prey transport mechanisms in blindsnakes and the evolution
TRANSCRIPT

1321
AMER. ZOOL., 41:1321–1337 (2001)
Prey Transport Mechanisms in Blindsnakes and the Evolution of UnilateralFeeding Systems in Snakes1
NATHAN J. KLEY2
Organismic & Evolutionary Biology Program, University of Massachusetts,Amherst, Massachusetts 01003
SYNOPSIS. Most snakes ingest and transport their prey via a jaw ratcheting mech-anism in which the left and right upper jaw arches are advanced over the prey inan alternating, unilateral fashion. This unilateral jaw ratcheting mechanism differsgreatly from the hyolingual and inertial transport mechanisms used by lizards,both of which are characterized by bilaterally synchronous jaw movements. Giventhe well-corroborated phylogenetic hypothesis that snakes are derived from lizards,this suggests that major changes occurred in both the morphology and motor con-trol of the feeding apparatus during the early evolution of snakes. However, mostprevious studies of the evolution of unilateral feeding mechanisms in snakes havefocused almost exclusively on the morphology of the jaw apparatus because therehave been very few direct observations of feeding behavior in basal snakes. In thispaper I describe the prey transport mechanisms used by representatives of twofamilies of basal snakes, Leptotyphlopidae and Typhlopidae. In Leptotyphlopidae,a mandibular raking mechanism is used, in which bilaterally synchronous flexionsof the lower jaw serve to ratchet prey into and through the mouth. In Typhlopidae,a maxillary raking mechanism is used, in which asynchronous ratcheting move-ments of the highly mobile upper jaws are used to drag prey through the oralcavity. These findings suggest that the unilateral feeding mechanisms that char-acterize the majority of living snakes were not present primitively in Serpentes,but arose subsequently to the basal divergence between Scolecophidia and Ale-thinophidia.
INTRODUCTION
Three fundamental modes of intraoralprey transport are recognized within Squa-mata. Most lizards use a hyolingual trans-port mechanism, in which cycles of tongueprotraction and retraction serve to ratchetprey through the mouth and towards thepharynx (Smith, 1984; Herrel et al., 1996;Schwenk, 2000). In some lizards, however,this lingual ratcheting mechanism is aug-mented or replaced by a cranioinertialtransport mechanism, in which rapid move-ments of the entire head are used to propelprey through the oral cavity (Gans, 1969;Bramble and Wake, 1985). This mode ofintraoral transport is of particular impor-tance in varanid lizards (Smith, 1986; Eliaset al., 2000), which share with snakes a
1 From the Symposium Motor Control of VertebrateFeeding: Function and Evolution presented at the An-nual Meeting of the Society for Integrative and Com-parative Biology, 3–7 January 2001, at Chicago, Illinois.
2 E-mail: [email protected]
highly reduced tongue that lacks a frictionalsurface (McDowell, 1972; Schwenk, 1988).Finally, snakes use gnathic (jaw-based)transport mechanisms, in which kinetic el-ements of the jaw apparatus are used toratchet prey into and through the mouth(Cundall and Greene, 2000). While bothhyolingual and cranioinertial transport arecommon among tetrapods, gnathic transportmechanisms are unique to snakes (Brambleand Wake, 1985).
Nearly all of what is currently knownabout gnathic transport in snakes derivesfrom studies of taxa belonging to Macro-stomata (Fig. 1), a large and diverse cladethat includes approximately eighty-five per-cent of the more than 2,500 species of ex-tant snakes (McDiarmid et al., 1999). Thesestudies have shown that most macrostoma-tans ingest and transport their prey via a‘‘pterygoid walk’’ mechanism (Boltt andEwer, 1964), in which reciprocating ratch-eting movements of the medial upper jawarches, combined with lateral rotations of

1322 NATHAN J. KLEY
FIG. 1. Phylogenetic hypothesis of relationships ofthe major groups of snakes, modified from Tchernovet al. (2000). Interrelationships of the three scoleco-phidian families (Leptotyphlopidae, Typhlopidae andAnomalepididae) after Cundall et al. (1993). Fossiltaxa are indicated by a dagger (†).
the entire head about the cranio-vertebraljoint, serve to advance the snake’s headover its prey (Dullemeijer, 1956; Albrightand Nelson, 1959a, b; Frazzetta, 1966; Kar-dong, 1977; Cundall and Gans, 1979; Cun-dall, 1983; Kardong, 1986). Because thisunilateral jaw ratcheting mechanism differsgreatly from both the hyolingual and cran-ioinertial transport mechanisms of lizards(Kardong and Berkhoudt, 1998; Cundall,1995), its origin has been somewhat enig-matic, and there have thus been numerousattempts made to determine the evolution-ary steps through which it arose (e.g., Gans,1961; Rieppel, 1980; Lee et al., 1999; Kar-dong and Bels, 2001). However, most stud-ies that have addressed the evolutionary or-igin of unilateral feeding mechanisms insnakes have been conducted from an almostexclusively anatomical perspective becausethe actual feeding mechanisms used by bas-al snakes have remained largely unknown.
Recently, Cundall (1995) provided thefirst detailed account of feeding behavior ina basal snake, describing a prey transport
mechanism that he termed ‘‘snout shifting’’in the anilioid Cylindrophis (Fig. 1). Likethe pterygoid walk, snout shifting involvesunilateral movements of the toothed ele-ments of the upper jaws, combined withside-to-side movements of the entire head,which together serve to advance the headover the prey. However, the upper jaws inCylindrophis (and in other anilioids) remaintightly bound to the ventral elements of thebony snout (e.g., vomers, septomaxillae) byseveral short, robust ligaments. These liga-ments prevent extensive translational move-ments of the jaws such as those which areassociated with the pterygoid walk in ma-crostomatans. Independent movements ofthe upper jaws are instead achieved throughlateral rotations of the entire snout complexabout the nasofrontal articulation (prokine-sis; Frazzetta, 1962) and independent trans-lational movements of the left and rightseptomaxilla-vomer complexes to whichthe upper jaws are bound (rhinokinesis;Cundall and Shardo, 1995). Thus, in certainrespects, Cylindrophis represents an inter-mediate functional stage between lizardsand macrostomatan snakes (Cundall, 1995);despite retaining a tight connection betweenthe upper jaws and the snout (as in lizards),prey is transported via a unilateral jawratcheting mechanism (as in macrostoma-tans).
The discovery of snout shifting in Cylin-drophis, and its likely presence in other an-ilioids such as Anilius (Gans, 1961; Cun-dall, 1995), suggests that the pterygoidwalk is unique to Macrostomata, but thatthe unilateral jaw displacement patterncommon to both snout shifting and the pter-ygoid walk is primitive for Alethinophidia(Fig. 1). Whether this unilateral jaw dis-placement pattern is unique to Alethinophi-dia is unknown, however, because the feed-ing mechanisms of the three families ofblindsnakes (Scolecophidia; Fig. 1) have re-mained largely unknown (Kardong et al.,1997). The feeding behavior of these tiny,secretive snakes is known only from a fewbrief accounts (Smith, 1957; Reid and Lott,1963; Thomas, 1985; Webb and Shine,1993b; Kley and Brainerd, 1999), and thusa detailed understanding of feeding me-chanics in the group is lacking. In this paper

