rna polymerase and transcription elongation factor … · rna polymerase and transcription...
TRANSCRIPT

RNA polymerase and transcription elongationfactor Spt4/5 complex structureBrianna J. Kleina,1, Daniel Boseb,1, Kevin J. Bakera, Zahirah M. Yusoffa, Xiaodong Zhangb,2, and Katsuhiko S. Murakamia,2
aDepartment of Biochemistry and Molecular Biology, Pennsylvania State University, University Park, PA 16802; and bDivision of Molecular Biosciences,Centre for Structural Biology, Department of Life Sciences, Faculty of Natural Sciences, Imperial College London, London SW7 2AZ, United Kingdom
Edited by* Roger D. Kornberg, Stanford University School of Medicine, Stanford, CA, and approved November 30, 2010 (received for review September15, 2010)
Spt4/5 in archaea and eukaryote and its bacterial homolog NusG isthe only elongation factor conserved in all three domains of lifeand plays many key roles in cotranscriptional regulation and inrecruiting other factors to the elongating RNA polymerase. Here,we present the crystal structure of Spt4/5 as well as the structureof RNA polymerase-Spt4/5 complex using cryoelectron microscopyreconstruction and single particle analysis. The Spt4/5 binds in themiddle of RNA polymerase claw and encloses the DNA, reminiscentof the DNA polymerase clamp and ring helicases. The transcriptionelongation complex model reveals that the Spt4/5 is an upstreamDNA holder and contacts the nontemplate DNA in the transcriptionbubble. These structures reveal that the cellular RNA polymerasesalso use a strategy of encircling DNA to enhance its processivityas commonly observed for many nucleic acid processing enzymesincluding DNA polymerases and helicases.
cryo-EM ∣ Spt4/5-DSIF-NusG ∣ X-ray crystallography
Transcription by RNA polymerase (RNAP) plays a central rolein gene expression, and this process is highly regulated at
many steps, including promoter recognition, transcription activa-tion, elongation, and termination. During transcription, severaltranscription elongation factors communicate with RNAP andregulate the velocity of RNA synthesis, transcriptional pausing,and termination (1). Recent genome-wide analyses of transcrip-tion elongation revealed the importance of transcription elonga-tion in the regulation of gene expression (2–4). For example, alarge fraction of transcribing eukaryotic RNAP II (Pol II) pausesnear promoters to facilitate rapid changes in gene expressionduring cell development.
Bacterial NusG is perhaps the best characterized transcriptionelongation factor in vivo and in vitro. Diverse functions of NusGhave been reported, e.g., Escherichia coli NusG reduces RNAPpausing and intrinsic termination (5, 6), whereas Bacillus subtilisand Thermus thermophilus NusG enhance pausing (7, 8). NusGconsists of the NusG amino-terminal (NGN) domain andKyprides–Onzonis–Woese (KOW) motif at the C-terminal do-main (hence also called KOW domain), and these two domainsfold independently and are connected by a flexible linker of 13amino acids (9, 10) (Fig. 1B and Fig. S1C). The NGN domain hasbeen assigned the function for regulating transcription elonga-tion. The KOW domain, on the other hand, plays important rolesin interacting with other proteins. For example, it contacts theelongation factor NusE, which is also the ribosomal S10 subunit.This interaction is essential for forming rRNA and λ gene anti-termination complexes, as well as for the coupling of transcrip-tion and translation. Furthermore, the KOW contacts Rho fortranscription termination (9, 11).
Eukaryotic Spt5 is the functional and structural counterpart ofbacterial NusG. In addition to the NGN and KOW domains, eu-karyotic Spt5 contains additional motifs including the N-terminalacidic region, four to five additional KOWmotifs, and C-terminalrepeats that are targets of regulatory kinases (12) (Fig. S1A). Eu-karyotic Spt5 associates with Spt4 to form a Spt4/5 heterodimer,which is also called the DRB (5,6-dichloro-1-β-D-ribofuranosyl-
benzimidazole) sensitivity-inducing factor (DSIF) in highereukaryotes (13). The Spt4/5 complex plays important roles inregulating transcription elongation both positively (14) and nega-tively (15).
Structural and biochemical studies of the archaeal RNAPtranscription system revealed that it is an excellent model fordissecting the molecular basis of eukaryotic transcription, as wellas providing unique data that may unify basic transcription me-chanisms across all three domains of life (16–18). The archaealtranscription apparatus, which includes RNAP and general tran-scription and elongation factors, is similar to the eukaryoticsystem. Archaeal Spt5 has only two domains, NGN and KOW,similar to the bacterial NusG, but it forms a heterodimer withSpt4 like the eukaryotic Spt4/5 complex (19) (Fig. S1). To under-stand how Spt4/5 regulates transcription elongation, we deter-mined the crystal structure of archaeal Spt4/5 and the complexstructure of RNAP-Spt4/5 using cryoelectron microscopy andsingle particle analysis.
