quality, boron distribution and floral
TRANSCRIPT

AN ABSTRACT OF THE THESIS OF
Nancy Woodworth Callan for the degree of Doctor of Philosophy
in Horticulture presented on December 15, 1976
Title: EFFECT OF BORON SPRAYS ON FRUIT SET, FRUIT
QUALITY, BORON DISTRIBUTION AND FLORAL
MORPHOLOGY OF 'ITALIAN' PRUNE (PRUNUS
DOMESTICA L.)
Abstract approved: / ■ ; Maxine M. Thompson
The 'Italian1 prune (Prunus domestica L. ) is a high-quality
cultivar grown widely in the Willamette Valley of Oregon despite its
tendency for erratic production. Cool temperatures following bloom
slow pollen tube growth and thus limit fruit set.
In this study, 'Italian' prune trees which were not deficient in
B by August mid-'shoot leaf analysis, the standard nutritional index,
responded to fall foliar B application with increased fruit set.
Both pre- and postharvest B sprays were effective in increasing
set. A prebloom spray, however, did not increase fruit set. Mid-
summer, or "blue" drop, was unaffected by B treatment.
In an attempt to determine the basis of the fruit set increase,
the B concentration of various tissues was determined. Boron con-
tent of dormant bud and spur tissues, flower buds, flowers, and

immature fruit flesh was increased by 50 to 200% by the fall B spray.
August mid-shoot leaf B levels were increased by fall B in only one
of the two years of the study.
Various morphological and physiological changes resulted from
the fall B spray. Size of flower buds and of flowers at anthesis, as
well as style and pedicel length, was decreased by the fall spray, and
bloom was delayed slightly. Germinability of pollen from fall B-
treated trees was reduced. B levels of neither the pollen nor of the
pistil had an effect on pollen tube growth in the style.
Polyphenoloxidase activity of acetone powders prepared from
immature fruit was increased in fruits which had received fall foliar
B. This was in contrast to the observation that fruit from fall B-
treated trees browned more slowly when cut than fruit from control
trees. A decrease in the incidence of internal browning of mature
fruit after fall foliar B the previous year was also observed.
Speculations are made about possible relationships between the
observed responses of 'Italian' prune to fall B and the role of B in
the metabolism of phenolic compounds and in auxin oxidation.

Effect of Boron Sprays on Fruit Set, Fruit Quality, Boron Distribution and Floral
Morphology of 'Italian' Prune (Prunus domestica L. )
by
Nancy Woodworth Callan
A THESIS
submitted to
Oregon State University
in partial fulfillment of the requirements for the
degree of
Doctor of Philosophy
Completed December 1976
Commencement June 1977

APPROVED:
Associate Professor of Horticulture in charge of major
Head of Deparyraent of Horticultu re
Dean of Graduate School
Date thesis is presented O-^* ) S - ) f 7(a
Typed by Mary Jo Stratton for Nancy Woodworth Callan

ACKNOWLEDGMENTS
I would like to thank my adviser, Dr. Maxine Thompson, for
her advice, moral support and friendship offered during this study.
I also thank the members of my graduate committee. Dr. Michael
Chaplin, Dr. Warren Kronstad and Dr. Fred Rickson for their help-
ful suggestions and aid. Consultation with and assistance by Dr. Mel
Westwood, Dr. Robert Stebbins and Dr. Daryl Richardson has been
invaluable, as has the help given me by Harold Bjornstad and Fred
Dixon. Finally, I would like to thank the members of the Oregon
Processed Prune and Plum Growers Commission for the financial
support which made this work possible.

TABLE OF CONTENTS
Page
INTRODUCTION 1
REVIEW OF LITERATURE 3
Fruit Set 3 Boron and Pollen 8 Boron Levels and Plant Response 11 Mobility of Boron within the Plant 13 Fruit Quality 14 Postulated Roles of Boron in the Plant 16
FRUIT SET OF 'ITALIAN' PRUNE AS AFFECTED BY SPRING AND FALL BORON SPRAYS 20
Introduction 21 Materials and Methods 22 Results and Discussion 26
BORON DISTRIBUTION IN 'ITALIAN' PRUNE TISSUES FOLLOWING SPRING AND FALL BORON SPRAYS 39
Introduction 40 Materials and Methods 40 Results and Discussion 43
THE EFFECT OF FALL FOLIAR BORON APPLICATION ON POLYPHENOLOXIDASE ACTIVITY AND INTERNAL BROWNING IN 'ITALIAN' PRUNE FRUIT 53
Introduction 54 Materials and Methods 55 Results and Discussion 57
BIBLIOGRAPHY 62

LIST OF TABLES
Table Page
1 Fruit set of 'Italian' prune, 1975, following postharvest and prebloom B sprays. 33
2 Fruit set of 'Italian' prune, 1976, following fall foliar B sprays. 34
3 Fruit set of 'Italian1 prune in six orchards in 1976 following 492 ppm B applied postharvest. 35
4 Germinability of 'Italian' prune pollen in deionized water with 10% sucrose following 492 ppm B postharvest. 36
5 Dry weight of flower buds from seven orchards collected at various stages of bud development after 492 ppm B applied postharvest. 37
6 Style and pedicel length of 'Italian' prune in two orchards after 492 ppm B postharvest. 38
7 Boron in 'Italian' prune dormant bud and spur tissue and in flower buds in six orchards after 492 ppm B postharvest. 48
8 Boron in 'Italian' prune dormant bud and spur tissue and in floral tissues in two stages of bud development and full bloom, 1976. 49
9 Boron in 'Italian' prune flower parts at full bloom and at petal fall, 1975, as affected by 492 ppm B applied postharvest 1974 and pre- bloom 1975. 50
10 Boron in 'Italian' prune mid-shoot leaves in August as affected by B application. 51
11 Boron in 'Italian' prune leaves from four locations on the tree in August 1976 as influenced by 492 ppm B applied postharvest, 1975. 52

Table Page
12 Polyphenoloxidase activity and B content of immature 'Italian' prune fruit flesh, July 1976, after 492 ppm B applied postharvest, 1975. 60
13 Yield and fruit quality at harvest, 1976, as affected by fall foliar B applications. 61

EFFECT OF BORON SPRAYS ON FRUIT SET, FRUIT QUALITY, BORON DISTRIBUTION AND FLORAL
MORPHOLOGY OF 'ITALIAN' PRUNE (PRUNUS DOMESTICA L. )
INTRODUCTION
The 'Italian, ' or 'Fellenberg, ' prune (Prunus domestica L. ) is
a high-quality cultivar for drying, canning and fresh market. The
fact that it does not consistently produce a full crop of fruit is a major
defect of this cultivar. Erratic cropping of 'Italian' is not confined to
the Willamette Valley of Oregon; reports from Russia (160, 161) and
Italy (118) also indicate this tendency. Thompson and Liu (166) found
'Italian' to be self-compatible and not to benefit from supplemental
hand or bee pollination under Willamette Valley conditions. Fruit set
failure was related to temperature. When temperatures averaged
10 Cor below during the 3-week postbloom period pollen tube growth
was delayed and ovule breakdown began before fertilization could
stimulate fruit set (166).
In 1972, a year of generally poor cropping for 'Italian,' M.H.
Chaplin (personal communication) found that a fall foliar B application
resulted in 100% greater yield from trees which were not deficient
according to August mid-shoot leaf analysis, the standard index of
nutrition. Research which ensued had these objectives:

1. To reaffirm the increased yield following a fall foliar B applica-
tion of 'Italian' prune trees with adequate B nutrition and to
elucidate the basis of this yield increase by taking fruit set counts
during fruit development.
2. To determine optimum timing relative to harvest and optimum
concentration of the fall B spray.
3. To discover the effect of fall foliar B on factors which could
influence fruit set such as pollen tube growth in the style, pollen
germinability and date of bloom.
4. To determine B levels in buds, flowers and flower parts the
spring after B application and in leaves the following August.
5. To determine the effect of the fall B spray on the fruit itself.
This included B levels and polyphenoloxidase activity in imma-
ture fruit and harvest quality factors of mature fruit.

REVIEW OF LITERATURE
Fruit Set
Fruit set is a complex phenomenon which can be affected by a
wide range of cultural and climatic conditions (46, 112, 188). Tem-
perature appears to be a major factor limiting fruit set in 'Italian'
prune, as stated in the introduction to this thesis. While low tem-
peratures prolong ovule longevity (64, 65) as well as retard pollen
tube growth rate, the effect on pollen tube growth appears most
influential in determining fruit set in plum (58, 167).
Pollen tube growth rate varies with species and cultivar as well
as with temperature. Plum pollen tube growth rate was reported to
be slower than that of apple or pear (58, 158). Almond cultivars also
varied in pollen tube growth rate (75). In 'Bartlett' pear, pollen tubes
reached the ovary in 4 days at 10oC and in only 2 days at 15.5 C
(105). In 'Anjou' pear, 5 and 3 days respectively, were needed at the
above temperatures (114).
Fruit set can be reduced by abnormalities in pollen or ovule.
Irregularities in embryo sacs and their degeneration were responsible
for poor fruit set in apricot (66). Since ovule breakdown began at the
chalazal end near the vascular connection in peach, Harrold (81) pro-
posed that vascular disruption was responsible for fruit drop.

Competition between fruits or between fruits and shoots can
influence fruit set. Competition between flowers was proposed by-
Bradbury (24) to be a possible cause of fruit set reduction in sour
cherry. She found that degeneration of aborting flowers commenced
before pollination. Reducing competition by thinning clusters to one
flower increased percent fruit set in this study and in another study
with 'Cornice' pear (183). At the time of initial fruit set, when both
fruit and shoots are drawing on nutrients stored in the tree, competi-
tion between fruit and shoot can also be a determining factor in fruit
set (22, 137, 177).
Transfer of pollen from anther to stigma is essential for fruit
set in 'Italian, ' which does not normally set fruit parthenocarpically.
Thompson and Liu (166) found that 'Italian' prune, a self-fertile culti-
var, did not benefit from supplemental hand pollination or from the
presence of bees. This indicated that adequate pollen transfer occurs
by wind and by anthers touching the stigma. Style length was impor-
tant in positioning the stigma and anthers for efficient self-pollination
(76) and length of style could also determine the length of time neces-
sary for the pollen tube to travel from stigma to ovary to fertilize the
egg. The rootstock to which the cultivar was grafted had a marked
influence on style length in plum (76), peach and apricot (55).
Growth regulators, both endogenous and applied, influence fruit
set (45, 46). Although plums do not generally set fruit without

fertilization, parthenocarpy has been induced by gibberellin in plum
(89) as well as in peach (48, 49), almond and apricot (48). Both
gibberellin and auxin were required for parthenocarpy in sweet cherry
(47). Plant hormones may also influence fruit set by directing the
movement of assimilates within the plant. Assimilates will be
mobilized toward areas of high auxin concentration (22, 142, 177).
Competition for nutrients between developing fruits and shoots can
occur. Schneider (137) has proposed that a NAA spray to the entire
tree thins apples by creating vegetative sinks which compete with the
fruit for assimilates. Auxin sprays will also thin stone fruits.
Another synthetic auxin, 3-CPA (3-chlorophenoxy propionamide) is
used to thin peaches, plums and apricots (M.N. Westwood, personal
communication). It appears that the balance between growth regulators
such as gibberellin and auxin at each stage of fruit set is as important
as hormone levels per se. For example, while gibberellin is present
in stone fruits at bloom (112), auxin concentration is not detectable
until after about 2 weeks after bloom (46, 127).
A delay in the time of bloom can increase fruit set under cer-
tain conditions since chances of favorable temperatures for fruit set
increase as the season progresses. Boron levels may influence the
time of bloom in fruit and nut trees. Boron excess has been shown
to delay bud break in peach (38, 50, 83, 95, 186) and apple (80). In
Georgia, bloom date of pecans was advanced by B toxicity (147) but

the authors felt that this was probably due to the effect of injury on
rest. Growth regulators may also delay bud break. Ethephon (2-
chloroethyl phosphonic acid), applied the previous fall, was found to
delay bloom in sweet cherry (54, 130), sour cherry (31) and plum
(54). SADH (succinic acid 2, 2-dimethylhydrazide) delayed bloom of
'Delicious' apple (159) and azalea (93), while gibberellic acid retarded
bud break of peach (43).
Mineral nutrition, especially of the elements nitrogen and
boron, can affect fruit set. Williams (187) found that cell division in
the nucellus of the ovule continued later in the spring in apple flowers
which had received a summer soil nitrogen application. Also, fruit
set the next spring was increased when compared with trees which had
not received late summer N. Westwood et al. (182) also found N to
be important for fruit set of pear.
Boron is important for both flower development and initial fruit
or seed set. Boron is needed by ovules in vitro as well as by intact
flowers (16, 120). Higher levels of B in the nutrient solution were
needed for reproductive growth than for vegetative growth of cotton
(85). The B levels.of both the pollen and pistil were related to
seed-setting ability in clover. When no deficiencies existed, the B
content of the pollen-bearing plant appeared to be more important
than that of the pistil-bearing plant (115). In contrast, the B content
of only the female parent was important for seed set in another study

involving clover (15) and in rape (82). A decrease in seed or fruit set
is often one of the first signs of B deficiency. Batjer et al. ( 8 ) found
that an incipient B deficiency of pear caused by an inability of tree
roots to take up B from cold soil in early spring causes blossoms
to wilt and die, or "blast." Vegetative growth is normal during the
remainder of the season but set may be reduced on affected trees.
Boron for flower and fruit development can be provided by a
spray or soil application. Preharvest, postharvest and prebloom
sprays increased B levels in 'Bartlett' pear flower bud clusters (44).
Both fall-and spring-applied sprays were effective in correcting the
"blossom blast" condition of pear described above (8, 92) although fall
sprays were most consistent (92). Spring-applied B sprays increased
'Anjou1 pear fruit set in one study (9) but not in another (53). Fruit
set of 'Magness' pear was not increased by a spring spray of B (135).
Spring-applied B sprays increased fruit set of 'Stayman' apple (25)
but not of other cultivars (25, 86). Fruit and seed set increases
were also observed after prebloom B sprays on grape (123) and after
soil applications or the addition of B to the nutrient solution in many
crops (15, 82, 115,. 181, 184).
Boron excess, on the other hand, may or may not reduce fruit
set. Apricot trees with marked symptoms of B injury have been
observed to bear normal crops but prune trees, while they may flower
profusely, are sometimes reduced in set by excess B (63).

