potential role of growth hormone in impairment of insulin signaling in skeletal muscle, adipose...
TRANSCRIPT

Potential Role of Growth Hormone in Impairment ofInsulin Signaling in Skeletal Muscle, Adipose Tissue,and Liver of Rats Chronically Treated with Arginine
Thais de Castro Barbosa, Jose Edgar Nicoletti de Carvalho, Leonice Lourenco Poyares,Silvana Bordin, Ubiratan Fabres Machado, and Maria Tereza Nunes
Department of Physiology and Biophysics, Institute of Biomedical Sciences, University of Sao Paulo, 05508-900 SaoPaulo, Sao Paulo, Brazil
We have shown that rats chronically treated with Arginine (Arg), although normoglycemic, exhibithyperinsulinemia and decreased blood glucose disappearance rate after an insulin challenge.Attempting to investigate the processes underlying these alterations, male Wistar rats weretreated with Arg (35 mg/d), in drinking water, for 4 wk. Rats were then acutely stimulated withinsulin, and the soleus and extensorum digitalis longus muscles, white adipose tissue (WAT), andliver were excised for total and/or phosphorylated insulin receptor (IR), IR substrate 1/2, Akt, Januskinase 2, signal transducer and activator of transcription (STAT) 1/3/5, and p85�/55� determination.Muscles and WAT were also used for plasma membrane (PM) and microsome evaluation of glucosetransporter (GLUT) 4 content. Pituitary GH mRNA, GH, and liver IGF-I mRNA expression wereestimated. It was shown that Arg treatment: 1) did not affect phosphotyrosine-IR, whereas itdecreased phosphotyrosine-IR substrate 1/2 and phosphoserine-Akt content in all tissues studied,indicating that insulin signaling is impaired at post-receptor level; 2) decreased PM GLUT4 contentin both muscles and WAT; 3) increased the pituitary GH mRNA, GH, and liver IGF-I mRNA expression,the levels of phosphotyrosine-STAT5 in both muscles, phosphotyrosine-Janus kinase 2 in extenso-rum digitalis longus, phosphotyrosine-STAT3 in liver, and WAT as well as total p85� in soleus,indicating that GH signaling is enhanced in these tissues; and 4) increased p55� total content inmuscles, WAT, and liver. The present findings provide the molecular mechanisms by which insulinresistance and, by extension, reduced GLUT4 content in PM of muscles and WAT take place afterchronic administration of Arg, and further suggest a putative role for GH in its genesis, consideringits diabetogenic effect. (Endocrinology 150: 2080–2086, 2009)
There is a growing body of evidence showing that dietaryconstituents can modulate events that govern gene expres-
sion at transcriptional and/or posttranscriptional level, and,therefore, affecting specific aspects of cellular function. Aminoacids are included in this category because they play crucial rolesin metabolism, development, growth, and reproduction of all lifeforms. Among them, L-Arginine (Arg) has been described as oneof the most important, considering its physiological and nutri-tional properties, as extensively reported (1).
Arg is a natural precursor of nitric oxide, which, by acti-vating the cyclic-GMP pathway, can modify the local bloodflux and/or activate kinases cascades, which might in turn lead
to physiological repercussions in the tissues where it is gen-erated (2). In fact, Arg was shown to revert the endothelialdysfunction commonly associated with the main cardiovas-cular risk factors, like diabetes, hypertension, hypercholes-terolemia, and obesity (3, 4).
Apart from these beneficial aspects, Arg supplementationmight be faced in the light of important limitations. We havepreviously demonstrated that acute (60 min, iv) and chronic (30d) oral administration of Arg increases GH mRNA expression inrats, indicating that GH synthesis is being stimulated (5, 6).However, although normoglycemic, rats treated chronically (30d) with Arg presented hyperinsulinemia and reduced blood glu-
ISSN Print 0013-7227 ISSN Online 1945-7170Printed in U.S.A.Copyright © 2009 by The Endocrine Societydoi: 10.1210/en.2008-1487 Received October 22, 2008. Accepted December 17, 2008.First Published Online December 23, 2008
Abbreviations: AU, Arbitrary unit; A35, 35 mg arginine/d/animal; Arg, arginine; EDL, ex-tensorum digitalis longus; eIF, eukaryotic translation initiation factor; GLUT, glucose trans-porter; IR, insulin receptor; IRS, insulin receptor substrate; JAK, Janus kinase; M, micro-some; PI3-K, phosphatidylinositol 3-kinase; PK, protein kinase; PM, plasma membrane;STAT, signal transducer and activator of transcription; WAT, white adipose tissue.
D I A B E T E S - I N S U L I N - G L U C A G O N - G A S T R O I N T E S T I N A L
2080 endo.endojournals.org Endocrinology, May 2009, 150(5):2080–2086
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 14 July 2014. at 15:20 For personal use only. No other uses without permission. . All rights reserved.

cose decay rate after an insulin challenge, which indicates theoccurrence of insulin resistance (6).
It is well known that Arg presents a GH secretagogue activitythat involves, primarily, an inhibitory effect upon somatostatinrelease (2, 7). Besides, Arg induced somatotropes depolarization(8), which might also contribute to its GH-releasing activity, aneffect that has already been observed in pancreatic �-cells (9).However, it was reported that rats chronically treated with GH,transgenic mice overexpressing GH, and acromegalic patientsdevelop insulin resistance (10–13), which is characterized by areduced translocation of glucose transporter (GLUT) 4 to themuscle and adipose cell surface, where this protein is the majorGLUT. Thus, one hypothesis relating Arg and insulin resistancecould involve enhanced GH activity.