Results and DiscussionX-ray Crystal Structure of Pyrococcus furiosus Spt4/5. Crystal struc-tures of archaeal Spt4/5 from Methanococcus jannaschii (19) andeukaryotic Spt4/5 from yeast (12) and human (20) revealed ahighly conserved overall structure and architecture of the Spt4and Spt5 dimerization surface. However, as these structurescontain only the NGN of Spt5, the orientation of the KOW re-lative to NGN remains unknown. Here we have determined thecrystal structure of the complete P. furiosus Spt4/5 at 1.8-Å reso-lution (Table S1). The P. furiosus Spt4 and Spt5 were producedindependently in E. coli and purified as an Spt4/5 complex tohomogeneity as determined by SDS-PAGE. The molecularweights of His6-tagged Spt4 and Spt5 are 11 and 17 kDa, respec-tively, and a native molecular mass of 25 kDa determined by sizeexclusion column chromatography indicates that Spt4/5 exist as aheterodimer in solution. The crystal structure of P. furiosus Spt4/5was determined by molecular replacement, and refinementstatistics are detailed in Table S1.
The overall structures of P. furiosus Spt4 and Spt5-NGN as wellas the dimerization interface are similar to their homologuesfrom M. jannaschii (19), yeast (12), and human (20) (Fig. 1 andFig. S1). P. furiosus Spt4 consists of a NGN-binding domain
Author contributions: X.Z. and K.S.M. designed research; B.J.K., D.B., and Z.M.Y.performed research; B.J.K., D.B., X.Z., and K.S.M. analyzed data; K.J.B. contributed newreagents/analytic tools; and X.Z. and K.S.M. wrote the paper.
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.
Data deposition: Coordinate and structure factor have been deposited in the ProteinData Bank, www.pdb.org (PDB ID code 3P8B). The cryo-EM map corresponding to theRNAP-Spt4/5 complex has been deposited in the EMDataBank, www.emdatabank.org(EMD ID code 1840).1B.J.K. and D.B. contributed equally to this work.2To whom correspondence may be addressed. E-mail: [email protected] [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1013828108/-/DCSupplemental.
546–550 ∣ PNAS ∣ January 11, 2011 ∣ vol. 108 ∣ no. 2 www.pnas.org/cgi/doi/10.1073/pnas.1013828108
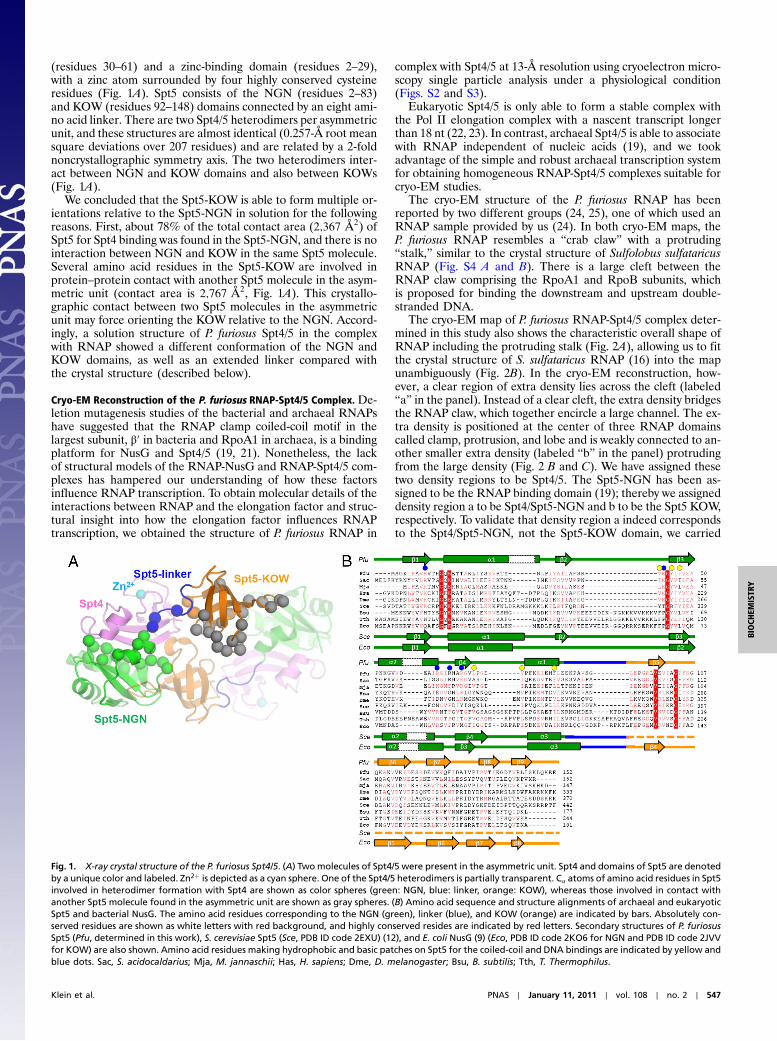
(residues 30–61) and a zinc-binding domain (residues 2–29),with a zinc atom surrounded by four highly conserved cysteineresidues (Fig. 1A). Spt5 consists of the NGN (residues 2–83)and KOW (residues 92–148) domains connected by an eight ami-no acid linker. There are two Spt4/5 heterodimers per asymmetricunit, and these structures are almost identical (0.257-Å root meansquare deviations over 207 residues) and are related by a 2-foldnoncrystallographic symmetry axis. The two heterodimers inter-act between NGN and KOW domains and also between KOWs(Fig. 1A).
