soil biology & biochemistry multiple climate change …aclassen/publications_files/2011...9700...
TRANSCRIPT

SSSAJ: Volume 75: Number 6 • November–December 2011
1
Soil Sci. Soc. Am. J. 75:2011 Posted online 26 Sept. 2011 doi:10.2136/sssaj2011.0135 Received 14 Apr. 2011. *Corresponding author ([email protected]). Supplementary material available online. © Soil Science Society of America, 5585 Guilford Rd., Madison WI 53711 USA All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher.
Multiple Climate Change Factors Interact to Alter Soil Microbial
Community Structure in an Old-Field Ecosystem
Soil Biology & Biochemistry
Microbial communities are an integral component of biogeochemical pro-cesses influencing the cycling of both C and nutrients in soils (Buckley and
Schmidt, 2003; Pendall et al., 2004; Bardgett et al., 2008). The composition and abundance of the soil microbial community influences the kinds of organic and in-organic materials that can be utilized as substrates, as well as the reaction rate of substrate utilization (e.g., Waldrop et al., 2000). Furthermore, the potential exists for changes in both the composition and function of various components of the soil microbial community in response to changes in climate, such as rising temperatures, increasing atmospheric CO2, and altered precipitation patterns (e.g., Pendall et al., 2004; Compant et al., 2010). While the direct effects of individual climatic change factors on soil communities have been demonstrated in a number of studies (e.g., Montealegre et al., 2002; Kandeler et al., 2008; Austin et al., 2009), we know less about how these climate drivers may interact to shape soil communities (Chung et al., 2007; Castro et al., 2010; Antoninka et al., 2009; Kardol et al., 2010b, 2011).
Sharon B. Gray*Bldg. 203, E-161 Biosciences DivisionArgonne National Lab.9700 S. Cass Ave.Argonne, IL 60439-4843andInstitute for Genomic BiologyUniv. of Illinois1206 W. Gregory Dr.Urbana, IL 61801
Aimée T. ClassenDep. of Ecology and Evolutionary Biology569 Dabney HallUniv. of TennesseeKnoxville, TN 37996
Paul KardolDep. of Forest Ecology and ManagementSwedish Univ. of Agricultural Sciences90183 Umeå, Sweden
Zhanna Yermakov R. Michael Miller
Bldg. 203, E-161 Biosciences DivisionArgonne National Lab.9700 S. Cass Ave.Argonne, IL 60439-4843
Climate change has the potential to alter both the composition and function of a soil’s microbial community, and interactions among climate change factors may alter soil communities in ways that are not possible to predict from experiments based on a single factor. This study evaluated the direct and interactive effects of three climate change factors—elevated CO2, altered amounts of precipitation, and elevated air temperature—on soil microbial commu-nities from an old-field climate change experiment being conducted at Oak Ridge, TN. Soil microbial community composition and biomass were determined by phospholipid fatty acid (PLFA) and neutral lipid fatty acid composition. We found that the interactive effects of precipitation and temperature treatments, as well as the interactive effects of precipitation and CO2 treatments, had significant impacts on microbial community composition. We found that total soil PLFA concentration, a measure of microbial biomass, was greater in the low-precipitation treatments, especially when low precipitation was combined with ambient CO2 concentrations or ambient temperature. Ordination analysis indicated that temperature was the most significant predictor of shifts in the soil microbial community composition, explaining approximately 12% of the variance in relative abundance of PLFA biomarkers. The elevated-temperature treatment increased the abundance of Firmicutes (low-guanine–cytosine Gram positive) and decreased the abundance of Gram-negative bacteria. Elevated temperature also reduced the abundance of the arbuscular mycorrhizal fungi PLFA biomarker 16:1π5c and saprophytic fungal PLFA biomarker 18:2π6,9. Overall, our data indicate that the interactions among climate change factors alter the composition of soil microbial communities in old-field ecosystems, suggesting potential for changes in microbial community function under predicted future climate conditions.
Abbreviations: AMF, arbuscular mycorrhizal fungi; BrSFA, branched, saturated fatty acid; BSI, bacterial stress index; CyFA, cyclopropyl fatty acid; GC, guanine and cytosine; MUFA, monounsaturated fatty acid; NLFA, neutral lipid fatty acid; OCCAM, Old-Field Community, Climate, and Atmosphere Manipulation; PLFA, phospholipid fatty acid.

6 SSSAJ: Volume 75: Number 6 • November–December 2011
Elevated temperature can directly alter the soil microbial community function by affecting temperature-sensitive micro-bial enzyme activity (von Lutzow and Kogel-Knabner, 2009). Elevated temperature can also influence soil microbial abun-dance and composition by altering net primary production and therefore the pool of available substrates utilized for microbial growth. Elevated temperature also increases the evaporative flux of water from the soil and therefore indirectly affects microorgan-isms through soil drying (Pendall et al., 2004; Zogg et al., 1997; Filella et al., 2004). The effects of elevated temperature on the total microbial biomass are variable (Zogg et al., 1997; Pendall et al., 2004), and groups of microorganisms differ in their responses to elevated temperature. For example, Firmicutes (low guanine and cytosine [GC], Gram positive) and Actinobacteria (high GC, Gram positive) have been observed to respond positively to elevated temperature, while Gram-negative bacteria respond negatively (Bardgett et al., 1999; Zogg et al., 1997). The response of the mycorrhizal fungal community to elevated temperature is variable and will probably be closely linked to the productiv-ity and C allocation responses of host plants (Bentivenga and Hetrick, 1992; Rillig et al., 2002; Heinemeyer and Fitter, 2004).
Rising atmospheric CO2 concentrations can alter the bio-mass and composition of soil microbial communities indirectly via increases in plant biomass production, increased C inputs from litterfall and root turnover, and mitigation of soil drying via reduced stomatal conductance of plants (Hu et al., 1999; Norby et al., 2001). Given the positive relationship between plant pro-ductivity and microbial biomass across many different soil types (Schimel, 1986; Burke, 1989), increases in plant productivity under elevated CO2 are expected to result in increased microbial biomass (e.g., Zak et al., 2000). Increased plant productivity, lit-ter inputs, and rhizodeposition, as well as changes in plant com-munity composition, can change the biochemical characteristics and quantity of resources that plants deliver to the soil under el-evated CO2 (e.g., Kandeler et al., 2008), differentially affecting groups of microorganisms. For example, increased rhizodeposi-tion of plants grown under elevated CO2 may increase the avail-ability of low-molecular-weight C substrates, favoring Gram-negative bacteria over Firmicutes or Actinobacteria (Kandeler et al., 2008; Milchunas et al., 2005; Drissner et al., 2007).
Changes in precipitation can affect the composition and functioning of the soil microbial community through differential drought tolerance among taxonomic and functional groups of mi-croorganisms. For example, more drought-tolerant microorgan-isms such as fungi, Firmicutes, and Actinobacteria usually benefit from dry conditions (Griffin, 1969; Drenovsky et al., 2004; Sylvia et al., 1998), while the abundance of Gram-negative bacteria usu-ally declines under soil drying (Harris, 1981; Nesci et al., 2004; Schimel et al., 2007). These changes lead to an increased ratio of fungi to bacteria under dry conditions, as measured by an in-creased microbial C/N ratio ( Jensen et al., 2003) and increased fungal/bacterial PLFA ratio (Drenovsky et al., 2004), resulting in a shift in community composition, which has important implica-tions for nutrient cycling and decomposition processes.
