reference copy - wetland and aquatic research center · pdf filereference copy do not remove...
TRANSCRIPT

REFERENCE COPY Do Not Remove from the Library
r - . db I m,
Biological Report 82 (1 1.55) Nationa l Wetlands Research Centel- TR EL-82-4 June 1986
700 Cajun Dome Bou!e*~:crd
Species Profiles: Life Histories and Environmental Requirements of Coastal Fishes and Invertebrates (Gulf of Mexico)
BLUE CRAB
Fish and Wildlife Service Coastal Ecology Group
Waterways Ex~er iment Station
U.S. Department of the Interior U.S. Army Corps of Engineers

B i o l o g i c a l Report 82(1155) TR EL-82-4 June 1986
Species P r o f i 1 es: L i f e Hi s t o r i e s and Environmental Requirements o f Coastal Fishes and Inve r teb ra tes (Gu l f o f Mexico)
BLllE CRAB
H a r r i e t M. Perry and Thomas D. McI lwain Gu l f Coast Research Laboratory
Ocean Springs, MS 39564
P r o j e c t Manager C a r r o l l Cordes P r o j e c t O f f i c e r
David Moran Nat ional Coastal Ecosystems Team U.S. F i s h and W i l d l i f e Serv ice
1010 Gause Boulevard S l i d e l l , LA 70458
Performed f o r Coastal Ecology Group
Waterways Experiment S t a t i o n U. S. Army Corps o f Engineers
Vicksburg, MS 39180
and
Nat iona l Coastal Ecosystems Team Research and Development F i s h and W i l d l i f e Serv ice
U. S. Department o f I n t e r i o r Washington, DC 20240

DISCLAIMER
The mention o f t r ade names does n o t c o n s t i t u t e endorsement o r recommendation o f use by t h e Federal Government.
This s e r i e s should be referenced as fo l l ows :
U. S. F i s h and W i l d l i f e Service. 1983-19-. Species p r o f i l e s : 1 i f e h i s t o r i e s and environmental requ i rements o f coas ta l f i s h e s and inver tebra tes . U. S. F i s h Wi ld l . Serv. B i o l . Rep. 82(11). U.S. Army Corps o f Engineers, TR EL-82-4.
Th is p r o f i l e should be c i t e d as fo l lows:
Perry, H.M., and T.D. McIlwain. 1986. Species p r o f i l e s : l i f e h i s t o r i e s and environmental requirements o f coas ta l f i s h e s and i nve r teb ra tes (Gu l f of Mexico)--blue crab. U.S. F i s h W i l d l . Serv. B i o l . Rep. 82(11.55). U.S. Army Corps o f Engineers, TR EL-82-4. 21pp.

PREFACE
Th is species p r o f i l e i s one o f a se r i es on coasta l aquat ic organisms, p r i n c i p a l l y f i s h , o f spo r t , commercial , o r eco log i ca l importance. The p r o f i 1 es a re designed t o p rov ide coasta l managers, engineers, and b i o l o g i s t s w i t h a b r i e f comprehensive sketch o f t h e b i o l o g i c a l c h a r a c t e r i s t i c s and environmental requirements o f t h e species and t o descr ibe how popu la t ions o f t he species may be expected t o r e a c t t o environmental changes caused by coasta l development. Each p r o f i l e has sec t ions on taxonomy, l i f e h i s t o r y , eco log i ca l r o l e , environmental requirements, and economic importance, i f appl i cab le . A t h r e e - r i n g b inde r i s used f o r t h i s se r i es so t h a t new p r o f i l e s can be added as they a re prepared. Th is p r o j e c t i s j o i n t l y planned and f inanced by t h e U.S. Army Corps o f Engineers and t h e U.S. F i sh and W i l d l i f e Service.
Suggestions o r quest ions regard ing t h i s r e p o r t should be d i r e c t e d t o one of t h e f o l l owing addresses.
I n fo rma t ion Trans fer S p e c i a l i s t Nat iona l Coastal Ecosystems Team U.S. F i s h and W i l d l i f e Serv ice NASA-Sl i d e l 1 Computer Complex 1010 Gause Boulevard S l i d e l l , LA 70458
U. S. Army Engineer Waterways Experiment S t a t i o n A t ten t i on : WESER-C Post O f f i c e Box 631 Vicksburg, MS 39180

CONVERSION TABLE
Metric t o U.S. Custmary
Mu1 t i p l y !.Y To Obtain
mill ine te r s (mn) centimeters (on) meters (m) ki 1 ometers ( km)
2 square meters (m ) 10.76 square ki 1 ometers ( km2) 0.3861 hectares (ha) 2.471
l i t e r s ( 1 ) cubic ineters (m3) cubic meters
inches inches f e e t miles
square f ee t square inil es acres
gal 1 ons cubic f e e t acre-feet
mill igrams (mg) 0.00003527 ounces grams ( g ) 0.03527 ounces kil ograms (kg) 2.205 pounds metric tons ( t ) 2205.0 pounds metric tons 1.102 short tons ki 1 ocal or ies ( kcal ) 3.968 Br i t i sh thennal uni ts
Cel s ius degrees 1.8(OC) + 32 Fahrenheit degrees
U.S. Customary to Metric
inches 25.40 inches 2.54 f e e t ( f t ) 0.3048 fa thorns 1.829 miles (mi) 1.609 nautical miles ( m i ) 1.852
square f e e t ( f t 2 ) acres 2 square miles (mi
gal 1 ons ( gal ) 3.785 cubic f e e t ( f t 3 ) 0.02831 acre-feet 1233.0
ounces (oz ) 28.35 pounds ( l b ) 0.4536 shor t tons ( t o n ) 0.9072 Brit ish thermal uni ts ( B t u ) 0.2520
mil 1 imeters cent imeters meters meters kil ometers ki 1 oqeters
square meters hectares square ki 1 ometers
1 i t e r s cubic meters cubic meters
grains ki 1 ograms metric tons ki 1 ocal o r i e s
Fahrenheit degrees 0.5556("F - 32) Celsius degrees

CONTENTS
Page
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . PREFACE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . CONVERSIONTABLE
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . ACKNOWLEDGMENTS
NOMENCLATURE/TAXONOMY/RANGE . . . . . . . . . . . . . . . . . . . . . . . MORPHOLOGY/IDENTIFICATION AIDS . . . . . . . . . . . . . . . . . . . . . REASON FOR INCLUSION I N THE SERIES . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . LIFE HISTORY . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Spawning
Larvae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Juven i les . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Adul ts . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Migra t ions GROWTH . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . ECOLOGICAL ROLE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . THE FISHERY
Commercial Harvest . . . . . . . . . . . . . . . . . . . . . . . . . . Recreat ional Harvest . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . I n c i d e n t a l Harvest Factors A f f e c t i n g Commercial Landings . . . . . . . . . . . . . . . .
ENVIRONMENTAL REQUIREMENTS . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . LITERATURECITED
iii i v v i

ACKNOWLEDGMENTS
The authors are g ra te fu l t o Gerald Adkins, Louisiana Department o f Wild1 i f e and F isher ies and t o Paul Hammerschmidt, Texas Parks and W i l d l i f e Department, f o r t h e i r reviews o f the manuscript. A special thanks t o Cindy Dickens f o r typing the manuscript and t o Linda La i rd f o r preparing the i l l u s t r a t i o n s .

Figure 1. Blue crab ( f rom Rathbun 1930).
NOMENCLATURE/TAXONOMY/RANGE MORPHOLOGY/IDENTIFICATION AIDS
S c i e n t i f i c name . . . . . Ca l l i nec tes sapidus Rathbun 1896
Pre fer red common name . . . Blue crab (Figure 1)
Other common names . . Common e d i b l e crab, e d i b l e b lue crab
Class . . . . . . . . . . . Crustacea Order . . . . . . . . . . . . Decapoda I n f r a o r d e r . . . . . . . . Brachyura Family . . . . . . . . . . Portunidae
Geographical range: D i s t r i b u t e d throughout t h e coasta l waters o f t h e Gu l f o f Mexico (Figure 2). W i l l iams (1974) de f ined t h e range as: Occasional l y Nova Scot ia , Maine, and nor thern Massachusetts t o nor thern Argentina, i n c l u d i n g Bermuda and t h e A n t i l l e s ; Oresund, Denmark; t h e Netherlands and adjacent North Sea; southwest France (found twice) ; Go l fo d i Genova; nor thern A d r i a t i c ; Aegean, western Black, and eas tern Mediterranean seas.
Wi l l iams (1974) prov ided a de ta i l e d morphological desc r i p t i on . Fronta l margin o f t he carapace w i t h f o u r i nne r o r b i t a l tee th . Antero- l a t e r a l margin o f carapace w i t h 9 spines o r t ee th , t he poster ior-most s t r o n g l y developed. Carapace about 2.5 t imes as wide as long, moderately convex and nea r l y smooth. Granula- t i o n s on t h e i n n e r b ranch ia l and card iac reg ions of t h e carapace.
Sex determined e x t e r n a l l y by t h e shape o f t h e abdomen. Abdomen o f t h e male T-shaped. Male gonopods (copu la tory organs) reach near ly t o o r extend beyond t h e t i p o f t h e abdomen. Immature females w i t h t r i a n g u l a r abdomen; abdomen o f mature females semi -c i rcu la r . M a t u r i t y i n males cannot be determined e x t e r n a l l y .
Color va r i ab le , w i t h shades o f g ray ish , b l u i s h , o r brownish green occurr ing. The propodi o f chelae o f

_ _ _ _ - - - - ----- dm --
GULF OF MEXICO
0 50 100 150 200
KILOMETERS
F igu re 2. B lue crabs a r e d i s t r i b u t e d throughout t h e coas ta l waters o f t h e G u l f o f Mexico and a r e most abun- dan t i n waters up t o 35 m. Greatest r epo r t ed commercial l and ings occur n o r t h o f 2 8 O N l a t i t u d e .

