rnai%therapeu-c%targe-ng%% alpha21%an-trypsin%(aat
TRANSCRIPT

Targeting Innovation
RNAi Therapeu-c Targe-ng Alpha-‐1 An-trypsin (AAT) Deficiency
Liver Disease
Steven B. Kanner, Ph.D. 1

2
Disclosures
! Employee and shareholder of Arrowhead Research Corpora-on

3
! AAT (SERPINA1) is an abundant serum protein synthesized primarily in liver
! Inhibits neutrophil proteases during lung inflamma-on (par-cularly smoking injury)
! Z mutant (single aa subs-tu-on, E342K) common disease variant with low secre-on
that polymerizes in liver, not blood
Alpha-‐1-‐an-trypsin (AAT) deficiency disease T
arget

4
! AAT (SERPINA1) is an abundant serum protein synthesized primarily in liver
! Inhibits neutrophil proteases during lung inflamma-on (par-cularly smoking injury)
! Z mutant (single aa subs-tu-on, E342K) common disease variant with low secre-on
that polymerizes in liver, not blood
! Autosomal co-‐dominant inheritance: ~1/3000 births in US and EU
! Associated with liver disease in children/adults, and lung disease (COPD) in adults
Alpha-‐1-‐an-trypsin (AAT) deficiency disease
Incide
nce

5
! AAT (SERPINA1) is an abundant serum protein synthesized primarily in liver
! Inhibits neutrophil proteases during lung inflamma-on (par-cularly smoking injury)
! Z mutant (single aa subs-tu-on, E342K) common disease variant with low secre-on
that polymerizes in liver, not blood
! Autosomal co-‐dominant inheritance: ~1/3000 births in US and EU
! Associated with liver disease in children/adults, and lung disease (COPD) in adults
! Polymeriza-on of Z-‐AAT protein in liver leads to globule forma-on
! Proteolysis and autophagy likely mechanisms of globule degrada-on
! Globules induce hepatocyte apoptosis, fibrosis, cirrhosis and hepatocellular carcinoma
Alpha-‐1-‐an-trypsin (AAT) deficiency disease
Pathology

6
! AAT (SERPINA1) is an abundant serum protein synthesized primarily in liver
! Inhibits neutrophil proteases during lung inflamma-on (par-cularly smoking injury)
! Z mutant (single aa subs-tu-on, E342K) common disease variant with low secre-on
that polymerizes in liver, not blood
! Autosomal co-‐dominant inheritance: ~1/3000 births in US and EU
! Associated with liver disease in children/adults, and lung disease (COPD) in adults
! Polymeriza-on of Z-‐AAT protein in liver leads to globule forma-on
! Proteolysis and autophagy likely mechanisms of globule degrada-on
! Globules induce hepatocyte apoptosis, fibrosis, cirrhosis and hepatocellular carcinoma
! No specific therapy for liver disease other than transplant
! Lung disease treated with M-‐AAT protein replacement, with no benefit to liver
Alpha-‐1-‐an-trypsin (AAT) deficiency disease
Therap
y

