contrast,hepatic stearoyl-CoA desaturase levels are induced in rat liver(14) by clofibrate, a known peroxisome proliferator.
Stearoyl-CoA desaturase (SCD; EC 1.14.99.5) is a keyenzyme in fatty acid biosynthesis. It catalyzes the A9-cisdesaturation of fatty acyl-CoA substrates, the preferred sub-strates being palmitoyl-CoA and stearoyl-CoA, which areconverted to palmitoleoyl- and oleoyl-CoA respectively. Oleicand palmitoleic acids are the major unsaturated fatty acids infat depots and membrane phospholipids. The peroxisomeproliferator-mediated increase in SCD1 activity correspondsto an increase in the levels of oleic acid incorporated intophospholipids relative to stearic acid (15). The ratio of stearicto oleic acid is one of the factors influencing cell membranefluidity and cell-cell interactions (16). Abnormal alteration ofthis ratio has been shown to play a role in various disease statessuch as aging, diabetes, neurological and vascular disease, andcarcinogenesis (16).The present study examined the effects of clofibrate and
gemfibrozil on SCD gene expression in mouse liver. Thepurpose of this study was to determine the mechanism throughwhich peroxisome proliferators regulate SCD1 gene expres-sion. We show that peroxisome proliferators induce SCD1mRNA and that this increase is due primarily to an increase intranscription of the SCD1 gene. A PPRE has been localized inthe SCD1 promoter that confers peroxisome proliferatorresponsiveness. Similar to other PPREs, this sequence has asubstantial identity to the known consensus sequence. Clofi-brate induction of SCD1 mRNA is inhibited by cycloheximideadministration, indicating that ongoing protein synthesis isnecessary for the induction. Clofibrate and arachidonic acidhave opposing effects on SCD1 mRNA levels. This suggeststhat regulation of the SCD1 gene by peroxisome proliferatorsand PUFAs occur through distinct mechanisms.
EXPERIMENTAL PROCEDURESMaterials. Restriction enzymes and other nucleic acid-
modifying enzymes were from Promega. The fat-free high-carbohydrate diet was obtained from United States Biochem-ical. Triarachidonin, trilinolein, clofibrate, gemfibrozil, andcycloheximide were purchased from Sigma. Lipofectamineand cell culture media were obtained from GIBCO/BRL. ThecDNA probe for human ,3-actin was obtained from D. Cleve-land (Johns Hopkins University). The f-actin used for theRNase protection was from Ambion. The albumin cDNAprobe used was obtained from K. Zaret (Brown University).
Abbreviations: SCD, stearoyl-CoA desaturase; PPAR, peroxisomeproliferator-activated receptor; PPRE, peroxisome proliferator-activated receptor response element; PUFA, polyunsaturated fattyacid; CAT, chloramphenicol acetyltransferase.
9443
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020

9444 Biochemistry: Miller and Ntambi
All plasmid DNAs were isolated by a modified SDS/NaOHmethod or the alkaline lysis method (17). The cDNA for themPPARa expression vector was obtained from J. Tugwood(Zeneca, Macclesfield, United Kingdom). The plasmidspCAT(An) and TKCAT(An) were obtained from HowardTowle (University of Minnesota). Oligonucleotides were ob-tained from Eppendorf.Animals and Treatments. Male BALB/c mice were ob-
tained from the Department of Biochemistry of the Universityof Wisconsin and from Harlan Sprague Dawley. Mice (4-6weeks old) were maintained on a complete diet of PurinaFormulab Diet 5008 for at least 2 days prior to the start of theexperiment. Mice were starved for 24 hr, refed Purina chowdiet for 24 hr, starved for 24 hr (18), and then fed the test dietat intervals shown in the figures and legends. This feedingschedule ensured that the mice ingested the food. In desig-nated experiments the chow diet was supplemented withclofibrate or gemfibrozil, and the fat-free diet was supple-mented with triarachidonin, butylated hydroxytoluene (0.1%)as an antioxidant, and clofibrate. Animals were given free accessto tap water at all times. In each experiment two mice weretreated per sample, and they were processed separately to con-firm results by duplication. Each experiment was repeated at leastthree times. Ten-week-old mice were used to confirm the effectin older animals. Cycloheximide (1 mg/100 g of body weight) wasintraperitoneally administered 12 hr prior to sacrifice.
Isolation and Analysis of Liver RNA. Total hepatic RNAwas isolated according to Chirgwin et al. (19). RNase protec-tion analysis was performed according to Melton et al. (20),using antisense RNA probes for SCD1, albumin, and actinsynthesized and hybridized to 15 ,ug of RNA as previouslydescribed (21). The protected RNA hybrids were precipitatedwith ethanol, analyzed by electrophoresis on a 7% polyacryl-amide/7 M urea sequencing gel, and visualized by autoradiog-raphy. Relative levels of hybridization were quantified by laserdensitometry using multiple exposures to confirm the results. Theintegrity of the RNA was verified by fractionating 20 ,ug of totalliver RNA on a 1% agarose gel containing 6.7% formaldehydeand visualizing the RNA by ethidium bromide staining.
