pharmacokineticsintheratofapanelofimmunotoxinsmadewithabri...
TRANSCRIPT

[CANCER RESEARCH 50, 7519-7526. December 1, 1990]
Pharmacokinetics in the Rat of a Panel of Immunotoxins Made with Abrin AChain, Ricin A Chain, Gelonin, and Momordin1
Edward J. Wawrzynczak,2 Alan J. Cumber, Raymond V. Henry, Julie May, David R. Newell,3 Geoffrey D. Parnell,
Nan R. Worrell, and J. Anthony ForresterDrug Targeting Laboratory, Section of Medicine ¡E.J. W., A. J. C., R. V. H., J. M., G. D. P.. N. R. W.. J. A. F.], and Section of Drug Development [D. R. N.J. Instituteof Cancer Research, Cotswold Road, Sulton, Surrey, SM2 5A/G, England
ABSTRACT
A panel of immunotoxins was constructed by chemically attaching theribosome-inactivating proteins abrin A chain, ricin A chain, gelonin, andmomordin to the monoclonal mouse IgG2a antibody Fib75 by means ofa disulfide linkage. All the immunotoxins were toxic in tissue culture tothe I.I human bladder carcinoma cell line expressing the antigen recognized by Fib75, inhibiting the incorporation of |'U|lcmine by 50% atconcentrations between 1 x IO"10M and 8 x 10"'°M. The pharmacoki-
netics of the immunotoxins in the normal Wistar rat was determinedfollowing i.v. administration. The concentrations of intact immunotoxinin serum samples taken at various intervals after injection for up to 24 hwere measured by solid-phase enzyme-linked immunosorbent assaysspecific for each of the four different ribosome-inactivating proteins. TheFib75 immunotoxins were cleared from the circulation with comparable,but not identical, biphasic kinetics best described by a two compartmentopen pharmacokinetic model. The a-phase half-lives of the panel, between 0.35 and 0.71 h, were similar. The #-phase half-life of Fib75-abrinA chain, 13.3 h, was significantly longer than the ¿f-phasehalf-lives ofFib75-ricin A chain, Fib75-gelonin, and Fib75-momordin, between 7.5and 8.6 h. Fib75-abrin A chain was found to be about 3- to 4-fold moreresistant than the other immunotoxins to breakdown by reduction of thedisulfide linkage between the A chain and the antibody with glutathionein vitro. This suggests that the longer serum half-life of Fib75-abrin Achain may have been due to greater stability against reduction in vivo.Analysis of serum samples obtained up to 24 h after injection of Fib75-abrin A chain revealed that the chemically intact immunotoxin presentin the circulation retained full cytotoxic activity. An abrin A chainimmunotoxin made with a different monoclonal mouse IgG2a antibodywas also found to be more stable against reduction by glutathione in vitrothan an analogous ricin A chain immunotoxin. Thus, abrin A chain maypossess unique molecular properties that endow immunotoxins made withthis A chain with greater stability in vivo than immunotoxins made withricin A chain or other ribosome-inactivating proteins.
INTRODUCTION
ITs4 are hybrid protein molecules consisting of a toxic protein
linked to an antibody. Many agents of this type can bindselectively to cells that express the target antigen recognized bythe antibody component and kill the cells by the intracellularaction of the toxic protein. Immunotoxins made using antibodies recognizing tumor-associated antigens have been shown toexert potent and selective cytotoxic effects in animal tumourmodels and are currently undergoing clinical trials in cancer
Received 6/12/90; accepted 8/30/90.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
' This work was supported by funds from the Medical Research Council and
the Cancer Research Campaign, United Kingdom.! To whom requests for reprints should be addressed.3Present address: University of Newcastle-upon-Tyne, Department of Clinical
Oncology, Cancer Research Unit, Medical School, Framlington Place, Newcastle-upon-Tyne, NE2 4HH. England.
*The abbreviations used are: IT, immunotoxin; ELISA, enzyme-linked im
munosorbent assay: HPLC, high performance liquid chromatography; PAGE,polyacry'amide gel electrophoresis; PBS. phosphate-buffered saline; RIP. ribosome-inactivating protein; SDS. sodium dodecyl sulfate; SPDP, /V-succinimidyl-3-(2-pyridyldithio)propionate.
patients (reviewed in Refs. 1-4).Ricin A chain, a RIP isolated from the plant toxin ricin, has
most frequently been used in the synthesis of ITs. The firstexperimental studies in which ricin A chain ITs were administered i.v. to rabbits and rats revealed that the IT was rapidlylost from the circulation (5-7). Subsequent studies clearly demonstrated that the cause of the initial rapid clearance afterinjection of the IT was specific receptor-mediated hepatic recognition of an oligomannose side chain present exclusively onthe A2 form of native ricin A chain (8-10). Three strategieshave been shown to minimize uptake of ricin A chain ITs bythe liver in vivo: (a) saturation of the hepatic receptor systemby coadministration of the IT and an excess of mannose-
containing protein or carbohydrate (8, 9, 11); (b) chemicaltreatment of the native ricin to destroy the mannose residues(10); and (c) the use of the A, form of native ricin A chainwhich lacks the oligomannose side chain (12).
An alternative approach to bypassing the problem of hepaticclearance is to construct ITs with RIPs other than ricin A chain.A large family of RIPs of diverse plant origin has now beenidentified (13). These RIPs resemble ricin A chain in size andmode of action but differ from ricin A chain and from oneanother in primary structure, pi, and the degree and type ofglycosylation. Pharmacokinetic studies have been reported forITs made with abrin A chain (14), pokeweed antiviral protein(15, 16), gelonin (16-21), saporin (17, 18, 22-25), and tricho-kirin (26). A common feature of the majority of these studiesis the finding that all types of IT have a much shorter half-lifein the bloodstream than the parent antibody. However, it isdifficult to make direct comparisons between the results of thevarious studies because of major differences in experimentaldetail. These differences include: (a) the nature of the antibodies; (b) the methods of IT preparation and purification; (c) theanimal species; (d) the times of the sampling points; (e) theprocedures for IT quantification; and (/) the methods of pharmacokinetic analysis.
In this study, we have measured the blood clearance of apanel of ITs prepared using a single monoclonal antibody,Fib75, which recognizes an integral membrane glycoprotein(M, 19,000) present on most differentiated normal and neo-plastic human cells and not expressed by rodent tissues. Cytotoxic ITs were prepared by similar chemical procedures usingabrin A chain, native ricin A chain, gelonin, and momordin.Samples of IT were injected into normal rats i.v. and bloodsamples were taken at identical times after injection. The concentration of IT in serum was determined using RIP-specificELISAs detecting only intact IT molecules. All the experimental data were analyzed by the same mathematical procedures.This study showed that the Fib75-abrin A chain IT had thelongest blood half-life of the ITs in the panel. The disulfidebond linking abrin A chain to the Fib75 antibody was found tobe less susceptible to reduction by glutathione in vitro than thedisulfide linkages in the ITs made with the other RIPs suggest-
7519
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

PHARMACOKINETICS OF IMMUNOTOXINS IN THE RAT
ing that the longer persistence of the abrin A chain IT may beattributed, in whole or in part, to a slower rate of breakdownin vivo.
MATERIALS AND METHODS
Materials
Castor bean cake derived from seeds of Ricinus communis of Kenyanorigin was obtained from Croda Premier Oils, Hull, Humberside,England. Seeds of Abrus precatorius were obtained from the Departmentof Botany, University of Ghana, Accra, Ghana. Seeds of Geloniummultiflorum and Momordica charantia were from United Chemicalsand Allied Products, Calcutta, India.
The murine hybridoma LICR-LOND Fib75 secreting a mouse monoclonal antibody of the IgG2a isotype was provided by Dr. R. A. J.Mcllhinney. The antibody Fib75 was purified from the ascitic fluid ofhybridoma-bearing mice by ammonium sulfate precipitation followedby chromatography on immobilized staphylococcal protein A usingstepwise elution. The mouse monoclonal antibody 2AL-1, raisedagainst vesicular stomatitis virus and also of the lgG2a isotype, waspurified by a similar procedure.