1323FEEDING IN BLINDSNAKES
FIG. 2. Schematic diagram of the apparatus used for magnified high-speed videography.
I describe the prey transport mechanismsused by representatives of two families ofScolecophidia (Leptotyphlopidae and Ty-phlopidae) in an effort to elucidate the phy-logenetic origin of unilateral feeding mech-anisms in snakes.
MATERIALS AND METHODS
The blindsnakes used for this study werepurchased from commercial herpetologicalsuppliers. Feeding behavior was observedin one species of Leptotyphlopidae (Lep-totyphlops dulcis (Baird and Girard), n 520) and two species of Typhlopidae (Ty-phlops lineolatus Jan, n 5 5; Rhinotyphlopsschlegelii (Bianconi), n 5 5). The snakeswere fed larvae and pupae of several spe-cies of ants (most frequently species ofCamponotus, Formica and Acanthomyops)collected in the vicinity of Amherst, Mas-sachusetts. Ant pupae ranged in size from1.3 3 3.0 mm to 3.4 3 8.4 mm, while thelarvae were generally smaller, often lessthan 1 mm in diameter.
Feeding trials for 10 individuals of Lep-totyphlops dulcis and 1 individual of Ty-phlops lineolatus were recorded using mag-nified high-speed videography (Fig. 2).(Unfortunately, all but one of the typhlopidsobserved during this study refused to feedunder the conditions required for this tech-nique.) Several hundred feedings were re-corded for each species. Because the mouthin blindsnakes is subterminal and counter-
sunk into the ventral surface of the head,feeding trials were recorded from a ventralperspective. The snakes were fed in a clearacrylic filming chamber (6 3 13 3 34 mm)positioned above an inverted Nikon SMZ-U dissecting microscope that was coupledto a Kodak EktaPro high-speed video sys-tem. Feeding sequences were recorded at250 fps (shutter speed, 1/500 sec) and thentransferred to S-VHS videotape using aPanasonic AG-1970 VCR. Selected videosequences were digitized using Adobe Pre-miere software on a Power Macintosh G3computer and jaw movements were ana-lyzed frame-by-frame using NIH Imagesoftware. Additional feeding trials were re-corded at 60 fps with a Sony DCR VX700digital camera. Finally, several feeding tri-als for T. lineolatus were recorded using vi-deofluoroscopy. X-ray videos were record-ed at 60 fps using a Sony DCR VX1000digital camera that was coupled to a Sie-mens radiographic unit equipped with a Si-recon image intensifier. In these feeding tri-als, ant pupae were injected with an aque-ous barium sulfate solution so that theycould be visualized throughout intraoraltransport and swallowing.
The cranial morphology of all three spe-cies of blindsnakes was studied primarilythrough microdissections and the examina-tion of cleared and stained skeletal prepa-rations. Microdissections were performedon 5 Leptotyphlops dulcis, 4 Typhlops li-

1324 NATHAN J. KLEY
neolatus and 3 Rhinotyphlops schlegelii toexamine the myology of the jaw apparatusin each species. Because of the extraordi-narily small size of L. dulcis (2.2–3.0 mmhead diameter, mean 5 2.6 mm), specimensof this species were generally dissected inwater or in 70% ethanol, and the entire headof each specimen was frequently immersedin Lugol’s iodine solution (Weigert’s vari-ation) to facilitate differentiation betweenclosely apposed muscle layers (Bock andShear, 1972). Following these dissections, 5L. dulcis, 3 T. lineolatus and 2 R. schlegeliiwere cleared and stained according to a pro-tocol modified from Hanken and Wassersug(1981) and 2 L. dulcis, 1 T. lineolatus and1 R. schlegelii were prepared as dried skel-etons using small dermestid beetle larvae.The bones, cartilages, ligaments and jointsof the jaw apparatus were examined andmanipulated in each of these skeletal prep-arations under a Nikon SMZ-U stereo dis-secting microscope. In addition, the headsof 3 specimens of L. dulcis were decalci-fied, dehydrated and embedded in low-vis-cosity nitrocellulose (Thomas, 1983) andserially sectioned at 30 mm in transverse,frontal and sagittal planes. Sections werestained with Hematoxylin and Picro-Pon-ceau (Humason, 1979; Thomas, 1983) andexamined using a Zeiss Axioskop com-pound microscope to verify the relation-ships of cranial elements determined fromalcoholic and cleared and stained speci-mens.
RESULTS
Magnified high-speed videography re-vealed that both Leptotyphlops dulcis andTyphlops lineolatus ingest and transporttheir insect prey using rapid jaw ratchetingmechanisms. However, the transport mech-anisms used by these species differ radical-ly from one another, and neither resemblesthe unilateral jaw ratcheting mechanismsused by alethinophidian snakes.
Leptotyphlops
Morphology of the jaw apparatus. Onlya brief description of the relevant structuresof the jaw apparatus of Leptotyphlops ispresented here. A more detailed account ofthe cranial morphology of L. dulcis will be
published elsewhere (Kley, in preparation).Descriptions of the cranial anatomy of otherspecies of Leptotyphlops are provided byHaas (1930, 1959), McDowell and Bogert(1954), List (1966), Brock (1932), and Ab-deen et al. (1991a, b, c).
One of the most peculiar features of thejaw apparatus in Leptotyphlops dulcis is thecomplete lack of teeth in the upper jaws(viz., the maxillae, palatines and ptery-goids; Fig. 3A). Within Serpentes, this con-dition is unique to Leptotyphlopidae(Greene, 1997). Furthermore, the upper jawarches are relatively immobile due to tightligamentous connections between the max-illae and other elements of the rigid snoutcomplex (especially the premaxilla and pre-frontals). As a result, ratcheting movementsof the upper jaws like those associated withprey transport in alethinophidian snakes arenot possible in Leptotyphlops. Also, in con-trast to most alethinophidians, the ligamen-tous connection between the posterior partof the pterygoid and the distal part of thequadrate is extremely weak, visible only inhistological sections as a faint band ofloosely arranged connective tissue. Thus,the lower jaw is functionally decoupledfrom the upper jaws in Leptotyphlops.
In contrast to the upper jaws, however,the lower jaw of Leptotyphlops bears teethand is highly kinetic. The mandible itself isrelatively short and is suspended from thebraincase by the exceptionally long, antero-ventrally directed quadrates (Fig. 3A). Theproximal end of each quadrate articulateswith the braincase in a relatively loose slid-ing joint. A single row of four or five teethis present on each dentary, representing theonly teeth in the skull. Because of thesomewhat cupped shape of the enlarged lat-eral flanges of the dentaries, these toothrows are oriented nearly transversely acrossthe anterior margin of the lower jaw (Fig.3B, left). Moreover, each mandibular ramusis divided into separate anterior and poste-rior halves by a highly mobile intramandi-bular joint. This joint is formed by contactbetween the posterior end of the splenial,which is closely applied to the ventromedialsurface of the dentary, and the anterior endof the angular, which runs along the ventro-lateral surface of the compound bone (Fig.
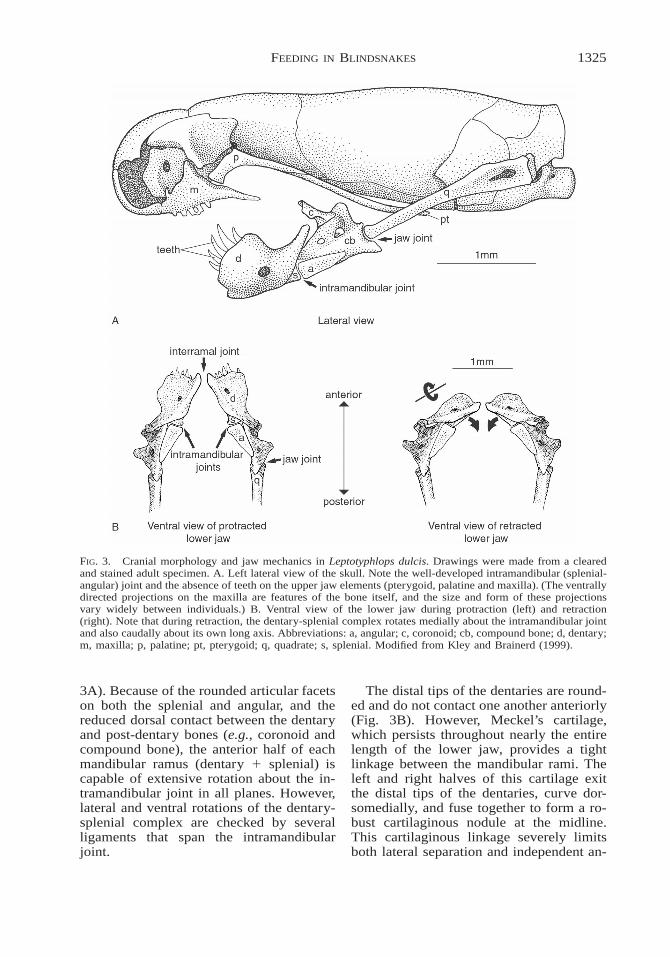
1325FEEDING IN BLINDSNAKES
FIG. 3. Cranial morphology and jaw mechanics in Leptotyphlops dulcis. Drawings were made from a clearedand stained adult specimen. A. Left lateral view of the skull. Note the well-developed intramandibular (splenial-angular) joint and the absence of teeth on the upper jaw elements (pterygoid, palatine and maxilla). (The ventrallydirected projections on the maxilla are features of the bone itself, and the size and form of these projectionsvary widely between individuals.) B. Ventral view of the lower jaw during protraction (left) and retraction(right). Note that during retraction, the dentary-splenial complex rotates medially about the intramandibular jointand also caudally about its own long axis. Abbreviations: a, angular; c, coronoid; cb, compound bone; d, dentary;m, maxilla; p, palatine; pt, pterygoid; q, quadrate; s, splenial. Modified from Kley and Brainerd (1999).
3A). Because of the rounded articular facetson both the splenial and angular, and thereduced dorsal contact between the dentaryand post-dentary bones (e.g., coronoid andcompound bone), the anterior half of eachmandibular ramus (dentary 1 splenial) iscapable of extensive rotation about the in-tramandibular joint in all planes. However,lateral and ventral rotations of the dentary-splenial complex are checked by severalligaments that span the intramandibularjoint.
The distal tips of the dentaries are round-ed and do not contact one another anteriorly(Fig. 3B). However, Meckel’s cartilage,which persists throughout nearly the entirelength of the lower jaw, provides a tightlinkage between the mandibular rami. Theleft and right halves of this cartilage exitthe distal tips of the dentaries, curve dor-somedially, and fuse together to form a ro-bust cartilaginous nodule at the midline.This cartilaginous linkage severely limitsboth lateral separation and independent an-