males b lue on t h e i n n e r and ou te r surfaces t i p p e d w i t h red. The f i n g e r s of chelae o f mature females orange t i pped w i t h purp le .
REASON FOR INCLUSION I N SERIES
The b lue crab f i s h e r i e s have become inc reas ing l y impor tan t i n t h e G u l f States. Reported land ings f o r t h e G u l f o f Mexico i n 1984 exceeded 5 1 m i l l i o n 1b w i t h an ex-vessel value approaching $15 m i 11 ion: increases o f 4 l% and 29%, respec t i ve l y , compared t o 1983. I n a d d i t i o n t o t he commercial hard crab f i s h e r y , t h e r e e x i s t a sub- s t a n t i a l r e c r e a t i o n a l f i s h e r y and an expandi ng commerci a1 and r e c r e a t i o n a l f i s h e r y f o r s o f t - s h e l l e d crabs.
Blue crabs occupy a v a r i e t y o f hab i ta t s . The upper, middle, and lower es tuary and adjacent marine area together c o n s t i t u t e the b lue crab h a b i t a t . Ea r l y l a r v a l stages are found i n t h e lower estuary and adja- cen t marine waters; s a l i n i t i e s i n excess o f 20.0 p p t a re requ i red f o r successful development. La te r stage zoeae e x i s t main ly i n t he open g u l f where t h e i r a rea l and v e r t i c a l d i s t r i - b u t i o n determine t h e i r eventual t r a n s p o r t shoreward. Blue crabs en te r t he estuary as megalopae, adopt ing a more ben th i c ex istence. The m o l t t o t he f i r s t c rab stage takes p lace w i t h i n t h e estuary. Factors a f f e c t i n g d i s t r i b u t i o n and s u r v i v a l i nc lude sub- stratum, food a v a i l a b i l i t y , a v a i l a b l e she1 t e r , water temperature, and sa l i n i t y . Because b lue crabs occupy a v a r i e t y o f h a b i t a t s and are an i n t e g r a l p a r t of t h e coasta l eco- system, maintenance o f t h e e n t i r e es tua r ine system i n a c o n d i t i o n s u i t a b l e f o r cont inued product ion i s o f pr ime importance.
lUn less otherwise noted, a1 1 s t a t i s t i - c a l da ta presented i n e i t h e r t he t e x t o r t ab les a re from F ishery S t a t i s t i c s of t h e Un i ted Sta tes and Current - - - Fishery ~ t a t i sti-arious years), both pub l ished by t h e Nat iona l Marine F i she r ies Serv ice.
LIFE HISTORY2
Spawning
Spawning o f b lue crabs i n nor thern g u l f waters i s p r o t r a c t e d w i t h egg- bear ing females occu r r i ng i n coasta l and es tua r ine waters i n t he spr ing , summer, and f a l l (Gunter 1950, Daugherty 1952, More 1969, Adkins 1972, Per ry 1975). Add i t i ona l l y , Adkins (1972) found evidence o f w i n t e r spawning i n o f f s h o r e Louis iana waters based on commercial catches o f "ber ry" crabs i n December, January, and February, and Daugherty (1952) noted t h a t crabs i n southern Texas may spawn year round i n years w i t h m i l d w in ters .
For most marine animals mat ing and spawni ng are synonymous ; however, i n t h e case o f t he b lue crab, t h e two events occur a t d i f f e r e n t t imes. The female mates i n t h e s o f t - s h e l l s t a t e f o l 1 owing t h e puber ta l o r te rmina l mo l t ( i n t h e female b lue c rab the cyc le of growth and m o l t i n g te rm i - nates w i t h a f i n a l ecdysis) . A f t e r inseminat ion, t he male cont inues t o c a r r y t h e female u n t i l her s h e l l has hardened. Spawning usual l y occurs w i t h i n 2 months o f mat ing i n t h e s p r i n g and summer. Females t h a t mate i n t he f a l l u s u a l l y delay spawning u n t i l t h e fo l l ow ing spr ing . Sperm t rans fe r red t o t h e female remain v i a b l e f o r a year o r more and are used f o r repeated spawnings.
The f e r t i l i z e d eggs are extruded and at tached t o f i n e setae on the endopo- d i t e s of t h e pleopods, forming an egg mass known as a "sponge," "berry," o r "pom-pom." As many as two m i l l i o n eggs may be present i n a s i n g l e sponge. The sponge i s i n i t i a l l y b r i g h t orange, becomi ng progres- s i v e l y darker as t h e l a rvae develop and absorb the yo l k . Jus t p r i o r t o hatching, t h e sponge i s b lack. The eggs hatch i n about 2 weeks. C h u r c h i l l (1921) and Van Engel (1958) have prov ided d e t a i l e d data on repro- duc t i on and spawning o f b lue crabs.
Z In fo rma t ion i n t h e f o l low ing sec t ions was taken from Perry e t a l . (1984).

Larvae
Costlow and Bookhout (1959) repo r ted seven zoeal stages and one megalopal stage (F igure 3). An e i g h t h zoeal stage was sometimes observed though s u r v i v a l t o t h e megalopal stage was rare . Development through the seven zoeal stages requ i red from 3 1 t o 49 days; t h e megalopal stage p e r s i s t e d from 6 t o 20 days. I n s a l i n i t i e s below 20.1 p p t t h e l a rvae r a r e l y surv ived t h e f i r s t mol t .
The l a r v a l l i f e h i s t o r y o f t he b l u e crab i n t h e Gu l f o f Mexico i s p o o r l y understood. A1 though Daugherty (1952), Menzel (1964), and Adkins (1972) s p e c i f i c a l l y discussed t h e d i s t r i b u t i o n o f b l u e crab la rvae, t h e poss ib i 1 i t y o f co-occurrence o f t he l a rvae o f t he l e s s e r b lue crab (C. s i m i l i s ) must be considered. The temporal and s p a t i a l over lap i n spawn- i n g h a b i t s of t h e two species (Perry 1975), coupled w i t h t he d i f f i c u l t y i n us ing the e a r l y morphological descr ip - t i o n s o f the b l u e crab l a rvae from t h e A t l a n t i c (Costlow and Bookhout 1959) t o re1 i a b l y i d e n t i f y g u l f speci- mens, suggests t h a t pub l ished accounts o f the seasona l i ty o f b lue crab l a rvae are quest ionable. Recognizing the d i f f i c u l t y i n separa t ing the two species, King (1971), Per ry (1975), and Andryszak (1979) d i d n o t d i f f e r e n - t i a t e between the l a rvae o f t he b l u e crab and the l e s s e r b lue crab.
Per ry and Stuck (1982a) noted t h a t e a r l y stage C a l l i n e c t e s zoeae (I and 11) were present i n M i s s i s s i p p i coas ta l waters i n t h e spr ing , summer, and fa1 1. Adkins (1972) repo r ted b lue crab l a rvae were present year-round i n Louis iana, b u t d i d no t separate the zoeal and megalopal stages. The sampling programs o f Menzel (1964) and Andryszak (1979) were o f l i m i t e d dura t ion . Per ry and Stuck (1982a) and Andryszak (1979) found o n l y t h e e a r l y stage zoeae abundant nearshore.
C a l l i nectes mega1 opae have been repo r ted t o oc ru r throughout t h e year.
Perry (1975) found megalopae i n M i s s i s s i p p i Sound i n a l l months; peak abundance was i n t h e l a t e summer-early f a l l and i n February. I n Texas coasta l waters, C a l l i n e c t e s megalopae have been found i n a1 1 seasons (Daugherty 1952, More 1969, King 1971). King (1971) noted th ree waves o f megalopae i n Cedar Bayou, Texas: t h e f i r s t from January through March, t h e second i n May and June, and t h e t h i r d i n October.
A t t e m ~ t s t o s e ~ a r a t e t h e meaa lo~ae - . o f C. 'sapidus a"d C. s i m i l i s , us ing t h e characters d e v e i o ~ e d bv Bookhout and Costlow (1977), have been 1 a rge l y unsuccessful because o f apparent mor- phol og i ca l d i f f e rences i n 1 arvae from t h e G u l f o f Mexico and A t l a n t i c Ocean. Stuck e t a l . (1981) and Stuck and Per ry (1982) provided charac ters usefu l i n d i s t i n g u i s h i n g the megalopae and e a r l y crab stages o f t he two species. Subsequent ana l ys i s of a rch ived p lank ton samples from M i s s i s s i p p i and Louis iana coasta l waters has fu rn ished i n fo rma t ion on the seasona l i ty o f C. sa idus mega- lopae i n t h e norther; g u h u c k and Per ry 1981). C a l l i nectes sapidus megalopae were r a r e l y found i n samples before May. These data suggest t h a t t h e r e ~ o r t e d w i n t e r peaks o f c a l l i nectes 1 arvae i n t h e no r the rn g u l f a re probably r e f e r a b l e t o C. s i m i l i s .
R e ~ o r t s on the v e r t i c a l d i s t r i b u t i o n of c a l l i nectes mega1 opae appear con- f l i c t i n g . Wi l l iams (1971), King (1971), Perry (1975), and Smyth (1980) repo r ted megalopae t o be most abundant i n sur face waters. I n con t ras t , 96% o f t he Ca l l i nec tes megalopae c o l l e c t e d by Tagatz (1968a) and a l l o f those c o l l e c t e d by Sand i fe r (1973) were from bottom waters. Stuck and Perry (1981) found t h a t or tun id meaa lo~ae (C. sapidus, C. ' s im i 1 i s , a i d ' ~ o r t u i u s spp.) showed no a f f i n i t y f o r sur face o r bottom waters. They noted t h a t t he m a j o r i t y o f l a r g e catches o f C. sapidus megalopae were taken on r i s i r i g o r peak t i des , whereas the mega1 opae

Figure 3. Larval stages o f t h e b lue crab (Stuck and Perry 1982).
of C. s i m i l i s and Portunus spp. were commonly c o l l e c t e d on bo th r i s i n g and f a l l i n g t i des .
L i t t l e i s known about t he mechani sms o f l a r v a l t r a n s p o r t and d ispersa l o f b lue crab zoeae i n t he nor thern g u l f . Based on the data o f Menzel (1964), Andryszak (1979), and Perry and Stuck (1982a), i t appears t h a t development through the l a t e zoeal stages (111 through V I I ) takes p lace i n o f f sho re waters. A t t h i s t ime, t h e l a rvae are sub jec t t o cur ren ts and may be t rans- po r ted considerable d is tances. Recruitment o f l a r vae back i n t o coasta l waters occurs du r ing the mega1 opal stage. Oester l i ng and Evi nk (1977) proposed a mechanism f o r l a r v a l d ispersa l i n nor theastern g u l f waters i n which b lue crab l a rvae were t rans- po r ted d is tances o f 300 km o r more. I f such t r a n s p o r t mechanisms do e x i s t i n t he g u l f , l a r vae produced by spawn- i n g females i n one Sta te may i n f a c t be respons ib le f o r rec ru i tmen t i n a d j o i n i n g States.
Juveni 1 es
Recruitment o f b lue crabs t o g u l f es tua r ies occurs du r ing the mega1 opal stage (More 1969, King 1971, Perry 1975, Perry and Stuck 1982b). The r e l a t i o n s h i p between numbers o f mega1 opae r e c r u i t e d and subsequent abundance o f j u v e n i l e crabs i s no t we1 1 defined. Perry and Stuck (1982b) noted t h a t l a rge catches o f b lue crab megalopae i n August and September were usual l y f o l 1 owed by an increased catch o f j u v e n i l e crabs (10.0 t o 19.9 mm) i n October o r November i n M i s s i s s i p p i es tuar ies ; however, incons is tenc ies between rec ru i tmen t o f megalopae and subsequent occurrence and abundance o f j u v e n i l e s were noted i n t h e sp r i ng and summer i n t h e i r samples. King (1971) found comparable popu la t i on d e n s i t i e s o f j u v e n i l e s between 2 years though rec ru i tmen t was markedly d i f f e r e n t . I n t e r p r e t a t i o n o f h i s data i s somewhat complicated by t h e taxonomic problems associated w i t h t h e