Chromatography media, Sephadex G-25 (F), Sephacryl S-200, Seph-arose 4B, protein A-Sepharose 4B, and CNBr-activated Sepharose 4B,were purchased from Pharmacia, Ltd., Milton Keynes, Buckinghamshire, England. TSK-G3000 HPLC columns were purchased fromAnachem, Ltd., Luton, Bedfordshire, England.
Sheep anti-mouse immunoglobulin-horseradish peroxidase (NA.931 ),sodium ['"IJiodide (IMS.30), and L-[4,5-'H]leucine (TRK.170) were
purchased from Amersham International pic., Amersham, Buckinghamshire, England. Emulsifier-Safe liquid scintillation cocktail was obtained from Canberra Packard Ltd., Pangbourne, Berkshire, England.lodo-Gen was purchased from Pierce (UK), Ltd., Chester, England.
lodoacetamide was obtained from BDH, Ltd., Poole, Dorset, England. Glutathione and o-phenylenediamine were purchased from Sigma
Chemical Co., Ltd., Poole, Dorset, England. SPDP was from Pharmacia. All other reagents were of the highest purity available.
Tissue culture media, RPMI 1640 and Dulbecco's modified Eagle's
medium, and fetal calf serum were purchased from Gibco, Ltd., Paisley,Scotland.
Flat-bottomed 96-well micro-ELISA plates (Immulon 2) were obtained from Dynatech Laboratories, Ltd., Billingshurst, Sussex, England. Sterile tissue culture plates (24 wells) were from Nunclon, Ltd.,Uxbridge, Middlesex, England.
Normal male albino Wistar/CBI rats, 8-16 weeks old, were suppliedby the M.R.C. Animal Breeding Unit, National Institute of MedicalResearch, London, England, and allowed free access to food and water.Male New Zealand White rabbits were obtained from J. & L. G.Phillips, Ltd., Petersfield, Hampshire, England.
Purification of Ribosome-inactivating Proteins
Ricin was purified from an aqueous extract of defatted castor beancake by ammonium sulfate precipitation, affinity chromatography onacid-treated Sepharose 4B, and gel permeation chromatography onSephacryl S-200 essentially as described by Nicolson and Blaustein (27)with the modifications previously described by Cumber et al. (28). RicinA chain was isolated from the toxin by reductive cleavage and furtherpurified by the method described by Forrester et al. (29). This preparation gave two bands with apparent molecular weights of about 30,000and 32,000 when examined by SDS-PAGE corresponding to the twodifferently glycosylated forms of the A chain.
Abrin was purified from defatted meal of A. precatorius seeds andabrin A chain was isolated from the toxin and purified by proceduresidentical with those described for ricin A chain (28, 29). The preparationgave a single band on SDS-PAGE with an apparent molecular weightof about 29,000.
Gelonin was prepared as described by Stirpe et al. (30). Momordinwas prepared according to the method of Barbieri et al. (31). Purified
gelonin and momordin both appeared as single bands with apparentmolecular weights of about 30,500 and 31,000, respectively, by SDS-PAGE.
Preparation and Characterization of Immunotoxins
Abrin A chain and ricin A chain were attached to the Fib75 antibodyusing the methodology described in detail by Cumber et al. (28). Briefly,2-pyridyl disulfide groups were first introduced into Fib75 at an averagederivatization level of about 1.8 groups/antibody by reaction withSPDP. The derivatized antibody was then reacted with an excess offreshly reduced toxin A chain. The reaction mixture was applied to acolumn of Sephacryl S-200 and the material which eluted at a positioncorresponding to IT (M, 180,000-210,000) was collected. Immunotoxins made with the 2AL-1 antibody were prepared by the same procedure.
Gelonin and momordin were modified by reaction with SPDP tointroduce an average of 1.0 2-pyridyl disulfide groups/RIP moleculeand were reduced with dithiothreitol before mixing with derivatizedantibody as above. The gelonin and momordin ITs were purified freeof unconjugated RIP by gel permeation chromatography on SephacrylS-200 as for the toxin A chain ITs.
The IT preparations were analyzed by SDS-PAGE and gel permeation HPLC. Electrophoresis was performed on 4.0-12.5% gradientpolyacrylamide gels in the presence of 0.2 g SDS/100 ml solution (32).Samples were prepared for electrophoresis in the absence of reducingagent to preserve the disulfide linkage between the antibody and RIP.Gel permeation HPLC was performed on a TSK-G3000SW column(7.5 mm inside diameter x 60 cm). Samples (0.1 ml) were applied tothe column and eluted at a flow rate of 0.03 ml/min. The runningbuffer was 20 mivi sodium phosphate-0. l M sodium sulfate, pH 6.8,containing 0.05 g NaNi/100 ml solution. The column was calibratedusing protein standards of known molecular weight.
The RIP content of the final IT preparations was determined asdescribed by Cumber et al. (28). The content of unconjugated antibodyand different IT species in the final preparations was estimated following SDS-PAGE by densitometry of Coomassie blue-stained proteinbands using a Bio-Rad Model 620 video densitometer.
Preparation of Affinity-purified Anti-RIP Antibody
Rabbit antisera to abrin A chain, ricin A chain, gelonin, and momordin were produced by the procedure described by Worrell et al.(17). Abrin A chain- and ricin A chain-specific antibody was isolatedfrom serum by affinity chromatography on columns of A. precatoriusagglutinin and R. communis agglutinin, respectively, linked to CNBr-activated Sepharose 4B. Gelonin- and momordin-specific antibody wasisolated from antiserum by affinity chromatography on a column of theappropriate RIP linked to CNBr-activated Sepharose 4B.
Measurement of Immunotoxin Concentration in Serum Samples by Enzyme-linked Immunosorbent Assay
The solid-phase ELISA procedure used was the method originallydescribed for detecting intact ITs made with ricin A chain (33). Briefly,affinity-purified anti-RIP antibody was first immobilized on micro-ELISA plates. Serum samples containing IT were added to the platesand, following incubation and washing, the bound IT was detectedusing anti-mouse immunoglobulin-horseradish peroxidase in combination with an o-phenylenediamine substrate solution developing colorat 492 nm. The mean absorbance at 492 nm of triplicate serum sampleswas used to calculate IT concentration at each time point by referenceto a standard curve spanning a concentration range of 0.125-15 ng/ml. The maximum standard deviation from the determined mean valuesranged between 5 and 9% in individual experiments.
Measurement of IT Breakdown in Vitro
ELISA. A stock solution of glutathione was prepared in PBS at aconcentration of 10 mivi and sterilized by passage through a 0.22-¿<mfiltration unit (Millex GV). The concentration of reduced glutathionein the stock solution was confirmed by measuring the amount of reactive
7520
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

PHARMACOKINETICS OF IMMUNOTOXINS IN THE RAT
sulfhydryl groups in dilutions of the stock solution using Ellman's
reagent (34). Sterile solutions of IT containing between 0.3 and 0.5 ugof conjugated RIP per ml were prepared in PBS containing 4% (w/w)alkylated rat serum to provide carrier proteins. Each IT solution (0.5ml) was mixed with an equal volume of 10 BIM glutathione solutionand incubated at 37°Cunder sterile conditions. Control samples were
treated in identical fashion except that the glutathione solution wasreplaced by PBS. Samples were removed after different times of incubation up to 8 h and mixed with an equal volume of 20 mM iodoacet-amide solution in PBS to block free sulfhydryl groups. The concentration of intact IT in each sample was determined by a solid-phase ELISAsimilar to the method described above. Rate constants for the slowerphase of the reaction were calculated assuming pseudo-first order
reaction kinetics.Gel Permeation HPLC. Solutions of glutathione were prepared in
PBS at concentrations of 0.1 M, 10 mM, and 1 mM and sterilized byfiltration. Sterile IT solutions containing between 100 and 170 ¿igofconjugated RIP per ml were prepared in PBS without the addition ofcarrier protein. Each IT solution (135 pi) was mixed with 15 ¿ilofglutathione solution at the three concentrations. After incubation for 1h at 37°Cunder sterile conditions, iodoacetamide was added to a final
concentration of 13 mM. Control samples were treated in identicalfashion except that the glutathione was omitted. Samples (0.1 ml) ofthe IT solutions were analyzed by gel permeation HPLC on a TSK-G3000SWXL column (7.8 mm inside diameter x 30 cm) equilibratedwith 20 mM sodium phosphate-0. l Msodium sulfate, pH 6.8, containing0.05 g of NaN VI 00 ml of solution at a flow rate of 0.4 ml/min. Theabsolute amounts of ricin A chain or abrin A chain released from intactIT by coincubation with glutathione were calculated by comparing thepeak heights of material eluting with the retention time of the uncon-jugated A chains and the peak heights from linear standard curvesgenerated with samples of the appropriate A chain at concentrations of1-15 MgA chain/ml. The total A chain content of the ITs was determined from the A chain released by incubation of the IT in the presenceof 10 mM glutathione for 5 h at 37°C.