Boron appears to be important for flower development rather
than for initiation of flowers. No influence of B levels above the
deficiency level was seen on flower initiation in cotton (85).
Boron and Pollen
Boron has been reported to enhance in vitro pollen germination
in many plants. Stimulation by B is not universal among plant species,
however. Species such as pear with high endogenous pollen B levels
generally gave a greater response to B in the germination medium
than those such as pine with low levels (151). Boron had no effect
on or inhibited pollen of Capsicum annuum (170), Crotalaria juncea,
and Corchorus capsularis (174) while concentrations up to 1200 ppm
were optimal for other species (173). Boron was reported to improve
in vitro pollen germination in pea (100), onion (98), cherry (17, 165),
apple and apricot (165), trumpet vine (124), eggplant (170), chrysan-
themum (168), pear (17, 149, 165) and Seta via sphacelata (52).
Thompson and Batjer (165) reported a stimulation of plum pollen by
B while Blaha and Schmidt (17) found plum and prune pollen did not
respond to the addition of B to the germination medium. However,
only two prune pollen samples were examined by Thompson and
Batjer, and germination was very low both in the control (6%) and
when B was added to the germination medium (10%).

9
Conflicting reports of B requirements for pollen germination
may be reconciled when plant B supply is considered. According to
Vasil (169, 170) pollen is generally deficient in B. He states that
. . . a. relatively reliable bioassay technique. . .for B can be developed by employing the pollen germination test. Plants which are deficient in B do not show good germina- tion of pollen but show considerably better germination after B has been supplied either through the roots or through cut ends of branches.
Pollen from B deficient pear trees was enhanced by the addition of B
to the germination medium but after 4 years of B fertilization it was
no longer necessary to add B to the medium (3). Likewise, response
of alsike clover (115) and Forsythia pollen (174) to the addition of B to
the germination medium was dependent on the plant's supply of B.
Boron sprays have also been reported to improve pollen germination.
Potted strawberry plants responded to boric acid applied to the flower
trusses by improved anther dehiscence and pollen germination (77).
Also, a foliar spray of B improved grape pollen germination (1Z3).
While endogenous or applied B appears necessary for optimum
pollen germination in many plants, an excess of B in the medium will
usually reduce pollen germinability. In most plants pollen germination
is enhanced by the addition of supra-optimal amounts of B to the
germination medium (171). Soil-induced B toxicity was reported to
reduce peach pollen germinability when compared with germination of
pollen from normal trees (95).

10
In contrast to the conflicting reports on the effect of B on pollen
germination, in nearly all plants studied pollen tube growth rate has
been accelerated by B in the germination medium. Visser (173)
stated that boric acid enhanced pollen tube growth rather than pollen
germination. Stanley and Loewus (152) agreed, adding that "pollen
tube membranes must be present for boron to exert a stimulatory
effect on the metabolism of glucose supplied exogenously to developing
pollen. "
The action of B in enhancing pollen tube growth may involve the
reaction of borate with compounds in the germination medium. Phe-
nolic compounds were shown to be deposited on the surface of the
pollen grain by the anther tapetum of Tulipa (162) and were found to
diffuse out of the pollen of several species into the germination medium
(150). Caffeic acid, a diphenol, inhibited pollen tube growth and
germination of several plant species (111). As B has been demon-
strated to form complexes with diphenols (192), it is possible that the
presence of B in the germination medium could prevent certain phe-
nolic compounds from inhibiting germination or pollen tube growth
in vitro.
Boron does not appear to influence pollen tube growth rate in the
style. The B content of neither the pollen nor the pistil was found to
influence the growth rate of clover pollen in the style (184).

11
Boron Levels and Plant Response
The standard technique of leaf analysis for determination of
plant mineral nutrition involves. sampling mid-shoot leaves in late
summer when levels of most elements fluctuate least. One of the first
workers to use mid-shoot leaves for B analysis was Burrell (34).
This practice arose out of convenience since these leaves were already
in use to index the nitrogen, magnesium and potassium status of fruit
trees. There is general agreement on the B levels desirable for opti-
mum growth in plum and prune trees. The Oregon State University
Fertilizer Guide for Prunes (153) states that 35-80 ppm B in mid-
shoot leaves is . optimal, below 25 ppm deficient and over 100 ppm
toxic. Leece (103) in Australia considers 25-60 ppm normal for plum
trees in midsummer with deficiency below 20 ppm. Hansen and
Proebsting (79) found that 'President' plum trees with 72-77 ppm B
in leaves showed injury. Leaf concentrations from 26-29 ppm were
found in both normal and deficient trees but below 25 ppm deficiency
symptoms were always found. Hernandez and Childers (83) found
toxicity symptoms in peach at levels of 100 ppm B or more.
It is difficult to specify B levels for optimum plant performance.
The greatest plant growth may occur at B concentrations which produce
mild toxicity symptoms on the leaves. Maximum plant response over-
lapped with onset of B toxicity symptoms when sunflower plants were
grown in nutrient solution (67). 'Valencia' orange trees suffered no

12
reduction in growth at B concentrations giving mild toxicity symptoms
(146). Optimum B concentration for reproductive growth may
generally be higher than for vegetative growth, as Montgomery (115)
found with alsike clover. Soybean growth was increased at B levels
producing leaf toxicity symptoms (42, 122). However, plant species
and cultivars vary widely in boron tolerance (68). Stone fruits are
less susceptible to boron toxicity than many other plants in that they
show fewer symptoms (179)- They also have an ability to divert B to
the stem and fruit (63).
Several studies have shown that the highest concentrations of B
occur in the reproductive organs. Boron accumulates in both male
and female tissues in apple (19), corn (21), lily (13), tobacco (14),
Qenothera (73), cherry, apple and pear (190, 191) and in buckwheat
(20). In Qenothera, the stigma contained higher B levels than the
style. In rye, more B was found in anthers and pollen than in female
tissues (104).
Seasonal variation in the B content of flower buds occurs.
Boron levels in buds change little during dormancy but as growth
begins in the spring B moves into the buds. At full bloom flowers
contained high concentrations of B (35, 74, 191). Pear, apple and
cherry fruit buds did not store sufficient B for their development but
drew on B from the soil or from reserves within the plant (191).

13
Mobility of Boron within the Plant
Many questions exist about the mobility of B within the plant
system. Boron has traditionally been grouped with the immobile
elements because deficiency symptoms first occur in the youngest
tissues (72). Under limited B supply most of the B was found in the
older, basal leaves of alfalfa (4, 56) and tomato (29). Boron did not
move out of senescing sugarcane leaves (116). Oertli and Richardson
(121) explained B immobility by a cyclic movement between leaf mar-
gin and leaf midrib and base, keeping B within the leaf. However,
some mobility of B within the plant has been reported. In broadbean
B was not normally transported to the radicle from cotyledons or the
epicotyl (117), but when cotyledons were provided with excess B it
was readily transported from this organ to the radicle (113). In
turnip (69), and in broccoli (12) B did not move out of the oldest
leaves of a plant but it was apparently translocated from younger but
mature leaves to apical tissues. Boron levels were similar in upper
and lower leaves in grapevines (140) and tobacco plants (176) deprived
of B. This would indicate redistribution within the plant. Stone
fruits are well known for their ability to transport B from the leaves
to the bark and then to the fruit under conditions of excess (62, 63).
When B was in limited supply the fruit exhibited B deficiency symp-
toms 'while the leaves contained adequate B (11). A loss of B from

14
the peach endocarp and a gain in flesh B occurred as fruits matured
(10).
Conflicting reports of B mobility can be more easily understood
when the solubility of the plant B is considered. Plants contain a
fixed amount of bound or insoluble B which remains constant under
conditions of varying B supply. There is little or no reutilization of
insoluble B. The soluble B content of plants, however, decreases as
B is withheld and drops to zero when the plants exhibit deficiency
symptoms (144). Differences in plant B solubility could be expected
among species and cultivars (Z, 113). Monocots, for example, have
a higher proportion of their B in soluble, or unbound form than dicots,
have a lower B requirement, and take longer to show B deficiency
symptoms (143). If the soluble B fraction, which is dependent on
plant B supply, is the only portion which is available for translocation,
conditions of plant B supply prior to an experiment as well as the
species or cultivar involved would have a considerable effect on
apparent B mobility.
Fruit Quality
Boron levels may affect fruit quality in various ways. Peaches
from trees suffering from B toxicity were insipid and flat in flavor (38,
186). Malformed peach fruits developed on trees given excess B
(83). Spring B sprays (25), as well as soil applications of borax (7),

15
reduced the storage life of apples. However, late summer B sprays
on 'Bartlett' pears had no effect on storage quality (44), nor did
spring sprays affect storage quality of 'Magness' pear (135). Foliar
sprays on 'Golden Delicious1 apple increased sugars and ascorbic
acid content (32). Boron application delayed maturity in 'Mclntosh'
(126) while 'Granny Smith1 apples remained green longer although
ripeness was not affected (30).
Boron sprays have also been useful in correcting some fruit
disorders. Spring B sprays reduced the incidence of bitter pit in
apple (60). Since calcium deficiency of the fruit has also been asso-
ciated with this disorder it is interesting to note Marsh's theory (108)
that B tends to keep Ca in an available condition. Boron sprays
applied in spring and summer did not decrease the amount of fruit
cracking, a disorder thought to be related to low calcium, in 'Italian'
prune (39)-
Internal browning of 'Italian1 prune is a complex physiological
disorder in which the fruit flesh near the pit breaks down and becomes
discolored (172). This disorder has shown variable response to B
applications. Spring B sprays reduced internal browning in one year
but not in another (172). A similar disorder of apricot was found to
be caused by B deficiency (33). Gibberellin sprays applied before
harvest to 'Early Italian' prune reduced internal browning (128, 129).
Summer applications of ethephon (2-chloroethyl phos phonic acid) and

16
SADH (succinic acid 2, 2-dimethylhydrazide) reduced browning in pro-
cessed peaches. Less discoloration of the pit cavity was found after
treatment with the above compounds alone or in combination (78).
Fresh or canned peaches from trees which had been treated with
SADH were found to be free from a benzaldehyde or "peach pit" odor
found in the control (141). SADH (36) and boron (101) have both been
proposed to direct glucose metabolism away from the pentose shunt,
from which phenolic acids arise.
Postulated Roles of Boron in the Plant
Many roles in plant metabolism have been proposed for B but
none have been established conclusively as its primary function.
Indeed, the ability of B to complex with hydroxyl-containing compounds
such as sugars, cellulose and polyphenols may allow it to enter into
many cellular associations. Briefly, some of the functions proposed
for B include carbohydrate translocation (70, 71, 72), starch-sugar
interconversions (59), cell wall metabolism (148), pectin synthesis
(189), plant osmotic balance (5), phosphate metabolism (133), calcium
metabolism (18, 94, 108), protein synthesis (175), synthesis of ATP
(132), RNA metabolism (90, 91), DNA metabolism (40), membrane
potentials (163), oxidative enzymes (107), metabolism of phenolic
compounds (1, 51, 101, 131), and regulation of auxin levels (41, 6l,
131, 145).

17
Since the developing flower bud is an area of active cell division
and growth, and since auxins may influence fruit set both alone and in
combination with other hormones, these aspects of B metabolism
will be considered.
Boron is extremely important for cell division and growth.
Several studies involving the early stages of B deficiency in roots
showed that cell division ceases at various intervals after B with-
drawal; in squash, 6.5 hours (40), in bean after 49 hours (185), and
in sunflower after 3 hours (97). In order to study DNA synthesis,
Cohen and Albert (40) examined tritiated thymidine uptake by squash
root meristems. Although mitotic figures were not seen after 6.5
hours without B, incorporation of label continued up to 20 hours.
When B was resupplied after B withdrawal, label uptake
resumed, indicating a temporary suspension of DNA synthesis during
the early period of deficiency.
Cessation of cell elongation is also an early feature of B
deficiency. Elongation stopped within 48 hours of B withdrawal in
Vicia faba (133) and after 6.5 hours in squash (40).
Boron appears to be involved in the metabolism of phenolic
compounds and, through these, auxin metabolism. The accumulation
of certain phenolic compounds in B deficiency has been well docu-
mented. Chlorogenic and caffeic acids accumulated in sunflower (51),
tomato (28), radish and lettuce (125); scopoletin and a glucose

18
derivative of gentisic acid in sunflower (178); and vanillic and ferulic
acids in oil palm (131).
Two explanations have been presented for the accumulation of
phenolic compounds in B deficiency. Rajaratnam and Lowry (131)
found a complete absence of one class of flavonoids, the leucoantho-
cyanins, in B-deficient tissues. They postulated that B was necessary
for leucoanthocyanin synthesis from phenolic precursors such as those
which accumulate in B deficiency. Gentisic acid is a simple phenolic
compound while the remainder of the above compounds represent
metabolic routes from cinnamic acid to compounds other than
flavonoids (134). Lee and Aronoff (101), on the other hand, presented
evidence for the role of B in partitioning glucose metabolism between
glycolysis and the pentose shunt. In B deficiency the pentose shunt is
favored and more phenolic acids are formed. Skol'nik (145) also felt
that fewer phenolic compounds were formed in the presence of B.
In 1968, Coke and Whittington (41) proposed that B deficiency
could be equivalent to a state of auxin toxicity. Boron deficiency
symptoms can be duplicated by auxin application (41, 97, 131, 133).
Auxin-like activity was high in extracts of 24-hour B deficient bean
root tips (41) and in extracts of primordia of oil-palm seedlings show-
ing little-leaf B deficiency symptoms (131).
Phenolic substances can be important in the regulation of IAA
oxidase (peroxidase) (138, 155, 157) and therefore in the control of
plant auxin levels (136, 164). In general, polyphenols, those with

19
ortho hydroxy groups, inhibit IAA oxidase while monophenols enhance
its activity (155). Chlorogenic and caffeic acids and scopoletin (87)
and ferulic and vanillic acids (138) inhibit IAA oxidase. Stonier et al.
(157) have isolated, "auxin protectors" of varying molecular weights.
These appear to be phenolic compounds which act as antioxidants
inhibiting the peroxidase-catalyzed oxidation of IAA. IAA was also
degraded in the presence of polyphenoloxidase and certain phenolic
substrates (27). The authors postulated that IAA was destroyed in a
reaction with intermediates of phenolic oxidation, since polyphenolox-
idase alone does not destroy IAA. Polyphenoloxidase was found to
eliminate "protector" activity (156).
In view of the abundance of phenolic compounds in Prunus
domestica (84) and the importance of auxin, both alone and in com-
bination with other hormones, for fruit set, this aspect of B meta-
bolism appears particularly relevant to a discussion of fruit set.
The remainder of the thesis is presented in the form of three
manuscripts, the first two to be submitted to J. Amer. Soc. Hort.
Sci, and the third to HortScience.