We concluded that the Spt5-KOW is able to form multiple or-ientations relative to the Spt5-NGN in solution for the followingreasons. First, about 78% of the total contact area (2;367 Å2) ofSpt5 for Spt4 binding was found in the Spt5-NGN, and there is nointeraction between NGN and KOW in the same Spt5 molecule.Several amino acid residues in the Spt5-KOW are involved inprotein–protein contact with another Spt5 molecule in the asym-metric unit (contact area is 2;767 Å2, Fig. 1A). This crystallo-graphic contact between two Spt5 molecules in the asymmetricunit may force orienting the KOW relative to the NGN. Accord-ingly, a solution structure of P. furiosus Spt4/5 in the complexwith RNAP showed a different conformation of the NGN andKOW domains, as well as an extended linker compared withthe crystal structure (described below).
Cryo-EM Reconstruction of the P. furiosus RNAP-Spt4/5 Complex. De-letion mutagenesis studies of the bacterial and archaeal RNAPshave suggested that the RNAP clamp coiled-coil motif in thelargest subunit, β′ in bacteria and RpoA1 in archaea, is a bindingplatform for NusG and Spt4/5 (19, 21). Nonetheless, the lackof structural models of the RNAP-NusG and RNAP-Spt4/5 com-plexes has hampered our understanding of how these factorsinfluence RNAP transcription. To obtain molecular details of theinteractions between RNAP and the elongation factor and struc-tural insight into how the elongation factor influences RNAPtranscription, we obtained the structure of P. furiosus RNAP in
complex with Spt4/5 at 13-Å resolution using cryoelectron micro-scopy single particle analysis under a physiological condition(Figs. S2 and S3).
Eukaryotic Spt4/5 is only able to form a stable complex withthe Pol II elongation complex with a nascent transcript longerthan 18 nt (22, 23). In contrast, archaeal Spt4/5 is able to associatewith RNAP independent of nucleic acids (19), and we tookadvantage of the simple and robust archaeal transcription systemfor obtaining homogeneous RNAP-Spt4/5 complexes suitable forcryo-EM studies.
The cryo-EM structure of the P. furiosus RNAP has beenreported by two different groups (24, 25), one of which used anRNAP sample provided by us (24). In both cryo-EM maps, theP. furiosus RNAP resembles a “crab claw” with a protruding“stalk,” similar to the crystal structure of Sulfolobus sulfataricusRNAP (Fig. S4 A and B). There is a large cleft between theRNAP claw comprising the RpoA1 and RpoB subunits, whichis proposed for binding the downstream and upstream double-stranded DNA.