Cytotoxicity Experiments in Tissue Culture
Cytotoxicity experiments using the EJ human bladder carcinomacell line were carried out essentially as described by Forrester et al.(29). Briefly, dilutions of IT solution were added to subconfluentmonolayer cultures of EJ cells in triplicate and incubated for 24 h.[3H]Leucine (1 jjCi) was added to each culture followed by incubation
for a further 24 h. At the end of this period, the incorporation of['Hjleucine was determined by liquid scintillation counting. The resultswere expressed as a percentage of the ['Hjleucine incorporated bycontrol cultures not receiving IT. The incorporation of [•'Hjleucineby
control cultures was greater than 80,000 cpm.
Blood Clearance Measurements
Immunotoxins. ITs were prepared in sterile solution at concentrationsof conjugated RIP between 109 and 187 ¿ig/ml.These preparationswere checked by gel permeation HPLC shortly before administrationto animals in order to ensure the absence of aggregated protein. Clearance studies »ereperformed as described by Worrell et al. (7) followinga single i.v. injection of 11-22 ^g of conjugated RIP. The concentrationof intact IT in serum samples was determined using the ELISA procedure described above. The experimental data were adjusted accordingto the amount of conjugated RIP injected and animal weight in eachexperiment and are expressed as the serum concentration based on theinjection of 10 /jg of conjugated RIP/300 g of rat weight.
Ribosome-inactivating Proteins. RIPs were radiolabeled to a specificactivity of 200-400 ^Ci '"I/mg protein using the lodo-Gen method
(35). In the case of abrin A chain and ricin A chain, the intact toxinswere labeled and the A chains were isolated subsequently using theusual procedure. Groups of rats, treated as above, received injectionsof I25I-RIP solution in PBS containing between 1.0 and 1.3 x 106cpm.
Samples of blood taken at intervals between 2 and 30 min after injection
were weighed and the radioactivity present in the samples was determined in a Packard 5266 gamma counter.
Pharmacokinetic Analysis
Blood clearance curves were fitted to the determined serum levels ofIT by a computerized nonlinear least-squares regression algorithm (36).The experimental data were most consistent with a two compartmentopen pharmacokinetic model described by the biexponential equation
C = Ac'"' + Be'1"
where C is the concentration at time / and A, B and «,ßare theconcentration and rate constants, respectively. The weighting function\/(Y + Y)2 was applied to all measurements (37). The C0 values werecalculated from the fitted curves by back extrapolation, i.e., Co = A +B. The «-and /3-phase half-lives were calculated as rw= 0.6931 /a or ß.The area under the serum concentration versus time curve at infinitetime after injection (ADC) was calculated as the integral of the curveusing the formula
ADC = A/a + B/ß
RESULTS
Characterization of the Fib75 Immunotoxins. Fib75 ITs madewith abrin A chain, ricin A chain, gelonin, and momordin weresynthesized using closely matched preparative conditions. Ineach case, the IT preparation was completely separated fromunconjugated RIP and partially resolved from unmodified antibody by a single step of gel permeation chromatography onSephacryl S-200. The four IT preparations all had a similarcomposition as judged by SDS-PAGE (Fig. 1). Densitometrywas used to calculate the mean percentage content of thedifferent molecular species and the standard deviations fromthe mean values. The predominant species present was thesingly substituted antibody-RIP conjugate [51 ±3% (SD)J,containing one RIP molecule linked to one antibody molecule,with lesser amounts of multiply substituted antibody-RIP con
jugates [23 ±12%] and of unconjugated Fib75 antibody [26 ±11%]. All the IT preparations were further analyzed by gelpermeation HPLC. Fig. 2 shows the elution profile of Fib75-abrin A chain which is representative of the elution profilesseen for all the ITs in the figure. This HPLC analysis showed
1
Fig. 1. SDS-PAGE analysis of Fib75 antibody and ITs. Samples were run ona 4-12.5% gradient polyacrylamide gel under nonreducing conditions. Proteinbands were visualized by Coomassie Brilliant Blue staining. Lane I, unconjugatedFib75; Lane 2, Fib75-abrin A chain; Lane 3, Fib75-ricin A chain: Lane 4. Fib75-gelonin; Lane 5, Fib75-momordin.
7521
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from
PHARMACOKlNETirS OF IMMUNOTOX1NS [N THE RAT
the presence of multiply substituted IT species (Fig. 2, Peak A),singly substituted IT (Fig. 2, Peak B), and unconjugated antibody (Fig. 2, Peak C) in proportions consistent with the analysisby SDS-PAGE and of minimal amounts of aggregated proteinor unconjugated RIP.
The Fib75 ITs were all toxic to the EJ human bladdercarcinoma cell line in tissue culture. The concentrations atwhich ['H]leucine incorporation by cells was reduced by 50%for the panel of ITs ranged between approximately 1 x 10 '"and 8 x 10"'°M.In direct comparisons of cytotoxicity in vitro,
such as in the representative experiment shown in Fig. 3, theITs differed consistently in potency. The order of potency was:Fib75-abrin A chain > Fib75-ricin A chain > Fib75-momordin> Fib75-gelonin. The difference in cytotoxicity between Fib75-abrin A chain and Fib75-ricin A chain was statistically significant: P < 0.02 by Student's / test. The unconjugated RIPs or
unconjugated Fib75 antibody exerted no significant cytotoxiceffects upon EJ cells at concentrations of 1 x 10~" M (not
shown).Pharmacokinetics of the Fib75 Immunotoxins in the Rat. The
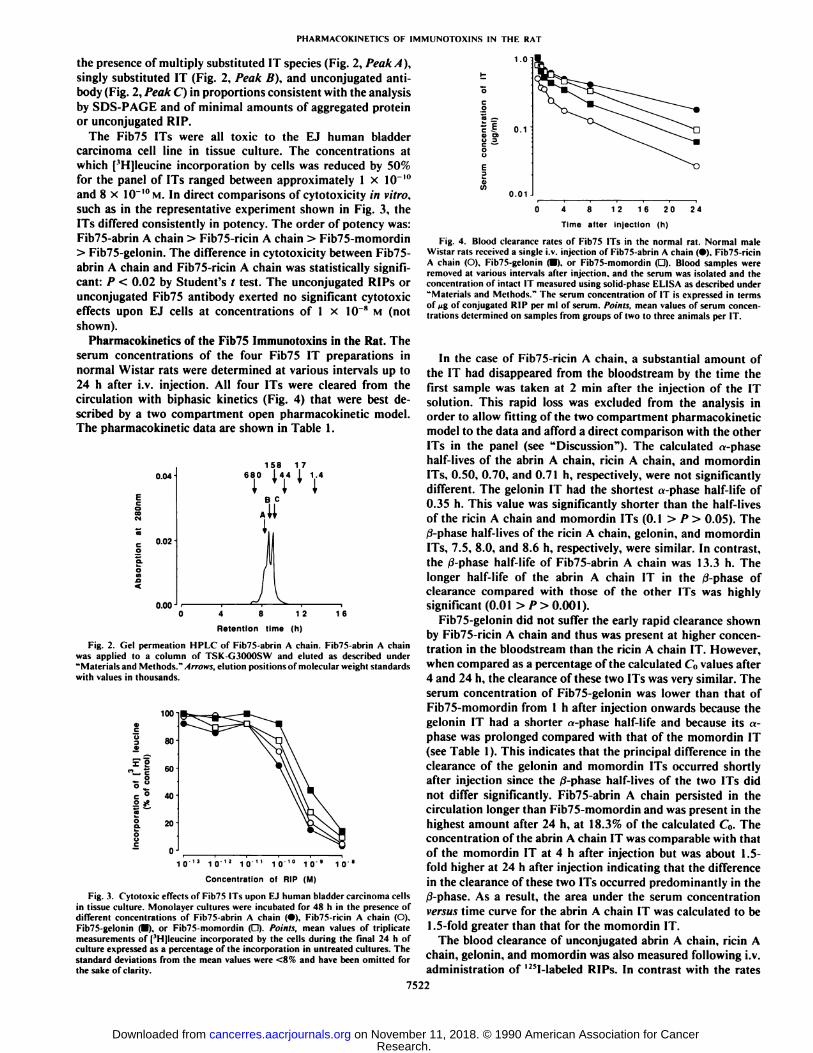
serum concentrations of the four Fib75 IT preparations innormal Wistar rats were determined at various intervals up to24 h after i.v. injection. All four ITs were cleared from thecirculation with biphasic kinetics (Fig. 4) that were best described by a two compartment open pharmacokinetic model.The pharmacokinetic data are shown in Table 1.