Isolation of Nuclei and in Vitro Run-on Transcription.Nuclei were isolated from fresh liver tissue of mice fed a chowdiet with or without 1% clofibrate for various time periods andused for gene transcription studies as previously described (21).
Construction, Transfection, and Assay of Expression ofSCD1-Chloramphenicol Acetyltransferase (CAT) ChimericGenes. The putative regulatory regions containing 4300, 1200,600, and 363 bp of the 5' flanking sequence were subclonedinto the BamHI sites of the promoterless plasmid pCAT(An)(22). The deletion constructs were generated by using appro-priate restriction enzymes followed by Bal 31. After bluntingof the ends with the Klenow fragment of DNA polymerase 1,BamHI linkers were added for insertion into the pCAT(An)plasmid. Oligonucleotides (corresponding to the -664/-642region of the SCD1 promoter) with BamHI ends were syn-thesized (5'-GATCCTAGATGTGAAGTTAGACCGAG-3'). They contained the PPRE of the SCD1 gene as shown inFig. 4. After annealing of the single-stranded oligonucleotides,the double-stranded PPRE was subcloned into the BamHI siteof TKCAT(An). HepG2 cells (106) were seeded onto 6-cmculture dishes in high-glucose (0.45%) Dulbecco's modifiedEagle's medium (DMEM) supplemented with 10% fetal bo-vine serum 1 day prior to transfection. Transfections werecarried out using Lipofectamine with 10 ,ug of reporter plasmidand 1 ,ug of mPPARa expression plasmid. RSVCAT andpCAT(An) were transfected as controls (23). After overnighttransfection, the medium was replaced with high-glucoseWilliams' medium E (GIBCO/BRL) containing 1 ,uM insulinand 1 nM dexamethasone with or without 150 ,uM clofibratefor 36 hr. Williams' medium E was used to maximize trans-fection efficiency and to maintain fatty acid-free conditions.
CAT activity was determined as described by Neumann et al.(24). Results were normalized to protein content.
RESULTSInduction of SCD1 mRNA by Peroxisome Proliferators.
Hepatic SCD activity was shown to increase with treatment ofclofibrate (14). To determine if the induction was due to anincrease in mRNA, we examined the effects of 0.5% and 1%clofibrate administered in a normal chow diet for 36 hr on theaccumulation of SCD1 mRNA in mouse liver. SCD1 andf3-actin mRNA levels were analyzed by RNase protection (21,25). The protected fragments were separated on a denaturingpolyacrylamide gel and the autoradiograms are shown. Clofi-brate (0.5% and 1%) significantly induced SCD1 mRNA (Fig.1A), without changing levels of f3-actin mRNA (Fig. 1B),showing that the clofibrate-mediated increase in SCD1 mRNAis specflic.To show that other peroxisome proliferation-inducing drugs
could induce SCD1 mRNA, we examined the effects of 1% and1.5% gemfibrozil, administered in a chow diet for 36 hr. SCD1mRNA levels were analyzed by RNase protection. The resultsshow that mouse liver SCD1 mRNA is induced in a dose-dependent manner by gemfibrozil (Fig. 1C). This furthersupports the observation that SCD1 mRNA levels are inducedby peroxisome proliferators.SCD1 mRNA Induction by Clofibrate Is Time Dependent
and Requires Protein Synthesis. To examine the inductiontime course of SCD1 mRNA by clofibrate, mice were starvedand refed a chow diet supplemented with 1% clofibrate for 0,6, 18, 24, 30, and 36 hr. SCD1 and albumin mRNA levels wereanalyzed by RNase protection assay, and the SCD1 autora-
A
B
clofibrateT + 0\20\1I
a 41 Z
SCDI_probe w :~~~~~~~~~~~~~~~.... ..
mRNA
actinprobe
actnmRNA
c.0 gemfibrozil
IT\oo\\1 0?2 ---..-le % oh.'ON
SCD1probe
-
0SCD1mRNA
FIG. 1. Effects of peroxisome proliferators on SCD1 mouse livermRNA. Mice were fasted and then fed Purina lab chow, fat-freehigh-carbohydrate diet (CHO), or Purina lab chow supplemented withclofibrate or gemfibrozil for 36 hr. Total RNA was isolated andanalyzed by RNase protection assay using an SCD1-specific comple-mentary riboprobe in response to clofibrate (A) or clofibrate andgemfibrozil (C). (B) A f-actin-specific complementary probe was usedas a control to show that f3-actin mRNA levels do not changesignificantly in response to clofibrate treatment. Essentially identicalresults were obtained from three separate experiments.
