rapid acclimation of juvenile corals to co2‐mediated...
TRANSCRIPT

Rapid acclimation of juvenile corals to CO2-mediatedacidification by upregulation of heat shock protein andBcl-2 genes
A. MOYA,*† ‡ L. HUISMAN,*§ S . FORET, *¶ J . -P . GATTUSO,† ‡ D. C. HAYWARD,¶ E. E . BALL*¶and D. J . MILLER***
*ARC Centre of Excellence for Coral Reef Studies, James Cook University, Townsville, Qld 4811, Australia, †Laboratoired’Oc�eanographie de Villefranche, INSU-CNRS, 181 Chemin du Lazaret, 06230 Villefranche-sur-mer, France, ‡SorbonneUniversit�es, UPMC Univ. Paris 06, Observatoire Oc�eanologique, 06230 Villefranche-sur-mer, France, §Section of Computational
Science, Universiteit van Amsterdam, Science Park 904, 1098 XH, Amsterdam, The Netherlands, ¶Evolution, Ecology and
Genetics, Research School of Biology, Australian National University, Bldg. 46, Canberra, ACT 0200, Australia, **School of
Pharmacy and Molecular Sciences, James Cook University, Townsville, Qld 4811, Australia
Abstract
Corals play a key role in ocean ecosystems and carbonate balance, but their molecular
response to ocean acidification remains unclear. The only previous whole-transcriptome
study (Moya et al. Molecular Ecology, 2012; 21, 2440) documented extensive disruption of
gene expression, particularly of genes encoding skeletal organic matrix proteins, in juve-
nile corals (Acropora millepora) after short-term (3 d) exposure to elevated pCO2. In this
study, whole-transcriptome analysis was used to compare the effects of such ‘acute’ (3 d)
exposure to elevated pCO2 with a longer (‘prolonged’; 9 d) period of exposure beginning
immediately post-fertilization. Far fewer genes were differentially expressed under the
9-d treatment, and although the transcriptome data implied wholesale disruption of
metabolism and calcification genes in the acute treatment experiment, expression of most
genes was at control levels after prolonged treatment. There was little overlap between
the genes responding to the acute and prolonged treatments, but heat shock proteins
(HSPs) and heat shock factors (HSFs) were over-represented amongst the genes respond-
ing to both treatments. Amongst these was an HSP70 gene previously shown to be
involved in acclimation to thermal stress in a field population of another acroporid coral.
The most obvious feature of the molecular response in the 9-d treatment experiment was
the upregulation of five distinct Bcl-2 family members, the majority predicted to be anti-
apoptotic. This suggests that an important component of the longer term response to ele-
vated CO2 is suppression of apoptosis. It therefore appears that juvenile A. milleporahave the capacity to rapidly acclimate to elevated pCO2, a process mediated by upregula-
tion of specific HSPs and a suite of Bcl-2 family members.
Keywords: Acropora millepora, Bcl-2, caspases, climate change, corals, heat shock proteins
Received 5 September 2014; revision received 17 November 2014; accepted 20 November 2014
Introduction
Whilst there is no doubt that coral reefs are in global
decline, it is not yet clear how much climate change
will influence their fate. One widely propagated view is
that the dual impacts of continuing high levels of CO2
emission—elevated seawater temperatures and decreas-
ing ocean pH—will, to use a phrase coined in a some-
what different context (Pandolfi et al. 2005), inevitably
push coral reefs down the ‘slippery slope to slime’. Sea
surface temperatures have risen by a global average of
0.6 °C over the last century (Roemmich et al. 2012) and
Correspondence: Aurelie Moya, Fax: +61 7 4781 6722;
E-mail: [email protected] and David J. Miller,
Fax: +61 7 4781 6078; E-mail: [email protected]
© 2014 John Wiley & Sons Ltd
Molecular Ecology (2015) 24, 438–452 doi: 10.1111/mec.13021

are projected to rise by a further 3.0 °C by the end of
the 21st century under the RCP8.5 scenario (Collins
et al. 2013). In addition, ocean pH has decreased by
0.1 unit since the 1950s and is projected to decrease a
further 0.2–0.3 units by the end of this century (Orr
2011). Similar projections based on earlier studies gave
rise to the publication of an influential study (Hoegh-
Guldberg et al. 2007), predicting the inevitable demise
of coral reefs under ‘business as usual’ CO2 emission
scenarios, and these predictions have been supported
by some recent mesocosm experiments (Dove et al.
2013). However, other studies, based on a diversity of
sources and a longer term perspective of hundreds of
million years, suggest that change will be slower and
less dramatic than suggested by the studies mentioned
above (Pandolfi et al. 2011). In view of these continuing
uncertainties, it is therefore crucial to understand the
response of corals to ocean acidification (OA) at the
molecular level.
A major limitation in predicting the fate of coral reefs
is our lack of understanding of the molecular responses
of corals, even to individual stressors, and this is partic-
ularly the case with respect to OA. Most research to
date on the impact of OA on corals has focused on
growth and rates of survival. Considerably less atten-
tion has been paid to the molecular mechanisms
involved in the response to elevated CO2, which is pre-
dicted to decrease calcification rates in corals and other
calcifying organisms, as more energy should be
required due to the lower availability of carbonate ions
(Gattuso et al. 1999; Doney et al. 2009).
In contrast with the paucity of molecular data avail-
able, the broader biological literature detailing coral
responses to OA is large and variable in both methodol-
ogies and results, even for early developmental stages.
The recent literature on this restricted topic is summa-
rized by Albright (2011) and is tabulated in Chua et al.
(2013a) and Cumbo et al. (2013), the latter listing
approximately 20 studies dealing with the effects of
warming and acidification, either alone or in combina-
tion. Although the experiments are heterogeneous in
design, length and execution, the general picture that
emerges is that warming has a much greater immediate
effect on larval development and survival than CO2
concentration does. In general, acidification alone led to
few and small effects. Amongst the exceptions are the
results of Albright et al. (2010) on Acropora palmata and
Albright & Langdon (2011) on Porites asteroides. In both
studies, OA had a significant effect on survival and
growth post-settlement. However, the reduction in
growth reported in these two studies is not necessarily
inconsistent with other studies which reported little or
no effect, as the former experiments lasted considerably
longer (50 d for A. palmata and 49 d for P. asteroides).
This much greater length of experiment would allow
small growth effects, not apparent in shorter experi-
ments, to summate into larger ones. Different experi-
mental designs were also the cause of some of the
variable responses of early developmental stages to OA,
with some unanticipated variables proving important.
For example, whether the tiles used for settlement were
conditioned under ambient or acidified conditions
resulted in quite different bacterial communities and
settlement successes (Doropoulos et al. 2012).
Few experiments have tested the effects of OA on
gene expression, and they involved a limited group of
candidate genes. These studies are summarized in
Table 1, with further discussion below when warranted.
Kaniewska et al. (2012) used EST microarrays (Grasso
et al. 2008) to investigate the impact of chronic (28 d)
exposure to elevated pCO2 on adult colonies of Acropora
millepora. They concluded that the most significant
impacts were on genes involved in respiration and sym-
biosis rather those directly involved in calcification. The
most comprehensive molecular investigation of the
impacts of elevated CO2 on corals (Moya et al. 2012)
studied the ‘acute’ (3 d) response of juvenile A. mille-
pora to 380 and 750 ppm CO2(air) (448 and 677 latm) on
the whole transcriptome and revealed considerable dis-
ruption in levels of expression of metabolic genes as
well as of many genes implicated in skeleton deposi-
tion. In that earlier study, primary polyps were exposed
for 3 d to elevated CO2 starting 1 day after settlement,
when calcification was just beginning. To investigate
what happens over a longer 9-day period, but with
exposure starting at fertilization, this study was carried
out using the same species, facilities and equipment. In
both cases, the experiment was terminated at 4 d after
settlement (total exposure time of 3 and 9 d for the
‘acute’ and ‘prolonged’ treatment experiments, respec-
tively). In each year, levels of gene expression were
compared between the two treatments (control and
acidified treatments) after mapping the individual reads
onto the reference transcriptome (Moya et al. 2012). Pre-
liminary analysis led us to focus specifically on the heat
shock and apoptotic repertoires, which have also been
implicated in the responses to thermal stress (Kenkel
et al. 2013; Polato et al. 2013) and in thermal stress toler-
ance of natural populations (Barshis et al. 2013; Palumbi
et al. 2014). This study is not only the first to document
the response of a complete coral gene set to longer term
CO2 stress, but also the first to dissect the responses of
individual heat shock protein (HSP) and Bcl-2 isoforms
in what are complex gene repertoires. This latter point
is critical, because these are typically large gene families
in metazoans, only a few members of which may
respond to specific stressors. For example, the Acropora
digitifera genome encodes 23 distinct HSP70 family
© 2014 John Wiley & Sons Ltd
RAPID ACCLIMATION OF JUVENILE CORALS TO OA 439

members (Shinzato et al. 2012a), only a small number of
which are likely to be stress responsive (Richter et al.
2010).