Alpha-‐1-‐an-trypsin deficiency (AATD): A protein folding, gain-‐of-‐func-on disease ! 95-‐98% of the clinical disease from Z-‐
mutant homozygosity ! ZZ muta-on leads to less secreted
protein with lower ac-vity ! Polymer forma-on and reten-on in
hepatocyte ER
American Thoracic Society Documents 825
Figure 1. Mechanism of inhi-bition of neutrophil elastase bythe alpha-1 antitrypsin (AAT)inhibitor. Elastase attacks thereactive center loop (yellow) ofAAT with the active serine ofthe elastase (small red sidechain), forming a link to theamino acid at the base of thereactive center (small green sidechain) of AAT. The resultingcleavage of the reactive loopallows it to snap back into themain ! sheet (red ribbon witharrows) of the AAT. This spring-like movement flings the teth-ered elastase to the oppositeend of the AAT molecule, dis-torting its active site and alter-ing its structure so that it canbe destroyed. The center of theactive site of the AAT moleculeis Met358. When Met358 is oxi-dized to methionine sulfoxideor sulfone, the association rateconstant of the inhibition ofneutrophil elastase by AAT ismarkedly reduced. Oxidants inthe lower respiratory tract arisefrom cigarette smoke, environ-mental oxidants, as well as neu-trophils and macrophages.Modified by permission fromCarrell and Lomas (295).
fewer than 60% of individuals with severe AAT deficiency de-velop significant airflow limitation (6). This suggests that in manycases, AAT deficiency alone is not enough to induce emphysema(34). It has also been suggested that pulmonary emphysema devel-ops when elastin fiber repair mechanisms are overwhelmed by amassive attack of elastases from inflammatory reactions (35).
A major pathogenic factor is cigarette smoke, which containsoxidants capable of inactivating AAT by converting active siteMet358 to methionine sulfoxide, with the association constant forNE being reduced about 2,000-fold. In addition, it has beenshown that Z AAT inhibits NE at a slower rate than doesM AAT (4.5 versus 9 " 106 M–1 second–1) (36). Furthermore,AAT polymers can be detected in the bronchoalveolar lavagefluid, as demonstrated in two of five subjects with emphysemarelated to PI*ZZ AAT deficiency (37). Because polymerizationobscures the AAT reactive loop, the conformational transitionmay impair the inhibitory activity. Therefore, in AAT deficiency,oxidants contained in cigarette smoke may further impair a quan-titatively and qualitatively less functional AAT. In addition,cigarette smoke and proteinases may both work to impair lungelastin resynthesis in the animal model of elastase-induced em-physema (38).
Cigarette smoke also recruits inflammatory cells. In AATdeficiency, more neutrophils are found within air spaces than inemphysematous lungs of individuals with normal AAT plasmalevels. This contributes to a greater NE load (39). This phenome-non might be attributable to the presence of neutrophil chemo-tactic factors, mainly leukotriene B4 released from alveolar mac-rophages (40).
In addition, neutrophils and macrophages may release a vari-
ety of metalloproteinases with the potential to degrade extracel-lular matrix components (41). A role for a human collagenasein alveolar injury in experimental emphysema has also beendemonstrated (42). Metalloproteinases are not inhibited by AATand may even inactivate it by limited proteolysis near the activesite (41). The C-terminal fragments of AAT released duringproteolytic inactivation are potent neutrophil chemotactic fac-tors (43, 44).
LABORATORY TESTS
Observation of a reduced or absent #1-globulin band on routineplasma protein electrophoresis should arouse suspicion of AATdeficiency and should be confirmed quantitatively and qualita-tively (see Table 3).
TABLE 3. RANGE OF SERUM LEVELS* OF ALPHA-1ANTITRYPSIN ACCORDING TO PHENOTYPE
Phenotype
Units PI*MM PI*MZ PI*SS PI*SZ PI*ZZ
$M 20–48 17–33 15–33 8–16 2.5–7mg/dl 150–350 90–210 100–200 75–120 20–45
Data from references 14, 47, and 298.* Serum levels given are measured using a typical commercial standard (mg/dl)
and the purified standard ($M) used in the U.S. Registry. A level of less than11 $M is associated with an increased risk for emphysema.
Janciauskiene, S. et al. (2013) Acute Phase Proteins
Globule formation à fibrosis à cirrhosis à HCC
7

Therapeu-c ra-onale
Inhibit Z-‐AAT produc-on by silencing AAT gene: ! Prevent accumula-on of disease causing Z-‐AAT protein ! Allow clearance of accumulated Z-‐AAT protein ! Prevent repeated cycle of cellular damage and -ssue repair ! Reverse liver fibrosis
Feldmann, G., et al. (1975) Gut
PiZZ phenotype (diseased) Pi-null phenotype (normal)
8

ARC-‐AAT: An RNAi therapeu-c using Dynamic PolyConjugate (DPC) technology
Unlocked Nucleobase Analog (UNA)
ARC-‐EX1
NAG Targe-ng Ligand
MLP
RNAi Trigger Targe-ng AAT mRNA
CDM Masking
Cholesterol
MLP: melian-‐like pep-de CDM: carboxy-‐dimethylmaleamide NAG: N-‐acetyl-‐galactosamine
9

