splicing alternativo y la progresión tumoral
TRANSCRIPT

Splicing Alternativo y La Progresión Tumoral
Resumen: El splicing alternativo es un mecanismo molecular clave para el aumento de la diversidad
funcional de los proteomas eucariotas. Un gran chico de los datos experimentales implica corte y
empalme aberrante en varias enfermedades humanas, incluyendo el cáncer. Ambas mutaciones en los
elementos de corte y empalme que actúan en cis y alteraciones en la expresión y / o actividad dad de
factores reguladores de empalme afectan drásticamente el perfil de empalme de muchos genes
asociados con el cáncer. Además, el perfil de empalme de varios genes cancerassociated se altera en
determinados tipos de cáncer argumentan a favor de un papel directo de las isoformas de corte y
empalme específicos en la progresión tumoral.
Descifrando los mecanismos que subyacen a corte y empalme aberrante en el cáncer puede ser crucial
para entender cómo se controla la maquinaria de empalme e integrado con otros procesos celulares, en
particular, la transcripción y vías de señalización. Por otra parte, la caracterización de la desregulación
de empalme en el cáncer conducirá a una mejor comprensión de la transformación maligna. Variantes de
splicing alternativo asociado al cáncer pueden ser nuevas herramientas para el diagnóstico y
clasificación de los tipos de cáncer y podrían ser los objetivos de las intervenciones terapéuticas
innovadoras basadas en enfoques de corrección de empalme altamente selectivos.
INTRODUCCIÓN
Durante la evolución del número de genes dejó de crecer en paralelo con la complejidad del proteoma. Por
lo tanto, el genoma humano contiene sólo 20,000-25,000 genes (Genoma Humano Consorcio
Internacional de Secuenciación 2004), un número no significativamente diferente de la contadas en
organismos menos complejos tales como erizos de mar (23.000) (Erizo de mar de Secuenciación del
Genoma Consorcio 2006) y el nematodo gusano (19 000). Por otra parte, el número de genes humanos no
es suficiente para dar cuenta de todas las proteínas reveladas por el análisis proteómico.¿Cómo se
pueden explicar estas paradojas? Recientes cDNA secuencia y los datos de microarrays han implicado
splicing alternativo (AS) como la principal fuente de diversidad funcional y proteómica en organismos
metazoos. Junto con otros promotores y sitios de poliadenilación, edición de ARN y el procesamiento
después de la traducción, COMO da lugar a un número estimado de al menos 100.000 proteínas humanas
diferentes.
El término "splicing alternativo" describe cualquier situación en la que un único transcrito primario (pre-
ARNm) se puede empalmar en más de un patrón para generar múltiples mRNAs maduros, distintas que
conducen a la expresión de isoformas de proteínas con diferentes propiedades estructurales y
funcionales. El "recordholder" para corte y empalme alternativo es un gen de Drosophila
llamado Dscam, con 38.000 variantes de empalme, más que el número de genes de Drosophila. En los
seres humanos, al menos, el 70% (y esta proporción podría ser aún más alto!) De los genes que codifican
para las transcripciones que se someten a splicing alternativo, que pone de relieve la importancia de
este mecanismo de regulación de la biología de la especie.