1326 NATHAN J. KLEY
teroposterior excursions of the tips of themandibular rami, but allows extensive ro-tation between the left and right anteriormandibular segments. In addition, this nod-ule of cartilage between the tips of the man-dibular rami serves as the site of origin forthe extraordinarily robust median tendon ofthe M. genioglossus. A large slip of thismuscle extends caudally beyond the lingualsheath to insert onto the posteriorly posi-tioned, Y-shaped hyoid which, on average,is located between the 14th and 19th pre-caudal vertebrae in Leptotyphlops dulcis.This hyoid portion of the M. genioglossus(a likely homologue of the M. mandibulo-hyoideus II of lizards) is unique to Lepto-typhlopidae among snakes (Langerbartel,1968; Groombridge, 1979).
Feeding mechanics and behavior. Whenpresented with ant larvae and pupae, indi-viduals of Leptotyphlops dulcis quickly be-gan to exhibit characteristic frenzied feed-ing behavior. The snakes began to feverish-ly sway their heads and necks from side toside in an attempt to locate potential fooditems. Once the snout contacted an ant larvaor pupa, the snake would slide the ventralsurface of its snout over the top of the preyuntil the prey item was positioned at or nearthe front of the mouth. In some instances,the snake would actually pin its preyagainst the substrate with its snout prior toinitiating ingestion, but this was not a nec-essary pre-requisite for successful inges-tion. Once the snake had positioned itsmouth near the prey, it would then open itsmouth widely and grasp the prey betweenits jaws.
In all feedings that were observed, Lep-totyphlops dulcis ingested and transportedant larvae and pupae using a rapid mandib-ular raking mechanism (Kley and Brainerd,1999). Immediately following mouth open-ing, the snakes initiated bilaterally synchro-nous cycles of lower jaw flexion in whichthe toothed anterior mandibular segmentswere rapidly (2–3 Hz) rotated in and out ofthe mouth to drag prey into and through theoral cavity (Fig. 3B). This mandibular rak-ing mechanism involved complex move-ments at the intramandibular joints. Eachanterior mandibular segment (dentary 1splenial) was rotated medially in a horizon-
tal plane about the splenial-angular (intra-mandibular) joint. As a result of this move-ment, both the intramandibular joints andthe distal ends of the quadrates were forcedlaterally. At the same time, however, eachanterior mandibular segment was also ro-tated caudally about its own long axis (Fig.3B, right). The combined result of thesemovements was that the transversely ori-ented dentary tooth rows were rotated cau-dally into the mouth, thereby dragging theprey toward the snake’s esophagus.
The rapid jaw movements associatedwith ingestion and prey transport in Lep-totyphlops were usually augmented by syn-chronized movements of the anterior por-tion of the trunk. During jaw protraction,the neck was flexed slightly in the verticalplane, forming a shallow arch over the sub-strate. Then, during subsequent jaw retrac-tion, the neck was straightened, therebypushing the braincase anteriorly as the low-er jaw was pulling the prey caudally. Thus,as in alethinophidian snakes, movements ofthe anterior trunk supplement jaw move-ments during prey transport in Leptotyph-lops. In contrast to the concertina-likemovements seen in alethinophidians (Cun-dall, 1995; Kley and Brainerd, 1996; Moon,2000), however, axial bending during preytransport in Leptotyphlops is restricted tothe vertical plane. Finally, it should be em-phasized that while the ventroflexion asso-ciated with prey transport in Leptotyphlopsmay result in the prey being forced againstthe substrate, this is neither a necessary nora common aspect of feeding behavior inthese snakes (contra Reid and Lott, 1963).
Once the prey was transported to the rearof the mouth, Leptotyphlops dulcis initiatedswallowing with a forceful compression ofthe pharynx. After the prey was forced intothe esophagus through pharyngeal com-pression, swallowing appeared to occur en-tirely through peristalsis. No axial bendingwas observed during swallowing.
Muscular control of jaw movements.Leptotyphlopid snakes are among the mosthighly miniaturized tetrapods. Even in thelargest species of Leptotyphlops, maximumhead diameter rarely exceeds 5 mm. Thus,electromyography of head muscles in thesesnakes was not feasible. However, micro-

1327FEEDING IN BLINDSNAKES
manipulations of both fresh and cleared andstained specimens provided useful, albeitlimited, insight into the muscular control ofthe mandibular raking mechanism.
The results of these manipulationsstrongly suggest that jaw retraction in Lep-totyphlops is powered primarily by the longand robust M. genioglossus, pars hyoidea,which runs from the cartilaginous nodulebetween the distal tips of the mandibularrami to the posteriorly positioned hyoid ap-paratus. Indeed, by applying a caudally di-rected force to the tendon of the M. genio-glossus, the movements of the anteriormandibular segments that were observedduring feeding were replicated almost pre-cisely. In contrast, jaw protraction in Lep-totyphlops appears to be powered mainly bythe M. geniohyoideus. This complex strapmuscle has a lateral head, originating fromthe lateral surface of the trunk muscles inthe cervical region, and a medial head, orig-inating from the hyoid. Both heads con-verge to insert via tendons onto the ventro-posterior surface of the dentary. The actionof the M. geniohyoideus is to rotate the an-terior mandibular segments laterally aboutthe intramandibular joints and also rostrallyabout their own long axes, thus rotating thedentary tooth rows outwards to their restingposition. It is likely, however, that contrac-tion of the M. intermandibularis complexand elastic recoil of Meckel’s cartilage alsocontribute to jaw protraction.
Typhlops and Rhinotyphlops
Morphology of the jaw apparatus. As forLeptotyphlops, only a brief description ofthe typhlopid jaw apparatus will be pre-sented here. More comprehensive treat-ments of the cranial anatomy of other ty-phlopid species are provided by Haas(1930), Smit (1949), List (1966) and Ior-dansky (1997).
In nearly every major respect, the mor-phology of the jaw apparatus of Typhlopslineolatus was found to agree completelywith that of Rhinotyphlops schlegelii. How-ever, the jaw morphology of both of thesetaxa differs profoundly from that of Lep-totyphlops dulcis. Most significantly, thelower jaws of Typhlops and Rhinotyphlopsare toothless and relatively rigid, whereas
the upper jaw arches bear teeth and arehighly mobile.
In both Typhlops and Rhinotyphlops, themandible is much longer than in Leptotyph-lops, and is suspended from the braincasevia the relatively short quadrates (Fig. 4A).The dentary is greatly reduced and lacksteeth, a condition that is unique to typhlo-pids among snakes. As in Leptotyphlops,the left and right halves of Meckel’s carti-lage are bound together by an expanded in-terramal cartilaginous nodule, an arrange-ment which prevents independent move-ments of the left and right mandibular rami.However, in sharp contrast to the conditionseen in Leptotyphlops, there is extensivecontact between the relatively long splenialand the large, triangular coronoid. Thus, theintramandibular joint is bridged completelyby the splenial (Fig. 4A), and consequently,the lower jaws of Typhlops and Rhinotyph-lops are relatively rigid and akinetic.
In contrast to the lower jaw, the upperjaws of both Typhlops and Rhinotyphlopsare exceptionally kinetic. The maxillae liehorizontally against the roof of the mouthwith their transversely oriented tooth rowsdirected posteriorly (Fig. 4A, B, C), and aresuspended largely by ligaments and mus-cles rather than through bony articulations.In particular, a robust ligament runningfrom the canaliculate anterior end of themaxilla to the posterolateral margin of thepremaxilla (the premaxillo-maxillary liga-ment) anchors the anterior end of the max-illa (Fig. 4A, C). Posteriorly, the maxilla issuspended by the M. retractor maxillae, alarge muscle that takes its origin from thelateral surface of the braincase and insertsonto the posterodorsal surface of the max-illa.
The palatine is the only bone that con-tacts the maxilla directly. This bone has acomplex arched shape, with a medial pro-cess that articulates with the posterolateralmargin of the vomer (Fig. 4C), and a lateralprocess that curves ventrally to insert intoa shallow groove on the dorsal surface ofthe maxilla (Fig. 4B). Near the midpoint ofthe palatine, there is a slight ventral processthat is embraced by the forked anterior endof the pterygoid (Fig. 4C). As in Lepto-typhlops, the pterygoid in Typhlops and