GLUT4 translocation requires activation of phosphatidylino-sitol 3-kinase (PI3-K) (14), which occurs upon binding to ty-rosine-phosphorylated insulin receptor substrates (IRSs). There-fore, PI3-K leads to activation of the serine/threonine kinase Akt/protein kinase (PK) B and atypical PK C (�PKC), that have beenimplicated in GLUT4 translocation in adipose and muscle cells,and, therefore, to glucose uptake, as well as in the control ofglucose output in liver (15).
The present study attempted to investigate the molecularmechanisms involved in the reduced insulin sensitivity observedin rats chronically treated with oral Arg. With this purpose wefirst evaluated total, as well as the basal and insulin-inducedphosphorylation activity of proteins involved in the transmissionof insulin signaling [insulin receptor (IR), IRS 1/2, and Akt] inskeletal muscles [soleus and extensorum digitalis longus (EDL)],white adipose tissue (WAT), and liver. We then assessed GLUT4protein content in plasma membrane (PM) and microsome (M)of muscles and WAT. Considering the potential role of GH in thegenesis of these events (11, 16), we further evaluated the expres-sion of pituitary GH mRNA, GH, and IGF-I mRNA in liver, aswell as the phosphorylation activity of some proteins involved inthe GH signaling pathway in skeletal muscles, WAT, and liver.
Materials and Methods
Animals and treatmentsMale Wistar rats weighing approximately 200 g obtained from our
own breeding colony were used. They were maintained on a standard ratchow containing 22% of protein (Nuvilab CR1; Nuvital Nutrientes S/A,Colombo, Paraná, Brazil), and tap water ad libitum in a room kept atconstant temperature (23 � 1 C) and on a 12-h light, 12-h dark (lightson at 0615 h) schedule. Arg treatment was performed by its addition indrinking water in a concentration of 35 mg Arg/d/animal (A35) (3, 17)for 4 wk, a dose approximately seven times lower than the one used toinduce GH secretion in humans, and considered safe for rats because theycan tolerate oral administration of 2.14–5.70 g Arg/kg body weight�d(18). We have also considered that only approximately half of the in-gested Arg remains in the plasma (19). The control group received justpure water.
The animals were settled in collective cages because no differenceswere detected in food and water intake in grouped or individually housedArg-treated rats (6).
The experiments included the study of the insulin sensitivity by eval-uating some steps of the insulin and GH signaling pathway, as well as ofGLUT4 and GH gene expression. At the end of the treatment period, each
animal was anesthetized with ketamine and xylazine (100 and 10 mg/kgbody, respectively) and killed by decapitation.
The experimental protocol (no. 132/05) conforms to ethical princi-ples in animal research adopted by the Brazilian College of Animal Ex-perimentation and was approved by the Institute of Biomedical Sciences/University of Sao Paulo-Ethical Committee for Animal Research.
Procedures
Evaluation of the total and phosphorylated IR, IRS 1/2, andAkt content
Soleus, EDL, WAT, and liver of rats were removed before and afteran iv administration of regular insulin (1 U) as described (20, 21).
The tissues were homogenized, and the samples were placed in a hotbuffer (96 C) [100 mM Trizma-Base (Sigma-Aldrich Corp., St. Louis,MO) (pH 7.5), 10 mM EDTA, 1% sodium dodecyl sulfate, 100 mM
sodium fluoride, 10 mM sodium pyrophosphate, and 10 mM sodiumorthovanadate] and centrifuged for 40 min at 4 C, at 13, 400 � g.Immunoblotting of the tissues protein extracts was performed using an-tiphosphotyrosine (1:200), anti-pAkt (1:500), and anti-IR (1:500) (SantaCruz Biotechnology, Inc., Santa Cruz, CA), anti-IRS2 (1:500) (ImunyBiotechnology Ltda, Campinas, Sao Paulo, Brazil), and anti-IRS1 (1:2000) and anti-Akt2 (1:1000) (Upstate Biotechnology Inc., Lake Placid,NY), according to the manufacturer’s recommendations. The band de-tection was performed using secondary peroxidase-conjugated antibody(1:10,000; Santa Cruz Biotechnology) and the Enhanced Chemilumi-nescence (ECL) kit (Amersham Biosciences, Buckinghamshire, UK).Blots were analyzed with Scion Image software (Scion Corp., Frederick,MD). Results were expressed as arbitrary units (AUs).
Evaluation of GLUT 4 content in PM and M fractions ofskeletal muscle and adipose tissues
Soleus and EDL muscles were homogenized in 10 mM Trizma, 1 mM
EDTA, and 250 mM sucrose (pH 7.4) buffer. PM and M fractions ofGLUT4 were separated by ultracentrifugation according to Yonemitsuet al. (22) with modifications, as described (23). The WAT samples weresubmitted to a subcellular fractionation according to Fabres-Machadoand Saito (24).
Immunoblotting of the tissues protein extracts was performed usinganti-GLUT4 antiserum (1:3000 Polyclonal Rabbit Raised Anti-GLUT4,AB1346; CHEMICON International, Inc., Temecula, CA) in 8% BSA/PBS solution, followed by the incubation with appropriated secondaryperoxidase-conjugated antibody. The band detection was performed aspreviously described.
Considering the total protein yield of each subcellular membranefraction (PM and M) and considering the respective tissue weight, PMand M GLUT4 protein per gram of tissue was calculated (AU/g tissue);and the sum of these data were considered the total GLUT4 content pergram of tissue. The percentage of total cellular GLUT4 present in the PMwas then calculated as indicated: (100 � PM GLUT4)/(PM GLUT4 � MGLUT4) (23).
Evaluation of GH mRNA expression by Northern blottinganalysis
Total RNA was extracted by the guanidinium thiocyanate-phenol-chloroform method, electrophoresed, transferred to a nylon membrane,and subjected to Northern blotting analysis, as described (6). The resultswere expressed as mean � SEM of GH mRNA/18S rRNA ratio.