Debido a su capacidad para generar la diversidad de proteínas, se espera que corte y empalme
alternativo para jugar un papel importante en la regulación de la expresión génica, una predicción que
se fundamenta en la observación de que la generación de espacio-temporal apropiado de las variantes
de corte y empalme está implicado en muchos procesos celulares y de desarrollo (incluyendo la
determinación del sexo, la apoptosis, la guía de axones, la excitación y contracción celular y muchos
otros). No es de extrañar, por tanto, que la desregulación de los programas de splicing alternativo está
estrechamente vinculada a trastornos genéticos humanos heredados y adquiridos. De hecho, trabajos
en los últimos años han empezado a reconocer no apropiado splicing alternativo como un modificador
genético durante la tumorigénesis. Muchos de los genes relacionados con el cáncer están regulados por

splicing alternativo. Ellos codifican para proteínas implicadas en todos los aspectos importantes de la
biología celular de cáncer, incluyendo el control del ciclo celular, la proliferación, la diferenciación, las
vías de transducción de señales, la muerte celular, la angiogénesis, la invasión, la motilidad y la
metástasis. Una firma común de las células cancerosas es una pérdida general de la fidelidad de
empalme, con la consiguiente reorganización de los perfiles de corte y empalme, e incluso cambiar a
isoformas de empalme específicos por lo general expresadas en otros tipos de células. Todos estos
acontecimientos pueden contribuir a la carcinogénesis. En particular, hay varios casos de corte y
empalme alternativo que se limita a los tipos específicos de cáncer, lo que implica claramente que la
isoforma de empalme en particular en la progresión tumoral. Eventos de empalme del cáncer-específicos
pueden ser también beneficioso para la terapia, ya que generan nuevos epítopos contra los que es posible
producir anticuerpos para inmunoterapia. Por lo tanto, existe un gran interés en el descubrimiento del
efecto de corte y empalme alternativo de la complejidad del transcriptoma de células cancerosas y en
la comprensión de cómo este mecanismo de regulación contribuye a la tumorigénesis. Esta revisión
discute (1) los mecanismos básicos de splicing alternativo, (2) los conocimientos actuales sobre los
mecanismos de regulación que rigen splicing alternativo y la desregulación en el cáncer, (3) las
consecuencias biológicas derivadas de la alteración de empalme de alguna relacionada con el cáncer
relevante genes y (4) las variantes de corte y empalme alternativos de cáncer asociados con un valor de
diagnóstico / pronóstico claro que puede proporcionar dianas terapéuticas potenciales.
Splicing alternativo
Genomas eucariotas se caracterizan por la presencia de genes "interrumpidas" que consiste en una
sucesión ordenada de codificación (exones) y no codificante (intrón) secuencias. En metazoos,
particularmente en los mamíferos, interrumpido cuenta los genes para la gran mayoría de los genes. Por
lo tanto, el empalme de ARN se produce en forma obligatoria, altamente regulado, proceso. A través de
corte y empalme de pre-ARNm, intrones se eliminan con precisión y los exones se unen entre sí para
reconstituir el marco de lectura y para generar ARNm traducibles que luego se exportan al citoplasma.
La maquinaria de empalme (llamado spliceosome), es la máquina molecular más grande hasta ahora se
describe en las células se compone de cinco ribonucleoproteínas nucleares pequeñas (snRNPs U1, U2,
U4, U5 y U6) y más de 100 polipéptidos diferentes. El spliceosome reconoce, mal conservadas, la
secuencia de elementos que actúan en cis cortos en los límites exón-intrón (5 'y 3' sitios de empalme
también conocidos como sitios de "aceptores" "donante" y) y utilizarlos para la reacción de cortar y
pegar ( dos trans-esterificación secuencial) que elimina el intrón.
Mediante el uso de varias combinaciones de donante y aceptor de los sitios de diferentes exones, a
través del proceso de corte y empalme alternativo que es posible producir distintos mRNAs de un pre-
ARNm única. Se han observado cinco patrones de empalme alternativos distintos: 1) un exón regulado
(casete), que a veces se incluye y, a veces excluido del ARNm; 2) múltiples cassette exones que son
mutuamente excluyentes, es decir, el ARNm maduro siempre contiene sólo una de varias posibles
opciones exón; 3) en casos raros, todo un intrón se conservan; sitios aceptores 4) donantes alternativa
y 5) pueden resultar en exones de diferente tamaño (fig. 1). Por último, los promotores alternativos y
sitios de poli-adenilación contribuyen a la heterogeneidad de las transcripciones codificadas por un solo
gen. Además de modificar las proteínas características, corte y empalme alternativo puede afectar a la
estabilidad de las transcripciones a través de la decadencia (NMD) vía ARNm nonsensemediated, un
mecanismo de CONTROL DE CALIDAD ARNm que depende de la maquinaria de traducción. Análisis
recientes sugieren que aproximadamente el 35% de todos los eventos de corte y empalme alternativos
en las células de mamíferos generar especies de ARNm que contienen codones de terminación
prematuros (PTC), que pueden ser degradados de manera eficiente a través de
NMD. Curiosamente, Ni han demostrado recientemente que los exones que contienen un codón de parada
son particularmente frecuentes y conservadas en los genes para factores de empalme que intervienen

en la elección de empalme. Estos exones con frecuencia se superponen elementos ultraconserved en los
genomas de mamíferos.
