rights / license: research collection in copyright - …38107/... · jedoch die massentierhaltung...
TRANSCRIPT

Research Collection
Doctoral Thesis
Selektionsversuch auf wirtschaftlich gewichtete Merkmale -Eierlös minus Futterkosten - beim Huhn
Author(s): Marguerat-König, Catherine
Publication Date: 1988
Permanent Link: https://doi.org/10.3929/ethz-a-000579974
Rights / License: In Copyright - Non-Commercial Use Permitted
This page was generated automatically upon download from the ETH Zurich Research Collection. For moreinformation please consult the Terms of use.
ETH Library

Diss. ETH Nr. 8694
Selektionsversuch auf wirtschaftlich gewichtete Merkmale - Eierlös minus Futterkosten - beim Huhn
ABHANDLUNG
zur Erlangung des Titels eines
Doktors der Technischen Wissenschaften
der
EIDGENÖSSISCHEN TECHNISCHEN HOCHSCHULE
ZÜRICH
vorgelegt von
CATHERINE MARGUERAT - KÖNIG
Dipl. Ing. Agr. ETH
geboren am 23. November 1953
von Lutry VD und Bern
Angenommen auf Antrag von
Prof. Dr. G. Stranzinger . Referent
Prof. Dr. J. Landis. Korreferent
Dr. Chr. Hagger. Korreferent
ADAG Administration & Druck AG
Zürich 1988

Herrn Prof. G. Stranzinger danke ich für die Oberlassung des Themas, für seine Unterstützung während der Ausführung der vorliegenden Arbeit und für die Obernahme des Referates.
Herrn Dr. Ch. Hagger drücke ich meinen speziellen Dank aus für das grosse Interesse, das er der Arbeit stets entgegenbrachte, für die zahlreichen anregenden Diskussionen und Hinweise, sowie für die Obernahme des Korreferates.
Bei Frau D. Steiger-Stafl bedanke ich mich ganz herzlich für ihre ausgezeichnete technische Mitarbeit und für die moralische Betreuung während der ganzen Zeit .
Herrn Professor Landis danke ich für die Obernahme des Korreferates und die lehrreichen Diskussionen.
Danken möchte ich ebenfalls Fräulein M. Dann für ihre gewissenhafte und exakte Arbeit mit den Tieren auf der Chamau .
Im weiteren bedanke ich mich bestens bei folgenden Personen:
Herrn A. Eimasse schatten
Denzler, im Labor
für des
seine Mithilfe Institutes für
beim Trocknen der Lebensmittelwissen-
Herrn Dir. H. Schneeberger der Forschungsanstalt Grangeneuve und seinen Mitarbeiterinnen und Mitarbeitern im Labor welche mir eine wertvolle Hilfe waren beim Bestimmen der Eizusammensetzung
Frau M. Stettler und Herrn Dr. W. Kropf für die spontane Durchsicht der Dissertation
allen Assistenten der Weinbergstrasse sowie Kolleginnen und Kollegen, die mir zu irgend einem Zeitpunkt ihre Unterstützung gezeigt haben
den Verantwortlichen des Laur - Fonds der ETH Bewilligung eines Beitrages an die Druckkosten.
für die
Meinem Mann, Guy Marguerat, danke ich besonders für seine Begleitung in all diesen Jahren.

Inhaltsverzeichnis
1. Einleitung
1.1. Einführung
1. 2. Zuchtziel
1. 3.
2.
2 .1.
2.2.
2. 3.
2.4.
2.5.
3.
3 .1.
3. 2.
3. 2 .1.
3. 2. 2.
3. 2. 3.
3. 3.
3. 4 .
3. 4 .1.
3.4.1.1.
3. 4 .1. 2.
3. 4 .1. 3.
3. 4. 1. 4.
3 .4 .1. 5.
3. 4. 2.
3.4.2.1.
3. 4. 2. 2.
3. 4. 2. 3.
Selektionskriterium
Literaturübersicht
Wirtschaftlichkeit
Futterverbrauch
Futterverwertung
Eizusammensetzung
a) in Abhängigkeit vom Eigewicht
b) in Abhängigkeit vom Alter
Befiederung und Körpertemperatur
Material und Methoden
Selektionskriterium
- Eierlös
- Futterkosten
Versuchsaufbau
Herkunft der Tiere
Versuchsanordnung
Populationsgrösse
Umwelt und Haltung
Datenerfassung
Erhebungen im Stall
Futterverzehr
Körpergewicht
Legeleistung
Befiederung
Körpertemperatur
Laboruntersuchungen
Eizusammensetzung
Trockensubstanzgehalt
Proteingehalt
Seite
2
6
6
7
8
1 2
1 2
1 7
19
22
22
24
24
24
27
28
30
30
30
30
30
31
33
33
33
34
35

3. 4. 2 .4.
3. 5 .
3. 5 .1 .
3.5.2.
3 . 5. 3.
3. 5.4.
4.
4. 1.
4. 1. 1.
4.1.1.1.
4. 1. 1 . 2.
4.1. 2 .
4 .1. 3 .
4. 1 . 3. 1 .
4.1. 3 .2.
4 .1 . 4.
4 . 1 . 5.
4 .1 . 6.
4 . 2.
4 . 2 .1.
4. 2 .1 .1 .
4.2 . 1.2.
4. 2 .1. 3.
4.2.1.4.
Energiegehalt
Statistische Auswertung
Berechnete Merkmale
Genetische Parameter
Wiederholbarkeit
Einflüsse auf den Futterverbrauch
Ergebnisse
Selektionskriterium und Futterverwertung
Mittelwerte und Standardabweichungen
Selektionskriterium in den drei Generationen
Korrelierte Merkmale: Eimasse, Futterver
brauch und Futterverwertung der drei Genera
tionen
35
35
35
36
39
40
41
41
41
41
44
Korrelierte Selektionserfolge 47
Genetische Parameter 51
Genetische Korrelationen und Heritabilitäten 51
in der Selektionsperiode
Genetische Korrelationen innerhalb der
zweiten Periode sowie zwischen den beiden
Perioden und die Heritabilitäten
Einfluss der Bef iederung
Einfluss der Körpertemperatur
Schlussfolgerungen
Eizusammensetzung
Phänotypische Parameter
Mittelwerte und Standardabweichungen der
Merkmale der Eizusammensetzung der drei
Generationen bei 30 und bei 50 Wochen
Wiederholbarkeiten
Phänotypische Korrelationen der Eizusammen
setzung in den drei Generationen bei 30 und
bei 50 Wochen
Korrelierte Selektionserfolge in der Ei
zusammensetzung bei 30 und bei 50 Wochen
54
57
60
60
63
63
63
66
67
69

4.2.1.5.
4.2.2.
4.2.2 . 1.
4.2.2.2.
4.2.2.3.
4. 2. 2 .4.
4.2.2.5.
4. 2. 3.
4. 3.
5.
6.
7.
Korrelierte Selektionserfolge in der Ei
zusammensetzung bei 30 Wochen und bei 50
Wochen unter Berücksichtigung des Alters
einflusses für die Generation F2 und F3
Genetische Parameter
Genetische Korrelationen der Eizusammen
setzung in den drei Generationen innerhalb
der beiden Perioden bei 30 und bei 50 Wochen
Genetische Korrelationen der Eizusammen
setzung zwischen den Merkmalen der zwei
Untersuchungsperioden
Heritabilitäten
73
77
77
81
83
85 Regressionen des Futterverbrauches auf
verschiedenen Merkmale der Eizusammensetzung
Genetische Korrelationen zwischen dem Selek- 89
tionskriterium und den Merkmalen der Eizusam
mensetzung
Schlussfolgerungen
Schlupfergebnisse
Zusammenfassung
Summary
Literaturverzeichnis
91
94
97
101
105

1
1. Einleitung
1.1. Einführung
In praktisch allen Bereichen der landwirtschaftlichen Tier
haltung entfällt der grösste Teil der Produktionskosten auf
das Futter. In der Eiproduktion liegt der Anteil der Futter
kasten an den Gesamtkosten mit 50 - 70 t sehr hoch. Der Fut
teraufwand ist deshalb besonders bei Legehennen, wo vor
wiegend nicht betriebseigen-.:;;. Futter eingesetzt wird, für
das wirtschaftliche Ergebnis des Unternehmers von grösster
Bedeutung.
Ab Mitte der Fünfzigerjahre wurde nicht mehr nur zur Selbst
versorgung produziert, sondern zunehmend auch für den öf
fentlichen Markt. Der anhaltende Konkurrenzdruck zwang die
Geflügelhalter vermehrt zu Rationalisierungsmassnahmen. Da
jedoch die Massentierhaltung beim Geflügel schon seit ei
niger Zeit bekannt ist, kam dieser Betriebszweig stark ins
Schussfeld der öffentlichen Kritik und muss sich heute sehr
bemühen, den Tierschutzauflagen mit Bestandeskorrekturen
und anderen Produktionsmassnahmen gerecht zu werden und
gleichzeitig konkurrenzfähig zu
Verbesserung der Futterverwertung
Anliegen der Geflügelhalter.
1 .2. Zuchtziel
bleiben. Die genetische
ist somit ein wichtiges
In der Liter atur werden ver schiedene Definitionen für den
Begriff "Zuchtziel" angegeben. James (1982) hat in seiner
Arbeit unterschieden zwischen einem Zuchtziel, welches
darauf tendiert, das Idealbild eines Tieres zu verwirklichen
und dem Bestreben die wirtschaftliche Leistung eines Tieres
zu verbessern. Das Ziel dieser Arbeit besteht darin, mit
Hilfe des Selektionskriteriums "Eierlös minus Futterkasten"
einen Beitrag zur genetischen Verbesserung der Futterver-

2
wertung zu leisten, und gleichzeitig korrelierte Verände
rungen in der Eizusammensetzung zu verfolgen .
1 . 3. Selektionskriterium
Unter dem Begriff Selektionskriterium versteht James (1982)
"Gew i sse Merkmale eines Tieres ode r eines Verwandten, welche
zur Entscheidung beitragen , ob ein Tier als Elterntier für
die nächste Gener ation in Frage kommt oder nicht". Um die
wirtschaftliche Effizienz zu verbessern, wurden deshalb die
gewichteten Merkmale Eierlös und Futterkosten von der 21. -
40 . Woche erhoben und und im Selektionskriterium berück
sichtigt. Diese Zeitspanne wurde gewählt um ein Genera
tionenintervall von einem Jahr einhalten zu können.
Die wirtschaftliche Effizienz kann auf verschiedene Arten
ausgedrückt werden:
E1 Ertrag - Aufwand
E2 Ertrag / Aufwand
E3 Aufwand / Er trag
wobei hier mit den Merkmalen Ertrag und Aufwand jeweils Ei
erlös und Futterkosten gemeint sind. Das Problem besteht nun
darin, diese zwei Faktoren möglichst optimal miteinander zu
kombinieren .
Mit Daten aus einem Selektionsexperiment auf verbesserte
Futterverwertung (Heil und Pirchner, 1979) verglich Heil
(1984) die beiden Quotienten (E 2 , E3 ). Er kam zum Schluss,
dass der Quotient Eimasse pro kg verbrauchtes Futter geeig
neter sei, weil der Variationskoeffizient geringer ist , die
"Restabweichungen" besser normalverteilt sind und somit die
Vorhersagen zuverlässiger werden. Die Wiederholbarkeit über
Kontrollabschnitte von 4 Wochen ist ebenfalls grösser was

3
bedeutet , dass die Zufallsschwankungen dieses Merkmales von
geringer Bedeutung sind.
Nach Dickerson (1976) kann der Züchter versuchen, den Auf
wand zu reduzieren, weil eine steigende Produktion länger
fristig zu Preissenkungen führen wird, der Marktanteil des
halb nicht beliebig vergrössert werden kann und ein zu
grosses Angebot somit den Ertrag senken wird. Unter der
Voraussetzung eines fixen Marktes schlägt er deshalb vor,
den Quotienten Aufwand/Ertrag als Selektionskriterium zur
Verbesserung der Wirtschaftlichkeit einer Population
einzusetzen . Harris (1970) hat ebenfalls dem Quotienten
Aufwand/Ertrag den Vorzug gegeben, wenn man von einem fixen
Marktanteil ausgeht und der Ertrag somit nicht gesteigert
werden
leres
kann. Genet i sehe Veränderungen
Erreichen des Mastendgewichtes
am Tier wie schnel
beim Mastgeflügel,
können aber die Marktlage stark beeinflussen, indem z.B. der
Marktanteil vergrössert werden kann weil die Verkaufspreise
sinken, und der Gesamtertrag somit gesteigert werden kann.
Dies ist aber nur realistisch für Leistungen welche von
Tieren mit kurzem Produktionsintervall (Hennen} erbracht
werden , nicht aber für solche welche mehr Zeit benötigen
(Rinder) . Mann muss aber annehmen, dass der Ertrag nie ein
konstanter Faktor bleiben wird.
Die Effizienzberechnung anhand der Differenz Ertrag minus
Aufwand ist einfacher als anhand eines Quotienten, ausser es
wäre möglich ein Modell aufzustellen, welches Änderungen in
Preis und Kosten auf Grund von genetischen Veränderungen in
der Population und politischen Interessen auf längere Zeit
vorausrechnen könnte. Die Berechnung der Wirtschaftlichkeit,
gestützt auf die Preise der beiden Grössen Aufwand und Er
trag, ist einem Quotienten, welcher nur von phänotypischen
Durchschnitten abhängig ist, vorzuziehen . James (1982) be
vorzugt die Differenz von Ertrag minus Aufwand , weil diese
einerseits vom effektiven Preisverhältnis und nicht vom
Durchschnitt der beiden Merkmale abhängig ist. Andererseits
bleibt die Differenz ohne Rücksicht auf den effektiven Markt

4
gleich gross, ob der Ertrag um eine Einheit sinkt oder der
Aufwand um eine Einheit steigt, währenddessen sich die Quo
tienten verändern . Diese Oberlegungen haben die Festlegung
des verwendeten Selektionskriteriums "Eierlös minus Futter
kasten beeinflusst.
Seit 1968 werden in der Schweiz jährlich Leistungsprüfungen
für Legehennen an der schweizerischen Zentralstelle für Ge
flügel in Zollikofen durchgeführt. Der Futterverbrauch pro
Ei sank langsam aber deutlich (5.6 % in 20 Jahren). Zwischen
1972/73 und 1984/86 verbesserte sich die Futterverwertung
(kg Futter/kg Eimasse) von 2.63 auf 2.36.
Der Futterbedarf der Legehennen ist vor allem von der Ei
masseproduktion , dem Körpergewicht und dessen Veränderungen
abhängig . Neuere Publikationen weisen darauf hin, dass diese
drei Faktoren 35% - 70% der Variation des Futterbedarfes er
klären und 20t - 30% davon alleine auf die Eimasse zurückzu
führen sind (Fairfull, et al, 1984; Bentsen, 1983; Pirchner,
1980; Hagger und Abplanalp , 1978; Arboleda et al, 1976).
Unter Berücksichtigung der Leistung, des Körpergewichtes und
dessen Veränderungen kann der Futterbedarf geschätzt werden.
Durch eine gezielte Selektion derjenigen Tiere, deren
Futterbedarf unter dem vorausgeschätzten liegt, verbessert
sich die Futterverwertung und damit die wirtschaftliche
Effizienz. Ober die Futterverwertung finden sich in der
Literatur viele Angaben ( Hou, 1985; Bordas et al, 1984,
1981, 1974; Bentsen, 1983; Wing et al , 1982; Heil, 1984;
Heil et al , 1979; Nordsk.og et al, 1972). Die meisten Ex
perimente zur Verbesserung dieses Merkmales beruhten jedoch
auf indirekter Selektion über höhere Eimasseproduktion oder
tiefere Körpergewichte, weil die Erhebung des Futterverbrau
ches zur Berechnung der Futterverwertung zeit- und kosten
aufwendig ist. In den letzten Jahren wurde vor allem daran
gearbeitet, einerseits das genetische Potential der Eipro
duktion zu erhöhen und das Körpergewicht zu verringern und

5
andererseits die Umweltbedingungen zu verbessern. In einem
Selektionsexperiment auf bessere Futterverwertung (Hou et
al, 1985) hat sich gezeigt, dass ein Selektionsindex (I) mit
den Selektionsmerkmalen Körpergewicht und Eimasse wohl die
Eiproduktion steigert, jedoch gleichzeitig auch das Körper
gewicht und den Futterverbrauch erhöht. Unter Einbezug des
Futterverbrauches in den Index (II) stiegen das Körperge
wicht und der Futterverbrauch weniger an und die Futterver
wertung wurde besser als bei der Selektion mit Index ( I).
Pauw (1986) schätzte in Modellrechnungen einen um 13t grös
seren Zuchterfolg in der Futterverwertung, wenn neben dem
Körpergewicht und der Eimasse zusätzlich der Futterver
brauch berücksichtigt wurde. Pirchner und Heil (1978) haben
in ihrer Arbeit gezeigt, dass die erzielten Fortschritte in
der Futterverwertung nicht nur auf eine Steigerung der Ei
masseproduktion und eine Reduktion des Körpergewichtes zu
rückgeführt werden können, da diese zwei Merkmale nur etwa
2/3 der Verbesserung in der Futterverwertung erklären. Es
kommen demzufolge noch andere Einflüsse wie der Erhaltungs
bedarf, die Befiederung, die Körpertemperatur, die Aktivi
tät, die Körperzusammensetzung, die Eizusammensetzung oder
der Hormonhaushalt in Frage. Aus den Arbeiten von Bentsen
(1983) sowie Bordas und Merat (1981) geht hervor, dass bei
direkter Selektion auf Futterverwertung nicht nur das Kör
pergewicht sinkt, sondern dass sich auch die Eizusammen
setzung verändert. Heil (1976) stellte fest, dass die ge
netischen Korrelationen zwischen Futterverwertung und Dot
tergewicht nur leicht positiv sind. Bei einer Selektion auf
Futterverwertung sei somit keine grosse Veränderung des
Dottergewichtes zu erwarten. Bei Linien, welche auf Lege
leistung selektioniert wurden, beobachtete Frankham ( 1972)
keinen Anstieg im Futterverzehr, dafür aber signifikant
weniger Körperfett im Vergleich zu den Kontrolltieren. Er
führte dies auf eine bessere Mobilisierung der Körperre
serven zur Eisynthese zurück.

5
2. Literaturübersicht
Um die wichtigsten mit dem Selektionskriterium korrelierten
Faktoren sowie deren zusammenhänge und Auswirkungen auf wei
tere wirtschaftliche Merkmale besser analysieren zu können,
werden zuerst einige relevante Experimente diskutiert. Es
wurden umfangreiche Arbeiten publiziert, welche sich mit dem
Futterverbrauch, der Futterverwertung oder den verschiedenen
Eiqualitätsmerkmalen befassen. Bei den meisten Publikationen
liegt der Schwerpunkt auf der züchterischen Verbesserung der
Wirtschaftlichkeit der Eiproduktion.
2.1. Wirtschaftlichkeit
Die fünf wichtigsten Faktoren , welche die Unterschiede im
Einkommen bei der Legehennenhaltung beeinflussen, sind: die
je Henne produzierte Eizahl , der erzielte Preis je verkauf
tes Ei , der tägliche Futteraufwand sowie die Kosten für die
Haltung und Infrastruktur je Henne . Tabelle 1 fasst den An
teil der verschiedenen Faktoren an de r Varianz des Einkom
mens , ausgedrückt in Prozent, zusammen. Werden diese ver
schiedenen Faktoren gemeinsam berücksichtigt , können nahezu
90 t der Varianz erklärt werden.
Tabelle 1 Prozentuale Varianzanteile verschiedener
Faktoren auf das Einkommen in der Haltung
von Legehennen nach Krax (1974) .
Faktor Varianzanteil (t)
Eizahl/Henne 29.9
Preis/verkauftes Ei 25.3
Futteraufwand/Henne/Tag 13.6
Stallkosten/Jahr 10 . 7
Futterkosten/Dutzend 10.2

7
Daraus wird ersichtlich , dass die Legeleistung einen bedeu
tenderen Einfluss auf das Einkommen hat als der tägliche
Futteraufwand. Aus dieser Sicht hätten nach Simon (1967) die
Einnahmen verbessernden Faktoren eine wesent 1 ich grössere
Bedeutung fü r die Einkommenserhöhung, als die kostensenken
den Faktoren. Pauw (1986) verglich mehrere Selektionsvarian
ten mit dem Ziel die Wirtschaftlichkeit zu verbessern. Nach
Pauw, könnte ein Merkmal wie Eierlös minus Futterkosten ge
eignet sein , falls, wie berechnet , dessen Heritabilität im
Bereich von 0. 26 liegt. In jedem Fall war der Selektions
e r folg grösser in Modellen, in welchen der Futterverbrauch
als Selektionskriterium miteinbezogen wurde.
Das hier gewählte Selektionskriterium erfasst somit gleich
zeitig mehrere wichtige Merkmale . Im Eierlös einerseits wer
den sowohl die Eizahl wie auch die Eigrösse, und in den Fut
terkosten ander erseits der Futterverbrauch und die Futter
ver wertung berücksichtigt.
2.2. Futterverbrauch
Nach (Bordas et Merat, 1981, 1974) wird der Futterverbrauch
zu 35 - 70 .\ durch drei mit ihm korrelierte Faktoren, das
Körpergewicht (1), mit einer phänotypischen Korrelation (rp)
von 0.57, die Eiproduktion (2) (rp• 0.49) und die Veränderung
des Körpergewichtes (3) (rp=0.42) beeinflusst. 30 - 70 .\der
Unterschiede im Futterverbrauch sind jedoch von zahlreichen
anderen Einflüssen abhängig. Diese sind vor allem die Rasse
(Arboleda, 1976; Bordas et Merat, 1984, 1974; Nordskog et
al., 1972;), das Alter (Leeson, 1978), der Gesundheitszu
stand der Tiere , die Befiederung (Damme , 1984; Tauson et
al. , 1984, 1980; Tullet et al. , 1980; O'Neill et al., 1971),
die Haltungsform {Wegner, 1985), verschiedene Umweltein
flüsse (Leeson, 1978) die Aufrechterhaltung der Körpertem
peratur (Lee et al, 1983; O'Neill, 1971; Tasaki, 1970), die

8
Aktivität (Akbar, 1985), sowie die Qualität und die Beschaf
fenheit des Futters. Ungeeignete Fütterungseinrichtungen,
welche die Futterverschwendung fördern (Heil et al. 1980),
können den Futterverbrauch ebenfalls beeinflussen .
zusammengefasst ausgedrückt ist der Futterverbrauch die
Summe aus dem Bedarf an Nährstoffen für die Erhaltung und
die Leistung sowie aus dem Luxuskonsum und den Futterver
lusten. Um der Diskussion über den Futterverbrauch einen
klar begrenzten Umfang zu geben, wird sich diese Arbeit nur
auf seine Beziehungen zu den Merkmalen Körpergewicht und Ei
masseproduktion beschränken. In diesem Zusammenhang ist die
Futterverwertung ein wichtiges Merkmal.
2.3. Futterverwertung
In der Literatur wird die Futterverwertung je nach Autor
verschieden definiert. Eine Gegenüberstellung der verschie
denen Definitionen wie Futterverwertungsrate und Futterum
wandlungsrate gibt Pym (1982).
Die Futterverwertungsrate entspricht einer Kombination des
Verhältnisses der produzierten Eimasse zum Futterverzehr,
ohne Berücksichtigung des Energiegehaltes und der von Brody
( 1945) definierten energetischen Effizienz (Futter-Energie
im output/Futter-Energie im input). Unter Futterumwandlungs
rate wird das umgekehrte Verhältnis, bzw. Futterverzehr pro
produzierte Eimasse, verstanden. Beide Definitionen bein
halten den Bedarf für die Erhaltung und die Produktion. Je
nachdem ob die Zeit berücksichtigt wird, bis eine festge
legte Futtermenge verbraucht ist, oder ob das Zeitintervall
zwischen 2 Messungen fixiert ist, ist die Futterverwertungs
rate oder die Futterumwandlungsrate das gebräuchlichere
Mass. Das meist angewendete Kriterium ist die Futterumwand
lungsrate, (Futter/Eimasse), welche jedoch oft als Futter
verwertung bezeichnet wird. Pauw (1986) bestätigte in seinen

9
Untersuchungen die Annahme, wonach durch die Selektion auf
Wirtschaft! ichkei t die Produktion der Eimasse ansteigt. In
den meisten Fällen erhöhte sich auch der Futterverbrauch.
Hagger und Abplanalp (1978) benutzten einen Selektionsindex
um den Quotienten Ertrag/Futterkasten ~u maximieren und
kamen zu folgendem Ergebniss. Die genetische Verbesserung
einer Legehennenherde wird erreicht über eine Erhöhung der
produzierten Eimasse durch Herabsetzen der Legereife, höhe
res Eigewicht und höhere Legerate, verbunden mit einem höhe
ren Futterkonsum und einem, nicht immer, reduzierten Körper
gewicht.
Heil und Pirchner (1979) selektierten zwei reziproke Kreu
zungen über drei Generationen einerseits auf bessere Futter
verwertung und andererseits auf höhere Eizahl. Sie kamen zum
Schluss, dass die auf Eizahl gezüchtete Linie mehr und
schwerere Eier legte und ein höheres Körpergewicht und einen
höheren Futterverbrauch pro Tag hatte. Die Futterverwertung
verbesserte sich von 2.45 auf 2.34. Durch die direkte Selek
tion auf Futterverwertung sanken das Eigewicht und das Kör
pergewicht sowie der Futterverbrauch pro Tag, währenddessen
die Legeleistung leicht anstieg. Die Futterverwertung nahm
über drei Generationen von 2.46 auf 2.22.ab.
Hau (1985) selektierte in zwei Linien mit zwei verschiedenen
Indices. Im ersten (I1) dienten das Körper- und das Eige
wicht sowie der Futterverbrauch, im zweiten (I2) das Körper
gewicht und die produzierte Eimasse als Informationsmerk
male. Es zeigte sich, dass bei Anwendung beider Selektions
indices mehr Eimasse produziert wurde, dass das Körperge
wicht und der Futterverbrauch bei nach I2 selektierten Hen
nen jedoch stark anstiegen. Auch er kam zum Schluss, dass
durch den Einbezug des Futterverbrauches in einen Index, die
Futterverwertung noch stärker verbessert werden konnte.