10
RNAi therapeu-cs using Dynamic PolyConjugate (DPC) technology
1. Rozema et al. (2007) Proc Natl Acad Sci USA 104:12982-‐12987 2. Wong et al. (2012) Nucleic Acid Ther 22:380-‐390 3. Wooddell et al. (2013) Mol Ther 21:973-‐985
DPC polymer and RNAi trigger aeach to their respec-ve cell surface targets
DPC polymer and RNAi trigger are internalized
DPC polymer and RNAi trigger are enclosed in endosomes. Low pH results in polymer unmasking
DPC polymer induces endosome disrup-on and release of RNAi trigger into cytoplasm
RNAi trigger engages RISC resul-ng in target knockdown

Screening for RNAi triggers
! 2,648 19-mer sequences identified, 840 human/cyno monkey cross-reactive, 46 selected for in vitro screening after application of specificity criteria
! EC50 values for highly potent UNA-containing sequences identified for ranking ! Screening of top Chol-RNAi triggers + ARC-EX1 in transgenic Z-AAT mice
Human liver cancer cell line
11
D ay
No
rma
liz
ed
AA
T i
n s
eru
m
0 7 1 4 2 1 2 8 3 5 4 20 .0 1
0 .1
1
S a lin e
8 m g /k g A R C -5 2 0 E x c + 8 m g /k g A D 0 0 2 5 4
8 m g /k g A R C -5 2 0 E x c + 8 m g /k g A D 0 0 3 6 9
8 m g /k g A R C -5 2 0 E x c + 8 m g /k g A D 0 0 3 7 0
8 m g /k g A R C -5 2 0 E x c + 8 m g /k g A D 0 0 2 9 3
8 m g /k g A R C -5 2 0 E x c + 8 m g /k g A D 0 0 2 9 5
D ay
No
rma
liz
ed
AA
T i
n s
eru
m
0 7 1 4 2 1 2 8 3 5 4 20 .0 1
0 .1
1
S a lin e
8 m g /k g A R C -5 2 0 E x c + 8 m g /k g A D 0 0 2 5 4
8 m g /k g A R C -5 2 0 E x c + 8 m g /k g A D 0 0 3 6 9
8 m g /k g A R C -5 2 0 E x c + 8 m g /k g A D 0 0 3 7 0
8 m g /k g A R C -5 2 0 E x c + 8 m g /k g A D 0 0 2 9 3
8 m g /k g A R C -5 2 0 E x c + 8 m g /k g A D 0 0 2 9 5
Z-AAT transgenic mice (PiZ)
Top leads
UNA Chol

12
RNAi trigger comparison in vitro
! UNA modifica-on confers lower mel-ng temperature
! UNA modifica-on confers measurable changes in potency in vitro
Lead series
RNAi trigger Version Tm EC50 in vitro (pM)Family A
254 Canonical 77.0 17.3369 UNA 59.9 17.9370 UNA 64.0 17.0
Family B143 Canonical 88.7 153.8305 UNA 73.8 >1000306 UNA 73.7 >1000
Family C145 Canonical 88.1 32.7307 UNA 79.9 32.0308 UNA 77.2 22.0
Family D378 Canonical 71.1 44.9379 UNA 60.8 57.4380 UNA 60.9 22.0

Incorpora-on of UNA results in improved profile Single dose activity in Z-AAT transgenic mice (PiZ)
! >90% knockdown using AAT-specific canonical (AD00254) or UNA trigger (AD00370) after a single dose
! Improved duration of effect with UNA-AAT 13
D a y
% h
AA
T i
n s
eru
m
(no
rma
liz
ed
to
pre
-do
se
)
0 1 0 2 0 3 0 4 00
2 0
4 0
6 0
8 0
1 0 0
1 2 0
1 4 0
S a l i n e
8 m g / k g c a n o n i c a l A A T - s i R N A
8 m g / k g A A T - U N A
D a y
% h
AA
T i
n s
eru
m
(no
rma
liz
ed
to
pre
-do
se
)
0 1 0 2 0 3 0 4 00
2 0
4 0
6 0
8 0
1 0 0
1 2 0
1 4 0
S a l i n e
8 m g / k g c a n o n i c a l A A T - s i R N A
8 m g / k g A A T - U N A
Saline AD00254 (canonical) AD00370 (UNA-‐AAT)