separa t ion o f C. sapidus and C. s i m i 1 i s mega1 opaey
Juven i l e b l u e crabs show wide seasonal and area l d i s t r i b u t i o n i n g u l f es tuar ies . L i v i ngs ton e t a l . (1976) found maximum numbers o f b l u e crabs i n Apalachicola Bay, F l o r i d a , i n t he w i n t e r and summer, n o t i n g t h a t an almost "continuous succession" o f juven i l e crabs entered t h e sampl i ng area du r ing the year. Per ry (1975) and Per ry and Stuck (1982b) found f i r s t crab stages i n a l l seasons, i n d i c a t i n g con t i nua l rec ru i tmen t t o t he j u v e n i l e popu la t i on i n M iss i ss ipp i . I n Lake Pontchar t ra i n, Louisiana, Darnel 1 (1959) noted t h a t rec ru i tmen t o f j u v e n i l e crabs was h ighes t i n t h e l a t e sp r i ng -ea r l y summer and i n t he f a l l .
A1 though j uven i 1 e crabs occur over a broad range o f s a l i n i t i e s , they are most abundant i n low t o interme- d i a t e s a l i n i t i e s c h a r a c t e r i s t i c o f middle and upper es tua r ine waters. Using temperature-sal i n i t y matr ices, Swingle (1971), Pe r re t e t a l . (1971), Christmas and Langl ey (1973), and Per ry and Stuck (1982b) determined the d i s t r i b u t i o n o f b lue crabs ( p r i m a r i l y j uven i 1 es). P e r r e t e t a1 . (1971) and Swingle (1971) found maximum abundance i n sa l i n i ti es be1 ow 5.0 p p t (Table 1). I n con t ras t , Christmas and Langley (1973) and Per ry
and Stuck (1982b) found h ighest average catches assoc ia ted w i t h s a l i n i t i e s above 14.9 p p t i n M i s s i s s i p p i (Table 1). From 1 year o f bag seine data, Hammerschmidt (1982) found no d i r e c t r e l a t i o n s h i p between catches o f j uven i 1 e crabs and s a l i n i t y i n Texas. Although s a l i n i t y in f luences d i s t r i b u t i o n , f a c t o r s such as bottom t ype and food a v a i l a b i l i t y a l s o p l a y a r o l e i n determining d i s t r i b u t i o n a l pa t te rns o f j uven i 1 e b l ue crabs.
The importance o f bottom type i n t h e d i s t r i b u t i o n o f j u v e n i l e b lue crabs i s w e l l es tab l ished. More (1969), Hol land e t a l . (1971), Adkins (1972), Per ry (1975), L i v i n g s t o n e t a l . (1976), and Per ry and Stuck (1982b) a l l noted the assoc ia t i on o f j u v e n i l e b lue crabs w i t h s o f t , mud sediments. Evink (1976) c o l l e c t e d t h e g rea tes t number o f i n d i v i d u a l s and biomass from mud bottoms and noted t h a t b lue crab biomass appeared t o be determined by faunal food a v a i l - a b i 1 i t y .
Adu l ts
The ovar ian stages descr ibed by Hard (1942) were used by Per ry (1975) t o de f i ne the reproduct ive s t a t e o f b l u e crab populat ions i n M i s s i s s i p p i . Recently mated females (Stage I ) and crabs w i t h developing ovar ies (Stage
Table 1. D i s t r i b u t i o n o f the b lue crab by s a l i n i t y i n t e r v a l s o f 5 p p t showing number o f samples (below) and catch per sample (above).
Sa l inity(ppt) - .-. . - . . - - - - - - -. . - - - -- -- -- . - - 0 .0-4.9 5. J -9 .9 10.0-14.9 15.0-19.9 20.0-24.9 25.0-29.9 30+ T o t a l
Modi f i e d f r o s ~ :
Swing le 1971 6.0 4.7 2 .6 2.3 (11) (15) (14) (19)
P e r r e t e t a l . 1971 12 6 6 6 (197) ( 185) (263) (278)
Ch~* i s tmas and 1.2 2.7 3.8 3 . 2 Langley 1973 (134) (07) (110) (99)
Perry and Stuck 7.6 7.8 7 . 1 8. 3 1982b (561) (423) ( r e ? ) (520)

11) were observed i n t h e spr ing , summer, and f a l l . Females w i t h mature ovar ies (Stage 111) occurred through- ou t t he year. The appearance o f b e r r i e d females (Stage I V ) i n March and Apr i 1 i n d i c a t e d t h a t ove rw in te r i ng Stage I11 females spawned when water temperatures began t o r i s e i n t h e spr ing. Stage I V crabs were most abundant du r i ng the middle and l a t e summer, corresponding w i t h t h e i n f l u x o f " g u l f " crabs from o f f sho re waters. Stage V crabs ( repeat spawners) appeared du r ing the summer, p r o v i d i n g evidence t h a t some females do spawn tw ice .
There i s a d i f f e r e n t i a l d i s t r i - b u t i o n o f male and female crabs i n r e l a t i o n t o s a l i n i t y ( C h u r c h i l l 1921, Gunter 1950, Darnel1 1959, Perry 1975). Adu l t males tend t o remain i n low s a l i n i t y waters w h i l e mature females p r e f e r t he h igher s a l i n i t i e s o f t h e lower es tuary and adjacent marine waters.
Churchi 11 (1921) noted t h a t t h e maximum age f o r b lue crabs i n Chesapeake Ray was about 3 years. Tagatz (1968a) repor ted t h a t t he maximum age o f b lue crabs i n the St. Johns River , F l o r i d a , was 4 years b u t noted t h a t few crabs su rv i ve pas t 2 years o f age.
Migra t ions
Movements o f b lue crabs w i t h i n es tua r ine systems a re r e l a t e d t o l i f e cyc le stages, season, and envi ron- mental cond i t i ons (Van Den Avyle and Fowler 1984).
Migra t ions o f females are u s u a l l y associated w i t h mat ing and gonadal matura t ion and spawning.
The m ig ra t i on pa t te rns o f b lue crabs i n the g u l f observed by More (1969) and Per ry (1975) were t y p i c a l o f t h e onshore/offshore movements charac- t e r i z e d i n previous s tud ies ( F i e d l e r 1930, Van Engel 1958, F i s c h l e r and Walburg 1962, Tagatz 1968a, Judy and
Dudley 1970). Oester l i n g and Evink (1977) and Stee le (1984) prov ided evidence o f an a1 ongshore movement o f females i n F l o r i d a coasta l waters. M ig ra to ry pa t te rns observed i n t h e i r s tud ies demonstrated movement o f females t o s i t e s n o r t h o f t h e i r mat ing estuary. Oester l i n g and Evink (1977) noted t h a t t he Apalachicola Bay reg ion appeared t o be a pr imary spawning ground f o r crabs a long t h e F l o r i d a pen insu lar g u l f coast. Steele (1984) repo r ted t h a t the concent ra t ion o f m ig ra to ry females i n t he Apalachicola Bay area was the r e s u l t o f a sa l i n i t y b a r r i e r c rea ted by ou t f l ow from the Apalachicola River . A hypothesis f o r r e d i s t r i bu- t i o n o f l a r vae t o southwestern F l o r i d a invo lved t r a n s p o r t o f zoeae i n sur face cu r ren ts associated w i t h Apalachicola R iver f l o w and the G u l f o f Mexico Loop Current.
GROWTH
Newcombe e t a l . (1949) est imated the number o f p o s t l a r v a l i n s t a r s f o r male and female b l u e crabs t o be 20 and 18, respec t i ve l y . I f the number o f mo l ts i s assumed t o be f i x e d i n b l u e crabs (Newcombe e t a l . 1949, Van Engel 1958), t he v a r i a b i l i t y i n t he average s i z e a t which m a t u r i t y i s a t t a i n e d i n t h e female, coupled w i t h t he observat ions t h a t unusual ly l a r g e b lue crabs a re found i n low s a l i n i t i e s , suggests t h a t envi ron- mental cond i t i ons i n f l uence the percentage increase i n s i z e pe r mol t . Blue crabs i n Chincoteague, Chesapeake, and Delaware Bays increase i n s i z e w i t h decreasing environmental sal i n i ty (Po r te r 1955, Cargo 1958). The data o f Newcombe (1945), Van Engel (1958), and Tagatz (1965, 1968a) a l so suggest a poss ib le negat ive c o r r e l a- t i o n o f s i z e w i t h t he s a l i n i t y o f t h e water i n which growth occurs. Van Engel (1958) be l i eved t h a t the osmoregulatory mechanism was involved; d i f f e rences i n the l e v e l s o f s a l t concent ra t ion between t h e crabs and t h e i r environment a f f e c t e d the uptake o f water , r e s u l t i n g i n increased

growth pe r mol t . Haefner and Shuster (1964), i n a study o f t he growth increments occu r r i ng d u r i n g the te rm ina l m o l t o f t h e female b lue crab under d i f f e r e n t sa l i n i t i e s , concluded t h a t " w i t h i n t h e parameters o f t he experiment, t h e s a l i n i t y v a r i a t i o n o f t h e environment i s n o t r e l a t e d t o percentage increase i n l e n g t h a t t he te rm ina l mol t. I' Tagatz (1968b) a1 so repor ted t h a t a decrease i n s a l i n i t y d i d n o t produce an increase i n s i z e and suggested t h a t some f a c t o r o the r than s a l i n i t y appeared t o account f o r l a r g e r crabs i n c e r t a i n waters.
Growth o f b lue crabs i s s t r o n g l y a f f e c t e d by temperature. One o f t h e more obvious e f f e c t s o f temperature on growth r a t e i s t h e l e n g t h o f t ime requ i red f o r crabs t o reach ma tu r i t y . Up t o 18 months i s necessary f o r matura t ion i n Chesapeake Bay (Van Engel 1958), w h i l e b lue crabs i n t he G u l f o f Mexico may reach m a t u r i t y w i t h i n a yea r (Per ry 1975, Tatum 1980).
I n t h e l abo ra to ry , L e f f l e r (1972) demonstrated t h a t t h e m o l t i n g r a t e (mol ts pe r u n i t o f t ime) increased r a p i d l y w i t h i nc reas ing temperature from 13.0 t o 27.0 OC. Th is increase cont inued a t a s lower r a t e between 27.0 and 34.0 O C and growth v i r t u a l l y ceased a t temperatures below 13.0 OC.
The growth pe r mo l t was s i g n i f i c a n t l y reduced above 20.0 OC. Thus, w h i l e t h e m o l t i n g r a t e increased w i t h temperature, t h e number o f mo l ts necessary t o a t t a i n a c e r t a i n s i z e a l s o increased. I f the maximum s i z e a b lue crab a t t a i n s i s assumed t o r e f l e c t t h e growth pe r m o l t r a t h e r than t h e number o f mol ts , envi ron- mental temperatures may, i n p a r t , be respons ib le f o r t h e v a r i a t i o n i n s i ze a t ma tu r i t y .
Per ry (1975) est imated growth by t r a c i n g modal progressions i n monthly width-frequency d i s t r i b u t i o n s f o r crabs i n M i s s i s s i p p i Sound. The est imated growth r a t e o f 24 t o 25 mm/ month i s somewhat h igher than r a t e s
found i n o the r g u l f es tuar ies . Adkins (1972) found growth i n Louis iana waters . t o be approximately 14 .mm/ month f o r young crabs, w i t h s l i g h t l y h igher r a t e s (15 t o 20 mm/month) as crabs exceeded 85.0 mm i n carapace w id th . Darnel 1 ' s (1959) growth e s t i - mate o f 16.7 mm/month f o r crabs i n Lake Pontchar t ra in fa1 1 s w i t h i n t he average repor ted by Adkins. More (1969) noted a growth r a t e o f 15.3 t o 18.5 mm/month i n Texas. P l o t t i n g t h e progression o f modal groups from February through August, Hammerschmidt (1982) repor ted h igher growth r a t e s f o r crabs i n Texas (21.4 and 25.2 mm/ month f o r seine and t r a w l samples, respec t i ve l y ) and a t t r i b u t e d these r a t e s t o t h e use o f seasonal r a t h e r than y e a r l y data. Tatum (1980) found seasonal changes i n t h e r a t e o f growth o f young b lue crabs i n Mobi le Bay, Alabama. He observed monthly r a t e s o f 19.0, 10.0, and 5.0 mm f o r crabs r e c r u i t e d i n Apr i 1, August, and December, respec t i ve l y .
ECOLOGICAL ROLE
Blue crabs feed on var ious crustaceans, molluscs, f i s h , d e t r i t u s , and on o the r b lue crabs. They are u s u a l l y charac ter ized as o p p o r t u n i s t i c ben th i c omnivores.
Young and a d u l t b lue crabs occur i n es tua r ine waters throughout t h e year and a re impor tan t p rey species f o r a v a r i e t y o f organisms. Many species o f b i r d s , i n c l ud i ng herons and d i v i n g ducks, feed on b lue crabs. Mammal i an predators i n c l ude humans and t h e raccoon. F i s h species feed ing on b l ue crabs i nc lude spot ted seat rou t , r e d drum, b lack drum, croaker, gars, sheepshead, and f reshwater and s a l t - water c a t f i s h .