Evaluation of GH protein expression by Western blottinganalysis
The pituitaries were removed, homogenized in 0.25 M sucrose, 2 mM
MgCl2, and 20 mM Tris-HCl buffer, and centrifuged at 100 � g for 10 minat 4 C; the supernatant was centrifuged at 800 � g for 10 min at 4 C.Immunoblotting of pituitary protein extracts was performed using anti-GHantibody (1:5000) (National Hormone and Pituitary Program, National
Endocrinology, May 2009, 150(5):2080–2086 endo.endojournals.org 2081
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 14 July 2014. at 15:20 For personal use only. No other uses without permission. . All rights reserved.
Institute of Diabetes and Digestive and Kidney Diseases, Torrance, CA),followed by the incubation with appropriated secondary peroxidase-con-jugated antibody. The band detection was performed as described.
IGF-I mRNA expression by real-time PCRTotal RNA was extracted of liver tissue (�50 mg), as described (6). Two
micrograms of total RNA were used to synthesize the first-strand cDNAusing oligo-deoxythymidine primers and the Moloney murine leukemia vi-rus reverse-transcriptase kit (Invitrogen Corp., Carlsbad, CA) according tothe manufacturer’s recommendations. Reverse-transcriptase reaction wasperformed at 70 C for 10 min, followed by 37 C for 60 min, and 10 min at95 C. Real-time quantitative PCR amplification was performed using theSYBR Green PCR Master Mix Kit (Applied Biosystems, Warrington, UK),and the primers for IGF-I or cyclophilin, which are listed in Table 1. Thereaction conditions consisted of two steps at 50 C for 2 min and 95 C for 10min, followed by 45 cycles of three steps: 20 sec denaturation at 95 C, 60 secannealing at 58 C, and 20 sec at 72 C. SYBR Green-based real-time PCRanalysiswasperformedwiththeABIPrism7300sequencedetector (AppliedBiosystems) according to the manufacturer’s instructions.
Evaluation of total and/or phosphorylated Janus kinase(JAK) 2, signal transducer and activator of transcription(STAT) 1, STAT3, STAT5, p85�, and p55� protein content
Control and Arg-treated rats were anesthetized, and soleus, EDL,WAT, and a piece of liver were removed and immediately homogenizedin a hot buffer, as described previously. Immunoblotting of tissue proteinextracts was performed using the antibodies: anti-STAT1 (1:500), anti-STAT3 (1:500), anti-STAT5 (1:500), and anti-pSTAT3 (1:500) (SantaCruz Biotechnology); anti-JAK2 (1:1,000), anti-pJAK2 (1:2,000), anti-pSTAT1 (1:2,000), anti-pSTAT5 A/B (1:1,000), and anti-p85� (1:2,000)(Upstate Biotechnology); followed by the secondary peroxidase-conju-gated antibody (1:10,000; Santa Cruz Biotechnology). The band detec-tion was performed as described.
Statistical analysisAll data were expressed as mean � SEM, and the significance level was
set at 5% (P � 0.05). The statistical analysis was performed by a two-wayANOVA, followed by Bonferroni posttest. Other data were analyzedusing an unpaired nonparametric Student’s t test (Prism GraphPad, Ver-sion: 4.0; GraphPad Software Inc., San Diego, CA).
Results
Effects of Arg supplementation on total andphosphorylated content of IR, IRS 1/2, and Akt
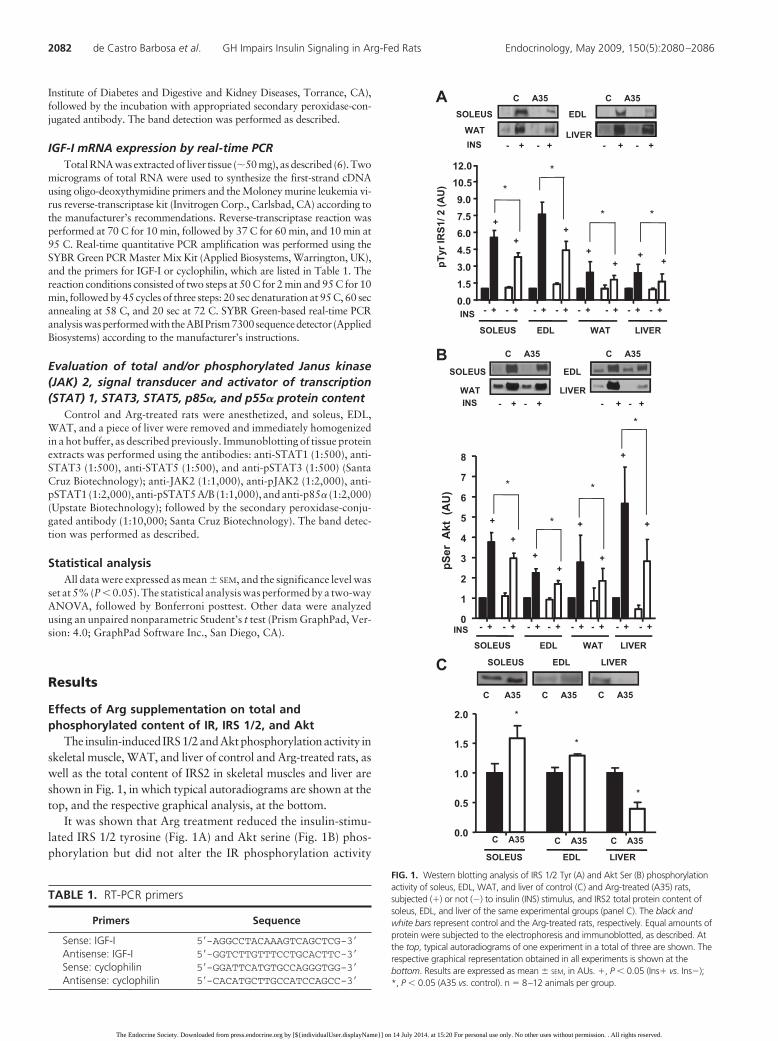
The insulin-induced IRS1/2andAktphosphorylationactivity inskeletal muscle, WAT, and liver of control and Arg-treated rats, aswell as the total content of IRS2 in skeletal muscles and liver areshown in Fig. 1, in which typical autoradiograms are shown at thetop, and the respective graphical analysis, at the bottom.