Proc. Natl. Acad. Sci. USA 93 (1996)
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020

Proc. Natl. Acad. Sci. USA 93 (1996) 9445
diogram is shown (Fig. 2A). The hybridization signals werequantitated by laser densitometry (Fig. 2B). Clofibrate in-duced SCD1 mRNA levels 3-fold within 6 hr and accumulatedto a maximum of 22-fold within 30 hr. High-carbohydratefat-free diet was administered as a control, to reaffirm induc-tion of SCD1 mRNA under these conditions (21). The expres-sion of SCD1 mRNA was induced 20-fold in response to ahigh-carbohydrate diet. The level of liver albumin mRNA didnot change significantly during the time course of clofibratefeeding, showing that the clofibrate-mediated increase inSCD1 mRNA is specific.The 24-hr induction lag of SCD1 mRNA by peroxisome
proliferators indicates a requirement for protein synthesis. Totest this possibility, and to gain further insight into the mo-lecular mechanism of the clofibrate-mediated SCD1 inductionin vivo, we examined the effect of cycloheximide on SCD1mRNA levels. Cycloheximide was injected into mice 18 hr afterclofibrate feeding, and total liver RNA was isolated for RNaseprotection analysis at the end of 30 hr. This resulted in a 77%inhibition of SCD1 mRNA (Fig. 2C). This suggests thatongoing protein synthesis is required for the in vivo inductionof the SCD1 gene by clofibrate. No significant changes in actin
A
SCD1_probe ....~~~~~~~~~~~~~~~~~~~~~. ..............
,,',.'W
20 -
15
10
5-
0 10 20 30...........0
< 0 3 chow + cofibrateZ 0I 6 6 hII. 0 6 18 24 30 36 hrs
-SCD1mRNA
10 20 30Time (hours)
N9-
40
or albumin (data not shown) mRNA were observed in re-sponse to these manipulations (Fig. 2D).
Effects of Clofibrate on the Rate of Transcription of theSCD1 Gene. Nuclear run-on transcription assays were per-formed to determine if the clofibrate-mediated increase insteady-state levels of SCD1 mRNA was due to an increase ingene transcription, as opposed to an increase in mRNAstability. Experiments were performed using liver nuclei iso-lated from mice fed a chow diet supplemented with 1%clofibrate for 0, 18, and 36 hr. SCD1 gene transcription wasmeasured by quantitating the amount of radiolabeled RNAhybridized to pC3, a 2-kb SCD1 cDNA (21). Fig. 3A shows thatSCD1 transcription increased in response to clofibrate. Asshown in Fig. 3B, the transcription rate of the SCD1 geneincreased 3-fold after 18 hr and 14-fold after 36 hr. The SCD1transcriptional induction by carbohydrates is 11-fold (21). ThepC3 cDNA can hybridize to both SCD1 and SCD2 genetranscripts (21). Therefore, we also analyzed the SCD300cDNA, a SCD2-specific probe, to distinguish the expression ofthe two isoforms. No hybridization occurred to the SCD300cDNA. Thus, the clofibrate-mediated increase observed wasthat of SCD1. ,B-Actin was used as a control to ensure that thechange in transcription activity measured for the SCD1 genedoes not reflect a general increase in transcription in responseto clofibrate. pGEM vector was used as a negative control fornonspecific hybridization. No change in transcriptional activitywas detected with the f3-actin plasmid cDNA, and radiolabeledRNA from the transcription assays showed no hybridization topGEM. These results indicate that clofibrate treatment causesa specific increase in SCD1 gene transcription in mouse liver.