Desde un punto de vista mecanicista de los diferentes tipos de splicing alternativo puede ser
simplemente visto como un problema de reconocimiento de empalme de sitio por el spliceosome donde
la decisión entre la inclusión y la omisión de un exón particular depende principalmente en el
reconocimiento y la utilización de los sitios de empalme que flanquean el exón. Exones empalmados
alternativamente se caracterizan a menudo por los sitios de empalme corto y degenerar. La debilidad
intrínseca de estos sitios, lo que refleja la afinidad reducida para las proteínas spliceosomal, es la causa
principal de corte y empalme alternativo. El reconocimiento de los exones alternativos es modulada por
una capa adicional de información proporcionada por un extensas y complejas redes de elementos que
actúan en cis auxiliares (sitio de ARN elementos de secuencia no-empalme), se hace referencia como
potenciadores y silenciadores de empalme, respectivamente, que promueven e inhiben el exón
reconocimiento (Fig. 2A). Estos son los elementos de regulación cortos (~ 10 nucleótidos) que se pueden
encontrar aislados o agrupados en el pre-mRNA y están presentes tanto en los exones (ESEs, Exonic
empalme potenciadores y ESSS, silenciadores exonic empalme) e intrones (ISE, Intronic potenciador
de empalme y ISS, Intronic empalme silenciadores). Los potenciadores de empalme bestcharacterized
son típicamente ricos en purina y la función al proporcionar sitios de unión para serina-arginina (SR)
factores, una familia (alrededor de una docena) de las proteínas de unión a ARN esenciales y abundante
altamente conservadas en las células de plantas y animales. Factores SR muestran múltiples papeles en
constitutiva de empalme y alternativa, así como en otros aspectos de la expresión génica. Todos los
miembros de esta familia comparten una estructura modular que consta de una o dos copias de un motivo
de ARN de reconocimiento N-terminal (RRM), seguido por un dominio C-terminal de longitud variable
rico en serina alterna - dipéptidos arginina (el dominio RS). Los RRM determinar la especificidad de
unión al ARN, mientras que el dominio RS media interacciones específicas entre proteínas que son
esenciales para la contratación del aparato de empalme. Sin embargo, dentro de la spliceosome funcional
también los dominios de RS pueden contactar directamente con el pre-mRNA. El carácter secuencial de
estos contactos sugiere que el dominio de las interacciones de RS con ARN promover spliceosome
asamblea.
Además, los residuos de serina del dominio RS son objetivos de extensas eventos de fosforilación que
influyen en las interacciones de proteínas, y regulan la actividad y la distribución subcelular de las
proteínas SR. Aunque se han mostrado varias quinasas, incluyendo SR proteínas quinasas (SRPKs) 1 y 2,
CLK / STY, de especificidad dual de la tirosina quinasa-regulado, CRKRS, ADN topoisomerasa I,
glucógeno sintasa quinasa-3 y AKT, para fosforilar las proteínas SR, la las vías de transducción de
señales que regulan splicing alternativo son aún poco conocidos.