Incorpora-on of UNA results in improved profile Single dose activity in normal cynomolgus monkeys
! 80% knockdown using AAT-specific canonical or 90% knockdown with UNA-AAT trigger after a single dose
! Improved duration of effect with UNA-AAT 14
W e e k s
No
rma
liz
ed
AA
T i
n s
eru
m
0 1 2 3 4 5 6 70
2 0
4 0
6 0
8 0
1 0 0
1 2 0
6 m g / k g A A T - A D 0 0 3 7 0 + 3 m g / k g A R C - E X 1 ( n = 2 )
6 m g / k g A A T - A D 0 0 2 5 4 + 3 m g / k g A R C - E X 1 ( n = 2 )
UNA-‐AAT (N = 2) AD00254 (canonical, N = 2)

Reduc-on in serum Z-‐AAT levels correlates with reduced Z-‐AAT mRNA levels
Reduc-on in Z-‐AAT mRNA is prolonged, correla-ng with serum Z-‐AAT levels
! PiZ mouse livers evaluated for mRNA on indicated days aher a single UNA-‐AAT dose
15

Phosphoryla-on of RNAi trigger guide strand and RISC loading
16
Endosome
RISC
RISC
5’-‐Chol OH-‐5’
5’-‐Chol OH-‐5’
5’-‐Chol P-‐5’
Release
CLP-‐1 kinase P-‐5’
RISC loading
P-‐5’
AAAAAA
mRNA loading and cleavage
Hepatocyte
5’-‐Chol OH-‐5’
Guide strand
Internaliza-on
P-‐5’: 5’-‐phosphoryla-on

RNAi trigger in RISC exhibits stability
Total lysate
RISC-‐Associated
17
! Ago2 immunoprecipita-on captures 5’-‐phosphorylated guide strand detected in PNA assay ! PiZ mice treated with single dose of UNA-‐AAT
Homogenize liver (Total lysate)
Ago2 immunoprecipitation (IP)
Hybridize PNA probe for AD00370
Detect RNAi trigger guide strand by AEX/HPLC
Wash
Elute
3 8 1 5 2 2 2 91
1 0
1 0 0
1 0 0 0
D a y
ng
GS
siR
NA
/
g t
iss
ue
3 8 1 5 2 2 2 90 . 1
1
1 0
1 0 0
D a y
ng
GS
siR
NA
/
g t
iss
ue
UNA-AAT ng P-‐5’ G
S/g -ssue
ng P-‐5’ G
S/g -ssue

18
Mul-ple dosing profile in PiZ mice
Mul-ple dosing once every 2 or 3 weeks induces significant reduc-on in Z-‐AAT
Control RNAi trigger UNA-‐AAT (q2w) UNA-‐AAT (q3w)
q3w Dosing:

Mul-ple dosing profile in NHP
19
! Cynomolgus monkeys (N = 2) were dosed q6w with 6 mpk UNA-‐AAT + 3 mpk ARC-‐EX1
Mul-ple dosing once every 6 weeks induces significant reduc-on in monkey AAT
W e e k s
% A
AT
in
se
rum
(no
rma
liz
ed
to
pre
-do
se
)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 40
2 0
4 0
6 0
8 0
1 0 0
1 2 0Dose

AAT deficiency liver disease mouse model The transgenic PiZ mouse model expressing human Z-‐mutant AAT recapitulates the human AATD associated liver phenotype:
! Hepatocytes produce high levels of human Z-‐AAT ! Hepatocytes are not able to efficiently process and secrete Z-‐AAT from hepatocytes ! Z-‐AAT forms polymers that accumulate in large “globules” within the hepatocytes by 1 – 2 months of age ! Globules are visualized with Periodic Acid Schiff (PAS) staining + diastase
Rudnick, D.A. et al., (2004) Hepatology 20
PiZZ Wild Type