The transcriptomic response of A. millepora juveniles
to prolonged CO2 stress is quite different to that
observed under acute CO2 stress. Whereas many genes
responded under acute exposure to CO2, suggesting
that growth and skeletogenesis would be severely dis-
rupted, expression data implied that most metabolic
processes had adjusted to 750 ppm CO2(air) (677 latm)
after 9 d of exposure. The acclimation to elevated pCO2
implied by these results appears to be facilitated by
higher levels of expression of a core set of HSPs and
Bcl-2 family members. Consistent with predictions, only
a small number of HSP70 family members respond to
CO2 stress; one of the two that are up-regulated under
prolonged CO2 stress is also implicated in thermal
stress tolerance in natural populations of Acropora hya-
cinthus (Barshis et al. 2013; Palumbi et al. 2014) and may
thus prove useful as a molecular marker for stress in
general. Although at least some Acropora species are
apparently able to rapidly acclimate to either CO2 or
thermal stress, it would be premature to immediately
revise the predictions concerning the future of coral
reefs, as multiple stressors may have more damaging
impacts, acclimation alone may not equip natural popu-
lations to survive frequent and severe thermal stress,
and species may differ in their ability to acclimate.
However, given the dominance of Acropora in the Indo-
Pacific, the ability of at least some species to rapidly
acclimate to stress constitutes a glimmer of hope for the
survival of corals and the maintenance of coral reefs.
Methods
Maintenance of coral material
Prior to the spawning event in November 2010, fifteen
adult colonies of Acropora millepora were collected off the
coast of Orpheus Island, Queensland, Australia (under
GBRMPA Permit No G10/33174.1). These were trans-
ported to the aquaculture facilities at James Cook Univer-
sity in Townsville where they were maintained in outdoor
flow-through aquaria filled with 5 lm filtered seawater.
Eggs and sperm were collected from each of these 15 colo-
nies individually, immediately after spawning, and used
to make three independent crosses (each cross containing
eggs and sperm from five colonies) corresponding to three
biological replicates. Each biological replicate was split
between a control and an experimental tank.
Experimental design
For the 9-d exposure experiment, embryos were trans-
ferred immediately after fertilization to seawater that
had been equilibrated with air containing 380 ppm
Table 1 Summary of published data on the effects of ocean acidification on gene expression in juvenile corals
Authors Species Genes
Length of
experiment (d)
Atmospheric
CO2 level (ppm) Result
Nakamura
et al. (2012)
Acropora digitifera HSF1
HSP70
HSP90
7 1000 No significant difference
No significant difference
No significant difference
Putnam et al. (2013) Pocillopora
damicornis
HSP70 9 600 Up-regulated
Ogawa et al. (2013) Acropora aspera CoCA2
CoCA3
Glycogen synthase
Glycogen
phosphorylase
GADPH
1, 4, 6, 14 194–756 General pattern: up-regulated
at 6 d and down-regulated
at 14 d
Kaniewska
et al. (2012)
Acropora millepora Microarray
containing 8606
unigene clusters
(Grasso et al. 2008)
0, 1, 28 600–790and 1010–1350
Discussed in the
present paper
Moya et al. (2012) Acropora millepora Entire transcriptome 3 750 and 1000 Discussed in the
present paper
Moya et al.
(present study)
Acropora millepora Entire transcriptome
(Moya et al. 2012)
3, 9 750
The Kaniewska et al. (2012) study is included as it was on Acropora millepora and is thus directly relevant to the present work. HSPs,
heat shock proteins; HSFs, heat shock factors.
© 2014 John Wiley & Sons Ltd
440 A. MOYA ET AL.

(pHNBS~8.16) or 750 ppm (pHNBS~7.96) CO2, reflecting
the control condition and medium CO2 scenario for
the 21st century. Note that, in the case of the short-
term (‘acute’) exposure experiment, larvae were culti-
vated under ambient conditions (380 ppm pCO2(air),
pHNBS~8.16) and transferred to the experimental
tanks 24 h after settlement for 3 d of CO2 exposure
(Moya et al. 2012). As in the previous experiment,
settlement was induced 5 d after fertilization by the
introduction of unglazed terracotta tiles that had
been conditioned with crustose coralline algae under
ambient seawater conditions and then autoclaved.
Figure 1 summarizes the experimental design used
for both the earlier (Moya et al. 2012) and present
experiments. Exposure to 750 ppm of CO2 had no
obvious effect on embryonic development, larval sur-
vivorship and metamorphosis of A. millepora larvae
(Chua et al. 2013a,b).
The desired CO2 concentration was produced using a
CO2 mixing system developed by Munday et al. (2009).
pH was measured on the National Bureau of Standards
(NBS) scale with a portable metre (Hach HQ11D) cali-
brated daily with NBS buffers (pH 4 and 7). Tempera-
ture, pH, oxygen concentration and total alkalinity were
monitored daily at 11:00. The seawater chemistry was
estimated using CO2SYS (Lewis & Wallace 1998). Average
pCO2(seawater) was estimated to be 448 and 677 latm for
the 9-d experiment. Seawater carbonate parameters are
shown in Table 2.
Sampling and RNA extraction
The primary polyps (~50 per biological sample and per
condition) were carefully removed from the tiles (1–3tiles per condition) with a sterile scalpel, pooled and
immediately snap-frozen in liquid nitrogen and stored
at �80 °C until further processed. Total RNA was
extracted from each sample of 50 using TRIzol Reagent
(Invitrogen) according to the Chomczynski method
(Chomczynski & Sacchi 1987) and dissolved in RNase-
free water. RNA quantity and quality were assessed
using a NanoDrop ND-1000 spectrometer and denatur-
ing gel electrophoresis using standard methods (Sam-
brook & Russell 2001). Before being shipped on dry ice
to the Macrogen sequencing facilities in Seoul, South
Korea, each RNA sample was precipitated in ethanol
and sodium acetate (29 and 0.19 sample volume,
respectively) and stored at �80 °C.
High-throughput sequencing and data analysis
mRNA isolation and library construction were per-
formed by Macrogen (South Korea). The libraries were
sequenced using the Illumina HiSeq2000 platform, pro-
ducing an average across the six samples of 19 million
reads per sample (100 bp) for the 9-d CO2 exposure.
Sequencing reads were mapped onto the A. millepora
transcriptome assembly (Moya et al. 2012) using the BOW-
TIE mapping software version 0.12.7 (Langmead et al.
2009). Differential gene expression was inferred based on
C1 C2 C3
Ac1 Ac2 Ac3
C1 C2 C3
Pr1 Pr2 Pr3
380 ppm CO2 (air)
750 ppm CO2 (air)
3 day elevated CO2
ACUTE
Sampling
380 ppm CO2 (air)
750 ppm CO2 (air)
Sampling
PROLONGED
Fertilization
PlanulaEgg
9 day exposure to elevated CO2
Basal platedeposition
Fully developedcalcification
Settlementday 5
Fig. 1 Summary of the experimental design. For the prolonged (9-d) exposure to pCO2, embryos were transferred immediately after
fertilization to seawater that had been equilibrated with air containing 380 ppm (pH~8.16) or 750 ppm (pH~7.96), reflecting the con-
trol condition and medium CO2 scenario for the 21st century. For the short-term (‘acute’) exposure described in Moya et al. (2012),
larvae were cultivated under ambient conditions and transferred to the experimental tanks 24 h after settlement for 3 d of CO2 expo-
sure. In both cases, settlement was induced 5 d after fertilization by the introduction of unglazed terracotta tiles conditioned with
crustose coralline algae under ambient seawater conditions. Four days later, primary polyps (~50 per condition, three biological
replicates per condition) were snap-frozen in liquid nitrogen prior to RNA extraction for RNA-seq analysis. C, Control; Ac, Acute;
Pr, Prolonged.
© 2014 John Wiley & Sons Ltd
RAPID ACCLIMATION OF JUVENILE CORALS TO OA 441

these mapping counts using the edgeR package (Robin-
son et al. 2010). Gene ontology (GO) enrichment analysis
was performed using the GOseq package (Young et al.
2010) that accounts for selection bias such as gene size or
expression level. The RNA-seq reads used in this study
have been submitted to the NCBI Gene Expression Omni-
bus (GEO) database under Accession no. GSE61114. The
corresponding data for the ‘acute’ treatment experiment
are available from the same source under Accession no.
GSE33016. Note that, rather than a common reference for
both the 3- and 9-d experiments, a distinct set of controls
was employed for the 9-day experiment.
For each gene category of interest, BlastP and
HMMER domain searches (e-value cut-off = 1e�5) were
performed on the A. millepora transcriptome (Moya et al.
2012). An additional blast onto the NCBI nr database
confirmed the identification of each sequence.
Results
Whole-transcriptome analysis was used to compare Acro-
pora millepora juveniles that had been exposed to 380 or
750 ppm CO2(air) (414 or 669 latm) for a period of 9-d
(‘prolonged’ exposure—this experiment), as shown sche-
matically in Fig. 1, and these data were compared with
results of a previous 3-d ‘acute’ exposure treatment, car-
ried out using the same species and experimental format
(Moya et al. 2012). The previous (Moya et al. 2012) and
present experiments allow comparison of gene expres-
sion levels in primary polyps at the same time point, rela-
tive to their respective controls, but after different
periods of exposure to elevated pCO2. In the acute treat-
ment, elevated CO2 stress was initiated 24 h after settle-
ment (Moya et al. 2012), whereas in the case of prolonged
exposure, fertilized eggs were immediately exposed to
elevated CO2, with the resulting polyps then harvested at
the same age as in the acute experiment.