20
FRUIT SET OF 'ITALIAN' PRUNE AS AFFECTED BY SPRING AND FALL BORON SPRAYS1
Nancy W. Callan2
Department of Horticulture, Oregon State University Corvallis, Or. 97331
Additional index words. pollen, anthesis, Prunus domestica L.
Abstract. A fall foliar B application was found to increase
fruit set of 'Italian' prune (Prunus domestica L. ). A prebloom B
spray had no effect on fruit set. Neither fall nor spring applications
influenced the amount of fruit lost in the midsummer or "blue" drop.
All trees involved in the experiment had adequate B by the standard
index of tree nutrition, August mid-shoot leaf analysis. Incipient B
deficiency, in which cold, heavy soil prevents B uptake in early
spring, did not appear to be involved.
Several morphological and physiological effects of the fall B
spray were observed. Among these were a decrease in the size of
flower buds and mature flowers accompanied by reduction of style and
Received for publication . Published with the approval of the director of the Oregon State University Experiment Station as Journal Series No. . From a dissertation submitted by the senior author in partial fulfillment of the requirements for the PhD degree at Oregon State University.
2 Gratitude is expressed to Harold Bjornstad and Fred Dixon for technical assistance. We also thank the Oregon Processed Prune and Plum Growers Commission for financial assistance in this study.

21
pedicel length, a delay in the time of bud break and a decrease in
pollen germinability. B level of pistil and pollen had no effect on
in vivo pollen tube growth rate.
The postulated roles of B in phenolic metabolism and auxin
oxidation are discussed in relation to the observed effects of fall B.
Introduction
The 'Italian' prune is noted for its erratic bearing in the
Willamette Valley and in other areas of the world (118, 160, 161).
Low temperatures at anthesis and during the post-bloom period delay
fertilization and thus limit fruit set. Thompson and Liu (167) found
that when the average temperature for the 3 weeks immediately follow-
ing bloom was below 10 C a poor crop resulted.
In 1972, a year of generally poor cropping for 'Italian,' M.H.
Chaplin (personal communication) found that 'Italian' prune yields
could be increased by application of a fall foliar B spray. A crop of
115.5 kg fruit per tree was produced on trees which had received fall
B while adjacent untreated trees bore 57.6 kg per tree. These trees,
with 29 and 27 ppm B respectively, were not deficient in B by August
mid-shoot leaf analysis, the standard index of B nutrition for prunes.
According to the Oregon State University Fertilizer Guide for Prunes

22
(153), below 25 ppm B indicates deficiency while 35-80 ppm is con-
sidered optimal.
The purpose of this study was to confirm the yield increases
after fall foliar B application of trees which are apparently not
deficient in B and to compare the effects of a fall B spray on fruit set
with those of a prebloom B application. We wished to discover whether
the increased yields were due to a greater initial fruit set or to a
decrease in the amount of midsummer or "blue" drop. We also
wished to learn whether the B spray affected in vivo pollen tube growth
rate. During the course of the investigation, differences due to the
fall B spray in factors such as pollen germinability, flower size,
including style and pedicel length, and date of bloom were noted.
In the years of this study, .1975 and 1976, most 'Italian' prune
orchards in the Willamette Valley bore good crops of fruit. Unlike
1972, temperatures did not appear to be limiting fruit set. The
3-week average postbloom temperatures at the OSU experimental
orchard were 12.3 C in 1975 and 12 C in 1976.
Materials and Methods
Experiments were conducted at the Oregon State University
Experimental Orchard near Corvallis, Oregon (OSU) and in six com-
mercial orchards in the Willamette Valley. All orchards were
'Italian' prune on seedling peach (OSU and Orchards 1, 2, 3, 4 and

23
5) or myrobalan plum (Orchard 6) rootstocks. A range of nutritional
conditions was represented by the orchards selected. OSU and
orchards 1, 3, 4 and 6 had adequate levels of both B and N by August
mid-shoot leaf analysis, while orchard 2 was deficient in both B and
N and orchard 5 was deficient in N. Boron was applied as a com-
mercial product containing 20.5% B (78% Na Bo0...MH-O and 20%
Na B O *5H O). Three hundred ppm wetting agent (alkyl aryl poly-
oxyethylene glycol and free fatty acids in isopropanol) was added in
1976. Application was made with a hand sprayer and trees were
sprayed to the point of drip. Randomized block design was used with
eight replications in the OSU orchard and five replications in each
of the commercial orchards.
In 1975, B treatments at OSU included: (1) 4 92 ppm B (2 lb
product/lOO gal) 2 weeks after harvest and (2) 492 ppm B at first white
stage, when petals were first visible in the flower bud (22).
In 1976, timing of the fall B spray relative to harvest and con-
centration of the s pray were varied. As brown rot, Monilinia sp.,
may be a problem in the Willamette Valley, wettable sulfur was added
to one preharvest spray to determine compatibility of B and S when
applied together. Boron treatments at OSU in 1976 were: (1) 492 ppm
B 1 week before harvest, (2) 492 ppm B + 4798 ppm S 1 week before
harvest, (3) 492 ppm B 2 weeks after harvest, (4) 246 ppm B 2 weeks

24
after harvest, and (5) 123 ppm B 2 weeks after harvest. In the com-
mercial orchards 492 ppm B was applied within 2 weeks after harvest.
In each orchard approximately 8, 000-10, 000 flowers were
counted. Four to six limbs, each bearing approximately 300-400 flow-
ers, were selected on each tree. Blossom counts were made pre-
bloom. At OSU, initial set counts were made 4-5 and 7 weeks after
full bloom, and preharvest counts were made after the "blue" drop in
mid-August. In the commercial orchards, fruits were counted 5-7
weeks after bloom and after the "blue" drop. Fruit set data were
analyzed as a split plot, with dates at which set counts were made the
main plots and B treatments the subplots.
Nutritional status of all trees involved in the experiment was
determined by sampling mid-shoot leaves in August before B treat-
ment. Boron determinations were made on a 3/4-meter direct read-
ing spectrometer using the method of Chaplin and Dixon (37). Nitrogen
was measured using an automated Kjeldahl technique (139)-
To measure in vivo pollen tube growth, pollen from trees receiv-
ing 492 ppm B as a postharvest spray was placed on stigmas of control
flowers and of flowers from trees which had received the spray.
Likewise, control pollen was placed on both kinds of stigma. Pollen
was obtained from anthers removed in the orchard the previous day
and allowed to dehisce on glassine paper overnight. All pollen was
checked for germinability. Flowers to be used for pistils were picked

25
from the orchard in early morning just before petals unfolded.
Approximately 50 flowers per treatment were pollinated. Pollinated
flowers were placed on moist filter paper in petri dishes and incu-
o bated at 10 C for 2 days. This temperature was chosen because it
has been found to limit fruit set (166). Emas culation was unnecessary
because, due to the high humidity in the petri dish, anther dehiscence
did not occur. Pollen tubes were measured using Martin's (110)
fluorescence technique. Styles were frozen in the aniline blue stain
for later examination. Tubes fluoresced much more brightly when
styles were preserved in this manner than when a fixative or soften-
ing solution was used. After freezing a softening solution is not
needed. Styles were squashed in stain under plastic coverslips and
the length of the longest tube recorded. A completely randomized
design was employed in data analysis.
o In vitro pollen germination was also tested at 10 C. Large
populations of pollen were used to eliminate differences due to the
"population effect, " a phenomenon whereby large numbers of pollen
stimulate germination (26). Pollen was allowed to germinate in hang-
ing drops of 10% sucrose with and without 10 ppm boric acid. One
hundred to 500 pollen grains for each of six replications were counted.
Data were analyzed as a split plot, with B treatments the main plots
and B in the germination medium subplots.

26
Styles and pedicels were measured for 25 full bloom flowers
from each of five trees which had received 492 ppm B as a post-
harvest spray and from each of five control trees. The OSU orchard
and orchard 6 were sampled. Randomized block design was used for
data analysis. Average bud or flower size was obtained from dry
weight measurements of samples of 200 buds or flowers collected at
various stages of floral development. Flowers and buds of uniform
external stage of development were taken at each sampling date.
Treatments were replicated by location and data analyzed by t-test.
Results and Discussion
In 1975 initial fruit set at the OSU orchard was increased after
a fall B application (Table 1). These initial set increases following the
fall spray carried through to harvest. There was no significant dif-
ference due to treatment in the amount of fruit lost in the midsummer
or "blue" drop. A spring spray had no effect on initial, blue drop, or
final fruit set.
In 1976 initial fruit set at the OSU orchard was increased by all
preharvest B sprays and by all concentrations of postharvest sprays
applied (Table 2). The addition of wettable sulfur to a preharvest
spray for brown rot control did not interfere with the effect of B on
fruit set. Initial set increases were again sustained through the
season and, as in 1975, "blue" drop was not influenced by B treatment.

27
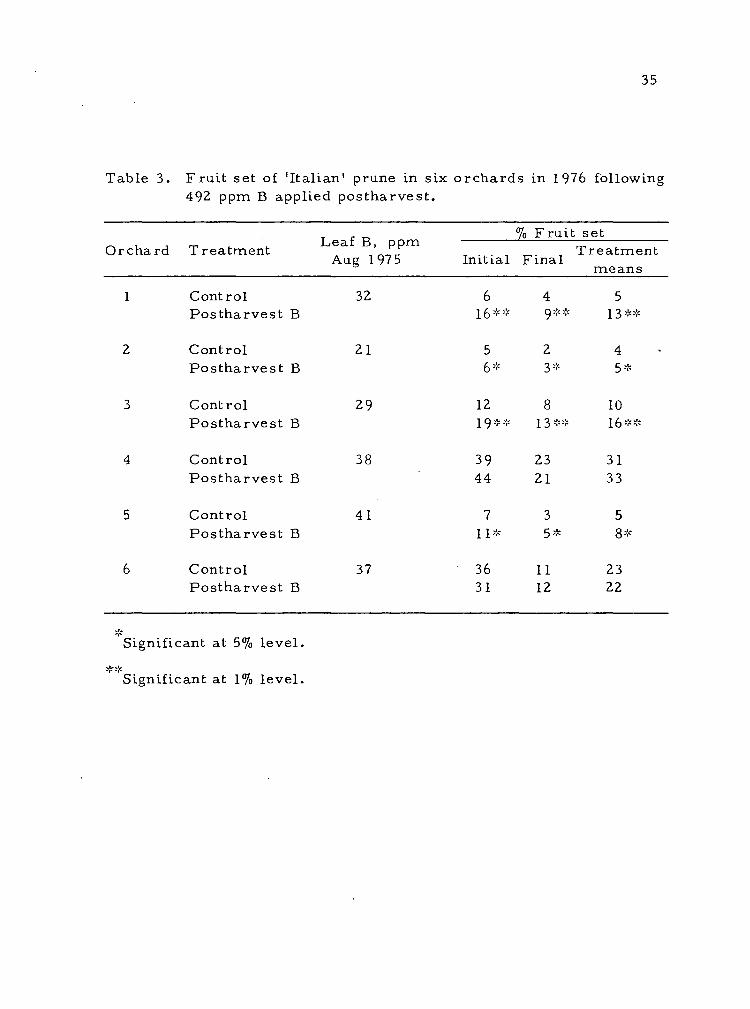
Four of the six commercial orchards also responded to the fall
B spray with increased fruit set in 1976 (Table 3). Of these, orchards
2 and 5, which were deficient in N, experienced very low fruit set.
This would be expected due to the need for N for fruit set (182, 187).
The other two orchards in which fruit set was increased after fall B,
orchards 1 and 3, had sufficient levels of both B andN. Fruit set
after fall B in these orchards was very good. Orchards 4 and 6,
which also had adequate B and N levels, had ample fruit set regard-
less of treatment. Apparently there were no limiting factors for
optimum fruit set in these orchards.
The presence of green leaves in the fall at the time of spraying
appears to be essential in 'Italian' for maximum effectiveness of B
application. Spring sprays have been found effective in correcting B
deficiencies of pear (8, 92) but even here a fall spray often proved the
most consistent (92). However, the increased fruit set in 'Italian'
prune in this study does not appear to be the result of correcting an
incipient B deficiency as in the "blossom blast" condition of pear. In
the latter case cold, heavy soil suppresses B uptake in the spring
and deprived flowers "blast" or shrivel and die. Control trees in this
study showed no signs of blossom dieback and at the OSU orchard had
adequate fruit set though lower than sprayed trees. Furthermore,
we would expect low flower bud B levels in control trees suffering
from an incipient B deficiency (8). Control flower bud levels were no