THE FISHERY f 1 uctuated w ide l y a1 though repor ted landings from 1975 t o 1980 have no t
Commerci a1 Harvest
Annual commercial 1 andi ngs o f b lue crabs from t h e Gu l f o f Mexico have been repo r ted s ince 1880 (Table 2).
Va r ia t i ons i n t h e abundance o f crabs due t o environmental f a c t o r s and disease, use o f more e f f i c i e n t gear, increased f i s h i n g e f f o r t , and t h e economic c o n d i t i o n o f t h e market a re r e f l e c t e d i n h i s t o r i c a l catches o f b l u e crabs. The f i s h e r y i n M i s s i s s i p p i and Alabama has been r e l a t i v e l y s tab le ; each s t a t e repor ted from 1.1 m i l l i o n t o 2.7 m i l l i o n l b annual ly . Louis iana cont inues t o be the l a r g e s t producer i n t h e g u l f , supp ly ing raw product t o Texas, M i s s i s s i p p i , and Alabama processing p lan ts . Landings f o r Louis iana have
approached the 1973 landings o f 23 m i l l i o n l b . F l o r i d a g u l f coas t landings have remained re1 a t i v e l y s t a b l e a t 13 m i l l i o n 1b a f t e r d e c l i n i n g from 2 1 m i l l i o n 1b i n 1965 t o 9 m i l l i o n 1b i n 1968. Landings i n Texas approached 9 m i l l i o n l b i n 1980. The percentage c o n t r i - bu t i ons o f each S ta te t o t he t o t a l Gu l f o f Mexico b l u e crab landings from 1960-1980 a re shown i n Table 3; t he g u l f coas t o f F l o r i d a and Louis iana have con t r i bu ted the h ighest per- centage s ince 1960. The percentage c o n t r i b u t i o n s o f g u l f landings t o t o t a l U.S. landings (1960-80) a re shown i n Table 4.
Seasonal f l u c t u a t i o n s i n repo r ted commercial landings a re s imi 1 a r among a1 1 the Gu l f States (F igure 4). Com-
Table 2. H i s t o r i c a l commercial l and ing s t a t i s t i c s by S ta te f o r hard s h e l l b lue crab, 1880-1980, i n thousands o f pounds and thousands o f d o l l a r s (from Perry e t a l . 1984).
Florida West Coast Alabama Mississippi Louisiana Texas Total
Year Quantity Value Quantity Value Quantity Value Quantity Value Quantity Value Quantity Value
1880 - - - - - - 288 7 36 1 324 8 1887 (2) (2) (2) (2 ) 38 1 837 13 11 1 4 (2) (2) 1888 3 (1) 96 6 16 (1) 85 1 1 3 115 4 1,081 2 3 1889 - - 48 1 842 14 189 5 1,079 20 -
1890 - - - - 33 1 85 1 1 3 191 5 1,075 19 1891 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1892 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1895 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1897 6 (1) 24 1 132 3 1,459 13 138 4 759 2 1 1898 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1899 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1901 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1902 1 (1) 75 2 235 5 312 16 4 3 2 1,666 25 1904 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1905 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1908 2 (1) 246 6 380 10 244 8 199 5 1,071 29 1915 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2)
- (2)
1918 - 96 3 216 6 282 10 193 11 787 30 1919 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1920 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1921 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1922 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2)
- ,1923 - 84 3 4 35 11 312 8 109 9 940 31 1924 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1925 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1926 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1927 12 1 32 1 2,426 62 1,091 5 1 121 9 3,682 124
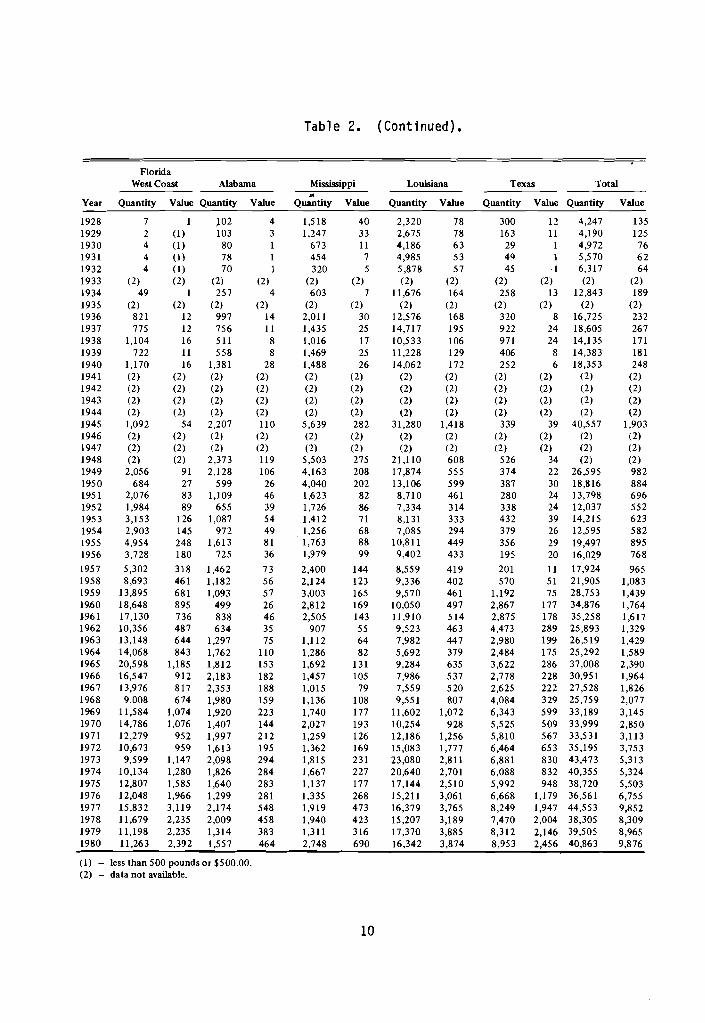
Table 2. (Continued).
Florida West Coast Alabama Mississippi Louisiana Texas Total
1
Year Quantity Value Quantity Value Quantity Value Quantity Value Quantity Value Quantity Value
1928 7 1 102 4 1,518 40 2,320 7 8 300 12 4,247 135 1929 2 (1) 103 3 1,247 33 2,675 7 8 163 11 4,190 125 1930 4 (1) 80 1 673 11 4,186 6 3 29 1 4,972 76 1931 4 (1) 7 8 1 454 7 4,985 53 49 1 5,570 6 2 1932 4 (1) 7 0 1 320 5 5,878 57 45 1 6,317 64 1933 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1934 49 1 257 4 603 7 11,676 164 258 13 12,843 189 1935 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1936 821 12 997 14 2,011 30 12,576 168 320 8 16,725 232 1937 775 12 756 11 1,435 25 14,717 195 9 22 24 18,605 267 1938 1,104 16 511 8 1,016 17 10,533 106 971 24 14,135 171 1939 722 11 558 8 1,469 25 11,228 129 406 8 14,383 181 1940 1,170 16 1,381 28 1,488 26 14,062 172 252 6 18,353 248 1941 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1942 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1943 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1944 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1945 1,092 54 2,207 110 5,639 282 31,280 1,418 3 39 39 40,557 1,903 1946 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1947 (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) (2) 1948 (2) (2) 2,373 119 5,503 275 21,110 608 526 34 (2) (2) 1949 2,056 91 2,128 106 4,163 208 17,874 555 374 22 26,595 982 1950 684 27 599 26 4,040 202 13,106 599 387 30 18,816 884 1951 2,076 83 1,109 46 1,623 82 8,710 461 280 24 13,798 696 1952 1,984 89 655 39 1,726 86 7,334 314 338 24 12,037 552 1953 3,153 126 1,087 54 1,412 71 8,131 333 432 39 14,215 623 1954 2,903 145 972 49 1,256 68 7,085 294 379 26 12,595 582 1955 4,954 248 1,613 81 1,763 88 10,811 449 356 29 19,497 895 1956 3,728 180 725 36 1,979 99 9,402 433 195 20 16,029 768 1957 5,302 318 1,462 73 2,400 144 8,559 419 201 11 17,924 965 1958 8,693 461 1,182 56 2,124 123 9,336 402 570 51 21,905 1,083 1959 13,895 681 1,093 57 3,003 165 9,570 461 1,192 75 28,753 1,439 1960 18,648 895 499 26 2,812 169 10,050 497 2,867 177 34,876 1,764 1961 17,130 736 838 46 2,505 143 11,910 514 2,875 178 35,258 1,617 1962 10,356 487 634 35 907 55 9,523 463 4,473 289 25,893 1,329 1963 13,148 644 1,297 75 1,112 64 7,982 447 2,980 199 26,519 1,429 1964 14,068 843 1,762 110 1,286 82 5,692 379 2,484 175 25,292 1,589 1965 20,598 1,185 1,812 153 1,692 131 9,284 635 3,622 286 37,008 2,390 1966 16,547 912 2,183 182 1,457 105 7,986 537 2,778 228 30,951 1,964 1967 13,976 817 2,353 188 1,015 79 7,559 520 2,625 222 27,528 1,826 1968 9,008 674 1,980 159 1,136 108 9,551 807 4,084 329 25,759 2,077 1969 11,584 1,074 1,920 223 1,740 177 11,602 1,072 6,343 599 33,189 3,145 1970 14,786 1,076 1,407 144 2,027 193 10,254 928 5,525 509 33,999 2,850 1971 12,279 952 1,997 212 1,259 126 12,186 1,256 5,810 567 33,531 3,113 1972 10,673 959 1,613 195 1,362 169 15,083 1,777 6,464 653 35,195 3,753 1973 9,599 1,147 2,098 294 1,815 231 23,080 2,811 6,881 830 43,473 5,313 1974 10,134 1,280 1,826 284 1,667 227 20,640 2,701 6,088 832 40,355 5,324 1975 12,807 1,585 1,640 283 1,137 177 17,144 2,510 5,992 948 38,720 5,503 1976 12,048 1,966 1,299 281 1,335 268 15,211 3,061 6,668 1,179 36,561 6,755 1977 15,832 3,119 2,174 548 1,919 473 16,379 3,765 8,249 1,947 44,553 9,852 1978 11,679 2,235 2,009 458 1,940 423 15,207 3,189 7,470 2,004 38,305 8,309 1979 11,198 2,235 1,314 383 1,311 316 17.370 3,885 8,312 2,146 39,505 8,965 1980 11,263 2,392 1,557 464 2,748 690 16,342 3,874 8,953 2,456 40,863 9,876
( I ) - less than 500 pounds or $500.00. (2) - data not available.