While much insight has been gained with regard to how changes in climate affect soil microbial communities, we have much to learn about how interactions among these variables might affect microbial communities. Interactive climate change effects on soil moisture (Dermody et al., 2007), soil temperature, and plant-mediated shifts in resource inputs such as changes in quantity and biochemical characteristics of litter or allocation of C to roots (Dukes and Hungate 2002) suggest a potential for downstream effects on the composition of soil communities (Hungate et al., 1996; Kardol et al., 2010b, 2011). We took ad-vantage of a multifactor climate change experiment that manipu-lated the atmospheric CO2, air temperature, and precipitation in an old-field community to examine how multiple climate change factors directly and interactively shape soil communities. We pre-dicted that the availability of soil moisture would have the largest influence on soil microbial communities and hence the precipi-tation treatments would moderate the impacts of warming and elevated atmospheric CO2 on the soil community composition.
MATeRiAlS And MeThodSSite description
The Old-Field Community, Climate, and Atmosphere Manipulation (OCCAM) experiment was established in 2002 at the Oak Ridge National Laboratory National Ecological Research Park in Oak Ridge, TN (35°54´12˝ N, 84°20´22˝ W). Soils at this site are developed in well-drained, slightly acidic, floodplain alluvium and are classified as Captina silt loam (a fine-silty, siliceous, active, mesic Typic Fragiudult) (Edwards and Norby, 1999). The average soil pH at the OCCAM site is 5.8 across treatments. Before establishment of the ex-perimental plots, the existing vegetation was killed using glyphosate herbicide [N-(phosphonomethyl)glycine], and dead plants as well as aboveground meristems were removed by scraping away the top 1 cm of soil using a hoe (Engel et al., 2009). In August 2002, following removal of the existing vegetation, seedlings of seven plant species common to old-field communities in the southeastern United States were transplanted into plots. These species included Plantago lanceolata L., a herbaceous forb; Andropogon virginicus L., a cespitose C4 bunchgrass; Festuca praten-sis Huds. subsp. apennina (De Not.) Hegi, syn. F. elatior L. var. apennina (De Not.) Hack., a C3 bunchgrass; Dactylis glomerata L., a C3 bunch-grass; Trifolium pratense L., a herbaceous legume; Solidago canadensis, a herbaceous forb; and Lespedeza cuneata (Dum. Cours.) G. Don, a N2–fixing dwarf shrub (see Engel et al., 2009; Kardol et al., 2010a,b).
This experimental facility has been extensively described elsewhere (Garten et al., 2008; Engel et al., 2009), but treatments are described brief-ly below. Open-top chambers (4-m diameter) with rain-exclusion shelters were constructed on 12 plots in each of three blocks. Treatments of ambient and elevated (300 μmol mol−1 above ambient) atmospheric CO2 concen-trations, ambient and elevated (3°C above ambient) air temperature, and differential irrigation (dry and wet plots) began in May 2003. The elevated CO2 treatment was achieved by injecting pure CO2 into the chamber ple-num (Engel et al., 2009). The daytime CO2 concentration in the ambient CO2 treatment averaged 395 μmol mol−1 in 2004 and 394 μmol mol−1 in 2005, and daytime CO2 concentration in elevated CO2 treatment aver-aged 703 μmol mol−1 in 2004 and 707 μmol mol−1 in 2005 (warming.

SSSAJ: Volume 75: Number 6 • November–December 2011 6
ornl.gov/OCCAM/data.html; verified 15 Aug. 2011). Temperature treat-ments were achieved through the use of evaporative coolers and in-line heating coils (Norby et al., 1997). The annual average air temperature in the ambient temperature treatment was 14.89 ± 0.17°C in 2004 and 14.96 ± 0.11°C in 2005, and the annual average air temperature in the elevated temperature treatments was 17.56 ± 0.18°C in 2004 and 17.19 ± 0.53°C in 2005 (warming.ornl.gov/OCCAM/data.html; verified 15 Aug. 2011). The CO2 and temperature treatments were imposed at a whole-plot level, and each whole plot was divided into two 6.3-m2 split plots that received either wet or dry irrigation treatments. Rainwater was collected onsite and added weekly to maintain the dry (2 mm wk−1) and wet (25 mm wk−1) split plots. The average annual precipitation at the field site is 136 cm, ap-proximately 26 mm wk−1 (Garten et al., 2009). Therefore, the wet treat-ment represents standard precipitation conditions at Oak Ridge, and the dry treatment represents a significant decrease in precipitation, which re-duced the soil volumetric water content in the top 15 cm from 26.2% in the wet treatments to 21.7% in the dry treatments (Wan et al., 2007). The absolute humidity in the plots was not affected by treatment (Wan et al., 2007). Treatments are designated as: ECAT, ACAT, ECET, and ACET, where EC is elevated CO2, AC is ambient CO2, ET is elevated tempera-ture, and AT is ambient temperature.
Soil Sampling and AnalysisSoil samples were collected in August 2004, October 2004, and June
2005 (0–15 cm) using a 5-cm-diameter soil corer. Three samples were col-lected at random locations within each split plot and stored at −20°C. Before processing, the samples were thawed, sieved to 4 mm, and roots were removed. The soils were freeze-dried (−50°C, 8 GPa) for 48 h in a Labconco Freezone 4.5 freeze-drier (Labconco, Kansas City, MO), sub-samples were composited to yield one sample per subplot per sampling date, and the composited samples were stored at room temperature until laboratory analysis of PLFA and neutral lipid fatty acid (NLFA) contents.
Lipids were extracted from a 2-g subsample of the composited, freeze-dried soil in a single-phase mixture of chloroform, methanol, and phosphate buffer (pH 7.4) at a ratio of 1:2:0.8, using an adaptation of the method described by Bligh and Dyer (1959). After 3 h, water and chloroform were added to separate the mixture into polar and nonpolar phases. Total lipids were extracted from the chloroform phase and phos-pholipids were separated from the neutral lipids using silicic acid col-umn chromatography (Allison et al., 2005). Phospholipids and neutral lipids were then saponified and the resulting fatty acids were methylated using a mild alkaline solution to form fatty acid methyl esters (FAMEs), and samples were frozen until analysis via gas chromatography.
Before gas chromatography analysis, samples were thawed and dissolved in a 24 mg L−1 solution of FAME 19:0 dissolved in hexane (Matreya, Pleasant Gap, PA). This known concentration of FAME 19:0 was used as an internal standard to quantify the concentration of PLFAs or NLFAs detected in the biological samples. Fatty acids were separated by capillary gas chromatography, using an HP 6890 gas chromatograph equipped with an HP7683 autosampler (Agilent Technologies, Palo Alto, CA). A 30-m HP-5MS column was used, with H2 as the carrier gas at a constant flow rate of 4 mL min−1. For each sample, a 1-μL splitless in-jection was made, with the inlet temperature set at 230°C and the inlet purged at 47 mL min−1 0.75 min after injection. The oven temperature
was held at 80°C for 1 min, increased at a rate of 20°C min−1 up to 155°C, and then increased at a rate of 5°C min−1 to a final temperature of 270°C. The oven temperature was then held at 270°C for 5 min. Individual fatty acids were detected by flame ionization at 350°C. Fatty acids were identi-fied by retention time in comparison to known standards and were quan-tified using a known concentration of FAME 19:0 as an internal standard.