The cryo-EM map of P. furiosus RNAP-Spt4/5 complex deter-mined in this study also shows the characteristic overall shape ofRNAP including the protruding stalk (Fig. 2A), allowing us to fitthe crystal structure of S. sulfataricus RNAP (16) into the mapunambiguously (Fig. 2B). In the cryo-EM reconstruction, how-ever, a clear region of extra density lies across the cleft (labeled“a” in the panel). Instead of a clear cleft, the extra density bridgesthe RNAP claw, which together encircle a large channel. The ex-tra density is positioned at the center of three RNAP domainscalled clamp, protrusion, and lobe and is weakly connected to an-other smaller extra density (labeled “b” in the panel) protrudingfrom the large density (Fig. 2 B and C). We have assigned thesetwo density regions to be Spt4/5. The Spt5-NGN has been as-signed to be the RNAP binding domain (19); thereby we assigneddensity region a to be Spt4/Spt5-NGN and b to be the Spt5 KOW,respectively. To validate that density region a indeed correspondsto the Spt4/Spt5-NGN, not the Spt5-KOW domain, we carried
Fig. 1. X-ray crystal structure of the P. furiosus Spt4/5. (A) Twomolecules of Spt4/5 were present in the asymmetric unit. Spt4 and domains of Spt5 are denotedby a unique color and labeled. Zn2þ is depicted as a cyan sphere. One of the Spt4/5 heterodimers is partially transparent. Cα atoms of amino acid residues in Spt5involved in heterodimer formation with Spt4 are shown as color spheres (green: NGN, blue: linker, orange: KOW), whereas those involved in contact withanother Spt5 molecule found in the asymmetric unit are shown as gray spheres. (B) Amino acid sequence and structure alignments of archaeal and eukaryoticSpt5 and bacterial NusG. The amino acid residues corresponding to the NGN (green), linker (blue), and KOW (orange) are indicated by bars. Absolutely con-served residues are shown as white letters with red background, and highly conserved resides are indicated by red letters. Secondary structures of P. furiosusSpt5 (Pfu, determined in this work), S. cerevisiae Spt5 (Sce, PDB ID code 2EXU) (12), and E. coli NusG (9) (Eco, PDB ID code 2KO6 for NGN and PDB ID code 2JVVfor KOW) are also shown. Amino acid residues making hydrophobic and basic patches on Spt5 for the coiled-coil and DNA bindings are indicated by yellow andblue dots. Sac, S. acidocaldarius; Mja, M. jannaschii; Has, H. sapiens; Dme, D. melanogaster; Bsu, B. subtilis; Tth, T. Thermophilus.
Klein et al. PNAS ∣ January 11, 2011 ∣ vol. 108 ∣ no. 2 ∣ 547
BIOCH
EMISTR
Y

out antibody labeling experiments against the 6xHis-tag at theN terminus of Spt4 as shown in Fig. 3. Briefly, we imaged the anti-body labeled RNAP-Spt4/5 complex using negative stained EM.The labeled particles were picked and aligned to the cryo-EMreconstruction of the complex. Euler angles were assigned tothese images by projection matching (Fig. 3 A–D). Comparisonswith the reprojections of the cryo-EM reconstructions alongthe same angles (Fig. 3 B and C) show a good correspondencebetween the particles and the reprojections. Additional densityregions could be observed that have a characteristic Y shape ofthe IgGs (Fig. 3 C and D). The antibodies are clearly colocalizedwith the density a, which we assigned to be the Spt4/5-NGN do-main (as indicated by magenta arrows), and have no overlap withdensity b (as indicated by orange arrows), which we have assignedto be the KOW domain.
Next, we carried out rigid body fitting of the X-ray structure ofP. furiosus Spt4/5 determined in this study (Fig. 2C), which
showed good agreement between the crystal structure and thedensity shape. We also carried out energy minimization calcula-tion for modeling the complex of RNAP coiled-coil motif andSpt4/5 (Fig. 2D). The hydrophobic patch of Spt5-NGN, includingY45 and F47, faces the tip of the coiled coil of the RpoA1 subunitas suggested by site-directed and deletion mutagenesis studies(19). In addition to the main interactions, our cryo-EM structureidentified an additional previously undescribed contact site; α1and α2 of Spt5-NGN fit into a cleft between the lobe and theprotrusion (Fig. 2C). There is no interaction observed betweenRNAP and Spt4 in our model. This observation is in good agree-ment with biochemical data that the Spt5-NGN is the sole factorto form a complex with RNAP (19). The Spt5-KOW fits nicelyinto density b (Fig. 2C), outside the main RNAP density. Tofit the Spt5-KOW into the cryo-EM map, the KOW was rotatedabout 90 ° toward the upstream DNA side starting from the crys-tal structure. No steric clash between Spt4 and the Spt5-KOWnorSpt4 and the Spt5-linker is observed for this KOW fitting. A bind-ing site of bacterial transcription elongation factor RfaH, which isa paralog of E. coli NusG, was mapped on the coiled coil ofRNAP, and the docking model of RNAP transcription elongationcomplex with RfaH was constructed (21). A location and orien-tation of RfaH relative to the coiled coil in the docking model aresimilar to those of the Spt5-NGN in the cryo-EM RNAP-Spt4/5complex structure.