0.04
c 0.02
0.00
158 17680 l44 I 1.4
8 12 1 6
Retention time (h)
Fig. 2. Gel permeation HPLC of Fib75-abrin A chain. Fib75-abrin A chainwas applied to a column of TSK-G3000SW and cluted as described under"Materials and Methods. "Arrows, elution positions of molecular weight standards
with values in thousands.
100
' oà 60
IO'13 10'12 10" 101° 10
Concentration of RIP (M)
10"
Fig. 3. Cytotoxic effects of Fib75 ITs upon FJ human bladder carcinoma cellsin tissue culture. Monolayer cultures were incubated for 48 h in the presence ofdifferent concentrations of Fib75-abrin A chain (•).Fib75-ricin A chain (O).Fib75-gelonin (•),or Fib75-momordin (D). Points, mean values of triplicatemeasurements of [3H]leucine incorporated by the cells during the final 24 h ofculture expressed as a percentage of the incorporation in untreated cultures. Thestandard deviations from the mean values were <8% and have been omitted forthe sake of clarity.
0.1
0.01 J
8 12 16 20 24
Time after injection (h)
Fig. 4. Blood clearance rates of Fib75 ITs in the normal rat. Normal maleWistar rats received a single i.v. injection of Fib75-abrin A chain (•).Fib75-ricinA chain (O). Fib75-gelonin (•).or Fib75-momordin (D). Blood samples wereremoved at various intervals after injection, and the serum was isolated and theconcentration of intact IT measured using solid-phase ELISA as described under"Materials and Methods." The serum concentration of IT is expressed in terms
of tig of conjugated RIP per ml of scrum. Points, mean values of serum concentrations determined on samples from groups of two to three animals per IT.
In the case of Fib75-ricin A chain, a substantial amount ofthe IT had disappeared from the bloodstream by the time thefirst sample was taken at 2 min after the injection of the ITsolution. This rapid loss was excluded from the analysis inorder to allow fitting of the two compartment pharmacokineticmodel to the data and afford a direct comparison with the otherITs in the panel (see "Discussion"). The calculated «-phasehalf-lives of the abrin A chain, ricin A chain, and momordinITs, 0.50, 0.70, and 0.71 h, respectively, were not significantlydifferent. The gelonin IT had the shortest «-phasehalf-life of0.35 h. This value was significantly shorter than the half-livesof the ricin A chain and momordin ITs (0.1 > P > 0.05). The0-phase half-lives of the ricin A chain, gelonin, and momordinITs, 7.5, 8.0, and 8.6 h, respectively, were similar. In contrast,the /i-phase half-life of Fib75-abrin A chain was 13.3 h. Thelonger half-life of the abrin A chain IT in the tf-phase ofclearance compared with those of the other ITs was highlysignificant (0.01 > P> 0.001).
Fib75-gelonin did not suffer the early rapid clearance shownby Fib75-ricin A chain and thus was present at higher concentration in the bloodstream than the ricin A chain IT. However,when compared as a percentage of the calculated C(1values after4 and 24 h, the clearance of these two ITs was very similar. Theserum concentration of Fib75-gelonin was lower than that ofFib75-momordin from 1 h after injection onwards because thegelonin IT had a shorter «-phase half-life and because its «-phase was prolonged compared with that of the momordin IT(see Table 1). This indicates that the principal difference in theclearance of the gelonin and momordin ITs occurred shortlyafter injection since the /i-phase half-lives of the two ITs didnot differ significantly. Fib75-abrin A chain persisted in thecirculation longer than Fib75-momordin and was present in thehighest amount after 24 h, at 18.3% of the calculated C„.Theconcentration of the abrin A chain IT was comparable with thatof the momordin IT at 4 h after injection but was about 1.5-fold higher at 24 h after injection indicating that the differencein the clearance of these two ITs occurred predominantly in the0-phase. As a result, the area under the serum concentrationversus time curve for the abrin A chain IT was calculated to be1.5-fold greater than that for the momordin IT.
The blood clearance of unconjugated abrin A chain, ricin Achain, gelonin, and momordin was also measured following i.v.administration of '"I-labeled RIPs. In contrast with the rates
7522
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

PHARMACOKINETICS OF IMMUNOTOXINS IN THE RAT
Table 1 Pharmacokinetic data for the blood clearance of the Fib75 immunotoxins
ITFib75-abrin
AFib75-ricin AFib75-geloninFib75-momordinI»*'
(h)0.50
±0.080.70 ±0.160.35 ±0.080.71 ±0.191*0
(h)13.3
±0.57.5 ±0.48.0 ±0.48.6 ±0.2/ero-lime
intercepts (% ofd)A38.3
±3.250.6 ±6.055.8 ±8.121.0 ±2.7B63.6
±1.749.0 ±3.546.4 ±2.880.3 ±2.0Blood
level (%ofCO4h51.8
35.633.464.324
h18.3
5.55.912.4AUC
kg/ml xh)12.1
2.95.77.9
* tv,a and /ufi, a- and tf-phase half-lives in blood; A and B, concentration constants in the biexponential equation
C = Be~"
for the a- and ff-phases of clearance, respectively: Al'C. area under the serum concentration versus time curve.
of clearance of the ITs, all the RIPs cleared very rapidly fromthe bloodstream with an initial half-life of less than 5 min (notshown).
Breakdown of Fib75 and Control Immunotoxins in Vitro. Thesusceptibility of the panel of Fib75 ITs to breakdown in thepresence of reducing agent was assessed by incubating a solutionof each IT in the presence of 5 mivi glutathione at 37°Cin vitro.