It was shown that Arg treatment reduced the insulin-stimu-lated IRS 1/2 tyrosine (Fig. 1A) and Akt serine (Fig. 1B) phos-phorylation but did not alter the IR phosphorylation activity
TABLE 1. RT-PCR primers
Primers Sequence
Sense: IGF-I 5�-AGGCCTACAAAGTCAGCTCG-3�Antisense: IGF-I 5�-GGTCTTGTTTCCTGCACTTC-3�Sense: cyclophilin 5�-GGATTCATGTGCCAGGGTGG-3�Antisense: cyclophilin 5�-CACATGCTTGCCATCCAGCC-3�
B
0
1
2
3
4
5
6
7
8
INS
pSer
Akt
(A
U)
- +
SOLEUS EDL WAT LIVER
- + - + - + - + - + - + - +
+
+
++
+
+
+
+
*
*
*
*
SOLEUS EDL
WAT LIVER
C A35 C A35
- + - +- + - +INS
ASOLEUS EDL
WAT LIVER- + - + - + - +
C A35 C A35
INS
0.01.53.04.56.07.59.0
10.512.0
INS
pTyr
IRS1
/ 2 (A
U)
- + - + - + - + - + - + - + - +
SOLEUS EDL WAT LIVER
+
*
+
++
++
+
*
* *
C SOLEUS EDL LIVER
C A35 C A35 C A35
SOLEUS EDL LIVER
0.0
0.5
1.0
1.5
2.0 *
*
*
C A35 C A35 C A35
FIG. 1. Western blotting analysis of IRS 1/2 Tyr (A) and Akt Ser (B) phosphorylationactivity of soleus, EDL, WAT, and liver of control (C) and Arg-treated (A35) rats,subjected (�) or not (�) to insulin (INS) stimulus, and IRS2 total protein content ofsoleus, EDL, and liver of the same experimental groups (panel C). The black andwhite bars represent control and the Arg-treated rats, respectively. Equal amounts ofprotein were subjected to the electrophoresis and immunoblotted, as described. Atthe top, typical autoradiograms of one experiment in a total of three are shown. Therespective graphical representation obtained in all experiments is shown at thebottom. Results are expressed as mean � SEM, in AUs. �, P � 0.05 (Ins� vs. Ins�);*, P � 0.05 (A35 vs. control). n � 8–12 animals per group.
2082 de Castro Barbosa et al. GH Impairs Insulin Signaling in Arg-Fed Rats Endocrinology, May 2009, 150(5):2080–2086
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 14 July 2014. at 15:20 For personal use only. No other uses without permission. . All rights reserved.

(data not shown), in all tissues studied, when compared with thecontrol group. As expected, insulin induced IR, IRS 1/2, and Aktphosphorylation, despite the treatment in use, which indicatesthe efficiency of the methodology used.
Arg treatment did not modify the total IR and Akt2 contentin the same tissues, even though it increased IRS1 content inWAT (1.52 � 0.15 vs. 1.0 � 0.03 AU; P � 0.05). Arg treatmentalso increased total IRS2 content in soleus and EDL muscles,whereas decreased it in liver, as shown in Fig. 1C.
Effects of Arg on total and PM GLUT4 content inskeletal muscle and adipose tissue
The results of this study are shown in Fig. 2, in which Fig. 2Arepresents the total GLUT4 content (M plus PM), and Fig. 2B,the PM GLUT4 content in soleus, EDL, and WAT of control andArg-treated rats. It was observed that Arg treatment induced asignificant reduction in the total GLUT4 content in soleus andWAT (EDL muscle: P � 0.09) (Fig. 2A) and in the PM GLUT4content of all tissues studied (Fig. 2B). GLUT4 content in Mfraction was not modified by the Arg treatment (Fig. 2A).
Effects of Arg on GH-IGF-I axisFigure 3 illustrates the effects of chronic Arg administration
on pituitary GH mRNA and protein expression (Fig. 3A), as wellas on liver IGF-I mRNA levels (Fig. 3B). It was observed that bothGH mRNA and protein levels increased significantly in the Argtreated; the same occurred with the IGF-I mRNA abundance inliver.
Effects of Arg supplementation on total and/orphosphorylated JAK2, STAT1, STAT3, STAT5, p85�, andp55� protein content
The effect of Arg treatment on the proteins involved in the GHsignaling pathway in skeletal muscles, WAT, and liver is shownin Fig. 4. It was observed that Arg supplementation induced anincrease in the tyrosine phosphorylation activity of STAT3 in WATand liver (Fig. 4A), without any noticeable changes in total STAT3content (data not shown); the same occurred with JAK2 tyrosinephosphorylation activity in EDL (2.13 � 0.45 vs. 1.0 � 0.22 AU;P � 0.05). Total JAK2 content was increased only in WAT of Arg-
B
C A35 C A35 C A350
25
50
75
100
*
SOLEUS EDL
**
WAT
PM G
LUT4
Pro
tein
Con
tent
(%)
A
C A35 C A35 C A350.0
0.5
1.0
1.5
*
PMM
SOLEUS EDL WAT
Tota
l GLU
T4/ g
of t
issu
e (A
U)
*
PMC A35
SOLEUS EDL WAT
C A35 C A35
M
FIG. 2. Western blotting analysis of total (M plus PM) and PM GLUT4 content insoleus, EDL, and WAT of control (C) and Arg-treated (A35) rats is shown in Aand B, respectively. A, Typical autoradiograms representative of GLUT4 proteinanalysis in M (black bar) and PM (white bar) fractions obtained from the specifiedtissues are shown at the top, and the graphical analysis representative of the sumof GLUT4 present in M and PM fractions is shown at the bottom. B, Thegraphical representation of the amount (%) of GLUT4 protein measured in thePM fraction of the soleus, EDL, and WAT of the same experimental groups isshown. Results are expressed as mean � SEM of total GLUT4/g tissue or PMGLUT4 content (%), in AUs. *, P � 0.05 (A35 vs. control).