Modeling of the Transcription Elongation Complex with Spt4/5. TheDNA binding channel of multisubunit RNAP is positioned be-tween the RNAP claw, and the space of the channel is modulatedby the position of the RNAP clamp. The crystal structures of tran-scription elongation complexes consisting of RNAP and theDNA/RNA hybrid, from T. thermophilus (26) and Pol II (27),showed that the clamp is in the closed position in the transcribingRNAP, but the channel is still partially open, perhaps explainingthe property for transcription to pause in the absence of elonga-tion factors. The cryo-EM reconstruction of the RNAP-Spt4/5
lobeprot.
jaw
lobe
prot.
A
clamp
jawprot.
wall
D/L
clamp
E/FE/F
Front90°Top
ba
ba
B
b
Front
b
Top
lobeprot.
jawV742 3
DC
ccclamp
2 1Y45
V42
F47L70 F82
R9 D267R41 Q254
1
rudder
KOW
Fig. 2. Surface representations of the P. furiosus RNAP-Spt4/5 complex re-construction. (A) Two different views of the RNAP-Spt4/5 complex reconstruc-tion: from the top (looking from above the upstream DNA binding channel)and from the front (looking into the active site cleft, downstream and up-stream DNAs are positioned on left and right, respectively). Positions of D/Land E/F subunits as well as RNAP domains (“prot”, protrusion) are indicated.(B) Partially transparent maps of A are shown with the S. sulfataricus RNAPcrystal structure fitted into the cryo-EM maps. RpoA1 and RpoA2, dark gray;RpoB, light brown; other RNAP subunits, light gray. Positions of extra den-sities assigned for the Spt4/Spt5-NGN (a, green) and Spt5-KOW (b, orange)are indicated. (C) A magnified view of the interface between RNAP andSpt4/5. This view is the same as a boxed area in B. RNAP and Spt4/5 are shownas surface and cartoon models, respectively. The cryo-EM map is shown asmesh. Spt5-NGN, green; Spt5-linker, blue; Spt5-KOW, orange; Spt4, violet.(D) The Spt5-NGN and coiled-coil interaction. The cartoon models of Spt5-NGN (green), Spt5-linker (blue), and Spt4 (violet) are shown with partiallytransparent surface view. The RNAP coiled coil is shown as a gray tube. Hy-drophobic residues of Spt5 involved in the coiled-coil interaction are depictedas sticks and colored yellow. Cα positions of the three hydrophobic residues(P261, L263, and I264) in the coiled coil are indicated as spheres.
Fig. 3. IgG labeling of Spt4 in the RNAP-Spt4/5 complex. Monoclonalanti-poly-Histidine IgG labeled particles were aligned to the cryo-EM recon-struction of the complex and angles assigned by projection matching.(A) Three-dimensional surface rendering of cryoreconstruction viewed fromthe assigned angles. Purple arrows indicate the location of Spt4/5-NGNdomain from the fitted model. Orange arrows indicate the location of theSpt5-KOW domain. (B) Reprojections from the 3D reconstruction alongthe assigned Euler angles. (C) Raw negatively stained immunocomplexes.White circles indicate the IgG density. Red outline shows the position ofreprojected particle from B. (D) Same as C with arrows, showing the positionof Spt4/5-NGN (magenta) and Spt5-KOW (orange). IgG density is also indi-cated (blue).
548 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1013828108 Klein et al.

complex in this study revealed that Spt4/5 binds between theRNAP claw, thereby completely encircling the DNA bindingchannel. This closure would prevent DNA from disengaging fromthe transcribing RNAP and may provide an explanation as to whySpt4/5 is able to enhance transcription processivity. Completeclosure of the DNA channel is reminiscent of the DNA slidingclamp in the DNA replication fork, the β-clamp in bacteria,and PCNA in eukaryotes, which enhances the processivity ofDNA polymerase (28). The enclosure of DNA by proteins is alsofound in many processive ring helicases such as RuvB, E1, LargeT-antigen, and MCM (minichromosome maintenance) (29).