Fatty acid nomenclature is in the form of A:BπC, where A is the num-ber of C atoms in the chain, B is the number of double bonds, and C is the position of the double bond from the methyl end of the PLFA (Tunlid and White, 1992; Allison and Miller, 2005). A cis geometry is indicated by the suffix c. The prefixes i, a, and me refer to iso, anteiso, and midchain methyl branching, respectively, and cy indicates a cyclopropyl ring structure.
data AnalysisIndividual PLFAs were summed to give an estimate of the total micro-
bial biomass (μmol kg−1 soil) in each sample. The average concentration of each individual PLFA is shown for all treatment combinations and sampling dates in Supplementary Table 1. To make inferences on the effects of climate change treatments on functionally similar microbial groups, we grouped signature fatty acids into biomarkers for the following microbial groups: Firmicutes, Gram-negative bacteria, Actinobacteria, arbuscular mycorrhizal fungi (AMF) and saprophytic fungi (Table 1). We further evaluated the energy storage of the AMF community using NLFAs (Olsson et al., 1997). Specifically, we estimated the total NLFA concentration for AMF species of the genera Glomus sensu lato using NLFA 16:1π5c (Graham et al., 1995; Olsson, 1999). We estimated the total NLFA concentration for AMF spe-cies of the genus Gigaspora as NLFA 22:1π9 + NLFA 20:1π9c (Graham et al., 1995; Bentivenga and Morton, 1996; Madan et al., 2002). We esti-mated treatment effects on the Gigaspora/Glomus species ratio as the ratio of these two sums of NLFAs. We estimated the ratio of Firmicutes to Gram-negative bacteria in each sample as the ratio of branched, saturated fatty acids to monounsaturated fatty acids (BrSFA/MUFA; Vestal and White, 1989; White et al., 1993). The degree of physiological stress experienced by the Gram-negative bacterial community was estimated using the bacterial stress index (BSI) for each sample: BSI = cy17:0 + cy19:0/16:1π7c + 18:1π7c (Guckert et al., 1986; Bossio et al., 1998). A further estimate of Gram-negative physiological stress was made using the ratio of cyclopropyl fatty
Table 1. Phospholipid fatty acids (PlFAs) signature assign-ments used in this study.
Group Biomarker PlFAs
Actinobacteria (high-GC gram positive) 10me16:0†, 10me18:0†
Bacteria 14:0‡, i15:0§, a15:0†¶, i16:0§, 16:1w7c†¶, 18:1w7c†¶, 10me18:0§, 10me16:0§, cy17:0§, a17:0†¶, i17:0†¶
Gram-negative bacteria 16:1w7c†¶, 18:1w7c†¶, cy17:0†¶
Firmicutes (low-GC Gram positive) i15:0†¶, i16:0†¶
Fungi Saprophytic 18:2w6,9c†¶, 18:1w9c‡Arbuscular mycorrhizal fungi 16:1w5c#, 20:1w9c††† Zelles (1999).‡ Zak et al. (1996).§ Frostegård and Bååth (1996).¶ Frostegård et al. (1993).# Olsson (1999).†† Madan et al. (2002).

6 SSSAJ: Volume 75: Number 6 • November–December 2011
acids to monounsaturated fatty acids [CyFA/MUFA = (cy17:0 + cy19:0)/(16:1π7c + 16:1π5c + 18:1π9c + 18:1π7c + 20:1π9c)] (Guckert et al., 1986; Ramos et al., 2001). An increase in the CyFA/MUFA ratio indicates a starvation response or a shift to stationary phase growth in Gram-negative bacteria (Ramos et al., 2001). We calculated the fungal/bacterial PLFA ratio as the sum of AMF and saprophytic fungi PLFAs divided by the sum of all bacterial PLFAs (Frostegård and Bååth, 1996).
Repeated measures analysis of variance with a variance components covariance structure (PROC MIXED in SAS ver-sion 9.2, SAS Institute, Cary, NC) was used to test for treat-ment effects on microbial groups, BSI, and the fungi/bacteria ratio across sampling dates. Analyses were conducted using the summed concentration (μmol kg−1) of PLFA markers belonging to each microbial group. Direct and interactive effects of CO2, temperature, and precipitation treatments were analyzed in a mixed-model ANOVA, where levels of CO2, temperature, and precipitation treatments were included as fixed effects and block as well as interactions between block and CO2, temperature, and precipitation treatments were included as random effects. Least squares means were separated using the pdiff option.
Treatment effects on the microbial community composition were analyzed by redundancy analysis (RDA) using relative abun-dance data, calculated as the concentration of each individual PLFA divided by the total concentration of PLFA in each sample. We tested the across-sampling-date effects of the treatments us-ing CO2, temperature, precipitation, and their interactions as explanatory variables and sampling date and block as covariables. To reflect our repeated measurements, treatment effects were tested using Monte Carlo permutation tests (999 permutations) restricted for split-plot design, i.e., records at different dates from the same plots (split plots) were not permuted, while whole plots were freely exchangeable (Lepš and Šmilauer, 2003). Samples were permuted within blocks. Individual treatment and interac-tion effects (marginal effects) were tested by manual selection of each individual term. Redundancy analyses were performed using CANOCO, version 4.5 (ter Braak and Šmilauer, 2002).
ReSulTSTreatment effects on Phospholipid Fatty Acid Concentrations in Microbial Groups
The total soil PLFA concentration was higher in dry treat-ments than wet treatments under ambient temperature, but pre-cipitation had no effect on the PLFA concentration under ele-vated temperature (F1,40 = 7.27, P = 0.01; Fig. 1; Supplementary Table 2). Additionally, there was a significant interaction be-tween CO2 and precipitation treatments, where total PLFA concentrations were higher in dry treatments exposed to ambi-ent CO2, but precipitation had no effect on total PLFA con-centrations under elevated CO2 (F1,40 = 4.07, P = 0.05; Fig. 1). Interestingly, total PLFA concentrations and concentrations of PLFA accounted for by specific microbial groups did not differ significantly among the August 2004, October 2004, and June 2005 collection dates, and there were no significant interactive effects of sampling date and treatment on total PLFA or PLFA of specific microbial groups (Supplementary Table 2).
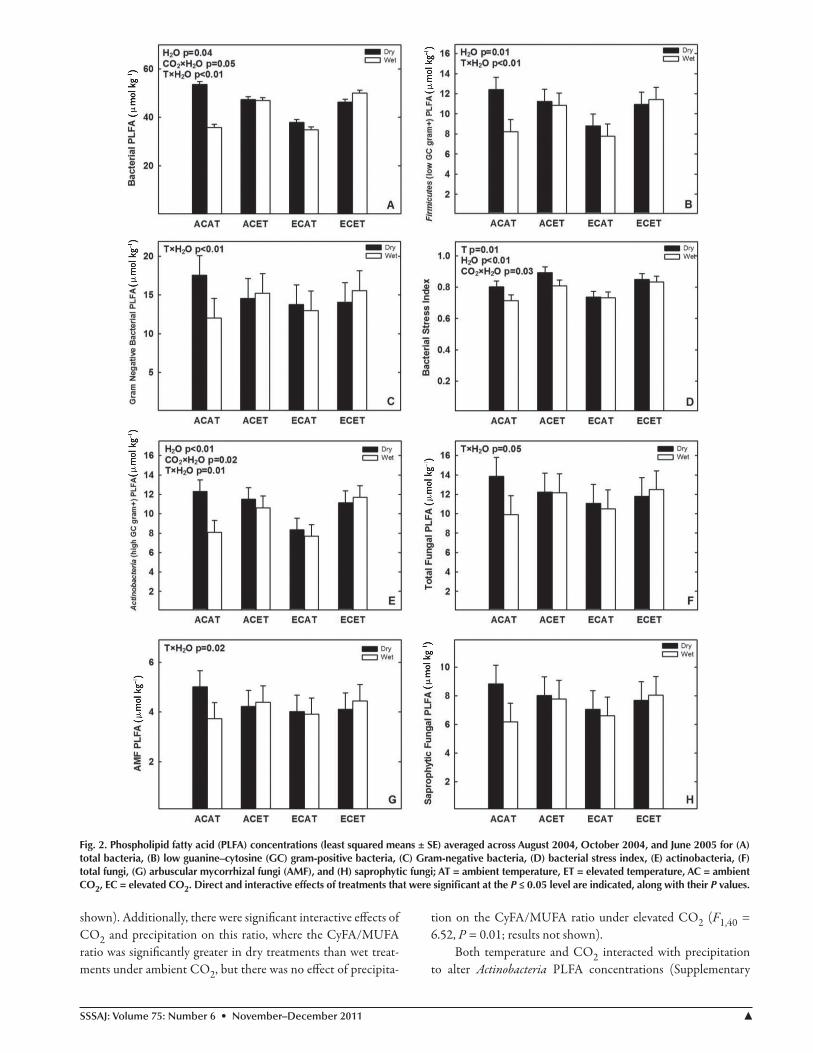
Similar to total PLFA (Fig. 1), the amount of PLFA ac-counted for by bacterial markers was higher in dry treatments than wet treatments exposed to ambient temperature, but pre-cipitation had no effect on total bacterial PLFA concentrations under elevated temperature (F1,40 = 7.98, P < 0.01; Fig. 2A). Additionally, total bacterial PLFA concentrations were higher in dry treatments exposed to ambient CO2, but precipitation had no effect on total PLFA concentrations under elevated CO2 (F1,40 = 4.2, P < 0.05; Fig. 2A).