28
lower in orchards which responded to the fall B spray with increased
set than in orchards which had good fruit set regardless of B treat-
ment (data to be presented in subsequent paper).
The observed increase in fruit set after fall B does not appear
to be the result of accelerated pollen tube growth rate. Pollen tube
growth in 'Italian' prune styles at 10 C was unaffected by either the
B nutrition of the pollen-bearing plant or of the pistil-bearing plant.
This is in agreement with a report by Whittington (184), who found
that parental B level did not influence in vivo pollen tube growth rate
in red clover.
Pollen from trees which had received the 492 ppm B posthar-
vest spray was poorer in germinability than pollen from untreated
trees (Table 4). No difference was seen in germinability due to the
addition of 10 ppm boric acid to the germination medium. Thompson
and Batjer (165) reported a stimulation of plum pollen germination by
the addition of B to the medium, while Blaha and Schmidt (17) found
no such response in plum and prune pollen. Conflicting reports of B
requirements for pollen germination may be reconciled when plant B
supply is considered. Several authors have found that germination of
pollen from plants low in B is enhanced by the addition of B to the
medium but when the plants have adequate B there is no longer a
benefit from this addition (3, 115, 174). In contrast, soil-induced B
toxicity in peach trees was reported to reduce pollen germination

29
when compared with pollen from normal trees (95). In this study of
'Italian' prune, pollen germinability was also decreased but this was
apparently not sufficient to offset the beneficial effects of fall B on
fruit set.
Time of bloom was also affected by fall foliar B. On treated
trees in the orchard at OSU, bloom was delayed 3-4 days in 1975 and
1-2 days in 1976. When branches were brought into the laboratory in
early March and forced at 20 C, flowers on branches given all fall B
treatments opened 1 day after the control. When flower buds and
flowers at comparable stages of external development were compared,
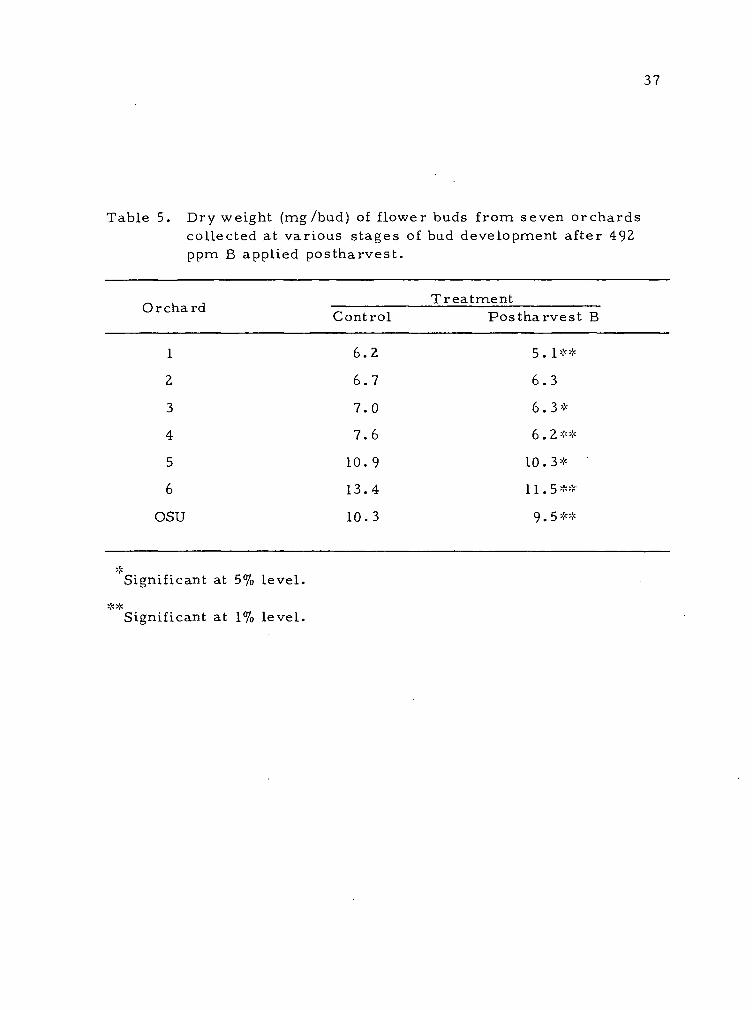
buds from trees receiving fall B applications were found to be smaller
on a dry weight basis than those from control trees (Table 5). This
is reflected in a decrease in style and pedicel length of mature
flowers (Table 6). The effect on floral morphology appears to be
general. The actual amount of decrease in style length, though
statistically significant, amounts to only about 3% of the total style
length. It is doubtful that this alone would be responsible for increas-
ing initial fruit set; that is, by shortening the length of time from
pollination to fertilization. In addition, in both 1975 and 1976
temperatures were not low enough to limit fruit set because of slow
pollen tube growth. Style length as influenced by rootstock was found
to be important in 'Italian' prune fruit set in Poland (76). The authors
proposed that shortened styles positioned anthers and stigmas for

30
more effective self-pollination. Thompson and Liu (166) found no
increase in fruit set with supplemental pollination in orchard 6 and
in five other 'Italian' prune orchards in the Willamette Valley, indi-
cating that pollen transfer is not a limiting factor.
The observed morphological and developmental effects induced
by high B levels reflect alterations in physiology which probably in-
clude hormonal balances. One of the postulated roles of B in the plant
is regulation of auxin levels. Coke and Whittington (41) and others
(131, 145) proposed that B deficiency is actually a state of auxin
toxicity. Auxin-like activity was shown to be high in extracts of
B-deficient plant materials, and B deficiency symptoms could be
duplicated by applying auxins (41, 97, 131, 133). One of the means by
which auxin levels may be controlled is the IAA oxidase (peroxidase)
system (136, 138, 157, 164). Several of the polyphenolic compounds
which accumulate in B deficiency in sunflower such as chlorogenic and
caffeic acids (51) will inhibit IAA oxidase (87). Chlorogenic acid is
one of the most abundant phenolics in Prunus domestica (84). By
controlling the synthesis (101, 145) or metabolism (131) of phenolic
compounds which will inhibit IAA oxidase, B may decrease auxin
levels in plant tissue.
Although the mechanism of the fruit set increase following fall
B is not known, balance between auxin and other hormones such as
gibberellins and cytokinins at specific stages in fruit set may be

31
involved (46, 88, 112). Stone fruits do not readily set fruit without
fertilization, but gibberellin induced parthenocarpy in plum (89).
peach (48, 49), almond and apricot (48). Both gibberellin and auxin
were required for parthenocarpic fruit set in 'Bing' cherry (47). An
alteration of auxin balance between shoot and fruit could also influence
fruit set. Boron accumulates in both leaf and flower buds after a fall
foliar B application (data not presented), so a decrease of auxin
levels might be expected to occur in developing leaves as well as
fruits. At the time of initial fruit set both developing shoots and
fruits are metabolic sinks for assimilates stored in the tree. Move-
ment of assimilates appears to be directed toward areas of high auxin
concentration (22, 142, 177). A decrease in leaf auxin levels could
cause leaves to be less competitive with flowers for available nutrients
and allow more nutrients to be directed to the flower, thereby increas-
ing fruit set. Schneider (137) has proposed that naphthaleneacetic acid
(NAA), when applied to the entire tree after bloom, thins apples by
creating vegetative sinks which compete with the fruit for nutrients.
Another synthetic auxin, 2-(3 chlorophenoxy)-propionamide (3-CPA), is
effective as a post-bloom thinner for peach, plum and apricot (M.N.
Westwood, personal communication).
A role for auxin can also be envisioned in some of the other
observed responses to fall B. It is not known whether style and pedi-
cel length is reduced due to a decrease in cell elongation or cell

32
division, but higher auxin levels in the controls could conceivably
cause styles and pedicels to be longer through stimulation of cell
elongation. Also, higher auxin levels in control trees could lead to
an advance in the time of bud break by inducing renewed cambial
activity (99). Interaction between auxin and other hormones such as
ethylene or cytokinin is also possible. Measurement of auxin and
other hormone levels in developing flowers and leaves after a fall B
spray would aid greatly in elucidating the physiological basis of the
observed fruit set increase, and would also contribute to our under-
standing of the role of B in the plant.

33
Table 1. Fruit set of 'Italian' prune, 1975, following postharvest and prebloom B sprays. Oregon State University Experimental Orchard, Corvallis, Or.
% Fruit set Treatment _, , A„„l „ , „ ,_„ ,_ , A ^„ Treatment
4 wk AFB 7 wk AFB 17 wk AFB means
Control 26 21 11 19
492 ppm B postharvest 1974 35** 31** 17** 27**
492 ppm B prebloom 1975 28 20 10 19
AFB = after full bloom.
Significantly different from control at 1% level.

34
Table 2. Fruit set of 'Italian' prune, 1976, following fall foliar B sprays. Oregon State University Experimental Orchard, Corvallis, Or.
Treatment % Fruit set
„• , A^„i _ , A ^„ ,„ , . „„ Treatment SwkAFB1 7 wk AFB 17 wk AFB means
Control 23 22 18
492 ppm B preharvest 32** 31* 13** 25**
492 ppm B + 4798 ppm S preharvest 33** 31** 12** 25**
492 ppm B postharvest 35* 32** 15** 27*=:
246 ppm B postharvest 30 = 2 9" 13" 24**
123 ppm B postharvest 32** 30** 12** 25**
1 AFB = after full bloom.
** Significantly different from control at 1% level.
35
Table 3. Fruit set of 'Italian' prune in six orchards in 1976 following 492 ppm B applied postharvest.
Treatment Leaf B, ppm
Aug 1975
% Fruil t set Orchard
Initial F 'inal Treatment
means
1 Control 32 6 4 5 Postharvest B 16** Q** 13**
2 Control 21 5 2 4 Postharvest B 6* 3* 5*
3 Control 29 12 8 10 Postharvest B 19** 13** 16**
4 Control 38 39 23 31 Postharvest B 44 21 33
5 Control 41 7 3 5 Postharvest B 11* 5* Q -v
6 Control 37 36 11 23 Postharvest B 31 12 22
Significant at 5% level.
Significant at 1% level.

36
Table 4. Ge rminability of 'Italian' prune pollen in deionized water with 10% sucrose following 492 ppm B postharvest. Oregon State University Experimental Orchard, Corvallis, Or.
Treatment of Germination « ^ 1 % Germination
pollen source medium
Control no added B 69
Control 10 ppm B 65
Postharvest B no added B 42
Postharvest B 10 ppm B 37
1 Effect of pollen source significant at 5% level. Effect of B in germination medium not significant.
37
Table 5. Dry weight (mg/bud) of flower buds from seven orchards collected at various stages of bud development after 492 ppm B applied postharvest.
Orchard
1
2
3
4
5
6
OSU
T rea tment Control Pos tharvest B
6.2 D . 1 ^ '^
6.7 6.3
7.0 6.3*
7.6 6.2* *
10.9 10.3*
13.4 11.5 **
10.3 9.5**
**
Significant at 5% level.
Significant at 1% level.

38
Table 6. Style and pedicel length of 'Italian' prune in two orchards after 492 ppm B postharvest.
Orchard Treatment Style, mm Pedicel, mm
OSU Control 11.7
Postharvest B 11.3**
Control 11.2 15.4
Postharvest B 10.9** 12.8**
r for style and pedicel = 0.4152.
Significant at 1% level.

39
BORON DISTRIBUTION IN 'ITALIAN' PRUNE TISSUES FOLLOWING SPRING AND FALL BORON SPRAYS1
Nancy W. Callan2
Department of Horticulture, Oregon State University Corvallis, Or. 97331
Additional index words. Prunus domestica L., flowers, flower buds, anther, ovary, style
Abstract. Boron levels in 'Italian' prune (Prunus domestica L. )
tissues following a fall foliar B application and a prebloom B applica-
tion were compared. Fall foliar B increased B levels in dormant bud
and spur tissues and in flower buds and flowers. A prebloom B spray
increased B levels of floral tissues to a lesser degree. The highest
B concentration was found in the ovary. Boron concentration of flow-
er buds in April following a fall B spray was as much as five times
the amount ordinarily found in mid-shoot leaves in August. August
mid-shoot leaf analysis revealed higher levels in leaves from trees
treated the previous fall in one year of the study but not in the other
year. In fall-treated plants, B appeared to be uniformly distributed
Received for publication . Published with the approval of the director of the Oregon State University Experiment Station as Journal Series No. . From a dissertation submitted by the senior author in partial fulfillment of the requirements for the PhD degree at Oregon State University.
2 Gratitude is expressed to Harold Bjornstad and Fred Dixon for technical assistance. We also thank the Oregon Processed Prune and Plum Growers Commission for financial assistance in this study.