In the cryo-EM reconstruction of the RNAP-Spt4/5 complex,the diameter of the channel for the downstream DNA is muchwider than that for upstream DNA (Fig. 2A). To obtain furtherstructural insight into how the transcription bubble is accom-modated in the RNAP-Spt4/5 complex and also how Spt4/5 influ-ences RNAP transcription, the DNA/RNA scaffold was modeledinto the RNAP-Spt4/5 complex. Although many crystal structuresof eukaryotic and bacterial elongation complexes have beendetermined, none of them shows a clear location for the nontem-plate DNA in the transcription bubble nor the upstream double-stranded DNA. Recent single-molecule FRET experimentsconstructed the complete elongation complex model of yeastPol II (30), and we have used this DNA/RNA scaffold for mod-eling the archaeal elongation complex. The yeast elongation com-plex model is overlaid with the archaeal RNAP using secondarystructure matching. The downstream DNA fits perfectly into theDNA binding channel whereas the upstream DNA had to betilted ∼30° toward the RNAP wall to avoid steric clash with thedensity corresponding to Spt4/5 (Fig. 4A). It is important to notethat the position of the upstream DNA has not been determined
structurally. Only a small region of the upstream DNA is pro-tected by RNAP in the DNA footprinting experiment, and alimited amount of DNA cross-linking was observed in this area(31). The upstream DNA in the elongation complex is thereforeintrinsically flexible and may occupy multiple positions duringtranscription elongation. Spt4/5 in the elongation complex is ide-ally positioned to function as an “upstream DNA holder” andmay be able to hold the upstream DNA in a confined location.
A basic region of Spt5-NGN is located near the nontemplateDNA within the transcription bubble (Fig. 4B). In this basicpatch, R9, K43, and R67 are highly conserved in archaea, butR61 and R64 are less conserved (Fig. 1B). Some of these residuesare conserved in eukaryotic Spt5 and bacterial NusG, but inter-estingly, each residue is conserved only in either eukaryote orbacteria; e.g., R9/K43/R67 and R61 are conserved in only eukar-yote and bacteria, respectively. The different amino acids in thisregion may explain their unique functions only found in particulardomains of life. The close proximity of the Spt5-NGN and thenontemplate DNA within the transcription bubble in the modelis able to explain the result whereby B. subtilis NusG contactsthe nontemplate DNA for stabilizing transcriptional pausing (7).
In addition to the interaction with proteins, the KOW motifhas been shown to bind DNA and RNA (32). In the model ofthe transcription elongation complex with Spt4/5 (Fig. 4A), theSpt5-KOW locates near the upstream DNA, which allows it tocontact DNA. A recent photo-cross-linking experiment with theDrosophila Pol II transcription elongation complex in the pre-sence of Spt4/5 has shown that the Spt5 subunit contacts a nas-cent transcript if the RNA length is longer than 18 nt (22). In ourmodel (Fig. 4A), the Spt5-KOW locates far from the 16 nt RNA,which just begins to emerge from the surface of RNAP suggestingthat eukaryotic Spt5 uses its second to fifth KOW motifs but notthe first KOW motif (Fig. S1A) for contacting nascent RNA inthe transcription elongation complex.
Concluding RemarksA recent single-molecule study of E. coli RNAP transcriptionrevealed that NusG enhances the processivity of transcriptionby increasing the pause-free speed of RNAP elongation andsuppressing the entry into long-lifetime pauses (6). In this study,the cryo-EM structure of the RNAP-Spt4/5 complex revealedthat the elongation factor encircles the DNA, similar to the DNAclamp in DNA polymerase, to promote processivity. Further-more, Spt4/5 is able to contact two different positions of DNAin the transcribing RNAP that include the upstream DNA andthe single-stranded nontemplate DNA (Fig. 4). The restrictionof the upstream DNA by Spt4/5 may facilitate (i) ejecting thetranscribed DNA template from the DNA channel and (ii) rean-nealing between the template and nontemplate DNA strands atthe upstream end of the transcription bubble (Fig. 4), promotingthe RNAP toward its posttranslocated state. The interactions be-tween the nontemplate DNA in the transcription bubble and thebasic patch of Spt5-NGN could prevent DNA secondary structureformation in the transcription bubble. Combinations of theseeffects by the transcription elongation factor would prevent ele-mental and backtracked pauses of RNAP transcription therebyimproving the processivity of transcription (1, 6). The interestingremaining questions are which interaction affects what aspect oftranscription and whether one interaction influences only oneset of parameters? The structure of archaeal RNAP-Spt4/5 com-plex presented in this study provides a structural framework forfurther biochemical and biophysical studies to answer thesequestions.
Materials and MethodsProtein Preparations and X-Ray Crystal Structure Determination. (i) Cloning ofP. furiosus Spt4 and Spt5 genes, (ii) Spt4 and Spt5 expressions and purificationof Spt4/5 complex, (iii) crystallization and determination of the X-ray crystal
A
u-DNAjaw
NGNKOW
cc
lobe prot.