The amount of intact IT remaining after different times ofexposure to the glutathione was measured using solid-phaseELISA. In each case, coincubation with glutathione led to aprogressive decline in the amount of intact IT (Fig. 5) whereasthere was no significant loss of IT as measured by the ELISAwhen glutathione was omitted from the incubation (not shown).The breakdown of the ITs in the presence of the large (approximately 500,000-fold) molar excess of glutathione used in theexperiment would have been predicted to follow pseudo-firstorder reaction kinetics giving a linear relationship between thelogarithm of the IT concentration and time. The biphasickinetics actually observed indicated that the IT preparationsmight be heterogeneous with respect to the rate of cleavage ofthe disulfide bond. In the more rapid phase of breakdown, thedecrease in the amount of intact IT was most pronounced inthe case of Fib75-gelonin, greater than 50% loss occurringwithin 1 h of incubation, compared with the loss of only about20% of Fib75-abrin A chain (Fig. 5). Fib75-ricin A chain andFib75-momordin broke down in similar amounts intermediatebetween those of the abrin A chain and gelonin ITs. In theslower phase of breakdown, the Fib75 ITs made with ricin Achain, gelonin, and momordin all broke down with similar rateconstants of between 0.14 and 0.17 h"1 whereas Fib75-abrin A
100 n
10
02468
Time of incubation (h)
Fig. 5. Instability of Fib75 ITs in the presence of glutathione. Solutions ofFib75-abrin A chain (•).Fib75-ricin A chain (O), Fib75-gelonin (•).and Fib75-momordin (O) were each incubated in the presence of 5 mM glutathione at 37°C
in vitro. The concentration of intact IT was determined after various times ofincubation using solid-phase ELISA as described under "Materials and Methods."
Points, mean values of triplicate determinations expressed as a percentage of theamount of intact IT at the start of the incubation; bars, SD from the mean valuesunless smaller than the points as plotted.
chain broke down with a 3-4-fold lower rate constant of 0.043h~'. The amount of Fib75-abrin A chain remaining was signif
icantly greater than that of the other ITs at all time points andmore than 50% of the starting amount of the abrin A chain ITwas left intact after 8 h of incubation. A similar difference inthe rates of breakdown of the abrin A chain and ricin A chainITs could be detected in the presence of glutathione concentrations as low as 0.2 mM (not shown).
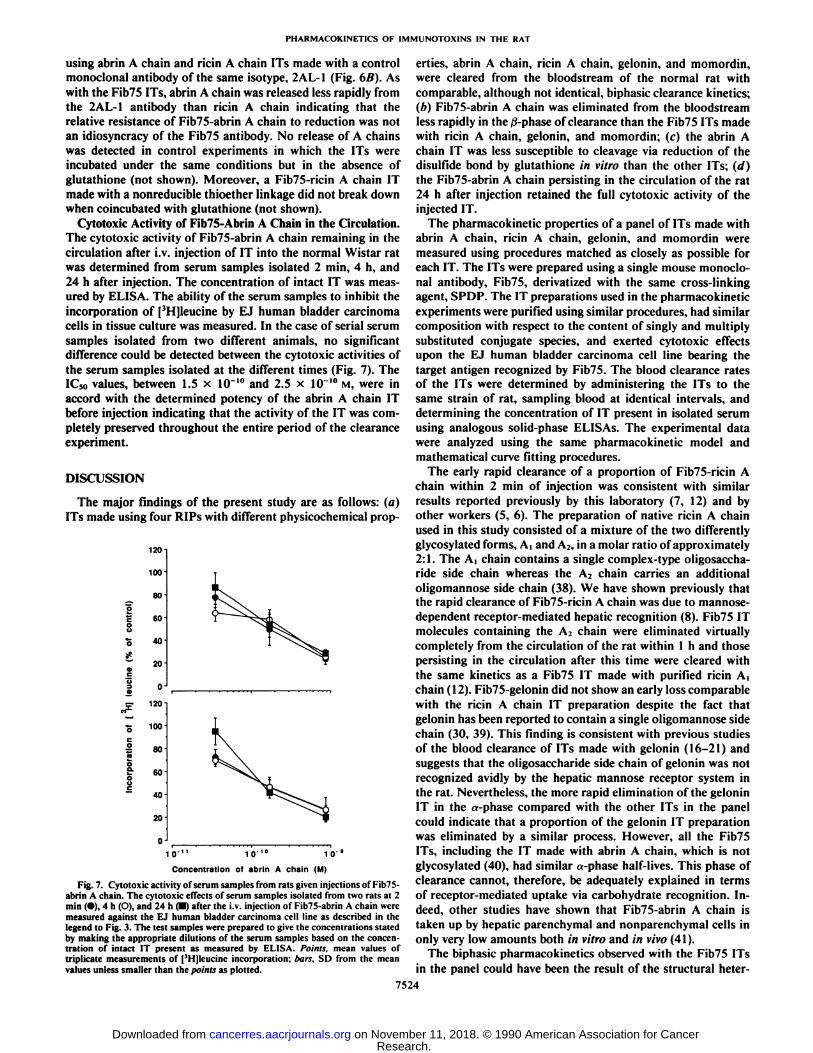
To confirm that the decrease in the amount of intact ITmeasured by the ELISA was, in fact, due to the cleavage of thedisulfide bond, the release of the A chains from Fib75-abrin Achain and Fib75-ricin A chain upon treatment with glutathionewas measured directly. Gel permeation HPLC was used tomeasure the absolute amount of A chain released upon incubation of the ITs in the presence of different concentrations ofglutathione for 1 h at 37°Cin vitro (Fig. 6A). The amount of A
chain released by glutathione was about 2- to 3-fold lower inthe case of Fib75-abrin A chain consistent with the greaterstability of the abrin A chain IT measured by ELISA. Therelease of A chain by glutathione was also measured directly
E
40
0
100
80
40
0 J
.1 1 10
Concentration of glutathione (mM)
Fig. 6. Release of A chain from Fib75 and 2AL-1 ITs made with abrin Achain and ricin A chain by glutathione. ITs made with abrin A chain (•)or ricinA chain (O) linked to the antibodies Fib75 (A) or 2AL-1 (A) were incubated inthe presence of different concentrations of glutathione for I h at 37"C in vitro.
The amount of A chain released was measured using gel permeation HPLC asdescribed under "Materials and Methods." Points are expressed as the percentage
of A chain released relative to the total A chain content.
7523
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from
PHARMACOKINETICS OF 1MMUNOTOXINS IN THE RAT
using abrin A chain and ricin A chain ITs made with a controlmonoclonal antibody of the same isotype, 2AL-1 (Fig. 6B). Aswith the Fib75 ITs, abrin A chain was released less rapidly fromthe 2AL-1 antibody than ricin A chain indicating that therelative resistance of Fib75-abrin A chain to reduction was notan idiosyncracy of the Fib75 antibody. No release of A chainswas detected in control experiments in which the ITs wereincubated under the same conditions but in the absence ofglutathione (not shown). Moreover, a Fib75-ricin A chain ITmade with a nonreducible thioether linkage did not break downwhen coincubated with glutathione (not shown).
Cytotoxic Activity of Fib75-Abrin A Chain in the Circulation.The cytotoxic activity of Fib75-abrin A chain remaining in thecirculation after i.v. injection of IT into the normal Wistar ratwas determined from serum samples isolated 2 min, 4 h, and24 h after injection. The concentration of intact IT was measured by ELISA. The ability of the serum samples to inhibit theincorporation of [3H]leucine by EJ human bladder carcinoma
cells in tissue culture was measured. In the case of serial serumsamples isolated from two different animals, no significantdifference could be detected between the cytotoxic activities ofthe serum samples isolated at the different times (Fig. 7). TheIC50 values, between 1.5 x 10"'°and 2.5 x 10"I0 M, were in
accord with the determined potency of the abrin A chain ITbefore injection indicating that the activity of the IT was completely preserved throughout the entire period of the clearanceexperiment.