The transcriptomic responses to the acute and 9-d CO2
treatment are largely discrete
The transcriptional response to 9-d CO2 exposure dif-
fers both quantitatively and qualitatively to that under
acute exposure. Using a cut-off of adjusted P < 0.05,
20% of A. millepora transcripts were differentially
expressed in the acute treatment, whereas only 4% of
transcript clusters responded in the 9-d treatment group
(Fig. 2A). The prolonged exposure resulted in a pre-
dominantly positive transcriptional response, whereas
similar numbers of transcript clusters were up- and
down-regulated under acute exposure to CO2. More-
over, the 3- and 9-d responses were largely discrete
Table 2 Summary of seawater parameters in the control and acidified treatments
pCO2(air) (ppm) pHNBS Total alkalinity (lmol/kg) Temperature (°C) Ωaragonite pCO2(seawater) (latm)
Control 8.16 � 0.01 2028.8 � 5.0 27.5 � 0.2 2.87 � 0.08 414.2 � 2.0
pCO2 acute 7.96 � 0.01 1876.7 � 4.0 28.0 � 0.1 1.80 � 0.05 669.2 � 2.0
Control 8.12 � 0.07 2076.2 � 140.0 26.5 � 0.3 2.80 � 0.48 447.5 � 64.5
pCO2 chronic 7.96 � 0.06 2015.5 � 129.0 26.5 � 0.2 1.99 � 0.28 677.4 � 90.8
The saturation states of aragonite (Ωa) and pCO2 were estimated from pHNBS and total alkalinity using the computer package CO2SYS.
Values are mean � SD. NBS, National Bureau of Standards.
Up-regulated Down-regulated Non-regulated
11%
9%
80%
Acute
3 day, 750 ppm CO2 (air)
<1%3%
97%
Prolonged
9 day, 750 ppm CO2 (air)
305
Ac
5334 Pr1422
Shared up-regulatedgenes
36
Shared down-regulatedgenes
Ac
4812125
Pr
(A)
(B)
Fig. 2 Effects of the elevated CO2 treatments on the Acropora
millepora transcriptome. Part (A) of the figure shows the per-
centages of transcripts up- (green) and down-regulated (red) in
response to acute (3-d) and prolonged (9-d) CO2 treatments
(adjusted P < 0.05). The Venn diagrams shown in (B) indicate
the numbers of up-regulated (green) and down-regulated (red)
transcripts in the acute and prolonged CO2 treatments
(adjusted P < 0.05), as well as those common to both. Ac,
Acute; Pr, Prolonged.
© 2014 John Wiley & Sons Ltd
442 A. MOYA ET AL.
(Fig. 2B); only 341 transcript clusters (0.65% of clusters;
305 and 36 up- and down-regulated, respectively)
responded under both acute and prolonged exposure to
CO2. Both sets of differentially expressed genes were
enriched in genes of unknown function (no blast hit
with a cut-off of 1e�5, possibly coral-specific genes),
compared to the transcriptome database. The percent-
ages of genes with unknown function were 56% and
47% in the 3- and 9-d treatments, respectively, com-
pared to 35% in the transcriptome database. Although
this result suggests the involvement of coral-specific
mechanisms, the fact that these genes lack annotation
precludes speculation concerning their nature.
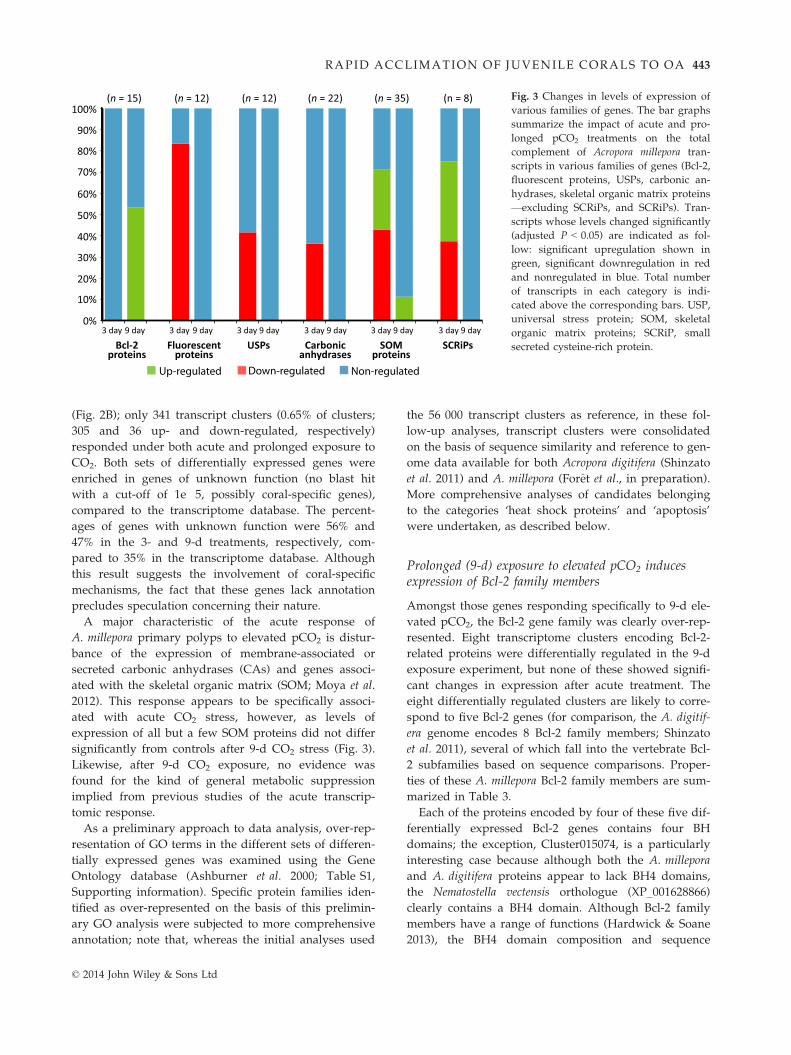
A major characteristic of the acute response of
A. millepora primary polyps to elevated pCO2 is distur-
bance of the expression of membrane-associated or
secreted carbonic anhydrases (CAs) and genes associ-
ated with the skeletal organic matrix (SOM; Moya et al.
2012). This response appears to be specifically associ-
ated with acute CO2 stress, however, as levels of
expression of all but a few SOM proteins did not differ
significantly from controls after 9-d CO2 stress (Fig. 3).
Likewise, after 9-d CO2 exposure, no evidence was
found for the kind of general metabolic suppression
implied from previous studies of the acute transcrip-
tomic response.
As a preliminary approach to data analysis, over-rep-
resentation of GO terms in the different sets of differen-
tially expressed genes was examined using the Gene
Ontology database (Ashburner et al. 2000; Table S1,
Supporting information). Specific protein families iden-
tified as over-represented on the basis of this prelimin-
ary GO analysis were subjected to more comprehensive
annotation; note that, whereas the initial analyses used
the 56 000 transcript clusters as reference, in these fol-
low-up analyses, transcript clusters were consolidated
on the basis of sequence similarity and reference to gen-
ome data available for both Acropora digitifera (Shinzato
et al. 2011) and A. millepora (Foret et al., in preparation).
More comprehensive analyses of candidates belonging
to the categories ‘heat shock proteins’ and ‘apoptosis’
were undertaken, as described below.
Prolonged (9-d) exposure to elevated pCO2 inducesexpression of Bcl-2 family members
Amongst those genes responding specifically to 9-d ele-
vated pCO2, the Bcl-2 gene family was clearly over-rep-
resented. Eight transcriptome clusters encoding Bcl-2-
related proteins were differentially regulated in the 9-d
exposure experiment, but none of these showed signifi-
cant changes in expression after acute treatment. The
eight differentially regulated clusters are likely to corre-
spond to five Bcl-2 genes (for comparison, the A. digitif-
era genome encodes 8 Bcl-2 family members; Shinzato
et al. 2011), several of which fall into the vertebrate Bcl-
2 subfamilies based on sequence comparisons. Proper-
ties of these A. millepora Bcl-2 family members are sum-
marized in Table 3.