10
Umfassende Literaturangaben weisen darauf hin, dass über
50 Jt der Varianz der Futterverwertung auf den Erhaltungs
bedarf , die Eimasse und die Veränderung des Körpergewichtes
zurückgeführt werden können (Fairfull et al., 1984; Bentsen
et al., 1983 ; Wing et al., 1982; Bordas et Merat, 1981;
Byerly et al., 1980; Pirchner , 1980; Hagger et al., 1978;
Hurnik et al. , 1977; Arboleda et al., 1976). Aus diesen
Untersuchungen geht hervor, dass die Futterverwertung durch
indirekte Selektion über Produktionssteigerung und Körper
gewichtreduzierung verbessert werden kann, dass der Erfolg
jedoch grösser ist, wenn das Merkmal Futterverbrauch in ei
nem Selektionsindex zur besseren Futterverwertung berück
sichtigt wird.
Wirtschaftlich ist es nun von Interesse , die Restvarianz des
Futterverz ehrs, definiert als Differenz zwischen beobach
tetem und auf Grund von Körpergewicht, Körpergewichtsver
änderung und Eiproduktion geschätztem Futterverzehr näher zu
untersuchen. Mögliche andere beeinflussende Faktoren müsssen
deshalb auch bei den folgenden Merkmalen gesucht werden: bei
der Befiederung (Leeson et al. 1978, Charles, 1976;) sowie
bei der Grösse der unbefiederten Teile wie Kamm, Kehllappen
oder Schenkel (Bordas et al., 1984, 1981, 1974), in der Ak
tivität (Akbar, 1985) , in der Futterverschwendung, wo täg
liche Unterschiede bis zu 100 g gefunden wurden (Heil et al.
1980) oder bei der Eizusammensetzung.
So stellten Romanoff et al . (1949) eine positive Korrelation
zwischen dem Restfutterverzehr und dem Verhältnis vom Ei
dotter zum Eiklar fest , weil der Trockensubstanzgehalt und
der Energiegehalt mit steigendem Eidot terantei 1 zunehmen.
Zum gleichen Ergebnis kam Pirchner ( 1985), da im Eidotter
mehr Nährstoffe als im Eiklar enthalten sind und dadurch
dessen Produktion mehr Futter benötigt. Bentsen (1983) und
Heil (1976) ermittelten ebenfalls positive Korrelationen,
wenn auch sehr schwache , zwischen der Futterumwandlungsrate

11
und dem Anteil Eidotter. Katle et al. (1984) weisen eben
falls auf eine signifikant positive phänotypische Korre
lation von r 0.19 zwischen Residualfutter (Abweichung
zwischen beobachtetem und berechnetem Futterkonsum) und dem
Anteil Eidotter hin. Diese Korrelationen wurden auf einen
signifikanten Unterschied im Metabolismus zurückgeführt, der
durch ein geringeres Akti vi tätsni veau der Hennen mit nied
rigerem Residualfutter zustande kam. Nach Pym (1982, 1979),
wird beim Broiler durch eine verbesserte Futterverwertung
möglicherweise der Energiegehalt im Körperzuwachs reduziert,
zum Beispiel durch eine Umverteilung des Wasser-, Fett- und
Proteingehaltes im Körper. Dies könnte sich auch dement
sprechend auf die Eizusammensetzung auswirken. Der ungleiche
Futterverzehr zwei sonst gleicher Hennen kann demzufolge
möglicherweise auch durch einen unterschiedlichen Eidotter
gehalt und mit ihm einem anderen Trockensubstanz-, Protein
und Energiegehalt zustandekommen.
Fairfull et al. (1984) weisen darauf hin, dass vielleicht
nicht alle Hennen den gleichen Futterbedarf haben um ent
weder 1 g Ei oder 1 g Körpergewicht zu produzieren.
Nach Fairful 1 et al. ( 1984) und Charles ( 1984) sind die
genetischen Korrelationen zwischen Restfutterverzehr und
Eigewicht al tersabhängig, d. h., sie erhöhen sich mit zu
nehmendem Alter der Henne. Deshalb sollte die Eiqual i tät
auch in einem möglichst späten Stadium untersucht werden.
Aus den erwähnten Arbeiten geht hervor, dass es sich lohnt,
das Merkmal Eizusammensetzung unter dem im Versuch ange
wandten Selektionskriterium zu untersuchen und dies sowohl
bei jüngeren wie bei älteren Tieren.

12
2.4. Eizusammensetzung
a) in Abhängigkeit vom Eigewicht
Schon in den fünfziger Jahren wurden Arbeiten über die Ei
qual itätsmerkmale publiziert. Dabei standen aber vor allem
das Eigewicht sowie die Anteile Eidotter und Eiklar am Eige
wicht im Vordergrund. Es war bekannt, dass das Eigewicht von
der Rasse der Tiere, vom Körpergewicht, vom Alter, von der
Legeleistung und anderen Einflüssen abhängig ist und dass
das Eigewicht seinerseits die Eizusammensetzung beeinflusst.
Die zahlreichen Faktoren, welche auf das Eigewicht einwir
ken , lassen sich jedoch auf Grund ihrer Verknüpfungen nicht
immer eindeutig auseinanderhalten.
Scott (1941) hat nachgewiesen, dass Hennen, welche Eier le
gen die schwerer sind als 3,95 % ihres Körpergewichtes, eine
schlechtere Schlupfrate aufweisen und, dass für eine gute
Schlupfrate das beste Verhältnis Eiklar zu Eidotter bei 2:1
liegt. Eine natürliche Selektion sollte deshalb extreme Ab
weichungen in den Eikomponenten verhindern, weil die Frucht
barkeit von solchen Eiern produzierenden Hennen sinkt. Ein
mittelgrosses Ei wiegt ungefähr 58 g und enthält 17 g Eidot
ter , 31 g Eiklar und 10 g Eischale mit 25 % Trockensubstanz,
52 % Rohprotein und einen Energiegehalt von 28.8 MJ/kg.
Hilfiker (1970) untersuchte die Schlupffähigkeit einer auf
hohes Anfangseigewicht (52 g) und einer auf tiefes Anfangs
eigewicht ( 44. 9 g) gezüchteten Linie. Die auf tiefes An
f angseigewicht gezüchtete Linie war der anderen in der
Schlupffähigkeit um 6.9 % in der zweiten und um 16 % in der
dritten Selektionsgeneration überlegen. Lerner (1951) prägte
den Ausdruck der "genetischen Homeostase", wonach Küken bei
einem optimalen Verhältnis vom Eidotter zum Eiklar am besten
schlüpfen werden und die Extreme durch natürliche Selektion
verschwinden. Aus Untersuchungen von Sainz (1983) an Eiern

13
von verschiedenen Spezies geht hervor, dass die Eigrösse
nicht a priori die Eizusammensetzung beeinflusst, sondern
dass genetische Rassenunterschiede vorhanden sind. Der
Dotteranteil variiert zwischen 25.5 t und 33 t (Burkhardt,
1971) wobei die Anteile Eidotter-Trockensubstanz und Eiklar
Trockensubstanz biologisch weniger von der Eigrösse abhängig
sind als ihre absoluten Gewichte (Hi 11, ,1966). Nach Marion
(1964) ist die Varianz in der Eizusammensetzung vor i allem
auf physiologische Veränderungen sowie das Alter, Umwelt
einf lüsse und die Selektionsrichtung zurückzuführen .
Durch Zucht auf höhere Eigewichte nehmen die Merkmale Dot
ter- und Eiklargewicht wegen ihrer positiven Korrelationen
mit dem Eigewicht zu (Chwalibog, 1985; Tawfik, 1981; Rodda,
1977; Hill, 1966; Jaffe, 1965; Chung, 1965;). Der Anteil Ei
dotter sinkt aber zu Gunsten des Anteils Eiklar . Den Resul
taten von Hilfiker (1970) kann man entnehmen, dass die Se
lektion auf hohes Anfangseigewicht vor allem die Eiklarmasse
und damit den Protein und Wassergehalt beeinflusste.
Ambrosen et al. (1981) beobachteten in Linien die während 4
Generationen auf Eigewicht oder Eizahl selektiert wurden,
signifikante Unterschiede in der Eigrösse und der Eizusam
mensetzung gegenüber der Kontrollinie. In der auf Eigewicht
selektierten Linie veränderte sich vor allem das Eigewicht.
Es nahm im Vergleich zur Kontrollinie stark zu und damit
verbunden lagen auch die Gewichte von Eidotter und Eiklar
über der Kontrollinie. In der auf Legeleistung selektierten
Linie veränderte sich vor allem die chemische Zusammenset
zung des Eiklares. Sein Protein- und Trockensubstanzgehalt
lagen unter den entsprechenden Werten der Kontrollinie. Der
Trockensubstanzgehalt der Gesamteimasse lag in der auf Eige
wicht selektierten Linie über demjenigen der Kontrollinie
und in der auf Legeleistung selektierten Linie war er gleich
hoch wie in der Kontrollinie. Dies mag erklärbar sein durch
die bei Hennen gleichen Alters festgestellten negativen Kor
relationen zwischen Legeleistung und Eigewicht einerseits,

14
und zwischen Eigewicht und Trockensubstanzgehalt anderer
seits.
Der Trockensubstanzgehalt liegt bei allen Untersuchungen
zwischen 24 und 31% und ist mit dem Eidotteranteil positiv
korreliert . Mit zunehmendem Eigewicht aber bei gleichem
Alter der Hennen sinken der Eidotteranteil und der Trocken
substanzgehalt in der Eimasse. Ambrosen et al. (1981) wiesen
darauf hin, dass genetisch bedingte grosse Eier einen höhe
ren Trockensubstanzgehalt im Eiklar enthalten als grosse
Eier , welche von älteren Hennen produziert wurden . Hilfiker
(1970) fand keine nachweisbaren Unterschiede im Trockensub
stanzgehalt des Eiklares zwischen einer auf hohes und einer
auf tiefes Eigewicht selektierten Linie. Nach Rodda (1977)
besteht eine positive genetische Korrelation zwischen Ei
dottergewicht und Trockensubstanzgehalt im Anteil Eidotter.
Dies ist umso erstaunlicher, als in der Natur normalerweise
jede Zunahme der produzierten Menge mit einer Aenderung der
Zusammensetzung, vor allem einer Zunahme des Wassergehaltes,
verbunden ist. Marion et al . (1965) haben auch gezeigt, dass
die Gesamteimasse von grossen Eiern einen tieferen Trocken
substanzgehal t aufweist als qie Eimasse von kleineren Eiern,
obwohl bei schweren Eiern der Trockensubstanzgehalt sowohl
im Eidotter wie im Eiklar höher ist. Dies ist darauf zurück
zuführen, dass das Verhältnis Eiklar/Eidotter bei schweren
Eiern grösser ist als bei leichteren. Die Grosseierlinie
hatte nach fünf Generationen im Durchschnitt 2 % mehr Eiklar
als die Kleineierlinie.
Bedeutend weniger Autoren haben sich mit chemischen Unter
suchungen des Eiinhaltes auseinandergesetzt und die meisten
Arbeiten stammen aus den letzten Jahren. Jaffe publizierte
1965 repräsentative Ergebnisse.

15
Tabelle 2 Zusammensetzung von Eidotter und Eiklar (nach
Jaffe, 1965).
Merkmal
Eidotter
Eiklar
Protein t
16.6
10.6
Fett t
32.6 50.0
0'1. o
Daraus geht deutlich der Unterschied zwischen Eiklar und Ei
dotter hervor, und es muss in allen Untersuchungen berück
sichtigt werden, dass 1 g Eidotter mehr Protein enthält als
g Eiklar. Weil jedoch der Anteil Eiklar am Ei ca. 63 t
gegenüber ca. 25 t Eidotter beträgt, stammt mehr Protein aus
dem Eiklar als aus dem Eidotter wie folgende Rechnung zeigt:
16.6 x 0 . 25 4.15 gegenüber 10.6 x 0.63 6.67.
Untersuchungen von Kirchgessner ( 1980) an Tieren mit ver
schiedenen Energie- und Proteinaufnahmen haben gezeigt, dass
mit steigender Proteinaufnahme das Eigewicht anstieg, der
Trockensubstanzgehal t der Eimasse leicht sank (von 32. 4 t
auf 31.6 t) und der Fettgehalt in der Trockensubstanz eben
falls von 39.3 % auf 37.3 t sank. Mit zunehmender Energie
versorgung nahm der Rohproteingehalt in der Ei trockensub
stanz etwas ab und der Energiegehalt verzeichnete nur einen
geringen Anstieg. Da sich mit steigendem Proteingehalt im
Futter auch das Eigewicht erhöht, lässt sich nach Kirch
gessner folgern, dass grosse Eier einen etwas höheren Roh
proteingehalt und tieferen Fettgehalt aufweisen. Die Korre
lationen betrugen +O. 48 zwischen Eigewicht und Prozent Ei
protein in der Trockensubstanz und -0.46 zwischen Eigewicht
und Energie in der Trockensubstanz. Nach Marion, (1965), ist
der unterschiedliche Eifettgehalt nicht alleine von der Ei
dottermasse abhängig, sondern auch vom Verhältnis der Lipide
zu anderen chemischen Stoffen im Eidotter. May (1960) schät
zte eine positive genetische Korrelation zwischen dem Eige-

15
wicht und dem Eiproteingewicht (rg ~ +0.8). Demzufolge ist
der Anteil Fett in grossen Eiern kleiner als in kleinen Ei
ern, da bei den ersten die Anteile H2o und Protein höher
sind und diese mit dem Energiegehalt negativ korreliert
sind. Nach Sibbald (1979) ist die Variation der Bruttoener
gie der Gesamteimasse, incl. Schale, zu 89 % vom Eigewicht
abhängig . Die negativen Korrelationen zwischen Energie- und
Proteingehalt bewirken mit steigendem Eigewicht eine Zunahme
des Proteingehaltes und eine Abnahme des Energiegehaltes in
der Trockensubstanz.
Bekannt ist seit längerem, dass die Heritabilität für das
Eigewicht hoch ist. Die h 2 - Schätzwerte liegen bei allen Au-
toren zwischen 0. 42 - 0. 60 (Scheinberg, 1953,
Hill , 1966, Rodda, 1977). Nach Cahaner (1979)
Jaffe, 1965,
sind die h 2-
Werte in selektierten Linien kleiner, weil die genetische
Varianz gegenüber der Kontrolllinie geringer ist. Nach Van
Tijen (1970) ist der h 2 -wert am Ende der Legeperiode meist
höher als zu Beginn, währenddessen Nagai et al. (1969) fan
den , dass die h 2-werte einiger Eiqualitätsmerkmale im Laufe
der Legeperiode sanken.
Wiederholbarkeiten wurden für die Merkmale Ei-, Eidotter
und Eiklargewicht bestimmt, und die Literaturangaben liegen
zwischen 0.55 0.76 resp. 0.42 0 . 78 und 0.55 0.70
(Ambrosen, et al. 1981, Rodda, 1977, Scheinberg, 1953). Die
niedrigste Wiederholbarkeit wurde von allen Autoren für das
Merkmal Eidottergewicht ermittelt. Mit zunehmendem Alter
wird diese grösser, weil der Dotter im Vergleich zum Eiklar
bei Legebeginn sehr unterschiedlich entwickelt ist. Mit
fortschreitendem Alter verändert er sich deshalb noch stär
ker.
Zusammenfassend kann gesagt werden, dass das Eigewicht mit
den Merkmalen der Eizusammensetzung, ausgedrückt als Quanti
tät, positiv korreliert ist, mit den gleichen Merkmalen in

17
Prozenten ausgedrückt, aber zum Teil negativ. Das Eiklar
gewicht scheint die Hauptkomponente des Eigewichtes zu sein.
Durch Selektion erzieltes höheres Eigewicht wird durch an
steigendes Eiklargewicht erreicht und dies vor allem über
einen zunehmenden Wassergehalt. Als Folge davon haben
grössere Eier von Hennen gleichen Alters einen höheren Pro
teingehalt und einen tieferen Energiegehalt.
b) in Abhängigkeit vom Alter
Bei Qualitätsmerkmalen muss berücksichtigt werden, dass ein
58 g schweres Ei einer 30 Wochen alten Henne eine andere Zu
sammensetzung aufweist als ein gleich schweres Ei einer 50
Wochen alten Henne. Der Alterseinfluss auf das Eigewicht und
die Eizusammensetzung wurde von verschiedenen Autoren unter
sucht. Schon in den 50er Jahren wiesen Peterson et al.
(1949) darauf hin, dass · die Zunahme des Eigewichtes vom er
sten zum zweiten Legejahr verhältnismässig grösser ist bei
Hennen, welche im ersten Jahr leichtere Eier legten.
Aus der Literatur ist bekannt, dass das Eigewicht mit zuneh
mendem Alter der Henne ansteigt. Anderson, (1978), wies in
seinen Untersuchungen nach, dass das Eigewicht in den ersten
16 Legewochen stärker zunimmt, als in den folgenden. Paral
lel dazu steigen sowohl die Mengen Frischeidotter und
Frischeiklar und Wasser, wie auch die Mengen Trockeneidot
ter und Trockeneiklar an . Ein durch den Alterseffekt schwer
er werdendes Ei verändert sich demzufolge ebenfalls in sei
ner Zusammensetzung: der Anteil Eidotter steigt an , während
dessen der Anteil Eiklar sinkt. Das Experiment von Hurnik
(1977), bestätigte ebenfalls den altersbedingten Anstieg des
Ei- sowie des Eidotter- und Eiklargewichtes. Mit zunehmendem
Alter und bis zu 60 Wochen fällt der Proteingehalt im Eiklar
stärker als im Eidotter (Hurnik, 1977) wobei zu berücksich
tigen ist, dass die absolute Menge Eiklar stärker zunimmt

18
und deshalb auch die absolute Menge an Protein. Der Fettge
halt im Eidotter dagegen steigt leicht an und der Cholest
eringehalt sinkt. Durch die Untersuchungen von Varadarajulu
et al. (1972) wurden die genannten Ergebnisse grösstenteils
bestätigt, mit All:!mahme des Alterseinflusses auf die Trok
kensubstanz des Eidotters. Marion et al. (1964) untersuchten
den Alterseffekt an zwei auf verschiedene Merkmale selek
tierten Linien: schwere Eier (Grosseierlinie), leichte Eier
(Kleineierlinie). Mit zunehmendem Alter wurden die Eier
schwerer und der Anteil Eidotter stieg an währenddessen der
Anteil Eiklar sank. Der Anteil Eidotter nahm in der Klein
eierlinie weniger zu. Der Trockensubstanzgehalt von Eidotter
und Eiklar sank in beiden Linien.
Cunningham (1960) hat saisonale Einflüsse und Altersein
flüsse getrennt voneinander untersucht und kam zum gleichen
Schluss wie die anderen Autoren. Eiklarvolumen, Eidotter
volumen sowie Eidotteranteil stiegen mit zunehmendem Alter
an, währenddessen der Eiklaranteil sank. Zusätzlich weist er
darauf hin, dass der Trockensubstanzgehalt im Eiklar wohl um
4.8 t sank , dass die Menge Trockensubstanz im Ei jedoch an
stieg. Hilfiker (1970) beobachtete in seinen Untersuchungen
ebenfalls eine altersbedingt stärkere Zunahme des Eidotters
als des Eiklares.
Chwal ibog ( 1985) bestätigte die altersbedingte Zunahme des
Eigewichtes zwischen der 26. bis zur 47. Lebenswoche ver
bunden mit einer Zunahme des Eidottergewichtes und des Ei
dotteranteiles . Gleichzeitig stieg der Quotient von Eidot
ter- zu Eiklargewichten von 41 % auf 48 t. Der Anteil Eidot
ter stieg von 28.8 t auf 32.6 t währenddessen der Anteil
Eiklar von 71.2 t auf 67 . 4 t sank, Der Energiegehalt stieg
in dieser Zeit von 7.1 MJ/kg Eimasse auf 7.5 MJ/kg Eimasse.
Fletcher (1981) fand mit zunehmendem Alter eine Zunahme des
Anteils Eidotter von 29 % auf 33 t und eine Abnahme des
Anteils Eiklar von 71 % auf 67 %.
Ambrosen ( 1981), hat in seinem Experiment gefunden, dass
durch den Alterseinfluss, gemessen zwischen der 26. und der

19
61. Woche, das Eigewicht, das Eiklargewicht, das Eidotter
gewicht und der Trockensubstanzgehalt im Eidotter anstiegen.
Der Proteingehalt sowie der Trockensubstanzgehalt im Eiklar
sanken.
Mit zunehmendem Alter und bis zu 60 Wochen fiel nach Hurnik
(1977) der Proteingehalt im Eiklar stärker als im Eidotter.
Dagegen stieg der Fettgehalt im Eidotter leicht an. Nach
Chwalibog (1985), stieg der Energiegehalt signifikant um
5.3 t.
2.5. Befiederung und Körpertemperatur
Der Gef iederzustand der Legehennen spielt sowohl für das
Tier selbst, als auch für wirtschaftliche Oberlegungen eine
wichtige Rolle. Bedingt durch eine schlechtere Isolierung
gegenüber der Umwelt, führen Kahlstellen zu einer erhöhten
Wärmeabgabe. Der dadurch entstehende Energieverlust kann
durch einen höheren Futterkonsum kompensiert werden.
Charles (1976) beurteilte die Befiederung mit einem 5 Punkte
System. Dies diente den meisten nachfolgenden Arbeiten in
einer mehr oder weniger abgeänderten Form als Grundlage zur
Klassierung der Befiederung. Leeson et al. (1978) wiesen in
ihren Arbeiten nach, dass "futtereffizente" Hennen ein sig
nifikant (p i 0.01) höheres Federgewicht hatten als inef
fiziente, obwohl sie sich in deren visuellen Beurteilung
nicht deutlich unterschieden. Der geringe Aussagewert der
Befiederungsklassierung mag darauf zurückzuführen sein, dass
dieses subjektive Beurteilungskriterium in der Praxis nicht
problemlos angewendet werden kann. Tauson et al. (1984)
betrachteten die Aussagekraft von Beurteilungsschemen und
kamen zum Schluss, dass immer die gleichen Personen bewerten
sollten, wenn Tiere in verschiedenem Alter beurteilt werden,
damit ein Punkteschema genügend Auskunft über den gesamten
Gefiederzustand gibt.

20
Tul let et al. ( 1980) wiesen in ihrer Untersuchung darauf
hin , dass die künstliche Entfiederung von Hals und Brust
( 1 7 % der befiederten Körperf 1 äche) den Futterverbrauch um
10% erhöhte , dass aber teilentfiederte Tiere mehr Eier leg
t en und ihr Körpergewicht gegenüber der Kontrollgruppe ge
ringer war . Die zusätzlich aufgenommene Energie genügte of
fensi c htlich nicht, die höhere Wärmeabgabe zu decken . Mög
licherweise verringern diese Tiere ihre Aktivität .
Hughes (1980) kam zu ähnlichen Resultaten. Sein Bewer
tungssystem basierte auf 6 Punkten (0= perfekt; 6= völlig
nackt). Er schreibt, dass der Verlust einer Befiederungs
einheit den Erhaltungsbedarf um 7 % erhöhte und dass Hennen
mit beschädigtem Gefieder mehr Eier legten, was jedoch nicht
dem Mehrkonsum an Futter entsprach. Die Erklärung dafür mag
folgende sein: Befiederung und Eiproduktion sind vom Hormon
haushalt abhängig . Schlechtere Legehennen haben eine gerin
gere Ovarialtätigkeit mit tieferem Oestrogen Niveau und
können deshalb Nackstellen besser neu befiedern. Nach Gonyou
et al.(1982) bewirkt die künstli c he Entfernung der Federn am
Nacken , Rücken und / oder Brust einen 5 % - 6 % erhöhten Fut
terkonsum sowie eine s ch lec hte r e Fu t t e rverwertung, aber kei
ne Veränderung des Kö r perg ewi c htes , der Eiprodukt i o n ode r
der Eimasse. Tiere, wel c he an Nacken, Brust und Rücken nicht
befiedert waren , wiesen einen 25 % höheren Futterkonsum auf,
als die befiederten. Aus den Arbeiten von Tauson et al.
(1980) geht hervor , dass ganz schlecht befiederte Tiere 27 %
mehr fressen als gut befiederte. Nach Conson (1986) besteht
eine signifikante positive Beziehung zwischen Gefiederbe
schaffenhei t und Futteraufnahme ( r P 0. 27). Hennen mit
starken Gef iederschäden haben demnach eine höhere Futter
aufnahme als Hennen mit guter Befiederung.
Grashorn et al. ( 1987) kamen in ihren Untersuchungen des
Gefiederzustandes brauner Legehybriden zum Schluss, dass die
Wi r tschaft! ichkei t der Eierzeugung durch den Bef iederungs
zustand beeinflusst wird und dass die Heritabilität mit

21
zunehmendem Alter steigt (h2M 0.41 bis 0.87). Dies weist
darauf hin, dass die genetische Veranlagung altersabhängig
zum Ausdruck kommt. Hennen mit höherer Legeleistung wiesen
auch eine schlechtere Befiederung auf.
Als Masstab für die Körpertemperatur gilt die Rektaltempe
ratur. Beim Huhn beträgt die Körpertemperatur 41 °c und wird
durch verschiedene Faktoren wie Lichteinwirkung, Tagesrhyt
mus , Geschlecht , Herkunft, individuelles Verhalten, Alter
und Eiproduktion beeinflusst. Laut Tasaki (1970) steigt die
basale Wärmeproduktion legender Henne mit einem Gewicht
zwischen 1.31 bis 2.16 kg beinahe linear zum Körpergewicht.
Bobr et al. (1977) fanden, dass die basale Temperatur in der .
Nacht 0.3 °e bis 0.7 °e tiefer liegt als bei Lichteinwirkung
und , dass sie bei Belichtungsbeginn bis in den späteren
Nachmittag leicht ansteigt, um dann wieder zu sinken. Der
beste Messzeitpunkt wäre demzufolge am späten Nachmittag
oder nachts.

22
3. Material und Methoden
3.1 . Selektionskriterium
Das im vorliegenden Versuch verwendete Selektionskriterium
entspricht dem erzielten Eierlös von der 21. bis zur 40.
Lebenswoche abzüglich der Futterkosten zwischen der 21. und
der 40. Lebenswoche . Es wird als "Ei erlös minus
Futterkosten" bezeichnet.
Ei erlös
Die Eier wurden nach Gewicht in 7 verschiedene Klassen
eingetei 1 t. In Tabelle 3 sind die verwendeten Eigewichts
klassen und Preise zusammengefasst.
Tabelle 3 Einteilung der Eier in Gewichtsklassen und
relative Preise .
Klasse
2
3
4
5
6
7
Eigewichte (g)
>70
65-69
60-64
55-59
50-54
45-49
<44
relative Preise
1 . 3
1. 15
1 . 1 0
1 • 0
0.9
0 . 7
0.5
Als absoluter Eierpreis der Gewichtsklasse 4 wurden Fr . 0.18
eingesetzt. Auf Grund der Relationen der 7 Gewichtsklassen
kann der Eierpreis für jede Klasse berechnet werden. Der
Futterpreis und der absolute Eierpreis haben keinen Einfluss
auf die Rangierung der Hennen für das im Versuch angewandte
Selektionskriterium.