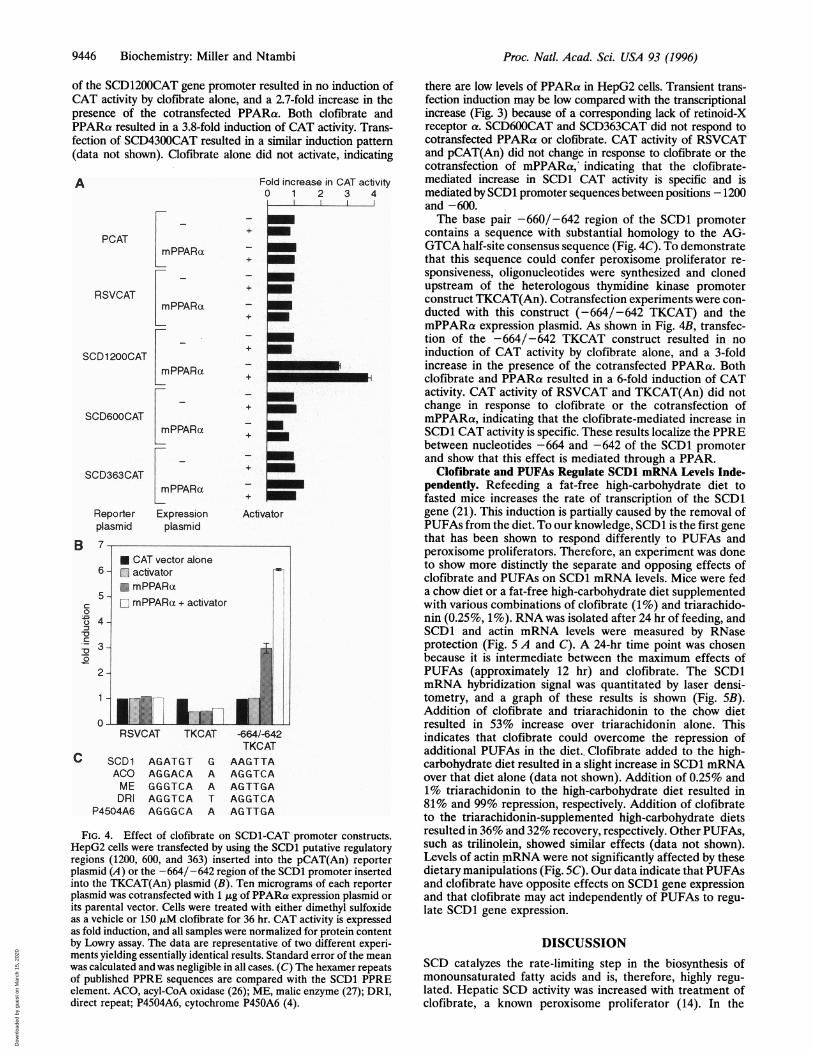
Effect of Clofibrate on SCD1-CAT Promoter Constructs. Tofurther demonstrate that the regulation of SCD1 by peroxi-some proliferators is transcriptional, fragments of the SCD1 5'flanking region containing putative regulatory elements werecloned into a CAT reporter plasmid for chimeric transfectionanalysis. Cotransfection experiments were conducted in HepG2cells with SCD4300CAT, SCD1200CAT, SCD600CAT, andSCD363CAT constructs and a mPPARa expression plasmid.This expression plasmid produces the a form of the mouse PPAR.mPPARa is known to be expressed and to mediate peroxisomeproliferator responses in liver. HepG2 cells were chosen becausethey are easily manipulated in culture and endogenously expressSCD1 gene (data not shown). As shown in Fig. 44, transfection
A
SCD1probe
0 18 36 hoursi. -:' -..... ...........! .: 'fpGEM -
SCD1 -
SCD2 -
f-Actin -
SCD1mRNA
D actinprobe -
:. ._-acbnmRNA
FIG. 2. Time course for clofibrate induction of liver SCD1 mRNA.Mice were fasted and then refed either a fat-free high-carbohydratediet (CHO) or Purina lab chow (fed for 36 hr) with or without 1%clofibrate. At times indicated, mice were sacrificed, and total liverRNA was isolated and analyzed by RNase protection using an SCD1-specific complementary probe (A). (B) The mRNA levels of SCD1 (a)and albumin (0) were quantitated by laser densitometry and are
presented as a fold increase in mRNA content over chow diet alone.Zero treatment is not shown in A. (C and D) Mice were starved andrefed a chow diet supplemented with 1% clofibrate; 18 hr afterclofibrate feeding, a single dose of cycloheximide (1 mg/100 g) was
intraperitoneally administered. Total liver RNA was isolated after 30hr of clofibrate feeding and analyzed by RNase protection using SCD1-(C) or actin- (D) specific complementary probes. Essentially identicalresults were obtained from three separate experiments.
0cc eCX 10.
55
u O
0 18 36 hours
FIG. 3. Effect of clofibrate on the transcription rate of SCD1 genein mouse liver. Nuclei were isolated from the livers of starved mice thathad been refed a chow diet supplemented with 1% clofibrate for 0, 18,or 36 hr. (A) Run-on transcription and hybridization with cDNAprobes and vector sequences were carried out as described (21).pGEM, pGEM plasmid; SCD1, pC3, a cDNA for mouse SCD gene;SCD2, SCD300, a cDNA for mouse SCD2 gene; and ,B-actin, human,B-actin. (B) The results were normalized to the value obtained at timezero and are presented as a fold change in transcription. Standarderror bars are obtained from two different experiments yieldingessentially identical results.
B <:za:Ec
0
._00
~0U-
C
Biochemistry: Miller and Ntambi
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020
9446 Biochemistry: Miller and Ntambi
of the SCD1200CAT gene promoter resulted in no induction ofCAT activity by clofibrate alone, and a 2.7-fold increase in thepresence of the cotransfected PPARa. Both clofibrate andPPARa resulted in a 3.8-fold induction of CAT activity. Trans-fection of SCD4300CAT resulted in a similar induction pattern(data not shown). Clofibrate alone did not activate, indicating
A
PCAT
RSVCAT
SCD 1 200CAT
SCD600CAT
SCD363CAT
Fold increase in CAT activity0 1 2 3 4i - - .