Table 3. Percent c o n t r i b u t i o n by Sta te t o t o t a l g u l f land ings o f b lue crab 1960-80.
F l o r i d a - Year west coast A1 abama M i s s i s s i p p i Louis iana Texas
merc ia l c rabb ing general l y begins i n March o r A p r i l as water temperatures r i s e above 15 OC. Greatest commercial catches u s u a l l y occur from May through August w i t h June o r J u l y as peak months. Reported landings then begin t o dec l i ne a long w i t h water tempera- tu re . These general t rends may s h i f t s l i g h t l y from month t o month, depending upon p r e v a i l i n g envi ron- mental and/or market cond i t ions .
Dominant commercial gear types used t o harvest hard b lue crabs i n t he g u l f are t raw ls , t r o t l i n e s and crab pots. Use o f po ts has increased g r e a t l y s ince 1948, w h i l e use o f t r o t l i n e s has decl ined.
aRecreati onal Harvest
Accurate da ta on t h e rec rea t i ona l catch o f crabs i n t he g u l f a re
lack ing . The s p o r t f i s h e r y i s thought t o c o n t r i b u t e s i g n i f i c a n t l y t o t o t a l f i s h i n g pressure, though est imates o f t he impact o f r e c r e a t i o n a l f i s h i n g on the resource vary w ide ly .
Tatum (1982) conse rva t i ve l y est imated t h a t t he rec rea t i ona l ca tch i n A1 abama equaled approximately 20% of t he annual commercial catch. From in te rv iews w i t h 810 spor ts f ishermen i n t he M i s s i s s i p p i coas ta l zone, He r r i ng and Christmas (1974) est imated a r e c r e a t i o n a l ca tch o f 50,000 I b o f hard crabs i n 1971. Compared t o commerci a1 1 andi ngs o f 1,259,230 I b f o r t h a t year, t h e spo r t s ca tch represented l ess than 4% o f t h e t o t a l . Data from a r e c r e a t i o n a l survey o f Gal veston Bay, .Texas, produced s imi 1 a r r e s u l t s . Benef i e l d (1968) est imated

8. ---- - .-. FLORIDA 7 - LOUISIANA
--TEXAS ----MISSISSIPPI
6 - -ALABAMA
5 .
1970 1971 1972 1073 1974 1975 1976 1977 1978 1979 1980 Years (by quarters)
Figure 4. Seasonal b lue crab landings by State, 1970-80 (Perry e t a l . 1984).
t he r e c r e a t i o n a l ca tch o f b lue crabs f o r accurate rec rea t i ona l ca tch from Galveston Bay t o be 33,125 1b s t a t i s t i c s i n o rder t o es t imate t o t a l o r 5.9% o f the commercial harves t from product ion . t h a t area.
I n Louis iana the s p o r t f i s h e r y landings were est imated t o exceed t h e commercial f i s h e r y landings by almost f o u r t imes. On the bas i s o f a s p o r t crab survey conducted by the Bureau o f Sports F i she r ies and W i l d l i f e i n 1968, t he est imated r e c r e a t i o n a l ca tch o f b lue crabs i n Louis iana was 29 m i 11 i o n l b (Linda1 1 and Hal 1 1970), compared t o commercial hard and s o f t crab re - po r ted landings o f 9.5 m i l l i o n 1b and 284,000 I b , respec t i ve l y . To ta l commerci a1 g u l f 1 andi ngs o f hard crabs f o r t he survey pe r iod were 25.7 m i l l i o n I b ; thus, t he est imated rec rea t i ona l ca tch i n Louis iana a lone exceeded the repor ted commerci a1 hard crab landings from a1 1 G u l f States i n 1968. These data emphasize t h e need
I n c i d e n t a l Harvest
I n a d d i t i o n t o t h e commercial and rec rea t i ona l hard and s o f t crab f i s h e r i e s , l a r g e numbers o f crabs are harvested as "by-catch" i n o the r f i s h e r i e s . Adkins (1972) noted t h a t commercial shrimp f ishermen i n Louis iana "eat , g i v e away, swap f o r supp l ies o r s e l l many o f t he crabs they ca tch w h i l e t r a w l i n g f o r shrimp." Adkins (1972) a l s o repor ted t h a t du r i ng t h e l a t e fa1 1 and w in te r , crabs a re f requent ly taken i n shrimp t r a w l s f o l l o w i n g s t rong c o l d f ron ts . He noted t h a t one shrimper, t r a w l i n g i n t h e mouth o f a deep bayou, caught 8,000 t o 9,000 1b o f crabp i n a s i n g l e day. These crabs were s o l d b u t

Table 4. Percent c o n t r i b u t i o n o f g u l f landings o f b lue c rab t o t o t a l U.S. b lue crab landings.
Year Percent 1960 23.3
no record o f t he t r a n s a c t i o n was made. Commerci a1 and r e c r e a t i o n a l b u t t e r f l y o r wing n e t (paupier) f ishermen a l so harves t 1 arge numbers o f crabs. According t o Adkins (1972) these "paupier" crabs a re "eaten, g iven t o f r iends , o r sold, thus n o t en te r i ng i n t o commercial landings. " Data on i n c i d e n t a l ca tch from o the r Gu l f States a re lack ing .
Factors A f f e c t i n g Commercial Landings
According t o Van Engel (1982), f l u c t u a t i o n s i n Chesapeake Bay land- ings r e s u l t p r i m a r i l y from v a r i a t i o n s i n year -c lass s t reng th and d i s t r i b u - t i o n o f t h e stock, bo th o f which he considered 1 a rge l y i n f 1 uenced by dens i ty - i ndependent environmental var iab les . Using s imple and m u l t i p l e c o r r e l a t i o n analyses t o determine the r e l a t i o n s h i p between environmental va r i ab les and subsequent harvest , Van Engel and H a r r i s (1979) found
t h a t t h ree parameters accounted f o r 86% of t h e v a r i a t i o n i n commercial 1 andi ngs from September 1965 through August 1975. These va r i ab les were i d e n t i f i e d as t h e c o o l i n g degree days a t Nor fo lk , V i r g i n i a , i n May o f t h e year of t he hatch; mer id iona l wind s t ress o f f Delaware Bay i n January f o l l o w i n g t h e year o f t he hatch; and an index o f j u v e n i l e crab abundance from the York R iver i n f a l l o f t h e year o f t h e hatch and i n t h e f o l l o w i n g sp r i ng and summer.
More (1969) l i s t e d changes i n rec ru i tmen t t o t h e f i s h e d popu la t i on and migra t ions t o and from f i s h i n g grounds as f a c t o r s i n f 1 uenci ng 1 and- ings i n Galveston Bay, Texas. I n F l o r i d a , Tagatz (1965) repor ted t h a t market cond i t i ons as w e l l as crab m ig ra t i ons and year -c lass s t r e n g t h were i n f 1 u e n t i a1 i n determi n i ng t h e l e v e l o f commercial catch. While v a r i a t i o n s i n year -c lass s t r e n g t h undoubtedly i n f l uence commercial harvest , t he use o f land ings data as an index o f a d u l t s tock abundance may be misleading.
The r e l a t i o n s h i p between com- merc ia l 1 andi ngs (b lue crabs, oys ters , penaeid shrimp) and long-term envi ron- mental f a c t o r s was i n v e s t i g a t e d by Meeter e t a l . (1979) f o r t he Apalachicola Bay es tua r ine system i n F lo r i da . They found t h a t w h i l e t he re were i n i t i a l i n d i c a t i o n s t h a t long- term f l o w from t h e Apalachicola R iver i n f 1 uenced annual commercial 1 andi ngs o f b lue crabs from F r a n k l i n County, when ca tch data from o the r species were p a r t i a l l e d out , r i v e r f l o w expla ined very 1 i t t l e o f t he annual v a r i a t i o n i n b l u e c rab harvest . The authors suggested t h a t u n i d e n t i f i e d socioeconomic va r i ab les and i n d i v i d u a l species s t r a t e g i e s r e l a t i v e t o sho r t - and long-term c l i m a t i c changes may i n p a r t be respons ib le f o r t h e lower "r" values observed w i t h p a r t i a l c o r r e l a - t i o n ana lys is . According t o Ly les (1976), f l u c t u a t i o n s i n t h e commercial ca tch o f b l u e crabs appear t o be