1328 NATHAN J. KLEY
FIG. 4. Cranial morphology and jaw mechanics in Typhlops lineolatus. Drawings were made from a clearedand stained adult specimen. A. Left lateral view of the skull. Note the horizontally positioned maxillae and theabsence of teeth on the lower jaw. B. Dorsal view of the skull, showing the sliding articulation between thelateral process of the palatine and the dorsal surface of the maxilla. C. Ventral view of the skull, showing thearticulation between the medial process of the palatine and the posterolateral edge of the vomer. Note the robustpremaxillo-maxillary ligaments which anchor the anterior ends of the maxillae. D. Ventral view of the skullduring maxillary raking, showing asynchronous retraction of the maxillae. The lower jaw has been removed topermit an unobstructed view of the upper jaws. Abbreviations: c, coronoid; cb, compound bone; d, dentary; m,maxilla; p, palatine; pml, premaxillo-maxillary ligament; pt, pterygoid; q, quadrate; s, splenial; v, vomer.
Rhinotyphlops is a long and slender bonethat is widely separated from the quadrate(Fig. 4C). Thus, the upper and lower jawsare functionally decoupled as in Leptotyph-lops. In both Typhlops and Rhinotyphlops,the pterygoid is suspended from the ventralsurface of the braincase by the robust M.protractor pterygoidei, which originatesfrom the ventral surface of the braincaseand courses posteroventrally to ensheathethe posterior end of the pterygoid.
Feeding mechanics and behavior. LikeLeptotyphlops, Typhlops and Rhinotyphlopsexhibited somewhat frenzied feeding be-havior when presented with ant brood.Soon after a snake detected the presence ofprey (presumably via chemosensory cuesobtained through tongue-flicking), it initi-
ated a tactile search for individual preyitems by rapidly sweeping its head fromside to side in an erratic fashion as it pro-gressed forward. Once the snake’s headcontacted an ant larva or pupa, the snakemaneuvered its head so that its mouth waspositioned directly over the prey. The snakewould then open its mouth to initiate cap-ture and ingestion.
In all feedings that were observed, Ty-phlops lineolatus and Rhinotyphlops schle-gelii ingested and transported their prey us-ing a rapid maxillary raking mechanism. Asthe mouth was opened over an ant larva orpupa, the snake initiated ratcheting move-ments of the upper jaws in which thetoothed maxillae were rapidly (3–5 Hz) ro-tated in and out of the mouth. These max-

1329FEEDING IN BLINDSNAKES
illary raking movements served to drag theprey caudally into the mouth and towardsthe throat.
The jaw movements associated withmaxillary raking in Typhlops and Rhino-typhlops were more variable than those in-volved in the mandibular raking mechanismof Leptotyphlops. Most frequently, both in-gestion and intraoral transport were char-acterized by asynchronous jaw movementsin which the left and right maxillae wereprotracted and retracted slightly out ofphase with one another (Fig. 4D). In someinstances, however, only a single maxillawas used during ingestion. This generallyoccurred when a snake was attempting toingest an ant larva or pupa that was posi-tioned to one side of its mouth rather thandirectly below it, or when a snake was try-ing to extract prey from a narrow, confinedspace.
Movements of the lower jaw were alsosomewhat variable. In the majority of feed-ings observed, ingestion was completedwith a single maxillary retraction. In suchinstances the lower jaw was abducted dur-ing maxillary protraction and then adductedduring maxillary retraction. However, in-gestion of larger prey required several max-illary protraction-retraction cycles. In theseinstances, the lower jaw remained partiallyadducted throughout most of the maxillaryprotraction phase, presumably to preventthe advancing maxillae from pushing theprey back out of the mouth. Finally, duringthe rapid ingestion of large numbers of rel-atively small prey, movements of the lowerjaw were largely independent of upper jawmovements. Under these circumstances, thelower jaw generally remained abducted asthe maxillae continued to rake prey into themouth. In this manner, multiple prey itemswere often ingested simultaneously.
It should be emphasized that no appre-ciable protraction of the lower jaw was everobserved in either Typhlops lineolatus orRhinotyphlops schlegelii, and the mandiblewas never used to scoop prey off of thesubstrate as predicted by Iordansky (1981,1990, 1997). However, prey items that wereinitially positioned unfavorably for efficientingestion were in some instances reorientedby the mandible. During maxillary retrac-
tion, prey were frequently flipped up overthe anterior margin of the lower jaw, an ac-tion that tended to align the long axis of theprey with that of the snake’s head.
Unlike the smaller Leptotyphlops, Ty-phlops and Rhinotyphlops often ingestedmultiple prey items before swallowingthem. Once the pharynx was filled, thesnake initiated swallowing with a forcefulcontraction of the throat muscles immedi-ately ventral to the pharynx, followed bywavelike muscular contractions propagatedalong the ventral surface of the neck. Theprey was then rapidly propelled along theremainder of the esophagus, presumably viaperistalsis. Like Leptotyphlops, Typhlopsand Rhinotyphlops exhibited no axial bend-ing during swallowing.
Muscular control of jaw movements.Several previous studies have analyzed thepotential movements of the jaw apparatusin typhlopids and their muscular control(Haas, 1930; Evans, 1955; Iordansky,1997). However, all of these studies werebased exclusively on anatomical evidence.My observations of feeding in live Ty-phlops lineolatus and Rhinotyphlops schle-gelii, combined with the brief accounts ofThomas (1985) and Webb and Shine(1993b) for Typhlops richardi and Rampho-typhlops nigrescens, respectively, providesome corroboration of the functional hy-potheses generated through these morpho-logical investigations.
High-speed video recordings of Typhlopslineolatus reveal that the tooth-bearing pos-terior ends of the maxillae are rotatedthrough an arc of greater than 908 duringjaw protraction. Given that neither of themuscles inserting onto the maxillae (M. re-tractor maxillae and M. pterygoideus) haveorigins that are anterior to the maxillae, theobserved rotations of these elements are in-ferred to be produced indirectly throughmovements of the pterygoid and palatinebones. As noted by Evans (1955), the ap-plication of an anteriorly directed force tothe pterygoid causes the arched palatine topivot about its medial connection with thevomer, resulting in the lateral process of thepalatine being displaced anteriorly and ven-trally, thereby erecting the maxilla. Theonly muscle capable of producing this an-