+
+
+
+
mPPARa
mPPARa
mPPARa
mPPARal-
LmPPARa
Reporter Expressionplasmid plasmid
+
Activator
RSVCAT TKCAT -664/-642TKCAT
SCD1 AGATGT G AAGTTAACO AGGACA A AGGTCAME GGGTCA A AGTTGADRI AGGTCA T AGGTCA
P4504A6 AGGGCA A AGTTGA
FIG. 4. Effect of clofibrate on SCD1-CAT promoter constructs.HepG2 cells were transfected by using the SCD1 putative regulatoryregions (1200, 600, and 363) inserted into the pCAT(An) reporterplasmid (A) or the -664/-642 region of the SCD1 promoter insertedinto the TKCAT(An) plasmid (B). Ten micrograms of each reporterplasmid was cotransfected with 1 gg of PPARa expression plasmid orits parental vector. Cells were treated with either dimethyl sulfoxideas a vehicle or 150 ,uM clofibrate for 36 hr. CAT activity is expressedas fold induction, and all samples were normalized for protein contentby Lowry assay. The data are representative of two different experi-ments yielding essentially identical results. Standard error of the meanwas calculated and was negligible in all cases. (C) The hexamer repeatsof published PPRE sequences are compared with the SCD1 PPREelement. ACO, acyl-CoA oxidase (26); ME, malic enzyme (27); DRI,direct repeat; P4504A6, cytochrome P450A6 (4).
there are low levels of PPARa in HepG2 cells. Transient trans-fection induction may be low compared with the transcriptionalincrease (Fig. 3) because of a corresponding lack of retinoid-Xreceptor a. SCD600CAT and SCD363CAT did not respond tocotransfected PPARa or clofibrate. CAT activity of RSVCATand pCAT(An) did not change in response to clofibrate or thecotransfection of mPPARa, indicating that the clofibrate-mediated increase in SCD1 CAT activity is specific and ismediated by SCD1 promoter sequences between positions -1200and -600.The base pair -660/-642 region of the SCD1 promoter
contains a sequence with substantial homology to the AG-GTCA half-site consensus sequence (Fig. 4C). To demonstratethat this sequence could confer peroxisome proliferator re-sponsiveness, oligonucleotides were synthesized and clonedupstream of the heterologous thymidine kinase promoterconstruct TKCAT(An). Cotransfection experiments were con-ducted with this construct (-664/-642 TKCAT) and themPPARa expression plasmid. As shown in Fig. 4B, transfec-tion of the -664/-642 TKCAT construct resulted in noinduction of CAT activity by clofibrate alone, and a 3-foldincrease in the presence of the cotransfected PPARa. Bothclofibrate and PPARa resulted in a 6-fold induction of CATactivity. CAT activity of RSVCAT and TKCAT(An) did notchange in response to clofibrate or the cotransfection ofmPPARa, indicating that the clofibrate-mediated increase inSCD1 CAT activity is specific. These results localize the PPREbetween nucleotides -664 and -642 of the SCD1 promoterand show that this effect is mediated through a PPAR.
Clofibrate and PUFAs Regulate SCD1 mRNA Levels Inde-pendently. Refeeding a fat-free high-carbohydrate diet tofasted mice increases the rate of transcription of the SCD1gene (21). This induction is partially caused by the removal ofPUFAs from the diet. To our knowledge, SCD1 is the first genethat has been shown to respond differently to PUFAs andperoxisome proliferators. Therefore, an experiment was doneto show more distinctly the separate and opposing effects ofclofibrate and PUFAs on SCD1 mRNA levels. Mice were feda chow diet or a fat-free high-carbohydrate diet supplementedwith various combinations of clofibrate (1%) and triarachido-nin (0.25%, 1%). RNA was isolated after 24 hr of feeding, andSCD1 and actin mRNA levels were measured by RNaseprotection (Fig. 5 A and C). A 24-hr time point was chosenbecause it is intermediate between the maximum effects ofPUFAs (approximately 12 hr) and clofibrate. The SCD1mRNA hybridization signal was quantitated by laser densi-tometry, and a graph of these results is shown (Fig. SB).Addition of clofibrate and triarachidonin to the chow dietresulted in 53% increase over triarachidonin alone. Thisindicates that clofibrate could overcome the repression ofadditional PUFAs in the diet.. Clofibrate added to the high-carbohydrate diet resulted in a slight increase in SCD1 mRNAover that diet alone (data not shown). Addition of 0.25% and1% triarachidonin to the high-carbohydrate diet resulted in81% and 99% repression, respectively. Addition of clofibrateto the triarachidonin-supplemented high-carbohydrate dietsresulted in 36% and 32% recovery, respectively. Other PUFAs,such as trilinolein, showed similar effects (data not shown).Levels of actin mRNA were not significantly affected by thesedietary manipulations (Fig. SC). Our data indicate that PUFAsand clofibrate have opposite effects on SCD1 gene expressionand that clofibrate may act independently of PUFAs to regu-late SCD1 gene expression.