40
in leaves from spur, shoot tip, mid-shoot and shoot base. In control
plants, B was higher in mid-shoot leaves than in leaves from spur,
shoot tip and shoot base. The suggestion is made that while August
mid-shoot leaf analysis is a useful indicator of the vegetative nutri-
tion of 'Italian' prune trees, analysis of bud or flower tissue might be
a better index of the B needs of the tree for reproductive growth.
Introduction
A fall foliar B application has been found to increase initial
fruit set of 'Italian' prune trees which were not B deficient by
August mid-shoot leaf analysis, the standard index of B nutrition.
Several morphological and physiological changes such as a decrease
in flower bud size, a delay in bloom and a decrease in pollen germin-
ability accompanied the set increase (Callan, first chapter of thesis).
The purpose of this study was to determine the distribution in
the plant of the applied B by analyzing B levels in various plant
tissues. Of special interest was the B concentration of floral tissues,
as these tissues often contain some of the highest B levels found in
plants (21, 191). The effect of B sprays on leaf levels the following
August was also of interest.
Materials and Methods
Experiments were conducted at the Oregon State University

41
Experimental Orchard near Corvallis, Oregon (OSU) and in six com-
mercial orchards in the Willamette Valley. All orchards were
'Italian' prune on seedling peach (OSU and orchards 1, 2, 3, 4 and 5)
or myrobalan plum (orchard 6) rootstocks. A range of nutritional
conditions was represented by the orchards selected. OSU and
orchards 1, 3, 4 and 6 had adequate levels of both B and N by August
mid-shoot leaf analysis, while orchard 2 was deficient in both B and
N and orchard 5 was deficient in N. Boron was applied as a com-
mercial product containing 20.5% B (78% Na Bo0..^H-O and 20% 2 8 13 2
Na B O *5H O). Three hundred ppm wetting agent (alkyl aryl
polyoxyethylene glycol and free fatty acids in isopropanol) was added
in 1976. Application was made with a hand sprayer and trees were
sprayed to the point of drip. Randomized block design was used with
eight replications in the OSU orchard and five replications in each of
the commercial orchards.
In 1975, B treatments at OSU included: (1) 492 ppm B (2 lb
product/100 gal) 2 weeks after harvest and (2) 492 ppm B at first
white stage, when petals were first visible in the flower bud (22).
In 1976, timing of the fall B spray relative to harvest and con-
centration of the spray were varied. As brown rot, Monilinia sp.,
may be a problem in the Willamette Valley, wettable sulfur was added
to one preharvest spray to determine compatibility of B and S when
applied together. Boron treatments at OSU in 1976 were: (1) 492 ppm

42
B 1 week before harvest, (2) 492 ppm B + 4798 ppm S 1 week before
harvest, (3) 492 ppm B 2 weeks after harvest, (4) 246 ppm B 2 weeks
after harvest, and (5) 123 ppm B 2 weeks after harvest. In the com-
mercial orchards 492 ppm B was applied within 2 weeks after harvest.
Boron determinations were made on a 3/4-meter direct reading
spectrometer using the method of Chaplin and Dixon (37). In 1975,
at OSU, eight replicates of 40 flowers each were sampled at fall bloom
and at petal fall. Anthers, ovaries and styles were analyzed sepa-
rately. Data from each sampling date were analyzed separately as
split plots, with flower parts the main plots and B treatments the
subplots. Short spurs (less than 2 cm) with 4-7 buds attached were
sampled in all orchards in Feb 1976. Ten spurs were collected
from each tree and samples composited for analysis. At OSU,
flower buds were collected at the tip green stage at which time
florets become visible within the bud scales, at the first white stage
when sepals separate and at full bloom (22). In the commercial
orchards, flower buds were collected at various stages before bloom.
Sixteen buds or flowers were collected from each tree and samples
composited for each, treatment. Bud weight was calculated from the
dry weight of the composite sample. Mid-shoot leaves were sampled
in August of 1975 and 1976. At OSU, 20-30 mid-shoot leaves were
taken per tree from 16 replications in 1975 and eight replications in
1976. In 1976 the two basal leaves on the shoot, the terminal shoot

43
leaf and two spur leaves were sampled from trees which had received
492 ppm B postharvest and from control trees in four replications.
Data were analyzed as a split plot, with B treatments the main plots
and leaf location the subplots.
Results and Discussion
Boron levels of dormant bud and spur tissues were greatly-
increased by fall foliar B sprays (Tables 7 and 8). When different
concentrations of B were applied, the amount of B found in bud and
spur tissue was roughtly proportional to the amount applied. At OSU,
applied before or after harvest, the highest concentration spray,
492 ppm B, approximately doubled the B concentration of these
tissues (Table 8). In the commercial orchards (Table 7), the 492
ppm B postharvest spray increased bud and spur B 50% to 200% over
the control. Boron concentration was higher in the bud than in the
spur. In control trees at OSU after leaf fall, buds contained 55 ppm B
and spur tissue 30 ppm B. At this time trees which had received fall
B as a 492 ppm postharvest spray contained 187 ppm B in buds and
95 ppm B in spurs. Differences due to tissue and to treatment are
significant to the 1% level. Since B appears to increase proportion-
ately in bud and spur tissues after fall B, combined bud and spur
tissue analysis should be a good indicator of the B status of dormant
tissue.

44
With the approach of anthesis B apparently moves into the flow-
er buds from subtending tissues. Little change occurred in B con-
centration of the buds despite the considerable increase in dry
weight which takes place between the green tip stage and full bloom
(Table 8). This is in agreement with reports by Woodbridge et al.
(191) who found that pear, apple and cherry buds did not store suffi-
cient B for their development but drew on B from the soil or from
reserves within the tree.
Boron levels in flower buds were increased considerably by the
fall B sprays, especially those of higher concentration. In each
orchard, after fall B treatment, the amount of increase in flower bud
B was approximately the same as that seen in bud and spur tissue.
Boron levels in anther, ovary and style, measured at full
bloom and at petal fall, were found to be higher after a fall or a
spring B spray than in the control (Table 9). A fall spray increased
B levels in the flower parts to a greater extent than did a prebloom
spray. Boron concentration of the ovary was higher than that of the
anther or of the style.
Young shoot B levels at the time of full bloom were also
increased by the fall B spray. Developing shoots 4-5 cm long from
orchard 6 had 44 ppm B in the control buds and 94 ppm after fall B.
Much of this B may be utilized in current season growth, however, as
B levels of mid-shoot leaves in August were increased by fall B in

45
only one of the two years studied (Table 10). In 1975 the amount of
increase in mid-shoot leaf B concentration after fall B was 6 ppm.
In 1976 none of the fall B applications tested increased mid-shoot
leaf B over that of the control. However, differences in B levels
due to the fall B spray were found in leaves from locations other
than mid-shoot, i.e., from spur, shoot base and shoot tip (Table 11).
In fall B-treated plants, B appeared to be uniformly distributed in
leaves from spur, shoot tip and shoot base (Table 11). In control
plants, B was higher in mid-shoot leaves than in leaves from spur,
shoot tip and shoot base. The cause of this is not known. Perhaps
the lower values of spur and shoot base leaves in control trees is a
result of B being drawn from them to the fruit. However, Leece
(10Z) found no difference in B content of leaves from fruiting and non-
fruiting wood in peach. It may also be that under more limited B
supply, as in the control trees, B content of leaves may reflect B
availability at the time they are developing. Spur and shoot base
leaves develop while the soil is still cold in early spring. Boron
uptake may have been hampered at this time and only trees which
had received fall B contained a large reserve of B readily available
for bud growth.
Preliminary studies show that B in the fall sprays is readily
taken up by the leaf and translocated to the bud and subtending tis-
sues. Boron is taken up quickly and passively into the intercellular

46
spaces in roots (23) and it is possible that this mechanism of uptake
can function in the leaf. Once in the leaf, B can move in both xylem
and phloem (121). Boron may be transported out of the leaf to the
bud and woody tissues in a complex with sugars or other polyhydroxy
compounds (70). The B, applied as a spray, apparently becomes
part of the "free" or "unbound" B pool. Shive (143) found that this
fraction of the plant B varied with B supply, dropping to zero as
deficiency symptoms arose. The amount of "bound" B in the plant
remained relatively constant. Mobility of B in stone fruits is well
known. Under conditions of excess B stone fruits will transport B
from the leaves to the bark and also to the fruit (63).
Fall B sprays are a means to achieve high bud and flower B
concentrations which stimulate fruit set in the spring. Due to the
dilution effect with growth, vegetative injury which would result from
high B levels later in the season is avoided. It is interesting to note
that, while response to fall foliar B could hardly be considered
toxicity because of the fruit set increase, two of the features of the
fall B response, a delay in bloom date and a decrease in pollen
germinability (Callan, first chapter of thesis), are characteristic of
B toxicity in peaches (38, 90, 186).
August mid-shoot leaf analysis is the standard index of 'Italian'
prune B nutrition. It indicates the B level, for best growth, perform-
ance and general tree health. However, these experiments and

47
studies by Montgomery (115) with alsike clover and Holley and Dulin
(85) with cotton indicate that optimum B levels for reproduction are
higher than those for vegetative growth. It may be possible to define
a B level for optimum reproductive growth, perhaps by utilizing
dormant bud and spur tissue or floral tissue as indicators.

48
Table 7. Boron (ppm) in 'Italian' prune dormant bud and spur tissue and in flower buds in six orchards after 492 ppm B post- harvest.
Orchard Treatment Bud and spur Bud, first white
Jan 1976 April 1976
Control 26 71 Postharvest B 66 166
Control 13 33 Postharvest B 38 101
Control 22 6l Postharvest B 45 150
Control 23 73 Postharvest B 50 162
Control 47 121 PostharvestB 68 171
Control 29 66 Postharvest B 76 158

49
Table 8. Boron (ppm) in 'Italian' prune dormant bud and spur tissues and in floral tissues in two stages of bud development and full bloom, 1976. Oregon State University Experimental Orchard, Corvallis, Or.
Bud and spur, Flower Treatment , Green First
Jan . , . Full bloom tip white
Control 37 100 88 87
492 ppm B preharvest 66 175 172 165
492 ppm B + 4798 ppm S preharvest 76 168 175 170
4 92 ppm B postharvest 66 205 185 186
246 ppm B postharvest 62 149 131 153
123 ppm B postharvest 49 118 105 138

Table 9. Boron (ppm) in 'Italian' prune flower parts at full bloom and at petal fall, 1975, as affected by 492 ppm B applied postharvest 1974 and prebloom 1975.
Full bloom Petal fall
Treatment Flower part Flower part
... _ _. . Treatment Treatment Anther Ovary Style Ovary Style means means
Control 47 59 52 53 64 50 57
Postharvest B 120 171 133 141 138 89 113
Prebloom B 63 98 79 80 90 69 80
Flower part means 77 109 88 97 69
Full bloom: LSD for flower part: 5% = 9, 1% = 12. LSD for B treatment: 5% = 11, 1% = 15.
Petal fall: LSD for flower part: 5% = 10, 1% = 14. LSD for B treatment: 5% = 18, 1% = 24.
o

51
Table 10. Boron (ppm) in 'Italian' prune mid-shoot leaves in August as affected by B application. Oregon State University Experimental Orchard, Corvallis, Or.
Treatment 1975 1976
2 3 Control 36 35
492 ppm B prebloom 38
492 ppm B preharvest 37
492 ppm B + 4798 ppm S preharvest 38
492 ppm B postharvest 42** 33
246 ppm B postharvest 31
123 ppm B postharvest 33
Data supplied by Dr. M.H. Chaplin, Horticulture Dept., OSU. 2
Mean of 16 replications. 3 Mean of 8 replications.
** Significantly different from control at 1% level.

52
Table 11. Boron (ppm) in 'Italian' prune leaves from four locations on the tree in August 1976 as influenced by 492 ppm B applied postharvest, 1975. Oregon State University Experimental Orchard, Corvallis, Or.
r . L. Treatment Leaf location
Control Postharvest B
Spur 28 33
Shoot base 27 34
Mid-shoot 36 34
Shoot tip 25 31
Mean 29 33
Treatment effects not significant.
LSD for leaf location within a given B treatment: 5% = 3, 1% = 4.

53
THE EFFECT OF FALL FOLIAR BORON APPLICATION ON POLYPHENOLOXIDASE ACTIVITY AND INTERNAL BROWNING IN 'ITALIAN' PRUNE FRUIT1
2 Nancy W. Callan Department of Horticulture, Oregon State University Corvallis, Or. 97331
Additional index words. Prunus domestica L.
Abstract. Polyphenoloxidase activity of acetone powders prepared
from immature 'Italian' prune (Prunus domestica L. ) fruit was higher
in fruits from trees treated with fall foliar B the preceding year than
in fruit from control trees. This was in contrast to the observation
that green fruits from fall B-treated trees browned more slowly when
cut than did controls. Further, after fall B there was a decrease in
the amount of internal browning, a physiological disorder involving
breakdown and darkening of the fruit flesh next to the stone. The
role of B in phenolic metabolism is discussed in relation to both of
these observations.
Received for publication . Published with the approval of the director of the Oregon State University Experiment Station as Journal Series No. . From a dissertation submitted by the senior author in partial fulfillment of the requirements for the PhD degree at Oregon State University.
2 Gratitude is expressed to Harold Bjornstad and Fred Dixon for technical assistance. We also thank the Oregon Processed Prune and Plum Growers Commission for financial assistance in this study.

54
Introduction
A fall foliar B application has been found to influence reproduc-
tive development of 'Italian' prune. Initial fruit set was increased by
fall B (Callan, first chapter of thesis). High levels of B were found
in buds and flowers the spring following a fall B spray (Callan, second
chapter of thesis).
Boron also appears to influence browning reactions. When flesh
of immature prune fruits was prepared for B analysis, the cut sur-
faces of fruit from trees which had received fall foliar B did not brown
as rapidly as those of fruit from control trees. Polyphenoloxidase
(PPO) is active in promoting browning in many fruits. Dilley (57)
states that "browning is the result of phenol oxidation and eventual
non-enzymatic polymerization of the quinones formed into tannins or
'melanins. ' " Polyphenoloxidase has been found to increase during B
deficiency in sunflower (107), tomato (96), and potato (154). In
addition, borate has been shown to be a competitive inhibitor of PPO
(180). Therefore, I wished to determine if a correlation existed
between the B content of immature prune fruit and its PPO activity.
I also wished to determine the effect of fall B on fruit quality, in
particular the complex physiological disorder called inte rnal brown-
ing. This disorder involves breakdown of the flesh and a darkening
next to the pit in mature fruit (172). A similar disorder of apricots
was caused by B deficiency (33), but attempts to prevent internal

55
browning in prunes by the use of B sprays have given variable
results (172).
Materials and Methods
Experiments were carried out at the Oregon State University
Experimental Orchard near Corvallis, Oregon. The orchard is
'Italian' prune on seedling peach rootstock. Boron was applied as a
commercial product containing 20.5% B (78% Na B00 _*4H_0 and 2 o 13 2
20% Na Bo0 -SH O). Three hundred ppm wetting agent (alkyl aryl 2 o 7 2
polyoxyethylene glycol and free fatty acids in isopropanol) was added.
Application was made with a hand sprayer and trees were sprayed to
the point of drip. Randomized block design with eight replications
was used. As brown fruit rot, Monilinia sp., may be a problem in the
Willamette Valley, wettable sulfur was added to one preharvest spray
to determine compatibility of B and S when applied together. Boron
treatments were: (1) 492 ppm B (2 lb product/100 gal) 1 week before
harvest, 1975 and again in 1976, (2) 492 ppm B + 4798 ppm S (4 lb /
100 gal) 1 week before harvest, 1975, and 984 ppm B 1 week before
harvest, 1976, (3) 492 ppm B 2 weeks after harvest, 1975, (4) 246
ppm B 2 weeks after harvest, 1975, and (5) 123 ppm B 2 weeks after
harvest, 1975i
Flesh of immature prune fruits from control trees and from
trees which had received a spray of 492 ppm B 2 weeks after harvest