Se han propuesto varios modelos para la función de ESEs y factores de SR (Fig. 2B). De acuerdo con
uno de estos modelos, las proteínas SR ESE enlazados a promover el exón definición directamente
mediante el reclutamiento de la maquinaria de empalme a través de interacciones específicas proteína-
proteína mediadas por el dominio RS. Otro modelo predice que la principal función de los factores de
SR ESE-consolidados es para antagonizar el efecto negativo en el empalme de una proteína inhibidora
que está enlazado a un elemento silenciador yuxtapuesta (ESS) (modelo de inhibidor). Exón inclusión o
saltarse se determina por el equilibrio de estas actividades de la competencia, que a su vez reflejan por
las concentraciones relativas de la unión al ARN activador cognado y proteínas represoras. Estos
modelos de mejora de empalme no son necesariamente excluyentes, ya que pueden reflejar las
diferentes necesidades en el contexto de diferentes exones.
Silenciadores de empalme identificados hasta la fecha parecen muy diverso. Ellos pueden actuar como
sitios de unión para factores que bloquean el acceso de la maquinaria de empalme a un sitio de

empalme. Entre las proteínas que interactúan con ESS y elementos ISS hay ribonucleoproteínas
nucleares heterogéneas (hnRNP), un grupo de proteínas de unión a ARN inicialmente reconocida como
factores que interactúan con transcripciones de ARN polimerasa II para formar partículas de
hnRNP. En geles bidimensionales se han descrito aproximadamente 30 manchas, denominado con las
letras del alfabeto de hnRNP A1 a través de U. De manera similar a los factores de SR, proteínas hnRNP
tienen una estructura modular en la que uno o más dominios de unión a ARN, generalmente en el extremo
N-terminal, se asocian a diferentes dominios "auxiliares". Tres tipos de dominios de unión a ARN (RRM,
hnRNP K y RGG dominio de homología de dominio, una región de la proteína rica en repeticiones Arg-
Gly-Gly) se han identificado en las proteínas hnRNP y se muestra para proporcionar un cierto nivel de
especificidad de unión de ARN. Los dominios auxiliares son muy diferentes en secuencia y controlar la
localización subcelular y la interacción con otras proteínas. Especificidad de ARN y proteína-proteína
interacciones de unión contribuyen a la asamblea de los complejos de ribonucleoproteína que son los
sustratos para la reacción de empalme subsiguiente. El mecanismo de acción hnRNPs depende de la
posición de sus sitios de unión a lo largo de la pre-ARNm. Además del modelo de inhibidor descrito
anteriormente, unido a hnRNPs ISS que flanquean un exón alternativo puede hacer que el exón a la
salida de bucle, lo que resulta en la omisión del exón (Fig. 2C). Por otra parte, factores inhibidores
unidos a ESS pueden polimerizarse por el exón y desplazar a las proteínas SR ESE determinada
(Fig. 2C).
Sólo en pocos casos los exones empalmados alternativamente son controlados por empalme reguladores
específicos de tejido. En la gran mayoría de los casos de eventos de empalme parecen ser controlado
por la abundancia relativa y / o la actividad de factores SR antagónicas ampliamente expresadas y
proteínas hnRNP a través de un mecanismo de combinatoria, con múltiples factores positivos y negativos
y los elementos de secuencia que influyen en el resultado final del empalme reacción. Esto se ejemplifica
por los efectos antagonistas sobre pre-mRNA de empalme de SF2/ASF, un factor de SR, y las proteínas
hnRNP A1: altos niveles de SF2/ASF inducen la inclusión exón mientras que los altos niveles de hnRNP
A1 promover la omisión de exón. Curiosamente, el nivel relativo de expresión de hnRNP A1 y SF2/ASF
se ha demostrado que cambiar durante el crecimiento de pulmón neoplásico. Otro ejemplo es SRp55 y
su factor de antagonista hnRNP E / PTB que el control del perfil de corte y empalme del factor de
crecimiento de fibroblastos recepto r (FGFR1). Saltarse de resultados α-exón en la producción de la
isoforma FGFR1-β que tiene una mayor afinidad por los factores de crecimiento de fibroblastos. El
aumento de expresión de esta isoforma se correlaciona con el cáncer en el páncreas y el cerebro y con
mal pronóstico en los tumores de mama y su sobreexpresión promueve la formación de tumores en
ratones desnudos. SRp55 se une a una ESE 69-nucleótidos y es necesario para α-exón inclusión mientras
PTB reconoce una secuencia aguas arriba de la α-exón y promueve la omisión de exón y la producción de
FGFR1-β.