wall
D/L
jaw
AC
d-DNA
E/F
clamp
d-DNA
u-DNA
NGN
KOW
70°
bubble
Front
B
Side
KOW
cc
5’
d-DNA
u-DNA
Spt4/Spt5-NGN
Basic region(R9, K43, R61, R64 and R67)
p p
Fig. 4. Model of the P. furiosus RNAP transcription elongation complexwith Spt4/5. (A) Two different views—front and side—of the RNAP cryo-EMreconstruction with the DNA/RNA model (template DNA, cyan; nontemplateDNA, black; RNA, red). Positions of D/L and E/F subunits, RNAP domains andSpt4/5 domains (Spt4/Spt5-NGN, NGN; Spt5-KOW, KOW) are indicated. Loca-tions of downstream (d-DNA) and upstream (u-DNA) double-stranded DNAsare indicated. A map of the side view was sliced to show a transcription bub-ble and the active site (AC) inside the DNA binding channel. (B) The interac-tion between Spt4/5 and nucleic acid in the transcribing RNAP. A magnifiedview showing only DNA/RNA, Spt4/Spt5-NGN, and coiled coil (cc) of clamp. Anorientation of this view is the same as A, Right. The surface of Spt4/5 is colorcoded according to electrostatic surface potential (negative, red; neutral,white; positive, blue).
Klein et al. PNAS ∣ January 11, 2011 ∣ vol. 108 ∣ no. 2 ∣ 549
BIOCH
EMISTR
Y

structure of P. furiosus Spt4/5, and (iv) P. furiosus RNAP purification and theRNAP-Spt4/5 complex formation are described in SI Text.
Cryo-Electron Microscopy. P. furiosus RNAP-Spt4/5 complex (2 μL of ∼0.1 mg∕mL) was applied to glow-discharged 300 mesh holey carbon grids (Quanti-foil). Excess sample was blotted and flash frozen in liquid ethane cooledby liquid nitrogen (−183 °C) to form vitreous ice. Grids were prepared undercontrolled environmental conditions (100% humidity, 4 °C) using a vitrobot(FEI). Data were collected at 50;000× magnification using a Philips CM200field emission gun (FEG) electron microscope operating at 200 kV underlow-dose conditions (10 e−∕Å2), and over a range of nominal defoci (−0.5 μmto −3.5 μm). Images were collected directly on a 4k × 4k CCD camera (F415from Tietz Video and Imaging Processing GmbH), giving a pixel size of 1.76 Å(Fig. S2). (i) Image processing, (ii) IgG labeling of Spt4 in the RNAP-Spt4/5
complex, and (iii) fitting of the S. sulfataricus RNAP and the P. furiosusSpt4/5 X-ray crystal structures are described in SI Text.
ACKNOWLEDGMENTS. We thank the staff at F1 of the MacCEHSS (Macromo-lecular Diffraction at Cornell High Energy Synchrotron Source) for supportcrystallographic data collection. We thank P. Babitzke, D. S. Gilmour, andL. B. Rothman-Denes for critical readings of the manuscript. We thankI. Artsimovitch for sending a coordinate of the Tth TEC-RhaH complex model.Figures were prepared using PyMOL (http://pymol.sourceforge.net/) andUniversity of California, San Francisco (UCSF) Chimera (33). This work wassupported by National Institutes of Health (NIH) Grant GM087350 (to K.S.M.)and Wellcome Trust (to X.Z). Center for Hybrid and Embedded SoftwareSystems is supported by the National Science Foundation (NSF) and NIH/National Institute of General Medical Sciences via NSF Award DMR-0225180, and the MacCHESS resource is supported by NIH/National Centerfor Research Resources Award RR-01646.
1. Landick R (2006) The regulatory roles and mechanism of transcriptional pausing.Biochem Soc Trans 34:1062–1066.
2. Muse GW, et al. (2007) RNA polymerase is poised for activation across the genome.Nat Genet 39:1507–1511.
3. GuentherMG, Levine SS, Boyer LA, Jaenisch R, Young RA (2007) A chromatin landmarkand transcription initiation at most promoters in human cells. Cell 130:77–88.
4. Nechaev S, et al. (2010) Global analysis of short RNAs reveals widespread promoter-proximal stalling and arrestof Pol II in Drosophila. Science 327:335–338.
5. Burova E, Hung SC, Sagitov V, Stitt BL, Gottesman ME (1995) Escherichia coli NusGprotein stimulates transcription elongation rates in vivo and in vitro. J Bacteriol177:1388–1392.
6. Herbert KM, et al. (2010) E coli NusG inhibits backtracking and accelerates pause-freetranscription by promoting forward translocation of RNA polymerase. J Mol Biol399:17–30.
7. Yakhnin AV, Yakhnin H, Babitzke P (2008) Function of the Bacillus subtilis transcriptionelongation factor NusG in hairpin-dependent RNA polymerase pausing in the trpleader. Proc Natl Acad Sci USA 105:16131–16136.