56
were analyzed for B content and PPO activity on July 5, 1976, just
after the midsummer or "blue" drop. Boron determinations were
made on a 3/4-meter direct reading spectrometer using the method
of Chaplin and Dixon (37). Twenty fruits from each of seven trees
of each treatment we re sampled for B content. Acetone powders
were prepared from 20 fruits from each of two trees receiving each
treatment. Fruit flesh was homogenized with 100 ml cold acetone,
filtered three times with 100 ml cold acetone and the residue air-
dried overnight. To assay enzyme activity, 0.1 g acetone powder was
mixed with 10 ml O.OZ M pH 6.9 phosphate buffer and allowed to stand
10 min before filtering. Two ml filtrate and 1 ml 0. 1 M catechol
were mixed rapidly in a spectrophotometer cell and the OD at 240 nm
recorded after 1/4, 1, 2, 3, 4 and 5 min. A standard containing only
buffer and catechol was used. Average change in OD/min was cal-
culated. This method is a modification of that described by Zocca
and Ryugo (193). All acetone powder preparations were run in dupli-
cate and the experiment was performed twice. Soluble N content of
acetone powder filtrate was calculated in triplicate using Kjeldahl
analysis (139) of freeze-dried samples. The soluble protein was
then estimated by dividing by 6.25%. The amount of soluble N in the
filtrate did not differ due to B treatment. Average change in OD/min/
mg soluble protein was calculated. Data were analyzed as a split plot,

57
with B treatments the main plots and the individual acetone powder
preparations the subplots.
At harvest in 1976, eight replicates of 25 fruit per sample were
weighed and examined for fruit quality. Fruits were sliced length-
wise and transversely and internal browning was recorded. To meas-
ure firmness, a Magness-Taylor pressure tester with an 8 mm
(5 /l6 in) head was used on a peeled cheek of each fruit. To measure
color, fruits were separated into two groups and recorded as red or
purple fruit. Percent soluble solids was measured with a hand
refractometer. Two drops of juice from the stylar end of 10 prunes
were composited for each reading.
Results and Discussion
Boron content of immature fruit flesh was increased by a fall
foliar B application. Fruit from B-treated trees was higher in B
than fruit from control trees (Table 12). Acetone powders from fall B
fruits also contained more B than those from control trees. Although
the flesh of fruits from B-treated trees appeared to brown less and
acetone powders were lighter in color than those of controls, PPO
activity in acetone powders prepared from these fruits was greater.
The observed difference in PPO activity may not be due to an
increase in the amount of PPO as a result of fall foliar B but rather to
a decrease in phenolic compounds. These are the substrate of PPO

58
and they polymerize to form the pigments in browning of cut surfaces.
Prunus domestica has been reported to be high in phenolics (84).
These compounds are known to interact with proteins and can cause
much difficulty in the isolation of active enzymes (106). Phenolic
compounds accumulate in B-deficient plants (Z8, 51, 125, 131, 178).
B has been reported to shift plant metabolism away from the pro-
duction of phenolic acids (101). It also may be involved with the
synthesis of flavonoids from their phenolic precursors (131). There-
fore, a reduction in the amount of phenolics could be expected with an
abundant supply of B. The lower PPO activity in acetone powders of
control fruit may have been due to a polymerization and binding of the
more abundant phenolic compounds to the enzyme during acetone pow-
der preparation. An inverse relationship between acetone powder
PPO activity and substrate levels exists in apple. As 'Golden
Delicious1 apples mature, activity of acetone powders increased as
the amount of phenolic substrate in apple flesh decreased (193).
The reduction in the incidence of internal browning of mature
fruit after fall B likewise may reflect a decrease in the amount of
substrate available for browning reactions. Internal browning of
mature fruit was decreased by all concentrations of fall foliar B
tested, whether applied before or after harvest the preceding year.
Trees receiving preharvest B sprays in 1975 were sprayed again
before harvest in 1976. However, as less browning was seen in fruit

59
from trees which had received only a single postharvest spray, it
appears that B application the previous fall was responsible for the
observed reduction in internal browning. Yield and/or fruit size was
increased by some concentrations and timings of fall B but color,
soluble solids and firmness were not affected by B treatment (Table
13).
Additional research should be done to determine whether the
observed differences in PPO activity and internal browning were due
to a difference in the amount of PPO present or to the amount of
phenolic substrates available for browning. This could be accom-
plished through the use of phenolic binding agents in enzyme
preparation (106). Research should also involve determination of
differences, qualitative or quantitative, in the phenolic compounds of
'Italian' prune after fall foliar B.

Table 12. Polyphenoloxidase activity and B content of immature 'Italian' prune fruit flesh, July 1976, after 492 ppm B applied postharvest, 1975.
B (ppm) in B (ppm) in mg Soluble ^^ . / ' „ ._ \ . 1 , AOD/ Polyphenoloxidase
Treatment fruit Tree acetone protein/ml i ,. .. ? „ , ,., mm1 activity^
flesh powder filtrate
Control 22 1
Postharvest B 38*
29 .375 .0052
27 .469 .0066
33* .613 .0254
34* .425 .0162
Significant at 5% level.
Significant at 1% level.
.0146
.0395**
AOD/min/mg soluble protein. /N = 420 nm. 2 Each figure is the mean of two trials in each of two experiments.
o
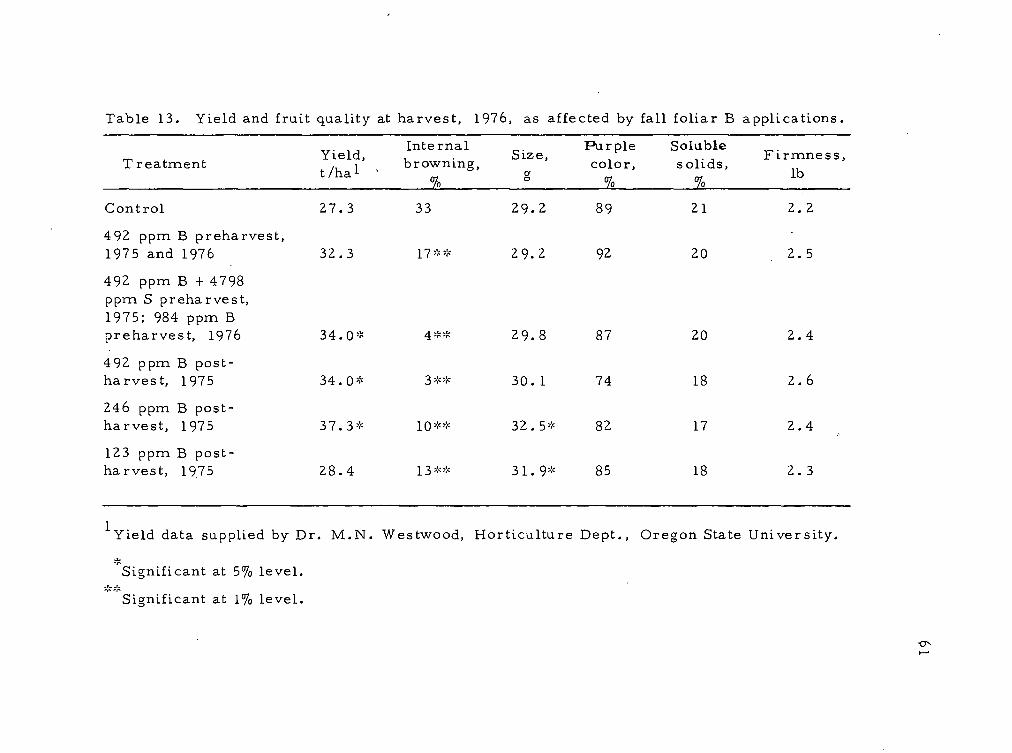
Table 13. Yield and fruit quality at harvest, 1976, as affected by fall foliar B applications.
„. , , Internal Purple Soluble _ _ . . Yield, Size, Firmness, Treatment ,, i browning, color, solids, t/ha % g % % lb
Control 27.3 33 29.2 89 21 2.2
492 ppm B preharvest, 1975 and 1976 32.3 17** 29.2 92 20 2.5
492 ppm B + 4798 ppm S preharvest, 1975; 984 ppm B preharvest, 1976 34.0* 4** 29.8 87 20 2.4
492 ppm B post- harvest, 1975 34.0* 3** 30. 1 74 18 2.6
246 ppm B post- harvest, 1975 37.3* 10** 32.5* 82 17 2.4
123 ppm B post- harvest, 19.75 28.4 13** 31.9* 85 18 2.3
Yield data supplied by Dr. M.N. Westwood, Horticulture Dept., Oregon State University.
Significant at 5% level.
Significant at 1% level.
•c^

62
BIBLIOGRAPHY
1. Acerbo, S., R. Kastori, H. Sochtig, H. Harms and K. Haider. 1973. Effect of boron on synthesis and transformation of lignin-precursors in Zea mays. Z. Pflanzen. 69:306-317.
2. Anderson, D.E. and K. Ohki. 1972. Cotton growth response and boron distribution from foliar application of boron. Agron. J. 64:665-667.
3. Antles, L. C. 1951. Eighteen years of commercial pollen research. Hoosier Hort. 33:68-74.
4. Baker, A.S. andR.L. Cook. 1956. Need of boron fertiliza- tion for alfalfa in Michigan and methods of determining this need. Agron. J. 48:564-568.
5. Baker, J.E., H.G. GauchandW.M. Dugger. 1956. Effects of boron on the water relations of higher plants. Plant Physiol. 31:89-94.
6. Ballard, J.K., E.L. Proebsting, R.B. Tukey and H. Mills. 1971. Critical temperatures for blossom buds. Prunes. Wash. State Univ. Extension Circ. 372.
7. Batjer, L.P. andM.M. Haller. 1942. F ruit maturity and growth of apple trees as affected by boron content. Proc. Amer. Soc. Hort. Sci. 40:29-33.
8. Batjer, L.P., B.L. Rogers and A. H. Thompson. 1953. "Blossom blast" of pears: an incipient boron deficiency. Proc. Amer. Soc. Hort. Sci. 62:119-122.
9- Batjer, L.P. andA.H. Thompson. 1949- Effect of boric acid sprays applied during bloom upon the set of pear fruits. Proc. Amer. Soc. Hort. Sci. 53:141-142.
10. Batjer, L.P. andM.N. Westwood. 1958. Seasonal trend of several nutrient elements in leaves and fruits of Elberta peach. Proc. Amer. Soc. Hort. Sci. 71:116-126.
11. Benson, N.R. andH.E. Dregne. 1949- Leaf analysis gives part of the answer. Better Fruit 43:37-38.

63
12. Benson, N.R., E.S. Degman and I. C. Chmelis. 1961. Translocation and re-use of boron in broccoli. Plant Physiol. 36:296-301.
13. Bertrand, G. and L. Silberstein. 1938. Distribution of boron in organs of the white lily. Compt. Rend. Acad. Sci. Paris 206:796-799. .
14. Bertrand, G. and L. Silberstein. 1940. Distribution of boron in the organs of Nicotiana rustica. Compt. Rend. Acad. Sci. Paris 210:70-73.
15. Bibbey, R.O. 1952. To determine the effect of boron on plant development and seed in alsike clover. Ontario Agr. Coll. and Exp. Farm 76th Ann. Rpt.:38.
16. Birnbaum, E.H., C.A. Beasley and W.M. Dugger. 1974. Boron deficiency in unfertilized cotton Gossypium hirsutum ovules grown in vitro. Plant Physiol. 54:931-935.
17. Blaha, J. and J. Schmidt. 1939- The effect of boron on the germination of pollen in fruit trees. Sbor. Cesk. Akad. Zem. 14:186-192.
18. Blanc, D. 1975. Etude de la nutrition en bore de I'oeillet americain. Ann. Agron. 26:327-336.
19. Bobko, E.V. 1935. Recherches dur le role du bore dans les plantes. Ann. Agron. 5:801-803.
20. Bobko, E.V. and T.D. Priadilshchikova. 1945. On the solubility of boron compounds in the plant. Compt. Rend. (Doklady) Acad. Sci. USSR 48:358-359.
21. Bobko, E.V. and V.V. Zerling. 1938. Influence of boron on the reproductive development of plants. Ann. Agron. 8:174- 184.
22. Bollard, E.G. 1970. The physiology and nutrition of develop- ing fruits. InA.C. Hulme (ed. ). The biochemistry of fruits and their products. Vol. I. Academic Press, London.
23. Bowen, J.E. and P. Nissen. 1976. Boron uptake by excised barley roots. I. Uptake into the free space. Plant Physiol. 57:353-357.

64
24. Bradbury, D. 1929. A comparative study of the developing and aborting fruits of Prunus cerasus. Amer. J. Bot. 16:525-542.
25. Bramlage, W.J. and A.H. Thompson. 196l. The effects of early-season sprays of boron on fruit set, color, finish and storage life of apples. Proc. Amer. Soc. Hort. Sci. 80:64-72.
26. Brewbaker, J. L. and B.H. Kwack. 1963. The essential role of calcium ion in pollen germination and pollen tube growth. Amer. J. Bot. 50:859-865.
27. Briggs, W.R. and P.M. Ray. 1965. An auxin inactivation system involving tyrosinase. Plant Physiol. 31:165-167.
28. Brown, J.C. and J.E. Ambler. 1973. Genetic control of uptake and a role of boron in tomato. Soil Sci. Soc. Am. Proc. 37:63-66.
29. Brown, J.C. andW^E. Jones. 1971. Differential transport of boron in tomato, Lycopersicon esculentum. Physiol. Plant. 25:279-282.
30. Bryden, S.D. 1940. Effect of borax on green color in Granny Smith apples. Agr. DayN.S. Wales 51:525-526.
31. Bukovac, M.J., F. Zucconi, R.P. Larsen and C. D. Kesner. 1969. Chemical promotion of fruit abscission in cherries and plums with special reference to (2- chloroethyl) phosphonic acid. J. Amer. Soc. Hort. Sci. 94:226-230.
32. Bulatovic, S. and M. Jovanovic. 1970. Effect of boron on some pomological characters of Golden Delicious apple. Proc. 18th Int. Hort. Cong. 176-177.
33. Bullock, R.M. andN.R. Benson. 1948. Boron deficiency in apricots. Proc. Amer. Soc. Hort. Sci. 51:199-204.
34. Burrell, A. B. 1958. Boron in apple leaves and fruits as influenced by sodium pentaborate sprays. Proc. Amer. Soc. Hort. Sci. 71:20-25.
35. Burstrom, H. 1948. The rate of the nutrient transport to swelling buds of trees. Physiol. Plant. 1:124-135.