23
Futterkosten
Für die Futterkosten wurde auf Grund von Angaben aus der
Bundesrepublik Deutschland ein Durchschnittspreis von
Fr. 65. -- /100 kg Futter eingesetzt. Um einen Vergleich
zwischen den Jahren zu ermöglichen, wurde er nicht der
Teuerung angepasst.
Die Rentabilität der Legehennen ist stark von den Produk
tionskosten, welche vorwiegend auf die Futterkosten zurück
zuführen sind, abhängig. Diese entsprechen dem Produkt von
Futterverbrauch und Futterpreis. Der Futterverbrauch setzt
sich aus dem Futterverzehr und den Futterverlusten zusammen.
Der Futterverzehr ist vom Körpergewicht, von der Veränderung
des Körpergewichtes sowie von der Produktion abhängig. Die
Rentabilität kann zusammengefasst folgendermassen darge
stellt werden:
Abbildung 1 Zusammenstellung der Faktoren welche die Rentabilität beeinflussen
Rentabilität
Produktionskosten Futterkosten Ei erlös
1 Futterpreis x Futterverbrauch
1 Eizahl, Eipreis,
Eigewicht
1 Futterverzehr Futterverluste
Körpergewicht Körpergewichtsveränderungen Eiproduktion
Eizusammensetzung Körpertemperatur Befiederung Aktivität

24
3.2. Versuchsaufbau
3.2.1. Herkunft der Tiere
Die Ausgangspopulation setzte sich aus einer Kreuzung zwi
schen Rhodeländer Hähnen x White Plymouth Rock Hennen zu
sammen. Rhodeländer sind amerikanischer Herkunft und ent
standen aus Kreuzungen zwischen roten malaisischen Kämpfern
und anderen Rassen asiatischen Ursprunges. Ein ausgewach
sener Hahn wiegt zwischen 3.3 - 4 kg und die Henne zwischen
2 . 6 - 3 kg. White Plymouth Rock stammen aus Nordamerika und
entstanden aus einer Kreuzung Brahman, Dominikanern und
Cochin. Ein ausgewachsener Hahn wiegt 3 - 3.6 kg , die Hennen
2 . 5 - 3 kg. Im Dezember 1982 wurden die Bruteier in Deutsch
land auf dem Basiszuchtbetrieb der Meisterhybriden in der
Forschungsanstalt Grub produziert. 50 Hähne mit je 6 ange
paarten Hennen lieferten je ungefähr 10 - 11 Bruteier . Diese
wurden im Januar 1983 in die Schweiz gebracht und kamen zum
Ausbrüten auf den Versuchsbetrieb "Chamau", der ETH Zürich.
Eine Kreuzung als Ausgangspopulation zu wählen , hat ver
schiedene Vor- und Nachteile (Hill , 1971). Die genetische
Streuung ist in Kreuzungen grösser als in reinen Linien .
Falls Heterosiseffekte, die in der Eiproduktion geradezu
eingeplant sind , auftreten, dann vor allem in Merkmalen mit
niedriger Heritabilität. Als Nachteile einer Kreuzung als
Ausgangspopulation müssen die möglichen nicht additiven
genetischen Effekte erwähnt werden, welche unter Umständen
den Selektionserfolg in der ersten Generation beeinflussen.
3.2.2. Versuchsanordnung
Für dieses Experiment standen auf dem Versuchsgut 1440
Einzelkäfige für ausgewachsene Tiere, wovo n 1200 für Hennen,
zur Verfügung. Angestrebt wird eine Verteilung von 480
Tieren pro Selekt ionsl inie und 240 in der Kontroll inie mit

2 5
einer Selektionsintensität von 15.4t auf der weiblichen
Seite.
Aus der Ausgangspopulation (F1) wurden 2 Selektionslinien
(L1 und L2) sowie eine Kontrollinie (K) erzeugt. Eine un
selektierte Kontrollinie erlaubt es, Umwelteinflüsse und
genetische Verbesserungen auseinanderzuhalten (Hill, 1972),
vorausgesetzt, dass sich die Kontrolle von Generation zu
Generation genetisch nicht verändert. Das Selektionsver
fahren und der Herdeumtrieb können wie folgt charakterisiert
werden:
Pro Selektions- und Kontrollinie werden je 20 Hähne und 80
Hennen als Zuchttiere eingesetzt,
- die männlichen Tiere werden zufällig ausgelesen, wobei
aber berücksichtigt wird, dass ein Hahn aus jeder Halb
geschwister Familie für die Erzeugung der nächsten
Generation verwendet wird,
- die weiblichen Zuchttiere der zwei Selektionslinien wurden
auf Grund ihrer Eigenleistung ausgelesen,
- in der Kontrollinie wird nicht selektiert und die Anzahl
Zuchttiere entspricht derjenigen der Selektionslinien,
- in der Kontrollinie soll von jedem Hahn ein zufällig aus
gelesener Sohn und von jeder Henne eine zufällig ausgele
sene Tochter zur Weiterzucht eingesetzt werden, was einer
seits erlaubt, den Inzuchtgrad möglichst gering zu halten
und andererseits die Leistung der Kontrollinie nicht durch
unbeabsichtigte Selektion zu beeinflussen .

25
Die Ausgangspopulat ion (F1) wurde wie folgt in zwei Se-
lektionslinien und eine Kontrollinie aufgeteilt. Aus jeder
Halbgeschwister-Familie
Familien der F1 wurde
Kontrollinie zugeordnet
und ein
jeweils
und die
bis zwei Vollgeschwister
zufäl l ig eine Henne der
restlichen Hennen wurden
zufällig auf die Selektionslinien und 2 verteilt. Die
Auswahl der Zuchttiere erfolgte in allen drei Linien nach 40
Wochen. In der ersten Generation wurden dieselben 20 Hähne
in allen drei Linien verwendet um für die drei Linien eine
möglichst ähnliche genetische Ausgangslage zu schaffen .
Diese Hähne wurden an die 80 besten Hennen der
Selektionslinien und an 80 zufällig ausgelesene Hennen der
Kontrollinie angepaart. Jedem Hahn wurden 4 Hennen aus jeder
Linie zugetei 1 t. Ab der F2 wurden pro Linie 20 Hähne zu
fällig ausgelesen und an 4 Hennen der gleichen Linie ange
paart . Das Symbol F1 wurde verwendet wei 1 es sich bei der
Ausgangspopulation , wie in 3.4.2. erwähnt , um eine F1-
Kre~zungsgeneration
und F3 für die
handelte .
nächsten
Selektio nslinien) beibehalten.
Analog wu r den
Generationen
die Symbole F2
(Kontroll- und
Di e e rste Generation wurde in 2 Bruten, die zweite Gene
ration in 3 Bruten und die dritte Generation in 4 Bruten
über küns tliche Besamung produziert. Den Hähnen wurde wenn
möglich 1 ml Sperma entnommen und unverdünnt je vier Hennen
eingespritzt.

27
3.2.3. Populationsqrösse
Abbi ldunq 2 gibt Auskunft über die Methode und die Anzahl
selektierter Tiere pro Generation.
Abbildung 2: Paarungsschema
Paarungsschema
Genera- Auf- Linie tion teilung
so! x J03f f--P
~F1 Bei 40 Wochen aus jeder HGF aus 1-2 VGF a~ie erste nach zuiW11szahl rangierte Henne.
Rest zufällig verteilt auf~L1 und auf ~L2
20 unsel. Hähne [
44
4
sel. Hennen aus L1 sel. Hennen aus L2 unsel. Hennen aus K
3 x 20~4 sel. Hennen aus L1 unsel. ~ 4 sel. Hennen aus L2 Hähne 4 unsel. Hennen aus K

28
Tabelle 4 zeigt die Aufteilung der Hennen im Alter von 40
Wochen i n den verschiedenen Generationen und Linien.
Tabelle 4 Anzahl Versuchstiere der drei Generationen (F1, F2, F3) , in den Selektionslinien (L1 und L2) und in der Kontrollinie (K), sowie die Anzahl Stichprobentiere für die Untersuchung der Eizusammensetzung.
Linie
Generation
F1
F2
F3
L1
404
422
L2 K
396 254
506 221
Total
905
1054
1149
davon auf Eizusammensetzung untersucht in den Perioden 21.- 40. Woche/41. - 60. Woche
303
220/200
227/215
Der Inzuchtgrad war nach der dritten Generation immer noch
praktisch Null.
3.3. Umwelt und Haltung
Das Versuchsgut der ETH besitzt einen eigenen Brut - und
Schlupfapparat sowie spezielle Gebäude für die Aufzucht und
die spätere Haltung von 1200 Legehennen und 240 Hähnen.
Während den ersten 14 Wochen werden die Küken auf Tiefstreu
bei künstlichem Licht in Gruppen zu ca. 110 Tieren aufgezo
gen. Danach werden die Tiere bis zur sechzigsten Lebenswoche
in fensterlosen Ställen in Einzelkäfigen verteilt auf drei
Etagen mit Einzelfütterungsvorrichtungen gehalten . Dabei
wird darauf geachtet, dass die drei Linien über beide Ställe
und alle Etagen gleichmässig verteilt sind. In beiden

2 9
Ställen beträgt die Temperatur ungefähr 20 °c, wobei in den
Wintermonaten auf ein Minimum von 18 °c geheizt werden kann
und in den heissen Sommermonaten nur die Möglichkeit besteht
den Boden zu befeuchten und die Ställe dadurch etwas zu
kühlen. Bis zur 40. Lebenswoche erhalten die Tiere 12
Stunden Tageslicht. Ab dann wird das Lichtprogramm jede
Woche um 30 Minuten bis zu einem Maximum von 14, 5 Stunden
verlängert.
Während der ersten 10 Lebenswochen werden die Hennen mit UFA
Kükenfut ter gefüttert , danach wird bis zum Legebeginn auf
UFA Junghennenfutter umgestellt und ab Legebeginn (20
Wochen) erhalten die Tiere UFA Legehennenfutter.
Der Gehalt des Legehennenfutters wurde jährlich an der
Forschungsanstalt Grangeneuve untersucht. Die Ergebnisse
sind Tabelle 5 zu entnehmen.
Tabelle 5 Durchschnittliche Ergebnisse der Analyse des
Legehennenfutters für die 3 Versuchsjahre
Trockensubstanzgehalt 90.4 .\'
Asche 14.6 %
Energie 11. 4 MJ/kg UE
Protein 1 7. 1 %
Fett 4.6 %
Die drei Proben unterschieden sich kaum in den drei Jahren.
Der Futtergehalt entsprach somit einer Stichprobe über drei
Jahre.

30
3.4. Datenerfassung
3.4.1. Erhebungen im Stall
3.4.1.1. Futterverzehr
Von der 21. bis und mit der 60. Lebenswoche wurde der
Futterverzehr durchgehend erhoben. Das Futter wurde in
Büchsen zu 300 g Nettoinhalt automatisch, mit einem Gerät
des Types 700-6 der Firma AWENA AG , abgefüllt . Die Hennen
erhielten erst eine neue Ration Futter, wenn der gesamte
Inhalt einer Büchse ohne Probleme in den individuellen
Futtertrog gegeben werden konnte, andernfalls wurde erst am
nächsten Tag gefüttert . Mit dieser Fütterungsmethode konnte
die Futterverschwendung gering gehalten werden. Jede
Futtergabe wurde am betreffenden Tag auf der Stallkarte des
Tieres eingetragen. Ueber eine Periode von 40 Wochen
ermöglicht dieses Aufzeichnungssystem ein recht genaues
Erfassen des Futterverbrauchs. Am Ende der Lebensperiode
wurde das Futter nicht zurückgewogen, da man es so richtete,
dass möglichst alles Futter aufgefressen wurde.
3.4.1.2. Körpergewicht
Das Gewicht der Hennen und Hähne wurde auf den Tag genau bei
einem Alter von 10, 20, 30, 40 und 60 Wochen erhoben und auf
+/ - 5 g gerundet.
3.4 . 1.3 . Legeleistung
In der Periode von der 21. bis zur 60. Wochen wurden die
Eier jeder
jeden Tag,
einzelnen Henne gewogen,
anschliessend wöchentlich
im ersten Legemonat
an 6 aufeinanderfol-
genden Tagen . Dies erlaubte sowohl die Festlegung des Alters
bei Legebeginn als auch die Berechnung der Legeleistung (P)
in Prozenten. Die erhobene individuelle Eizahl wurde für die

3 1
Auswertung in Abschnitten von 30 Tagen wie folgt korrigiert.
Für die Tage an welchen keine Erhebung durchgeführt wurde ,
wurde eine im Intervall von 0 bis gleichverteilte Zufa_lls
zahl (Z) gezogen. War diese Zahl Z 5 P (P• Legerate des zu
korrigierenden Abschnittes) so wurde am fehlenden Tag ein Ei
mit dem durchschnittlichen Gewicht der Periode eingesetzt .
Die Summe aller einzelnen Eigewichte ergab die pro Henne
produzierte Eimasse in den Zeitabschnitten von der 21. bis
zur 40. und der 41. bis zur 60. Woche.
3.4.1.4. Befiederung
Der Gefiederzustand wurde subjektiv erfasst . Die Klassierung
wurde mit einer 5 Punkte Skala vorgenommen. Dabei galten für
die Einteilung der Tiere folgende Anhaltspunkte:
Tabelle 6 Schema für die Klassierung der Befiederung
1 • völlig intaktes Federkleid,
2 = leichte Gefiederschäden im Halsbereich und auf der
Flügeloberfläche,
3 • deutliche entfiederte Nacktstellen (Durchmesser ca 3 cm)
am Hals und am Nacken oder auf den Flügeln,
4 • Tier nicht mehr vollständig bedeckt, grössere
Nacktstellen am Hals, Nacken und auf der Brust
5 • grosse Nacktstellen vom Hals direkt übergehend auf die
Brust (geierhaftes Aussehen).
Bewertet wurden die Tiere zum ersten Mal im Alter von 40
Wochen und zum zweiten Mal im Alter von 60 Wochen.

32
Abbildung 3: Methode zur Bestimmung der Eizusammensetzung
+ Qt> BECHER.GLAS ES
FRISC.HMASSE ~OC.KENMASSE
ALU - SCHALE L Y0'PHILl51 Fi:aT lt8h
~
~
TIEGEL kP,LORIMEIER l<~~EL-+OSS
BOMBE
TS 2. °lo ENERGIE TS M:J/KS PROTEll'-J TS 01&

33
3.4.1.5. Körpertemperatur
Die Rektaltemperaturen wurden mit Hilfe eines Temperatur
messgerätes des Typs QT 100, Prexe, der Firma Wullschleger
AG, ausgerüstet mit einer Sonde, gemessen. Dank der mög-
1 ichst konstant gehaltenen Stalltemperatur befanden sich
alle Tiere im Bereich der thermoneutralen Zone und mussten
keinen zusätzlichen Regelmechanismus z.B. Bewegung oder
Fettabbau, zur Aufrechterhaltung der Körpertemperatur
aktivieren. Die Temperatur wurde nur bei Hennen erhoben die
alle im gleichen Alter und dem gleichen Lichtprogramm aus
gesetzt waren. Zudem wurde darauf geachtet, dass 90 t der
Hennen ihr Ei bei der Temperaturmessung schon gelegt hatten.
In der F3 Generation wurden die Daten .im Alter von 30 und 50
Wochen bei all denjenigen Tieren erhoben, welche auch in die
Eiuntersuchung einbezogen waren. Für die F2 Generation
standen nur die Messungen bei 50 Wochen zur Verfügung.
3.4.2. Laboruntersuchungen
Die Eiqualitlitsmerkmale wurden in allen 3 Generationen im
Alter von 30 Wochen untersucht. Bei der zweiten und dritten
Generation wurden sie zusätzlich noch im Alter von 50 Wochen
erhoben, um den Alterseinfluss mitberücksichtigen zu können.
Aus jeder Vollgeschwistergruppe wurde zufällig eine Henne
für den Versuch ausgewählt, wobei darauf geachtet wurde,
dass die Tiere auch gleichmässig über alle Bruten verteilt
waren. Von jeder dieser Stichprobenhennen wurden während 6
aufeinanderfolgenden Tagen die Eier gesammelt und die 4
resp. 3 letzten im Labor untersucht.
3.4.2.1. Eizusammensetzung
Abbildung 3 zeigt schematisch die angewandte Methode zur Be
stimmung der Eizusammensetzung. Alle 4, resp. 3 Eier einer

34
Henne wurden einzeln gewogen und aufgeschlagen um das Ei -
klar, den Eidotter und die Schale separat zu wägen. Ober
einem Becherglas wurde das Eiklar vom Eidotter getrennt. Der
freie Eidotter wurde über ein Fi 1 terpapier gerollt um ihn
möglichst vom Eiklar zu befreien. Die Eischale wurde mit
Hilfe eines Kleenex ausgetrocknet und anschliessend sofort
gewogen. Das Eiklargewicht wurde als Differenz zwischen
Eigewicht und Eidotter und Eischale bestimmt. Dieses
Verfahren wurde angewendet , wei 1 eine Wägung des Eiklares
keinen genauen Wert ergab , da die Verluste auf dem Filter
papier und in der Eischale beträchtlich sein können . Am
Schluss wurden alle Eier einer Henne im gleichen Becherglas
zusammengemischt .
3.4.2 .2 . Trockensubstanzgehalt
Die Frischeimasse, die Summe von 4 resp. 3 Eiern einer
Henne, wurde wieder gewogen, über Nacht eingefroren und
anschliessend während 48 h im Gefriertrockner des Institutes
für Lebensmittelwissenschaft der ETH Zürich gefrierge
trocknet . Danach wurde sie während 6 Stunden offen stehen
gelassen und nochmals gewogen. Dies ergab die erste Trocken
substanzmasse. Die Masse wurde nicht sofort gewogen , wei 1
die Zeitspanne zwischen dem Wägen der ersten Probe und der
letzten im Minimum h betrug und somit das ausgesprochen
hygroskopische Protein bei den letzen Proben zuviel H2o aus
der Luft hätte aufnehmen können. Diese erste Trockensub
stanzmasse (TS 1 ) wurde noch einmal mit einem elektrischen
Küchenmesser gemahlen und in luftdichte Gläser abgefüllt.
Anschliessend wurden Trockensubstanz-, Protein- und
Energiegehalt an der Eidg. Forsc hungsanstalt, Grangeneuve ,
bestimmt. Ungefähr 2 g dieser Trockensubstanz wurden an
schliessend während 3 h bei 104 °C Hitze getrocknet. Damit

35
wurde die zweite Trockensubstanz ermittelt (TS 2 ). Die de
finitive Trockensubstanz (TS3 ) entspricht dem Produkt TS 1 x
TS 2 .
3.4.2.3. Proteingehalt
Für die Rohproteinbestimmung der gefriergetrockneten Eimasse
wurden 5 mg genau gewogen und der Gehalt mit einem Kjelfoss
Gerät halbautomatisch nach der Weender Analyse bestimmt.
3.4.2.4.Energiegehalt
Eine Menge von 1 g gefriergetrockneter Eisubstanz wurde zu
Tabletten gepresst. Der Energiegehalt, ausgedrückt in MJ/kg
TS, wurde mit einem automatischen Kalorimeter des Types
AC200 bestimmt.
3.5. Statistische Auswertung
3.5.1. Berechnete Merkmale
Da sich die genetischen Parameter im Laufe einer Produk
tionsperiode verändern können, wurde das Legejahr in 2
Abschnitte eingeteilt. Der 1. Abschnitt beinhaltet alle
Daten der Tiere von der 21. bis zur 40 . Lebenswoche und der
2. Abschnitt alle Angaben von der 41. bis zur 60. Lebens
woche. Für jeden Abschnitt wurden die Leistungsmerkmale
getrennt berechnet.
Die Eimasse entspricht der Summe aller einzelnen Eier eines
Abschnittes. Windeier wurden miteinbezogen, da die diesbe
züglichen Angaben auf den Legekarten nicht eindeutig inter
pretiert werden konnten.
Der Anteil Eidotter ergibt sich aus der Summe der 4 resp. 3
separat gewogener Eidotter geteilt durch die Anzahl Eier.

36
Aus der berechneten Differenz Eigewicht minus Eischale minus
Eidotter für die 4 resp. 3 Eier wurde der Anteil Eiklar
berechnet .
3.5.2. Genetische Parameter
Für die Schätzung der Least Squares Konstanten und der ge
netischen Parameter wurde Harveys (1976) Least Squares and
Maximum Likelihood General Purpose Program verw~ndet.
Die Heritabilitäten wurden nach folgender allgemeiner Form
der Varianzanalyse zur Schätzung der Varianz und Kovarianz
komponenten berechnet (Falconer, 1984):
Varianz- Freiheitsursache grade
Hähne
Hennen
Rest
2 C1 d
2 C1
s
FG
s-1
d-s
e-d
- C1
Durchschnitts quadrate
DQ
2 e
Erwartungswerte E (DQ)
2 C1
2e C1
2e C1 e
Die Heritabilität ist somit aus der Väterkomponente durch
folgende Beziehungen gegeben:
4 X 2
C1 s

3 7
Je nach Generation dienten die folgenden Modelle als
Grundlage:
Für die Generation F1 wurde Modell I verwendet, da die
Linien noch nicht aufgeteilt waren:
Yijkln
Yij kln µ
s. 1
d .. 1)
µ + si + dij + Stall k + Brut 1 + eijkln
Beobachtungswert am Ei nze ltier in Generation F1
Allgemeiner Mittelwert
zufälliger Effekt des i-ten Hahnes (i = 1, ... ,50)
zufälliger Effekt der j-ten Henne innerhalb des
i-ten Hahnes
Stallk =fixer Effekt der k-ten Stallseite (k = 1 , ... ,6)
Brut 1 = fixer Effekt der 1-ten Brut (1 = 1,2) 2
eijkln = zufälliger Restfehler, N (0 , o e)
Für die Generation F2 wurden die Linien ins Modell II
einbezogen, die gleichen 20 Hähne waren jedoch noch über
alle drei Linien verteilt.
Modell II:
Yijklmn µ + si + dij + Stall k + Brut 1 + Liniem + eijklmn
Yijklmn = Beobachtungswert am Einzeltier in Generation F2
µ Allgemeiner Mittelwert
s . zufälliger Effekt des i-ten Hahnes ( i = 1 ' ... '20) 1
d .. zufälliger Effekt der j-ten Henne innerhalb des 1)
i-ten Hahnes
Stallk = fixer Effekt der k-ten Stallseite (k = 1' ... '6)
Brut 1 = fixer Effekt der 1-ten Brut ( 1 = 1' . .. 3)
Liniem . fixer Effekt der m-ten Linie (m = 1' . .. 3)
eijklmn = zufälliger Restfehler, N ( 0, 2 e) Cl

38
Im Modell III Fur die Generation F3 wurde berücksichtigt,
dass die Hähne nur innerhalb Linie verwendet wurden.
Modell III:
yijklmn
Yijklmn }J.
s . mi
d .. miJ
Stallk
Brut 1 Linie
m
eijklmn
µ + Liniem + smi + dmij + Stall k 7 Brut 1 +
eijklmn
Beobachtungswert am Einzeltier in Generation F3
Allgemeiner Mittelwert
zufälliger Effekt des i-ten Hahnes der m-ten
Linie (i = 1, . . . ,60)
zufälliger Effekt der j-ten Henne innerhalb des
i-ten Hahnes der m-ten Linie
fixer Effekt der k-ten Stallseite (k = 1, . .. ,6)
fixer Effekt der 1-ten Brut (1 = 1, ... ,4)
fixer Effekt der m-ten Linie (m = 1, . . . ,3)
zufälliger Restfehler , N ( 0 , o 2 ) e
Zur Berechnung der genetischen Parameter der Eizusammen
setzung wurde folgendes Modell ohne Berücksichtigung der
Henne verwendet , wei 1 nur Nachkomme pro Vollgeschwister-
familie für die Untersuchung herangezogen wurde.
Y ijkn µ + si + Stallj + Untersuchungstagk + eijkn
yijkn
µ
s. 1
Stall . J
Untersuchungstagk
eijkn
Beobachtungswert des n-ten Nachkommen
des Hahnes i im Stall j am Untersuch
ungstag k
Allgemeiner Mittelwert
= zufälliger Effekt des i-ten Hahnes
fixer Effekt des j-ten Stalles
= fixer Effekt des Untersuchungstages k
= zufälliger Restfehler, N (0, o 2 ) e

39
3.5.3. Wiederholbarkeit
Für die Wiederholbarkeiten (R) der Messungen eines gleichen
Merkmales an einem Tier wurde folgende Schätzmethode
angewandt:
Modell V:
Yijk = µ + Analysetagi + Tierij + eijk
Yijk Beobachtungswert am Einzeltier
µ allgemeiner Mittelwert
Analysetagi = fixer Effekt des Analysetages i
Tierij = Tier j innerhalb Analysetag i
eijk zufälliger Restfehler N (0 , o 2e)
2 o T
R -- -------2 2
o T + o e
wobei:
Varianz zwischen Individuen 2 o T
2 o e = Varianz der einzelnen Messungen innerhalb Individuen
Der Selektionserfolg wurde als Abweichung zur Kontrollinie
mit den Linieneffekten der Modelle II und III geschätzt.

4 0
3.5.4. Einflüsse auf den Futterverbrauch
Modell VI wurde verwendet , um die Faktoren mittels Regres
sionsanalyse zu erörtern, welche den Futterverbrauch beein
flussen:
Mode_l_LYJ_
FCijkl = µ + Fei + sj + b1(BWijkl) + bz(BWCHijkl) +
b3(AFEjikl) + ~bn(COVijkl) + eijkl
FCijkl µ
Fe . 1
s. J
BWijkl
BWCHijkl
AFEjikl
~covijkl
= Futterverbrauch eines bestimmten Tieres (kg)
Allgemeiner Mittelwert
fixer Effekt der Federnklassen(i=1, .. 4)
zufälliger Effekt des j-ten Hahnes
= Körpergewicht bei 30, resp. 50 Wochen (kg)
Körpergewichtsveränderung zwischen der 21. und
der 40. Lebenswoche, resp. zwischen der 41. und
der 60 . Lebenswoche (kg)
Alter bei Legebeginn (Tage)
andere im Modell einbezogene kontinuierliche
Kovariablen
Folgende Kovariablen wurden in den Modellen VII X
zusätzlich berücksichtigt :
Modelle:
(VII)
(VIII)
(IX)
(X)
wobei
Eimasse inklusiv Schale (EM)
Energie in der Eimasse ohne Schale (EN)
Gesamt Eidottermasse (YM) und Eiklarmasse (AM)
Energie und Protein (P) in der Eimasse ohne
Schale
Partielle Regressionskoeffizienten
= zufälliger Restfehler, N (0, o 2 ) e

41
4. Ergebnisse
4.1. Selektionskriterium und Futterverwertung
Die Selektion hatte zum Ziel, die Wirtschaftlichkeit der
Eiproduktion zu verbessern. Zu diesem Zwecke wurde das
Selektionskriterium "Eierlös minus Futterkosten" zwischen
der 21. und der 40. Woche angewendet.
In einem ersten Teil wird der Einfluss der Selektion auf das
Selektionskriterium sowie auf die korrelierten Merkmale,
Futterverbrauch, Futterverwertung, Körpergewicht und Eipro
duktion, diskutiert.
Der Hauptteil der Untersuchung bestand jedoch in der Erfas
sung der durch die Selektion verursachten Veränderungen in
der Eizusammensetzung. Dieser Aspekt wird später behandelt.
Als Mass für die Futterverwertung wurde das Verhältnis des
Futterverbrauches zur produzierten Eimasse verwendet. Die
durch indirekte Selektion verbesserte Futterverwertung ist
somit von grösster Bedeutung für den Selektionserfolg des im
Versuch angestrebten Zuchtzieles.
4.1.1. Mittelwerte und Standardabweichungen
4 . 1.1.1. Selektionskriterium in den drei Generationen
Das Selektionskriterium "Eierlös minus Futterkosten" wurde
für jedes Jahr und jede Linie in der ersten Periode (21.-40.
Woche) berechnet. In der zweiten Periode (41. - 60. Woche)
wurde ein entsprechendes Merkmal definiert als "analoge
Differenz". Die Ergebnisse der drei Generationen sind in
Tabelle 7 zusammengefasst.
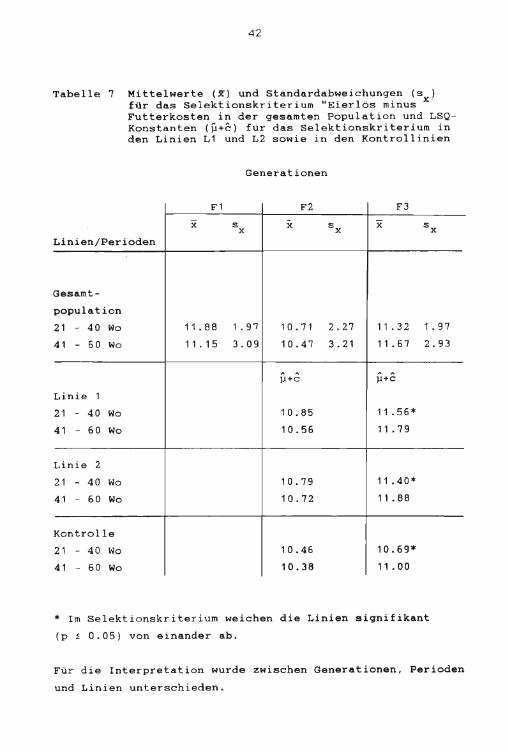
4 2
Tabelle 7 Mittelwerte (X) und Standardabweichungen (s ) für das Selektionskriterium "Eierlös minus x Futterkasten in der gesamten Population und LSQKonstanten (µ+c) für das Selektionskriterium in den Linien L1 und L2 sowie in den Kontrollinien
Generationen
F1 F2 F3
x s :X sx x s X X
Linien/Perioden
Gesamt-
population
21 - 40 Wo 11. 88 1. 91 1 0. 71 2.27 11 . 3 2 1 . 97
41 - 50 Wo 11 . 1 5 3 . 09 10.47 3. 21 11. 57 2.93
µ+c ,J+C
Linie 1
21 - 40 Wo 10.85 11. 56*
4.1 - 60 Wo 10.56 11 . 7 9
Linie 2
21 - 40 Wo 10.79 11. 40*
41 - 60 Wo 10.72 11. 88
Kontrolle
21 - 40 Wo 10.46 10.69*
41 - 60 Wo 10.38 11. 00
* Im Selektionskriterium weichen die Linien signifikant
(p i 0 . 05) von einander ab.
Für die Interpretation wurde zwischen Generationen, Perioden
und Linien unterschieden.