GH mRNAGH protein
0.0
0.5
1.0
1.5
2.0
*
*
C A35A35
GH
exp
ress
ion
(AU
)
C
GH mRNA
18S rRNAGH protein
C A35
A
B
C A350.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
*
IGF-
I mR
NA
exp
ress
ion
(AU
)
FIG. 3. Northern and Western blotting analysis of pituitary GH mRNA and GHcontent, and RT-PCR analysis of liver IGF-I mRNA expression of control (C) andArg-treated (A35) rats, are shown in A and B, respectively. A, Typicalautoradiograms representative of GH mRNA, 18S rRNA, and GH obtained frompituitaries are shown at the top, and the graphical analysis representative of thehybridization of rat GH and 18S rRNA transcripts ratio and GH protein contentobtained by densitometric analysis of the exposed films, is shown at the bottom.B, The graphical representation of IGF-I mRNA content in liver of the sameexperimental groups is shown. Data are expressed as mean � SEM, in AUs(arbitrary units). *, P � 0.05 (A35 vs. control). n � 11–17 animals per group.
Endocrinology, May 2009, 150(5):2080–2086 endo.endojournals.org 2083
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 14 July 2014. at 15:20 For personal use only. No other uses without permission. . All rights reserved.

treated rats (1.5 � 0.2 vs. 1.0 � 0.05 AU; P � 0.05), even thoughtyrosine-phosphorylated JAK2 content remained unaltered.
Figure 4B shows that Arg treatment increased the tyrosine-phosphorylated STAT5 content in soleus and EDL muscles,whereas reduced it in WAT. No changes were detected in totalSTAT5 content, as well as in the total and tyrosine-phosphory-lated STAT1 protein content in all tissues studied (data notshown).
Arg treatment was shown to enhance the PI3-K regulatorysubunit p85 total content in soleus muscle, and to decrease it inWAT and liver, as shown the Fig. 4C. However, the contraryoccurred in soleus, EDL, WAT, and liver PI3-K regulatory sub-unit p55 total content, which was increased, as pointed out inFig. 4D.
Discussion
The current study presents the molecular mechanisms by whichdietary supplementation with Arg induces insulin resistance, andprovides evidence for a potential role of GH in this process.
First, we evaluated the total and phosphorylated proteins in-volved in insulin signaling pathway, such as IR, IRS 1/2, and Akt,in skeletal muscle, adipose tissue, and liver of control and Arg-treated rats. We observed that, whereas IR basal and insulin-stimulated tyrosine phosphorylation content remained unaf-fected, insulin-stimulated IRS 1/2 tyrosine and Akt serinephosphorylation activity were decreased in all tissues studied ofthe Arg-treated rats, indicating that Arg treatment impaired in-sulin signaling transmission at a post-receptor level. These eventswere observed even in the presence of increased levels of totalIRS1 and IRS2 content, as well as unaltered IR and Akt totalcontent.
This is consistent with most studies pointing that the molec-ular mechanism responsible for insulin resistance in target tissueslies at a post-receptor step (25). It is noteworthy that the Aktserine phosphorylation was reduced in liver of Arg-treated rats,even before the insulin stimulus, and that, whereas in muscleand adipose tissues the augment in the Akt phosphorylation inresponse to insulin was around 20 –30% lower than in controlrats, in liver it was reduced at 50%, a finding that suggests thatliver plays a major role in the insulin resistance settled. Thisis reinforced by the decreased total content of IRS2, in liver ofArg-treated rats, because it is considered the major protein inthe insulin-signaling pathway in this tissue (12).
One possibility to be considered for this insulin signal trans-duction disturbance is that rats under chronic Arg treatmentpresent hyperinsulinemia (6), which has been related to a de-crease in IRS1 tyrosine phosphorylation activity and an increasein IRS1 degradation in adipocytes (26). Furthermore, prolongedexposure to insulin was shown to exacerbate the insulin resis-tance state because it is associated with serine phosphorylationof IRS1 and with the inhibition of PI3-K activity (27).
These proteins are essential in signaling mechanisms im-plicated in the various biological responses to insulin becausethey are involved in Akt serine phosphorylation, GLUT4translocation to PM and glucose uptake, primarily in skeletalmuscle and adipose tissue, as well as in reducing hepatic glu-cose output (26, 27).
Because the clearance of circulating glucose depends mainlyon insulin-induced GLUT4 translocation to PM in muscle andadipose cells, in which this protein is the major GLUT (28), wehave subsequently focused our study on the evaluation of totalGLUT4 content and GLUT4 distribution in PM and M of soleus,EDL, and WAT.
Arg treatment induced a significant reduction in soleus, EDL,and WAT PM GLUT4 content, which strongly indicates thatglucose uptake rate is impaired. The parallel reduction in thetotal GLUT4 content, mainly in soleus and WAT, might con-tribute to the decreased GLUT4 availability in PM because itappears to be the major determinant in the target tissues’ abilityto respond to insulin (28, 29). These results are in accordancewith the impairment of the insulin signaling exhibited by theseanimals, presently described.