65
36. Chaplin, M.H. 1968. The influence of succinic acid 2,2- dimethylhydrazide (Alar) on fruit ripening and vegetative growth of the sweet cherry (Prunus avium L. ). Ph.D. Thesis. Mich. State Univ.
37. Chaplin, "M. H. and A.R. Dixon. 1974. A method for analysis of plant tissue by direct reading spark emission spectroscopy. Applied Spectroscopy 28:5-8.
38. Cibes, H.R., E. Hernandez and N. F. Childers. 1955. Boron toxicity induced in a New Jersey peach orchard. Part I. Proc. Amer. Soc. Hort. Sci. 66:13-20.
39. Cline, R.A. and G. Tehrani. 1973. Effects of boron and calcium sprays and of mulch on cracking in 'Italian' prune. Can. J. Plant Sci. 53:827-831.
40. Cohen, M.S. and L.S. Albert. 1974. Autoradiographic examinations of meristems of intact boron-deficient squash roots treated with tritiated thymidine. Plant Physiol. 54:766- 768.
41. Coke, L. and W.J. Whittington. 1968. The role of boron in plant growth. IV. Interrelationships between boron and indol- 3yl-acetic acid in the metabolism of bean radicles. J. Exp. Bot. 19:295-308.
42. Collings, G.H. 1927. The influence of boron on the growth of the soybean plant. Soil Sci. 23:83-105.
43. Corgan, J.N. and F.B. Widmoyer. 1971'. The effects of gibberellic acid on flower differentiation, date of bloom and flower hardiness of peach. J. Amer. Soc. Hort. Sci. 96:54- 57.
44. Crandall, P. C. andC.G. Woodbridge. 1968. Absorption of boron by 'Bartlett' pears. Proc. 19th Ann. Fert. Conf. 117- 118.
45. Crane, J.C. 1964. Growth substances in fruit setting and development. Ann. Rev. Plant Physiol. 15:303-326.
46. Crane, J.C. 1969. The. role of hormones in fruit set and development. HortScience 4:8-11.

66
47. Crane, J.C. and J.R. Hicks. 1968. Further studies on growth-regulator induced parthenocarpy in the 'Bing' cherry. Proc. Amer. Soc. Hort. Sci. 92:113-118.
48. Crane, J.C, P.E. Primer and R. C. Campbell. I960. Gibberellin induced parthenocarpy in Prunus. Proc. Amer. Soc. Hort. Sci. 75:129-137.
49- Crane, J.C, CA. Reibeiz and R. C Campbell. 1961. Gibberellin-induced parthenocarpy in the J.H. Hale peach and the probable cause of "button" production. Proc. Amer. Soc. Hort. Sci. 78:111-118.
50. Daines, R.H., S. Bachelde r and C E. Bartley. 1953. Effect of nutrition on the incidence of bacterial spot of peach. Hort. News 34:2594-2597.
51. Dear, J. and S. Aronoff. 1965. Relative kinetics of chloro- genic and caffeic acids during the onset of boron deficiency in sunflower. Plant Physiol. 40:458-459.
52. DeBruyn, J.A. 1966. The in vitro germination of pollen of Setavia sphacelata. II. Relationships between boron and certain cations. Physiol. Plant 19:322-324.
53. Degman, E.S. 1953. Effect of boron sprays on fruit set and yield of 'Anjou' pears. Proc. Amer. Soc. Hort. Sci. 62:167- 172.
54. Dennis, F.G., Jr. 1976. Trials of ethephon and other growth regulators for delaying bloom in tree fruits. J. Amer. Soc. Hort. Sci. 101:241-245.
55. Dezso, S. 1974. Influence of the flower-organization of Prunus species by rootstocks. Bot. Kozlem. 61:117-120.
56. Dible, W.T. and K.C. Berger. 1952. Boron content of alfalfa as influenced by boron supply. Proc. Soil Sci. Soc. Amer. 16:60-62.
57. Dilley, D.R. 1970. Enzymes. In A.C Hulme (ed.). Biochemistry of fruits and their products. Academic Press, New York.
58. Dorsey, M.J. 1919. Relation of weather to fruitfulness in the plum. J. Apr. Res. 17:103-126.

67
59. Dugger, W.M., Jr., T.E. Humphreys and B. Calhoun. 1967. The influence of boron on starch phosphorylase and its sig- nificance in translocation of sugars in plants. Plant Physiol. 32:364-370.
60. Dunlap, D.B. andA.H. Thompson. 1959- Effe ct of boron sprays on the development of bitter-pit in the York Imperial apple. Md. Agr. Exp. Sta. Bui. A-102.
6l. Dyar, J.J. and K. L. Webb. 196l. A relationship between boron and auxin in C Physiol. 36:672-676. boron and auxin in C*'* translocation in bean plants. Plant
62. Eaton, F.M. 1942. Deficiency, toxicity and accumulation of boron in plants. J. Agr. Res. 69:237-277.
63. Eaton, F.M., R.D. McCallum and M. S. Mayhugh. 1941. Quality of irrigation waters of the Hollister area of California with special reference to boron content and its effect on apricots and prunes. U.S. Dept. Agr. Tech. Bui. 746.
64. Eaton, G.W. 1959. A study of the megagametophyte of P. avium and its relation to fruit setting. Can. J. Plant Sci. 39:466-476.
65. Eaton, G.W. 1962. Further studies on sweet cherry embryo sacs in relation to fruit setting. Rpt. Hort. Expt. Sta. and Products Lab., Vineland, Ontario, Canada. 26-38.
66. Eaton, G.W. and A.M. Jamont. 1965. Embryo sac develop- ment in the apricot, Prunus armeniaca L. cv. Constant. Proc. Amer. Soc. Hort. Sci. 86:95-101.
67. Eaton, S.V. 1940. Effects of boron deficiency and excess on plants. Plant. Physiol. 15:95-107.
68. El-Sheikh, A.M., A. Ulrich, S. K. AwadandA.E. Mawardy. 1971. Boron tolerance of squash, melon, cucumber and corn. J. Amer. Soc. Hort. Sci. 96:536-537.
69. Ferguson, W. and L. E. Wright. 1940. Micro-element studies with special reference to the element boron. Sci. Agr. 20:470- 487.
70. Gauch, H.G. 1972. Inorganic plant nutrition. Dowden, Hutchison and Ross, Stroudsburg, Pa.

71. Gauch, H.G. andW.M. Dugger, Jr. 1953. The role of boron in the translocation of sucrose. Plant Physiol. 28:457-467.
72. Gauch, H.G. andW.M. Dugger, Jr. 1954. The physiological action of boron in higher plants: a review and interpretation. Md. Agr. Exp. Sta. Bui. A-80.
73. Glenk, H. and W. Wagner, i960. Untersuchung uber die borverteilung in einigen Oenotheren. Ber. Deut. Botan. Ges. 73:463-470.
74. Gouny, P. and F. Huguet. 1964. Biochemical changes occur- ring in pear buds before and during flowering. C.R. Acad. Agric. Fr. 50:391-399.
75. Griggs, W.H. and B.T. Iwakiri. 1975. Pollen tube growth in almond flowers. Calif. Agr. 29:4-7.
76. Grzyb, Z.S. and S.W. Zagaja. 1975. The influence of root- stock on the morphological changes in Italian prune flowers. Acta Horticulturae 48:7-11.
77. Guttridge, C.G. andJ.M. Turnbull. 1975. Improving anther dehiscence and pollen germination in strawbe rry with boric acid and salts of divalent cations. Hort. Res. 14:73-79.
78. Hair, B.L., L.W. Van Blaricom and R. A. Baumgardner. 1976. Discoloration of processed peaches as influenced by SADH and ethephon. HortScience 11:105-106.
79. Hansen, C.J. and E. L. Proebsting. 1949. Boron require- ments of plums. Proc. Amer. Soc. Hort. Sci. 53:13-20.
80. Hansen, P. 1974. The effect of boron upon leaf development and growth of the apple cultivar 'Cox's Orange Pippin.' J. Hort. Sci. 49:211-216.
81. Harrold, T.J. 1935. Comparative study of the developing and aborting fruits of Prunus persica. Bot. Gaz. 96:505-520.
82. Hasler, A. and A. Maurizio. 1949. The effect of boron on seed yield and nectar secretion in rape (B_. napus L.). Phytopath. Atschr. 15:193-207.

69
83. Hernandez, E. andN.F. Childers. 1956. Boron toxicity induced in a New Jersey peach orchard. Part II. Proc. Am. Soc. Hort. Sci. 67:121-129.
84. Hillis, W.E. and T. Swain. 1959. Phenolic constituents of Prunus domestica. III. Identification of the major constituents in the tissues, of Victoria plum. J. Sci. Food Agric. 10:533- 537.
85. Holley, K.T. and T. G. Dulin. 1939- Influence of boron on flower-bud development in cotton. J. Agr. Res. 59:541-545.
86. Horsfall, F., Jr. andG.M. Shear. 1950. Effect of boron sprays on set of apples. Va. Agr. Expt. Sta. Ann. Rpt. p. 46.
87. Imbert, M.P. and L..A. Wilson. 1972. Effects of chlorogenic acid and caffeic acid on IAA oxidase preparations from sweet potato root. Phytochemistry 11:2671-2676.
88. Jackson, D.I. 1968. Gibberellin and the growth of peach and apricot fruit. Aust. J. Biol. Sci. 21:209-215.
89- Jackson, D.I. 1968. Gibberellin and growth in stone fruits. Induction of parthenocarpy in plum. Aust. J. Biol. Sci. 21:1103-1106.
90. Jackson, J.F. and K.S.R. Chapman. 1975. The role of boron in plants, p. 213-225. In_C.A. Lamb, O. G. Bentley and J. M. Seattle (eds.). Trace elements in soil-plant-animal systems. Academic Press, New York.
91. Johnson, D. L. and L.S. Albert. 1967. Effect of selected nitrogen bases and boron on the ribonucleic acid content, elongation and visible deficiency symptoms of tomato root tips. Plant Physiol. 42:1307-1309.
92. Johnson, F., D.F. Allmendinger, V.L.. Miller and D. Polley. 1954. Fall application of boron sprays as a control for blossom blast and twig dieback of pears. Phytopathology 45:110-114.
93. Joiner, J.N. and C.A. Conover. 1974. Influence of pruning and growth-retarding compounds on forcing azaleas. Hort- Science 9:541-542.
94. Jones, H.E. and G. D. Scarseth. 1944. The calcium-boron balance in plants as related to boron needs. Soil Sci. 57:15-23.

70
95. Kamali, A.R. andN.F. Childers. 1970. Growth and fruiting of peach in sand culture as affected by boron and a fritted form of trace elements. J. Amer. Soc. Hort. Sci. 95:562-656.
96. Klein, R.M. 1951. The relation of gas exchange and tyro- sinase activity of tomato tissues to the level of boron nutrition of the plants. Arch. Bioch. 30:207-214.
97. Kouchi, H. and K. Kumazawa. 1975. Anatomical responses of root tips to boron deficiency. I. Effects of boron deficiency on elongation of root tips and their morphological charac- teristics. Soil Sci. Plant Nutr. 21:21-28.
98. Kwan, S.C., A.R. Hamson and W. F. Campbell. 1969. The effects of different chemicals on pollen germination and tube growth in Allium cepa L.. J. Amer. Soc. Hort. Sci. 94:561- 562.
99. Larson, P.R. 1962. Auxin gradients and the regulation of cambial activity, p. 97-118. InT.T. Kozlowski (ed.). Tree growth. Ronald Press, New York.
100. Layne, R.E.C. andD.J. Hagedorn. 1964. Effect of boron and agar on germination of pea pollen in sucrose media. Crop Sci. 4:39-42.
101. Lee, S. and S. Aronoff. 1967. Boron in plants: a biochemical role. Science 158:798-799.
102. Leece, D.R. 1972. Diagnostic leaf analysis for stone fruit. Errors associated with standard and simplified sampling pro- cedures. Aust. J. Exp. Agr. and Anim. Husb. 12:315-322.
103. Leece, D.R. 1975. Diagnostic leaf analysis for stone fruits. IV. Plum. Aust. J. Exp. Agr. and Anim. Husb. 15:112-117.
104. Lohnis, M.P.. 1940. Cited in; Stanley, R.G. andH.A. Linskens. 1974. Pollen biology, biochemistry, management. Springer-Verlag, New York. 307 p.
105. Lombard, P.B., M.N. Westwood and M. Thompson. 1971. Effective pollination - the facts of life behind uniform pear cropping. Ore. Hort. Soc. Ann. Rpt. 62:31-36.
106. Loomls, W.D. 1974. Overcoming problems of phenolics and quinones in the isolation of plant enzymes and organelles.
Methods in Enzymology 31:528-544.

71
107. MacVicar, R. andR.H. Burris. 1948. Relation of boron to certain plant oxidases. Arch. Biochem. 17:31-39.
108. Marsh, R.P. 1942. Comparative study of the calcium-boron metabolism of representative dicots and monocots. Soil Sci. 53:75-78.
109. Marshall, R.E. 1919. Report of three years' results in plum pollination in Oregon. Proc. Amer. Soc. Hort. Sci. 16:42-49.
110. Martin, F.W. 1953. Staining and observing pollen tubes in the style by means of fluorescence. Stain Tech. 34:125-127.
111. Martin, F.W. 1972. In vitro measurement of pollen tube growth inhibition. Plant Physiol. 49:924-925.
112. Martin, G. C. 1973. Peach fruit set and abscission. Acta Hort. 34:345-351.
113. Mcllrath, W. J. 1965. Mobility of boron in several dicotyle- donous species. Bot. Gaz. 126:27-30.
114. Mellenthin, W.M., C.Y. Wang and S. Y.Wang. 1972. Influence of temperature on pollen tube growth and initial fruit development in 'd'Anjou' pear. HortScience &:;557-559.
115. Montgomery, F.H. 1951. The effect of boron on the growth and seed production of alsike clover, Trifolium hybridum L. Can. J. Bot. 29:597-606.
116. Mukherjee, K.L. 1969- Microelement composition of sugar- cane leaves during their growth and senescence. J. Indian Bot. Soc. 47:180-184.
117. Neales, T.F. I960. Some effects of boron on root growth. Aust. J. Biol. Sci. 13:232-248.
118. Nicotra, A., C. Damiano, D. Cobianchi, L. Moser and W. Faedi. 1976. Indagine pomologica ed agronomica su 91 varieta di susino. Frutticoltura 38:5-52.
119. Nitsch, J. P. 1970. Hormonal factors in growth and develop- ment, p. 427-472. In_R.M. Klein (ed.). The biochemistry of fruits and their products, Vol. I. Academic Press, London.