43
Beim Vergleich der drei Generationen muss jedoch berück
sichtigt werden, dass die Generation F1 eine kommerzielle
Kreuzung Rhode-Island x White Rock war. Beim Kreuzen zweier
Rassen ist mit einer gewissen Heterosis zu rechnen; in die
sem Versuch konnte sie jedoch nicht berechnet werden.
Die Abnahme des durchschnittlichen Selektionskriteriums
( 11 . 88) von der F1 als Gesamtpopulation zur unselektierten
Kontrollinie (10.46) in der F2 ist unter anderem auf einen
Heterosisverlust zurückzuführen. In der F2 ist schon eine
leichte Verbesserung des Selektionskriteriums der Selek
tionslinien (10.82) gegenüber der Kontrollinie (10.46)
festzustellen. In der F3 sind die Selektionslinien bereits
signifikant besser (11 . 48) als die Kontrollinie (10.69).
In den zwei ersten Generationen (F1 und F2) war das Selek
tionskriterium höher als die "analoge Differenz" . In der
dritten Generation (F3) verhielt es sich umgekehrt, die
"analoge Differenz" lag über dem Selektionskriterium. Diese
Beobachtungen galten für alle drei Linien der Generationen
F2 und F3.
Weiter konnte festgestellt werden, dass die Kontrollinie in
beiden Perioden sowohl
schlechtesten war. Die
in der Generation F2 wie in F3 am
durchschnittliche Abweichung beider
Linien von der Kontrollinie wurde definiert als Selektions
erfolg. In der Generation F2 betrug der Selektionserfolg für
das Selektionskriterium im Durchschnitt beider Linien 3.3 t
und für die "analoge Differenz" 2.4 t. In der Generation F3 waren beide Selektions! inien bereits
signifikant besser als die Kontrollinie. In der ersten
Periode betrug der Selektionserfolg im Selektionskr i ter i um
6.9 t und in der zweiten Periode für die "analoge Differenz"
7. 1 t. Diese Resultate zeigen, dass in der Generation F2
einerseits das Selektionskriterium (21. - 40. Woche) für die
Gesamtpopulation höher war als die "analoge Differenz" (41.
60. Woche) und dass andererseits die Abweichungen zur
Kontrollinie mit zunehmendem Alter kleiner wurden. In der

44
Generation F3 hingegen war das Selektionskriterium für die
Gesamtpopulation tiefer als die "analoge Differenz" und die
Abweichungen von der Kontrollinie wurden mit zunehmendem Al
ter grösser. Um daraus weitere Schlüsse ziehen zu können,
und weil sich das Selektionskriterium aus den Faktoren Eier
lös (Menge x Preis) minus Futterkosten (Menge x Preis) zu
sammensetzte, wurden die Merkmale Eimasse, Futterverbrauch
und Futterverwertung eingehender analysiert.
4 .1 .1. 2. Korrelierte Merkmale: Eimasse, Futterverbrauch
und Futterverwertung in den drei Generationen.
Die Resultate sind für die verschiedenen Merkmale in Tabelle
8 zusammengefasst.
In Bezug auf die Gesamtpopulation geben die Zahlen aus Ta
belle 8 wichtige Hinweise zur Erklärung der Veränderung im
Selektionskriterium. Wie schon im Selektionskriterium zu
sehen war, hatte die Heterosis in der ersten Generation auch
einen grossen Einfluss auf andere Leistungen.
In der ersten Generation produzierten alle Hennen zusammen
während der ersten Periode 0.3 t mehr Eimasse als während
der zweiten Periode; der Futterverbrauch der ersten Periode
lag 7 t unter demjenigen der zweiten Periode mit dem Resul
tat, dass die Futterverwertung in der ersten Periode um 9 t
besser war als in der zweiten Periode.
In der zweiten Generation lag die Eimasseproduktion in der
ersten Periode schon um 0.9 t über derjenigen in der zweiten
Periode, der Futterverbrauch war nur noch 3.2 t geringer und
die Futterverwertung der ersten Periode lag 6.2 t über der
jenigen der zweiten Periode.
In der dritten Generation hingegen produzierten die Hennen
in der ersten Periode 4.3 t weniger Eimasse als in der zwei
ten Periode, der Futterverbrauch war demzufolge um 7 . 1 t ge
ringer und die Futterverwertung um 4.2 t besser.

45
Die grosse Differenz in Bezug auf die Futterverwertung
zwischen den beiden Perioden der ersten Generation hat
verschiedene Ursachen. Im Laufe des Legejahres veränderte
Tabelle 8 Phänotypische Mittelwerte (x) oder LSQKonstanten <P+c) der Merkmale Eimasse (EM}, Futterverbrauch (FV) und Futterverwertung (FVW)
Merkmal Eimasse Futterverbrauch Futterverwertun2
Generation F1 F2 F3 F1 F2 F3 F1 F2 F3
x :X x x x x :X x x Linien/ Perioden
Gesamt-population
21 - 40 Wo 7. 16 6. 91 6.90 16 . 21 16.56 15.70 2 . 29 2.42 2.30
41 - 60 Wo 7. 14 6 . 85 7.21 17.42 17. 1 0 16.92 2 . 52 2.58 2.40
µ+~ :U+C ,.. ,..
;l+C V.+c ).l+c µ+c
Linie 1
21 - 40 Wo 6.95 6.95 16.53 15 . 64 2.40 2.27
41 - 60 Wo 6.87 7.22 17. 1 0 16 . 79 2.58 2.39
Linie 2
21 - 40 Wo 6.94 6.93 16.54 15.77 2.41 2.31
41 - 60 Wo 6. 91 7.32 17.03 17 . 08 2.54 2.39
Kontrolle
21 - 40 Wo 6.89 6. 72 16.68 15.64 2.45 2.34
41 - 60 Wo 6.85 6.93 17. 21 16. 71 2.59 2.46
sich die Produktion der Eimasse kaum (-0.3t). Der Futter
verbrauch nahm jedoch über die Legeperiode stark zu (6.7%),
was zu einem erheblichen Unterschied in der Futterverwertung

45
zwischen den beiden Perioden führte . In der zweiten Genera
tion nahm die Eimasseproduktion zwischen 21 und 60 Wochen
ebenfalls kaum ab (-0.9 t), der Futterverbrauch erhöhte sich
jedoch weniger als in der Generation F1 ( 3 . 2 t), was die
kleinere Differenz zwischen den beiden Perioden für das
Merkmal Futterverwertung erklärt . In der dritten Generation
hingegen stieg die Eimasseproduktion innerhalb der Legeper
iode um 4. 3 t an. Weil, bedingt durch die höhere Eimasse
produktion auch der Futterverbrauch um 7. 1 t zunahm, fiel
die Differenz in der Futterverwertung zwischen den beiden
Perioden geringer aus.
Diese Beobachtungen führen zur Frage, weshalb die Hennen der
ersten und der zweiten Generation mit zunehmendem Alter pro
Zeiteinheit weniger Eimasse produzierten und in der dritten
Generation das Gegenteil zutraf . Auskunf t dazu gibt eine Zu
sammenstellung der Legeleistung in Tabelle 1 im Anhang . In
den zwei ersten Generationen hat die durch Heterosis beding
te Frühreife mitgespielt. In der ersten Periode wurde da
durch die produzierte Eimasse grösser. Die Hennen begannen
schon mit 137.7 , resp. mit 135 Tagen zu legen , also 7 Tage
früher als in Generation F3 (142.8 Tage) . Obwohl die Eizahl
in allen drei Generationen von einer Periode zur anderen
sank , war der Rückgang in der Eizahl in der F3 Generation
etwas geringer , und gleichzeitig lag die Zunahme des Eige
wichtes um 1 t über derjenigen der Generationen F1 und F2.
Auf Grund des früheren Legebeginnes produzierten die Hennen
der F2 Generation in den ersten drei Legemonaten sogar mehr
Eier als die Hennen der F3 Generation in der gleichen Per
iode. Ab dem vierten Legemonat jedoch sank die Leistung der
Tiere der F2 Generation unter diejenige der Tiere der F3
Generation und blieb darunter. Obschon die F2 Hennen in der
ersten Periode mehr Eier legten als die F3 Hennen, erreich
ten sie in der zweiten Periode nicht mehr das Produktions
niveau der F3 Hennen. Insgesamt produzierten die Hennen der
F3 Generation, obwohl sie 7 Tage später mit Legen begann,
nur zwei Eier weniger als diejenigen der Generation F2.

47
4.1.2. Korrelierte Selektionserfolge
Die direkten und indirekten Selektionserfolge für die Merk
male Selektionskriterium, "analoge Differenz", Eimasse,
Futterverbrauch und Futterverwertung wurden als Abweichung
der Selektionslinien von der Kontrollinie mit den Modellen
II und III geschätzt. Der Selektionserfolg der n-ten Gene
ration enthält den Selektionserfolg der n-1 vorangehenden
Generationen.
Die Selektionserfolge waren positiv für das Selektions
kriterium resp. die "analoge Differenz" in beiden Perioden
und Generationen und vergrösserten sich von der F2 zu F3. In
der ersten Periode der dritten Generation war der korrelier
te Selektionserfolg auf die Futterverwertung bereits signi
fikant. Die Selektionslinien hatten in beiden Generationen
gegenüber den entsprechenden Kontrollinien eine höhere Ei
masseproduktion und eine bessere Futterverwertung. Der Fut
terverbrauch der Selektionslinien war in der zweiten Genera
tion tiefer als derjenige der Kontrollinien.
In Tabelle 9 sind die Selektionserfolge für die wichtigsten
Merkmale aufgeführt .
Die Linie 1 veränderte sich im Selektionskriterium von einer
Generation zur andern stärker als die Linie 2. Da die Unter
schiede aber sehr gering ausfielen, waren die Ursachen kaum
abzuklären. Die Differenz könnte auf die frühere Legereife
der ersten Linie sowie auf ihr leicht höheres Anf angseige
wicht zurückgeführt werden (Tabelle 1 im Anhang).
Allgemein waren die Selektions! inien den Kontroll inien in
allen wirtschaftlichen Merkmalen überlegen. Der Unterschied
im Selektionskriterium war in der dritten Generation bereits
signifikant und konnte durch den Anstieg der produzierten
Eimasse , das Produkt aus Eigewicht und Eizahl, verbunden mit

48
einer geringeren Zunahme des Futterverbrauches und einem
tieferen Körpergewicht grossenteils erklärt werden.
Tabelle 9
Merkmal
Generation
Linie/
Periode
Linie 1
21 - 40
41 - 60
Linie 2
21 - 40
41 - 60
Durchschni t1
21 - 40
41 - 60
Selektionserfolge für das Selektionskriterium und korrelierte Selektionserfolge für verschiedene Merkmale
Selektions- Eimasse Futter- Futterver-kriterium verbrauch wertung
(kg) (kg)
F2 F3 F2 F3 F2 F3 F2 F3
. 39 . 90 . 07 .23 - .15 -.01 -.05 -.07
. 17 .78 .02 .38 - . 12 .08 -.01 - . 07
.34 .72 . 06 . 21 - .14 . 13 -.03 -.03
.34 .88 .06 . 29 - . 19 . 37 -.06 - .08
.36 .79 . 06 . 22 - .15 .07 -.04 -.05
. 25 .83 .04 .34 - . 16 .22 -.03 -.07

4 9
Die Graphiken 1 und 2 veranschaulichen die höhere Eiproduk
t ion der Selektionslinien , verbunden mit einer besseren Per
sistenz . Eine Arbeit von Von dem Hagen (1986) , könnte dazu
mögliche Hinweise liefern. Die genannte Untersuchung weist
auf zusammenhänge zwischen der Leistungsminderung bei Lege
hennen und Leukosevirusinfektionen hin. Aviäre Leukoseviren
vermögen die Legeleistung von Hennen bis ~ur 45. resp. 71.
Woche um 23 bis 30 Eier zu senken. Die Reaktion auf eine In
fektion ist aber, wie die Infektionsempfänglichkeit, nicht
bei allen Individuen identisch. Möglicherweise wurden in
diesem Experiment durch die Selektion auf Eierlös minus Fut
terkosten indirekt eher resistentere und/oder virusfreie
Tiere ausgelesen. Deshalb vergrössert sich eventuell die
Differenz in der Legeleistung zwischen der Kontrollinie und
den Selektionslinien von Jahr zu Jahr. Da die Versuchstiere
jedoch nicht auf Leukose untersucht wurden, liefert diese
Arbeit nur eine Hypothese zur Erklärung der tieferen Eipro
duktion der Kontrollinie, aber keinen Beweis für ihre Unter
legenheit.
Die Selektionslinien der zweiten Generation benötigten trotz
der höheren Eimasseprodukt ion weniger Futter als die Kon
troll inie. Unter Berücksichtigung der Tatsache, dass der
Futterverbrauch von der Eimasse und dem Körpergewicht ab
hängt, konnte der geringere Futterverbrauch der Selektions
linien teilweise durch ihr signifikant tieferes Körperge
wicht (Tabelle 2 im Anhang) erklärt werden. In der dritten
Generation lag der Futterverbrauch der Selektionslinien über
demjenigen der Kontrollinie und könnte mit der hohen Eimas
seproduktion erklärt werden, obwohl das Körpergewicht der
Selektionslinien noch leicht unter demjenigen der Kontrol
linie lag.

Graphik 1 Legeleistung
"' 0
"' " 1
,lp
,-,~-,-,
Graphik 2 Leg e l eis tun g
50
5 0
F2 Generation der
100
der
100
150
1984 F2
19-l'l SELEKTION 1
~ SELEKTION 2
tr---6 KONTROLLE
200
LEGET AG
Ge ner a t io n F3
1985 ° l9-l'l SELEKTION
(9---(9 SEJ. EKTI ON 2
tr---6 KONTROLLE
---,-,,-250 JOO

51
4.1.3. Genetische Parameter in der Selektionsperiode
4.1.3.1. Genetische Korrelationen und Heritabilitäten in der
Selektionsperiode
Die Korrelationen zwischen zwei Merkmalen sind dann von Be
deutung, wenn durch die Selektion auf ein Merkmal Verände
rungen in einem anderen Merkmal herbeigeführt werden. Diese
korrelierten Selektionserfolge in wirtschaftlich wichtigen
Merkmalen wurden deshalb auch untersucht.
Die genetischen Korrelationen mussten auf Grund der unter
schiedlichen Populationsstrukturen mit den Modellen I - III
berechnet werden. Die Schätzungen beruhen auf den Väterkom
ponenten. Die Werte der Tabelle 10 sind ein mit der Varianz
der Schätzung gewichteter Durchschnitt der drei Generatio
nen.
Die genetischen Korrelationen zwischen dem Selektionskrite
rium und den übrigen untersuchten Merkmalen bestätigen eini
ge Beziehungen, welche in den korrelierten Selektionserfol
gen zu beobachten waren. Die zusammenhänge zwischen Selek
tionskri terium und Eimasse sind hoch positiv und zwischen
Selektionskriterium und Futterverwertung hoch negativ. Sie
sind unbedeutend zwischen dem Selektionskriterium und dem
Futterverbrauch, resp. dem Körpergewicht bei 20 Wochen.

5 2
Tabelle 10 genetische Korre2ationen und Standardfehler (in Klammer) sowie h -Werte auf der Diagonalen für die wirtschaftlich wichtigen Merkmale zwischen dsr 21. - 40. Woche für das Selektionskriterium (SK) , die Eimasse, (EM), die Eizahl (EZ}, das Eigewicht (EW}, den Futterverbrauch (FV), die Futterverwertung (FVW} und das Körpergewicht bei 20 Wochen (KG20}.
Merkmal SK EM EZ EW FV FVW KG20
SK .23 .89 .35 .74 .09 -.92 -.09
(. 06) (. 03} ( . 11 ) (. 07) { .12) (. 19) ( . 12)
EM .24 .47 .64 .57 -.57 . 19
(. 05} ( . 1 0} (. 08} (. 09) (. 20) ( .12)
EZ . 23 -. 56 .17 -.43 .08
( . 05) ( .10) ( . 13) (. 20) ( .13)
EW 1 . 0 .35 - . 16 .14
(. 12} ( .10} (. 15) (. 10)
FV . 40 .57 .64
( . 07) ( . 14) (. 07)
FVW . 09 . 42
(. 03) (. 15)
KG20 .58
(. 09}
Die genetische Korrelation zwischen dem Selektionskriterium
und dem Eigewicht ( 0. 74) ist bedeutend höher als diejenige
zwischen dem Selektionskriterium und der Eizahl (0.35). Zwi
schen dem Selektionskriterium und dem Eigewicht war sie so

53
eng, dass durch die Se).ektion die Eimasse trotz der nega
tiven Korrelation zwischen Eizahl und Eigewicht grösser wur
de (Tabe! le 9) . Die enge Beziehungen zwischen dem Selek
tionskr i terium und der Futterverwertung muss auf die hohe
Korrelation zwischen Selektionskriterium und Eimasse zurück
geführt werden, da die Korrelation Selektionskriterium
Futterverbrauch sehr tief ist. Dies lässt somit den Schluss
zu, dass auf Grund des Selektionskriteriums effizientere
Hennen ausgelesen wurden .
Der Futterverbrauch wies die höchste Korrelation mit dem
Körpergewicht auf (.64). Diese lag im Bereiche der von
Pirchner (1985) geschätzten Korrelation (rg = .72) und be
stätigte die enge Beziehung zwischen Futterverbrauch und
Körpergewicht. Demzufolge benötigten schwerere Hennen mehr
Futter, produzierten jedoch mehr Eimasse und wiesen dement
sprechend eine bessere Futterverwertung auf, was im Selek
tionserfolg zum Ausdruck kam. Die Korrelation zwischen Fut-
terverbrauch und Eimasse (rg = .57) war leicht
von Pirchner (1985) (rg = .3) oder von Damme,
.35 - . 46) publizierten Werte . Die positiven
höher als die
(1984), (r = g Korrelationen
zwischen dem Körpergewicht und der Futterverwertung, der Ei
masse und dem Eigewicht deuten darauf hin , dass wegen der
positiven Korrelationen zwischen dem Selektionskriterium und
der produzierten Eimasse, das Körpergewicht langfristig se
lektionsbedingt ansteigen wird. Da die Selektion jedoch die
Eimasseproduktion stärker beeinflusst als das Körpergewicht,
wird wohl der Futterverbrauch ansteigen, die Futterverwer
tung wird sich aber dennoch verbessern und demzufolge zu
einer weiteren Verbesserung des Selektionskriteriums führen .
Die geschätzten Heri tabil i täten sind in Tabelle ·10 auf der·
Diagonalen aufgeführt. Sie waren, mit Ausnahme derjenigen
für die Futterverwertung, relativ hoch und lagen im Bereiche
der von Damme (1984) publizierten Werte. Die von verschied
enen anderen Autoren geschätzten Heritabilitäten sind in
Tabelle 11 zusammengestellt.

5 4
Tabelle 11 Heritabilitäten einiger Merkmale nach
verschiedenen Autoren
Merkmal eigener
Versuch
Eimasse .24
Eigewicht 1 . 0
Futterverbrauch .4
Futterverwertung .09
Körpergewicht . 58
Hagger
( 1978)
.30 -. 70
Pirchner
(1985)
.32
.58
. 1 2
.17
. 80
Damme
(1984)
.34
.73
. 51
.23
Nordskog
( 1972)
. 20
Einzig für das Mer k mal Eigewicht fielen die Schätzwerte weit
höher aus als bei den i n Tabelle 11 zitierten Autoren.
Der Schätzwert für die Heritabilität des Selektionskriteri
ums lag im Bereich der We r te für die Eimasse und Eizahl. Auf
Grund der engen positiven Korrelati on mit der Eimasse darf
in der folgenden Generati on mit einer weiteren Verbesserung
des Selektionskriteriums gerechnet werden.
4 . 1.3 .2. Genetische Korrelationen innerhalb der zweiten
Periode (41. - 60. Woche) sowie zwischen den beiden
Pe r ioden (21. - 40 . Woche und 41 . - 60 . Woche)
und die Heritabilitäten.
In Tabelle 12 sind die genetischen Korrelationen zwischen
verschiedenen Merkmalen innerhalb der zweiten Periode sowie
zwisc hen der ersten und der zweiten Periode zusammenge
ste llt. Die Heritabili t äten der verschiedenen Merkmale sind
für die zweite Periode in Klammer aufgeführt.

55
Tabelle 12 Genetische Korrelationen (r ) zwischen der Periode 21. - 40. Woche (l)gund der Periode 41.-60. Woche (2), sowie die Korrelationen und die Heritabilitäten (in Klammer) in der zweiten Periode für die Merkmale: Selektionskriterium (SK), "analoge Differenz" (D), Eimasse (EM), Eizahl (EZ), Eigewicht (EW), Futterverbrauch (FV), Futterverwertung (FVW) und Körpergewicht bei 20 Wochen (KG20) und bei 60 Wochen (KG60) .
Merkmal
Periode
SK
D
EM
EZ
EW
FV
FVW
1
2
2
2
2
2
2
KG20
KG60 2
D EM EZ
2 2 2
.89 .80 . 65
(.27) .94 .81
.83 .88 .67
(. 32) . 78
EW
2
.68
.49
. 60
. 60
. 31 .25 .79 - .64
(.22) -.28
FV
2
.11
.19
.48
.64
.05
.41
FVW
2
-.76
-.85
- .45
-.59
- .20
-.52
.56 .68 -.06 .95 .40 -.30
( .90) .28 -.24
.02 .39 . 17 .30 .90 .52
-.74 -.49 -.43 -.18
-.16 -.06 -.05 . 14
( .55) .57
.52
. 53
1. 00
( .18)
.43
KG60
2
- . 19
-.30
.18
- . 01
- . 01
- . 15
. 18
. 16
.80
.6 7
.63
.65
.86
(. 60)
Die Korrelationen sind recht hoch und liegen für die zweite
Periode im Bereiche derjenigen der ersten Periode . In der
ersten Periode war das Selektionskriterium am höchsten mit

56
der Futterverwertung korreliert (-.92) , gefolgt von der Ei
masse (.89) , vom Eigewicht ( . 74) und von der Eizahl ( . 35).
In der zweiten Periode hingegen wurden für die "analoge Dif
ferenz" die höchsten Korrelationen mit der Eimasse ermittelt
(. 94) '· gefolgt von der Futterverwertung (-. 85), der Ei zahl
(. 81) und dem Eigewicht (. 49). Die Rangfolge der Korrela-
tionen zwischen
wicht hat sich
umgekehrt .
Selektionskriterium und Eizahl
in der zweiten Periode ( 41 .
resp. Eige-
60. Woche)
Die "analoge Differenz'' wurde stärker von der Eizahl als vom
Eigewicht beeinflusst . Dies bringt zum Ausdruck, dass Hennen
welche zu Beginn der Selekti onsperiode einen hohen Anteil
kleiner Eier legten , bei der Selektion nicht berücksichtigt
wurden . Somit wäre es interessant Hennen mit flacheren Eige
wichtskurven zu züchten deren Niveau zu Beginn der Legeper
iode jedoch höher wäre. Bei ziemlich konstantem Eigewicht in
der zweiten Periode verbes s er te folglich vorwiegend die Ei
zahl die " anal oge Differenz". Das gleiche Phänomen kommt
auch i n den Korrelati o nen zwischen der Eimasse und der Ei
zahl in d er ersten Periode (.47) gegenüber der zweiten Per
iode (.78) sowie zwischen der Eizahl und dem Eigewicht der
ersten Periode (- . 56) gegenüber (-.28) der zweiten Periode
zum Ausdruck.
Vergleicht ma n noch die Korrelation zwischen Eizahl und Fut
terverbrauch in der ersten Periode ( . 17) mit derjenigen in
der zwe iten Periode ( .41) so wird deutlich, dass der Futter
verbrauch in der zweiten Periode enger mit der Eizahl korre
liert ist als in der ersten Periode. Die Korrelationen des
Selektionskriteriums mit den übrigen Merkmalen in der zwei
ten Periode sind relativ hoch ausser für die Merkmale Fut
terverbrauc h und Körpergewicht. Sie liegen unter denjenigen
der "analogen Differenz" ausser für das Merkmal Eigewicht,
was wiederum bedeutet, dass das Selektionskriterium vom
Eigewicht, und die "analoge Differenz" von der Eizahl beein
flusst wird . Die Heritabilitäten der zweiten Periode weichen
kaum von denjenigen der ersten Periode ab.

5 7
Die Korrelationen zwischen gleichen Merkmalen in beiden Per
ioden waren positiv. Sie waren hoch bei den Merkmalen Ei
masse, Eizahl, Eigewicht und Futterverwertung, mittel für
das Eigewicht und tief für das Körpergewicht und den
Futterverbrauch. Eier von Hennen mit hoher Legeleistung wa
ren in der ersten Periode leichter und die Legeleistung der
ersten Periode schien sich negativ auf die Eigewichte in der
zweiten Periode auszuwirken.
4.1.4. Einfluss der Befiederung
Bis anhin wurden die wirtschaftlichen Merkmale und ihre Ein
flüsse auf das Selektionskriterium diskutiert. Als möglichen
weiteren Einfluss auf den Futterkonsum und somit auf das
Selektionskriterium wurde in der Literatur die Befiederung
erwähnt (Tullet, 1980; Tauson, 1980; Hughes, 1980; Charles,
1976). Mit Hilfe des von Charles (1976) publizierten und
angepassten Punktierschemas ( 1 gut, 4 = schlec ht) wurde
die Befiederung für die Linien in beiden Perioden und beiden
Generationen bestimmt.
Tabelle 13 Unterschiede in der Befiederung zwischen den Selektionslinien und der Kontrollinie in beiden Perioden der Generationen F2 und F3.
1. Periode 2. Periode
Generation F2 F3 F2 F3
Linie L1 L2 L1 L2 L1 L2 L1 L2
.05 .03 .23 . 16 -.01* -.06* .22 . 11
Aus Tabelle 13 wird ersichtlich, dass die Selektionslinien
in der ersten Periode weniger gut befiedert waren als die

58
entsprechenden Kontroll inien und dass sich die Bef iederung
mit zunehmendem Selektionszyklus leicht verschlechterte. In
der zweiten Periode der zweiten Generation, wiesen die Tiere
der Selektionslinien eine signifikant bessere Befiederung
auf als die Tiere der Kontrollinie. In der F3 wiederum waren
die selektierten Tiere schlechter befiedert als die unselek
tierten.
Die Selektion scheint eine Verschlechterung der Befiederung
zu bewirken . Die Tiere waren dadurch schlechter isoliert,
ihr Wärmeverlust wurde grösser und sie frassen möglicher
weise dementsprechend mehr. Tabelle 14 zeigt die Abhängig
keit des Futterkonsumes von der Befiederungsklasse , korri
giert auf Kör pergewicht, Gewichtsdifferenz, Eimasse und Al
ter bei Legebeginn , geschätzt aus Modell VII. Die Klasse 1
wurde gleich Nul 1 gesetzt und die übrigen Klassen wurden
dann mit dieser verglichen.
Tabelle 14 Abweichungen im Futterverbrauch (kg) zwischen
den verschiedenen Befiederungsklasssen
Generation
Periode
Klasse
2
3
4
0.000
. 22 3
. 306
. 424
F2
0.000
.252
.239
. 371
F 3
0.000
. 135
. 188
.513
F 2
2
0.000
.398
.669
.997
F3
2
0 . 000
. 036
.233
.673
Hieraus wird ersichtlich, dass ein relativ grosser Unter
schied zwischen den verschiedenen Befiederungsklassen in Be
zug auf den Futterkonsum besteht . Es zeigt sich , dass die
schlechter befiederten Hennen generell mehr fressen . Am

5 9
grössten ist der Unterschied in der zweiten Periode der
zweiten Generation , wo die am schlechtesten befiederten Hen
nen beinahe 1 kg mehr fressen als die gut befiederten. Weil
in der zweiten Periode der zweiten Generation die Selek
tionslin i en signifikant besser befiedert waren als die
Kontrollinie, war ihr Futterverbrauch auch dementsprechend
geringer und die Futterverwertung umso besser.
Tullet (1980) hatte in se i ner Arbeit mit teilentfiederten
Tieren nachgewiesen , dass deren Futterkonsum erhöht war.
Ihre Legeleistung war jedoch grösser und ihr Körpergewicht
geringer . Zu ähnlichen Resultaten kam Hughes (1980) indem er
feststellte, dass Hennen mit beschädigtem Gefieder mehr Eier
legten , was jedoch nicht dem Mehrkonsum entsprach . Die in
der vorliegenden Arbeit untersuchten Selektionslinien
zeichneten sich sowohl deutlich durch eine schlechtere
Befiederung wie auch durch höhere Legeraten, Eigewichte und
eine grössere Eimasse sowie tiefere Körpergewichte aus und
bestätigen die Ergebnisse anderer Untersuchungen. Da sowohl
die Befiederung, wie auch die Eiproduktion hormonell
gesteuert werden, haben schlechtere Legerinnen möglicher
weise eine tiefere Ovarialtätigkeit, können somit Nackt
stel len besser befiedern und werden dementsprechend besser
klassiert . Auch Conson (1986) kam zum Schluss, dass eine
hoch signifikante positive Beziehung zwischen Gefieder
beschaffenheit und Futterkonsum besteht. In Bezug auf diesen
Punkt wichen die Resultate der hier untersuchten Selektions
linien nicht von anderen Resultaten ab. Lediglich Gonyou
( 1982) fand, dass eine Entfernung der Federn an Rücken,
Nacken und Hals den Futterkonsum um 5 - 6 t erhöhte, dass
sich jedoch weder das Körpergewicht noch die Eimasse
produktion und die Futterverwertung dadurch veränderten. Es
handelte sich in der genannten Arbeit nicht um eine gene
tische Veränderung der Befiederung, sondern um einen Umwelt
einfluss (Entfiederung). Diese Folgerungen sind deshalb mit
den hier aufgeführten Resultaten nicht vergleichbar.