Estudios recientes indican que las vías de señalización puede controlar decisiones de empalme al afectar
a la distribución subcelular y / o la actividad de los reguladores de empalme. Muchos factores SR y
proteínas hnRNP servicio de transporte continua y rápidamente entre el núcleo y el citoplasma, el cual
da a conocer una función citoplasmática de estas proteínas, por ejemplo, en la traducción del ARN. En
este sentido, Michlewski mostró que el factor de empalme SF2/ASF estimula la iniciación de la
traducción mediante la contratación directamente el objetivo mamífero de la rapamicina (mTOR) a un
subconjunto de mRNAs. Notablemente, varios tratamientos de estrés perturbar la distribución núcleo-
citoplasma de algunos reguladores de empalme. Por ejemplo, la exposición de las células a estímulos de
estrés, tales como choque osmótico o la irradiación con UVC, da como resultado una marcada acumulación
citoplásmica de hnRNP A1, concomitante con un aumento en la fosforilación de proteínas. Estos efectos
están mediados por la MKK (3/6) - ruta de p38 y se correlacionan con los cambios en el patrón de corte
y empalme alternativo de un E1A reportero de corte y empalme de pre-ARNm de adenovirus. Por otra
parte, varios reguladores de empalme, incluyendo SF2/ASF, son secuestradas en los órganos de

resistencia nuclear después de los tratamientos que activan la respuesta de choque térmico. Esta
capacidad para modular la actividad de los reguladores de empalme se abre la excitante posibilidad de
que condiciones estresantes, como aquellos en el microambiente del tumor, puede influir en el perfil de
empalme de un número de genes y por lo tanto afectar identidad de la célula.
Nuestro conocimiento de los mecanismos moleculares que subyacen a corte y empalme alternativo
todavía es limitado. De hecho, hay muchos elementos exonic y intrónica para que los mediadores que
actúan en trans están aún por identificar y, en la mayoría de los casos, circuitos de regulación que las
decisiones de control de corte y empalme y la conexión con las vías de transducción de señales, no se
conocen bien. Descifrando el complejo entramado que subyace alternativa de empalme elección sitio
regulador sigue siendo un reto importante en esta área de investigación.
ABERRANTE SPLICING ALTERNATIVO EN CÁNCER, GENES RELACIONADOS
De ancho análisis del genoma indican que ~ 75% de los genes humanos codifican al menos dos isoformas
empalmados alternativamente. Por lo tanto, corte y empalme alternativo parece ser la norma para los
genes humanos y no es sorprendente que los defectos de empalme en los genes específicos están
causalmente ligados a trastornos genéticos. Wellcharacterized ejemplos son la atrofia muscular espinal,
distrofia miotónica, la retinitis pigmentosa, síndrome de Frasier, la hemofilia A, * La talasemia, fibrosis
quística atípica y algunas enfermedades neurodegenerativas. Se ha calculado que aproximadamente el
15% de mutaciones puntuales que causan enfermedades hereditarias hacen alterar el procesamiento de
pre-ARNm al afectar canónica 5 'y 3' sitios de empalme, sitios de ramificación o señales de
poliadenilación. Esta cifra es probablemente una subestimación ya que no tiene en cuenta las mutaciones
en elementos reguladores de empalme (potenciadores y silenciadores) que pueden prevenir la
interacción con los reguladores de empalme (hnRNPs y factores SR). De hecho, al menos el 50% de 50
sustituciones de una sola base que causan la omisión de exón en genes humanos interrumpir
potenciadores de empalme. Con frecuencia, los defectos de empalme debido a mutaciones genéticas
introducen codones de parada prematura y dirigen el ARNm de la degradación por la vía de
descomposición mediada por el antisentido (NMD). El efecto final es que, en lugar de un polipéptido
truncado, no se produce ninguna proteína.