governed more by economic cond i t i ons than by a s c a r c i t y o f crabs. Moss (1981) noted t h a t landings do n o t necessari l y r e f 1 e c t populat ions , b u t may on ly r e f 1 e c t economic f l uctua- t i ons .
ENVIRONMENTAL REQUIREMENTS
Both dens i ty - i ndependent and density-dependent va r iab les operate t o i n f l uence l a r v a l and j u v e n i l e popu la t i on 1 evel s. The vu l nerabi 1 i t y o f b lue crabs t o changing environ- mental cond i t i ons i s perhaps greates t du r ing the l a r v a l and j u v e n i l e stages. While cu r ren t and pas t crab research has emphasized the r o l e of t he nursery area as a l i m i t i n g f a c t o r i n deter - mining the success o f a year-c lass o r modal group, cond i t i ons t h a t a f f e c t the i n i t i a l movement o f la rvae and post la rvae toward the estuary must a1 so be considered. The d i f f e r e n t i a l d i s t r i b u t i o n o f e a r l y and l a t e stage zoeae, though i t helps assure mix ing o f populat ions, subjects recru i tment t o the vagaries o f o f f sho re t ranspor t . The r o l e t h a t o f f sho re recru i tment p lays i n determining l e v e l s o f young on es tuar ine nursery grounds i s c u r r e n t l y under i nves t i ga t i on .
Laboratory s tud ies on Ca l l i nec tes la rvae i n d i c a t e t h a t there i s a behavioral bas is f o r t he v e r t i c a l d i s t r i b u t i o n o f b lue crab zoeae. Accordi ng t o Sul k i n (1981), "exper i - ments i n d i c a t e t h a t du r ing t h e course o f b lue crab zoeal development changes occur i n c r i t i c a l behavioral responses which, through ontogeny, produce a c h a r a c t e r i s t i c p a t t e r n o f d i f f e r e n t i a l v e r t i c a l d i s t r i b u t i o n . " From these observat ions he developed a d ispersa l - based recru i tment model f o r the Middle A t l a n t i c B i g h t which inc luded mechanisms f o r bo th the es tuar ine r e t e n t i o n o f la rvae and the r e c r u i t - ment o f la rvae from of fshore. He noted t h a t s i g n i f i c a n t r e t e n t i o n o f l a rvae i s most l i k e l y t o occur i n s t r a t i f i e d es tuar ies which are wide w i t h respect t o depth near the mouth.
I n such an estuary, la rvae released a t a depth below the pycnocl i n e would be re ta ined. Most f i e l d data i n d i - cate, however, t h a t C. sapidus la rvae are re leased t o surTace waters and, consequently, t ranspor ted of fshore. As these zoeae progress i n t h e i r development, they move t o deeper waters which have pronounced landward d r i f t . This d r i f t concentrates l a t e - stage zoeae and megalopae near the mouths o f estuar ies. The 1 i t e r a t u r e i n d i c a t e s t h a t recru i tment t o t h e estuary occurs i n the megalopal stage (Tagatz 1968a, More, 1969, King 1971, Perry 1975). The Sul k i n (1981) model p r e d i c t s t h a t f o r la rge, s t r a t i f i e d es tuar ies , t he re i s a low b u t s t a b l e base- 1 evel o f recru i tment v i a re ten- t i o n t h a t i s augmented by a h i g h l y v a r i a b l e degree o f rec ru i tmen t from of fshore ; i t i s the l e v e l o f t h i s o f fshore recru i tment t h a t i s responsib le f o r t he annual v a r i a t i o n s i n recru i tment success. I n small e r es tua r ies t h a t are s t r a t i f i e d aper i - od i c a l l y , o r i n which s t r a t i f i c a t i o n i s less s tab le , b lue crab recru i tment would be more s e n s i t i v e t o t h e uncer- t a i n t i e s o f t he o f f sho re l a r v a l pool and recru i tment would be more v a r i - able. Mechanisms o f 1 a rva l t r a n s p o r t and the e f f e c t s o f changing environ- mental cond i t ions on s u r v i v a l o f la rvae i n the Gu l f o f Mexico are aspects o f t h e l i f e h i s t o r y o f b lue crabs t h a t have rece ived l i t t l e a t t e n t i o n from b i o l o g i s t s .
Once the megalopae have reached the estuary, t h e major concerns f o r su rv i va l are r e l a t e d t o maintenance o f adequate h a b i t a t and favorab le en- v i ronmental cond i t ions on the nursery grounds.
Va r ia t i ons i n s a l i n i t y , tempera- tu re , po l 1 u tants , p redat ion , disease, h a b i t a t 1 oss, and food ava i 1 ab i 1 i t y a1 1 a f f e c t su rv i va l . The d i v e r s i t y o f these parameters and t h e i r poss ib le s y n e r g i s t i c e f f e c t s make prec ise i d e n t i f i c a t i o n o f t he i n f l uence o f s p e c i f i c var iab les d i f f i c u l t . Addi- t i o n a l l y , t he e f f e c t s o f va r iab les

such as s a l i n i t y may be i n t r i n s i c ( phys io l og i ca l ) o r e x t r i n s i c ( a f f e c t i n g t h e compos i t ion o f t h e b i o t i c environment). Van Engel (1982) suggested t h a t temperature, s a l i n i t y , and substratum a r e t h e p r ima ry f a c t o r s a f f e c t i n g growth, s u r v i v a l , and d i s - t r i b u t i o n o f b l ue crabs i n Chesapeake Bay. S a l i n i t y has been i d e n t i f i e d as a determinant a f f e c t i n g b l u e c rab abundance i n Texas bays (Hoese 1960, More 1969). More (1969) found an i nve rse r e l a t i o n s h i p between s a l i n i t y and t h e abundance o f j u v e n i l e crabs and noted t h a t low c rab s tocks on t h e lower Texas coas t f rom 1963 t o 1965 were assoc ia ted w i t h drought condi - t i o n s . I n c o n t r a s t , L i v i n g s t o n e t a l . (1976) noted t h a t temperature and s a l i n i t y m igh t n o t be as c r i t i c a l i n t h e determi n a t i o n o f es tua r i ne popula- t i o n l e v e l s as a r e b i o l o g i c a l para- meters. They observed t h a t b i o l o g i c a l ~ a r a m e t e r s r e l a t e d t o t r o p h i c phenomena may p l a y an impo r tan t r o l e i n es tua r i ne p o p u l a t i o n s h i f t s .
B lue c rab m o r t a l i t i e s assoc ia ted w i t h chemical and b i o l o g i c a l po l l u - t a n t s , sediment, temperature, s a l i n i t y , and d i sso l ved oxygen were d iscussed by Van Engel (1982). One o f t h e most ser ious inc idences o f chemical p o l 1 u t i o n a f f e c t i n g t h e b l u e c rab f i s h e r y occur red i n V i r g i n i a and was assoc ia ted w i t h t h e re l ease o f t h e c h l o r i n a t e d hydrocarbon Kepone i n t o t h e James R i ve r f rom t h e l a t e 1950's t o l a t e 1975. Closure o f t h e r i v e r t o commercial f i s h i n g had a severe nega t i ve e f f e c t on t h e i n d u s t r y throughout t h e Chesapeake Bay. The annual m o r t a l i t y o f young and a d u l t b l ue crabs due t o exposure t o Kepone remai ns unknown ; however, bo th com- merc ia l l and ings and j uven i 1 e c rab abundance have been lower i n t h e James than i n t h e York o r Rappahannock R ivers f o r t h e pas t 15 years (Van En e l 1982). Lowe e t a1 . (1971) r e p o r t e j Mi r e x ( c l o s e l y r e1 a ted t o Kepone) t o be t o x i c t o b l u e crabs e i t h e r as a c o n t a c t o r stomach poison.
Low l e v e l s o f d i sso l ved oxygen n o t o n l y cause m o r t a l i t y o f b l u e crabs b u t a l s o impede m ig ra t i on . Death i n c rab t r a p s due t o anoxia i s a se r i ous problem i n many areas. Tatum (1982) r e p o r t e d t h a t oxygen-de f ic ien t bot tom waters covered as much as 44% o f Mob i le Bay, Alabama, i n t h e summer o f 1971, w i t h some area f ishermen r e p o r t i n g t h a t as much as 75% o f t h e i r ca t ch was dead. Low l e v e l s o f d i s s o l v e d oxygen i n t h e deeper waters o f Chesapeake Bay and assoc ia ted t r i b u t a r i e s d u r i n g t h e summer months have a l s o been i m p l i c a t e d i n t r a p death. P e r i o d i c k i l l s o f b l u e c rabs f o l 1 owing excessive f reshwater runo f f and t h e subsequent d e p l e t i o n o f oxygen due t o r a p i d decomposit ion o f o rgan ic ma t te r were repor ted ' by Van Engel (1982).
Other mor ta l i t i e s o f b l u e c rabs have been r e l a t e d t o extreme c o l d o r t o sudden drops i n temperature (Gunter and H i ldebrand 1951, Van Engel 1978 c i t e d by Rhodes and Bishop 1979, Van Engel 1982, Couch and M a r t i n 1982) and t o r e d t i d e s (Wardle e t a l . 1975, Gunter and Ly les 1979).
Mass m o r t a l i t i e s o f b l u e c rabs occurred i n South Carol i na , Nor th Carol ina, and Georgia i n June 1966 and i n South Caro l ina and Georgia i n June 1967. While t h e pathoqenic amoeba (Paramoeba pernici'osa) " was a1 luded t o as a poss ib l e cause o f t he mor ta l i t i e s , t he re was some imp1 i c a - t i o n t h a t p e s t i c i d e s may have been involved. According t o Newman and Ward (1973), b l ue crab m o r t a l i t i e s o f g rea te r and l e s s e r magnitude have occurred du r i ng May and June w i t h Paramoeba i nvo l ved i n the m a j o r i t y o f t h e k i l l s t h a t were inves t iga ted .
Large numbers o f dead crabs have p e r i o d i c a l l y 1 i t t e r e d t he beaches o f Louis iana (Adkins 1972) and Miss is - s i p p i (Per ry 1975). The vas t m a j o r i t y o f these crabs were h e a v i l y fou led, spent females.


LITERATURE CITED
Adkins, G. 1972. A study o f t h e b lue crab f i s h e r y i n Louisiana. La. W i ld l . Fish. Comm. Tech. B u l l . 3: 1-57.
Andryszak, B. L. 1979. Abundance, d i s t r i b u t i o n and p a r t i a l descr ip- t i o n s o f rep tan t decapod crustacean la rvae c o l l e c t e d from n e r i t i c Louisiana waters i n Ju ly , 1976. Master 's Thesis. Louisiana S ta te Un ive rs i t y , Baton Rouge. 102 pp.
Benef ie ld. R. L. 1968. Survev o f t h e b lue crab ( c a l l i nectes " sapidus Rathbun) spo r t f i s h e r y o f t he Galveston Bay system 1968. Tex. Parks Wi ld l . , Coastal Fish. Proj . Rep. 1968: 35-44.
Bookhout, C. G., and J. D. Costlow, Jr . 1977. Larval deve lo~ment o f Ca l l i nec tes s i m i l i s rea red i n the labora torv . B u l l . Mar. Sci. 27: 704-
Cargo, D. G. 1958. Sizes o f crabs compared. Mary1 and Tidewater News 13(3): 1-4.
Christmas, J. Y., J r . and W. Langley. 1973. Estuar i ne i nve r teb ra tes , Miss i ss ipp i . Pages 255-317 i n J. Y. Christmas, J r . , ed. Gu l f 3 Mexico Estuar ine Inventory and Study, M iss i ss ipp i . Gu l f Coast Research Laboratory, Ocean Springs , Miss.
Church i l l , E. P., Jr . 1921. L i f e h i s t o r y o f t he b lue crab. B u l l . U. S. Bur. Fish. 36:95-128.
Costlow, J. D., J r . , and C. G. Bookhout. 1959. The l a r v a l devel- opment o f C a l l i nectes sapidus Rathbun reared i n the labora torv . B i o l . B u l l . (Woods Hole) 116(3): 373-396.
Couch, J. A. , and S. Mart in. 1982. Protozoan symbionts and re1 a ted diseases o f t he b lue crab, Ca l l i nectes sapidus Rathbun from the A t l a n t i c and Gu l f coasts o f t he Uni ted States. I n : H. M. Perry and W. A. Van E g e l , eds. Proc. Blue Crab Colloquium, October 18-19, 1979, B i 1 o x i , Miss i ss ipp i . Gu l f States Mar. F ish. Comm. 7: 71-80.
Darne l l , R. M. 1959. Studies o f t he l i f e h i s t o r y o f the b lue crab (Ca l l i nectes sapidus Rathbun) i n Louis iana waters. Trans. Am. Fish. SOC . 88(4) : 294-304.
Daugherty, F. M., J r . 1952. The b lue crab i nves t i ga t i on , 1949-50. Tex. J. Sci. 4(1):77-84.
Evink, G. L. 1976. Some aspects o f t h e b io logy o f the b lue crab, Ca l l i nectei- sapidus Rathbun, on F l o r i d a ' s g u l f coast. Master 's Thesis. u n i v e r s i t y o f F lo r i da , Ga inesv i l l e . 67 pp.
F i e d l e r , R. H. 1930. So lv ing the quest ion o f crab migrat ions. Fish. Gaz. 47(6): 18-21.