DISCUSSION
The major findings of the present study are as follows: (a)ITs made using four RIPs with different physicochemical prop-
120
100
80
60
40
20
0
120
100
80
60
40
20
10" 10'
Concentration of abrin A chain (Mi
Fig. 7. Cytotoxic activity of serum samples from rats given injections of Fib75-abrin A chain. The cytotoxic effects of serum samples isolated from two rats at 2min (•),4 h (O), and 24 h (•)after the i.v. injection of Fib75-abrin A chain weremeasured against the EJ human bladder carcinoma cell line as described in thelegend to Fig. 3. The test samples were prepared to give the concentrations statedby making the appropriate dilutions of the serum samples based on the concentration of intact IT present as measured by ELISA. Points, mean values oftriplicate measurements of [3Hjleucine incorporation: bars, SD from the mean
values unless smaller than the points as plotted.
erties, abrin A chain, ricin A chain, gelonin, and momordin,were cleared from the bloodstream of the normal rat withcomparable, although not identical, biphasic clearance kinetics;(b) Fib75-abrin A chain was eliminated from the bloodstreamless rapidly in the ß-phaseof clearance than the Fib75 ITs madewith ricin A chain, gelonin, and momordin; (c) the abrin Achain IT was less susceptible to cleavage via reduction of the(lisiillide bond by glutathione in vitro than the other ITs; (d)the Fib75-abrin A chain persisting in the circulation of the rat24 h after injection retained the full cytotoxic activity of theinjected IT.
The pharmacokinetic properties of a panel of ITs made withabrin A chain, ricin A chain, gelonin, and momordin weremeasured using procedures matched as closely as possible foreach IT. The ITs were prepared using a single mouse monoclonal antibody, Fib75, derivatized with the same cross-linkingagent, SPDP. The IT preparations used in the pharmacokineticexperiments were purified using similar procedures, had similarcomposition with respect to the content of singly and multiplysubstituted conjugate species, and exerted cytotoxic effectsupon the EJ human bladder carcinoma cell line bearing thetarget antigen recognized by Fib75. The blood clearance ratesof the ITs were determined by administering the ITs to thesame strain of rat, sampling blood at identical intervals, anddetermining the concentration of IT present in isolated serumusing analogous solid-phase ELISAs. The experimental datawere analyzed using the same pharmacokinetic model andmathematical curve fitting procedures.
The early rapid clearance of a proportion of Fib75-ricin Achain within 2 min of injection was consistent with similarresults reported previously by this laboratory (7, 12) and byother workers (5, 6). The preparation of native ricin A chainused in this study consisted of a mixture of the two differentlyglycosylated forms. A, and A2, in a molar ratio of approximately2:1. The A, chain contains a single complex-type oligosaccha-ride side chain whereas the A2 chain carries an additionaloligomannose side chain (38). We have shown previously thatthe rapid clearance of Fib75-ricin A chain was due to mannose-dependent receptor-mediated hepatic recognition (8). Fib75 ITmolecules containing the A2 chain were eliminated virtuallycompletely from the circulation of the rat within l h and thosepersisting in the circulation after this time were cleared withthe same kinetics as a Fib75 IT made with purified ricin A,chain ( 12). Fib75-gelonin did not show an early loss comparablewith the ricin A chain IT preparation despite the fact thatgelonin has been reported to contain a single oligomannose sidechain (30, 39). This finding is consistent with previous studiesof the blood clearance of ITs made with gelonin (16-21) andsuggests that the oligosaccharide side chain of gelonin was notrecognized avidly by the hepatic mannose receptor system inthe rat. Nevertheless, the more rapid elimination of the geloninIT in the «-phasecompared with the other ITs in the panelcould indicate that a proportion of the gelonin IT preparationwas eliminated by a similar process. However, all the Fib75ITs, including the IT made with abrin A chain, which is notglycosylated (40), had similar «-phasehalf-lives. This phase ofclearance cannot, therefore, be adequately explained in termsof receptor-mediated uptake via carbohydrate recognition. Indeed, other studies have shown that Fib75-abrin A chain istaken up by hepatic parenchyma! and nonparenchymal cells inonly very low amounts both in vitro and in vivo (41).
The biphasic pharmacokinetics observed with the Fib75 ITsin the panel could have been the result of the structural heter-
7524
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

PHARMACOKINETICS OF IMMUNOTOXINS IN THE RAT
ogeneity inherent in IT preparations synthesized by chemicalprocedures. First, the experiments measuring the rate of breakdown of the Fib75 ITs in vitro indicated the presence of ITmolecules with different susceptibilities to reduction by gluta-thione. Thus, the a-phase of clearance could reflect the preferential elimination of the fraction of IT molecules in which thedisulfide bond is most readily cleaved. However, in a differentstudy, Fib75-ricin A chain ITs made with a disulfide linkage orwith a nonreducible thioether linkage had comparable «-phasehalf-lives (33). Second, IT molecules within a single preparationdiffer with regard to the position of attachment of the RIPmolecule to the antibody and may differ with respect to therelative configuration of the two components even when linkedvia the same position. The more rapid clearance of a proportionof IT molecules could have been due to an impairment to theability of the antibody component to persist in the circulation.This might occur either by enhancement of interactions betweenthe conjugated antibody and normal tissues mediated via natural mechanisms such as Fc receptor recognition or by novelinteractions made possible by structural changes to the antibodyas a result of conjugation.
The clearance rates of Fib75-ricin A chain, Fib75-gelonin,and Fib75-momordin were very similar in the /3-phase of clearance suggesting that the different modes of construction necessarily used in the synthesis of ITs with toxin A chains on theone hand, compared with single-chain RIPs on the other, didnot have a major influence upon the /3-phase half-life. In contrast, the j3-phase half-life of Fib75-abrin A chain was significantly longer than that of the other ITs in the panel. A possibleexplanation for the shorter 0-phase half-life of ITs made withricin A chain, gelonin, and momordin is that these RIPs mayall contain a complex-type oligosaccharide side chain (38, 39)and that the presence of this structural feature somehow enhances their clearance. One way to test this hypothesis wouldbe to determine the effect of removing the oligosaccharide sidechain from the RIPs upon the rate of clearance of IT. However,it has been reported that the complex-type oligosaccharide sidechain of ricin A chain cannot be cleaved enzymically withoutloss of ribosome-inactivating activity (38). Similarly, we wereunable to remove the oligosaccharide side-chains from geloninand momordin without first denaturing the RIPs.5 We have
recently found that a Fib75 IT made with an aglycosyl recombinant ricin A chain had a .; phase half-life similar to that ofFib75-ricin Ai-chain suggesting that the presence of the complex-type oligosaccharide side chain had no influence in thisphase of clearance.6
The rate at which the disulfide bond linking the antibody andRIP can be cleaved by reduction has been shown previously toinfluence IT half-life in the /3-phase. Thus, ITs made with cross-linking agents such as SPDB (33) and 4-succinimidyloxycar-bonyl-«-methyl-«-(2-pyridyldithio)toluene (42), which containhindered disulfide bonds that are less susceptible to chemicalreduction, have significantly longer ß-phasehalf-lives than thecorresponding ITs made with nonhindered linkages. In thisstudy, we found that Fib75-abrin A chain was cleaved lessrapidly (by 3-4-fold) by glutathione in vitro than the Fib75 ITsmade with ricin A chain, gelonin, or momordin. Direct evidencefor the slower cleavage of the abrin A chain IT was obtained by
' Unpublished results.6 E. J. Wawrzynczak, A. J. Cumber, R. V. Henry, and G. D. Parncll. Compar
ative biochemical, cytotoxic and pharmacokinetic properties of immunotoxinsmade with native ricin A chain, ricin A, chain and recombinant ricin A chain,submitted for publication.
measuring the actual release of A chain by glutathione using aquantitative HPLC method. Since glutathione is the most abundant reducing molecule in the circulation (43), this findingstrongly suggests that the longer half-life of Fib75-abrin A chainin the 0-phase of clearance compared with the other Fib75 ITsmay be due, either in part or in whole, to a slower rate ofcleavage of the disulfide bond in vivo. In our experiments, ahigh concentration of glutathione was used to give readilymeasurable rates of breakdown for comparison. The concentration of glutathione in peripheral blood is relatively low but it ispresent at higher levels in the liver, the site of glutathionesynthesis (43). It is therefore possible that the major site of ITbreakdown by reduction is within the hepatic circulation.