DISCUSSIONSCD catalyzes the rate-limiting step in the biosynthesis ofmonounsaturated fatty acids and is, therefore, highly regu-lated. Hepatic SCD activity was increased with treatment ofclofibrate, a known peroxisome proliferator (14). In the
B 7-
C0
-
Q
0
-0Co
c
Proc. Natl. Acad. Sci. USA 93 (1996)
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020

Proc. Natl. Acad. Sci. USA 93 (1996) 9447
present study, the mechanism responsible for this inductionwas investigated. To determine if the induction was due to anincrease in mRNA, we examined the effects of clofibrate andgemfibrozil administered in a normal chow diet for 36 hr on theaccumulation of SCD1 mRNA in mouse liver. Both drugsactivated SCD1 mRNA synthesis equally well (Fig. 1). A timecourse study showed that SCD1 mRNA induction was appar-ent within 6 hr and reached a maximum of 22-fold after 30 hr.Cycloheximide treatment showed that ongoing protein syn-thesis is required for SCD1 mRNA induction by peroxisomeproliferators (Fig. 2). Nuclear run-on transcription assaysshowed that the increase of SCD1 mRNA was a consequenceof an increase in the rate of SCD1 gene transcription (Fig. 3).Transfection analysis revealed that the PPRE was at positions-664/-642 of the SCD1 promoter (Fig. 4). This element wasinitially speculated to be present on the basis of sequencehomology (11 of 15 base pairs) with the PPRE consensussequence (28). Supplementation of a chow diet or fat-freehigh-carbohydrate diet with clofibrate and/or arachidonic acidshowed that clofibrate and PUFAs have opposing effects onSCD1 mRNA levels (Fig. 5). The response of the SCD1 geneis unlike that of lipogenic genes, such as S14 and fatty acidsynthase, which are repressed by both peroxisome proliferatorsand PUFAs (5).
Peroxisome proliferation-activating drugs lower blood lipidlevels by stimulating increased hepatic uptake of triacylglyc-erides (1), which, in turn, results in transcriptional regulationof genes involved in lipid homeostasis. The triacylglyceride/fatty acid balance is influenced by changes in three major liverpathways: peroxisomal ,B-oxidation, fatty acid biosynthesis,and phospholipid biosynthesis. Gene transcription of the en-zymes involved in (3-oxidation and phospholipid biosynthesisare induced up to 20-fold in response to these drugs (2-4). Tolimit fatty acid accumulation, lipogenic enzymes such asacetyl-CoA carboxylase, fatty acid synthase, and S14 arerepressed (5, 29, 30), whereas stearoyl-CoA desaturation,palmitoyl-CoA elongation, and A6 and A5 desaturation activ-ities are induced by administration of peroxisome proliferators(14, 15, 31).The observable effects of administering peroxisome prolif-
erators to rodents are increased size of organelles, includingperoxisomes, mitochondria, and the endoplasmic reticulum,and enlargement of the liver itself (12, 32). Hepatocytes needto increase the supply of phospholipids, which are required forthe rapid proliferation of organelles and the plasma mem-brane. Phospholipids contain two fatty acids, the one atposition 2 (sn-2) usually being unsaturated. Peroxisome pro-liferators cause an influx of fatty acids, providing an immediatesupply of PUFAs. However, PUFAs are preferentially brokendown by peroxisomal 13-oxidation (33), causing a depletion ofthese PUFAs. Under conditions of essential fatty acid defi-ciency, SCD is activated, because it catalyzes the synthesis ofthe only unsaturated fatty acids that mammals can synthesizede novo (34). Therefore, the induction of SCD may be neces-sary to compensate for a lack of PUFAs in the membrane andto enhance phospholipid biosynthesis. Previous studies haveshown that clofibrate results in an increase in 18:1 found inphosphatidylcholine (15), supporting this hypothesis.PUFAs activate PPARs and were hypothesized to be the
endogenous activator of this orphan receptor (12). Lipogenicgenes, such as S14 and fatty acid synthase, are repressed byboth peroxisome proliferators and PUFAs (5, 35). It has beenspeculated that there is a PPRE-like binding site in the knownPUFA response element of the fatty acid synthase gene. Thus,PPAR and PUFA effects might be mediated through the sametrans-acting factor (28, 36). This theory was partially refutedbecause PPARs are activated by both saturated and unsatur-ated fatty acids. Therefore, PPARs do not seem to have thespecificity to mediate PUFA-specific repression of lipogenicgenes (37, 38).
A
SCD1_ Oaprobe *...