The temperature × precipitation interaction on total bacte-rial PLFA appeared to be driven by both Gram-negative bacteria and Firmicute PLFA concentrations. Firmicute PLFA concentra-tions were higher in the dry treatment than the wet treatment under ambient temperature, but precipitation had no effect on Firmicute PLFA concentrations under elevated temperature (F1,40 = 7.39, P < 0.01; Fig. 2B). Gram-negative bacterial PLFA concentrations showed similar interactive effects of temperature and precipitation (F1,40 = 7.46, P < 0.01; Fig. 2C). The BrSFA/MUFA ratio was significantly greater in the elevated tempera-ture treatment than the ambient temperature treatment, indicat-ing that the elevated temperature increased the abundance of Firmicutes relative to Gram-negative bacteria (F1,6 = 10.94, P = 0.02; Fig. 3). Additionally, the BrSFA/MUFA ratio was signifi-cantly greater in dry plots than wet plots, indicating that reduced precipitation increased the abundance of Firmicutes relative to Gram-negative bacteria (F1,40 = 10.05, P = 0.003; Fig. 3).
The BSI was significantly higher under elevated relative to ambient temperature treatments (F1,6 = 12.0, P = 0.01; Fig. 2D). The BSI was also significantly higher in dry treatments than wet treatments (F1,40 = 8.46, P < 0.01; Fig. 2D); however, a signifi-cant CO2 × precipitation interaction revealed that the effect of precipitation was only significant in combination with ambient CO2 (CO2 × H2O F1,40 = 5.18, P = 0.03; Fig. 2D). Similar re-sults were found for the CyFA/MUFA ratio, which was signifi-cantly higher in the elevated temperature treatment than the am-bient temperature treatment (F1,6 = 8.08, P = 0.03; results not
Fig. 1. Total phospholipid fatty acid (PlFA) concentrations (least squared means ± standard error of the means averaged across August 2004, october 2004, and June 2005; AT = ambient temperature, eT = elevated temperature, AC = ambient Co2, eC = elevated Co2. direct and interactive effects of treatments that were significant at the P ≤ 0.05 level are indicated, along with their P values.
SSSAJ: Volume 75: Number 6 • November–December 2011 6
shown). Additionally, there were significant interactive effects of CO2 and precipitation on this ratio, where the CyFA/MUFA ratio was significantly greater in dry treatments than wet treat-ments under ambient CO2, but there was no effect of precipita-
tion on the CyFA/MUFA ratio under elevated CO2 (F1,40 = 6.52, P = 0.01; results not shown).
Both temperature and CO2 interacted with precipitation to alter Actinobacteria PLFA concentrations (Supplementary
Fig. 2. Phospholipid fatty acid (PlFA) concentrations (least squared means ± Se) averaged across August 2004, october 2004, and June 2005 for (A) total bacteria, (B) low guanine–cytosine (GC) gram-positive bacteria, (C) Gram-negative bacteria, (d) bacterial stress index, (e) actinobacteria, (F) total fungi, (G) arbuscular mycorrhizal fungi (AMF), and (h) saprophytic fungi; AT = ambient temperature, eT = elevated temperature, AC = ambient Co2, eC = elevated Co2. direct and interactive effects of treatments that were significant at the P ≤ 0.05 level are indicated, along with their P values.

6 SSSAJ: Volume 75: Number 6 • November–December 2011
Table 2; Fig. 2E). Actinobacteria PLFA concentrations were higher in the dry, ambient temperature treatment than the wet, ambient temperature treatment but precipitation had no effect under elevated temperature (F1,40 = 6.49, P = 0.01; Fig. 2E). There was also a significant CO2 × precipitation interaction where Actinobacteria PLFA concentrations were higher in dry than wet treatments under ambient CO2, but precipitation had no effect under elevated CO2 (F1,40 = 6.31, P = 0.02; Fig. 2E).
The fungal community was generally less responsive to cli-mate change treatments than the bacterial community. There were significant temperature × precipitation interactions on the total fungal PLFA concentration, where the concentration of total fun-gal PLFA was higher in dry than wet treatments under ambient temperature, but precipitation had no effect under elevated tem-perature (F1,40 = 4.23, P < 0.05; Fig. 2F). This response was driven by a significant temperature × precipitation interaction on AMF PLFA, where AMF PLFA concentrations were greater in dry than wet treatments under ambient temperature, but precipitation had no effect under elevated temperature (F1,40 = 5.62, P = 0.02; Fig. 2G). We also found that the dry treatment altered the NLFA com-position of the AMF community by increasing the Gigaspora/Glomus NLFA ratio, and this effect was driven by the August 2004 sampling date (precipitation × date F1,40 = 9.73, P = 0.0004; Fig. 5). Saprophytic fungal PLFA concentrations showed a similar re-sponse to the interactive effects of elevated temperature and dry treatment, but for saprophytic fungi, the temperature × precipita-tion interaction was not significant (F1,40 = 3.53, P = 0.07; Fig. 2H). There were no significant single or interactive effects of the climate change treatments or sampling date on the fungal/bacte-rial PLFA ratio (data not shown).
Multivariate AnalysisTogether, the climate change treatments explained 21% of
the total variation in PLFA composition. Temperature had the strongest effect on PLFA composition (explaining almost 12% of the variation, Table 2), as indicated by its correlation with the
first redundancy analysis axis (Fig. 4). The temperature effect was dependent on the CO2 treatment (Table 2). The ordina-tion diagram shows that PLFA biomarkers for Firmicutes (i16:0 and i15:0) and Actinobacteria (10me18:0 and 10me16:0) were associated with elevated temperature treatments, while PLFA biomarkers for AMF (16:1π5 and 20:1π9) and Gram-negative bacteria (16:1π7 and 18:1π7, which Graham et al. [1995] and others have also described as commonly found in AMF) were associated with ambient temperature treatments (Fig. 4). The precipitation treatment had fewer significant effects than the temperature treatment; however, there was a significant CO2 × precipitation effect explaining about 4% of the variation (Table 2), with wet, elevated CO2 treatments separating from dry, am-bient CO2 treatments. Most notably, the i15:0 biomarker for Firmicutes was strongly associated with dry, ambient CO2 treat-ments (Fig. 4).
diSCuSSionSignificant interactive effects of temperature and precipita-
tion as well as interactive effects of CO2 and precipitation im-pacted the biomass and composition of the soil microbial com-munity in our experiment. Surprisingly, and contrary to what we predicted, total PLFA concentrations were higher in dry than wet treatments, particularly when combined with ambient tem-perature or ambient CO2 treatments. Furthermore, temperature explained the greatest proportion of the variation in the relative abundance of PLFA biomarkers (Table 2); elevated temperature resulted in a microbial community with a higher proportion of Firmicutes and a lower proportion of fungi and Gram-negative bacteria. This change in composition of the bacterial community was also demonstrated by the CyFA/MUFA ratio.