Baseline 5 weeks old
UNA-AAT 13 weeks old
Saline 13 weeks old
UNA-Control 13 weeks old
0 1 2 3 4 5 6 7 8 9 10 11 12 13
end
Week:
Dose:
21
Preven-on of globule forma-on in young PiZ mice
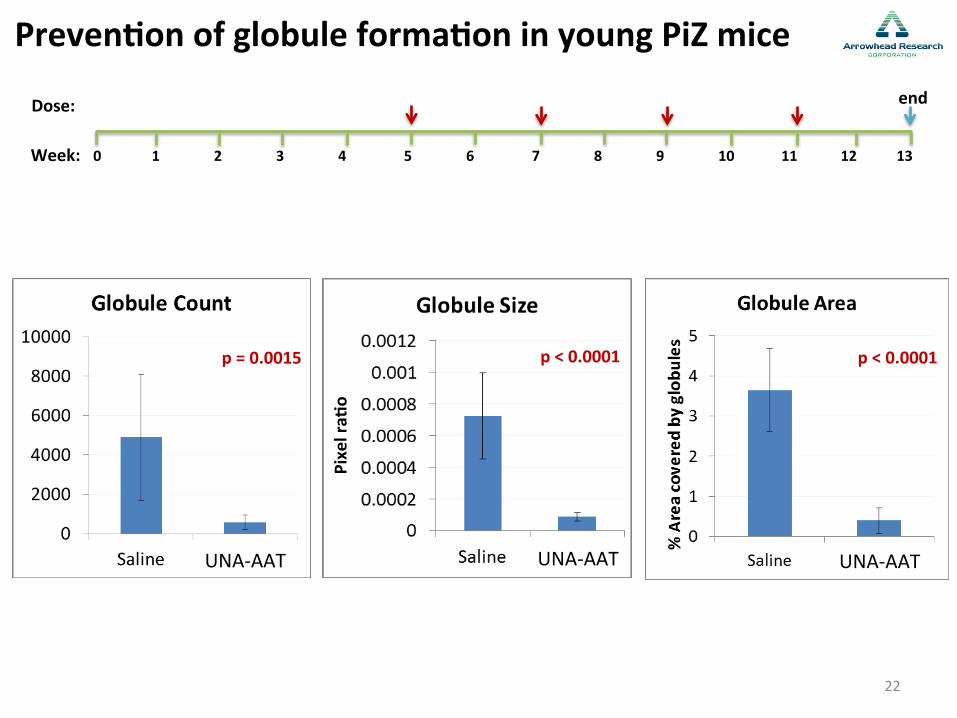
p < 0.0001 p = 0.0015 p < 0.0001
22
UNA-‐AAT UNA-‐AAT UNA-‐AAT
Pixel ra-
o
Preven-on of globule forma-on in young PiZ mice
0 1 2 3 4 5 6 7 8 9 10 11 12 13
end
Week:
Dose:

23
Preven-on of globule forma-on in young PiZ mice
0 1 2 3 4 5 6 7 8 9 10 11 12 13
end
Week:
Dose:
Western blot
Saline UNA-‐AAT
Soluble
Insoluble
UNA-‐AAT