Considering that insulin exerts a direct control upon tran-script translation, by activation of eukaryotic translation initi-ation (eIF) and elongation factors, mainly eIF2B, eIF4E, andelongation factor 2, it is expected that the insulin resistance
A BWAT LIVER
C A35 C A35
SOLEUS EDL WAT
0.0
0.5
1.0
1.5
2.0
2.5
*
*
*pTyr
STA
T5 (A
U)
C A35 C A35 C A350
1
2
3 *
pTyr
STA
T3 (A
U)
C CA35 A35
WAT LIVER
*
WATSOLEUS EDL
C A35 C A35 C A35
DWAT LIVER
SOLEUS EDL
C A35 C A35
WAT LIVERSOLEUS
C A35 C A35 C A35
SOLEUS EDL WAT LIVER
0.0
0.5
1.0
1.5
2.0
2.5
**
p55
Tota
l Pro
tein
Con
tent
(AU
)
*
*
C A35 C A35 C A35C A35SOLEUS WAT LIVER
0.0
0.5
1.0
1.5*
* *
p85
Tota
l Pro
tein
Con
tent
(AU
)
C A35 C A35 C A35
C
FIG. 4. Western blotting analysis of STAT3 (panel A) and STAT5 (panel B)phosphorylation activity in WAT and liver, and skeletal muscle and WAT,respectively, of control (panel C) and Arg-treated (A35) rats, as well as of thetotal p85� (panel C) content in soleus, WAT, and liver, and p55� (panel D)content in all tissues studied. The black and white bars represent control and theArg-treated rats, respectively. Equal amounts of protein were subjected to theelectrophoresis and immunoblotted, as described. At the top, typicalautoradiograms of one experiment in a total of three are shown. The respectivegraphical representation obtained in all experiments is shown at the bottom.Results are expressed as mean � SEM, in AUs (arbitrary units). *, P � 0.05 (A35vs. control). n � 5 animals per group.
2084 de Castro Barbosa et al. GH Impairs Insulin Signaling in Arg-Fed Rats Endocrinology, May 2009, 150(5):2080–2086
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 14 July 2014. at 15:20 For personal use only. No other uses without permission. . All rights reserved.

installed could impair the protein synthesis, which could ex-plain the reduced total GLUT4 content observed in the tissuesunder evaluation, where this protein is highly expressed (30).That might be the reason why the effect of Arg was less prom-inent in the EDL muscle because GLUT4 expression is knownto be lower in fast glycolytic muscles in comparison to slowoxidative ones (29).
Another potential candidate for the insulin resistance inducedby Arg chronic treatment could be the GH, whose secretion wasshown to be increased by acute Arg administration, and whosediabetogenic effects have been extensively reported (11, 16).Thus, the third part of our study was aimed to evaluate the effectof the chronic Arg treatment in the activity of the GH-IGF-I axis.
The data obtained show that Arg treatment increased GHmRNA expression, confirming our previous report (5, 6), andalso the pituitary GH content (6). One possible explanation forthis finding might involve the well-known inhibitory effect of Argupon somatostatin release (2, 7), which causes an increase in so-matotropes’ cAMP levels, and subsequently, activation of PKA andphosphorylationofpituitary transcriptional factors, likePIT1, thatcauses marked alterations in GH gene transcription, as described(31). Inhibition of somatostatin release mediated by Arg could alsoleadtoapositivetonusofGHRHonsomatotropes,ortoanincreasein steady-state levels of GHRH (32).
In addition, Arg can exert a direct effect on the pituitaryand induce GH gene expression, as shown by in vitro studiesdeveloped in our laboratory (5), a finding that is still underinvestigation.
Although the GH secretagogue activity of acute Arg adminis-tration is well established, we decided to evaluate this issue in Argchronically treated rats, by determination of the IGF-I mRNA ex-pression in liver because GH is the most important regulator ofhepatic IGF-I synthesis. Increased hepatic IGF-I mRNA expressionwas found, confirming the Arg effect on GH release.
Thus, considering these results, we can hypothesize that inArg-treated rats, the cells are being subjected to increased levelsof GH, which might lead to insulin resistance. In fact, the acti-vation of the GH signaling pathways in insulin target tissues ofthese rats was evidenced by the increase in tyrosine phosphory-lation of JAK2 in EDL, of STAT3 in WAT and liver, and STAT5in skeletal muscle. Moreover,an increase in the soleus total contentof p85� regulatory subunit of PI3-K, which is known to be up-regulated by GH, was detected, which might contribute to the in-sulin resistance because it competes in a dominant-negative fashionwith the p85-p110 heterodimer for binding to IRS1 protein (33,34). In addition, GH was shown to play a role in inducing insulinresistance via signaling cross talk with insulin. However, this issueis still contradictory and remains unclarified (11, 12, 35).
As far as we know, p55 is not under the control of GH, how-ever, an increase in its total content was observed in skeletalmuscle, WAT, and liver of rats under Arg treatment. This proteinacts as a negative regulator of PI3-K activity, further indicatingthat the insulin signaling is impaired in these tissues (36).
The relationship between GH and insulin resistance noted inthis investigation is reinforced by studies that described insulinresistance in rats chronically treated with GH, in transgenic miceoverexpressing GH, and in acromegalic patients (10–12). In
these models GH excess was shown to decrease IRS1 tyrosinephosphorylation and IRS1/PI3-K association in skeletal muscle,adipose tissue, and liver, making these tissues resistant to exog-enous insulin (11, 13, 35). In addition, GH has been shown toindirectly activate c-Jun N-terminal kinase/stress-activated pro-tein kinase activity, which is known to mediate feedback inhi-bition of the insulin-signaling cascade (37).