Our data indicate that temperature influenced the effects of precipitation on the soil microbial biomass. The total PLFA concentration was higher in the dry treatment than the wet treat-ment, but this effect was only significant when soils were exposed to ambient temperature. These results were somewhat surprising, given that previous work in this system showed that, rather than responding to precipitation treatments, bacterial abundance (as indicated by total copies of rRNA genes) responded primarily
Fig. 3. Ratio of branched, saturated fatty acids (BrSFA) to monounsaturated fatty acids (MuFA) averaged across August 2004, october 2004, and June 2005; AT = ambient temperature, eT = elevated temperature, AC = ambient Co2, eC = elevated Co2. direct and interactive effects of treatments that were significant at the P ≤ 0.05 level are indicated, along with their P values.
Table 2. Results from redundancy analysis (RdA) testing the effect of treatments Co2, temperature (T), h2o, and their interactions on the relative abundance of phospholipid fatty acid biomarkers. explained variance (marginal effects) is the sum of all canonical eigenvalues.
explanatory variable F ratio P value explained variance
%
CO2 1.92 0.16 2.5
T 9.99 <0.01* 11.8
H2O 2.76 <0.01* 3.6
CO2 ´ T 2.81 0.02* 3.7
CO2 ´ H2O 2.85 0.01* 3.7
H2O ´ T 1.64 0.21 2.2
CO2 ´ H2O ´ T 0.82 0.50 1.1
* Significant at P ≤ 0.05 based on permutation tests restricted for sampling date and block.

SSSAJ: Volume 75: Number 6 • November–December 2011 6
to the interactive effects of CO2 and temperature, while fungal abundance (as indicated by total copies of rRNA genes) responded to the direct effects of temperature (Castro et al., 2010). Nematode data from this experi-ment, however, suggests that grazing of the soil microbial community by nematodes could explain some of the reduction in total microbial biomass under wet conditions (Kardol et al., 2010b). The total PLFA values aver-aged across our three sampling dates were negatively correlated with total nematode numbers in soils collected from OCCAM in October 2007; however, it is important to note that this value included plant-feeding nematodes as well as microbial-feed-ing nematodes (Kardol et al., 2010b; Pearson’s r = −0.70, P = 0.06).
The interactive effects of temper-ature and precipitation on bacterial-feeding nematodes may explain why the concentrations of PLFA biomark-ers for the total bacterial community were greater in the dry, ambient tem-perature treatments. The number of bacterial-feeding nematodes in this soil community decreased in dry treatments, but this effect was not apparent in elevated CO2 and ambient temperature treatments (Kardol et al., 2010b). While we did observe an increase in PLFA concentration for bacterial markers in the dry treatment, this effect was magnified in the am-bient temperature treatments. Similar to total PLFA, we found a strong negative correlation between total bacterial PLFA aver-aged across our three sampling dates and the number of bacterial-feeding nematodes in soils collected from this field site in October 2007 (Kardol et al., 2010b; Pearson’s r = −0.82, P = 0.01). Firmicutes and Actinobacteria are known to be inherently resistant to drying due to their strong, interlinked peptidoglycan cell walls and their ability to synthesize osmotic protectants (Schimel et al., 2007; Sylvia et al., 1998), which is consistent with our observa-tions that the dry treatment resulted in increased PLFA concentra-tions in both of these microbial groups. The strong negative corre-lation between soil bacterial PLFA concentrations and nematode abundance (Kardol et al., 2010b) indicates that top-down control in response to reduced grazing by nematodes, rather than a direct physiological response to climate change treatments, may have in-fluenced the bacterial community response.
Generally, Gram-negative bacteria are poorly adapted to drought because they rely on inducible osmotic protectants to pre-vent desiccation (Schimel et al., 2007). During times of metabolic stress, Gram-negative bacteria convert cis-monoenoic fatty acids to CyFAs, which allows greater membrane stability (Guckert et
al., 1986; Bossio et al., 1998). Consistent with previous findings (Steinberger et al., 1999), the BSI was higher in elevated relative to ambient temperature treatments. This was probably a response to the combined stresses of increasing temperature and the reduction in soil moisture that occurred under the elevated temperature treat-ment in our experiment (Dermody et al., 2007). Elevated tempera-ture also increased the CyFA/MUFA ratio, indicating a starvation
Fig. 4. ordination diagram (species-treatment biplot) of redundancy analysis (RdA) for the composition of phospholipid fatty acid (PlFA) biomarkers. Treatments are Co2, temperature (T), and precipitation and their interactions. Sampling date and block were included as covariables. Percentages along the axes correspond to the amount of explained variability in composition. For clarity, treatment interactions are not shown and only the best-fitting PlFA biomarkers are plotted.
Fig. 5. Ratio of neutral lipid fatty acid (nlFA) biomarkers for Gigaspora species to nlFA biomarkers for Glomus species, averaged across August 2004, october 2004, and June 2005; AT = ambient temperature, eT = elevated temperature, AC = ambient Co2, eC = elevated Co2. direct and interactive effects of treatments that were significant at the P ≤ 0.05 level are indicated, along with their P values.

6 SSSAJ: Volume 75: Number 6 • November–December 2011
response or a shift to stationary phase growth (Guckert et al., 1986). This observation is consistent with treatment effects on the BSI.
We also measured several significant interactive effects of pre-cipitation and CO2 (Supplementary Table 2; Fig. 1–2 and 5). The total PLFA was greater in dry than wet treatments, but this effect was not significant under elevated CO2 (Table 2; Fig. 1). Under ambient CO2, the concentration of Actinobacteria PLFA was sig-nificantly higher in dry treatments but under elevated CO2 wet and dry treatments did not differ. These patterns probably resulted from the mitigation of soil drying under elevated CO2, as previ-ously documented at our experimental site (Dermody et al., 2007). Mitigation of soil drying under elevated CO2 was associated with reduced stomatal conductance, which probably decreased transpi-ration of old-field plants grown under elevated CO2 (Dermody et al., 2007). Dessication-tolerant Actinobacteria may have had a competitive advantage over other soil microbes under ambient CO2 when dry conditions reduced soil moisture but not under elevated CO2 when the impacts of the dry treatment on the soil water content were reduced. Mitigation of soil drying by elevated CO2 may have also contributed to a lack of precipitation effects on bacterial-feeding nematodes (Kardol et al., 2010b) in the elevated CO2 treatment at OCCAM, which, in turn, probably contribut-ed to the interactive effects of CO2 and precipitation on bacterial PLFA concentrations. This trend was also observed in total bacte-rial PLFA and Firmicutes PLFA, although the interactive effects of CO2 and precipitation on Firmicutes were not significant.
Similar to the results for total PLFA, the BSI was higher in dry than wet treatments but only under ambient CO2. This previ-ously noted interaction between CO2 and precipitation demon-strates a novel mechanism by which mitigation of soil drying by elevated CO2 may decrease the degree of physiological stress ex-perienced by Gram-negative bacteria due to reduced precipitation.