En los últimos años ha surgido un vínculo entre el desarrollo del cáncer y la desregulación del splicing
alternativo. Una característica común de las células cancerosas, de hecho, es la desregulación general
de empalme, que puede conducir a la expresión de las variantes específicas de tumores y que es más
probable es promovida por alteraciones en las vías de señalización y por las variaciones en la
concentración, localización y actividad de trans -actuando empalme reguladores. Todavía se discute si
estos cambios en los perfiles de corte y empalme son simplemente un "ruido" que ocurren en las células
cancerosas o tener un papel directo en la tumorigénesis. Como cuestión de hecho, recientes avances en
biología molecular y celular indican que los perfiles de corte y empalme alterados de genes críticos
pueden tener un impacto sobre todos los principales aspectos de la biología de las células del
cáncer. Esto puede ser debido a la inactivación de onco-supresor o para obtener de la función de las
proteínas implicadas en la susceptibilidad al cáncer y en la progresión tumoral.
Las mutaciones que actúan en cis
Estos se heredan o mutaciones somáticas que afectan el proceso de empalme. De acuerdo con su posición
y el efecto sobre empalme, estas mutaciones se pueden dividir en dos subclases. Subclase I (60% de
los casos) de empalme comprende mutaciones en los sitios de ayuste invariantes y abolir completamente
reconocimiento de exón. Estas mutaciones se asocian con enfermedades graves.Subclase II se asocia a
menudo con un fenotipo relativamente suave e incluye mutaciones en los motivos variantes (tales como
el corte y empalme del tracto poli-pirimidina alternativa) y mutaciones intrónicas que generan donante
o aceptor de los sitios crípticos. Un ejemplo proviene del gen p53 supresor de tumores, es decir, el gen

más comúnmente mutado en los cánceres humanos. Las isoformas de p53 se originan a través de splicing
alternativo y se diferencian por su función de supresor de tumor. Curiosamente, las mutaciones
"silenciosas" en el gen p53 se prevé que afectará a corte y empalme, ya que crean sitios de empalme en
el centro de un exón. Vale la pena considerar, sin embargo, que muchos de detección de mutaciones se
centraron en los exones e ignoradas regiones no codificantes. A continuación, se discuten otros ejemplos
de mutaciones que afectan empalme de oncogenes y genes supresores de tumores.
Las mutaciones en el gen APC resultado en la poliposis adenomatosa familiar, así (FAP). Recientemente,
se ha informado de que una forma atenuada de FAP es causada por una inserción de una única T entre
la segunda y la tercera posición del intrón 4 del gen APC que conduce a la omisión del exón 4 y para la
expresión predicho de una proteína truncada . Dos mutaciones adicionales en los intrones 3 y 4 tienen
el efecto idéntico.
También mutaciones en elementos reguladores de empalme, como ESE y las secuencias del SEE, pueden
perturbar perfiles de empalme. Un buen ejemplo es el gen BRCA1 supresor de tumores que codifica una
proteína implicada en una variedad de procesos celulares incluyendo la regulación transcripcional, la
recombinación, la reparación del ADN y la apoptosis de las células. Las mutaciones en el gen BRCA1 son
marcadores bien conocidos de susceptibilidad a cáncer de ovario y de mama, siendo la última la neoplasia
maligna más frecuente entre las mujeres. Mazoyer demostrado que una mutación sin sentido heredado
en el exón 18 interrumpe un elemento ESE (el sitio de unión para el factor de SF2/ASF SR) y provoca
la omisión de exón. El análisis por ordenador con el programa ESEfinder ha identificado 23 ESEs
altamente conservadas en los 22 exones del gen BRCA1. Alrededor del 60% de estos motivos contienen
sustituciones de nucleótidos reportadas en el Breast Cancer Information Core sugiere la posibilidad de
que la orientación de los perfiles de empalme pueden ser los mecanismos a través de los que las
mutaciones afectan la función BRAC1. El gen BRCA1 codifica al menos treinta distintas variantes de
empalme que son expresados diferencialmente en los tejidos. La mayoría de las transcripciones
empalmados alternativamente mantener el marco de lectura abierto del ADNc original y tienen el