Each of the proteins encoded by four of these five dif-
ferentially expressed Bcl-2 genes contains four BH
domains; the exception, Cluster015074, is a particularly
interesting case because although both the A. millepora
and A. digitifera proteins appear to lack BH4 domains,
the Nematostella vectensis orthologue (XP_001628866)
clearly contains a BH4 domain. Although Bcl-2 family
members have a range of functions (Hardwick & Soane
2013), the BH4 domain composition and sequence
Bcl-2proteins
Fluorescentproteins
USPs Carbonicanhydrases
SOMproteins
SCRiPs3 day 9 day 3 day 9 day 3 day 9 day 3 day 9 day 3 day 9 day 3 day 9 day
100%
90%
80%
70%
60%
50%
40%
30%
20%
10%
0%
(n = 15) (n = 12) (n = 12) (n = 22) (n = 35) (n = 8)
Up-regulated Down-regulated Non-regulated
Fig. 3 Changes in levels of expression of
various families of genes. The bar graphs
summarize the impact of acute and pro-
longed pCO2 treatments on the total
complement of Acropora millepora tran-
scripts in various families of genes (Bcl-2,
fluorescent proteins, USPs, carbonic an-
hydrases, skeletal organic matrix proteins
—excluding SCRiPs, and SCRiPs). Tran-
scripts whose levels changed significantly
(adjusted P < 0.05) are indicated as fol-
low: significant upregulation shown in
green, significant downregulation in red
and nonregulated in blue. Total number
of transcripts in each category is indi-
cated above the corresponding bars. USP,
universal stress protein; SOM, skeletal
organic matrix proteins; SCRiP, small
secreted cysteine-rich protein.
© 2014 John Wiley & Sons Ltd
RAPID ACCLIMATION OF JUVENILE CORALS TO OA 443
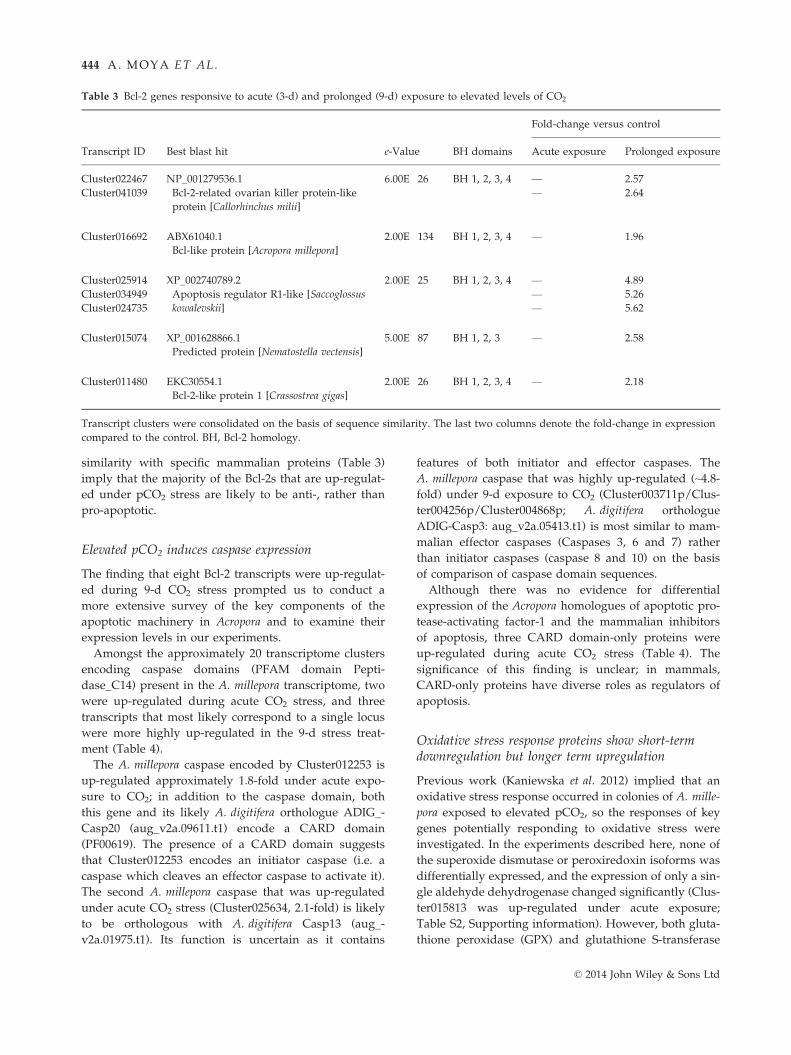
similarity with specific mammalian proteins (Table 3)
imply that the majority of the Bcl-2s that are up-regulat-
ed under pCO2 stress are likely to be anti-, rather than
pro-apoptotic.
Elevated pCO2 induces caspase expression
The finding that eight Bcl-2 transcripts were up-regulat-
ed during 9-d CO2 stress prompted us to conduct a
more extensive survey of the key components of the
apoptotic machinery in Acropora and to examine their
expression levels in our experiments.
Amongst the approximately 20 transcriptome clusters
encoding caspase domains (PFAM domain Pepti-
dase_C14) present in the A. millepora transcriptome, two
were up-regulated during acute CO2 stress, and three
transcripts that most likely correspond to a single locus
were more highly up-regulated in the 9-d stress treat-
ment (Table 4).
The A. millepora caspase encoded by Cluster012253 is
up-regulated approximately 1.8-fold under acute expo-
sure to CO2; in addition to the caspase domain, both
this gene and its likely A. digitifera orthologue ADIG_-
Casp20 (aug_v2a.09611.t1) encode a CARD domain
(PF00619). The presence of a CARD domain suggests
that Cluster012253 encodes an initiator caspase (i.e. a
caspase which cleaves an effector caspase to activate it).
The second A. millepora caspase that was up-regulated
under acute CO2 stress (Cluster025634, 2.1-fold) is likely
to be orthologous with A. digitifera Casp13 (aug_-
v2a.01975.t1). Its function is uncertain as it contains
features of both initiator and effector caspases. The
A. millepora caspase that was highly up-regulated (~4.8-fold) under 9-d exposure to CO2 (Cluster003711p/Clus-
ter004256p/Cluster004868p; A. digitifera orthologue
ADIG-Casp3: aug_v2a.05413.t1) is most similar to mam-
malian effector caspases (Caspases 3, 6 and 7) rather
than initiator caspases (caspase 8 and 10) on the basis
of comparison of caspase domain sequences.
Although there was no evidence for differential
expression of the Acropora homologues of apoptotic pro-
tease-activating factor-1 and the mammalian inhibitors
of apoptosis, three CARD domain-only proteins were
up-regulated during acute CO2 stress (Table 4). The
significance of this finding is unclear; in mammals,
CARD-only proteins have diverse roles as regulators of
apoptosis.
Oxidative stress response proteins show short-termdownregulation but longer term upregulation
Previous work (Kaniewska et al. 2012) implied that an
oxidative stress response occurred in colonies of A. mille-
pora exposed to elevated pCO2, so the responses of key
genes potentially responding to oxidative stress were
investigated. In the experiments described here, none of
the superoxide dismutase or peroxiredoxin isoforms was
differentially expressed, and the expression of only a sin-
gle aldehyde dehydrogenase changed significantly (Clus-
ter015813 was up-regulated under acute exposure;
Table S2, Supporting information). However, both gluta-
thione peroxidase (GPX) and glutathione S-transferase
Table 3 Bcl-2 genes responsive to acute (3-d) and prolonged (9-d) exposure to elevated levels of CO2
Transcript ID Best blast hit e-Value BH domains
Fold-change versus control
Acute exposure Prolonged exposure
Cluster022467 NP_001279536.1
Bcl-2-related ovarian killer protein-like
protein [Callorhinchus milii]
6.00E�26 BH 1, 2, 3, 4 — 2.57
Cluster041039 — 2.64
Cluster016692 ABX61040.1
Bcl-like protein [Acropora millepora]
2.00E�134 BH 1, 2, 3, 4 — 1.96
Cluster025914 XP_002740789.2
Apoptosis regulator R1-like [Saccoglossus
kowalevskii]
2.00E�25 BH 1, 2, 3, 4 — 4.89
Cluster034949 — 5.26
Cluster024735 — 5.62
Cluster015074 XP_001628866.1
Predicted protein [Nematostella vectensis]
5.00E�87 BH 1, 2, 3 — 2.58
Cluster011480 EKC30554.1
Bcl-2-like protein 1 [Crassostrea gigas]
2.00E�26 BH 1, 2, 3, 4 — 2.18
Transcript clusters were consolidated on the basis of sequence similarity. The last two columns denote the fold-change in expression
compared to the control. BH, Bcl-2 homology.
© 2014 John Wiley & Sons Ltd
444 A. MOYA ET AL.

isoforms showed differential expression during acute
CO2 stress. Five distinct GPX isoforms were identified in
A. millepora, two of which were down-regulated under
acute CO2 stress (Cluster018808 and Cluster011976,
�1.66 and �3.39-fold, respectively, Table S2, Supporting
information). Both Omega and Pi isoforms of GST
responded in the same way, suggesting coordinated
downregulation of at least some antioxidant defences
under acute CO2 stress, potentially facilitating a more
appropriate stress response, such as apoptosis.
Under 9-d exposure to CO2, the expression of both
the GPX and GST isoforms was at control levels, but
there was upregulation of both catalase (about twofold)
and both subunits of NADPH oxidase (1.9- and 2.2-fold,
respectively, Table S2, Supporting information) suggest-
ing induction of enzymatic antioxidant defences.
Two other multigene families, those encoding the GFP-
related (FPs) and universal stress proteins (USPs), were
over-represented amongst the genes differentially
expressed in the present analysis, but the functions of
these proteins remain unclear. A total of 12 distinct mem-
bers of the FP family were identified as encoded by the
A. millepora transcriptome (Table S3, Supporting infor-
mation). Comparison with data for A. digitifera (Shinzato
et al. 2012b) combined with BlastP analysis implies that
the 12 A. millepora FP transcripts encode two cyan FPs,
five green FPs, two chromoproteins and three red FPs.