In summary, the data obtained in this study refine and con-firm the insulin resistance evidenced in our previous study, pro-viding support for the reduced GLUT4 content in PM of musclesand WAT of chronically Arg-treated rats, and pointing to a roleof GH in the genesis of these events.
It is worth mentioning that in the present study, we charac-terized an insulin resistance state in a stage in which the increasedinsulin secretion seemed to account for normoglycemia. How-ever, in more advanced stages, alterations in glucose homeostasiswill probably occur and lead to the type 2 diabetes, a disease thatis expanding with epidemic proportions. This has to be consid-ered because Arg has been largely used as a dietary supplement,intended to enhance performance.
Acknowledgments
We thank the Laboratorios Baldacci (Sao Paulo-Brazil) for kindly pro-viding the amino acid arginine.
Address all correspondence and requests for reprints to: MariaTereza Nunes, Ph.D., Associate Professor, Department of Physiologyand Biophysics, Institute of Biomedical Sciences, University of Sao Paulo,05508-900 Sao Paulo, Sao Paulo, Brazil. E-mail: [email protected].
This work was supported by a grant from the Fundacao de Amparoa Pesquisa do Estado de Sao Paulo (02/07384-4) (to U.F.M. and M.T.N.).M.T.N., U.F.M., and S.B. are recipients of Conselho Nacional de Pes-quisa e Desenvolvimento fellowships. T.d.C.B. and J.E.N.d.C. are re-cipients of Fundacao de Amparo a Pesquisa do Estado de Sao Paulofellowships.
Disclosure Summary: The authors have nothing to disclose.
References
1. Morris Jr SM 2007 Arginine metabolism: boundaries of our knowledge. J Nutr137(6 Suppl 2):1602S–1609S
2. Ghigo E, Ceda GP, Valcavi R, Goffi S, Zini M, Mucci M, Valenti G, Cocchi D,Muller EE, Camanni F 1994 Low doses of either intravenously or orally admin-isteredarginineareable to enhancegrowthhormone response togrowthhormonereleasing hormone in elderly subjects. J Endocrinol Invest 17:113–117
3. Blum A, Hathaway L, Mincemoyer R, Schenke WH, Kirby M, Csako G,Waclawiw MA, Panza JA, Cannon RO 2000 Oral L-arginine in patients withcoronary artery disease on medical management. Circulation 101:2160–2164
4. Clarkson P, Adams MR, Powe AJ, Donald AE, McCredie R, Robinson J,McCarthy SN, Keech A, Celermajer DS, Deanfield JE 1996 Oral L-arginineimproves endothelium-dependent dilation in hypercholesterolemic youngadults. J Clin Invest 97:1989–1994
5. Adriao M, Chrisman CJ, Bielavsky M, Olinto SC, Shiraishi EM, Nunes MT2004 Arginine increases growth hormone gene expression in rat pituitary andGH3 cells. Neuroendocrinology 79:26–33
6. Castro-Barbosa T, Lourenco-Poyares L, Fabres-Machado U, Nunes MT 2006Chronic oral administration of arginine induces GH gene expression and in-sulin resistance. Life Sci 79:1444–1449
7. Alba-Roth J, Muller OA, Schopohl J, von Werder K 1988 Arginine stimulatesgrowth hormone secretion by suppressing endogenous somatostatin secretion.J Clin Endocrinol Metab 67:1186–1189
8. Villlalobos C, Nunez L, Garcia-Sancho J 1997 Mechanisms for stimulation
Endocrinology, May 2009, 150(5):2080–2086 endo.endojournals.org 2085
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 14 July 2014. at 15:20 For personal use only. No other uses without permission. . All rights reserved.

of rat anterior pituitary cells by arginine and other amino acids. J Physiol502(Pt 2):421– 431
9. Bolea S, Pertusa JA, Martín F, Sanchez-Andres JV, Soria B 1997 Regulation ofpancreatic �-cell electrical activity and insulin release by physiological aminoacid concentrations. Pflugers Arch 433:699–704
10. Hansen I, Tsalikian E, Beaufrere B, Gerich J, Haymond M, Rizza R 1986Insulin resistance in acromegaly: defects in both hepatic and extrahepatic in-sulin action. Am J Physiol 250(3 Pt 1):E269–E273
11. Smith TR, Elmendorf JS, David TS, Turinsky J 1997 Growth hormone inducedinsulin resistance: role of the insulin receptor, IRS-1, GLUT-1 and GLUT-4.Am J Physiol 272(6 Pt 1):E1071–E1079
12. Cho Y, Ariga M, Uchijima Y, Kimura K, Rho JY, Furuhata Y, Hakuno F,Yamanouchi K, Nishihara M, Takahashi S 2006 The novel roles of liver forcompensation of insulin resistance in human growth hormone transgenic rats.Endocrinology 147:5374–5384
13. Dominici FP, Cifone D, Bartke A, Turyn D 1999 Alterations in the early stepsof the insulin signaling system in skeletal muscle of GH transgenic mice. Am JPhysiol 277(3 Pt 1):E447–E454
14. Shepherd PR, Withers DJ, Siddle K 1998 Phosphoinositide 3-kinase: the keyswitch mechanism in insulin signaling. Biochem J 333(Pt 3):471–490
15. Cheatham B, Kahn CR 1995 Insulin action and the insulin signaling network.Endocr Rev 16:117–142
16. Davidson MB 1987 Effect of growth hormone on carbohydrate and lipidmetabolism. Endocr Rev 8:115–131