1330 NATHAN J. KLEY
terior translation of the pterygoid is the M.protractor pterygoidei, which originatesfrom the ventral surface of the braincaseand courses posteroventrally to ensheathethe caudal end of the pterygoid. Thus, assuggested by previous authors (Haas, 1964;Cundall and Rossman, 1993; Iordansky,1997), erection of the maxilla appears tooccur through the action of the M. protrac-tor pterygoidei.
My observations of the cranial muscula-ture of Typhlops lineolatus and Rhinotyph-lops schlegelii, together with those of Haas(1930) for T. punctatus, T. lumbricalis andRamphotyphlops bituberculatus and thoseof Iordansky (1997) for T. lumbricalis andT. vermicularis strongly suggest that theprimary retractor of the upper jaw is the M.retractor maxillae. This is a very largemuscle of uncertain homology (Lakjer,1926; Haas, 1930, 1973) that is unique toTyphlopidae. It has a broad origin over thelateral surface of the braincase and insertsonto the posterodorsal portion of the max-illa. While other muscles are likely to par-ticipate in upper jaw retraction (e.g., M.pterygoideus and M. retractor pterygoidei),the size, position and fiber orientation of theM. retractor maxillae, together with its di-rect insertion onto the maxilla, suggest thatit is largely responsible both for retractingthe upper jaw and rotating the maxilla backto its resting position.
DISCUSSION
Although the foraging strategies (Wat-kins et al., 1967; Gehlbach et al., 1971;Webb and Shine, 1992) and dietary habits(Punzo, 1974; Webb and Shine, 1993a;Webb et al., 2000; Torres et al., 2000) ofscolecophidian snakes have been studied insome detail, the feeding behavior of thesediminutive serpents has been known onlyfrom a few brief accounts (Smith, 1957;Reid and Lott, 1963; Thomas, 1985; Webband Shine, 1993b; Kley and Brainerd,1999). The actual mechanisms by whichblindsnakes capture, ingest, transport andswallow their prey have remained largelyunknown (Cundall and Rossman, 1993;Greene, 1997). However, previous studiesof the cranial morphology of scolecophidi-ans have proposed three different feeding
mechanisms that might be used by thesesnakes: 1) suction feeding (Haas, 1964,1968; Groombridge, 1979); 2) lingual trans-port (McDowell, 1972); and 3) gnathic(jaw-based) transport (Haas, 1930, 1962,1964, 1968; Iordansky, 1981, 1997). Nei-ther suction feeding nor lingual feedinghave ever been documented in any snakeand I found no evidence for either in Lep-totyphlops, Typhlops or Rhinotyphlops. In-stead, all three species observed during thecourse of the present study used their jawsto capture, ingest and transport their insectprey. However, both the mandibular rakingmechanism of Leptotyphlops and the max-illary raking mechanism of Typhlops andRhinotyphlops differ significantly from thejaw ratcheting mechanisms of alethinophi-dian snakes.
Mandibular raking
The mandibular raking mechanism ofLeptotyphlops differs profoundly from thefeeding mechanisms of other snakes in thatprey is transported by bilaterally synchro-nous movements of the lower jaw ratherthan by independent movements of the up-per jaws. Among the more than 2,500 liv-ing species of snakes, mandibular transportmechanisms are known elsewhere only in asmall number of cochleophagous (snail-eat-ing) colubrids of the subfamilies Dipsadi-nae and Pareatinae (Cundall and Greene,2000). In these taxa, the mandible is usedto pull snails from their shells. However,the snail extraction mechanisms used bythese highly specialized colubrids involveunilateral ratcheting movements of the leftand right mandibular rami (Sazima, 1989)and thus bear little resemblance to the bi-laterally synchronous mandibular rakingmechanism of Leptotyphlops. Furthermore,bilaterally synchronous jaw movements aregenerally restricted to prey capture in ale-thinophidians (e.g., Frazzetta, 1966; Kar-dong, 1974; Cundall, 1987; Cundall andDeufel, 1999). Only rarely do such move-ments occur during prey transport, andwhen they do, it is only during the transi-tion between intraoral transport and swal-lowing, a period during which the prey ismoved not by the jaws, but by concertina-like movements of the trunk in the cervical

1331FEEDING IN BLINDSNAKES
region (Cundall, 1995; Kley and Brainerd,1996; Kardong and Berkhoudt, 1998).
The muscle activity patterns associatedwith mandibular raking in Leptotyphlopsremain completely unknown. Given the bi-laterally synchronous jaw movements thatcharacterize this mechanism, it seems likelythat bilateral activation of the hypoglossalmuscles powering jaw retraction (M. genio-glossus, pars hyoidea) and protraction (M.geniohyoideus) is involved. Regardless ofmuscle activity patterns, however, move-ments of the lower jaw in Leptotyphlops aremorphologically constrained to be bilater-ally synchronous. As in typhlopids (Bellairsand Kamal, 1981; Young, 1998), the leftand right halves of Meckel’s cartilage inLeptotyphlops extend beyond the distal tipsof the mandibular rami and fuse together toform a robust interramal nodule of carti-lage. This cartilaginous link between thetips of the mandibular rami permits hinge-like rotations between the dentaries, but al-lows almost no lateral or anteroposteriorseparation of the mandibular tips. In addi-tion, during jaw retraction, caudally direct-ed forces generated through contraction ofthe M. genioglossus are distributed evenlybetween the left and right distal mandibularsegments because this muscle originatesfrom a median tendon that arises from theinterramal cartilaginous nodule. For thesereasons, independent movements of the leftand right halves of the lower jaw are notpossible in Leptotyphlops.
Maxillary raking
Like mandibular raking in Leptotyphlops,maxillary raking in Typhlops and Rhino-typhlops represents a highly specializedmechanism for the rapid ingestion andtransport of large numbers of small insectprey. In contrast to mandibular raking, how-ever, maxillary raking bears at least somesimilarity to the feeding mechanisms of al-ethinophidian snakes. In particular, prey istransported into and through the mouth viaindependent ratcheting movements of theupper jaws. However, many more differ-ences than similarities can be found be-tween the feeding mechanisms of typhlo-pids and alethinophidians.
Perhaps the most striking differences be-
tween the feeding mechanisms of typhlo-pids and alethinophidians relate to the mor-phology and function of the upper jaws inthe two groups. The vast majority of ale-thinophidian snakes have a continuous rowof relatively long, recurved teeth along eachmedial upper jaw arch. In most taxa, recip-rocating translational movements of thesetoothed palatopterygoid arches are primar-ily responsible for transporting preythrough the oral cavity (Cundall andGreene, 2000). In typhlopids, however, thepterygoids and palatines are toothless andtranslational movements of the pterygoidsserve only to protract and retract thetoothed maxillae. Prey transport in typhlo-pids is thus brought about exclusivelythrough rotational movements of the highlymobile maxillae.
Other important differences between thefeeding mechanisms of typhlopids and al-ethinophidians relate to the structure of thelower jaw and its functional associationwith the upper jaws. In alethinophidiansnakes, the distal tips of the dentaries arequite separate from one another and arejoined together only by highly variable ar-rays of connective tissues (Young, 1998;Bellairs, 1984). Although the biomechani-cal properties of these interramal connec-tive tissues have not yet been critically ex-amined, in most taxa they permit consid-erable separation between the distal tips ofthe mandibular rami. Consequently, eachhalf of the lower jaw can be protracted orretracted independently of the other. Fur-thermore, due to a ligamentous connectionbetween the caudal tip of the pterygoid andthe quadratomandibular joint (the pterygo-quadrate ligament), unilateral movementsof the upper and lower jaws on each sideof the head are at least partially coupled toone another in most taxa (e.g., Albright andNelson, 1959b; Cundall and Gans, 1979).In contrast, the interramal connection in ty-phlopids is quite rigid. As noted previouslyby Bellairs and Kamal (1981), the twohalves of Meckel’s cartilage are bound to-gether in the interramal region by an en-larged cartilaginous nodule, thereby pre-venting independent movements of the leftand right mandibular rami. However, the ri-gidity of the lower jaw in typhlopids does