F isch le r , K. J., and C. H. Walburg. 1962. Blue crab movement i n coasta l South Carol i na , 1958-59. Trans. Am. Fish. Soc. 91(3):275-278.
Gunter, G. 1950. Seasonal popu la t i on changes and d i s t r i b u t i o n s as r e l a t e d t o sa l i n i t y , o f c e r t a i n i nverte- b ra tes o f t h e Texas Coast, i n c l u d i n g t h e commercial shrimp. Publ. I n s t . Mar. Sci . Univ. Tex. 1(2):7-51.
Gunter, G., and H. H. Hi ldebrand. 1951. Des t ruc t i on o f f i s h e s and o ther organisms on t h e south Texas coast by t h e c o l d wave o f January 28 - February 3, 1951. Ecol ogy 32(4): 731-736.
Gunter, G. , and C. H. Ly les. 1979. Local i z e d p lank ton blooms and j u b i - lees on the g u l f coast. G u l f Res. Rep. 6(3): 297-299.
Haefner, P. A., J r . , and C. N. Shuster, J r . 1964. Length i nc re - ments du r ina te rm ina l mo l t o f t he female b l ;e crab. C a l l i nectes sapidus, i n d i f f e r e n t - s a l i n i t y environments. Chesapeake Sci.
Hammerschmidt, P. C. 1982. Popula- ti on t rends and commercial harves t o f the b l ue crab C a l l i nectes sapidus Rathbun. i n Texas bavs Seotember 1978 -~ugus t 1979.
., Tex. Parks
W i l d l . , Coastal Fish. Branch Manage. Data Ser. 38: 1-69.
Hard, W. L. 1942. Ovarian growth and o v u l a t i o n i n t h e mature b l u e crab. C a l l i nectes sapi dus Rathbun. Chesapeake B i o l . Lab. Publ . 46: 1-17.
Herr ing, R . , and J. Y. Christmas, J r . 1974. Blue crabs f o r fun. . .and food. Miss. Game Fish. March-Apri l , 1974: 12-14.
Hoese, H. D. 1960. B i o t i c changes i n a bay associated w i t h t he end o f a drought. L i m ~ o l . Oceanogr. 5(3): 326- 336.
Hol land, J. S., D. V. A ld r i ch , and K. Strawn. 1971. E f f e c t s o f temperature and sa l i n i t y on growth, food conversion, s u r v i v a l and temperature res i s tance o f juven i 1 e b l ue crabs, C a l l i nectes sapi dus Rathbun. Tex. A&M Univ. Sea Grant Publ. TAMU-SG-71-222:l-166.
Judy, M. H. , and D. L. Dudley. 1970. Movements o f tagged b lue crabs i n Nor th Carol i na waters. Commer. Fish. Rev. 32(11):29-35.
King, B. D . , 111. 1971. Study o f migra tory pa t te rns o f f i s h and s h e l l f i s h through a na tu ra l pass. Tex. Parks W i l d l . Dep. Tech. Ser. 9: 1-54.
L e f f l e r , C. W. 1972. Some e f f e c t s o f temperature on t h e growth and metabol ic r a t e o f j u v e n i l e b lue crabs, C a l l i nectes sapidus, i n t h e 1 aboratory. Mar. B i 01. 14: 104-110.
Linda1 1, W. N. , and J. R. Hal 1. 1970. F ishery resources: r e p o r t o f t h e commercial f i s h e r y work u n i t (b lue crab sect ion) . Nat iona l Marine F i she r ies Serv ice B i o l o g i c a l Labora- t o r y , St. Petersburg, F lo r i da . pp. 163-188.
L iv ings ton, R. J. , G. J. Koby l i nsk i , F. G. Lewis 111, and P. F. Sheridan. 1976. Long-term f 1 u c t u a t i ons o f ep ibenth ic f i s h and i n v e r t e b r a t e popul a t i ons i n Apal ach ico l a Bay, F lo r i da . U.S. Na t l . Mar. F ish. Serv. Fish. B u l l . 74(2): 311-321.
Lowe, J. I. , P. R. Par r ish , A. J. Wilson, J r . , P. D. Wilson, and T. W. Duke. 1971. E f f e c t s o f mirex on se lec ted es tua r ine organisms. Trans. 36 th N. Am. W i l d l . Nat. Resour. Conf: 171-186.
Lyles, C. H. 1976. A Management Plan f o r M i s s i s s i p p i ' s Marine F isher ies . M i s s i s s i p p i Marine Conservation Commission, B i 1 o x i . 103 pp.

Meeter, D. A. , R. J. L iv ings ton, and G. C. Woodsum. 1979. Long-term c l imatol og i ca l cycles and popul a- t i o n changes i n a r iver-dominated es tuar ine system. - In: R. J. L i v i ngston, ed. Ecological processes i n coasta l and marine systems. Mari ne Science 10: 315-338.
Menzel, R. W. 1964. Report on pre- l i m i n a r y s tud ies on t h e b lue crab i n A l l i g a t o r Harbor and adjacent Gu l f o f Mexico w i t h some observa- t i o n s on stone crab larvae. Oceanographic I n s t i t u t e , F l o r i d a Sta te Un ive rs i t y , Tal lahassee. 12 PP.
More, W. R. 1969. A c o n t r i b u t i o n t o the b io loav o f t he b lue crab (Ca l l i n e c t G sapidus Rathbun) i n Texas. w i t h a d e s c r i ~ t i o n o f t h e f i shery . Tex. ~ a r k s ' w i l d l . Dep. Tech. Ser. 1: l-31.
Moss, C. G. 1981. Nat ional Blue Crab I n d u s t r y Associat ion, r e p o r t from the Gu l f States. Mimeo. 20 PP.
Newcombe, C. L. 1945. The b io logy and conservat ion o f t he b lue crab, C a l l i nectes sapidus Rathbun. Va. Fish. Lab. Educ. Ser. 4: l-39.
Newcombe, C. L., M. D. Sandoz, and R. Rogers-Tal be r t . 1949. D i f f e r e n t i a l growth and moul t ing c h a r a c t e r i s t i c s of t he b lue crab. Ca l l i nectes Rathbun. J. Exp. Zool.
Newman, M., and G. W. Ward, J r . 1973. An ep izoo t i c o f b lue crabs, C a l l i nectes sapidus, caused by Paramoeba pe rn i ciosa. J. I nve r teb r . Path01 . 22: 329-334.
on t h e Apalachicola Drainage System, 23-24 Apr i 1 1976, Gai nesvi 11 e, F lo r i da . Fla. Mar. Res. Publ. 26: 101-121.
Per re t , W. S. , W. R. Latapie, J. F. Po l la rd , W. R. Mock, G. B. Adkins, W. J. Gaidry, and J. C. White. 1971. Fishes and inve r teb ra tes c o l 1 ected i n t r a w l and seine samples i n Louisiana es tuar ies . Sect ion I. Pages 39-105 i n Cooperative Gul f o f Mexico - estuar ine i nven to ry and study. Phase I V . B i 01 ogy. Louis iana W i l d l i f e and F i she r ies Commission, Baton Rouge.
Perry, H. M. 1975. The b lue crab f i s h e r y i n M iss i ss ipp i . Gu l f Res. Rep. 5(1): 39-57.
Perry, H. M., and K. C. Stuck. 1982a. The l i f e h i s t o r y o f t h e b lue crab i n M iss i ss ipp i w i t h notes on l a r v a l d i s t r i b u t i o n . I n : H. M. Perry and W. A. Van Engel, eds. Proc. Blue Crab Colloquium, Oct. 16-18, 1979, B i l o x i , Miss i ss ipp i . Gu l f States Mar. Fish. Comm. 7:17-22.
Perry, H. M., and K. C. Stuck. 1982b. F isher ies moni to r ing and assessment p r o j e c t repo r t , b lue crab sect ion. Completion Report (PL 88-309, P r o j e c t 2-296-R). Gu l f Coast Research Laboratory, Ocean Springs , Miss.
Perry, H. M. , G. Adkins, R. Condrey, P. Hammerschmidt, S . Heath, J. Herr ing, C. Moss, G. Perkins, and P. Steele. 1984. A p r o f i l e o f t he b lue crab f i s h e r y o f t he Gu l f o f Mexico. Gu l f States Mar. Fish. Comm., Tech. Rep. 9:l-84.
Oester l ing, M. L., and G. L. Evink. Por ter , H. J. 1955. V a r i a t i o n i n 1977. Re la t ionsh ip between morphometry o f t he a d u l t female F l o r i d a ' s b lue crab popu la t ion b lue crab, C a l l i nectes sapidus and Apalachicola Bay. I n : Rathbun. Master 's Thesis. R. J. L iv ings ton and E. A. JOYG, U n i v e r s i t y o f Delaware, Newark. eds. Proceedings o f t h e Conference 69 PP.