Z-AAT serum protein and mRNA reduction after a single ARC-AAT dose in older PiZ mice
24
Single injection significantly reduces AAT mRNA and serum protein
S a line
C o n trol-U
N A
D a y 3
D a y 8
D a y 15
D a y 22
D a y 29
Re
lati
ve
hA
AT
mR
NA
Ex
pre
ss
ion
A A T -U N A
0%
25%
50%
75%
1 00 %
1 25 %
0 5 1 0 1 5 2 0 2 5 3 0D a y
Se
rum
Z-h
AA
T(n
orm
ali
zed
to
pre
-do
se
)
S a lin e
C o n tro l-U N A
A A T -U N A
0%
25%
50%
75%
100%
125%
150%
! 6 month old PiZ mice treated with a single dose
UNA-‐AAT
UNA-‐AAT
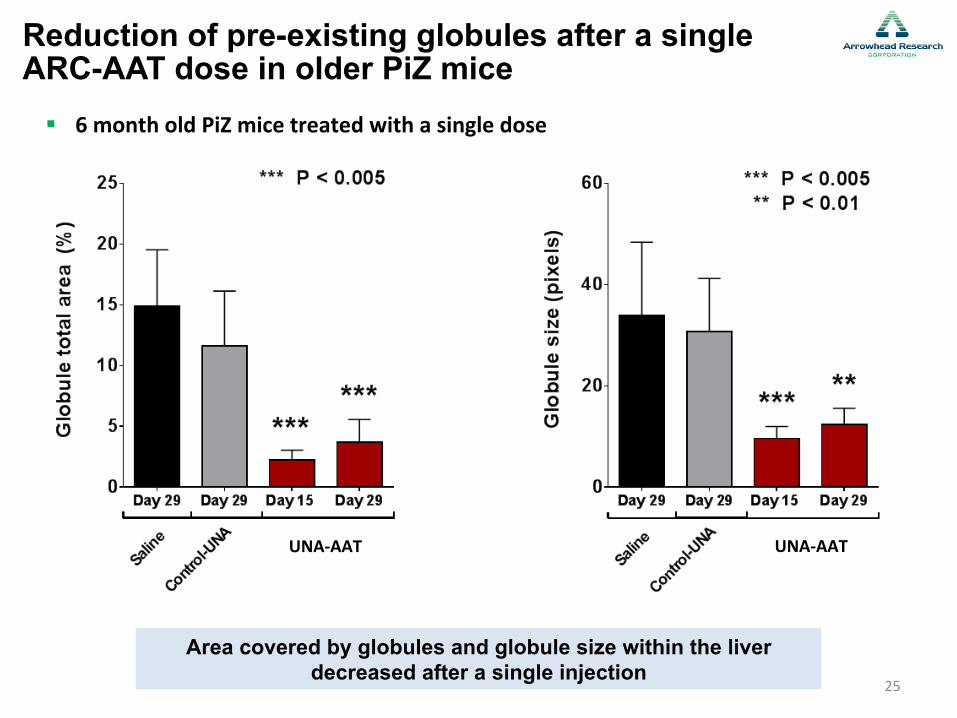
Area covered by globules and globule size within the liver decreased after a single injection
Reduction of pre-existing globules after a single ARC-AAT dose in older PiZ mice
25
! 6 month old PiZ mice treated with a single dose
UNA-‐AAT UNA-‐AAT

ARC-‐AAT: Summary
! AATD is a large scale orphan disease affecting >100,000 individuals in the US and a larger number in Europe
! Improved lung outcomes and resultant longer survival in adults is leading to a surge in recognized hepatic disease, including the emergence of HCC as an important issue
! AATD-related liver disease has clinically severe manifestations in pediatric patients
! AATD-related liver disease should be readily addressable by RNAi-based therapeutics
! Injection of ARC-AAT in transgenic mice expressing human PiZ AAT demonstrates this approach clears Z-AAT globules that cause fibrosis and HCC
! Injection of ARC-AAT in NHPs suggests once monthly dosing is sufficient to maintain 80 - 90% knockdown
26

! AATD is a large scale orphan disease affecting >100,000 individuals in the US and a larger number in Europe
! Improved lung outcomes and resultant longer survival in adults is leading to a surge in recognized hepatic disease, including the emergence of HCC as an important issue
! AATD-related liver disease has clinically severe manifestations in pediatric patients
! AATD-related liver disease should be readily addressed by RNAi-based therapeutics
! Injection of UNA-AAT in transgenic PiZ mice demonstrates this approach reduces the globules that lead to fibrosis and HCC
! Injection of UNA-AAT in NHPs suggests once every six-week dosing is sufficient to maintain 80 - 90% knockdown
27
ARC-‐AAT: Summary

Acknowledgements
Christine Wooddell Qili Chu Holly Hamilton Darren Wakefield Lauren Almeida Julia Hegge Zhao Xu Jacob Griffin Anthony Perillo-Nicholas Jason Klein Vladimir Subbotin Guofeng Zhang Alex Sokoloff Ryan Peterson Aaron Anderson Christine Hazlett Edie Doss Timothy Choi
Saint Louis University Jeffrey Teckman, M.D. Keith Blomenkamp
David Rozema Dawn Christianson James Hamilton David Lewis
Arrowhead Research Corp.