potencial para codificar proteínas funcionales. En particular, las células de mama y de ovario expresan
un conjunto común de variantes que sugiere la intrigante posibilidad de que comparten algunas vías de
regulación que en su lugar está ausente en líneas celulares de leucemia que muestran un conjunto
diferente de isoformas. Muchas mutaciones sin sentido en la región de codificación están asociados con
la omisión de exón. Por lo tanto, un punto de mutación doble en el exón 7 de la neurofibromatosis
1 (NF1) gen altera los sitios de unión de consenso para los factores de SR, SC35 y SF2/ASF, y promueve
la omisión de exón. El gen NF1 muestra uno de la tasa de mutación más alta entre los genes asociados
con trastornos humanos; simplemente sobre la base de la secuencia de ADN, se predijo que el 37% de
estas mutaciones genómicas podría determinar defectos de empalme. Sin embargo, cuando esta
predicción se verificó mediante el análisis de los mRNAs maduros, se hizo evidente que hasta el 50% de
las mutaciones del gen NF1 están asociados con defectos de empalme. El mensaje negativo que surge de
este análisis es que todavía no somos capaces de inferir el perfil empalme simple de la secuencia primaria
del ADN y que no tenemos que depender de ARN e incluso secuencias de la proteína para entender los
efectos de cualquier mutación genética.
Similares observaciones se han hecho con una serie de otros genes asociados a tumores como el BRCA2,
FHIT, KIT, MLH1, MDM2, MSH2 y LKB1. En particular, todos estos casos, y muchos otros que se
describen de forma continua, representan una extensión del concepto según el cual la progresión del
cáncer se debe a una serie de mutaciones genéticas estables que perturban la estructura, la función o
la abundancia de proteínas críticas. En este sentido, defectos de empalme pueden ser vistos como una
de las rutas a través de mutaciones genéticas que causan la tumorigénesis. Sin embargo, corte y empalme
alternativo de los oncogenes o supresores tumorales también podría verse afectada por mutaciones en
reguladores de empalme que implica una actividad de estos factores como oncoproteínas o supresores

tumorales, dependiendo de sus funciones antagónicas sobre la selección del sitio de empalme. Por
ejemplo, el gen SRp55 (SFR6) se encuentra mutado en cánceres de mama y colorrectal. Curiosamente,
SRp55 controles el perfil de empalme de varios genes asociados a tumores, entre los
que CD44 y KIT. Además algunas proteínas hnRNP se han clasificado como oncogenes. En el 90% de los
liposarcomas mixoides humanos, el (12; 16) t translocación genera una fusión entre el gen hnRNP P2, que
codifica una proteína multifuncional implicada en la transcripción, corte y empalme y exportación de
ARNm, y el gen CHOP, que codifica para una CCAAT / proteína de unión al potenciador implicado en la
eritropoyesis, la diferenciación de adipocitos, la detención del crecimiento y G1-S progresión del ciclo
celular. El producto de la fusión de hnRNP P2-CHOP contiene el dominio amino terminal de la
transcripción de activación de hnRNP P2 y el dominio de unión a ADN de CHOP y su sobre-expresión en
ratones desnudos resultados en la formación de tumores.
Los cambios en los factores que actúan en trans
Más interesante, desde nuestro punto de vista, es la observación, reportado por muchos estudios en los
últimos 20 años, que la mayoría de las alteraciones de empalme asociados con el cáncer no se asocian
con cambios de nucleótidos en los genes afectados, lo que implica modificaciones en la expresión y / o
actividad de factores de regulación de empalme. De hecho, los cambios en el repertorio de los factores
de SR y proteínas hnRNP se producen con frecuencia en los tumores y se acompañan de alteraciones en
la abundancia relativa de los productos de corte y empalme alternativos, una firma típica de las células
cancerosas con efectos predecibles sobre el comportamiento celular. Como cuestión de hecho, las líneas
de células cancerosas muestran un alto nivel de eventos de empalme alternativos que no se han
conservado entre el ser humano y el ratón y no se expresan en tejidos normales el fortalecimiento de
la idea de que un cambio en el nivel de empalme de los reguladores en células de cáncer puede afectar
gravemente en programas de expresión génica.