B .G < 4C0z
m)cc 302c
02R ic1Q(,)r- IL%L#tagpctr.obe . ..
probe- "I'
+chow +CHO
6\oc:,\ , R° c:)
..... ,.,.,,,,,,,.,, _~~~~~~~~~~SCRD... ..:::':'..SS'S!
actn_ ~~~~mRNA
FIG. 5. Clofibrate and PUFAs regulate SCD1 mRNA levels inde-pendently. Mice were starved and refed a chow diet or a fat-freehigh-carbohydrate diet (CHO) supplemented with various combina-tions of clofibrate (1% clof) and triarachidonin (0.25% AA, 1% AA)for 24 hr. Total liver RNA was isolated and analyzed by RNaseprotection using SCD1- (A) or actin- (C) specific complementaryprobes. (B) The mRNA levels of SCD1 were quantitated by laserdensitometry and are presented as a fold increase in mRNA contentover chow. The data are representative of three different experimentsyielding essentially identical results.
In vivo studies in this paper conclusively show that PUFAsand peroxisome proliferators have opposing effects on theexpression of the SCD1 gene. Transient transfections havelocalized the SCD1 PUFA response element to an area of theSCD1 promoter that is distinct from the PPRE (K. Waters,C.W.M., and J.M.N., unpublished data). Our results furthersuggest that PUFAs are not the endogenous activator ofPPARs. However, cotransfection of PPARa and treatmentwith 300 ,uM linolenic acid showed a slight increase inSCD4300CAT activity (data not shown). This indicates thatunphysiological concentrations of PUFAs can be used toinduce PPARs, as shown in other systems. It has also beenhypothesized that the induction of SCD1 may be a response toa PUFA deficiency, and therefore may be a secondary effectof peroxisome proliferator action (15). However, cotransfec-tion data presented here show that PPARa is directly involvedin the SCD1 transcriptional induction.
In vivo studies showed activation of peroxisomal (3-oxidationgenes was 9- to 15-fold within 1 hr after clofibrate adminis-tration, and it increased to a maximum within a few hours (37).A significant SCD1 mRNA induction is visible only after 24 hrof clofibrate supplementation, suggesting that a protein crucialfor this transcriptional induction is synthesized during thistime. This hypothesis is supported by the observation thatprotein synthesis is required for maximum SCD1 induction byperoxisome proliferators. Differential regulation using aPPAR isoform, varied heterodimerization with other steroidhormones, or effect of some unknown protein on PPARbinding could result in induction of the SCD1 gene. Also, oneof the two PPAR mouse isoforms expressed in liver could bedirectly induced during this lag, suggesting the possibility ofmultiple signaling pathways (10).The ratio of oleic to stearic acid is important in maintenance
of membrane fluidity (16). Small changes in this ratio, and inthe amount of available PUFAs, can affect the ability of the cell
Biochemistry: Miller and Ntambi
II
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020

9448 Biochemistry: Miller and Ntambi
to respond to external stimuli. Inability of the cell to maintainan appropriate ratio has been linked to carcinogenesis (16).Hepatic tumors are induced as a result of long-term peroxi-some proliferator administration to mice and rats (1). This maybe due, in part, to an increase in SCD1 activity and analteration of oleic to stearic acid ratios. All of the implicationsof changing the activity of SCD are not known. However, theopposing effects of PUFAs and clofibrate on SCD1 transcrip-tion have led to new insight into the mechanism ofPPAR generegulation. Further studies will be needed to elucidate thephysiological ramifications of induction of SCD1 transcriptionand to identify the endogenous PPAR ligands.
We thank David Casimir and Katrina Waters for useful discussionsand comments. This work was supported by National Institutes ofHealth Grant DK42825.
1. Reddy, J. K. & Lalwai, N. D. (1983) CRC Crit. Rev. Toxicol. 12,1-58.
2. Marcus, S. L., Miyata, K. S., Zhang, B., Subramani, S., Rachu-binski, R. A. & Capone, J. P. (1993) Proc. Natl. Acad. Sci. USA90, 5723-5727.
3. Alvares, K., Fan, C., Dadras, S. S., Yeldandi, A. V., Rachubinski,R. A., Capone, J. P., Subramani, S., lannaccone, P. M., Rao,M. S. & Reddy, J. K. (1994) Cancer Res. 54, 2303-6.