With the exception of the two chromoproteins, all of the
FP genes identified were down-regulated specifically
under acute exposure to CO2 (on average 4.4-fold, see
Fig. 3 and Table S3, Supporting information).
The USPs were originally identified in the context of
bacterial stress response (Nystr€om & Neidhardt 1992;
reviewed in Kvint et al. 2003). In Hydra, those USP
genes for which expression data are available are
expressed in the endodermal epithelium, which acts as
a barrier for protection against intruding microbes,
suggesting potential roles of some USPs in this defen-
sive barrier (Bosch et al. 2009). Thirteen USP genes have
been identified in A. millepora (Foret et al. 2011), five of
which, including the calcification candidate Amil10,
were down-regulated in response to acute exposure to
CO2 by an average of 1.8-fold (Fig. 3 and Table S4, Sup-
porting information).
Differential expression of heat shock proteins iscommon to both acute and prolonged exposure toelevated pCO2
Although the responses to acute (3-d) and prolonged
(9-d) CO2 stress were largely discrete, HSPs and their
associated cochaperones (HSP40 and sacsin family
members) and transcription factors (the heat shock fac-
tors; HSFs) were over-represented amongst the rela-
tively few genes that comprised the common response
(Fig. 2B, Table 5 and Table S5, Supporting informa-
tion). A total of nine HSP-related transcripts (4 HSP20,
3 HSP70 and 2 HSP40 transcriptome clusters, corre-
sponding to 3 HSP20, 2 HSP70 and 1 HSP40 genes)
showed strong upregulation under both acute and 9-d
pCO2 stress (Table 5). All of the five A. millepora HSF
transcriptome clusters identified were likewise up-
Table 4 Caspases and CARD-only genes responsive to acute (3-d) and prolonged (9-d) exposure to elevated levels of CO2
Yellow background shading indicates caspases; white background indicates CARD-only proteins. Transcript clusters were consoli-
dated on the basis of sequence similarity. The last two columns denote the fold-change expression compared to the control.
© 2014 John Wiley & Sons Ltd
RAPID ACCLIMATION OF JUVENILE CORALS TO OA 445

Table 5 Heat shock protein genes responsive to acute (3-d) and prolonged (9-d) exposure to elevated levels of CO2
Transcript ID HMMER Best blast hit e-Value
Fold-change versus
control
Acute
exposure
Prolonged
exposure
HSP20 Cluster009259 HSP20 ABA42877.1
Small heat shock protein, partial [uncultured
cnidarian]
3.00E�155 6.81 3.66
Cluster017128 7.36 5.77
Cluster022217 HSP20 9 2 ABA42878.1
Small heat shock protein [uncultured cnidarian]
4.00E�178 8.07 7.88
Cluster020497 HSP20 9 2 ABA42878.1
Small heat shock protein [uncultured cnidarian]
2.00E�81 9.50 3.97
HSP40 Cluster010480 DnaJ
CTDII
XP_005112000.1
dnaJ homologue subfamily B member 1-like
[Aplysia californica]
5.00E�112 1.78 —
Cluster015487 DnaJ ABC84495.1
heat shock protein 40 [Locusta migratoria]
9.00E�111 8.87 9.00
Cluster018170 CTDII 8.66 9.02
Cluster008833 DnaJ
CTDII
XP_005296130.1
dnaJ homologue subfamily B member 1
[Chrysemys picta bellii]
2.00E�95 — 3.63
Cluster008505 DnaJ XP_007537836.1
PREDICTED: dnaJ homologue subfamily C
member 17 [Erinaceus europaeus]
3.00E�11 2.00 —
Cluster009800 DnaJ
zf-C2H2_jaz
EKC33886.1
DnaJ-like protein subfamily C member 21
[Crassostrea gigas]
3.00E�117 — 4.98
HSP70 Cluster007731 HSP70 XP_002731913.1
heat shock cognate 71 kDa protein-like
[Saccoglossus kowalevskii]
3.00E�137 5.96 7.19
Cluster005163 1.82 2.06
Cluster005613 HSPA4_like_NDB BAD89541.1
heat shock protein 70 [Pocillopora damicornis]
0.00E+00 4.48 2.72
Cluster006034 HSP70 XP_006823582.1
heat shock 70 kDa protein 12A-like [Saccoglossus
kowalevskii]
8.00E�179 3.73 —
HSP90 Cluster010802 HATPase_c_3 NP_999808.1
heat shock protein gp96 precursor
[Strongylocentrotus purpuratus]
0.00E+00 2.19 —Cluster005735 HSP90 1.60 —
Cluster022600 HSP90 2.60 —
Sacsin Cluster005249 HEPN XP_002740585.1
sacsin-like [Saccoglossus kowalevskii]
0.00E+00 1.83 —
Cluster010514 3.22 —Cluster017656 5.94 —
Cluster003566 HEPN 0.00E+00 3.27 —Cluster007958 3.65 —
© 2014 John Wiley & Sons Ltd
446 A. MOYA ET AL.

regulated under both acute and prolonged exposure to
elevated CO2 (Table 5 and Table S5, Supporting infor-
mation).
In addition to the common response, other HSPs
responded to either 3- or 9-d elevated CO2. Six HSP
transcripts (2 HSP40, 1 HSP70 and 3 HSP90,
corresponding to 2 HSP40, 1HSP70 and the single
HSP90B1/gp96 gene) were specifically up-regulated
under 3-d CO2 stress, and 2 HSP40 (2 genes) were
up-regulated only after 9-d treatment (see Table 5). A
large proportion (14) of the 22 sacsin transcriptome
clusters (8 of the 18 genes) identified were up-regulated
specifically, and a single transcript (1 gene) was
down-regulated under acute CO2 stress.
Discussion
The observed extensive differences in gene expression
patterns between the ‘acute’ (3-d) and ‘prolonged’ (9-d)
exposure treatments were unexpected and surprising
given that both reflect relatively short-term responses.
Whereas acute CO2 stress suppressed metabolism and
altered the expression of many genes encoding CA and
SOM proteins (Moya et al. 2012), expression of most
genes in those categories was at control levels in the
prolonged treatment experiment. Ogawa et al. (2013)
reported that two CAs in adult Acropora aspera were
up-regulated at 6 d, but down-regulated at 14 d. As the
length of the present experiment was intermediate
Table 5 Continued
Transcript ID HMMER Best blast hit e-Value
Fold-change versus
control
Acute
exposure
Prolonged
exposure
XP_008106211.1
PREDICTED: sacsin isoform X3 [Anolis
carolinensis]
Cluster005325 3.62 —
Cluster001033 — XP_004631088.1
PREDICTED: sacsin isoform X1 [Octodon degus]
0.00E+00 2.61 —
Cluster001972 HEPN XP_003200093.2
sacsin-like isoform X1 [Danio rerio]
0.00E+00 2.24 —
Cluster009368 HEPN XP_005516322.1
sacsin [Pseudopodoces humilis]
9.00E�66 �3.71 —
Cluster004016 HEPN EOA99087.1
sacsin, partial [Anas platyrhynchos]
0.00E+00 2.08 —Cluster010311 1.85 —
Cluster009514 2.08 —Cluster013781 3.74 —
Cluster013781 HEPN XP_007234568.1
sacsin isoform X2 [Astyanax mexicanus]
0.00E+00 3.74 —
Cluster007316 HEPN XP_006927228.1
sacsin [Felis catus]
0.00E+00 2.15 —
Cluster000374 HEPN XP_007234567.1
sacsin isoform X1 [Astyanax mexicanus]
0.00E+00 — —
Cluster003923 1.87 —Cluster002694 — —
HSF Cluster007375 HSF-type
DNA-binding
ABR15461.1
HSF [Haliotis asinina]
3.00E�65 3.07 4.45
Cluster007882 3.19 4.20
Cluster007957 3.06 4.27
Cluster009578 2.95 4.13
Cluster003398 3.12 7.58
Transcript clusters were consolidated on the basis of sequence similarity. The last two columns denote the fold-change expression
compared to the control. HSPs, heat shock proteins; HSFs, heat shock factors.
© 2014 John Wiley & Sons Ltd
RAPID ACCLIMATION OF JUVENILE CORALS TO OA 447

between those two time points, there is no basis for
commenting on the consistency/inconsistency of the
results of the two experiments.