72
120. Numi, Y. 1973. Influence of iron, boron and sugars in viable seed production in excised ovule culture in Petunia hybrida. J. Jap. Soc. Hort. Sci. 42:163-169-
121. Oertli, J.J. andW.F. Richardson. 1970. The mechanism of boron immobility in plants. Physiol. Plant. 23:108-116.
122. Oertli, J.J. and J.A. Roth. 1969. Boron nutrition of sugar beet, cotton and soybean. Agron. J. 61:191-195.
123. Okamoto, G. and A. Kobayashi. 1971. Effects of shoot pinching and boron spray on the nutrient content and berry set of Muscat Alexandria II. J. Jap. Soc. Hort. Sci. 40:212-224.
124. O'Kelley, J.C. 1957. Boron effects on growth, oxygen uptake and sugar absorption by germinati pollen. Amer. J. Bot. 44:239-244.
125. Perkins, H.J. 1957. Chemistry of boron in plants. Thesis, Iowa State College. Cited in; Rajaratham, J.A. and J.B. Lowry. 1974. The role of boron in the oil palm (Elaeis guineensis). Ann. Bot. 38:193 = 200.
126. Phillips, W.R. andF.B. Johnston. 1942. The effect of boron application on the subsequent storage and physiological behavior of Mclntosh apples. Sci. Agr. 23:451-460.
127. Powell, L.E. and C. Pratt. 1966. Growth promoting sub- stances in the developing fruit of peach (Prunus persica L. ) . J. Hort. Sci. 41:331-348.
128. Proebsting, E.L.. 1971. Quality of Early Italian prunes. Proc. Wash. State Hort. Assn. 1J-3J.
129. Proebsting, E.L., Jr. andH.H. Mills. 1966. Effect of gibberellic acid and other growth regulators on quality of Early Italian prunes (Prunus domestica L.). Proc. Amer. Soc. Hort. Sci. 89:135-139-
130. Proebsting, E.L., Jr. andH.H. Mills. 1976. Ethephon increases cold hardiness of sweet cherry. J. Amer. Soc. Hort. Sci. 101:31-33.
131. Rajaratnam, J.A. and J.B. Lowry. 1974. The role of boron in the oil-palm (Elaeis guineensis). Ann. Bot. 38:193-200.

73
132. Reed, H.S. 1947. A physiological study of boron deficiency in plants. Hilgardia 17:377-383.
133. Robertson, G.A. and B.C. Loughman. 1974. Reversible effects of boron on the absorption and incorporation of phosphate in Vicia faba L. New Phytol. 73:291-298.
134. Robinson, T. 1975. The organic constituents of higher plants. Cordus Press, Amherst, Mass. 347 p.
135. Rogers, B.L., A.H. Thompson, L.E. Scott and G. J. Stadelbacher. 1976. Effects of boron and succinic acid 2, 2-dimethylhydrazide (SADH) on fruitfulness and storage behavior of 'Magness' pears. Fruit Varieties Journal 30:74- 77.
136. Russell, D.W. andA.W. Galston. 1969- Blockage by gibberellic acid of phylochrome effects on growth, auxin responses and flavonoid synthesis in etiolated pea internodes. Plant Physiol. 44:1211-1216.
14 137. Schneider, G.W. 1975. C-sucrose translocation in apple. J. Amer. Soc. Hort. Sci. 100:22-24.
138. Schneider, E.A. and F. WLghtman. 1974. Metabolism of auxin in higher plants. Ann. Rev. Plant Physiol. 25:487-513.
139. Schuman, G.E., M.A. Stanley and D. Knudson. 1973. Automated total nitrogen analysis of soil and plant samples. Soil Sci. Soc. Amer. 37:480-481.
140. Scott, L.E. and A. L. Schrader. 1947. Effect of alternating conditions of boron nutrition upon growth and boron content of grapevines in sand culture. Plant Physiol. 22:526-537.
141. Senter, S. D., B. G . Lyon and B. D. Horton. 1975. Effects of different concentrations of succinic acid 2, 2-dimethylhydrazide on the flavor of puree from fresh and canned freestone peaches. J. Food Sci. 40:1103-1104.
142. Seth, A.K. and P.F. Wareing. 1967. Hormone-directed transport of metabolites and its possible role in plant senescence. J. Exp. Bot. 18:64-77.
143. Shive, J.W. 1941. Significant roles of trace elemente in the nutrition of plants. Plant Physiol. 16:435-445.

74
144. Skok, J. 1958. The role of boron in the plant cell. p. 227- 243. In_C.A. Lamb, O.G. Bentley and M. J. Beattie (ed.). Trace elements. Academic Pres , New York.
145. Skol'nik, M. Ya. 1974. General conception of the physiologi- cal role of boron in plants. Fiziologiya Rastenii 21:174-186.
146. Smith, P.F. and W. Reuther. 1951. The response of young Valencia orange trees to differential boron supply in sand culture. Plant Physiol. 26:110-114.
147. Sparks, D. and J.A. Payne. 1976. Bud break in pecan following boron toxicity. HortScience 11:494.
148. Spurr, A.R. 1957. Boron in morphogenesis of plant cell walls. Science 126:78-80.
149. Stanley, R.G. andE.A. Lichtenberg. 1963. The effect of various boron compounds on in vitro germination of pollen. Physiol. Plant. 16:337-346.
150. Stanley, R.G. andH.F. Linskens. 1965. Protein diffusion from germinating pollen. Physiol. Plant. 18:47-53.
151. Stanley, R.G. andH.F. Linskens. 1974. Pollen; biology, biochemistry, management. Springer-Verlag, New York. 307 p.
152. Stanley, R.G. andF.A. Loewus. 1964. Boron and myo- inositol in pollen pectin biosynthesis. p. 128-132. In_ H.F. Linskens (ed. ). Pollen physiology and fertilization. North Holland, Amsterdam.
153. Stebbins, R.L., H.G. Gardner and M. H. Chaplin. 1972. Oregon State University Fertilizer Guide for Prunes, FG24.
154. Steinbeck, O. 1951. Untersuchungen uber Bormangeler scheinungen bei Kartoffeln. Bodenultur 5:57-60.
155. Stenlid, G. 1963. The effect of flavonoid compounds on oxidative phos phorylation and on the enzymatic destruction of indole acetic acid. Physiol. Plant. 16:110-120.

75
156. Stonier, T., R.W. Singer and H. Yang. 1970. Studies on auxin protectors. IX. Inactivation of certain protectors by polyphenol oxidase. Plant Physiol. 46:454-457.
157. Stonier, T. 1970. The role of auxin protectors in autonomous growth. In_M.L.. Hirth and G. Morel (eds.). Les cultures de tissus de plantes. Colloq. Int. Cent. Nat. Rech. Sci. 193:423-435.
158. Stott, K.G., C.J. Jeffries and C. Jago. 1973. Pollination and fruit set in plums. Long Ashton Report 1973. p. 21-22.
159. Sullivan, D.T. andF.B. Widmoyer. 1970. Effects of succinic acid 2, 2-dimethylhydrazide (Alar) on bloom delay and fruit development of Delicious apples. HortScience 5:91-92.
160. Surkova, L.I. andK.P. Skipina. 1963. An embryological study of the plum Vengerka Ital'janskaya. Spornik Nauc. Rabot. Soc. Opyt. Sta. Subtrop. Juz. Plod. Kul'tur 17:145- 152.
161. Surkova, L.I. andK.P. Skipina. 1966. The embryology of certain plum varieties. Nauc. Dokl. Vyss. Sk. Biol. Nauk. 3:124-127.
162. Sutfield, R. and R. Wiermann. 1974. The significance of the anther- tapetum for the accumulation of phenylpropanoids on pollen grains. Ber. Dt. Bot. Ges. 87:167-174.
163. Tanada, T. 1974. Boron-induced bioelectrical field change in mung bean hypocotyl. Plant Physiol. 53:775-776.
164. Tomaszewski, M. and K.V. Thimann. 1966. Interactions of phenolic acids, metallic ions and chelating agents on auxin- induced growth. Plant Physiol. 41:1443-1454.
165. Thompson, A-H. andL.P. Batjer. 1950. The effect of boron in the germinating medium on pollen germination and pollen tube growth for several deciduous tree fruits. Proc. Amer. Soc. Hort. Sci. 56:227-229.
166. Thompson, M.M. and L.J. Liu. 1972. Pollination and erratic bearing in 'Italian' prune. J. Amer. Soc. Hort. Sci. 97:489-491.

76
167. Thompson, M.M. and L.J. Liu. 1973. Temperature, fruit set and embryo sac development in 'Italian1 prune. J. Amer. Soc. Hort. Sci. 98:193-197.
168. Tsukamoto, Y. and S. Matsubara. 1968. Studies on germina- tion of Chrysanthemum pollen. II. Occurrence of a germina- tion promoting substance. Plant and Cell Physiol. 9:237-245.
169. Vasil, I.K. I960. Studies on pollen germination of certain cucurbitaceae. Amer. J. Bot. 47:239-247.
170. Vasil, I.K. 1964. Studies of pollen germination of certain solanaceae. Bull. Torrey Bot. Club 91:370-377.
171. Vasil, I.K. 1964. Effect of boron on pollen germination and pollen tube growth, p. 107-119- In_H.F. Linskens (ed.). Pollen physiology and fertilization. North-Holland, Amster- dam.
172. Verner, L., W.J. Kochan, C.E. Loney, D.C. Moore and A. L. Kamal. 1962. Internal browning of Italian prunes. Idaho Agric. Expt. Sta. Res. Bui. 56. 38 p.
173. Visser, T. 1955. Germination and storage of pollen. Meded. Landbouwhogesch. Wageningen 55:1.
174. Visser, T. 1956. Proc. Kon. Nederl. Akad. Wetensch. Ser. C. 59:685. Cited in; Stanley, R.G- and H.F. Linskens (eds. ). Pollen biology, biochemistry and management. Springer-Verlag, New York. 307 p.
175. Wadleigh, C.H. andJ.W. Shive. 1939. A microchemical study of the effects of boron deficiency in cotton seedlings. Soil Sci. 47:33-48.
176. Wallace, A. 1971. Boron deficiency in 'Golden Bright1
tobacco, p. 158. In_A. Wallace (ed. ). Regulation of the micronutrient status of plants by chelating agents and other factors. UCLA, Los Angeles, Calif.
177. Wareing, P.F. and J. Patrick. 1975. Source-sink relations and the partition of assimilates in the plant. Int. Biol. Programme 3:481-499.

77
178. Watanabe, R., W. Chorney, J. SkokandS.H. Wender. 1964. Effect of boron deficiency on polyphenol production in the sunflower. Phytochemistry 3:391-393.
179- Weber, I.E. 1935. Histological characteristics of plants grown in toxic concentrations of boron. J. Agr. Res. 50:189- 194.
180. Weser, U. 1968. The inhibition of O-diphenol oxidase activity by borate and germanate. Hoppe-Seyler's Z. Physiol. Chem. 349:982-988.
181. Wester, R.E. and R. MacGruder. 1941. Effect of boron on plant growth and dry seed yield in lima bean (Phaseolus lunatus L. ). Proc. Amer. Soc. Hort. Sci. 38:472-474.
182. Westwood, M.N., E.S. Degman and J. H. Grim. 1964. Effects of long-term fertilizer and management practices on growth and yield of pears grown on a clay adobe soil. Ore. State Univ. Exp. Sta. Tech. Bull. 82.
183. Westwood, M.N. andH.O. Bjornstad. 1974. Fruit set as related to girdling, early cluster thinning and pruning of 'Anjou' and 'Cornice' pear. HortScience 9:342-344.
184. Whittington, W. J. 1957. The role of boron in plant growth. I. The effect on general growth, seed production and cytologi- cal behavior. J. Exp. Bot. 8:353-367.
185. Whittington, W.J. 1959. The role of boron in plant growth* II. The effect of growth of the radicle. J. Exp. Bot. 10:93-103.
186. Williams, C.F. and O. Veerhoff. 1940. Response of peach trees to boron. Proc. Amer. Soc. Hort. Sci. 52:88-96.
187. Williams, R.R. 1965. The effect of summer nitrogen applications on the quality of apple blossoms. J. Hort. Sci. 40:31-41.
188. Williams, R.R. and D. Wilson (eds.). 1970. Towards regulated cropping. Grower Books, London. 6l p.
189. Winfield, M.E. 1945. The role of boron in plant metabolism. III. The influence of boron on certain enzyme systems. Aust. J. Expt. Biol. Med. Sci. 23:267-272.

190. Woodbridge, C.G. andP.C. Crandall. 1967. The boron content of 'Bartlett' buds and flower clusters. Proc. Wash. State Hort. Assn. p. 187-191.
191. Woodbridge, C.G., A. Venegas andP.C. Crandall. 1971. The boron content of developing pear, apple and cherry- flower buds, J. Amer. Soc. Hort. Sci. 96:613-615.
192. Zittle, C.A. 1951. Reaction of borate with substances of biological interest. Adv. Enzymol. 12:493-527.
193. Zocca, A. and K. Ryugo. 1975. Changes in polyphenoloxi- dase activity and substrate levels in maturing 'Golden Delicious' apple and other cultivars. HortScience 10:586-587,