The slower rate of breakdown of the IT made with abrin Achain in vitro could be associated with the physicochemicalproperties of this RIP. Abrin A chain has a considerably moreacidic isoelectric point, pi 4.6 (40), than ricin A chain, gelonin,and momordin, pis 7.5, 8.15, and 8.6, respectively (39, 40). Itis possible that a localized electrostatic interaction betweenabrin A chain and the Fib75 antibody could induce the hybridmolecule to adopt a more compact conformation in which thedisulfide bond is partially shielded from attack by reducingagents. A similar explanation was suggested previously for theapparently slower breakdown of an IT made with saporin (44)although subsequent pharmacokinetic analyses claimed thatthere was no difference in the rate of splitting compared withan analogous ricin A chain IT (24). An alternative explanationis that the proximity of negatively charged side chains of aminoacid residues of abrin A chain in the vicinity of the disulfidebond causes electrostatic repulsion of glutathione which isnegatively charged at physiological pH. Such a mechanism ofprotection would be a unique feature of abrin A chain since allthe other RIPs that have been used for IT construction carry anet positive charge at physiological pH. The relative resistanceto reduction of the abrin A chain IT made with Fib75 was nota peculiarity of the Fib75 antibody because an abrin A chain ITmade with another mouse monoclonal antibody of the sameisotype (IgG2a) was also cleaved less readily in vitro than thecorresponding ricin A chain IT. It remains to be determinedwhether this phenomenon is restricted to antibodies of theIgG2a isotype.
The ability of an IT made with abrin A chain to persist longerin the circulation than ITs made with ricin A chain, gelonin,and momordin would be of no advantage were IT moleculesthat are reduced less readily in vitro to possess a diminishedcytotoxic potency. The analysis of samples obtained from ratsup to 24 h after injection of Fib75-abrin A chain clearly showedthat the IT present in the circulation remained fully active. Thecytotoxic activity of ITs during the course of blood clearanceexperiments has rarely been reported. In one study, it was foundthat the rate of disappearance of a ricin A chain IT was mirroredexactly by the rate of loss of cytotoxic activity (9). In contrast,clearance studies of a gelonin IT revealed that the cytotoxicactivity of the IT remaining in the bloodstream was substantially diminished (20).
In conclusion, we have demonstrated that an IT made withabrin A chain had the highest cytotoxic potency and the longestserum half-life of a panel of ITs made with different RIPs andthat the IT retained full cytotoxic activity in the circulation.Our results suggest that the slower clearance of the abrin Achain IT may have been due to its greater resistance to reductionby glutathione. These findings indicate that abrin A chain canform ITs with greater stability than ITs made with other RIPs
7525
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

PHARMACOKINETICS OF IMMUNOTOXINS IN THE RAT
and merits further investigation as a component in the assemblyof ITs which are designed to have pharmacokinetic propertiesoptimal for therapy.
REFERENCES
1. Blakey, D. C., Wawrzynczak, E. J., Wallace, P. M., and Thorpe. P. E.Antibody-toxin conjugates: a perspective. In: H. Waldmann (ed.). Monoclonal Antibody Therapy, pp. 50-90. Basel: S. Karger AG. 1988.
2. FitzGerald, D. J., and Pastan. 1. Targeted toxin therapy for the treatment ofcancer. J. Nati. Cancer Inst.. 81: 1455-1463. 1989.
3. Frankel, A. E. (ed.). Immunotoxins. Norwell, MA: Kluwer Academic Publishers, 1988.
4. Vitella, E. S., Fulton. R. J., May, R. D.. Till. M., and Uhr. J. W. Redesigningnature's poisons to create anti-tumor reagents. Science (Washington DC),238: 1098-1104, 1987.
5. Jansen, F. K.. Blythman, H. E., Bourrie, B., Carriere, D., Casellas, P.,Dussossoy, D., Gros. O.. Laurent, J. C.. Liance, M. C., Poncelet. P., Richer,G., and Vidal, H. Significance of the kinetics of immunotoxin cytotoxicity.In: C. Gregoriadis, G. Poste, J. Senior, and A. Trouet (eds.). Receptor-mediated Targeting of Drugs, pp. 147-178. New York: Plenum PublishingCorp., 1984.
6. Raso, V., and Basala, M. Monoclonal antibodies as cell targeted carriers ofcovalently and non-covalently attached toxins. In: G. Gregoriadis, G. Poste.J. Senior, and A. Trouet (eds.). Receptor-mediated Targeting of Drugs, pp.119-138. New York: Plenum Publishing Corp., 1984.
7. Worrell, N. R., Cumber, A. J., Parnell, G. D., Ross, W. C. J., and Forrester.J. A. Fate of an antibody-ricin A chain conjugate administered to normalrats. Biochem. Pharmacol., 35:417-423. 1986.
8. Worrell. N. R., Skilleter, D. N., Cumber, A. J., and Price, R. J. Mannosereceptor dependent uptake of a ricin A chain-antibody conjugate by rat livernon-parenchymal cells. Biochem. Biophys. Res. Commun., 137: 892-896,1986.
9. Bourrie. B. J. P., Casellas, P., Blythman, H. E., and Jansen. F. K. Study ofthe plasma clearance of antibody-ricin A chain immunotoxins. Evidence forspecific recognition sites on the A chain that mediate rapid clearance of theimmunotoxin. Eur. J. Biochem.. 155: 1-10. 1986.
10. Blakey, D. C, Watson, G. J., Knowles, P. P., and Thorpe, P. E. Effect ofchemical deglycosylation of ricin A chain on the in vivo fate and cytotoxicactivity of an immunotoxin composed of ricin A chain and anti-Thyl.lantibody. Cancer Res., 47:947-952, 1987.
11. Byers, V. S., Pimm, M. V., Pawluczyk, I. Z. A., Lee, H. M., Scannen, P. J.,and Baldwin, R. W. Biodistribution of ricin toxin A chain-monoclonalantibody 79IT/36 immunotoxin and influence of hepatic blocking agents.Cancer Res., 47: 5277-5283, 1987.
12. Cumber, A. J., Parnell, G. D., Henry, R. V., Forrester, J. A., and Wawrzynczak. E. J. Mannose residues mediate the rapid clearance in vivo of a ricin Achain immunotoxin. Biochem. Soc. Trans., 17: 137-138, 1989.
13. Stirpe. F., and Barbieri. L. Ribosome-inactivating proteins up to date. FEBSLett., 195: 1-8, 1986.
14. Thorpe. P. E., Blakey, D. C., Brown. A. N. F., Knowles, P. P.. Knyba, R. E.,Wallace, P. M., Watson, G. J.. and Wawrzynczak, E. J. Comparison of twoanti-Thyl.l-abrin A chain immunotoxins prepared with different cross-linking agents: antitumor effects, in vivo fate, and tumor cell mutants. J.Nail. Cancer Inst.. 79: 1101-1111, 1987.
15. K.iiii.iktisliii.nl. S., and Houston, L. L. Immunological and biological stabilityof immunotoxins in vivo as studied by the clearance of disulfide-linkedpokeweed antiviral protein-antibody conjugates from blood. Cancer Res., 45:2031-2036, 1985.
16. Sivam, G., Pearson, J. W., Bohn, W., Oldham. R. K., Sadoff, J. C., andMorgan, A. C., Jr. Immunotoxins to a human melanoma-associated antigen:comparison of gelonin with ricin and other A chain conjugates. Cancer Res.,Â¥7:3169-3173, 1987.
17. Letvin, N. L., Goldmacher, V. S., Ritz, J., Yelz, J. M.. Schlossman, S. F.,and Lambert. J. M. In vivoadministration of lymphocyte-specific monoclonalantibodies in nonhuman primates. In vivo stability of disulfide-linked immunotoxins. J. Clin. Invest., 77: 977-984, 1986.