Upregulation of multiple Bcl-2 family members was
the single most obvious feature of the 9-d exposure
treatment group. Five distinct Bcl-2 proteins were
up-regulated in the 9-d treatment group, whereas no
proteins of this type were differentially expressed in the
acute treatment group. Based on comparisons with the
well-characterized mammalian proteins, four of the five
Bcl-2 proteins are likely to be anti-apoptotic, indicating
that suppression of apoptosis is a key component of
acclimation to elevated pCO2. Conversely, two caspase
genes were up-regulated after acute exposure to ele-
vated pCO2, suggesting that apoptosis may be activated
in this treatment group. However, in apparent contra-
diction to the above interpretation of the Bcl-2 data, a
distinct caspase gene showed significant upregulation
specifically in the prolonged treatment group. Interpret-
ing the significance of the caspase data is complicated
by the ambiguous relationships of the cnidarian pro-
teins with the well-defined functional groups of verte-
brate caspases. In the case of the Bcl-2 family, several of
the coral proteins showed convincing sequence similar-
ity with specific classes from bilaterians, but the coral
caspase repertoire is far more difficult to interpret. For
example, the Acropora protein Cluster002669p has cas-
pase 8-like specificity despite having caspase 3-like resi-
dues at what are otherwise diagnostic positions
(Sakamaki et al. 2014). Although phylogenetic analyses
were equivocal, the presence of a CARD domain in one
of the caspases (Cluster012253) up-regulated during
acute CO2 stress suggests that it is likely to be an initia-
tor caspase. The caspase induced under 9-d CO2 stress
has clear counterparts in other anthozoans, and despite
equivocal relationships with the vertebrate CASP clas-
ses, these are likely to be effectors rather than initiators.
Whilst the caspase data are difficult to interpret at
this stage, the elevated expression of a suite of anti-
apoptotic Bcl-2 proteins implies suppression of apopto-
sis in the 9-d treatment group. Based on four coral spe-
cies, Tchernov et al. (2011) presented evidence of the
ability to survive moderate bleaching when the caspase-
mediated apoptotic cascade is down-regulated. Under
prolonged elevated CO2, the activation of anti-apoptotic
molecules could serve as an important mechanism
moderating the apoptotic response, as suggested by
Kvitt et al. (2011).
Fluorescent proteins (FPs) show great diversity in cor-
als (Alieva et al. 2008; Shinzato et al. 2012b) and can be
amongst the most abundant proteins (Oswald et al.
2007); however, our understanding of the role of this pro-
tein family is far from complete. One proposal is that FPs
function as a component of the oxidative stress response
through superoxide quenching (Bou-Abdallah et al. 2006)
or hydrogen peroxide scavenging (Palmer et al. 2009).
Previous work has shown that some FPs are strongly
down-regulated by heat stress (Dove et al. 2006; Smith-
Keune & Dove 2008; Rodriguez-Lanetty et al. 2009). A
similar pattern is shown in this study under acute expo-
sure to pCO2, supporting downregulation of antioxidant
defences in the short-term; however, functional studies
of individual FPs are needed to clarify their roles.
Although responses to the acute and prolonged treat-
ments were in large part mutually exclusive, HSPs and
related cochaperones and transcription factors were
over-represented amongst the small number of genes
constituting the common response (Table 5). Beyond
the common response, other members of the HSP reper-
toire responded specifically to either acute or prolonged
CO2 stress (Table 5). Nakamura et al. (2012) for Acropora
digitifera and Putnam et al. (2013) for Pocillopora damicor-
nis report no differential response by members of the
HSP70 family to acidification or acidification plus heat-
ing, respectively (note though that these authors stud-
ied effects only in presettlement stages), but we cannot
determine whether those responses are consistent with
those reported here for Acropora millepora. In the case of
A. digitifera, the sequences are not available, whilst in
the case of Pocillopora, it is unclear to which of four
A. millepora isoforms (Cluster005613, Cluster005163,
Cluster011524, Cluster007731) the HSP70 from P. dami-
cornis studied in Putnam et al. (2013; BAD89541.1) corre-
sponds (all had >70% identity and e-value = 0). The
first two of these A. millepora transcripts (Cluster005613,
Cluster005163) were up-regulated in both acute and
prolonged treatments, whereas the others were not dif-
ferentially regulated in either case.
There are a number of differences between the behav-
iour of individual genes in the study of Kaniewska et al.
(2012) compared to the present study. For example, the
downregulation of two proteins that are likely to have
anti-apoptotic activity (Bcl-2 and API-5) reported by
Kaniewska et al. (2012) contrasts with the upregulation
of a suite of anti-apoptotic Bcl-2 proteins reported here.
However, there are many differences in the design of
the two studies. Perhaps the most significant of these is
that Kaniewska et al. (2012) were studying adult
A. millepora containing symbionts, which greatly com-
plicates both the analysis and its interpretation, while
the present study was of early developmental stages
lacking symbionts. Only further study will clarify the
reason for such differences.
Whereas few studies have addressed gene expres-
sion changes in corals under elevated pCO2, thermal
stress-induced changes have received much greater
attention. The categories of genes responding to
thermal stress include HSPs and members of the apop-
© 2014 John Wiley & Sons Ltd
448 A. MOYA ET AL.

totic repertoire, and as a result, models proposed to
account for the observed changes in gene expression in
response to acidification and heating are similar.
Indeed, Polato et al. (2013) state ‘In larval and adult
acroporids, expression of heat shock proteins is typi-
cally characterized by short-term upregulation followed
by a decline in expression after prolonged exposure to
high temperatures’. However, the superficial similarity
in molecular responses may be an artefact of not tak-
ing into account the complexity of the stress response/
apoptotic repertoire—for example, at least twelve dis-
tinct HSP70 isozymes and over 20 HSP40/DNAJ pro-
teins are present in A. millepora. Here we demonstrate
that, although a core set of HSPs responds to both
short and longer term exposure to elevated pCO2, dis-
tinct HSPs respond specifically to either duration of
exposure.
Heat shock proteins have been extensively explored as
potential stress markers in corals, but with limited con-
sideration of their functional diversity (Morris et al.
2013). A preliminary survey of the HSP and stress
response repertoire of Acropora has been published
(Shinzato et al. 2012a), but the available annotation is lim-
ited. The available data can best be interpreted in the
context of conservation of function more broadly, at the
class level. Typically, the HSP70 and HSP90 classes each
consist of constitutive and inducible variants, the former
dealing with the instability of many cellular proteins
even under normal physiological conditions. On the
other hand, the HSP40/DNAJ and other small HSP pro-
teins—sometimes referred to as ‘holdases’, whereas
HSP70/HSP90 are ‘foldases’ (Richter et al. 2010)—are the
first line of the chaperone response to proteins in non-
native conformations and are typically stress induced.
The cochaperone ‘holdases’ stimulate the ATPase activity
of the ‘foldases’ (HSP70, HSP90), stabilizing the ‘fol-
dase’/substrate interaction (Richter et al. 2010).
A substantial proportion of the small HSP isoforms
identified in A. millepora were up-regulated under acute
CO2 stress, including 15 of the 22 sacsin transcripts
(Table 5). However, most of the small HSPs up-regulated
under acute CO2 stress were at control levels in the 9-d
treatment group, although some (all four differentially
expressed HSP20 transcripts; two HSP40 transcripts)
remained at higher than control levels. The ‘foldase’
activity elevated in both the 3- and 9-d treatment groups
was limited to only three of the seventeen HSP70 tran-
scripts identified, the ‘holdase’ component consisting of
four HSP20 and two HSP40 types (Table 5).
The consensus view is that the chaperone-based
stress response is largely generic—because the HSPs
respond to the presence of unfolded protein, rather than
to specific stress sensors, the expectation is that the
same foldases should respond to CO2 stress as to any
other type of stress. However, this expectation is not
consistently fulfilled when our results are compared to
previous studies. For example, Leggat et al. (2011)
reported strong upregulation of HSP90 and HSP70 in
heat-stressed A. aspera, but the A. millepora orthologues
of these isoforms (the HSP90A1 protein DY584045.1—
Cluster009669 and GO000475.1—Cluster010972, respec-
tively) were not differentially regulated in our experi-
ment. The HSP70 variant up-regulated in heat-stressed
A. millepora larvae (Rodriguez-Lanetty et al. 2009) and
in immune challenged adults of the same species
(Brown et al. 2013) corresponds to the A. millepora
Cluster006117 (a likely gp78 orthologue), which was not
differentially expressed in response to CO2. DeSalvo
et al. (2008) reported upregulation of an HSP90 variant
in Montastraea faveolata during heat stress; whilst this
sequence is most similar to A. millepora HSP90A1, the
EST is too short to be unequivocally identified.
Although the data discussed above do not match the
expectations of a common response to all stressors,
other results, including those from the most comprehen-
sive field-based study on coral thermal tolerance con-
ducted to date (Barshis et al. 2013), are consistent with a
generic foldase response. The A. millepora HSP90B1/
gp96 variant, which is up-regulated under acute CO2
stress (Table 5), has also been shown to be up-regulated
in an acute thermal stress experiment on larvae of
A. millepora (Rodriguez-Lanetty et al. 2009). A clear
gp96 orthologue (CAC38753.1) was likewise up-regulated
in the octocoral Dendronephthya klunzingeri subjected to
heat stress (Wiens et al. 2000). Analyses of field popula-
tions of Acropora hyacinthus revealed that orthologues of
the A. millepora HSF (contig145533 and contig209263)
and one of the HSP70 types (contig187488 in A. hyacin-
thus, corresponding to Cluster 007731 and Cluster
005163 in A. millepora) were consistently more highly
up-regulated in more thermotolerant than in less ther-
motolerant colonies (Barshis et al. 2013). As it is also a
component of the common CO2 stress response of
A. millepora, this HSP70 variant, rather than others, is a
good candidate general stress response marker for cor-
als. The second HSP70 component of the common CO2
stress response in A. millepora (Cluster005613) was also
up-regulated in heat-stressed Acropora palmata larvae
(Contig_13281 and Contig_23857 in the microarray
analysis of Polato et al. 2013), which is again consistent
with a generic foldase stress response.