50
4.1.5. Einfluss der Körpertemperatur
In der dritten Generation wurden im Alter von 16 , resp . 30
und 50 Wochen die Körpertemperaturen bei je 236 ausgewählten
Tieren gemessen. Die erhobenen Daten sind in Tabelle 15
aufgeführt.
Tabelle 15 Körpertemperaturen gemessen im Alter von 16, 30
und 50 Wochen .
Alter Anzahl Temperatur (OC) L1 L2 K
Wochen Hennen 0
16 236 41 . 81 41. 86 41 . 81 41 . 76
30 236 41 .54 41. 53 41 . 55 41 .54
50 228 41. 36 41. 35 41 . 38 41 . 32
Die Hennen der Selektionslinien haben eine nicht signifikant
höhere Körpertemperatur als die Kontrollinie . Ein signifi
kanter Einfluss des Alters auf die Körpertemperatur konnte
jedoch festgestellt werden . Je älter eine Henne , desto tie
fer ih r e Körpertempera t ur. Diese Resultate können nur ein
zusätzlicher Hinweis für den erhöhten Futterverbrauch sein .
Die geringe Anzahl Tiere erlaubt es aber nicht aus diesen
Angaben konkrete Schlüsse zu ziehen.
4.1 .6. Schlussfolgerungen
Nach zwei, auf Eier lös minus Futterkosten selektierten Ge-
nerationen, konnte das Selektionskriterium signifikant
( p ~ 0. 05) verbessert werden. Diese Verbesserung war auf
eine Überlegenheit der Selektionslinien gegenüber der Kon
troll inie in den Merkmalen Legeleistung, Eigrösse, Eimasse
und Futterverwertung bei tieferem Körpergewicht zurückzu
führen. Die Futterverwertung der Gesamtpopulation veränderte
sich in der Periode von der 21. bis zur 40. Woche von der

51
ersten zur dritten Generation nicht. In der zweiten Periode,
41. bis 60 . Woche, hingegen verbesserte sich die Futterver
wertung von der ersten zur dritten Generation von 2.52 auf
2.40. Die Selektionslinien der dritten Generation hatten in
der ersten Periode eine um 2 t bessere und in der zweiten
Periode eine um 3 t bessere Futterverwertung als die Tiere
der Kontrollinie.
Die genetischen Korrelationen zeigten, dass sich bei anhal
tender Selektion auf Eierlös minus Futterkosten die korre
lierten Merkmale weiterhin verbessern werden und in den
nächsten Generationen mit zunehmenden Eigewichten, Legelei
stungen und grösserer Eimasse, verbunden mit einem anstei
genden Körpergewicht gerechnet werden kann. Ab welchem Eige
wicht die Folgen für die Brutergebnisse negativ sein werden,
ist noch nicht abgeklärt. Auf Grund von Untersuchungen von
Hilfiker (1970) und Scott (1941) muss aber beim Erreichen
des Optimums mit sinkenden Brutergebnissen gerechnet werden.
Die hier vorliegenden Resultate zeigen, dass eine Selektion
auf Eierlös minus Futterkosten zu einer besseren Futterver
wertung führt. Sie bestätigen nicht die Ergebnisse eines Ex
perimentes von Pirchner (1985), wo die Selektion von Kreu
zungstieren auf eine bessere Futterverwertunggsrate das Kör
pergewicht und die Eigewichte reduzierte und nur die Lege
rate leicht verbesserte. In der gleichen Untersuchung
schätzte Pirchner jedoch wirtschaftlich günstige Korrelatio
nen zwischen Futterverwertung, Eimasse und Eigewicht.
Ähnliche Korrelationen wurden auch in dieser Arbeit ge
schätzt. Im Experiment von Heil und Pirchner (1979) stiegen
bei direkter Selektion auf Eiproduktion das Eigewicht, die
Eizahl, das Körpergewicht, wie auch der tägliche Futterver
brauch. Der korrelierte Selektionserfolg auf die Futterver
wertung lag tiefer als bei direkter Selektion auf Futter
verwertung welche zu tieferen Ei- und Körpergewichten und zu
einer höheren Legeleistung führt. Pauw ( 1986) bekräftigte

5 2
die Annahmen, nach welchen eine Selektion auf Wirtschaft
lichkeit die Produktion von Eimasse erhöht und das Körper
gewicht unter Einbezug des Futterverbrauches abnimmt.
Das Selekt ionskr i ter i um Ei erlös minus Futterkosten scheint
in diesem Versuch demzufolge gegenüber anderen erwähnten
Selektionskriterien die günstigsten Auswirkungen auf Eimasse
und Futterverwertung zu haben, indem einerseits die Eimasse
produktion ansteigt , dies auf Grund der besseren Legelei
stung und der höheren Eigewichte und andererseits das Kör
pergewicht gesunken ist, was zu einer besseren Futterver
wertung führt.
Aus der Arbeit von Heil und Pirchner (1979) konnte entnommen
werden, dass die genetische Verbesserung der Futterverwer
tung durch Selektion nur teilweise durch die Leistungsver
besserung erklärt werden kann. Durch die Berücksichtigung
der Merkmale Eiproduktion und Futterverbrauch alleine werden
womöglich andere wichtige, nicht zu vernachlässigende Merk
male wie z.B. die Eizusammensetzung , nicht erfasst.
Korrelationen zwischen dem Selektionskriterium und diesen
Merkmalen könnten zu nicht vorhergesehenen Auswirkungen
führen.
Weil durch die Selektion das Eigewicht anstieg und sich die
Futterverwertung gleichzeitig verbesserte, war es von Inter
esse, die interne Eizusammensetzung der Eier zu analysieren
um weitere Hinweise zur Erklärung des Selektionserfolges im
Selektionskri terium zu erhalten. Die Frage nach der Verän
derung der Eizusammensetzung wurde anhand von Untersuchungen
an drei Generationen und zu verschiedenen Zeitpunkten, im
Alter von 30 Wochen und im Alter von 50 Wochen, abgeklärt.

53
4.2. Eizusammensetzung
In der Literatur gibt es bereits eine beachtliche Anzahl von
Hinweisen bezüglich des Eigewichtes, der Eizusammensetzung
und der Korrelationen zwischen diesen beiden Eigenschaften
(Sainz et al. , 1983; Ambrosen et al., 1981; Hill, 1966;
Marion et al., 1965; Chung et al., 1965; Cunningham et al.,
1960 ; ).
Die Variation des Eigewichtes wird auf die Linien sowie auf
die einzelne Henne zurückgeführt . Den grössten Einfluss auf
die Eigrösse von Legehennen hat jedoch deren Alter. Alle
Autoren sind sich einig , dass die Eigrösse die Eizusammen
setzung beeinflusst. Durch Selektion bedingte Veränderungen
der Eigrösse lassen somit erwarten, dass sich auch die Eizu
sammensetzung verändern wird . Obwohl das Gesamtei ungefähr
75 t Wasser enthält, ist es eine hochwertige Proteinquelle
und wird oft für Vergleiche mit anderen Nahrungsmitteln
herbeigezogen. Es war deshalb von Interesse, d i e Auswir
kungen der Selektion auf die Eigrösse und die Eizusammen
setzung zu erfassen.
4.2.1. Phänotypische Parameter
4 . 2.1.1. Mittelwerte und Standardabweichungen der Merkmale
der Eizusammensetzung der drei Generationen bei 30
respektiv bei 50 Wochen.
Die Ergebnisse bezüglich der Eizusammensetzung bas i eren auf
Untersuchungen an drei Generationen, bestehend aus der
Bas i spopulation und de~ zwei selektierten Generationen mit
je zwei Selektionslinien und einer Kontrollinie. In den drei
Generationen wurden aus der Gesamtpopulation von je 25 t bis
30 t zufällig ausgelesenen Hennen die Eier im Alter von 30,
resp. 50 Wochen untersucht .

54
Den Berechnungen liegt Modell VIII zu Grunde. Die Analysen
umfassen alle Stichprobentiere unabhängig davon ob sie in
beiden Erhebungen, 30 und 50 Wochen, innerhalb einer Genera
tion erfasst werden konnten oder nicht. Bei denjenigen Tie
ren, welche nicht in beiden Auswertungen vertreten waren,
handelte es sich um Hennen , die in der ersten oder zweiten
Stichprobe an 5 aufeinanderfolgenden Sammeltagen nicht ein
Minimum von drei ganzen Eiern gelegt hatten , die mauserten
oder abgegangen waren.
Um die drei Generationen zu charakterisieren, sind die Werte
für die verschiedenen Merkmale als Durchschnitt der Selek
tions- und der Kontrollinien innerhalb der drei Generationen
einander in Tabelle 16 gegenüber gestellt .
Auf Grund der Eigewichte lassen sich diese Untersuchungen
mit solchen aus der Literatur vergleichen (Sainz et al.,
1983: Ambrosen et al. , 1981 ; Burkhardt , 1971; Chung et al. ,
1965 ; Marion et al. , 1965 ; Skala , 1962). Im Alter von 30
Wochen wiegt ein Ei durchschnittlich 59.5 g gegenüber 66 . 9 g
im Alter von 50 Wochen. Im Alter von 30 Wochen setzen sich
die in den drei Generationen untersuchten Eier im Durch
schnitt aus 38.3 g Eiklar , resp. 64.4t, und 14.9 g Eidotter,
resp. 25.3 t zusammen. Der Trockensubstanzgehalt beträgt
23.1 t , der Proteingehalt 52.4 t und der Energiegehalt
29.1 MJ / kg Trockensubstanz . Im Alter von 50 Wochen legen die
Hennen durc hschnittlich schwerere Eier (66 . 9 g) mit mehr
Eiklar (41.5 g) und mehr Eidotter (18.8 g). Der Trockensub
stanz gehalt und der Energiegehalt sind bei 50 Wochen höher
( 24 . 2 % resp. 29.8 MJ / kg), und der Proteingehalt ist tiefer
(49.4 %) . In der dritten Generation ist das Eigewicht bei 30
und bei 50 Wochen höher als in der zweiten Generation. Der
Eiklaranteil nimmt von der zweiten zur dritten Generation
ebenfalls zu , währenddessen der Eidotteranteil bei 30 und
bei 50 Wochen von der zweiten zur dritten Generation sinkt.

55
Tabelle 16 Mittelwerte (:X) und Standardabweichungen (sx) der Eibestandteile der drei untersuchten Generationen im Alter von 30 und von 50 Wochen.
Generation F1 F2 F3
X: s X
x sx x Merkmal
Eigewicht g 30 61 . 0 3.7 58.6 4.0 58.9
50 65.6 5.2 68.2
Eiklargewicht g 30 39.2 2 . 9 37.4 3 . 2 38.3
50 40.5 3.9 42 . 5
Eiklaranteil t 30 64 . 3 1 . 5 63.8 1. 9 65.0
50 61. 7 2.3 62.2
Eidottergewicht g 30 15.4 1 . 0 15. 1 1. 2 14.4
50 18.6 1.8 18.9
Eidotteranteil t 30 25 . 3 1 . 4 25 . 9 1. 8 24.6
50 28.3 2.2 27.7
Trockensubstanz- 30 23.3 0.9 23.4 0.9 22.7
gehalt {TS) t 50 24.3 1 . 1 24 .1
Proteingehalt 30 52.4 1.5 51 . 7 1. 8 53 . 0
in der TS t 50 49.0 2 . 0 49.7
Energiegehalt 30 29.2 0.3 29.2 0 . 4 28.9
in der TS MJ/kg 50 29.8 0.4 29.7
Der Trockensubstanzgehalt sinkt von der zweiten zur dritten
Generation in den Untersuchungen bei 30 und bei 50 Wochen ,
innerhalb Generation steigt er mit zunehmendem Alter jedoch
an .

55
Der Proteingehalt in der Trockensubstanz nimmt von der zwei
ten zur dritten Generation sowohl bei 30 wie bei 50 Wochen
zu, und sinkt innerhalb der Generation mit zunehmendem Al
ter. Dies ist verbunden mit einer Abnahme des Energiege
haltes von Generation F2 zu F3 bei 30 und bei 50 Wochen,
währenddessen innerhalb einer Generation der Energiegehalt
mit zunehmendem Alter ansteigt. Diese Veränderungen der
durchschnittlichen Werte deuten auf einen Einfluss des Al
ters sowie der Selektion hin.
4.2.1 .2. Wiederholbarkeiten
In der ersten untersuchten Generation wurden vier und ab der
zweiten Generation drei Eier pro Henne an möglichst aufein
anderfolgenden Tagen gesammelt , gewogen und in ihre Einzel
komponenten zerlegt. Dies erlaubte es , die Wiederholbarkei
ten für die verschiedenen Merkmale in allen drei Generatio
nen und soweit vorhanden, in beiden Perioden (30 und 50 Wo
chen) zu berechnen. In Tabelle 17 sind die Ergebnisse zusam
mengefasst.
Tabelle 17 Wiederholbarkeiten verschiedener Merkmale de r Eizusammensetzung im Alter von 30 und 50 Wochen für die drei untersuchten Generationen
Generation F1 F2 F3
Alter 30 30 50 30 50
Merkmal
Eigewicht g 0.789 0.834 0.829 0.845 0.826
Eiklar g 0.789 0.835 0.807 0.851 0.797
Eidotter g 0.739 0.825 0.852 0.842 0.843
Eischale g 0.743 0.710 0.700 0.793 0.687

57
Diese Wiederholbarkeiten sind relativ hoch und liegen leicht
höher als die in der Literat.ur gefundenen Angaben ( Ambrosen
et al, 1981; Rodda et al, 1977; Hill et al, 1966; Scheinberg
et al , 1953;). Von Generation zu Generation nimmt die bei 30
Wochen errechnete Wiederholbarkeit zu. Dies mag auf die
grössere Obung bei der praktischen Durchführung zurückzu
führen sein. Der leicht sinkende Trend von der "30 Wochen
Untersuchung" zur "50 Wochen-Untersuchung" kann mit der un
regelmässigeren Legetätigkeit der Hennen bei 50 Wochen er
klärt werden. Mit 30 Wochen sind die Hennen in ihrer besten
Produktionsphase und legen ausgeglichenere Eier als bei 50
Wochen, wo einige Tiere schon nicht mehr jeden Tag ein Ei
produzieren. Diese Wiederholbarkeiten sowie Angaben aus der
Literatur (Kirchgessner
Nagai et al , 1969; Hill
Marion et al, 1965,1964;
Scheinberg et al, 1953)
et al, 1980; Tawfik et al, 1976;
et al, 1966; Chung et al, 1965;
Skala et al, 1962; May et al, 1960;
erlauben es, die weiteren Be rech-
nungen auf Grund der Durchschnittswerte von vier, resp. drei
gesammelten Eier durchzuführen.
4.2.1.3. Phänotypische Korrelationen der Eizusammensetzung
in den drei Generationen bei 30 und bei 50 Wochen
In Tabelle 18 sind die phänotypischen Korrelationen der Ei
zusammensetzung in den drei Generationen bei 30 und bei 50
Wochen einzeln aufgeführt.
In Tabelle 18 lassen sich deutlich zwei Gruppen unterschei
den. Positi'.r korreliert sind einerseits die Merkmale Eige
wicht (EG g), Eiklaranteil (EK t) und Proteingehalt (P t)
und andererseits die Merkmale Eidotteranteil (ED%), Ener
giegehalt (E MJ/kg) und Trockensubstanzgehalt (TS t). Ener
giegehalt und Proteingehalt beziehen sich immer auf die
Trockensubstanz.
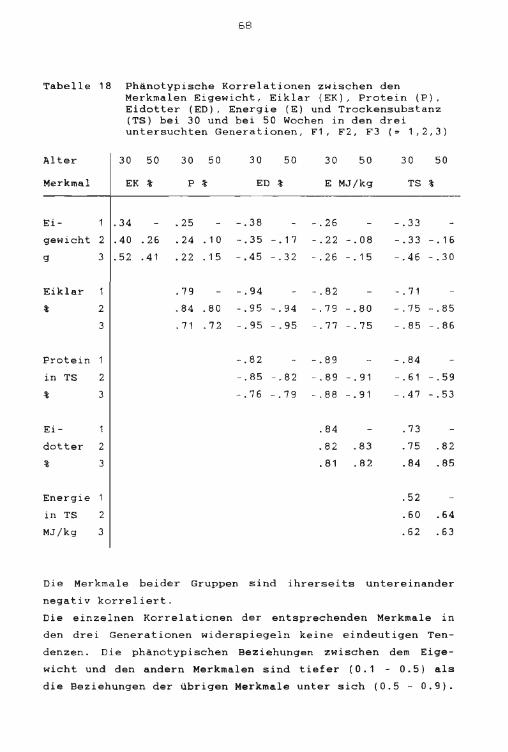
68
Tabelle 18 Phänotypische Korrelationen zwischen den Merkmalen Eigewicht, Eiklar (EK) , Protein (P), Eidotter (ED) , Energie (E) und Trockensubstanz {TS) bei 30 und bei 50 Wochen in den drei untersuchten Generationen , F1, F 2, F3 ( = 1 '2 ' 3)
Alter 30 50 30 50 30 50 30 50 30 50
Merkmal EK % p % ED % E MJ/kg TS %
Ei- . 34 .25 -. 38 - .26 -.33
gewicht 2 .40 .26 .24 .1 0 -.35 - . 17 - .2 2 - .08 - .33 - .16
g 3 .5 2 . 41 .22 .15 -.45 -.32 - .26 - . 15 - .46 - . 30
Eiklar . 7 9 -.94 - .82 - .71
% 2 .84 .80 - .95 -.94 -. 79 -.80 -.75 -.85
3 . 71 . 72 -. 95 - .95 - .77 - .75 -.85 -. 86
Protein -.8 2 -.89 -.84
in TS 2 - .85 -. 82 - .8 9 - . 91 - . 61 - . 59
% 3 -.76 -. 7 9 -.88 - . 91 - .47 - . 53
Ei- .84 .73
dotter 2 .82 . 83 .75 .82
% 3 . 81 . 82 .84 . 85
Energie .52
in TS 2 .60 .64
MJ / kg 3 .62 .63
Die Merkmale beider Gruppen sind ihrerseits untereinander
negativ korreliert.
Die einzelnen Korrelationen der entsprechenden Merkmale in
den drei Generationen widerspiegeln keine eindeutigen Ten
denzen. Die phänotypischen Beziehungen zwischen dem Eige
wicht und den andern Merkmalen sind tiefer (0.1 - 0.5) als
die Beziehungen der übrigen Merkmale unter sich (0.5 - 0.9).

69
Es scheint, dass die Korrelationen zwischen Eigewicht und
den übrigen Merkmalen von der F1 zur F3 enger werden. Die
aus den drei Korrelationen berechneten Mittelwerte liegen
für die verschiedenen Merkmale im Bereich der von anderen
Autoren angegebenen Werte (Tawfik, 1981; Hilfiker, 1970;
Jaffe, 1965).
Ein Vergleich der phänotypischen Korrelationen zwischen dem
Eigewicht und den anderen Merkmalen bei 30 und bei 50 Wochen
deutet auf Werte hin, die mit zunehmendem Alter sinken. Die
Beziehungen zwischen den übrigen Merkmalen verändern sich
kaum.
Aus den beobachteten Korrelationen zwischen dem Eigewicht
und den anderen Merkmalen kann geschlossen werden, dass die
Eizusammensetzung mit zunehmendem Alter stärker vom Alter
abhängt als von der Eigrösse. Je jünger eine Henne, desto
enger die Beziehungen zwischen dem Eigewicht und den übrigen
Merkmalen. Mehner (1983) weist auf einen höheren Wasserge
halt des Eiklars mit steigendem Eigewicht hin und begründet
ihn damit, dass bei höherem Gesamteigewicht die Leistung des
Ovars und der Uterusschleimhautdrüsen verstärkt sind und
eine Verzögerung der Kalzifikation in den ersten Stunden der
Sehwellphase, wo das Ei noch stark Flüssigkeit aufnehmen
kann, zu einer Erhöhung des Gesamteigewichtes führt. Vier
bis f\inf Stunden nach der Ovulation kommt das Ei in den
Uterus wo es noch 20 Stunden verbleibt, bis sich die Schale
um die Membranen gebildet hat. Der Grund für eine solche
Verzögerung kann in der relativ grossen Oberfläche eines
grossdotterigen Eies liegen.
4.2.1 . 4 . Korrelierte Selektionserfolge in der Eizusammen
setzung bei 30 und bei 50 Wochen
Die korrelierten Selektionserfolge, definiert als Abwei
chungen der Selektionslinien (L1, L2) von der Kontrollinie,

7 0
sind für die zwei selektierten Generationen bei 30 und bei
50 Wochen in Tabelle 19 zusammengefasst.
Tabelle 19 Korrelierte Selektionserfolge bei 30 und bei 50 Wochen für verschiedene Merkmale der Eizusammensetzung
Generation
Linie
Merkmal
Eigewicht
g
Eiklar
Protein
Eidotter
Energie
MJ/kg
Trocken
substanz t
* p i 0.05
** p i 0 . 01
w 0
c h e
30
50
30
50
30
50
30
50
30
50
30
50
F2
L1 L2
. 41 . 58
1.12 1.27
. 14 .15
1.16** . 70**
. 34 . 60
.84* .61*
- .36 - . 31
- 1.09** - .72**
- .14 -.11
- .18* -.08*
- .18 -.08
- .61** -.37**
F3
L1 L2
2.57** 2.07**
1 . 63 1. 41
1.00** 1.15**
. 65 . 76
. 96** . 65**
.73* .18*
- 1.01 * * -. 97**
- . 74 - . 61
- . 17* -.14*
- .16* ·· .07*
54** - . 64**
38 - .47
Die Daten wurden für den Einfluss der Brut und des Stalles
korrigiert. Die Selektionserfolge in der dritten Generation
(F3) enthalten auch diejenigen der zweiten Generation (F2).
Die zwei Selektionslinien verhalten sich, wie erhofft, sehr
ähnlich.