1332 NATHAN J. KLEY
not impede movement of the upper jaws.Due to the relatively wide separation be-tween the pterygoid and quadrate and theloss of the pterygo-quadrate ligament (Ior-dansky, 1997), the upper and lower jaws arefunctionally decoupled in Typhlopidae.
Finally, it is important to emphasize thatthe synchronization of jaw movements isdifferent in typhlopids and alethinophidi-ans. Alethinophidians generally use unilat-eral ratcheting movements of their jaws todrag themselves forward over their prey(Gans, 1961). The upper and lower jaws onone side of the head are opened and pro-tracted, and then closed and retracted.These movements are then mirrored by thecontralateral jaws. In contrast, typhlopidsmost commonly drag prey into and throughthe mouth using asynchronous movementsof the upper jaws. That is, the maxillae areprotracted and retracted slightly out ofphase with one another rather than in analternating, reciprocating pattern.
The neuromuscular control of maxillaryraking in typhlopids remains unexplored.Given the degree to which feeding kine-matics (especially patterns of maxillaryprotraction and mandibular abduction) aremodulated according to prey size and po-sition, it is likely that the motor patternsassociated with maxillary raking exhibitconsiderable variability as well. UnlikeLeptotyphlopidae, however, Typhlopidaeincludes a small number of ‘‘giant’’ species(e.g., Rhinotyphlops schlegelii, Typhlopspunctatus) that reach nearly 1 m in lengthand more that 2.5 cm in diameter (Fitz-Simons, 1962; Roux-Esteve, 1974). Largeadults of such species might afford the op-portunity to use electromyography to studythe activity of head muscles during feedingin these snakes. EMG data from typhlopidswould not only provide a more detailed un-derstanding of the maxillary raking mech-anism itself, but could also be comparedwith similar data from alethinophidiansnakes (e.g., Cundall and Gans, 1979; Kar-dong and Berkhoudt, 1998) and scleroglos-san lizards (e.g., Smith, 1982; Herrel et al.,1999) in an attempt to elucidate the modi-fications in motor control patterns that wereassociated with the shift to jaw-based preytransport mechanisms in the early evolution
of snakes. Finally, the sensory pathways in-volved in the modulation of feeding kine-matics in typhlopids remain unknown andare likely to represent a profitable line ofinquiry in future studies of typhlopid feed-ing. Given that ingestion in Typhlops andRhinotyphlops is usually initiated only afterthe snake’s snout has come into contactwith the prey, it seems likely that the integ-umentary mechanoreceptors present on thehead scales of typhlopids (Aota, 1940;Young and Wallach, 1998) represent an im-portant source of sensory feedback duringfeeding. Denervation experiments couldprovide insight into the role of these cuta-neous tactile organs in modulating feedingbehavior.
The origin of unilateral feeding in snakes
As discussed above, the leptotyphlopidmandibular raking mechanism and the ty-phlopid maxillary raking mechanism bothdiffer in significant ways from the snoutshifting and pterygoid walk mechanisms ofanilioids and macrostomatans, respectively.In particular, neither mandibular raking normaxillary raking are characterized by theunilateral pattern of jaw displacement thatunderlies the feeding mechanisms of ale-thinophidians. Although feeding behaviorin the third scolecophidian family, Anom-alepididae, remains entirely unknown, theclose phylogenetic relationship between an-omalepidids and typhlopids (Cundall et al.,1993; Tihen, 1945; Robb and Smith, 1966;List, 1966), and the similar form and posi-tion of the maxillae in these two families(List, 1966), suggest that anomalepididsalso transport prey using a maxillary rakingmechanism. Furthermore, many of the mor-phological features that facilitate unilateralfeeding in Alethinophidia (e.g., loose inter-ramal connection, kinetic snout, pterygo-quadrate ligament, etc.) are absent in An-omalepididae (Haas, 1964, 1968), as theyare in Leptotyphlopidae and Typhlopidae.Thus, available evidence suggests that,among extant snakes, unilateral feeding ispresent only in Alethinophidia.
When the feeding mechanisms of scole-cophidian snakes are placed within a phy-logenetic context (Fig. 5), two hypothesesemerge concerning the evolution of unilat-

1333FEEDING IN BLINDSNAKES
FIG. 5. The phylogenetic distribution of prey transport mechanisms in snakes. Phylogenetic hypothesis adaptedfrom Tchernov et al. (2000) and Cundall et al. (1993). Fossil taxa are excluded.
eral feeding: (1) unilateral feeding arose inthe common ancestor of Anilioidea andMacrostomata (i.e., within Alethinophidia);or (2) unilateral feeding arose in the com-mon ancestor of Scolecophidia and Alethin-ophidia (i.e., it is primitive for Serpentes),and was subsequently lost in Scolecophidia.It is clear that the feeding mechanisms ofblindsnakes are highly derived and doubt-less differ considerably from those of an-cestral snakes. Therefore neither of thesetwo hypotheses can be strongly rejectedbased on the morphological and behavioraldata that are currently available. However,when the criterion of parsimony is used toevaluate these hypotheses, the first scenariois favored over the second. For this reason,I accept the hypothesis that unilateral feed-ing evolved within Alethinophidia (Fig. 5).
Why unilateral feeding mechanisms areunique to Alethinophidia remains unknown.Perhaps the most conspicuous difference infeeding biology between Scolecophidia andAlethinophidia relates to the prey on whichthese snakes feed. Scolecophidians feedpredominantly on social insects. In mostspecies for which detailed dietary infor-mation is available, ant larvae and pupaerepresent the most important food resource(Webb and Shine, 1993a; Webb et al.,2000). In some species, termites are alsofrequently consumed (Punzo, 1974; Bratts-
trom and Schwenkmeyer, 1951). In con-trast, most alethinophidians feed on rela-tively large vertebrate prey (Greene, 1983,1997). It might therefore be inferred thatunilateral transport evolved as an adaptationfor handling large and potentially danger-ous prey, as this mode of transport ensuresthat a constant grip is maintained on strug-gling prey as it is ratcheted through themouth. Indeed, many non-venomous colu-broid snakes ingest and transport their preywhile it is still alive, and in the case of largecolubrids such as Drymarchon, formidableprey such as large rats and small rabbits canbe eaten safely in this manner (unpublishedobservation, N.J.K.). However, constrictionappears to have arisen very early in ale-thinophidian evolution (Greene and Bur-ghardt, 1978) and basal alethinophidiansrarely ingest their prey without first killingit through suffocation. Thus, it appears un-likely that unilateral transport arose as anadaptation for the transport of live, strug-gling prey.
It might also be hypothesized that theunilateral pattern of jaw displacement seenin alethinophidians arose simply as a by-product of the structural modifications ofthe skull associated with the ingestion oflarge-diameter prey in these snakes. For in-stance, the liberation of the mandibular tipshas long been recognized as an important

1334 NATHAN J. KLEY
morphological innovation in the early evo-lution of snakes that served to increase po-tential gape size (Gans, 1961). However,loosening of the interramal linkage not onlyallowed the tips of the mandibular rami tospread apart laterally, but also permitted theleft and right halves of the lower jaw to beprotracted and retracted independently ofone another. Therefore, if mandibular lib-eration occurred as a result of selection forincreased gape size, its role in unilateraltransport would be interpreted as being ex-aptive rather than adaptive (sensu Gouldand Vrba, 1982). Like the adaptive hypoth-esis presented above, however, this exaptivehypothesis seems unlikely in the light ofavailable evidence. First, the evolution ofincreased gape size in Alethinophidia hasoccurred primarily through structural mod-ifications of the lower jaw, suspensoria andintermandibular soft tissues (Gans, 1961;Lee et al., 1999; Cundall and Greene,2000). In contrast, many of the modifica-tions in cranial morphology that facilitateunilateral feeding have occurred in the up-per jaws, and therefore have had little effecton gape size. Second, although nearly allalethinophidians feed on relatively massiveprey, the ability to ingest exceptionallylarge-diameter prey is restricted primarilyto certain clades within Macrostomata (e.g.,Viperidae, Pythonidae, Boidae, etc.); basalalethinophidians feed predominantly on rel-atively narrow, elongate prey such as eels,caecilians, amphisbaenians, other snakes,and earthworms (Greene, 1983; Rajendran,1985). Thus, unilateral transport evolvedbefore many of the structural modificationsassociated with increased gape size arosewithin Macrostomata.
While the adaptive significance of uni-lateral transport remains unclear, its appar-ent absence in Scolecophidia suggests thatit is a uniquely alethinophidian innovation.Future studies of the evolution of unilateralfeeding mechanisms in snakes should there-fore be focused on functional comparisonsof the feeding apparatus in basal alethino-phidians and closely related scleroglossanlizards. In particular, electromyographicstudies using bilateral electrode placementwould be especially valuable in assessingthe degree to which motor patterns became
modified in association with the shift to aunilateral jaw displacement pattern.
ACKNOWLEDGMENTS
I am grateful to E. L. Brainerd for hersupport, encouragement and assistancethroughout this project. W. E. Bemis, K.Schwenk and E. J. Hilton provided valuableguidance during the preparation of histolog-ical sections. F. A. Jenkins and A. W.Crompton kindly permitted me to use theradiographic facility at the Museum ofComparative Zoology. For the loan of spec-imens used for comparative purposes, Ithank J. Hanken and J. P. Rosado (Museumof Comparative Zoology) and H. K. Vorisand A. Resetar (Field Museum of NaturalHistory). E. L. Brainerd, K. Schwenk andW. E. Bemis critically reviewed the manu-script. The lab in which this work was donewas supported by NSF IBN-9875245 to E.L. Brainerd.
REFERENCES
Abdeen, A. M., A. M. Abo-Taira, and M. M. Zaher.1991a. Further studies on the ophidian cranial os-teology: The skull of the Egyptian blind snakeLeptotyphlops cairi (Family Leptotyphlopidae). I.The cranium. A: The median dorsal bones, bonesof the upper jaw, circumorbital series and occipitalring. J. Egypt. Germ. Soc. Zool. 5:417–437.
Abdeen, A. M., A. M. Abo-Taira, and M. M. Zaher.1991b. Further studies on the ophidian cranial os-teology: The skull of the Egyptian blind snakeLeptotyphlops cairi (Family Leptotyphlopidae). I.The cranium. B: The otic capsule, palate and tem-poral bones. J. Egypt. Germ. Soc. Zool. 5:439–455.
Abdeen, A. M., A. M. Abo-Taira, and M. M. Zaher.1991c. Further studies on the ophidian cranial os-teology: The skull of the Egyptian blind snakeLeptotyphlops cairi (Family Leptotyphlopidae). II.The lower jaw and the hyoid apparatus. J. Egypt.Germ. Soc. Zool. 5:457–467.
Albright, R. G. and E. M. Nelson. 1959a. Cranial ki-netics of the generalized colubrid snake Elapheobsoleta quadrivittata. I. Descriptive morphology.J. Morphol. 105:193–240.
Albright, R. G. and E. M. Nelson. 1959b. Cranial ki-netics of the generalized colubrid snake Elapheobsoleta quadrivittata. II. Functional morphology.J. Morphol. 105:241–292.
Aota, S. 1940. An histological study on the integumentof a blind snake, Typhlops braminus (Daudin),with special reference to the sense organs andnerve ends. J. Sci. Hiroshima Univ. 7(B):193–208.
Bellairs, A. d’A. 1984. Closing address, with com-ments on the organ of Jacobson and the evolutionof Squamata, and on the intermandibular connec-