Taken together, these results imply that, despite ini-
tial metabolic disruption, A. millepora juveniles have
remarkable capacity for acclimation to elevated pCO2
with a short response time. Although the initial
response to elevated pCO2 involves suppression of
metabolism and disrupted expression of genes
involved in skeleton deposition, after 9-d exposure,
© 2014 John Wiley & Sons Ltd
RAPID ACCLIMATION OF JUVENILE CORALS TO OA 449

these categories of genes had returned to control lev-
els. Acclimation to pCO2 involves up-regulated expres-
sion of a suite of anti-apoptotic Bcl-2 and chaperone
genes. In this context, the inducible HSP70 variant
(Cluster187488) may be particularly significant, because
it is not only a core component of the CO2 stress
response repertoire, but also is implicated in acclima-
tion to thermal stress in field populations of A. hyacin-
thus (Palumbi et al. 2014).
It remains uncertain whether coral populations can
adapt or shift in distribution on timescales that will
allow them to survive projected rates of climate change
(Hoegh-Guldberg et al. 2007; Pandolfi et al. 2011; Bars-
his et al. 2013; Palumbi et al. 2014). The short timescale
for acclimation to pCO2 observed in this study is there-
fore particularly significant. Clearly, field populations
can also rapidly acclimate to thermal stress (Palumbi
et al. 2014), suggesting that climate change impacts on
coral reefs may not be as fast as projected by some
authors (Hoegh-Guldberg et al. 2007; Dove et al. 2013).
These findings provide a glimmer of hope given the
ecological significance of species of the genus Acropora
throughout the Indo-Pacific. However, a number of
caveats apply to such extrapolations (Moya et al. 2012).
First, although these experiments imply that rapid accli-
mation to single stressors may occur, the combination
of changes in several stressors, such as OA and warm-
ing, could lead to larger impacts and less potential for
acclimation. Second, a healthy coral colony comprises
not only the animal host, but also one or more Symbi-
odinium strains and a complex bacterial community,
any one of which may be the weakest link in determin-
ing the survival of the colony. Third, Doropoulos et al.
(2012) have shown that crustose coralline algae, which
harbour bacterial epiphytes providing cues for coral
settlement, are very sensitive to OA. A decline in their
abundance, along with the decline in abundance of
their associated bacteria, could greatly reduce coral set-
tlement. Finally, coral communities will face not only
gradual changes in environmental parameters, but also
extreme thermal anomalies (Depczynski et al. 2013). It
is unclear whether acclimation alone can enable coral
populations to survive more frequent extreme events.
We are in the midst of an ongoing experiment on a
complex system that exhibits some built-in resilience
and adaptability. Whether this will be adequate for the
survival of coral reefs or whether they will disappear
within this century, as some have predicted, remains to
be seen.
Acknowledgments
This research was supported by the Australian Research Council
through Discovery Grant DP1095343 to D.J.M., E.E.B. and S.F.,
and via the Centre of Excellence for Coral Reef Studies, and by a
Marie Curie International Outgoing Fellowship (grant agree-
ment # PIOF-GA-2008-235142 project title AMICAL) to A.M.
This work is a contribution to the ‘European Project on Ocean
Acidification’ (EPOCA), which received funding from the
European Community’s Seventh Framework Program (FP7/
2007–2013) under grant agreement # 211384. The authors thank
Andrew Baird and William Leggat for the use of aquaculture
facilities, Gergely Torda, Chia-Miin Chua and Peter Cross for
their help with field trips and experiments, and Philip Munday
for developing the CO2-mixing system. The authors are also
grateful to Andrew Negri, Eneour Puill-Stephan, the SEASIMS
aquaculture facilities at AIMS for providing larvae for conduct-
ing pilot experiments, and Kazuhiro Sakamaki (Kyoto Univer-
sity) for commenting on the manuscript.
References
Albright R (2011) Reviewing the effects of ocean acidification
on sexual reproduction and early life history stages of reef-
building corals. Journal of Marine Biology, 2011, 1–14.Albright R, Langdon C (2011) Ocean acidification impacts mul-
tiple early life history processes of the Caribbean coral Porites
astreoides. Global Change Biology, 17, 2478–2487.Albright R, Mason B, Miller M, Langdon C (2010) Ocean acidi-
fication compromises recruitment success of the threatened
Caribbean coral Acropora palmata. Proceedings of the National
Academy of Sciences, USA, 107, 20400–20404.Alieva NO, Konzen KA, Field SF et al. (2008) Diversity and
evolution of coral fluorescent proteins. PLoS One, 3, e2680.
Ashburner M, Ball CA, Blake JA et al. (2000) Gene Ontology:
tool for the unification of biology. Nature Genetics, 25, 25–29.Barshis DJ, Ladner JT, Oliver TA, Seneca FO, Traylor-Knowles
N, Palumbi SR (2013) Genomic basis for coral resilience to
climate change. Proceedings of the National Academy of Sciences,
110, 1387–1392.Bosch TC, Augustin R, Anton-Erxleben F et al. (2009) Uncover-
ing the evolutionary history of innate immunity: the simple
metazoan Hydra uses epithelial cells for host defence. Devel-
opmental and Comparative Immunology, 33, 559–569.Bou-Abdallah F, Chasteen ND, Lesser MP (2006) Quenching of
superoxide radicals by green fluorescent protein. Biochimica
et Biophysica Acta (BBA)-General Subjects, 1760, 1690–1695.Brown T, Bourne D, Rodriguez-Lanetty M (2013) Transcrip-
tional activation of C3 and HSP70 as part of the immune
response of Acropora millepora to bacterial challenges. PLoS
One, 8, e67246.
Chomczynski P, Sacchi N (1987) Single-step method of RNA
isolation by acid guanidinium thiocyanate–phenol–chloro-form extraction. Analytical Biochemistry, 162, 156–159.
Chua CM, Leggat W, Moya A, Baird AH (2013a) Temperature
affects the early life history stages of corals more than near
future ocean acidification. Marine Ecology Progress Series, 475,
85–92.Chua CM, Leggat W, Moya A, Baird AH (2013b) Near-future
reductions in pH will have no consistent ecological effects
on the early life-history stages of reef corals. Marine Ecology
Progress Series, 486, 143–151.Collins M, Knutti R, Arblaster J et al. (2013) Long-term climate
change: projections, commitments and irreversibility. In: Cli-
mate Change 2013: The Physical Science Basis. Contribution of
© 2014 John Wiley & Sons Ltd
450 A. MOYA ET AL.

Working Group I to the Fifth Assessment Report of the Intergovern-
mental Panel on Climate Change (eds Stocker TF, Qin D, Plattner
G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V,
Midgley PM), pp. 1029–1136. Cambridge University Press,
Cambridge.
Cumbo VR, Fan TY, Edmunds PJ (2013) Effects of exposure
duration on the response of Pocillopora damicornis larvae to
elevated temperature and high pCO2. Journal of Experimental
Marine Biology and Ecology, 439, 100–107.Depczynski M, Gilmour JP, Ridgway T et al. (2013) Bleaching,
coral mortality and subsequent survivorship on a West Aus-
tralian fringing reef. Coral Reefs, 32, 233–238.DeSalvo MK, Voolstra CR, Sunagawa S et al. (2008) Differential
gene expression during thermal stress and bleaching in the
Caribbean coral Montastraea faveolata. Molecular Ecology, 17,
3952–3971.Doney SC, Fabry VJ, Feely RA, Kleypas JA (2009) Ocean acidi-
fication: the other CO2 problem. Annual Review of Marine Sci-
ence, 1, 169–192.Doropoulos C, Ward S, Diaz-Pulido G, Hoegh-Guldberg O,
Mumby PJ (2012) Ocean acidification reduces coral recruit-
ment by disrupting intimate larval-algal settlement interac-
tions. Ecology Letters, 15, 338–346.Dove S, Ortiz JC, Enriquez S et al. (2006) Response of holosym-
biont pigments from the scleractinian coral Montipora monas-
teriata to short-term heat stress. Limnology and Oceanography,
51, 1149–1158.Dove SG, Kline DI, Pantos O, Angly FE, Tyson GW, Hoegh-
Guldberg O (2013) Future reef decalcification under a busi-
ness-as-usual CO2 emission scenario. Proceedings of the
National Academy of Sciences, USA, 110, 15342–15347.Foret S, Seneca F, De Jong D et al. (2011) Phylogenomics
reveals an anomalous distribution of USP genes in metazo-
ans. Molecular Biology and Evolution, 28, 153–161.Gattuso J-P, Allemand D, Frankignoulle M (1999) Photosynthe-
sis and calcification at cellular, organismal and community
levels in coral reefs: a review on interactions and control by
carbonate chemistry. American Zoology, 39, 160–183.Grasso LC, Maindonald J, Rudd S et al. (2008) Microarray
analysis identifies candidate genes for key roles in coral
development. BMC Genomics, 9, 540.