71
Die Selektionserfolge waren in beiden Generationen positiv
für die Merkmale Eigewi c ht, Eiklaranteil und Proteingehalt,
währenddessen Eidotte ranteil , Energiegehalt und Trockensub
stanzgehalt durch die Selektion zurückgingen. Mit Ausnahme
des Merkmal es Eigewicht, wi c hen die Selektionslinien in der
selekt i erten Generati on F 2 bei 50 Wochen in allen Merkmalen
signifikant von d e r Kontroll inie ab. In der Generation F3
wurden sign i fikante Abwei c hungen v on der Kontroll inie für
a lle, i m Alter von 30 Wochen untersuchten Merkmale gefunden.
Die gröss ten Abwei c hungen s i nd in der F 2 beim Merkmal Ei
do tterante il festzustellen , nämlich - 1 .1 % bei 30 Wochen
resp . -3.1 % bei 50 Wochen gefolgt von den Merkmalen Eige
wi c ht mit 1 .0 % r esp . 2 . 0 %, Trockensubstanzgehalt mit
- 0 . 9 % resp . -2 . 0 %, Proteingehalt mit 0.8 t resp. 1.4 t,
Eiklar gehalt mi t 0 .2 % resp. 1 . 6 %, und zuletzt vom Ener
giegehalt mit - 0 . 3 % resp. -0 . 3 % bei 50 Wochen .
In der zweiten selektierten Generation (F3) lagen die Selek
tionslinien bei 30 Woc hen im Merkmal Eigewicht um 3.9 t bei
30 Wochen und 2.3 % bei 50 Wochen über der entsprechenden
Kontroll inie. Eine ebenfalls starke Veränderung konnte im
Merkmal Eidotteranteil festgestellt werden , nämlich -3.9 %
resp. -2.1 t gefolgt v om Trockensubstanzgehalt mit -2.6 %
resp. -1 . 7 t , dem Eiklaranteil mit 1.7 % resp. 1.1 % gefolgt
vom Proteingehalt mit 1.5 % resp. 1.0 % und zuletzt vom
Energiegehalt mit - 0.3 % resp. -0.5 %.
Aus der Literatur (Ambrosen et al. , 1981 ; Rodda et al. ,
1977) ist bekannt , dass mit zunehmendem Eigewicht der Anteil
Eiklar ansteigt , währenddessen der Anteil Eidotter sinkt .
Das wurde auch in dieser Arbeit gefunden . Dadurch, dass der
Anteil Eidotter durchschnittlich nur 25 % der Eimasse
beträgt, im Vergleich zum Eiklar mit einem Anteil von 64 %,
wirken sich Veränderungen im Eidotteranteil stärker aus. Die
korrelierten Veränderungen im Protein , in der Energie und
der Trockensubstanz liegen in erwarteter Richtung. Je

72
grösser die Eimasse , desto höher de r Proteingehalt und desto
tiefer der Energiegehalt und der Trockensubstanzgehalt.
In Linien welche entweder auf Eizahl oder auf Eigewi c ht se
lektiert wurden fanden Ambrosen et al. ( 1981} einen um 1 %
höheren Trockensubstanzgehal t im Eidotter der Selektions-
1 inien gegenüber der Kontrollinie, am höchsten für die Eige
wichtlinie. Der Trockensubstanzgehalt im Eiklar hingegen
sank durch die Selektion in der Eizahllinie und veränderte
sich kaum in der auf Eigewicht selektierten Linie. Der Trok
kensubstanzgehal t im Gesamtei aber veränderte sich bei der
auf Eizahl selektierten Linie kaum gegenüber der Kontrol
linie , während e r bei der auf Eigewich t selektierten Linie
über dem Gehalt der Kontrollinie lag . Die in diesem Experi
ment erzeugten Linien legen mehr und schwerere Eier , mit
einem tieferen Trockensubstanzgehalt im Gesamtei. Der Ener
giegehalt scheint durch die Selektion am wenigsten beein
f 1 usst worden zu sein , findet man doch nach der zweiten
selektierten Generation einen kaum veränderten Energiege
halt. Ambrosen et al. (1981) fanden in den auf hohes Eige
wicht selektierten Linien keine signifikanten Unterschiede
im Energiegehalt der Eidotter, obwohl deren Gewichte ange
stiegen waren. Dies stimmt somit mit den vorliegenden Unter
suchungen übere in. Sie stellten hingegen eine Veränderung
der Eizusammensetzung bezüglich Cholesterolgehalt im Ei
dotter bei denjenigen Linien fest, welche auf hohe Eizahl
gezüchtet wurden. Der Gehalt lag unter unter demjenigen der
Kontrollinie. Nach Marion et al. (1965) bestehen Unterschie
de im Energiegehalt der Eier verschiedener Linien welche je
doch nicht einfach auf die Eidottergrösse zurückgeführt wer
den können, sondern auch vom Verhältnis der Fette zu den
übrigen Eidotterbestandteilen wie Protein und Wasser ab
hängig sind. Dies bedeutet, dass die relativ geringe Abnahme
des Energiegehaltes nicht a priori auf eine selektionsbe
dingte Veränderung im Fettgehalt, sondern auch auf die Zu
nahme an Protein und Wasser, teilweise auch im Eidotter,
zurückzuführen ist . Bei effizienteren Futterumwandlern

7 3
scheint eine Tendenz zu höherer Wassereinlagerung zu be
stehen.
zusammenfassend kann gesagt werden, dass in beiden Genera
tionen bei 30 und bei 50 Wochen die gleiche Reihenfolge, mit
Ausnahme des Merkmales Eiklar, für die korrelierten Verände
rungen in der Eizusammensetzung gefunden wurde. Sie waren am
bedeutendsten für den Anteil Eidotter gefolgt vom Eigewicht,
der Trockensubstanz, dem Proteingehalt, dem Eiklaranteil und
dem Energiegehalt. Das Ausmass der Abweichungen veränderte
sich jedoch von der Generation F2 zu F3. In der Generation
F2 wurden die Abweichungen mit zunehmendem Alter grösser und
in der F3 kleiner.
4.2.1.5. Korrelierte Selektionserfolge in der Eizusammen
setzung bei 30 Wochen und bei 50 Wochen unter Be
rücksichtigung des Alterseinflusses für die Genera
tionen F2 und F3.
Wie in Tabelle 16 gezeigt wurde, haben die Eier je nach
Alter der Henne eine andere Zusammensetzung.
Mit zunehmendem Alter steigt das Eigewicht an und bewirkt
eine Verschiebung der Ei zusammensetzung. Mit fortgeschr i t
tenem Alter einer Henne werden ihre Eier einerseits eidot
ter- und energiereicher und enthalten mehr Trockensubstanz.
Andererseits sinkt der Anteil Eiklar und mit ihm der Pro
teingehalt. Diese Veränderungen wurden in beiden untersuch
ten selektierten Generationen beobachtet.
Ein Vergleich der beiden Generationen (F2 und F3) in Tabelle
20 zeigt jedoch, dass sich die Eizusammensetzung, gerechnet
als Durchschnitt der Selektionslinien und der Kontrollinie,
in der dritten Generation altersbedingt stärker veränderte.

74
Tabelle 20 Veränderung der Eizusammensetzung zwischen 30 Wochen und 50 Wochen für die zweite (F2) und die dritte (F3) Generation in Prozent
Generation
Merkmal
Eigewicht g
Eiklargewicht g
Eiklaranteil %
Eidottergewicht g
Eidotteranteil %
Trockensubstanz-
gehalt (TS) %
Proteingehalt in
der TS %
Energiegehalt in
der TS MJ/kg
F2
+11%
+ 8%
- 3%
+19%
+ 8%
+ 4%
- 5%
+ 2%
F3
+14%
+10%
- 4%
+24%
+11%
+ 6%
- 7%
+ 2%
Weil einerseits gröss ere Ei e r mehr Eiklar enthalten und an
dererseits Eier von älteren Hennen eidotterreicher sind,
steigen, als Folge des stark zunehmenden Eigewichtes , sowohl
die Menge Eiklar als auch die Menge Eidotter an, wobei letz
tere stärker anstieg . Dies ist einerseits auf das kleinere
Verhältnis Eidotter zu Eimasse gegenüber Eiklar zu Eimasse
zurückzuführen und andererseits auf den Einfluss des Alters .
Aus dieser Tatsache lässt sich der damit verbundene höhere
Anstieg des Trockensubstanz- und weniger ausgeprägt, des
Energiegehaltes erklären , sowie auch der tiefere Protein
gehalt im Alter von 50 Wochen gegenüber 30 Wochen.
Weiter zeigen die vorliegenden Resultate , dass je tiefer der
Dotteranteil bei Legebeginn, desto grösser die altersbeding
ten Schwankungen.

75
Die Literaturangaben über den Alterseinfluss auf Eigrösse
und Eizusammensetzung ( Ambrosen et al., 1981; Fletcher et
al., 1981; Anderson et al., 1978; Hurnik et al., 1977;
Marion et al., 1964; Cunningham et al., 1960) bestätigen die
in den Tabellen 16 und 20 zusammengestellten Resultate.
Marion et al. ( 1964) untersuchten nach 4 Generationen die
altersbedingten Veränderungen in der Eizusammensetzung von
Linien welche auf grosse, resp. kleine Eier gezüchtet worden
waren. Sie stel 1 ten dabei fest, dass die Ei gewichte einer
Henne , die bei Legebeginn schon relativ schwere Eier legt,
mit dem Alter weniger ansteigen, als solche von Hennen mit
kleinerem Anfangseigewicht. Der Eiklaranteil der schwereren
Eier sank weniger als derjenige von kleineren, währenddessen
der Eidotteranteil der schwereren Eier im Laufe einer Lege
periode stärker zunahm, weil dieser zu Beginn auf Grund der
grösseren Eier relativ kleiner war. Je tiefer der Eidotter
anteil zu Beginn der Legeperiode, desto grösser sein An
stieg . Hurnik et al. (1977) haben in ihren Untersuchungen an
gefriergetrocknetem Eiklar und Eidotter nachgewiesen, dass
mit zunehmendem Alter das Gewicht von gefriergetrocknetem
Eidotter stärker anstieg, als dasjenige von Eiklar. Mit an
deren Worten ist der altersbedingte Anstieg des Eigewichtes
vor al lern auf die Zunahme des Trockensubstanzgehal tes im
Eidotter und den ziemlich konstant bleibenden Wassergehalt
im Eiklar zurückzuführen.
Vergleicht man den Durchschnitt der Selektionslinien mit den
Resultaten der Kontrollinie (Tabelle 21), so stellt man in
diesem Versuch folgendes fest: die Eier der Selektionslinien
sind in beiden Generationen bei 30 und bei 50 Wochen schwe
rer und haben mehr Eiklar und Protein, jedoch weniger Eidot
ter, Energie und Trockensubstanz als die Eier in den ent
sprechenden Kontrollinien. In der zweiten Generation erhöhte
sich in den Selektionslinien im Vergleich zur Kontrollinie
das Eigewicht im Laufe der Legeperiode stärker. Deshalb
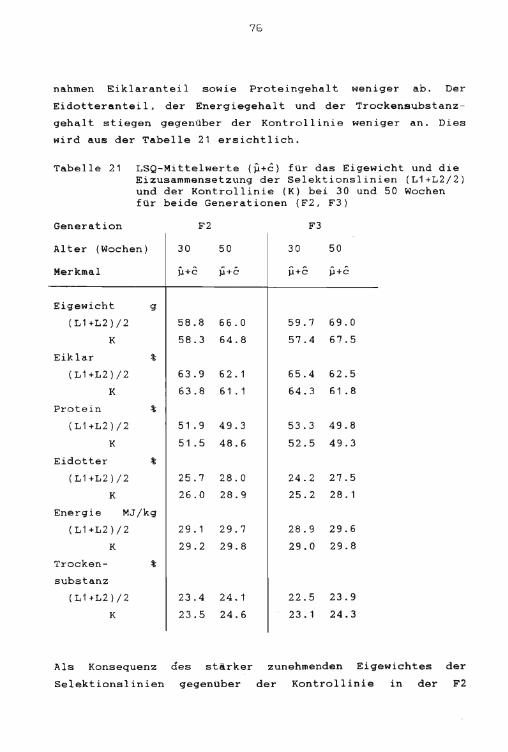
75
nahmen Eiklaranteil sowie Proteingehalt weniger ab. Der
Eidotteranteil, der Energiegehalt und der Trockensubstanz
gehalt stiegen gegenüber der Kontrollinie weniger an. Dies
wird aus der Tabelle 21 ersichtlich.
Tabelle 21 LSQ-Mittelwerte (}i+cJ für das Eigewicht und die Eizusammensetzung der Selektionslinien (L1+L2/2} und der Kontrollinie (K} bei 30 und 50 Wochen für beide Generationen (F2, F3}
Generation F2 F3
Alter (Wochen} 30 50 30 50
Merkmal ii+c ii+c il+c jj.+c
Eigewicht g
(L1+L2} /2 58.8 66 . 0 59.7 69.0
K 58.3 64.8 57.4 67.5
Eiklar %
(L1+L2} /2 63.9 62 .1 65.4 62.5
K 63.8 61 . 1 64.3 61. 8
Protein %
(L1+L2}/2 51 . 9 49.3 53.3 49.8
K 51 . 5 48.6 52.5 49 . 3
Eidotter %
(L1+L2}/2 25.7 28.0 24.2 27.5
K 26.0 28.9 25.2 28. 1
Energie MJ/kg
(L1+L2} / 2 29.1 29.7 28.9 29.6
K 29.2 29.8 29 . 0 29.8
Trocken- %
substanz
(L1+L2} /2 23.4 24 .1 22.5 23.9
K 23.5 24.6 23.1 24.3
Als Konsequenz des stärker zunehmenden Eigewichtes der
Selektionslinien gegenüber der Kontrollinie in der F2

77
vergrösserten sich die Abweichungen sowohl für den Eiklar
und den Proteingehalt wie auch für den Eidottergehalt und
die von ihm abhängigen Merkmale Energie und Trockensubstanz
gehal t. Mit zunehmendem Alter unterscheiden sich deshalb die
Eier der Selektions! inien der zweiten Generation in ihrer
Zusammensetzung immer stärker von denjenigen der Kontrol
linie (vgl. Tabelle 19).
In der Generation F3 hingegen nahm das Eigewicht der Selek
tionsl inien weniger zu als in der Kontrollinie, deshalb san
ken Eiklar- und Proteingehalt stärker und der Eidotteranteil
sowie der Trockensubstanzgehalt stiegen stärker an. In der
Generation F3, war das Eigewicht der Selektionslinien bei
Legebeginn wahrscheinlich schon sehr hoch, deshalb stieg es
während der Legeperiode weniger an. Weil das Eigewicht weni
ger zunahm, veränderte sich die altersbedingte Zusammen
setzung um so mehr. Die Eier der Selektionslinien wurden
einerseits eiklar- und proteinärmer und andererseits ei
dotter-, energie- und trockensubstanzreicher. Somit ver
ringerten sich die Abweichungen von der Kontrollinie zwi
schen 30 und 50 Wochen (vgl. Tabelle 19).
Diese Erhebungen lassen den Schluss zu, dass je grösser die
Unterschiede im Eigewicht, bei gleichem Alter , desto grösser
die Abweichungen in der internen Zusammensetzung, und je
grösser die altersbedingte Zunahme im Eigewicht, desto ge
ringer die Veränderungen in der Zusammensetzung und umge
kehrt.
4.2.2. Genetische Parameter
4.2.2.1. Genetische Korrelationen der Eizusammensetzung in
den drei Generationen innerhalb der beiden Perioden
bei 30 und bei 50 Wochen
Im vorigen Kapitel wurden die Selektionserfolge der ver
schiedenen Merkmale besprochen und dabei konnte beobachtet

78
werden, dass sogenannte Merkmalsgruppen bestehen. Dieses
Kapitel befasst sich mit den genetischen Korrelationen
zwischen zwei Merkmalen, deren Beziehungen entweder negativ
oder positiv sind. Die Erfassung der korrelierten Merkmale
ist im Zusammenhang mit Veränderungen, die durch die Selek
tion bewirkt werden, von grosser Bedeutung, weil die gene
tische Verbesserung eines Merkmales zu korrelierten Verän
derungen in anderen Merkmalen führt. Die genetische Ursache
einer Korrelation kann Kopplung oder Pleiotropie sein, wobei
ein Gen, oder verschiedene eng gekoppelte Gene mehrere Merk
male beeinflussen. Auf Grund der verschiedenen Strukturen
der Populationen zu Beginn des Experimentes mussten die ge
netischen Parameter mit den entsprechenden Modellen ge
schätzt werden. Die drei Schätzungen eines Parameters wur
den mit ihren Standardfehlern gewichtet und zu einem einzi
gen Wert zusammengefasst. Die genetischen Korrelationen sind
für die verschiedenen Merkmale in Tabelle 22 zusammenge
stellt.
Die tiefsten genetischen Korrelationen bestehen zwischen dem
Merkmal Eigewicht und den übrigen Merkmalen. Die zwei Grup
pen bestehend aus den Merkmalen Eigewicht, Eiklar und Prote
in einerseits, sowie Eidotter, Energie und Trockensubstanz,
andererseits sind untereinander deutlich negativ korreliert.
Die Beziehungen sind eng und liegen im Bereich von -0.67 bis
-0.99 mit Ausnahme für das Merkmal Eigewicht (0.53). Es kon
nte ebenfalls festgestel 1 t werden, dass die Korrelationen
zwischen den Merkmalen Eigewicht und Eiklaranteil in beiden
Perioden höher waren, als zwischen den Merkmalen Eigewicht
und Eidotter. Mit zunehmendem Alter sinken die genetischen
Korrelationen zwischen dem Merkmal Eigewicht und den übri
gen. Für das Merkmal Eiklar bleiben sie unverändert. Im Ge
gensatz dazu sind die Korrelationen für alle anderen unter
suchten Merkmale beim Alter von 50 Wochen höher als bei 30
Wochen.

79
Tabelle 22 Gewichtete genetische Korrelationen und Standardfehler (s.e.) zwischen den Merkmalen Eigewicht (EW), Eiklar (EK), Protein (P)' Eidotter (ED), Energie (E) und Trockensubstanz {TS) bei 30 Wochen ( 1/1 ) und bei 50 Wochen (2/2)
Merkmal EK s.e. p s . e. ED s.e. E* s.e. TS s.e .
Alter 1/1 1/ 1 1/1 1/1 1/1
2/2 2/2 2/2 2/2 2/2
Eigewicht g
1/1 . 75 . 1 7 .51 .16 -.55 . 20 -.53 . 19 -.68 .27
2/2 .65 . 19 . 33 .29 -.41 .28 -.32 .30 -.36 .26
Eiklar t
1/1 .94 .05 -.98 .32 -.97 .30 -.92 .28
2/2 .95 .05 -.98 . 32 - . 97 . 30 -.92 .28
Protein
1 /1 -.90 .23 -.99 .23 -.67 .27
2/2 - . 94 .36 -.99 . 36 -.78 . 31
Eidotter t
1/1 .99 .03 .90 .05
2/2 .95 .05 .93 .04
Energie MJ/kg
1 /1 .82 . 12
2/2 .88 .10
* Die genetischen Korrelationen der Untersuchungen bei 50
Wochen beruhen für das Merkmal Energie nur auf der zweiten
Generation, weil bei der F3 bei 50 Wochen eine negative
Varianzkomponente geschätzt wurde.
Die Standardfehler liegen im mittleren Bereich und sind bei
den positiven Korrelationen geringer als bei den negativen.
Die genetischen Korrelationen bestätigen die im Kapitel
4.2.1.3. diskutierten phänotypischen Beziehungen und liegen

80
sogar etwas darüber. Im Vergleich zu den Angaben von Tawfik
(1981) sind die gefundenen Korrelationen deutlich höher aus
gefallen, sie liegen jedoch im Bereiche der von Hill (1965)
publizierten Werte .
Auf Grund der verschiedan gerichteten Korrelationen zwischen
Eigewicht und Eiklaranteil resp. Eidotteranteil ist zu er
warten, dass ein positiv korrelierter Selektionserfolg bei
der Eimasse zu einer relativen Änderung zugunsten des Ei
klar- auf Kosten des Dotteranteiles führen wird. Weil eine
deutlich positive Korrelation zwischen Eigewicht und Prote
ingehalt besteht, kann mit ansteigendem Eigewicht auch eine
Zunahme des Proteingehaltes erwartet werden. Die positiven
Korrelationen zwischen Eidotteranteil und Energie- resp.
Trockensubstanzgehalt lassen wegen der negativen Beziehung
zwischen Eigewicht und Dot terantei 1 auf eine Verminderung
des Energie- und des Trockensubstanzgehaltes schliessen.
Diese Vermutungen konnten mit diesem Selektionsexperiment
bestätigt werden . Aus den Beziehungen des Eigewichtes zu den
Komponenten des Eies lässt sich ableiten, dass schwerere
Eier relat i v mehr Eiklar (rg +0.75 ± 0.17) und weniger
Eidotter (rg = - 0 . 55 ± 0 .2 ) enthalten.
Hurnik (1976) fand einerseits , dass das Gewicht von frischem
Eidotter und von frischem Eiklar mit zunehmendem Alter an
steigt , dass andererseits mit zunehmendem Alter j edoch das
Gewicht von gefriergetrocknetem Eidotter stärker anstei~t
als dasjenige von gefriergetrocknetem Eiklar, welches zuerst
leicht ansteigt um dann wieder auf gleiches Niveau zu sinken
wie zu Beginn der Legeperiode . Er zieht daraus den Schluss,
dass die altersbedingten Veränderungen im Trockensubstanz
gehal t von frischem Eidotter und Eiklar unterschiedlich sind
und dass der offensichtliche Anstieg in getrocknetem Eidot
ter vor allem zu Beginn der Legeperiode darauf schliessen
lässt, dass ein bedeutender Teil der Zunahme des Eigewichtes

81
während einer Legeperiode auf einen ansteigenden Trockensub
stanzgehalt im Eidotter und einen zunehmenden Gehalt an
Wasser im Eiklar zurückzuführen ist. Demzufolge muss im
Eiklar der Anteil an anderen Substanzen wie Protein zum
Beispiel, sinken. Der Fettgehalt im Eidotter nimmt nach
Hurnik (1976) mit zunehmendem Alter stark zu.
Ein korrelierter Selektionserfolg im Eigewicht kann nach
einiger Zeit zu Veränderungen der genetischen Korrelationen
führen, da Kopplungsungleichgewichte oder pleiotrope Gene,
welche beide Merkmale positiv beeinflussen, durch die Selek
tion fixiert werden können und somit die Varianz dieser
Merkmale verringert wird.
4.2.2.2 . Genetische Korrelationen der Eizusammensetzung
zwischen den Merkmalen der zwei Untersuchungs
perioden
Da bekannt ist, dass sich die Eizusammensetzung im Laufe des
ersten Legejahres verändert, wurden die Korrelationen
zwischen Merkmalen, welche sowohl bei 30 wie bei 50 Wochen
erhoben wurden, berechnet.
Die in Tabelle 23 zusammengefassten gewichteten genetischen
Korrelationen beruhen auf Angaben der Generation F2 und der
Generation F3 mit Ausnahme der Beziehungen für das Merkmal
Energie. Die genetischen Korrelationen zwischen der ersten
und der zweiten Periode sind mit Ausnahme des Merkmales
Eigewicht sehr hoch (rg= 0.6 - .99) und die Standardfehler
sind gering bis mittel.
Das Eigewicht der Untersuchungsperiode bei 30 Wochen scheint
in enger Beziehung zum Eigewicht bei 50 Wochen zu stehen.
Die Beziehung zwischen dem Eigewicht bei 30 Wochen und der
Eizusammensetzung bei 50 Wochen ist jedoch weniger eng. Die
genetischen Korrelationen zwischen den Merkmalen der Ei
zusammensetzung in der ersten Periode und ihren entsprechen-
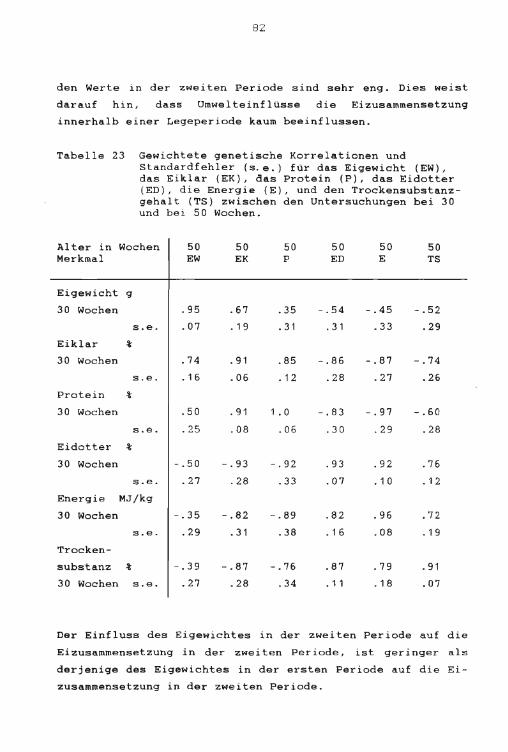
82
den Werte in der zweiten Periode sind sehr eng. Dies weist
darauf hin, dass Umwelteinflüsse die Eizusammensetzung
innerhalb einer Legeperiode kaum beeinflussen.
Tabelle 23 Gewichtete genetische Korrelationen und Standardfehler (s. e.) für das Eigewicht (EW), das Eiklar (EK), das Protein (P)' das Eidotter (ED), die Energie (E) , und den Trockensubstanz-gehalt (TS) zwischen den Untersuchungen bei 30 und bei 50 Wochen.
Alter in Wochen 50 50 50 50 50 50 Merkmal EW EK p ED E TS
Eigewicht g
30 Wochen . 95 .67 . 35 - .54 -.45 -.52
s.e. . 07 . 19 .31 . 31 .33 .29
Eiklar %
30 Wochen .74 . 91 . 85 -.86 -.87 -.74
s . e. . 16 .06 . 12 .28 .27 . 26
Protein %
30 Wochen .50 . 91 1 . 0 -.83 - . 97 -.60
s.e. .25 . 08 .0 6 .30 .29 .28
Eidotter %
30 Wochen -.50 -.93 -.92 .93 .92 .76
s . e. .27 .28 .33 . 07 . 10 . 1 2
Energie MJ/kg
30 Wochen - .35 -.82 -.89 . 82 .96 .72
s.e . . 29 . 31 .38 . 16 . 08 . 19
Trocken-
substanz % -.39 -.87 -.76 .87 . 79 .91
30 Wochen s.e. .27 .28 . 34 . 11 . 18 .07
Der Einfluss des Eigewichtes in der zweiten Periode auf die
Eizusammensetzung in der zweiten Periode, ist geringer als
derjenige des Eigewichtes in der ersten Periode auf die Ei
zusammensetzung in der zweiten Periode.

83
4.2.2.3. Heritabilitäten
Wegen der verschiedenen Populationsstrukturen wurden die h 2 -
Werte für die verschiedenen Merkmale gewichtet. Sie sind in
Tabelle 24 enthalten.
Die h 2 Werte für die Energie in der F3 konnten wegen der ne
gativen Varianzkomponenten nicht berechnet werden.
Die Werte sind relativ hoch und weisen tiefe Standardfehler
auf . Bei 30 Wochen ist die Heritabilität am höchsten für den
Eiklargehalt (0 . 75) , gefolgt von Dottergehalt, Proteinge
halt, Energiegehalt, Eigewicht und dem Trockensubstanzgehalt
mit dem tiefsten Wert von 0.4 .
Die Ergebnisse liegen im Rahmen der in der Literatur ange
gebenen Werte. Hilfiker (1970) schätzte für das durch
schnittliche Eigewicht eine Heritabilität von 0.63 in einer
auf kleines Anfangseigewicht selektierten Linie und von 0.73
in einer auf hohes Anfangseigewicht selektierten Linie.
Hagger (1977) ermittelte Werte zwischen 0.37 und 0.73, Damme
(1984) je nach Modellwahl zwischen 0.58 und 0.73 und Rodda
et al. (1977) einen Wert von 0 . 42. Jaffe (1965) gab einen
Wert von 0.7 an, Scheinberg (1953) 0.56, Cahaner (1979)
0 . 64 während Hill (1966) Werte von 0.55 aufführte. Nagai et
al. (1969) und Tawfik et al. (1976) beobachteten, dass die
h 2 Werte für das Eigewicht und die Merkmale der
Eizusammensetzung im Laufe einer Legeperiode sanken. Tawfik
stellt gleichzeitig fest, dass Heritabilitäten im dritten
Legemonat, bei höchster Legetätigkeit, am tiefsten sind und
dann leicht anstiegen um wieder zu sinken. Er erklärt das
mit einer generell hohen Leistungsbereitschaft von
Junghennen wodurch genetische Unterschiede nicht so stark
zum Tragen kommen .

84
Tabelle 24 Heritabilitäten (h2) und Standardfehler (s.e.) für Eigewicht und Eizusammensetzung bei 30 und 50 Wochen
Merkmal 30 Wochen 50 Wochen
h2 s.e . h2 s.e .
Eigewicht g .60 . 15 .69 .22
Eiklar .\ . 75 .15 .70 .22
Protein .\ . 70 .15 .36 .20
Eidotter .\ . 70 . 15 .59 . 22
Energie MJ/kg . 66 . 15
Trocken- .\ . 40 .13 . 85 .22
substanz
Dies wird auch in diesem Versuch bestätigt; der h 2 Wert für
das Me r kmal Eigewicht ist bei 30 Wochen geringer als bei 50
Wochen. Die Werte für die Merkmale Protein- und Eidotter
gehal t und in etwas geringerem Ausmass für den Anteil Ei
klar sinken mit zunehmendem Alter mit Ausnahme des Merkmales
Trockensubstanzgehalt. Aus diesen hohen h 2 Werten und wegen
der hohen genetischen Korrelationen lässt sich folgern , dass
in den nächsten Generationen mit Veränderungen in der Eizu
sammensetzung gerechnet werden kann, bis zu dem Moment wo
das Eigewicht oder die Eizusammensetzung das Optimum über
schreitet. Dies kann zum Beispiel dann der Fall sein, wenn
die Eizusammensetzung nicht mehr den Anforderungen für gute
Schlupffähigkeit entspricht, oder sich die genetischen
Korrelationen als Auswirkung der Selektion oder wegen
zufälliger Drift verändern.