Multivariate analysis indicated that elevated temperature increased the relative abundance of Firmicutes and decreased the relative abundance of AMF, saprophytic fungi, and Gram-negative bacteria. These results are supported by the trend of the BrSFA/MUFA ratio to increase in the elevated temperature treatment, indicating an increased ratio of Firmicutes to Gram-negative bac-teria. It was somewhat surprising that the wet treatment did not increase the AMF PLFA concentration, given that the wet treat-ment increased the plant community aboveground biomass at OCCAM by 80% (Kardol et al., 2010b). Our findings suggest, however, that climate change factors may have indirectly affected the fungal community via changes in nematode abundance. The saprophytic fungal PLFA concentration was negatively correlated with the density of fungal-feeding nematodes in the soil at this field site (Kardol et al., 2010b; Pearson’s r = −0.63, P = 0.09).
In addition to altering the abundance of fungal-derived lipids in the soil, environmental change may also alter the allocation pat-tern of energy-rich lipids to storage structures and spores of AMF (Bååth, 2003; Gavito and Olsson, 2003). We demonstrated that the dry treatment caused an increase in the Gigaspora/Glomus NLFA ratio (Fig. 5). Reduced soil moisture in the dry plots prob-ably preferentially stimulated the allocation of host photosyn-
thate to storage lipids and the production of spores in members of the AMF genus Gigaspora. Although variation exists for the kinds of neutral lipids that various AMF groups use as storage re-serves, there is a tendency for members of the Glomaceae to ac-cumulate the neutral lipid 16:1π5c, whereas members of the genus Gigaspora accumulate 20:1π9 and 22:1π9 (Graham et al., 1995; Olsson, 1999; Madan et al., 2002). We found that the NLFAs that accumulated under low precipitation were those NLFAs associ-ated with members of the various species that comprise the genera associated with Glomus sensu stricto and Scutellospora. Even so, the NLFAs that accumulated in association with AMF species belong-ing to the genus Gigaspora showed a significantly larger increase in the allocation of energy to spore production in response to the reduced precipitation treatment (Fig. 5).
Our results indicate that the interactions of reduced precipita-tion, elevated temperature, and elevated CO2 can significantly alter the biomass and composition of soil microbial communities. The strength of these interactive effects was demonstrated by the obser-vation that the dry treatment caused a 48% increase in total PLFA when combined with ambient CO2 and ambient temperature but did not affect the total PLFA in other treatment combinations (Fig. 1). This experiment provides unique evidence linking the previously reported reduction of soil drying under elevated CO2 (Dermody et al., 2007) to the mitigation of dry treatment effects on soil micro-organisms—for example, the desiccation-resistant Actinobacteria PLFA concentration increased in dry treatments but this effect was not significant under elevated CO2, where soil drying was reduced. Additionally, we demonstrated that previously reported changes in soil nematode populations in our experiment (Kardol et al., 2010b) were negatively correlated with the PLFA concentrations of vari-ous microbial groups. These results suggest that, in addition to di-rectly affecting microbial populations on the physiological level, elevated CO2, elevated temperature, and altered precipitation may affect microbial community biomass through top-down control. Recently, De Deyn et al. (2011) showed that PFLA biomarkers for individual microbial groups differentially responded to changes in plant community structure. Therefore, it is probable that climate-change-induced shifts in the plant community also have contrib-uted to the observed changes in microbial community composition in our experiment. Kardol et al. (2010b) found that the precipita-tion treatments explained the greatest amount of variation (12%) in the plant community composition at OCCAM—more than tem-perature or CO2 treatments—and that the dry treatment shifted plant cover dominance from Lespedeza cuneata to Festuca pratensis. Interestingly, they also found that enzyme activities in soils under L. cuneata were reduced relative to soils under F. pratensis, suggesting lower microbial biomass beneath L. cuneata (Kardol et al., 2010b). These data suggest that the shift to F. pratensis dominance that oc-curred under the dry treatment at OCCAM may have played a role in the increased PLFA concentrations under the dry treatment that were observed in this study (Fig. 1). Changes in the relative abun-dance of individual microbial groups suggest that, with time, climate change factors may alter the function of soil microbial communities. This may be particularly important in terms of specialized roles per-

SSSAJ: Volume 75: Number 6 • November–December 2011 6
formed by Gram-negative bacteria, such as nitrification or CH4 oxi-dation, as our experiment demonstrated that elevated temperature increased the Firmicutes/Gram-negative bacteria ratio and increased indicators of stress in the Gram-negative bacterial community.
ACKnowledGMenTSWe thank S. Wan and P. Allan for field assistance; R.J. Norby and J.A. Weltzin were integral in establishing the OCCAM experiment. Research was sponsored by the U.S. Dep. of Energy, Office of Science, Biological and Environmental Research Program under Contract no. DE-AC02-06CH11357 to Argonne National Laboratory and Grant no. DE-FG02-02ER63366 to the University of Tennessee; S.B. Gray was supported by the U.S. Dep. of Energy’s Global Change Education Program. Work was conducted in collaboration with Oak Ridge National Laboratory, which is managed by UT Battelle, LLC, for the U.S. Dep. of Energy under Contract DE-AC05-00OR22725.
ReFeRenCeSAllison, V.J., and R.M. Miller. 2005. Soil grinding increases the relative abundance
of eukaryotic phospholipid fatty acids. Soil Sci. Soc. Am. J. 69:423–426. doi:10.2136/sssaj2005.0423
Allison, V.J., R.M. Miller, J.D. Jastrow, R. Matamala, and D.R. Zak. 2005. Characterization of environmental and edaphic factors affecting soil microbial community structure in a tallgrass prairie chronosequence. Soil Sci. Soc. Am. J. 69:1412–1421. doi:10.2136/sssaj2004.0252
Antoninka, A., J.E. Wolf, M. Bowker, and A.T. Classen. 2009. Linking above- and belowground responses to global change at community and ecosystem scales. Global Change Biol. 15:914–929. doi:10.1111/j.1365-2486.2008.01760.x
Austin, E.E., H.F. Castro, C.W. Schadt, K.T. Sides, and A.T. Classen. 2009. Assessment of 10 years of CO2 fumigation on soil microbial communities and function in a sweetgum plantation. Soil Biol. Biochem. 41:514–520. doi:10.1016/j.soilbio.2008.12.010
Bååth, E. 2003. The use of neutral lipid fatty acids to indicate the physiological conditions of soil fungi. Microb. Ecol. 45:373–383. doi:10.1007/s00248-003-2002-y
Bardgett, R.D., C. Freeman, and N.J. Ostle. 2008. Microbial contributions to climate change through carbon cycle feedbacks. ISME J. 2:805–814. doi:10.1038/ismej.2008.58
Bardgett, R.D., E. Kandeler, D. Tscherko, P.J. Hobbs, T.M. Bezemer, T.H. Jones, and L.J. Thompson. 1999. Below-ground microbial community development in a high temperature world. Oikos 85:193–203. doi:10.2307/3546486
Bentivenga, S.P., and B.A.D. Hetrick. 1992. Seasonal and temperature effects on mycorrhizal activity and dependence of cool- and warm-season tallgrass prairie grasses. Can. J. Bot. 70:1596–1602. doi:10.1139/b92-201
Bentivenga, S.P., and J.B. Morton. 1996. Congruence of fatty acid methyl ester profiles and morphological characters of arbuscular mycorrhizal fungi in Gigasporaceae. Proc. Natl. Acad. Sci. 93:5659–5662. doi:10.1073/pnas.93.11.5659
Bligh, E.G., and W.J. Dyer. 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37:911–917. doi:10.1139/o59-099
Bossio, D.A., K.M. Scow, N. Gunapala, and K.J. Graham. 1998. Determinants of soil microbial communities: Effects of agricultural management, season and soil type on phospholipid fatty acid profiles. Microb. Ecol. 36:1–12. doi:10.1007/s002489900087
Buckley, D.H., and T.M. Schmidt. 2003. The structure of microbial communities in soil: Patterns of microbial distribution, their dynamic nature and the lasting impact of cultivation. Environ. Microbiol. 5:441–452. doi:10.1046/j.1462-2920.2003.00404.x
Burke, I.C. 1989. Control of nitrogen mineralization in a sagebrush steppe landscape. Ecology 70:1115–1126. doi:10.2307/1941380
Castro, H.F., A.T. Classen, E.E. Austin, R.J. Norby, and C.W. Schadt. 2010. Soil microbial community responses to multiple experimental climate change drivers. Appl. Environ. Microbiol. 76:999–1007. doi:10.1128/AEM.02874-09
Chung, H., D.R. Zak, P.B. Reich, and D.S. Ellsworth. 2007. Plant species richness, elevated CO2, and atmospheric nitrogen deposition alter soil microbial community composition and function. Global Change Biol. 13:980–989. doi:10.1111/j.1365-2486.2007.01313.x
Compant, S., M.G.A. van der Heijden, and A. Sessitsch. 2010. Climate change effects on beneficial plant–microorganism interactions. FEMS Microbiol. Ecol. 73:197–214.