1335FEEDING IN BLINDSNAKES
tion in Squamata. In M. W. J. Ferguson (ed.), Thestructure, development and evolution of reptiles,pp. 665–683. Zoological Society of London andAcademic Press, London. (Symp. Zool. Soc. Lon-don No. 52.)
Bellairs, A. d’A. and A. M. Kamal. 1981. The chon-drocranium and the development of the skull inRecent reptiles. In C. Gans and T. S. Parsons(eds.), Biology of the Reptilia, Vol. 11, Morphol-ogy F, pp. 1–263. Academic Press, New York.
Bock, W. J. and C. R. Shear. 1972. A staining methodfor gross dissection of vertebrate muscles. Anat.Anz. 130:222–227.
Boltt, R. E. and R. F. Ewer. 1964. The functional anat-omy of the head of the puff adder, Bitis arietans(Merr.). J. Morphol. 114:83–106.
Bramble, D. M. and D. B. Wake. 1985. Feeding mech-anisms of lower tetrapods. In M. Hildebrand, D.M. Bramble, K. F. Liem, and D. B. Wake (eds.),Functional vertebrate morphology, pp. 230–261.Belknap Press, Cambridge.
Brattstrom, B. H. and R. C. Schwenkmeyer. 1951.Notes on the natural history of the worm snake,Leptotyphlops humilis. Herpetologica 7:193–196.
Brock, G. T. 1932. The skull of Leptotyphlops (Glau-conia) nigricans. Anat. Anz. 73:199–204.
Cundall, D. 1983. Activity of head muscles duringfeeding by snakes: A comparative study. Amer.Zool. 23:383–396.
Cundall, D. 1987. Functional Morphology. In R. A.Seigel, J. T. Collins, and S. S. Novak (eds.),Snakes: Ecology and evolutionary biology, pp.106–140. McGraw-Hill, New York.
Cundall, D. 1995. Feeding behaviour in Cylindrophisand its bearing on the evolution of alethinophidiansnakes. J. Zool., London 237:353–376.
Cundall, D. and A. Deufel. 1999. Striking patterns inbooid snakes. Copeia 1999:868–883.
Cundall, D. and C. Gans. 1979. Feeding in watersnakes: An electromyographic study. J. Exp. Zool.209:189–208.
Cundall, D. and H. W. Greene. 2000. Feeding insnakes. In K. Schwenk (ed.), Feeding: Form,function and evolution in tetrapod vertebrates, pp.293–333. Academic Press, San Diego.
Cundall, D. and D. A. Rossman. 1993. Cephalic anat-omy of the rare Indonesian snake Anomochilusweberi. Zool. J. Linn. Soc. 109:235–273.
Cundall, D. and J. Shardo. 1995. Rhinokinetic snoutof thamnophiine snakes. J. Morphol. 225:31–50.
Cundall, D., V. Wallach, and D. A. Rossman. 1993.The systematic relationships of the snake genusAnomochilus. Zool. J. Linn. Soc. 109:275–299.
Dullemeijer, P. 1956. The functional morphology of thehead of the common viper Vipera berus (L.).Arch. Neerl. Zool. 11:386–497.
Elias, J. A., L. D. McBrayer, and S. M. Reilly. 2000.Prey transport kinematics in Tupinambis teguixinand Varanus exanthematicus: Conservation offeeding behavior in ‘chemosensory-tongued’ liz-ards. J. Exp. Biol. 203:791–801.
Evans, H. E. 1955. The osteology of a worm snake,Typhlops jamaicensis (Shaw). Anat. Rec. 122:381–396.
FitzSimons, V. F. M. 1962. Snakes of Southern Africa.Purnell and Sons, Cape Town and Johannesburg.
Frazzetta, T. H. 1962. A functional consideration ofcranial kinesis in lizards. J. Morphol. 111:287–320.
Frazzetta, T. H. 1966. Studies on the morphology andfunction of the skull in the Boidae (Serpentes).Part II. Morphology and function of the jaw ap-paratus in Python sebae and Python molurus. J.Morphol. 118:217–296.
Gans, C. 1961. The feeding mechanism of snakes andits possible evolution. Amer. Zool. 1:217–227.
Gans, C. 1969. Comments on inertial feeding. Copeia1969:855–857.
Gehlbach, F. R., J. F. Watkins, II, and J. C. Kroll. 1971.Pheromone trail-following studies of typhlopid,leptotyphlopid, and colubrid snakes. Behaviour40:282–294.
Gould, S. J. and E. S. Vrba. 1982. Exaptation—a miss-ing term in the science of form. Paleobiology 8:4–15.
Greene, H. W. 1983. Dietary correlates of the originand radiation of snakes. Amer. Zool. 23:431–441.
Greene, H. W. 1997. Snakes: The evolution of mysteryin nature. University of California Press, Berkeley.
Greene, H. W. and G. M. Burghardt. 1978. Behaviorand phylogeny: Constriction in ancient and mod-ern snakes. Science 200:74–76.
Groombridge, B. C. 1979. A previously unreportedthroat muscle in Scolecophidia (Reptilia: Serpen-tes), with comments on other scolecophidianthroat muscles. J. Nat. Hist. 13:661–680.
Haas, G. 1930. Uber das Kopfskelett und die Kau-muskulatur der Typhlopiden und Glauconiiden.Zool. Jahrb. Abt. Anat. Ontog. Tiere 52:1–94.
Haas, G. 1959. Bemerkungen uber die Anatomie desKopfes und des Schadels der Leptotyphlopidae(Ophidia), speziell von L. macrorhynchus Jan.Vierteljahresschr. Naturforsch. Ges. Zurich. 104:90–104.
Haas, G. 1962. Remarques concernant les relationsphylogeniques des diverses familles d’ophidiensfondees sur la differenciation de la musculaturemandibulaire. Colloq. Internat. Cent. Nat. Rech.Sci. 104:215–241.
Haas, G. 1964. Anatomical observations on the headof Liotyphlops albirostris (Typhlopidae, Ophidia).Acta Zool. 45:1–62.
Haas, G. 1968. Anatomical observations on the headof Anomalepis aspinosus (Typhlopidae, Ophidia).Acta Zool. 49:63–139.
Haas, G. 1973. Muscles of the jaws and associatedstructures in the Rhynchocephalia and Squamata.In C. Gans and T. S. Parsons (eds.), Biology of theReptilia, Vol. 4, Morphology D, pp. 285–490. Ac-ademic Press, New York.
Hanken, J. and R. Wassersug. 1981. The visible skel-eton. Funct. Photogr. 16:22–26, 44.
Herrel, A., J. Cleuren, and F. De Vree. 1996. Kine-matics of feeding in the lizard Agama stellio. J.Exp. Biol. 199:1727–1742.
Herrel, A., F. De Vree, V. Delheusy, and C. Gans.1999. Cranial kinesis in gekkonid lizards. J. Exp.Biol. 202:3687–3698.