18. Letvin, N. L., Chalifoux, L. V., Reimann, K. A., Ritz, J.. Schlossman. S. F.,and Lambert. J. M. In vivoadministration of lymphocyte-specific monoclonalantibodies in nonhuman primates. Delivery of ribosome-inactivating proteinsto spleen and lymph node T cells. J. Clin. Invest.. 77: 666-673. 1986.
19. Scott. C. F., Jr., Lambert, J. M., Goldmacher, V. S.. Blaltler. W. A., Sobel.R.. Schlossman. S. F., and Benacerraf, B. The pharmacokinetics and toxicityof murine monoclonal antibodies and of gelonin conjugates of these antibodies. Int. J. Immunopharmacol., 9: 211-225, 1987.
20. Scott, C. F., Jr., Goldmacher, V. S., Lambert. J. M., Jackson, J. V., andMclntyre, G. D. An immunotoxin composed of monoclonal antitransferrinreceptor antibody linked by a disulfide bond to the ribosome-inactivating
protein gelonin: potent in vitro and in vivo effects against human tumors. J.Nati. Cancer Inst., 79: 1163-1172, 1987.
21. Scott. C. F., Jr., Goldmacher, V. S., Lambert, J. M., Chari, R. V. J., Bolender,S., Gauthier, M. N., and Blatller. W. A. The antileukemic efficacy of animmunotoxin composed of a monoclonal anti-Thy-1 antibody disulfide linkedto the ribosomc-inactivating protein gelonin. Cancer Immunol. Immunother..25:31-40, 1987.
22. Stirpe, F., Derenzini. M., Barbieri, L.. Farabegoli, F.. Brown, A. N. F.,Knowles, P. P., and Thorpe, P. E. Hepatotoxicity of immunotoxins madewith saporin, a ribosome-inactivating protein from Saponaria officinalis.Virchows Arch. B Cell Pathol., 53: 259-271. 1987.
23. Glennie, M. J., McBride, H. M., Stirpe, F., Thorpe. P. E.. Worth, A. T., andStevenson, G. T. Emergence of immunoglobulin variants following treatmentof a B cell leukemia with an immunotoxin composed of antiidiotypic antibodyand saporin. J. Exp. Med., 766: 43-62, 1987.
24. Blakey, D. C.. Skilleler, D. N., Price, R. J., Watson, G. J., Hart. L. I..Newell. D. R.. and Thorpe, P. E. Comparison of the pharmacokinetics andhepatotoxic effects of saporin and ricin A chain immunotoxins on murineliver parenchyma! cells. Cancer Res., 48: 7072-7078, 1988.
25. Siena. S., Lappi, D. A., Brcgni, M., Formosa, A., Villa, S., Soria, M.,Bonnadonna, G., and Gianni. A. M. Synthesis and characterization of anantihuman T-lymphocyte specific saporin immunotoxin (OKTl-sap) with invivo stability into nonhuman primates. Blood. 72: 756-765. 1988.
26. Casellas, P., Dussossoy, D., Falasca. A. !.. Barbier!. L., Guillemot, J. C.,Ferrara, P., Bolognesi, A., Cenini, P., and Stirpe. F. Trichokirin, a ribosome-inactivating protein from the seeds of Trichosanthes kirilowii Maximowicz.Purification, partial characterisation and use for preparation of immunolox-ins. Eur. J. Biochem., 176: 581-588, 1988.
27. Nicolson, G. L., and Blaustem, J. The interaction of Ricinus communisagglutinin with normal and tumor cell surfaces. Biuchim. Biophys. Acta, 266:543-547, 1972.
28. Cumber, A. J., Forrester, J. A., Foxwell, B. M. J., Ross, W. C. J., andThorpe, P. E. Preparation of antibody-toxin conjugates. Methods Enzymol.,112: 207-225, 1985.
29. Forrester. J. A., Mclnlosh. D. P., Cumber. A. J., Parnell, G. D.. and Ross,W. C. J. Delivery of ricin and abrin A chains to human carcinoma cellsfollowing covalent linkage to monoclonal antibody LICR-LOND-Fib75.Cancer Drug Deliv., /: 283-292, 1984.
30. Stirpe, F.. Olsnes, S., and Pihl, A. Gelonin, a new inhibitor of proteinsynthesis, nontoxic to intact cells. J. Biol. Chem., 255: 6947-6953, 1980.
31. Barbieri, L.-Zamboni, M., Montanaro, L..Sperti, S., and Stirpe. F. Inhibitionof protein synthesis in vitro by proteins from the seeds of Momordicacharantia (bitter pear melon). Biochem. J.. 186: 443-452, 1980.
32. Laemmli, U. K. Cleavage of structural proteins during the assembly of thehead of bactcriophage T4. Nature (Lond.). 227: 680-685. 1970.
33. Worrell, N. R., Cumber, A. J., Parnell, G. D., Mirza, A., Forrester, J. A.,and Ross, W. C. J. Effect of linkage variation on pharmacokinctics of ricinA chain-antibody conjugates in normal rats. Anti-Cancer Drug Design, /:179-188, 1986.
34. I Mm.in G. L. Tissue sulfhydryl groups. Arch. Biochem. Biophys.. 82: 70-77. 1959.
35. Fraker, P. J., and Speck, J. C. Protein and cell membrane iodination with asparingly soluble chloroamide. Biochem. Biophys. Res. Commun., 80: 849-857, 1978.
36. Jennrich, R. I., and Sampson, P. F. Application of stepwise regression tonon-linear least squares estimation. Technometrics. 10: 63-72, 1968.
37. Ottaway, J. H. Normalization in the fitting of data by iterative methods.Biochem. J., 134: 729-732, 1973.
38. Foxwell, B. M. J., Donovan, T. A., Thorpe, P. E., and Wilson, G. Theremoval of carbohydrates from ricin with endoglycosidases H, F and D and«-mannosidase. Biochim. Biophys. Acta, 840: 193-203, 1985.
39. Falasca. A., Gaspcri-Campani, A., Abbondanza, A., Barbieri, L., and Stirpe,F. Properties of the ribosome-inactivating proteins gelonin, Momordicacharantia inhibitor, and dianthins. Biochem. J., 207: 505-509, 1982.
40. Olsnes, S., Refsnes, K., Christensen, T. B., and Pihl, A. Studies on thestructure and properties of the lectins from Abrus precatorius and Ricinuscommunis. Biochim. Biophys. Acta, 405: 1-10, 1975.
41. Skilleter, D. N., Price, R. J.. Parnell, G. D., and Cumber, A. J. The lowuptake of an abrin A chain immunotoxin by rat hepatic cells in vivo and invitro. Cancer Lett., 46: 161-166, 1989.
42. Thorpe, P. E., Wallace, P. M., Knowles, P. P.. Reif, M. G.. Brown, A. N.F., Watson, G. J.. Knyba, R. E.. Wawrzynczak, E. J., and Blakey, D. C. Newcoupling agents for the synthesis of immunotoxins containing a hindereddisulfide bond with improved stability in vivo. Cancer Res., 47: 5924-5931,1987.
43. Meister, A., and Anderson, M. E. Glutathione. Annu. Rev. Biochem.. 52:711-760, 1983.
44. Thorpe, P. E.. Brown, A. N. F., Bremner, J. A. G., Foxwell, B. M. J., andStirpe, F. An immunotoxin composed of monoclonal anti-Thyl.l antibodyand a ribosome-inactivating protein from Saponaria officinalis. Potent anti-tumor effects in vitro and in vivo. J. Nati. Cancer Inst., 75: 151-159, 1985.
7526
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

1990;50:7519-7526. Cancer Res Edward J. Wawrzynczak, Alan J. Cumber, Raymond V. Henry, et al. with Abrin A Chain, Ricin A Chain, Gelonin, and MomordinPharmacokinetics in the Rat of a Panel of Immunotoxins Made
Updated version
http://cancerres.aacrjournals.org/content/50/23/7519
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/50/23/7519To request permission to re-use all or part of this article, use this link
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from