De Deyn, G.B., H. Quirk, and R.B. Bardgett. 2011. Plant species richness, identity and productivity differentially influence key groups of microbes in grassland soils of contrasting fertility. Biol. Lett. 7:75–78. doi:10.1098/rsbl.2010.0575
Dermody, O., J.F. Weltzin, E.C. Engel, P. Allen, and R.J. Norby. 2007. How do elevated CO2, warming, and reduced precipitation interact to affect soil moisture and LAI in an old field ecosystem? Plant Soil 301:255–266. doi:10.1007/s11104-007-9443-x
Drenovsky, R.E., D. Vo, K.J. Graham, and K.M. Scow. 2004. Soil water content and organic carbon availability are major determinants of soil microbial community composition. Microb. Ecol. 48:424–430. doi:10.1007/s00248-003-1063-2
Drissner, D., H. Blum, D. Tscherko, and E. Kandeler. 2007. Nine years of enriched CO2 changes the function and structural diversity of soil microorganisms in a grassland. Eur. J. Soil Sci. 58:260–269. doi:10.1111/j.1365-2389.2006.00838.x
Dukes, J.S., and B.A. Hungate. 2002. Elevated carbon dioxide and litter decomposition in California annual grasslands: Which mechanisms matter? Ecosystems 5:171–183. doi:10.1007/s10021-001-0063-7
Edwards, N.T., and R.J. Norby. 1999. Below-ground respiratory responses of sugar maple and red maple saplings to atmospheric CO2 enrichment and elevated air temperature. Plant Soil 206:85–97. doi:10.1023/A:1004319109772
Engel, E.C., J.F. Weltzin, R.J. Norby, and A.T. Classen. 2009. Responses of an old-field plant community to interacting factors of elevated [CO2], warming, and soil moisture. J. Plant Ecol. 2:1–11. doi:10.1093/jpe/rtn026
Filella, I., J. Peñuelas, L. Llorens, and M. Estiarte. 2004. Reflectance assessment of seasonal and annual changes in biomass and CO2 uptake of a Mediterranean shrubland submitted to experimental warming and drought. Remote Sens. Environ. 90:308–318. doi:10.1016/j.rse.2004.01.010
Frostegård, Å., and E. Bååth. 1996. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fertil. Soils 22:59–65. doi:10.1007/BF00384433
Frostegård, Å., E. Bååth, and A. Tunlid. 1993. Shifts in the structure of soil microbial communities in limed forests as revealed by phospholipid fatty acid analysis. Soil Biol. Biochem. 25:723–730. doi:10.1016/0038-0717(93)90113-P
Garten, C.T., Jr., A.T. Classen, and R.J. Norby. 2009. Soil moisture surpasses elevated CO2 and temperature as a control on soil carbon dynamics in a multi-factor climate change experiment. Plant Soil 319:85–94. doi:10.1007/s11104-008-9851-6
Garten, C.T., Jr., A.T. Classen, R.J. Norby, D.J. Brice, J.F. Weltzin, and L. Souza. 2008. Role of N2–fixation in constructed old-field communities under different regimes of CO2, temperature, and water availability. Ecosystems 11:125–137. doi:10.1007/s10021-007-9112-1
Gavito, M.E., and P.A. Olsson. 2003. Allocation of plant carbon to foraging and storage in arbuscular mycorrhizal fungi. FEMS Microbiol. Ecol. 45:181–187. doi:10.1016/S0168-6496(03)00150-8
Graham, J.H., N.C. Hodge, and J.B. Morton. 1995. Fatty acid methyl ester profiles for characterization of glomalean fungi and their endomycorrhizae. Appl. Environ. Microbiol. 61:58–64.
Griffin, D.M. 1969. Soil water in the ecology of fungi. Annu. Rev. Phytopathol. 7:289–310. doi:10.1146/annurev.py.07.090169.001445
Guckert, J.B., M.A. Hood, and D.C. White. 1986. Phospholipid ester-linked fatty-acid profile changes during nutrient deprivation of Vibrio cholerae: Increases in the trans/cis ratio and proportion of cyclopropyl fatty acids. Appl. Environ. Microbiol. 52:794–801.
Harris, R.F. 1981. Effect of water potential on microbial growth and activity. p. 23–96. In J.R. Parr et al. (ed.) Water potential relations in soil microbiology. SSSA Spec. Publ. 9. SSSA, Madison, WI.
Heinemeyer, A., and A.H. Fitter. 2004. Impact of temperature on the arbuscular mycorrhizal (AM) symbiosis: Growth responses of the host plant and its AM fungal partner. J. Exp. Bot. 55:525–534. doi:10.1093/jxb/erh049
Hu, S., M.K. Firestone, and F.S. Chapin III. 1999. Soil microbial feedbacks to atmospheric CO2 enrichment. Trends Ecol. Evol. 14:433–437. doi:10.1016/S0169-5347(99)01682-1
Hungate, B.A., J. Canadell, and F.S. Chapin III. 1996. Plant species mediate changes in soil microbial N in response to elevated CO2. Ecology 77:2505–2515. doi:10.2307/2265749
Jensen, K.D., C. Beier, A. Michelsen, and B.A. Emmett. 2003. Effects of experimental drought on microbial processes in two temperate heathlands

6 SSSAJ: Volume 75: Number 6 • November–December 2011
at contrasting water conditions. Appl. Soil Ecol. 24:165–176. doi:10.1016/S0929-1393(03)00091-X
Kandeler, E., A.R. Mosier, J.A. Morgan, D.G. Milchunas, J.Y. King, S. Rudolph, and D. Tscherko. 2008. Transient elevation of carbon dioxide modifies the microbial community composition in semi-arid grassland. Soil Biol. Biochem. 40:162–171. doi:10.1016/j.soilbio.2007.07.018
Kardol, P., C.E. Campany, L. Souza, R.J. Norby, J. Weltzin, and A.T. Classen. 2010a. Climatic change alters plant dominance patterns and evenness in an experimental old-field ecosystem. Global Change Biol. 16:2676–2687. doi:10.1111/j.1365-2486.2010.02162.x
Kardol, P., M.A. Cregger, C.E. Campany, and A.T. Classen. 2010b. Soil ecosystem functioning under climate change: Plant species and community effects. Ecology 91:767–781. doi:10.1890/09-0135.1
Kardol, P., W.N. Reynolds, R.J. Norby, and A.T. Classen. 2011. Climate change effects on soil microarthropod abundance and community structure. Appl. Soil Ecol. 47:37–44. doi:10.1016/j.apsoil.2010.11.001
Lepš, J., and P. Šmilauer. 2003. Multivariate analysis of ecological data using CANOCO. Cambridge Univ. Press, Cambridge, UK.