4. Muerhoff, A. S., Griffen, K. J. & Johnson, E. F. (1992) J. Biol.Chem. 267, 19051-19053.
5. Jump, D. B., Ren, B., Clarke, S. & Thelen, A. (1995) Prostaglan-dins, Leukotrienes Essent. Fatty Acids 52, 107-111.
6. Kawashima, Y., Mizuguchi, H., Musoh, K. & Kozuka, H. (1994)Biochim. Biophys. Acta 1212, 311-318.
7. Issemann, I. & Green, S. (1990) Nature (London) 347, 645-650.8. Dreyer, C., Krey, G., Keller, H., Givel, F., Helftenbein, G. &
Wahli, W. (1992) Cell 68, 879-887.9. Tontonoz, P., Hu, E., Graves, R. A., Budavar, A. I. &
Spiegelman, B. M. (1994) Genes Dev. 8, 1224-1234.10. Zhu, Y., Alvares, K., Huang, Q., Rao, M. S. & Reddy, J. K. (1993)
J. Biol. Chem. 268, 26817-26820.11. Kliewer, S. A., Forman, B. M., Blumberg, B., Ong, E. S., Borg-
meyer, V., Mangelsdorf, D. J., Umesono, K. & Evans, R. M.(1994) Proc. Natl. Acad. Sci. USA 91, 7355-7359.
12. Gottlicher, M., Widmark, E., Li, Q. & Gustafssin, J. (1992) Proc.Nati. Acad. Sci. USA 89, 4653-4657.
13. Sher, T., Yi, H., McBride, 0. W. & Gonzalez, F. S. (1993)Biochemistry 32, 5598-5604.
14. Kawashima, Y., Hanioka, N., Matsumura, M. & Kozuka, H.(1983) Biochim. Biophys. Acta 752, 259-264.
15. Kawashima, Y., Musoh, K. & Kozuka H. (1990) J. Biol. Chem.265, 9170-9175.
16. Tebbey, P. W. & Buttke, T. M. (1992) Biochim. Biophys. Acta1171, 27-32.
17. Birnboim, H. C. & Doly, J. (1979) Nucleic Acids Res. 7, 1513-1523.
18. Thiede, M. A. & Strittmatter, P. (1985) J. Biol. Chem. 260,14459-14463.
19. Chirgwin, J. M., Przybyla, A. E., MacDonald, R. J. & Rutter,W. J. (1979) Biochemistry 18, 5294-5299.
20. Melton, D. A., Krieg, P. A., Rebaghiati, M. R., Maniatis, T.,Zinn, K & Green, M. R. (1984) NucleicAcids Res. 12,7035-7056.
21. Ntambi, J. M. (1992) J. Bio. Chem. 267, 10925-10930.22. Kadesch, T. & Berg, P. (1986) Mol. Cell Biol. 6, 2593-2601.23. Felgner, P. L., Gadek, T. R., Holm, M., Roman, R., Chan, H. W.,
Wenz, M., Northrop, J. P., Ringold, G. M. & Danielsen, M.(1987) Proc. Natl. Acad. Sci. USA 84, 7413-7417.
24. Neumann, J. R., Morency, C. A. & Russian, K. 0. (1987) Bio-Techniques 5, 444-447.
25. Kaestner, K. H., Ntambi, J. M., Kelly, T. J., Jr., & Lane, M. D.(1989) J. Biol. Chem. 264, 14755-14761.
26. Tugwood, J. D., Issemann, I., Anderson, R. G., Bundell, K. R.,McPheat, W. L. & Green, S. (1992) EMBO J. 11, 433-439.
27. Castelein, H., Gulick, T., Declercq, P. E., Mannaerts, G. P.,Moore, D. D. & Baes, M. I. (1994) J. Biol. Chem. 269, 26754-26758.
28. Landschulz, K T., Jump, D. B., MacDougald, 0. A. & Lane,M. D. (1994) Biochem. Biophys. Res. Commun. 200, 763-768.
29. Hertz, R., Kalderon, B. & Bar-Tans, J. (1993) Biochimie 75,257-261.
30. Kochan, Z., Bukato, G. & Swierezynski, J. (1993) Biochem.Pharmacol. 46, 1501-6.
31. Kawashima, Y., Hirose, A. & Kozuka H. (1984) Biochim. Biophys.Acta 795, 543-551.
32. Reddy, J. K. & Krishnakantha, T. P. (1975) Science 190, 787-789.33. Reddy, J. K. & Mannaerts, G. P. (1994) Annu. Rev. Nutr. 14,
343-70.34. Jeffcoat, R. (1979) Essays Biochem. 15, 1-36.35. Blake, W. L. & Clarke, S. D. (1990) J. Nutr. 120, 1727-1729.36. Girard, J., Perdereau, D., Foufelle, F., Prip-Buus, C. & Ferre, P.
(1994) FASEB J. 8, 36-42.37. Keller, H., Matifoudi, A., Dreyer, C., Hihi, A. K., Medin, J.,
Ozato, K. & Wehli, W. (1993)Ann. N.Y Acad. Sci. 684, 157-173.38. Reddy, J. K, Goel, S. K., Nemali, M. R., Carrino, J. J., Laffler,
T. G., Reddy, M. K., Sperbeck, S. J., Osumi, T., Hashimoto, T.,Lalwani, N. D. & Rao, M. S. (1986) Cell Biol. 83, 1747-1751.
Proc. Natl. Acad. Sci. USA 93 (1996)
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020