85
4.2.2.4. Regression des Futterverbrauches auf verschiedene
Merkmale der Eizusammensetzung
Die geschätzten multiplen linearen Regressionskoeffizienten
erlauben es, die zusammenhänge zwischen dem Futterverbrauch
und verschiedenen anderen Merkmalen zu erfassen.
Das Interesse solcher Untersuchungen für die Praxis 1 iegt
darin, dass die Regressionskoeffizienten als Merkmal zur
Schätzung des Futterverbrauches eingesetzt werden könnten.
Die Regressionen des Futterverbrauches auf die verschiedenen
Merkmale wurden mit den Stichprobentieren der Ei zusammen
setzungsuntersuchungen durchgeführt und sind in Tabelle 25
für die erste Periode der Generationen F1, F2 und F3 und in
Tabelle 26 für die zweite Periode der Generationen F2 und F3
aufgeführt.
Vier verschiedene Modelle wurden zur Schätzung des Futter
verbrauches miteinander verglichen. In allen Modellen wurde
der Futterverbrauch als Funktion des Körpergewichtes, der
Körpergewichtsveränderung und des Alters bei Legebeginn (nur
erste Periode) betrachtet. Je nach Modell (VII - X) wurden
die Eimasse, die Energie in der Eimasse, die Dottermasse,
die Klarmasse, die Proteinmasse oder die Schalenproduktion
miteinbezogen. Die Variation im Bestimmtheitsmass der ver
schiedenen Modelle war gering. Das Bestimmtheitsmass liegt
bei allen 4 Modellen um 0.8 und ist demzufolge hoch. 85 t
der Varianz des Futterverbrauches in der ersten Periode
gegenüber 80 t in der zweiten Periode konnten durch die im
Modell berücksichtigten Variablen erklärt werden. In der
ersten Periode (21. - 40. Woche) der ersten Generation lag
der Regressionskoeffizient für das Körpergewicht bei durch
schnittlich 3.28 kg mehr Futter pro zusätzliches kg Körper
gewicht, gegenüber 3.51 kg und 3.53 kg in den folgenden, wo
bei dafür keine Erklärung gefunden werden konnte. Der Re
gressionskoeff izient sank für den zusätzlichen Futterver-

85
brauch pro kg Körpergewichtsveränderung zwischen der 21. und
der 40 . Woche von 2.66 kg auf 2.25 kg resp. 1.47 kg. Dieser
Trend kann womöglich durch eine Veränderung in der Effizienz
zustande gekommen sein . Das Alter bei Legebeginn hatte nur
einen geringen Einfluss auf den Futterverbrauch. wie aus
Modell IX ersichtlich wird, braucht es bis zu vier mal mehr
Futter um Dottermasse zu produzieren als Klarmasse. Diese
Werte helfen die Tatsache erklären, dass mit zunehmendem
Eidotteranteil der Trockensubstanzgehalt im Eidotter an
steigt , währenddessen mit zunehmendem Eiklargehalt der
Wassergehalt im Eiklar zunimmt. Es braucht somit weniger
Futter um ein zusätzliches kg Eiklar gegenüber einem zu
sätzliches kg Eidotter zu produzieren. Aus Modell VIII wird
ersichtlich, dass 0 . 12 kg Futter benötigt werden, um ein
zusätzliches MJ Energie zu produzieren . Interessant ist es
hingegen festzustellen , dass in der e r sten Periode (21.
40. Woche) zwischen den Generationen Unterschiede bestehen
bezüglich Futterbedarf pro zusätzliches kg Protein. Braucht
es in der ersten Generation 1.6 kg Futter pro zusätzliches
kg Protein, werden in der zweiten 6.34 kg und in der dritten
4.43 kg benötigt .
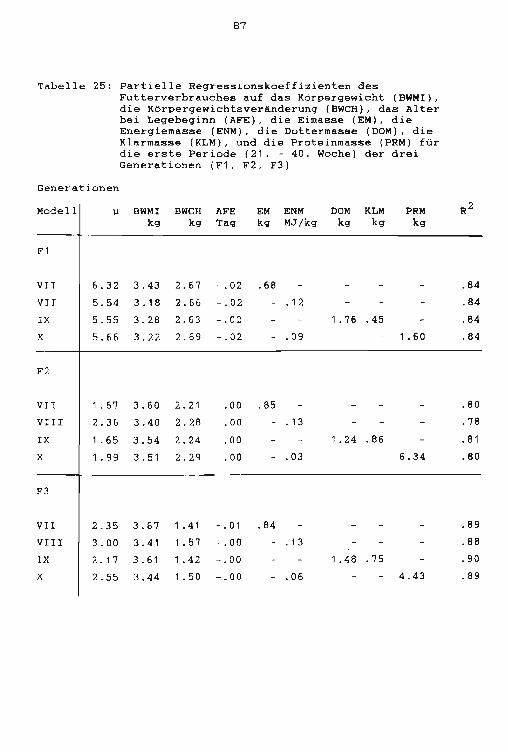
87
Tabelle 25: Partielle Regressionskoeffizienten des Futterverbrauches auf das Körpergewicht (BWMI), die Körpergewichtsveränderung (BWCH), das Alter bei Legebeginn (AFE), die Eimasse (EM), die Energiemasse (ENM), die Dottermasse (DOM), die Klarmasse (KLM), und die Proteinmasse (PRM) für die erste Periode ( 21. - 40. Woche) der drei Generationen (F1, F2, F3)
Generationen
Modell µ BWMI BWCH AFE EM ENM DOM KLM PRM R2 kg kg Tag kg MJ/kg kg kg kg
F1
VII 6.32 3.43 2.67 -.02 .68 .84
VII 5.54 3 .18 2.66 -.02 - . 12 .84
IX 5 . 55 3.28 2.&3 -.02 1. 76 .45 .84
X 5 . 66 3.22 2.69 -.02 - . 09 1.60 .84
F2
VII 1. 67 3.60 2.21 .00 .85 .80
VIII 2.36 3.40 2.28 .00 - . 13 .78
IX 1. 65 3.54 2.24 .00 1. 24 . 86 .81
X 1 . 99 3.51 2.29 .00 - .03 6.34 .80
F3
VII 2.35 3.67 1 . 41 - . 01 .84 .89
VIII 3.00 3.41 1 .57 - .00 - . 13 .88
IX 2. 17 3.61 1. 42 -.00 1. 48 .75 .90
X 2.55 3.44 1 .50 -.00 - . 06 4.43 .89

8 8
Tabelle 26: Partielle Regressionskoeffizienten des Futterverbrauches auf das Körpergewicht (BWMI), die Körpergewichtsveränderung (BWCH), die Eimasse (EM), die Energiemasse (ENM) , die Dottermasse (DOM) , die Klarmasse (KLM), und die Proteinmasse (PRM) für die zweite Periode ( 41 . -60. Woche) der zwei Generationen (F2, F3)
Generationen
Modell µ BWMI BWCH EM ENM DOM KLM PRM R2 kg kg kg MJ/kg kg kg kg
F2
VII 3 . 50 3.46 .54 . 73 .75
VIII 3 . 75 3.38 .34 . 11 . 75
IX 3 . 38 3.43 .53 1 . 1 7 . 69 .75
X 3.55 3 . 44 . 3 6 . 06 3.41 . 75
F3
VII 3 .9 5 3.26 2.08 .6 2 .84
VIII 3.9 6 3 . 05 2. 01 . 11 . 85
I X 3. 68 3 . 15 2 .06 1. 56 .42 .85
X 3. 91 3.07 2 .0 2 . 08 1 . 70 .85
Wie Tabell e 26 ze i gt , hat si c h in der zweiten Periode (41 .-
60. Woc he ) der Reg r ess ionskoeffizient für das Kö rpergewicht
kaum verändert und beträgt 3.4 kg pro zusätzliches kg Kör
perge wi c h t in der Generation F2 gegenüber 3.1 kg in der Ge
neration F3 .
Ein grosser Unterschied zwischen den Regressionskoeffi
zienten der Generati onen F2 und F3 ist beim Merkmal Körper
gewichtsveränderung feststellbar. In der zweiten Generation
betrug der Futterverbrauch durchschnittlich nur 0.45 kg pro
zusätzliches kg Körpergewichtsveränderung gegenüber 2.04 kg
in der dritten Generation. Diese sehr grossen Unterschiede
lassen sich nicht erklären , können aber möglicherweise durch

89
eine andere Körperzusammensetzung (Fetteinlagerungen) oder
eine andere Futterzusammensetzung in der zweiten gegenüber
der dritten Generation beeinflusst worden sein.
Unter dem Einfluss der Selektion nahm das Eigewicht zu und
die Eizusammensetzung veränderte sich, indem der Anteil Ei
klar anstieg und der Anteil Eidotter sank. Auf Grund des
höheren Anteiles Eiklar mit höherem Wassergehalt sank der
Futterbedarf pro zusätzliches kg Eimasse von der zweiten zur
dritten Generation (0.73 resp. 0.62). Werden für die zweite
Periode die um die Merkmale Energie und Protein erweiterten
Modelle beider Generationen miteinander verglichen, so kann
festgestellt werden, dass der Einfluss des Energiegehaltes
des Eies auf den Futterverbrauch gleich war, während der
Einfluss der Proteinmasse auf den Futterverbrauch in der
Generation F3 deutlich tiefer lag als in der Generation F2.
Dies verhält sich genau umgekehrt zur ersten Periode, wo der
zusätzliche Bedarf an Futter für die zusätzliche Produktion
von Protein von Generation zu Generation stark anstieg. Es
scheint als ob sich mit zunehmendem Alter und Anzahl Gene
rationen die Protein- und Energiegehalte immer weniger ver
ändern werden und dass die Gewichtszunahme im Eiklar und im
Eidotter tatsächlich vor allem auf einen erhöhten Wasser
gehalt zurückzuführen sind . Die Variationen von Generation
zu Generation in den Regressionskoeff izienten ist nicht er
klärbar.
4.2.2.5. Genetische Korrelationen zwischen dem Selektions
kriterium und den Merkmalen der Eizusammensetzung
In Tabelle 27 wurden die Korrelationen zwischen den ver
schiedenen Eizusammensetzungsmerkmalen und dem Selektions
kriterium resp . der "analogen Differenz" aufgeführt. Für die
erste Periode in der dritten Generation konnten keine Werte

90
angegeben werden, da die genetische Varianz des Selektions
kri teriums im untersuchten Material sehr klein war und die
Korrelati o nen somit ausserhalb der Grenzen lagen.
Anhand dieser Tabelle kann einzig festgestellt werden, dass
positive Korrelationen zwischen dem Selektionskriterium,
resp. der "analogen Differenz" und den Merkmalen Eigewicht,
Eiklaranteil und Proteingehalt bestehen. Negative Korre
lationen wurden zwischen dem Selektionskriterium, resp. der
"analogen Differenz" und den Merkmalen Anteil Eidotter und
Energie- und Trockensubstanzgehal t gefunden . Die entspre
chenden Standardfehler sind jedoch relativ hoch. Es können
somit keine eindeutigen Tendenzen festgestellt werden, weder
innerhalb einer Generation, noch innerhalb Perioden oder
zwischen den beiden Perioden.
Tabelle 27 Genetische Korrelationen und Standardfehler (s.e.) zwischen dem Selektionskriterium, resp. der analogen Differenz und den Merkmalen der Eizusammensetzung in den drei Generationen (F1 , F2, F3)
Selektio nskriterium analoge Differenz
Generation F1 s . e. F2 s . e. F2 s.e . F3 s.e.
Merkmal
Eigewicht g . 5 1 .32 .93 . 51 .49 .44 .28 . 51
Eiklar % . 56 . 32 .82 . 51 .56 .39 . 49 .49
Protein % .20 .34 .78 .54 .35 .44 .23 1 . 2
Eidotter % -.63 .48 - . 72 .80 - . 61 . 51 -. 54 .88
Energie MJ/kg -.4 7 .42 -.44 . 71 -.48 .49
Trocken-
substanz % -1 . 0 1 . 0 -.23 . 70 - .60 . 49 -.7 7 .77

91
4.2.3. Schlussfolgerungen
Durch die Selektion auf "Eierlös minus Futterkosten" konnte
ein korrelierter _Selektionserfolg im Eigewicht bei 30 Wochen
im Durchschnitt beider Selektionslinien von +0.49 g in der
ersten und von +2. 32 g in der zweiten Generation erzielt
werden. Die Eier wurden dadurch eiklarreicher (0.14 t resp.
1 . 75 t) und proteinreicher (0.47 t resp. 0.85 %). Der Anteil
Eidotter sank um 0.35 t resp. 0.99 t, der Energiegehalt um
0. 1 3 resp. 0. 16 t und der Trockensubstanzgehal t um 0. 13 .t,
resp. 0.59 t.
Der Heterosisef fekt auf das Eigewicht war gross in der
ersten Generation. Von der ersten zur zweiten Generation
musste ein Rückgang des Eigewichtes festgestellt werden, der
als Heterosisverlust interpretiert werden kann.
Zwischen den Merkmalen Eigewicht, Eiklaranteil und Protein
gehalt bestehen hohe, positive genetische Korrelationen. Sie
sind am geringsten zwischen dem Eigewicht und dem Proteinge
halt ( +O. 51) und am höchsten zwischen Eiklarantei 1 und Pro
teingehalt (+0.94). Diese drei Merkmale, Eigewicht, Eiklar
anteil und Proteingehalt, bilden eine Gruppe. Hohe positive
genetische Korrelationen im Bereich von 0.82 - 0.99 konnten
ebenfalls bei der anderen Gruppe, bestehend aus den Merkma
len Eidotteranteil, Energiegehalt und Trockensubstanzgehalt,
gezeigt werden. Zwischen den beiden Gruppen bestehen jedoch
deutlich negative Beziehungen. Die tiefsten Korrelationen
wurden zwischen dem Merkmal Eigewicht und den übrigen Merk-
malen ermittelt (0.51 0. 75). Die Korrelationen bei 30
Wochen zwischen dem Eigewicht und den übrigen Merkmalen
waren etwas höher als bei 50 Wochen.
Die Korrelationen zwischen den anderen Merkmalen veränderten
sich kaum von 30 zu 50 Wochen oder nahmen nur leicht zu. Die
Eizusammensetzung scheint demzufolge bei älteren Hennen von
der Eigrösse und zusätzlich vom Alter abhängig zu sein.
Grundsätzlich steigen mit zunehmendem Alter das Eigewicht,

92
der Anteil Eidotter sowie der Energiegehalt und der Trocken
substanzgehalt auf Kosten des Anteiles Eiklar und des Pro
teingehaltes. Je grösser die Zunahme im Eigewicht, als Folge
des Alterungseffektes, desto geringer die Abnahme im Anteil
Eiklar und im Proteingehalt und desto geringer die Zunahme
im Anteil Eidotter, im Energie- sowie im Trockensubstanz
gehalt.
In der ersten selektierten Generation liegt das Eigewicht
der Selektionslinien bei 30 Wochen um 0.5 g über demjenigen
der Kontrollinie. Im Laufe der Legeperiode ist die Gewichts
zunahme für die Selektionslinien höher als für die Kontrol
l inie. Demzufolge sinken mit zunehmendem Alter der Eiklar
gehal t und der Proteingehalt weniger stark, und der Eidot
teranteil sowie der Trockensubstanzgehalt nehmen weniger zu .
Weil einerseits in der Generation F2 die Eier der Selek
tionslinien bei 30. Wochen grösser und auch eiklar- und pro
teinreicher waren, und zudem einen tieferen Eidottergehalt
aufwiesen mit weniger Energie und Trockensubstanz als die
Eier aus der Kontrollinie, und sich andererseits die Eizu
sammensetzung der Selektionslinien auf Grund der höheren
Gewichtszunahme zwischen 30 Wochen und 50 Wochen gegenüber
der Kontrollinie weniger veränderte, waren die korrelierten
Selektionserf olge in der Generation F2 bei 50 Wochen grösser
als bei 30 Wochen.
In der zweiten selektierten Generation (F3) liegt das Eige
wicht der Selektionslinien bei 30 Wochen deutlich über dem
jenigen der Kontrollinie. Es nimmt während der Legeperiode
jedoch weniger an Gewicht zu und demzufolge sinken der Ei
klaranteil und der Proteingehalt stärker, währenddessen beim
Eidotterantei 1 sowie beim Trockensubstanzgehal t eine grös
sere Zunahme festgestellt wurde. Die Selektionslinien schei
nen zu Beginn der Legeperiode ein Eigewichtsniveau erreicht
zu haben , welches sie nicht mehr in gleichem Ausmass stei
gern konnten wie in der Generation F2. Somit veränderte sich
die Eizusammensetzung der Selektionslinien stärker gegenüber

93
der Kontrollinie und die Abweichungen zur Kontrollinie waren
bei 50 Wochen geringer als bei 30 Wochen.
Weil anders gesagt die Kontrollinie mit den tieferen Eige
wichten zu Legebeginn Eier mit höheren Eidotteranteilen und
damit höherem Energie- und Trockensubstanzgehal t legte als
die Selektionslinien, und sie ihr Eigewicht während der Le
geperiode mehr steigerten, nahmen der Anteil Eidotter sowie
der Trockensubstanzgehalt weniger zu. Der Anteil Eiklar und
der Proteingehalt sanken dementsprechend weniger. Dies er
klärt die geringere Differenz zwischen Selektionslinien und
Kontrollinien bei 50 Wochen gegenüber 30 Wochen.
Die Untersuchungen in beiden Perioden haben gezeigt, dass
das Eigewicht bei 30 Wochen mit demjenigen bei 50 Wochen
hoch korreliert ist (0.95). Ebenfalls im Bereich von 0.93 -
0.99 liegen die Korrelationen der verschiedenen Eizusammen
setzungsmerkmale bei 30 Wochen und ihren entsprechenden
Merkmalen bei 50 Wochen. Etwas weniger eng sind die Be-
ziehungen zwischen den
Wochen und den jeweils
Eizusammensetzungsmerkmalen bei 30
verschiedenen Merkmalen bei 50 Wo-
eben. Am tiefsten erwiesen sich die Beziehungen zwischen dem
Eigewicht bei 30 Wochen und seiner Zusammensetzung bei 50
Wochen.
Die engsten Korrelationen bestehen zwischen Eigewicht bei 30
Wochen und dem Eiklaranteil bei 50 Wochen gefolgt vom Eidot
teranteil, dem Trockensubstanzgehalt, dem Energiegehalt und
dem Proteingehalt. Das Eigewicht ist bei 50 Wochen stark von
demjenigen bei 30 Wochen abhängig, seine Zusammensetzung
wird mit zunehmendem Alter jedoch immer stärker vom Alters
einfluss gesteuert. Wird durch indirekte Selektion die Ei
grösse beeinflusst, so wirkt sich das auch auf seine Ge
wichtsveränderung im Laufe einer Legeperiode aus, und seine
Zusammensetzung wird dadurch beeinflusst. Es scheint, dass
sich mit zunehmenden Selektionsgenerationen und zunehmendem
Alter die Protein- und Energiegehalte im Ei immer weniger
stark verändern und dass die Gewichtszunahme im Gesamtei

94
wahrscheinlich auf eine höhere Wassereinlagerung zurückzu
führen ist. Pym et al. (1979) haben in ihren Untersuchungen
auf bessere Futterverwertung bei Mastgeflügel gezeigt, dass
schlechtere Futterverwerter fettreichere Gewebe aufweisen
als die besseren Futterverwerter, welche mehr Protein und
Wasser ins Gewebe einlagern. Da mit der Selektion auf "Eier
lös minus Futterkasten" auch die Futterverwertung verbessert
wurde , kann eine Verschiebung in der Eizusammensetzung zu
stande gekommen sein.
Die berechneten Heritabilitäten sind hoch und lassen den
Schluss zu, dass in Zukunft mit einem weiteren Anstieg des
Eigewichtes gerechnet werden kann. Es wird sich möglicher
weise innerhalb einer Legeperiode auf Grund seines hohen
Anfangsgewichtes immer weniger stark verändern, was die
Differenz in seiner Zusammensetzung gegenüber der Kontrol-
1 inie immer kleiner werden lassen wird.
Die Regressionen des Futterverbrauches auf die Eimasse haben
gezeigt, dass diese mit zunehmendem Selektionszyklus in der
zweiten Periode geringer werden. Die Regression des Futter
verbrauches auf die Gewichtszunahme schwankt sehr stark. Die
Regression auf die Dottermasse zeigt ebenfalls, dass diese
eine wichtige Ursache des Futterverbrauches ist und zwar so
wohl in der ersten wie auch in der zweiten Periode beider
Generationen. Obwohl sich der Energiegehalt vor allem auf
die Dottermasse konzentriert ist, veränderten sich die Re
gressionen des Futterverbrauches auf den Energiegehalt
nicht. Der Proteingehalt, Bestandtei 1 sowohl des Eikl ares
wie auch des Eidotters, erklärt einen wichtigen Teil des
Futterverbrauches.
4.3. Schlupfergebnisse
Untersuchungen von Scott (1941) und Hilfiker (1970) haben
darauf hingewiesen, dass zu grosse oder zu kleine Eier eine
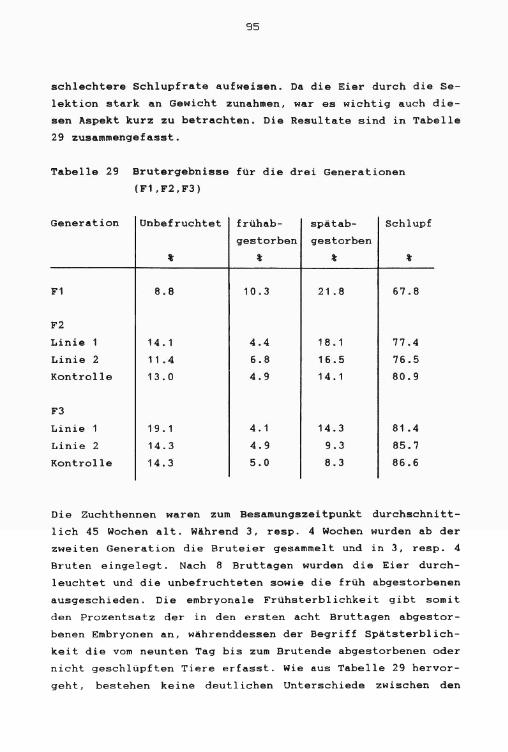
95
schlechtere Schlupfrate aufweisen. Da die Eier durch die Se
lektion stark an Gewicht zunahmen, war es wichtig auch die
sen Aspekt kurz zu betrachten. Die Resultate sind in Tabelle
29 zusammengefasst.
Tabelle 29 Brutergebnisse für die drei Generationen
(F1,F2,F3)
Generation Unbefruchtet früh ab- spätab- Schlupf
gestorben gestorben
t t t t
F1 8.8 10.3 21. 8 67.8
F2
Linie 14. 1 4.4 18. 1 77.4
Linie 2 11. 4 6.8 16.5 76.5
Kontrolle 13. 0 4.9 14. 1 80.9
F3
Linie 1 9 . 1 4 . 1 14.3 81 . 4
Linie 2 14.3 4.9 9.3 85.7
Kontrolle 14.3 5.0 8.3 86.6
Die Zuchthennen waren zum Besamungszei tpunkt durchschnitt-
1 ich 45 Wochen alt. Während 3, resp. 4 Wochen wurden ab der
zweiten Generation die Bruteier gesammelt und in 3, resp. 4
Bruten eingelegt. Nach 8 Bruttagen wurden die Eier durch
leuchtet und die unbefruchteten sowie die früh abgestorbenen
ausgeschieden . Die embryonale Frühsterblichkeit gibt somit
den Prozentsatz der in den ersten acht Bruttagen abgestor
benen Embryonen an, währenddessen der Begriff Spätsterblich
keit die vom neunten Tag bis zum Brutende abgestorbenen oder
nicht geschlüpften Tiere erfasst. Wie aus Tabelle 29 hervor
geht, bestehen keine deutlichen Unterschiede zwischen den

95
Selektionslinien und der Kontrollinie in den zwei selek
tierten Generationen in Bezug auf unbefruchtete Eier oder
frühabgestorbene Embryonen. Im Prozentsatz spätabgestorbener
und demzufolge auch in der Schlupfrate sind jedoch Unter
schiede zu erkennen. In den selektierten Generationen sind
die Schlupfergebnisse für die Kontrollinie am besten. Die
Embryonen der selektierten Linien stammten aus durchschnitt
lich schwereren Eiern und schienen erst gegen Ende der Brut
in Schwierigkeiten zu geraten oder konnten bis zum 22. Tag
nicht schlüpfen. Diese Resultate bestätigen somit die
zitierten Untersuchungen. Die höhere Sterblichkeit bei der
F1 kann möglicherweise auf Transporteinflüsse von Deutsch
land in die Schweiz zurückgeführt werden .

97
5. Zusammenfassung
Das Ziel dieser Arbeit war die Untersuchung des Selektions
kriteriums "Eierlös minus Futterkosten" zwischen der 21. und
der 40 . Woche als Mittel zur wirtschaftliche Verbesserung
der Eiproduktion in der Geflügelhaltung. Der Eierlös wurde
auf Grund der in der Selektionsperiode produzierten Eier be
rechnet . Die Futterkosten beziehen sich auf das in der glei
chen Periode verzehrte Futter, multipliziert mit dem Futter
preis. Zusätzlich wurden die Auswirkungen der Selektion auf
korrelierte biologische und wirtschaftliche Merkmale er
fasst. Die Erhebungen des individuellen Futterverbrauches,
der Körpergewichte, der Eizahl und der Eigewichte von drei
Generationen zu je ca. 1200 Hennen verteilt auf 2 Selek
tionslinien (2 x ca. 480) und Kontrollinie (ca. 240),
standen zur Verfügung. Die Untersuchungen der Eizusammen
setzung wurden an einer Stichprobe zu je ca. 250 zufällig
ausgelesenen Tieren in allen drei Generationen im Alter von
30 Wochen und in den Generationen F2 und F3 zusätzlich noch
im Alter von 50 Wochen durchgeführt.
Die Untersuchungen führten zu folgenden Hauptergebnissen:
1. Das Selektionskriterium (21. - 40. Woche) lag in der F2
für beide selektierten Linien im Durchschnitt um 3.3 t
über demjenigen der Kontrollinie und in der Generation
F3 um 6.9 t. ~ie "analoge Differenz" (41. - 60. Woche)
der Selektionslinien lag in der Generation F2 2.4 t und
in der Generation F3 7.1 t über derjenigen der Kontrol
linie.
2. Die korrelierten Selektionserfolge betrugen in der
ersten Periode für die Merkmale Eimasse 0.06 kg in der
F2 und 0. 22 kg in der F3. Der Futterverbrauch der F2
lag -0.15 kg unter demjenigen der Kontrollinie und in

98
der F3 0 . 07 kg darüber. Die Futterverwertung der Se
lektionsline war in der F2 um -0.04 kg und in der F3 um
-0.05 kg besser als in der Kontrollinie.
3. In der zweiten Periode lag die produzierte Eimasse der
Selektionslinie für die Generation F2 um 0.04 kg und in
der F3 um 0. 34 kg über derjenigen der entsprechenden
Kontrollinie. Der Futterverbrauch der Selektionslinie
der F2 lag -0.15 kg unter demjenigen der Kontrollinie.
In der F3 lag er 0.23 kg über demjenigen der Kontrol
linie. Die Futterverwertung war -0.03 kg besser in der
F2 und -0.07 kg in der F3.
4. Die Heritabilitäten verschiedener Merkmale und deren
genetischen Korrelationen (in Klammern) mit dem Selek
tionsmerkmal betrugen: Eimasse: 0 . 24 (0.89); Eizahl:
0.23 (0.35); Eigewicht: 0.99 (0 . 74); Futterverbrauch:
0.40 (0 . 09); Futterverwertung: 0.09 (-0.92);
Körpergewicht: 0.58 (0.09).
5. Die Heritabilität des Selektionsmerkmales "Eierlös
minus Futterkosten" betrug 0.23 und für die "analoge
Differenz" 0.27.
6. Die Befiederungsbewertung auf Grund einer Befiederungs
klassierung von 1 - 4 , war nicht signifikant schlechter
für die Selektionslinien. Die durchschnittlichen Werte
über alle Linien zeigten, dass Hennen mit der schlech
testen Befiederung (4) in der ersten Periode der Gene
rationen F1, F2, resp. F3, 0.42 kg, 0.37 kg resp. 0.51
kg mehr Futter benötigten. In der zweiten Periode be
nötigten die am schlechtesten befiederten Hennen 1 . 0,
resp. 0.67 kg mehr Futter.
7. Die Wiederholbarkeiten der Merkmale Eigewicht, Eiklar,
Eidotter, und Eischale wurden mit Daten von drei Eiern
pro Henne berechnet. In der ersten Periode lagen sie
zwischen 0.74 und 0.79. Sie waren etwas tiefer in der
zweiten Periode bei 50 Wochen.
8. Positiva phänotypische Korrelationen fanden sich einer
seits zwischen den Merkmalen Eigewicht und · Eiklaranteil

99
oder Proteingehalt und andererseits zwischen dem Eidot
teranteil und dem Energiegehalt oder dem Trockensub
stanzgehal t. Die Merkmale dieser beiden Gruppen sind
untereinander negativ korreliert.
9. Mit zunehmendem Alter sanken die Korrelationen zwischen
dem Eigewicht und den übrigen Merkmalen, während die
Beziehungen zwischen den übrigen Merkmalen unter sich
enger wurden.
1 0. Die korrelierten Selekt ionserfolge waren positiv für
die Merkmale Eigewicht, Eiklaranteil und Proteingehalt
und negativ für die Merkmale Eidotterantei 1, Energie
und Trockensubstanzgehalt. Sie wichen in der ersten
Periode der dritten Generation bereits signifikant ab.
Von der Generation F2 zur Generation F3 stiegen sie für
das Eigewicht von 0.50 g auf 2.32 g, für den Eiklaran
teil von 0.14 t auf 1.11 t, und für den Proteingehalt
von 0.47 t auf 0.81 t und veränderten sich für den Dot
tergehalt von -0.34 t auf -0.99 t, für den Energiege
halt von -0.13 MJ/kg auf -0 . 16 MJ/kg und für den Trok
kensubstanzgehalt von -0.13 t auf -0.59 t.
11. Zwischen 30 und 50 Wochen stiegen in der F2 resp. der
F3 das Eigewicht um 11 t, resp. 14 t, der Eidotteran
teil um 8 t resp. um 11 t, der Energiegehalt um 2 t
resp . um 3 t, und der Trockensubstanzgehal t um 4 t
resp. um 6 t. Hingegen sanken der Anteil Eiklar vom 3 t
resp. um 4 t und der Proteingehalt um 5 t resp. um 7 t.
Die Energie blieb unverändert.
12 . Die gewichteten genetischen Korrelationen zwischen den
untersuchten Merkmalen Eigewicht, Eiklar , Protein, Ei
dotter, Energie und Trockensubstanz waren in beiden
Perioden hoch (0.51 0.99). Sie waren am geringsten
zwischen dem Merkmal Eigewicht und allen übrigen. Mit
zunehmendem Alter wurden sie höher, mit Ausnahme der
jenigen zwischen dem Eigewicht und den übrigen, wo es
sich gerade umgekehrt verhielt.