17. Collier SR, Casey DP, Kanaley JA 2005 Growth hormone responses to varyingdoses of oral arginine. Growth Horm IGF Res 15:136–139
18. Wu G, Bazer FW, Cudd TA, Jobgen WS, Kim SW, Lassala A, Li P, Matis JH,Meininger CJ, Spencer TE 2007 Pharmacokinetics and safety of arginine sup-plementation in animals. J Nutr 137(6 Suppl 2):1673S–1680S
19. Windmueller HG, Spaeth AE 1976 Metabolism of absorbed aspartate, aspar-agine, and arginine by rat small intestine in vivo. Arch Biochem Biophys 175:670–676
20. Saad MJ, Folli F, Kahn JA, Kahn CR 1993 Modulation of insulin receptor,insulin receptor substrate-1, and phosphatidylinositol 3-kinase in liver andmuscle of dexamethasone-treated rats. J Clin Invest 92:2065–2072
21. Araujo EP, De Souza CT, Gasparetti AL, Ueno M, Boschero AC, Saad MJ,Velloso LA 2005 Short-term in vivo inhibition of insulin receptor substrate-1expression leads to insulin resistance, hyperinsulinemia, and increased adi-posity. Endocrinology 146:1428–1437
22. Yonemitsu S, Nishimura H, Shintani M, Inoue R, Yamamoto Y, Masuzaki H,Ogawa Y, Hosoda K, Inoue G, Hayashi T, Nakao K 2001 Troglitazone in-duces GLUT 4 translocation in L6 myotubes. Diabetes 50:1093–1101
23. Mori RC, Hirabara SM, Hirata AE, Okamoto MM, Machado UF 2008Glimepiride as insulin sensitizer: increased liver and muscle responses to in-sulin. Diabetes Obes Metab 10:596–600
24. Fabres-Machado U, Saito M 1995 The effect of adipose cell size on the mea-
surement of GLUT 4 in white adipose tissue of obese mice. Braz J Med Biol Res28:369–376
25. White MF 1997 The insulin signalling system and the IRS proteins. Diabeto-logia 40(Suppl 2):S2–S17
26. Zhande R, Mitchell JJ, Wu J, Sun XJ 2002 Molecular mechanism of insulin-induced degradation of insulin receptor substrate 1. Mol Cell Biol 22:1016 –1026
27. Paz K, Hemi R, LeRoith D, Karasik A, Elhanany E, Kanety H, Zick Y 1997 Amolecular basis for insulin resistance: elevated serine/threonine phosphoryla-tion of IRS-1 and IRS-2 inhibits their binding to the juxtamembrane region ofthe insulin receptor and impairs their ability to undergo insulin-induced ty-rosine phosphorylation. J Biol Chem 272:29911–29918
28. Herman MA, Kahn BB 2006 Glucose transport and sensing in the maintenanceof glucose homeostasis and metabolic harmony. J Clin Invest 116:1767–1775
29. Kern M, Wells JA, Stephens JM, Elton CW, Friedman JE, Tapscott EB, PekalaPH, Dohm GL 1990 Insulin responsiveness in skeletal muscle is determined byglucose transporter (GLUT4) protein level. Biochem J 270:397–400
30. Kimball SR, Farrell PA, Jefferson LS 2002 Role of insulin in translationalcontrol of protein synthesis in skeletal muscle by amino acids or exercise.J Appl Physiol 93:1168 –1180
31. Chung HO, Kato T, Tomizawa K, Kato Y 1998 Molecular cloning of pit-1cDNA from porcine anterior pituitary and its involvement in pituitary stim-ulation by growth hormone-releasing factor. Exp Clin Endocrinol Diabetes106:203–210
32. Sugihara H, Minami S, Okada K, Kamegai J, Hasegawa O, Wakabayashi I1993 Somatostatin reduces transcription of the growth hormone gene in rats.Endocrinology 132:1225–1229
33. del Rincon JP, Iida K, Gaylinn BD, McCurdy CE, Leitner JW, Barbour LA,Kopchick JJ, Friedman JE, Draznin B, Thorner MO 2007 Growth hormoneregulation of p85� expression and phosphoinositide 3-kinase activity in adi-pose tissue: mechanism for growth hormone-mediated insulin resistance. Di-abetes 56:1638–1646
34. Barbour LA, Mizanoor Rahman S, Gurevich I, Leitner JW, Fischer SJ, RoperMD, Knotts TA, Vo Y, McCurdy CE, Yakar S, Leroith D, Kahn CR, CantleyLC,FriedmanJE,DrazninB2005 IncreasedP85� is apotentnegative regulatorof skeletal muscle insulin signaling and induces in vivo insulin resistance as-sociated with growth hormone excess. J Biol Chem 280:37489–37494
35. Nielsen C, Gormsen LC, Jessen N, Pedersen SB, Møller N, Lund S, Jørgensen JO2008 Growth hormone signaling in vivo in human muscle and adipose tissue:impact of insulin, substrate background, and growth hormone receptor blockade.J Clin Endocrinol Metab 93:2842–2850
36. Chen D, Mauvais-Jarvis F, Bluher M, Fisher SJ, Jozsi A, Goodyear LJ, Ueki K,Kahn CR 2004 p50�/p55� phosphoinositide 3-kinase knockout mice exhibitenhanced insulin sensitivity. Mol Cell Biol 24:320–329
37. Lee YH, Giraud J, Davis RJ, White MF 2003 c-Jun N-terminal kinase (JNK)mediates feedback inhibition of the insulin signaling cascade. J Biol Chem278:2896–2902
2086 de Castro Barbosa et al. GH Impairs Insulin Signaling in Arg-Fed Rats Endocrinology, May 2009, 150(5):2080–2086
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 14 July 2014. at 15:20 For personal use only. No other uses without permission. . All rights reserved.