1336 NATHAN J. KLEY
Humason, G. L. 1979. Animal tissue techniques. 3rded. W. H. Freeman and Co., San Francisco.
Iordansky, N. N. 1981. [Cranial kinetism and functionsof the jaw muscles in Typhlops vermicularis. Acontribution to the problem of origin of the Ty-phlopidae]. Zool. Zh. 60:97–108. (In Russian).
Iordansky, N. N. 1990. Evolution of cranial kinesis inlower tetrapods. Neth. J. Zool. 40:32–54.
Iordansky, N. N. 1997. Jaw apparatus and feeding me-chanics of Typhlops (Ophidia: Typhlopidae): A re-consideration. Russian J. Herpetol. 4:120–127.
Kardong, K. V. 1974. Kinesis of the jaw apparatus dur-ing the strike in the cottonmouth snake, Agkistro-don piscivorus. Forma et Functio 7:327–354.
Kardong, K. V. 1977. Kinesis of the jaw apparatus dur-ing swallowing in the cottonmouth snake, Agkis-trodon piscivorus. Copeia 1977:338–348.
Kardong, K. V. 1986. Kinematics of swallowing in theyellow rat snake, Elaphe obsoleta quadrivittata:A reappraisal. Jap. J. Herpetol. 11:96–109.
Kardong, K. V. and V. L. Bels. 2001. Functional mor-phology and evolution of the feeding apparatus insquamates. In H. M. Dutta and J. S. Datta Munshi(eds.), Vertebrate functional morphology, pp.173–219. Science Publishers, Inc., Enfield, NewHampshire.
Kardong, K. V. and H. Berkhoudt. 1998. Intraoraltransport of prey in the reticulated python: Testsof a general tetrapod feeding model. ZACS 101:7–23.
Kardong, K. V., T. L. Kiene, and V. Bels. 1997. Evo-lution of trophic systems in squamates. Neth. J.Zool. 47:411–427.
Kley, N. J. and E. L. Brainerd. 1996. Internal concer-tina swallowing: a critical component of alethin-ophidian feeding systems. Amer. Zool. 35:81A.
Kley, N. J. and E. L. Brainerd. 1999. Feeding by man-dibular raking in a snake. Nature 402:369–370.
Lakjer, T. 1926. Studien uber die Trigeminus-versorgteKaumuskulatur der Sauropsiden. C. A. Rietzel,Copenhagen.
Langerbartel, D. A. 1968. The hyoid and its associatedmusculature in snakes. Ill. Biol. Monogr. 38:1–156.
Lee, M. S. Y., G. L. Bell, and M. W. Caldwell. 1999.The origin of snake feeding. Nature 400:655–659.
List, J. C. 1966. Comparative osteology of the snakefamilies Typhlopidae and Leptotyphlopidae. Ill.Biol. Monogr. 36:1–112.
McDiarmid, R. W., J. A. Campbell, and T. A. Toure.1999. Snake species of the world. A taxonomicand geographic reference. Herpetologists’League, Washington.
McDowell, S. B. 1972. The evolution of the tongue ofsnakes, and its bearing on snake origins. In T.Dobzhansky, M. Hecht, and W. Steere (eds.), Evo-lutionary biology, Vol. 6, pp. 191–273. Appleton-Century-Crofts, New York.
McDowell, S. B. and C. M. Bogert. 1954. The system-atic position of Lanthanotus and the affinities ofthe anguinomorphan lizards. Bull. Am. Mus. Nat.Hist. 105:1–142.
Moon, B. 2000. The mechanics of swallowing and the
muscular control of diverse behaviours in gophersnakes. J. Exp. Biol. 203:2589–2601.
Punzo, F. 1974. Comparative analysis of the feedinghabits of two species of Arizona blind snakes,Leptotyphlops h. humilis and Leptotyphlops d.dulcis. J. Herpetol. 8:153–156.
Rajendran, M. V. 1985. Studies in uropeltid snakes.Madurai Kamaraj University, Madurai.
Reid, J. R. and T. E. Lott. 1963. Feeding of Lepto-typhlops dulcis dulcis (Baird and Girard). Herpe-tologica 19:141–142.
Rieppel, O. 1980. The evolution of the ophidian feed-ing system. Zool. Jahrb. Abt. Anat. Ontog. Tiere103:551–564.
Robb, J. and H. M. Smith. 1966. The systematic po-sition of the group of snake genera allied to An-omalepis. Nat. Hist. Misc. 184:1–8.
Roux-Esteve, R. 1974. Revision systematique des Ty-phlopidae d’Afrique Reptilia-Serpentes. Mem.Mus. Nat. Hist. Nat. Ser. A 87:1–313.
Sazima, I. 1989. Feeding behavior of the snail-eatingsnake, Dipsas indica. J. Herpetol. 23:464–468.
Schwenk, K. 1988. Comparative morphology of thelepidosaur tongue and its relevance to squamatephylogeny. In R. Estes and G. Pregill (eds.), Phy-logenetic relationships of the lizard families, pp.569–598. Stanford University Press, Stanford.
Schwenk, K. 2000. Feeding in lepidosaurs. In K.Schwenk (ed.), Feeding: Form, function and evo-lution in tetrapod vertebrates, pp. 175–291. Aca-demic Press, San Diego.
Smit, A. L. 1949. Skedelmorfologie en -kinese vanTyphlops delalandii (Schlegel). S. Afr. J. Sci. 45:117–140.
Smith, H. M. 1957. Curious feeding habit of a blindsnake, Leptotyphlops. Herpetologica 13:102.
Smith, K. K. 1982. An electromyographic study of thefunction of the jaw adducting muscles in Varanusexanthematicus (Varanidae). J. Morphol. 173:137–158.
Smith, K. K. 1984. The use of the tongue and hyoidapparatus during feeding in lizards (Ctenosaurasimilis and Tupinambis nigropunctatus). J. Zool.,London 202:115–143.
Smith, K. K. 1986. Morphology and function of thetongue and hyoid apparatus in Varanus (Varani-dae, Lacertilia). J. Morphol. 187:261–287.
Tchernov, E., O. Rieppel, H. Zaher, M. J. Polcyn, andL. L. Jacobs. 2000. A fossil snake with limbs. Sci-ence 287:2010–2012.
Thomas, K. 1983. A nitrocellulose embedding tech-nique for vertebrate morphologists. Herpetol. Rev.14:80–81.
Thomas, R. 1985. Prey and prey processing in blindsnakes of the genus Typhlops. Amer. Zool. 25:14A.
Tihen, J. A. 1945. Notes on the osteology of typhlopidsnakes. Copeia 1945:204–210.
Torres, J. A., R. Thomas, M. Leal, and T. Gush. 2000.Ant and termite predation by the tropical blind-snake Typhlops platycephalus. Insectes Soc. 47:1–6.
Watkins, J. F. II, F. Gehlbach, and R. S. Baldridge.1967. Ability of the blind snake, Leptotyphlops

1337FEEDING IN BLINDSNAKES
dulcis to follow pheromone trails of army antsNeivamyrmex nigrescens and N. opacithorax.Southwest. Nat. 12:455–462.
Webb, J. K. and R. Shine. 1992. To find an ant: Trail-following in Australian blindsnakes (Typhlopi-dae). Anim. Behav. 43:941–948.
Webb, J. K. and R. Shine. 1993a. Dietary habits ofAustralian blindsnakes (Typhlopidae). Copeia1993:762–770.
Webb, J. K. and R. Shine. 1993b. Prey-size selection,gape limitation and predator vulnerability in Aus-tralian blindsnakes (Typhlopidae). Anim. Behav.45:1117–1126.
Webb, J. K., R. Shine, W. R. Branch, and P. S. Harlow.2000. Life-history strategies in basal snakes: Re-production and dietary habits of the Africanthread snake Leptotyphlops scutifrons (Serpentes:Leptotyphlopidae). J. Zool., London 250:321–327.
Young, B. A. 1998. The comparative morphology ofthe intermandibular connective tissue in snakes(Reptilia: Squamata). Zool. Anz. 237:59–84.
Young, B. A. and V. Wallach. 1998. Description of apapillate tactile organ in the Typhlopidae. S. Afr.J. Zool. 33:249–253.