100
13. Die Heritabilitäten lagen für die Merkmale der Eizusam
mensetzung zwischen 0.40 und 0.75 in der ersten Periode
und zwischen 0.36 und 0.85 in der zweiten Periode.
14. Die partiellen Regressionskoeffizienten des Futterver-
brauches auf verschiedene Merkmale in den drei Genera-
tionen (F1, F2, F3) und Perioden ( 21. - 40. Woche . 1 ' 41. - 60. Woche = 2) waren:
Generation F1 F2 F3
Periode 1 2 2
Körpergewicht kg 3.28 3.51 3.43 3.53 3 .13
Körpergewichtsver
änderung kg 2.66 2.26 0.44 1. 48 2.04
Eimasse kg .68 .85 .73 .84 .62
Energie MJ/kg . 11 . 08 .09 .10 . 1 0
Dottermasse kg 1. 76 1. 24 1 . 1 7 1. 48 1.56
Klarmasse kg .45 .86 .69 .75 .42
Proteinmasse kg 1 . 60 6.34 3.41 4.43 1. 70
15. Zwischen dem Selektionskriterium, resp. der "analogen
Differenz" und den Merkmalen Eigewicht, Eiklaranteil
und Proteingehalt bestanden positive Korrelationen.
Negative Korrelationen wurden zwischen dem Selektions
kriterium, resp. der "analogen Differenz" und den Merk
malen Eidotter, Energie und Trockensubstanz gefunden.
16. Die Schlupfergebnisse der Kontroll inie waren leicht
besser als in den Selektionslinien.

101
6. Summary
The objective of this thesis was to explore the selection
criterion "income minus feed cost between the 21st and 40th
week as a mean for economic improvement of egg production in
poultry breeding. The income from egg production is
calculated as a function cf number of eggs produced during
the selection period. Feed costs were calculated as the
amount of feed consumed during the same period, multiplied
by the feed price. Effects of the selection on correlated
biological and economic traits were also investigated.
Reccrds of individual feed consumption, body weight, number
of eggs and egg weight from three generations of approx.
1200 hens in two selection lines (2 x 480) and cne ccntrol
line ( 240) were available. The investigations of egg
composition were made with random samples cf 250 hens at 30
weeks of age in each of the three generations and in the
second and third generation also at 50 weeks.
The main results were :
1. The selection criterion ( 21. - 40. week) of the two
selection lines was 3.3 t above the control line in
generation F2 and 6.9 t in generation F3. The
"analogous difference" (41. 60 . week) of the
selection lines was 2.4 t above the control line in F2
and 7 . 3 t in F3.
2. The correlated selection responses in egg mass were 0.06
kg in generation F2 and 0.22 kg in generation F3. Feed
consumption of selection lines was -0.15 kg in F2 and
0 . 07 kg in F3. Feed conversion in the selection 1 ines
was improved by -0. 04 kg in F2 and -0. 05 kg in F3
compared to the control line.

102
3. In the second period (41. - 60 week) the egg mass of the
selection 1 ines was 0. 04 kg above the control line in
F2 and 0.34 kg in F3. Feed consumption of the selection
lines was -0.15 kg in F2 and 0 . 23 in F3 compared to the
control line . Feed conversion improved by -0. 03 kg in
F2 and by -0.07 kg in F3.
4. The heritabilities of different traits and their genetic
correlations (in parentheses) with the selection
criterion were: egg mass: 0.24 (0.89); egg number: 0.23
(0.35); egg weight: 0.99 (0.74); feed consumption: 0.40
(0.09); feed conversion : 0.09 ( - 0.92); body weight:0.58
( 0. 09).
5. The her i tabi 1 i ty for the selection criterion "income
minus feed cost" (21. - 40 week) was 0.23 and for the
criterion "analogous difference" (41. - 60. week) 0.27.
6. The mean feathering score , on a scale of 1 - 4 was
poorer in the selected lines, but not significantly.
Pooled data from all lines indicated that hens with
higher scores (poorer feathering) required 0.42 kg,
0 .3 7 kg and 0.51 kg more feed in the first period in
generations 1,2,3 respectively. In the second period cf
F2 and F3, 1.0 kg and 0.67 kg more feed was consumed by
the poorer feathered hens.
7 . The repeatabilities of the traits egg weight, egg
albumen , egg yolk and egg shell were calculated from
measures of three eggs per hen. In the f irst period
they were between 0.74 and 0.79. They were lower in the
second period at the age of 50 weeks.
8. Positive phenotypic correlations were found between the
traits egg weight and percentage cf albumen or protein,
as wel 1 as between percentage of yolk and energy or

103
percentage of dry matter. These two groups of trai ts
were negatively correlated.
9. The correlations between egg weight and the other traits
decreased with increasing age whereas the correlations
among the other traits increased.
10. The correlated selection responses were positive for the
trai ts egg weight, percentage of albumen and protein
content and negative for the traits percentage of yolk,
energy and percentage of dry matter. The response was
signif icant
generation.
in the first period of the third
Selection responses increased from ge-
neration F2 to generation F3 for egg weight from 0.50 g
to 2.32 g; from 0 .14 t to 1 . 11 t for albumen; from
0.47 t to 0.81 t for protein. They decreased from
-0.34 t to -0.99 t for yolk, from -0 .13 MJ/kg to -0.16
MJ/kg for energy and from -0 .13 t to 0.59 t for dry
matter.
11 . The egg weight increased between 30 and 50 weeks by 11 t
in F2 and 14 t in F3; the percentage of yolk increased
by 8 t in F2 and by 11 t in F3; the dry matter by 4 t
in F2 and by 6 t in F3. The percentage of albumen
decreased by 3 t in F2 and by 4 t in F3 and the
percentage of protein by 5 t in F2 and by 7 t in F3.
Energy remained unchanged.
12. The weighted genetic correlations between the traits egg
weight, albumen, protein, yolk, energie and dry matter
were high in both periods (0.51 0.99). The lowest
correlations were found between egg weight and the
others. All genetic correlations increased with
increasing age except f or those between egg weight and
the others which decreased.

104
13. The heritabilities for the egg composition traits were
between 0.40 and 0.75 in the first period and between
0.36 and 0.85 in the second period.
14. The partial regression coefficients of feed consumption
on the other traits in the three generations (F1, F2,
F3) and periods (21. - 40 week • 1 ; 41 - 60 week • 2)
were:
Generation F1 F2 F3
period 1 2 2
body weight kg 3.28 3.51 3.43 3.53 3 .13
body weight change kg 2.66 2.26 0.44 1 .48 2 . 04
egg mass kg .68 .85 .73 .84 . 62
energy MJ/kg . 11 .08 . 09 .10 . 10
yolk mass kg 1. 76 1 .24 1 . 17 1 .48 1. 56
albumen mass kg .45 .86 . 69 .75 .42
protein mass kg 1 . 60 6.34 3.41 4.43 1. 70
15. Positive genet ic c orrelations were found between the
selection criterio n, o r the analogous difference and
the traits egg weight, albumen an protein. They were
negative between the selection criterion, or the
analogous difference and the traits yolk, energy and
dry matter .
16. The percentage of eggs hatched was slightly greater for
the control line compared to the selection lines .

105
7. Literaturverzeichnis
Abplanalp, H., Peterson, S.J ., Okamoto, S. and Napolitano , D. (1984): Heterosis recombination effects and genetic variability of eqq composition in inbred lines of white Leghorns and - their crosses. Br. Poult . Sei., 25: 361-367.
Akbar, M.K., Gavora, J.S. and Fairfull, R. W. (1985): Relationship between activity at a young age and feed efficiency in chickens . Poult. Sei. , 64: 1402-1404.
Ambrosen, T. and Rotenberg, S. (1981): External and internal quality and chemical composition of hen eggs as related to hen age and selection for production traits. Acta Agric. Scand., 31: 139-152.
Anderson, G.B., Bolten, W., Jones, R.M. and Draper, M.H. (1978): Effect of age of the laying hen on the composition of the egg. Br. Poult. Sei., 19: 741 -745.
Arboleda, C.R., Harris, D. L. and Nordskog, A.W. (1976): Efficiency of selection in layer type chickens by using supplementary information on feed consumption. II Application to net income. Theor. Appl . Genet., 48: 75-83.
Bentsen , H.B. (1983): Genetic variation in feed efficiency of laying hens at constant body weight and egg production. I., II. Agric. Scand., 33: 289-319.
Bobr, L.W. and Sheldon, B . L . (1977): Analysis of ovulationoviposition patterns in the domestic fowl by telemetry measurements of deep body temperature. Aust. J. Biol. Sei., 30: 243-257.
Bordas , A. and Merat, P. (1974): Variabilite genetique et correlations phenotypiques caracterisant la consommation alimentaire de poules pondeuses apres correction pour le poids corporel et la ponte. Ann. genet. sel. anim., 6: 369-379.
Bordas, A. and Merat, P. (1981): Genetic variation and phenotypic correlations of food consumption of laying hens corrected for body weight and production. Br. Poult. Sei ., 22: 25-33 .
Bordas, A. and Merat, P. ( 1984): Reponses 1 iees dans une experience de selection sur la consommation alimentaire "residuelle" de coqs et poules Rhede Island. Proc. XVII World's Poult. Congress, Helsinki, Aug. 8 - 12.
Brody, S. (1945): Bioenergetics and growth: 371. zitiert nach Pym.
Burkhardt, A.D. ((1971): Qualitätseigenschaften von Flüssigei. Dissertation Hohenheim, Nr. 55.
Byerly, T.C., Kessler, J.W., Gous, R.M. and Thomas O.P. (1980): Feed requirements for egg production. Poult. Sei., 59: 2500-2507 .
Cahaner, A., Woodars, A.E. and Abplanalp, H (1979): Genetic parameters of egg production , body weight and egg size in the redlegged partridge. Br. Poult. Sei., 20: 541-549 .

105
Charles, D.R. (1976): The economic importance of feathering. Gleadthorpe experimental husbandry farm poul try booklet: 25-27. Agricul tural development and advisory service, England. zitiert nach Damme.
Charles, D. R. (1984): A model of egg production. Br. Poult. Sei., 25: 309-321.
Chung, R. A. and Stadelman W.J., (1965): A study of variations in the structure of the hen's egg. Br. Poult. Sei., 6: 277-282.
Chwalibog, A. (1985): Studies of energy metabolism in laying hens. Beretning fra. No. 578.
Clark, T. B. (1940): The relation of production and egg weight to age in white Leghorn fowls. Poult. Sei., 19: 61-66.
Conson, M.L. (1986): Zur Gefiederbeschaffenheit unterschiedlich aufgezogener Legehybriden. DGS, 23: 679-683.
Cunningham, F.E. Cotteril, O.J. and Funk, E.M. (1960): The effect of season and age of bird. 1. on egg size, quality and yield. Poult. Sei., 39:289-299.
Damme, K. ( 1984): Genetische und phänotypische Beziehungen zwischen Produktionsmerkmalen und dem Energiestoffwechsel von Legehennen. Dissertation an der Universität Hohenheim.
Damme, K. Pirchner, F. (1984): Genetische Unterschiede in der Befiederung von Legehennen und Beziehungen zu Produktionsmerkmalen. Arch . Geflügelk., 6/48: 215-222.
Dickerson, G. E. (1976): Growth and productivity. Meat animals: 449. zitiert nach James.
Fairfull, R.W., and Chambers, J.R. (1984): Breeding for feeding efficiency: poultry. Can. J. Amim. Sei., 64: 513-527.
Falconer , D.S. (1984): Einführung in die Quantitative Genetik. Deutsche Obersetzung.
Fletcher, D.L. , Britton, W.M., Rahn, A.P. and Savage, S.I. (1981): The influence of layer fleck age on egg compone~t yields and solids content. Poult. Sei., 60: 983-987 .
Frankham , R. (1972): Changes in feed consumption and gross feed efficiency of lines selected for part-record egg production. Poult. Sei., 50: 1194-1196.
Gonyou, H.W. and Morrison, W.D. (1983): Effects of feathering and insulative jackets on production laying hens at low temperature. Br. Poultry Sei., 311-317.
deby
24:
Grashorn, M. und Fleck , D.K. (1987): Genetisch-statistische Untersuchungen des Befiederungszustandes an weissen ( LSL) und braunen ( LB) Legehybr iden, Lehmann Information, Dezember 1987.
Hagger, C. (1977): Phänotypische und genetische Untersuchungen zur Futterverwertung der Legehenne. Dissertation ETH, Zürich, 5887.

Hagger, C. und Abplanalp, records for the genetic costs in laying flocks Sei., 19: 651-667.
107
H. (1978): Food consumption improvement of income over f eed of whi te Leghorns. Br. Poul t.
Harris, D.L. (1970): Breeding for efficiency in livestock production: Defining economic objectives. J. anim. Sei., 30: 860-865.
Harvey, W.R. (1976): Mixed model least squares and maximum likelihood computer program LSM-76. Ohio State University, Columbia.
Heil, G. (1976): Schätzung der genetischen Parameter für die Futterverwertung von Legehennen in Gruppenkäfigen und ihre Beziehungen zu anderen Leistungs- und Eiqualitätsmerkmalen. Dissertation No. 153, Techn. Universität, München.
Heil, G. (1984): Zur Eignung des Verhältnisses Futterverbrauch:Eimasse oder Eimasse:Futterverbrauch für quantitativ genetische Untersuchungen. Arch. Geflügelk., 48: 66-69.
Heil, G. and Pirchner F. (1979): Selection on feed efficiency in lay hens: direct and correlated response. EAAP, Harrogate, England.
Heil, G. and Hartmann, W. (1980): feed wastage in strains of crossbred hens from Leghorn lines selected for egg production and feed eff iciency. Europ. Geflügelkonferenz Hamburg, Vol. II: 147- 155.
Hilfiker, J. (1970): Beeinflussung einer Geflügelpopulation durch die Selektion auf hohes resp. kleines Eigewicht bei Legebeginn. Dissertation ETH, Zürich, 4567.
Hill, W.G. (1971): Theoretical aspects of crossbreeding. Ann. Genet. sel. anim., 3: 32-34.
Hill, W.G. (1972): Estimation of genetic change. I. General theory and design of control populations. Animal breeding abstracts Vol. 40, No. 1: 1-15.
Hill, A.T., Krueger, W.F. and Quisenberry, J.H. (1966): A biometrical evaluation of component parts of eggs and their relationship to other economically important traits in a strain of white Leghorns. Poult. Sei., 45: 1162-1185.
Hou, Y.H., Harpal, S. and Nordskog, A.W. (1985): Improving feed efficiency in laying hens using individual feed records in a selection index. Poult. Sei., 64: 118.
Hughes, B.0.(1980): Feather damage in hens caged individually. Br. Poult. Sei., 21: 149-154.
Hurnik, J.F., Summers, J.D., Reinhart, B.S. and Swierczewska, E.M. (1977):Effect of age on the performance of laying hens during the first year of production. Poult. Sei., 56: 222-230.
Jaffe, W. P. (1965): The relationships between egg weight and yolk weight. Br. Poult. Sei., 5: 295-298.
James, J.W. (1982): Economic aspects of developing breeding objectives: General considerations. Future Developments in the genetic improvement of animals, Part 5 Economic aspects of Developing breeding objectives: 107-118.

108
Katle, J., Bentsen , H.B. and Braastad, B. O. (1984): Correlated traits with residual feed consumption. Proc. XVII World's Poult. Congress, Helsinki.
Kirchgessner, M. and Voreck, 0. (1980): Zum Einfluss unterschiedlicher Energie- und Proteinversorgung auf die chemische Zusammensetzung von Hühnereiern. Arch. Geflügelk., 44: 89-94.
Krax, H. (1974): Geflügelproduktion. Paul Parey Verlag, Hamburg und Berlin.
Lee, B.D. , Morrison, W.P. and Leeson , S. (1983): Effects of feather cover and insulative jackets on metabolic rate of laying hens . Poult. Sei., 62: 1129-1132.
Leeson, S. and Morrison, W.D. (1978): Effect of feather cover on feed efficiency in laying birds. Poult. Sei., 57: 1094-1096 .
Lerner, I.M. and Cruden D. (1951): The heritability of egg weight, the advantages of mass selection and of early measurements. Poultry Sei., 30: 34-41.
Marion , W.W. Nordskog, A.W., Tolman , H.S. and Forsythe, R.H. (1964): Egg composition as influenced by breeding, egg size, age and season. Poult. Sei., 43: 255-264.
Marion, J.E., Woodroof, J . G. and Cook, R.E. (1965) Some physical and chemical properties of eggs from hens of five different stocks. Poult. Sei., 44: 529-535 .
May, K.N. and Stadelman, W. J. (1960): Some factors affecting components of eggs from adult hens. Paul t. Sei., 39: 560-565.
Mahner, A. (1983) : Eierproduktion und Eiqualit~t. Handbuch der Geflügelphysioligie, Teil II. Karger Verlag.
Merat, P. et Bordas, A. (1974): Consommation alimentaire d'animaux a plumage reduit (gene Na) au normal en presence au en 1 'absence du gene de nanisme dw. Ann . Genet. Sel. anim. , 6(1): 17-28.
Nagai, J. and Gowe, R.S. (1969): genetic control of quality. 1. Sources of variation. 2. Selection maximum rate of improvement. Br. Paul t. Sei., 10: 350, 351-358.
egg for
337-
Nordskog , A. W. , French, H. , Arbol eda C. R and Casey D. W. (1972): Breeding for efficiency of egg production. World's Poult. Sei. J., 29: 175-188.
O'Neill, S., Balnave, D. and Jackson, N. (1971): The influence of feathering and environmental temperature on the heat production and efficiency of utilization of metabolizable energy by the mature cockerl. J. Agr. Sei., Camb. 77: 293-305. zitiert nach Chwalibog.
Pauw, R. (1986): Züchterische Beeinflussung der Futterausnutzung. DGS 19: 555-559.
Peterson , C.F. and Lampaun, C.E. (1949): Relationship of egg size during f irst year of production to percentage increase in second year. Poult. Sei., 28: 302. zitiert nach Mehner.
Pirchner , F. (1980): Feed efficiency. 6. Europ . Geflügelkonferenz Hamburg, Vol. I:,29-42.

10 9
Pirchner , F. ( 1985): Genetics of efficiency of food conversion for egg production. Poultry genetics and breeding . Edit . by W.G. Hill , J . M. Manson and D. Hewitt. Br. Poult . Sei. Ltd.
Pi r chner F. und Heil, G. (1978) : Züchterische Bearbeitung der Futterverwertung bei Legehennen . Aktuelle Themen der Tierernährung und Veredlungswirtschaft: 159-164. zitiert nach Damme .
Pym, R. A.E . (1982): Selection results for efficiency of food utilisation in livestock and poultry. 2nd World Congress of genetics, Madrid, Vol V: 231-244.
Pym, R . A.E.and Nicholls, P.J. (1979): Selection for food conversion in broilers: direct and correlated responses to selection for body weight gain, food consumption and food conversion ratio. Br. Poult. Sei. , 20 : 73-86.
Pym , R . A. E . and Solvyns, A. J . (1979): Selection for food conversion in Broilers: Body composition of birds selected for increased body- weight gain , food consumption and food conversion ratio. Br . Poult. Sei ., 20: 87-97.
Rodda , D. D., Friars , G.W., Gavora , J . S. and Merritt, E.S. ( 1977) : Genetic parameter estimates and strain comparisons of egg compositional traits . Br. Poult. Sei., 18: 459-473 .
Romanoff , A. L. and Romanoff A.J. (1949): The avian egg. Wiley and sons. zit i ert nach Bordas.
Sainz, F. , Gonzalez, M. , Roca, P . and Alemany, M. (1983) : Physical and chemical nature of eggs from six breeds of domestic fowl. Br. Poult. Sei., 24: 301-309.
Scheinberg, S . L. Ward, H. and Nordskog, A.W. (1953): Breeding for egg quality. 1. Heritability and repeatability of egg weight and its components. Poult . Sei., 32: 504-509 .
Scott , H. M. and Warren, D. C. (1941): The relation of total weight and the weight of the component parts of the egg to hatching power. Poultry Sei . , 20: 75-78.
Sibbald, I.R. (1979): The gross energy of avian eggs. Poultry Sei . , 58: 404-409.
Simon, D.L . (1976): Bestimmungsgründe für Einkommensunterschiede aus der Legehennenhal tung. Agrarwirtschaft 16 : 48-60. zitiert nach Krax.
Skala , J .H. and Swanson, M. H. ( 1962): Studies of variation in initial quality of chicken eggs. 1. Physical measurements of albumen and yolks . Poult. Sei., 41: 1533-1536.
Tasaki, I. and Sasa, Y. (1970): Energy metabolism in laying hens. Energy metabolism of farm animals . EAAP Pub. 13: 197-200 .
Tauson , R. and Svensson , S.A. (1980): Influence of plumage condition on the hen's feed requirement. Swedish J. agric. res ., 10: 35-39.
Tauson, R. , Ambrosen, T . and Elwinger , K. (1984): Evaluation of procedures for scoring the integument of laying hens, independent scoring of plumage condition. Acta agric. Scand ., 34: 400-408.

110
Tawfik, E. S., Horst, P. and Petersen, J. (1976) Untersuchungen an Legehennen über genetische Fundierung und Beziehung von Legeleistung, Legereife Körpergewicht und Kriterien der Eibeschaffenheit. 1. Mitteilung: genetische Fundierung. Arch. Geflügelk., 40: 181-187.
Tawfik, E. S., Horst, P. and Peterson, J. (1981) Untersuchungen an Legehennen über genetische Fundierung und Beziehungen von Legeleistung, Legereife Körpergewicht und Kriterien der Eibeschaffenheit. 2. Mitteilung: phänotpische und genetische Beziehungen. Arch. Geflügelk., 45: 166-175.
Tullet, S.G., Macleod, M.G. and Jewitt, T.R. (1980): The effect of partial defeathering an energy metabolism in the laying fowl. Br. Poult. Sei., 21: 241-245.
Van Tijen, W. F. and Kuit, A. R. (1970): The heritability cf characteristics cf egg quality, their mutual correlation and the relationship with productivity. Arch. Geflügelk., 6: 201-210.
Varadarajulu, P. and Cunningham, F.E. (1972): A study cf selected characteristics of hen's egg yolk. 3. Influence cf breed, strain and age of bird . Poult. Sei., 51: 1327-1331.
Von dem Hagen, D. und Bielesch, M. (1986): Leukoseviren bei Legehybriden. DGS, 22: 651-652.
Wegner, R.M. (1978): Untersuchungen zur Leistung von Legehennen in verschiedenen Haltungssystemen und Käfigformen. DGS, 33: 834-838.
Wing, T.L. and Nordskog, A.W. (1982): Use cf individual feed records in a selection program for egg production efficiency. I. Heritability cf the residual component cf feed efficiency. Poult. Sei., 61: 226-230.

111
Anhang
Tabelle 1 : Legeleistung der Hennen in den drei Generationen
Gene- Alter bei Ei zahl Ei zahl Eigewicht Eigewicht ration Legebeginn 21.-40. 41 . -60. 30 Wochen 50 Wochen Linie
F1 137 . 7 120.9 106.7 59 . 3 67. 1
F2 L1 134 . 7 120.5 105.8 57 . 7 65.0 L2 135 . 1 120.4 106 . 2 57.6 65. 1 K 135 . 1 120 . 7 106.2 57.1 64 . 7
"' 135.0 120. 5 106. 1 57.5 65.0
F3 L1 141 . 4 117 . 0 107.2 59.5 67.5 L2 144 . 1 117. 0 108.8 59.4 67.4 K 142.2 116 . 0 104.8 57.8 66 . 3
"' 142.8 117. 0 107 . 0 58 . 9 67 . 0
Tabelle 2: Körpergewichte der Hennen der drei Generationen ( F1, F2, F3), der Selektionslinien ( L1, L2) und der Kontrollinie (K)
Generationen Linien
Gewichte F1 F2 F3
L1 L2 K L1 L2 K
20 Wochen 2050 2010 2007 2037 2019 2014 2022
30 Wochen 2191 2204 2215 2253 2144 2157 2169
40 Woc hen 2365 2337 2349 2398 2313 2320 2339
50 Wochen 2447 2429 2446 2510 2425 2446 2472
60 Wochen 2532 2517 2538 2615 2548 2574 2607
Differenzen 20-40 13 l 14 l 15 l 15 l 13 l 13 l 14 l 40 - 60 7 l 7 l 7 l 8 l 9 l 1 0 l 1 0 •

Name und Vorname:
Geboren:
Heimatort:
1960 - 19'70:
19'70 - 19'73:
19'73 - 19'74:
19'74 - 19'79:
seit 19'79:
Marguerat-König Catherine
23 . 11.53 in Genf
Lutry, Forel , Bern
Primarschule und Sekundarschule in Bern:
Gymnasium in Neuenburg
Abschluss: Maturität Typ B
Handels-Fachschule Dr. Rischik, Bern
Abschluss: Diplom als Chefsekretärin
Landwirtschaftliches Studium an der ETH
Zürich, Fachrichtung Tierproduktion
Abschluss: Diplom als Ingenieur Agronom
Unterricht an der Landwirtschaftlichen Schule
Grangeneuve, Freiburg
Teilzeitassistentin in der Gruppe Tierzucht des
Institutes für Nutztierwissenschaften der ETH Zürich