Hardwick JM, Soane L (2013) Multiple functions of BCL-2 family
proteins. Cold Spring Harbor Perspectives in Biology, 5, 1–24.Hoegh-Guldberg O, Mumby PJ, Hooten AJ et al. (2007) Coral
reefs under rapid climate change and ocean acidification. Sci-
ence, 318, 1737–1742.Kaniewska P, Campbell PR, Kline DI et al. (2012) Major cellular
and physiological impacts of ocean acidification on a reef
building coral. PLoS One, 7, e34659.
Kenkel CD, Goodbody-Gringley G, Caillaud D, Davies SW,
Bartels E, Matz MV (2013) Evidence for a host role in
thermotolerance divergence between populations of the mus-
tard hill coral (Porites astreoides) from different reef environ-
ments. Molecular Ecology, 22, 4335–4348.Kvint K, Nachin L, Diez A, Nystr€om T (2003) The bacterial
universal stress protein: function and regulation. Current
Opinion in Microbiology, 6, 140–145.Kvitt H, Rosenfeld H, Zandbank K, Tchernov D (2011) Regula-
tion of apoptotic pathways by Stylophora pistillata (Anthozoa,
Pocilloporidae) to survive thermal stress and bleaching. PLoS
One, 6, e28665.
Langmead B, Trapnell C, Pop M, Salzberg SL (2009) Ultrafast
and memory-efficient alignment of short DNA sequences to
the human genome. Genome Biology, 10, R25.
Leggat W, Seneca F, Wasmund K, Ukani L, Yellowlees D, Ains-
worth TD (2011) Differential responses of the coral host and
their algal symbiont to thermal stress. PLoS One, 6, e26687.
Lewis E, Wallace DWR (1998) Program Developed for CO2 Sys-
tem Calculations. ORNL/CDIAC-105. Carbon Dioxide Infor-
mation Analysis Center, Oak Ridge National Laboratory U.S.
Department of Energy, Oak Ridge, Tennessee.
Morris JP, Thatje S, Hauton C (2013) The use of stress-70 pro-
teins in physiology: a re-appraisal. Molecular Ecology, 22,
1494–1502.Moya A, Huisman L, Ball EE et al. (2012) Whole transcriptome
analysis of the coral Acropora millepora reveals complex
responses to CO2-driven acidification during the initiation of
calcification. Molecular Ecology, 21, 2440–2454.Munday PL, Dixson DL, Donelson JM et al. (2009) Ocean acidi-
fication impairs olfactory discrimination and homing ability
of a marine fish. Proceedings of the National Academy of Sci-
ences, 106, 1848–1852.Nakamura M, Morita M, Kurihara H, Mitarai S (2012) Expres-
sion of HSP70, HSP90 and HSF1 in the reef coral Acropora
digitifera under prospective acidified conditions over the next
several decades. Biology Open, 1, 75–81.Nystr€om T, Neidhardt FC (1992) Cloning, mapping and
nucleotide sequencing of a gene encoding a universal
stress protein in Eschericha coli. Molecular Microbiology, 6,
3187–3198.Ogawa D, Bobeszko T, Ainsworth T, Leggat W (2013) The
combined effects of temperature and CO2 lead to altered
gene expression in Acropora aspera. Coral Reefs, 32, 895–907.Orr JC (2011) Recent and future changes in ocean carbonate
chemistry. In: Ocean Acidification (eds Gattuso J-P, Hansson
L), pp. 41–66. Oxford University Press, Oxford.
Oswald F, Schmitt F, Leutenegger A et al. (2007) Contributions
of host and symbiont pigments to the coloration of reef cor-
als. FEBS Journal, 274, 1102–1122.Palmer CV, Modi CK, Mydlarz LD (2009) Coral fluorescent
proteins as antioxidants. PLoS One, 4, e7298.
Palumbi SR, Barshis DJ, Traylor-Knowles N, Bay RA (2014)
Mechanisms of reef coral resistance to future climate change.
Science, 344, 895–898.Pandolfi JM, Jackson JBC, Baron N et al. (2005) Are US Coral
reefs on the slippery slope to slime? Science, 307, 1725–1726.Pandolfi JM, Connolly SR, Marshall DJ, Cohen AL (2011) Pro-
jecting coral reef futures under global warming and ocean
acidification. Science, 333, 418–422.Polato NR, Altman NS, Baums IB (2013) Variation in the tran-
scriptional response of threatened coral larvae to elevated
temperatures. Molecular Ecology, 22, 1366–1382.Putnam HM, Mayfield AB, Fan TY, Chen CS, Gates RD (2013)
The physiological and molecular responses of larvae from
the reef-building coral Pocillopora damicornis exposed to near-
future increases in temperature and pCO2. Marine Biology,
160, 2157–2173.Richter K, Haslbeck M, Buchner J (2010) The heat shock
response: life on the verge of death.Molecular Cell, 40, 253–266.Robinson MD, McCarthy DJ, Smyth GK (2010) edgeR: a Bio-
conductor package for differential expression analysis of dig-
ital gene expression data. Bioinformatics, 26, 139–140.
© 2014 John Wiley & Sons Ltd
RAPID ACCLIMATION OF JUVENILE CORALS TO OA 451

Rodriguez-Lanetty M, Harii S, Hoegh-Guldberg O (2009) Early
molecular responses of coral larvae to hyperthermal stress.
Molecular Ecology, 18, 5101–5114.Roemmich D, Gould WJ, Gilson J (2012) 135 years of global
ocean warming between the Challenger expedition and the
Argo Programme. Nature Climate Change, 2, 425–428.Sakamaki K, Shimizu K, Iwata H et al. (2014) The apoptotic ini-
tiator caspase-8: its functional ubiquity and genetic diversity
during animal evolution. Molecular Biology and Evolution, 31,
3282–3301.Sambrook J, Russell DW (2001) Molecular Cloning: A Laboratory
Manual. Cold Spring Harbor Laboratory, Cold Spring Har-
bor, New York.
Shinzato C, Shoguchi E, Kawashima T et al. (2011) Using the
Acropora digitifera genome to understand coral responses to
environmental change. Nature, 476, 320–323.Shinzato C, Hamada M, Shoguchi E, Kawashima T, Satoh N
(2012a) The repertoire of chemical defense genes in the
coral Acropora digitifera genome. Zoological Science, 29, 510–517.
Shinzato C, Shoguchi E, Tanaka M, Satoh N (2012b) Fluores-
cent protein candidate genes in the coral Acropora digitifera
genome. Zoological Science, 29, 260–264.Smith-Keune C, Dove S (2008) Gene expression of a green
fluorescent protein homolog as a host-specific biomarker of
heat stress within a reef-building coral. Marine Biotechnol-
ogy, 10, 166–180.Tchernov D, Kvitt H, Haramaty L et al. (2011) Apoptosis and
the selective survival of host animals following thermal
bleaching in zooxanthellate corals. Proceedings of the National
Academy of Sciences, 108, 9905–9909.Wiens M, Ammar MS, Nawar AH et al. (2000) Induction of
heat-shock (stress) protein gene expression by selected nat-
ural and anthropogenic disturbances in the octocoral Den-
dronephthya klunzingeri. Journal of Experimental Marine
Biology and Ecology, 245, 265–276.Young M, Wakefield M, Smyth G, Oshlack A (2010) Gene
ontology analysis for RNA-seq: accounting for selection bias.
Genome Biology, 11, R14.
A.M., L.H., S.F., E.E.B. and D.J.M. analysed the data.
A.M. and L.H. performed the experiments. A.M., L.H.,
S.F., J.P.G. and D.J.M. conceived and designed the
experiments. A.M., S.F., J.P.G., D.C.H., E.E.B. and
D.J.M. contributed to reagents, materials and analysis
tools, A.M., L.H., S.F., J.P.G., D.C.H., E.E.B. and D.J.M.
wrote the study and designed figures.
Data accessibility
The RNA-seq reads used in this study have been
submitted to the NCBI Gene Expression Omnibus
(GEO) database under Accession no. GSE61114.
Sequences of all genes mentioned in this study can
be found in the transcriptome of Acropora millepora
(Moya et al. 2012).
Supporting information
Additional supporting information may be found in the online ver-
sion of this article.
Fig. S1. Level of agreement between biological replicates.
Table S1. Over-representation of Gene Ontology (GO) terms in
the set of differentially expressed genes responding to acute (3-
d) and prolonged (9-d) exposure to elevated levels of CO2.
Table S2. The response of oxidative stress proteins to acute (3-
d) and prolonged (9-d) exposure to elevated levels of CO2.
Table S3. The response of fluorescent proteins (FPs) to acute
(3-d) and prolonged (9-d) exposure to elevated levels of CO2.
Table S4. The response of universal stress proteins (USP) to
acute (3-d) and prolonged (9-d) exposure to elevated levels of
CO2.
Table S5. The response of heat shock proteins (HSP) and heat
shock factors (HSF) to acute (3-d) and prolonged (9-d) expo-
sure to elevated levels of CO2.
© 2014 John Wiley & Sons Ltd
452 A. MOYA ET AL.