Madan, R., C. Pankhurst, B. Hawke, and S. Smith. 2002. Use of fatty acids for identification of AM fungi and estimation of the biomass of AM spores in soil. Soil Biol. Biochem. 34:125–128. doi:10.1016/S0038-0717(01)00151-1
Milchunas, D.G., J.A. Morgan, A.R. Mosier, and D.R. LeCain. 2005. Root dynamics and demography in shortgrass steppe under elevated CO2, and comments on minirhizotron methodology. Global Change Biol. 11:1837–1855. doi:10.1111/j.1365-2486.2005.001009.x
Montealegre, C.M., C. van Kessel, M.P. Russelle, and M.J. Sadowsky. 2002. Changes in microbial activity and composition in a pasture ecosystem exposed to elevated atmospheric carbon dioxide. Plant Soil 243:197–207. doi:10.1023/A:1019901828483
Nesci, A., M. Etcheverry, and N. Magan. 2004. Osmotic and matric potential effects on growth, sugar alcohol and sugar accumulation by Aspergillus section Flavi strains from Argentina. J. Appl. Microbiol. 96:965–972. doi:10.1111/j.1365-2672.2004.02246.x
Norby, R.J., M.F. Cotrufo, P. Ineson, E.G. O’Neill, and J.G. Canadell. 2001. Elevated CO2, litter chemistry, and decomposition: A synthesis. Oecologia 127:153–165. doi:10.1007/s004420000615
Norby, R.J., N.T. Edwards, J.S. Riggs, C.H. Abner, S.D. Wullschleger, and C.A. Gunderson. 1997. Temperature-controlled open-top chambers for global change research. Global Change Biol. 3:259–267. doi:10.1046/j.1365-2486.1997.00072.x
Olsson, P.A. 1999. Signature fatty acids provide tools for determination of the distribution and interactions of mycorrhizal fungi in soil. FEMS Microbiol. Ecol. 29:303–310. doi:10.1111/j.1574-6941.1999.tb00621.x
Olsson, P.A., E. Bååth, and I. Jakobsen. 1997. Phosphorus effects on the mycelium and storage structures of an arbuscular mycorrhizal fungus as studied in the soil and roots by analysis of fatty acid signatures. Appl. Environ. Microbiol. 63:3531–3538.
Pendall, E., S. Bridgham, P.J. Hanson, B. Hungate, D.W. Kicklighter, D.W. Johnson, B.E. Law, Y. Luo, J.P. Megonigal, M. Olsrud, M.G. Ryan, and S. Wan. 2004. Below-ground process responses to elevated CO2 and temperature: A discussion of observations, measurement methods, and models. New Phytol. 162:311–322. doi:10.1111/j.1469-8137.2004.01053.x
Ramos, J.L., M.T. Gallegos, S. Marques, M.I. Ramos-Gonzales, M. Espinosa-Urgel, and A. Segura. 2001. Responses of Gram-negative bacteria to certain environmental stressors. Curr. Opin. Microbiol. 4:166–171. doi:10.1016/
S1369-5274(00)00183-1Rillig, M.C., S.F. Wright, M.R. Shaw, and C.B. Field. 2002. Artificial climate
warming positively affects arbuscular mycorrhizae but decreases soil aggregate water stability in an annual grassland. Oikos 97:52–58. doi:10.1034/j.1600-0706.2002.970105.x
Schimel, D.S. 1986. Carbon and nitrogen turnover in adjacent grassland and cropland ecosystems. Biogeochemistry 2:345–357. doi:10.1007/BF02180325
Schimel, J., T.C. Balser, and M. Wallenstein. 2007. Microbial stress-response physiology and its implications for ecosystem function. Ecology 88:1386–1394. doi:10.1890/06-0219
Steinberger, Y., L. Zelles, Q.Y. Bai, M. von Lutzow, and J.C. Much. 1999. Phospholipid fatty acid profiles as indicators for the microbial community structure in soils along a climatic transect in the Judean Desert. Biol. Fertil. Soils 28:292–300. doi:10.1007/s003740050496
Sylvia, D.M., J.J. Fuhrman, P.G. Hartel, and D.A. Zuberer. 1998. Principles and applications of soil microbiology. Prentice Hall, Upper Saddle River, NJ.
ter Braak, C.J.F., and P. Šmilauer. 2002. CANOCO reference manual and CanoDraw for Windows user’s guide: Software for canonical community ordination. Microcomputer Power, Ithaca, NY.
Tunlid, A., and D.C. White. 1992. Biochemical analysis of biomass, community structure, nutritional status, and metabolic activity of microbial communities in soil. p. 229–262. In G. Stotzky and J.-M. Bollag (ed.) Soil biochemistry. Vol. 7. Marcel Dekker, New York.
Vestal, J.R., and D.C. White. 1989. Lipid analysis in microbial ecology: Quantitative approaches to the study of microbial communities. BioScience 39:535–541. doi:10.2307/1310976
von Lutzow, M., and I. Kogel-Knabner. 2009. Temperature sensitivity of soil organic matter decomposition: What do we know? Biol. Fertil. Soils 46:1–15. doi:10.1007/s00374-009-0413-8
Waldrop, M.P., T.C. Balser, and M.K. Firestone. 2000. Linking microbial community composition to function in a tropical soil. Soil Biol. Biochem. 32:1837–1846. doi:10.1016/S0038-0717(00)00157-7
Wan, S., R.J. Norby, J. Ledford, and J.F. Weltzin. 2007. Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland. Global Change Biol. 13:2411–2424. doi:10.1111/j.1365-2486.2007.01433.x
White, D.C., D.B. Ringelberg, D.B. Hedrick, and D.E. Nivens. 1993. Rapid identification of microbes from clinical and environmental matrices by mass spectrometry. p. 8–17. In C. Fenselau (ed.) Identification of microbes by mass spectrometry. Symp. Ser. 541. Am. Chem. Soc., Washington, DC.
Zak, D.R., K.S. Pregitzer, J.S. King, and W.E. Holmes. 2000. Elevated atmospheric CO2, fine roots and the response of soil microorganisms: A review and hypothesis. New Phytol. 147:201–222. doi:10.1046/j.1469-8137.2000.00687.x
Zak, D.R., D.B. Ringelberg, K.S. Pregitzer, D.L. Randlett, D.C. White, and P.S. Curtis. 1996. Soil microbial communities beneath Populus grandidentata grown under elevated atmospheric CO2. Ecol. Appl. 6:257–262. doi:10.2307/2269568
Zelles, L. 1999. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterization of microbial communities in soil: A review. Biol. Fertil. Soils 29:111–129. doi:10.1007/s003740050533
Zogg, G.P., D.R. Zak, D.B. Ringelberg, N.W. MacDonald, K.S. Pregitzer, and D.C. White. 1997. Compositional and functional shifts in microbial communities due to soil warming. Soil Sci. Soc. Am. J. 61:475–481. doi:10.2136/sssaj1997.03615995006100020015x