Rathbun, M. J. 1930. The cancro id crabs of America o f the f a m i l i e s Euryal i dae, Portunidae, Ate1 ecy- c l i dae, Cancridae and Xanthidae. U.S. Na t l . Mus. B u l l . 152:l-609.
Rhodes, R. J. , and J. M. Bishop. 1979. Management p lann ing p r o f i 1 e f o r the South Caro l i na b lue crab f i s h e r y . F i n a l complet ion r e p o r t f o r SC Coastal Grant - in -a id Award Cont rac t No. 04-8-M01-365. October 1979. 6 1 pp.
Sandi fer , P. A. 1973. D i s t r i b u t i o n and abundance o f decapod crustacean l a rvae i n t he York R ive r Estuary and adjacent lower Chesapeake Bay, V i r g i n ia , 1968-1969. Chesapeake Sci . 14(4): 235-257.
Smyth, P. 0. 1980. Ca l l i nec tes (Decapoda: Portunidae) 1 arvae i n the Middle A t l a n t i c B igh t , 1975- 77. U.S. N a t l . Mar. F ish. Serv. F ish. B u l l . 78(2): 251-265.
Steele, P. 1984. Report on b lue crab tagg ing study conducted by t h e F l o r i d a Department o f Natura l Resources. Paper presented a t t he f a l l meeting o f t he G u l f S ta tes Mari ne F i she r ies Commission, New Or1 eans, Louisiana. October, 1984.
Stuck, K. C. , and H. M. Perry. 1981. Observations on the d i s t r i b u t i o n and seasonal i t y o f p o r t u n i d mega1 opae i n M i s s i s s i p p i coas ta l waters. Gul f Res. Rep. 7(1): 93-95.
Stuck, K. C . , and H. M. Perry. 1982. Morphological c h a r a c t e r i s t i c s o f b lue crab 1 arvae, C a l l i nectes sapidus Rathbun, from the no r the rn G u l f o f Mexico. G u l f States Mar. Fish. Comm. Completion r e p o r t 000-011. 53 pp.
Stuck, K. C . , S. Y. Wang and H. M. Perry. 1981. A p r e l i m i n a r y study o f t he i d e n t i f i c a t i o n o f meaa lo~ae and e a r l v c rab staaes o f ~ a r l i n e c t e s sapid& and C. G m i l i s from M i s s i s s i ~ ~ i coas ta i waters. J. Miss. Acad. ' ~ c i . 26: 121.
Su lk in , S. D. 1981. The behav iora l bas i s f o r b lue crab rec ru i tmen t i n m id -A t l an t i c es tuar ies . Univ. Md. Sea Grant Pub1 . UM-SG-TS-81-07: 1- 62.
Swingle, H. A. 1971. B io logy o f A1 abama es tuar ine areas - coopera- t i v e G u l f o f Mexico es tua r ine inventory . Ala. Mar. Res. B u l l . 5: 1-123.
Tagatz, M. E. 1965. The f i s h e r y f o r b lhe crabs i n t h e St. Johns R iver , F l o r i d a , w i t h spec ia l re ference t o f l u c t u a t i o n i n y i e l d between 1961 and 1962. U.S. F i s h W i l d l . Serv. Spec. Sci. Rep. Fish. 501:l-11.
Tagatz, M. E. 1968a. B io logy o f t he b l ue crab, C a l l inec tes sapidus Rathbun, i n t he St. Johns R iver , F lo r i da . U.S. F i sh Wi ld l . Serv. Fish. B u l l . 67(1): 17-33.
Tagatz, M. E. 1968b. Growth o f juven i 1 e b l ue crabs, C a l l i nectes sapidus Rathbun, i n t he St . Johns R iver , F lo r i da . U.S. F i s h W i l d l . Serv. F ish. B u l l . 67(2): 281-288.
Tatum, W. M. 1980. The b lue crab f i s h e r y o f Alabama. Pages 211-220 i n H. Loyacano and J. Smith, eds. - Symposium on t h e na tu ra l resources o f t he Mobi le Estuary, Alabama. MASGP-80-022
Tatum, W. M. 1982. The b lue crab f i s h e r y o f A1 abama. I n : H. M. Perry and W. A. Van ~ n g 3 , eds. Proc. Blue Crab Colloquium, Oct. 18-19, 1979, B i l o x i , M iss i ss ipp i . G u l f States Mar. Fish. Comm. 7: 23-28.

Van Den Avyle, M. J. , and D. L. Fowler. 1984. Species p r o f i l e s : 1 i f e h i s t o r i e s and environmental requ i re- ments o f coasta l f i s h e s and inver tebra tes (South A t l a n t i c ) - - b lue crab. U. S. F i sh Wi ld l . Serv. FWS/OBS-82/11.19. U. S. Army Corps o f Engineers, TR EL-82-4. 16 pp.
Van Engel, W. A. 1958. The b lue crab and i t s f i s h e r y i n Chesapeake Bay. Pa r t I . Reproduction, e a r l y develop- ment, growth and migra t ion . Comm. Fish. Rev. 20(6):6-17.
Van Engel, W. A. 1978. Cl imato- l o g i c a l e f f e c t s on d i s t r i b u t i o n , ca tch and abundance o f t h e b lue crab. Paper presented a t t h e Nat iona l She1 l f i s h e r i e s Associat ion Meeting, New Orleans, La.
Van Engel, W. A. 1982. Blue crab mor ta l i t i e s associated w i t h pes t i c ides , herb ic ides , tempera- tu re , s a l i n i t y , and d isso lved oxygen. I n : H. M. Perry and W. A. vanEnge1, eds. Proc. Blue Crab Col loquium, Oct. 18-19, 1979, B i l o x i , M iss i ss ipp i . Gu l f States Mar. Fish. Comm. 7:89-92.
Van Engel, W. A. , and R. Har r is . 1979. Crabs and c l imate. Progress r e p o r t t o Nat ional Marine F i she r ies Service, October 1, 1979 through September 30, 1980. Contract No. 03-78-D01-35. 10 pp.
Wardle, W. J., S. M. Ray, and A. S. A ld r i ch . 1975. M o r t a l i t y o f marine organisms associated w i t h o f f sho re summer blooms o f t he t o x i c d i n o f l a g e l l a t e Gonyaulax moni 1a ta Howell a t Galveston. Texas. Paaes 257-263 i n ~ r o c e e d i n$s F i r s t inter- na t iona l Conference on Toxic Dino- f l a g e l l a t e Blooms. Massachusetts Science and Technology Foundation, Wakef i e l d .
Wi l l iams, A. B. 1971. A ten-year study o f meroplankton i n Nor th Carol i na es tuar ies : annual occurrence o f some brachyuran developmental stages. Chesapeake Sci. 12:53-61.
Wi l l iams. A. B. 1974. The swimmina crabs * o f t h e genus c a l l inecte; (Decapoda: Portunidae). U. S. Nat l . Mar. Fish. Serv. Fish. B u l l . 72(3): 685- 798.

PAGE 4. MI. a d s ~ m n l a I L Rmwr( Data
I 10171 -101
I SPECIES PROFILES: LIFE HISTORIES AND ENVIRONMENTAL REQUIREMENTS OF COASTAL FISHES AND INVERTEBRATES (GULF OF MEXICO) -- BLUE CRAB
I REPORT DOCUMENTATION I 1. "C'OnT *- 12. Y. Rw~r*nc's ~rcassaon NO. I
12. Seonwnna O m a n l u t h Nama and Addnu ('3
7. AUthoN.)
H. M. Pe r ry and T. D. McI lwa in i. hdomu~ne Oqan~rat~on ~ a n m and Abdns.
G u l f Coast Research Labora to ry East Beach Ocean Spr ings, MS 39564
- -
Na t iona l Coastal Ecosys tems Team U. S. Army Corps of Engineers F i sh and W i l d l i f e Se rv i ce Waterways Experiment S t a t i o n U.S. Department o f t h e I n t e r i o r P.O. Box 631 Washington, DC 20240 Vicksburg, MS 39180
a h d o m m g Organwatlon Rem. No.
10. h ~ u t / ~ a s r ~ o r b Unit NO.
11. bntrmet(O a Crant(G) NO.
t o
1L Su~plamantav Net- '
U.S. Army Corps o f Engineers Report No. TR EL-82-4
I& A b s t M (Limit: 200 wro.)
Species p r o f i l e s a r e summaries o f t h e 1 i t e r a t u r e on taxonomy, morphology, range, 1 i f e h i s t o r y , and environmental requirements o f c o a s t a l a q u a t i c species. They a r e designed t o a s s i s t i n env i ronmenta l impact assessment. The b l u e crab, C a l l i n e c t e s sapidus, i s common i n t i d a l marsh e s t u a r i e s and coas ta l waters o f t h e G u l f o f Mexico, occupying a v a r i e t y o f h a b i t a t s depending upon t h e p h y s i o l o g i c a l requirements of each p a r t i c u l a r s tage i n i t s l i f e h i s t o r y . Spawning occurs from s p r i n g through f a 1 1 i n h i g h sa l i n i t y e s t u a r i n e and/or coas ta l waters . Development th rough t h e 7 toea1 stages r e q u i r e s approx imate ly 31 days and occurs o f f s h o r e . The megalopal s tage i s u s u a l l y completed w i t h i n a week. Recru i tment t o t h e es tua ry occurs d u r i n g the megalopal stage. M o l t t o t h e f i r s t c r a b takes p lace w i t h i n t h e es tua ry . Juven i l es e x h i b i t wide seasonal and a r e a l d i s t r i b u t i o n . Growth i s r a p i d and b l u e crabs i n t h e Gulf o f Mexico may reach m a t u r i t y w i t h i n a year . Factors a f f e c t i n g growth and s u r v i v a l i n c l u d e food a v a i l a b i l i t y , p reda t ion , substratum, a v a i l a b l e h a b i t a t , temperature, s a l i n i t y and p o l l u t a n t s . B lue crabs do n o t conform t o s p e c i f i c t r o p h i c l e v e l s and a r e c h a r a c t e r i z e d as o p p o r t u n i s t i c b e n t h i c omnivores. T h e i r d i v e r s e feed ing h a b i t s and t h e i r importance as p rey spec ies f o r a v a r i e t y of organisms make them an i n t e g r a l p a r t of coas ta l ecosystems.
- 17. Docvmmnt Analpis a. ~ e s c n p t o n
Es tua r ies Feeding h a b i t s F i s h e r i e s Crabs Oxygen Sal i n i t y L i f e c y c l e s Contaminants Temperature Growth
L ld*ntlfi*n10O*n-fnd~d Tarm.
Blue c rab C a l l i n e c t e s sapidus Temperature requirements S a l i n i t y requirements H a b i t a t requirements
c COUTI E*ldlCrouo
REGION 1 Regional Director U.S. Fish and Wildlife Service Lloyd Five Hundred Building, Suite 1692 500 N.E. Multnornah Street Portland, Oregon 97232
REGION 4 Regional Director U.S. Fish and Wildlife Service Richard B. Russell Building 75 Spring Street, S.W. Atlanta, Georgia 30303
REGION 2 REGION 3 Regional Director Regional Director U.S. Fish and Wildlife Service U.S. Fish and Wildlife Service P.O. Box 1306 Federal Building, Fort Snelling Albuquerque, New Mexico 87 103 Twin Cities, Minnesota 55 1 1 1
REGION 5 REGION 6 Regional Director Regional Director U.S. Fish and Wildlife Service U.S. Fish and Wildlife Service One Gateway Center P.O. Box 25486 Newton Corner, Massachusetts 02158 Denver Federal Center
Denver, Colorado 80225
REGION 7 Regional Director U.S. Fish and Wildlife Service 101 1 E. Tudor Road Anchorage, Alaska 99503

TAKE PRIDE in Amerzcd
DEPARTMENT OF THE IWTERIOR U.S. FISH AND Wl lDL lA SERVICE
As the Nation's principal conservation agency. the Department of the Interior has respon- sibility for most of our .nationally owned public lands and natural resources. This includes fostering the wisest use of our land and water resourcm, protecting our fish and wildlife, preserving thsenvironmental and cultural values of our national parks and historical places, and providing for the enjoyment of life through outdoor recreation. The Department as- sesses our energy and mineral resources and works to assure that their development is in the best interests of all our people. The Department also has a major responsibility for American Indian reservation communities and for paople who live in island territories under U.S. administration.