8. Sevostyanova A, Artsimovitch I (2010) Functional analysis of Thermus thermophilustranscription factor NusG. Nucleic Acids Res 38:7432–7445.
9. Mooney RA, Schweimer K, Rosch P, Gottesman M, Landick R (2009) Two structurallyindependent domains of E coli NusG create regulatory plasticity via distinct interac-tions with RNA polymerase and regulators. J Mol Biol 391:341–358.
10. Knowlton JR, et al. (2003) A spring-loaded state of NusG in its functional cycle is sug-gested by X-ray crystallography and supported by site-directed mutants. Biochemistry42:2275–2281.
11. Burmann BM, et al. (2010) A NusE:NusG complex links transcription and translation.Science 328:501–504.
12. Guo M, et al. (2008) Core structure of the yeast Spt4-Spt5 complex: A conservedmodule for regulation of transcription elongation. Structure 16:1649–1658.
13. Wada T, et al. (1998) DSIF, a novel transcription elongation factor that regulates RNApolymerase II processivity, is composed of human Spt4 and Spt5 homologs. Genes Dev12:343–356.
14. Chen Y, et al. (2009) DSIF, the Paf1 complex, and Tat-SF1 have nonredundant,cooperative roles in RNA polymerase II elongation. Genes Dev 23:2765–2777.
15. Wu CH, et al. (2003) NELF and DSIF cause promoter proximal pausing on the hsp70promoter in Drosophila. Genes Dev 17:1402–1414.
16. Hirata A, Klein BJ, Murakami KS (2008) The X-ray crystal structure of RNA polymerasefrom Archaea. Nature 451:851–854.
17. Hirata A, Murakami KS (2009) Archaeal RNA polymerase. Curr Opin Struc Biol19:724–731.
18. Kostrewa D, et al. (2009) RNA polymerase II-TFIIB structure and mechanism oftranscription initiation. Nature 462:323–330.
19. Hirtreiter A, et al. (2010) Spt4/5 stimulates transcription elongation through the RNApolymerase clamp coiled-coil motif. Nucleic Acids Res 38:4040–4051.
20. Wenzel S, Martins BM, Rosch P, Wohrl BM (2010) Crystal structure of the human tran-scription elongation factor DSIF hSpt4 subunit in complex with the hSpt5 dimerizationinterface. Biochem J 425:373–380.
21. Belogurov GA, et al. (2007) Structural basis for converting a general transcriptionfactor into an operon-specific virulence regulator. Mol Cell 26:117–129.
22. Missra A, Gilmour DS (2010) Interactions between DSIF (DRB sensitivity inducingfactor), NELF (negative elongation factor), and the Drosophila RNA polymerase IItranscription elongation complex. Proc Natl Acad Sci USA 107:11301–11306.
23. Cheng B, Price DH (2008) Analysis of factor interactions with RNA polymerase IIelongation complexes using a new electrophoretic mobility shift assay. Nucleic AcidsRes 36:e135.
24. Carlo SD, Lin S-C, Taatjes DJ, Hoenger A (2010) Molecular basis of transcriptioninitiation in Archaea. Transcription 1:1–9.
25. Kusser AG, et al. (2008) Structure of an archaeal RNA polymerase. J Mol Biol376:303–307.
26. Vassylyev DG, Vassylyeva MN, Perederina A, Tahirov TH, Artsimovitch I (2007)Structural basis for transcription elongation by bacterial RNA polymerase. Nature448:157–162.
27. Gnatt AL, Cramer P, Fu J, Bushnell DA, Kornberg RD (2001) Structural basis oftranscription: an RNA polymerase II elongation complex at 3.3 Å resolution. Science292:1876–1882.
28. Pomerantz RT, O’Donnell M (2007) Replisome mechanics: insights into a twin DNApolymerase machine. Trends Microbiol 15:156–164.
29. Enemark EJ, Joshua-Tor L (2008) On helicases and other motor proteins. Curr OpinStruc Biol 18:243–257.
30. Andrecka J, et al. (2009) Nano positioning system reveals the course of upstream andnontemplate DNAwithin the RNA polymerase II elongation complex.Nucleic Acids Res37:5803–5809.
31. Korzheva N, et al. (2000) A structural model of transcription elongation. Science289:619–625.
32. Steiner T, Kaiser JT, Marinkovic S, Huber R, Wahl MC (2002) Crystal structures oftranscription factor NusG in light of its nucleic acid- and protein-binding activities.EMBO J 21:4641–4653.
33. Pettersen EF, et al. (2004) UCSF Chimera—a visualization system for exploratoryresearch and analysis. J Comput Chem 25:1605–1612.
550 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1013828108 Klein et al.