Un ejemplo destacado para ilustrar la forma en corte y empalme aberrante en el cáncer puede ser
modulada por la actividad alterada o expresión de factores de empalme es proporcionado porCD44, una
glicoproteína transmembrana involucrados-las interacciones célula-célula y célula-matriz. Varias
isoformas de CD44 se generan a través de la incorporación de la variable de 10 exones alternativos (v1-
v10) en su dominio extracelular proximal. Estándar CD44, que carecen todos los exones alternativos, se
expresa predominantemente en los tejidos normales, mientras que las isoformas de CD44, en particular,
los que contienen variante exones V5, V6 y V7, están sobre-expresan en diversos tumores y han sido
implicados en la invasión de células tumorales y la metástasis.La producción de las diferentes isoformas
de CD44 se relaciona con los cambios en la abundancia de las proteínas SR y varios factores de
empalme (incluyendo hnRNPA1, SRp55, SF2/ASF, Tra-2 beta, YB-1 y Sam68) que han demostrado para
regular los exones variantes específicas. Varias líneas de evidencia indican que CD44 empalme está
regulada en respuesta a estímulos extracelulares.Por lo tanto, la Materia y col. [68] encontró que la
proteína de unión de ARN Sam68 (Src asociada en la mitosis 68-kDa) promueve v5 inclusión en respuesta
a la activación de la vía de Ras-Raf-Mek-Erk. Aunque los detalles mecanicistas son incompletos, se ha
sugerido que la interacción de fosforilados Sam68 con el exón v5 bloquea la actividad represiva de
hnRNP A1, ya sea mediante la prevención de la interacción de hnRNP A1 con el elemento de ESS (por
impedimento estérico), o contrarrestando el inhibitorio efecto de hnRNP A1 une a la
ESS. Recientemente, Cheng y Sharp identificados SRm160 como otro factor de empalme Ras regulado
responsable de inclusión de exón v5 en CD44 transcripciones. Es importante destacar, que también
mostraron que el silenciamiento de SRm160 disminuye la invasividad celular, que une este regulador de
empalme a la tumorigénesis.
Otro buen ejemplo de modulación de corte y empalme por vías de señalización viene del gen de la
fibronectina (FN), un componente de la matriz extra-celular y un factor determinante en el control de
la proliferación, la migración, invasión y metástasis comportamiento de las células tumorales. EDA es

una isoforma de empalme FN generado por la inclusión de un solo exón (exón EDA, también conocido
como EDI o EIIIA). Esta isoforma de empalme está pobremente expresado en tejidos normales de
adultos, mientras que está presente en los embriones, así como durante la curación de heridas y en
algunos tumores. La inclusión del exón EDA es desencadenada por la activación de la quinasa / AKT
Ras/PI3- por factores de crecimiento. AKT fosforila directamente la SR proteínas 9G8 y SF2/ASF,
que a su vez se unen a la exón EDA y promover su reconocimiento por parte del aparato de empalme. La
activación de la misma vía por la insulina regula la actividad de otra proteína SR, SRp40, y estimula la
inclusión de un exón alternativo en la proteína quinasa C (PKC) II pre-ARNm. En total, estos resultados
apoyan la posibilidad de que la desregulación de la vía de Ras/PI3-kinase/AKT, por ejemplo como
resultado de mutaciones en sus componentes, podría tener consecuencias dramáticas en el perfil de
empalme de cualquier pre-mRNAs regulados por 9G8, SF2/ASF, SRp40 y tal vez otras proteínas SR. Una
hipótesis atractiva es que los exones que responden a esta vía de señalización pertenecen a un conjunto
de genes que funcionan cooperativamente para modular la fisiología de la célula de acuerdo con la función
biológica de la molécula de señalización. La identificación de estos exones, por lo tanto, será de vital
interés. Una indicación a favor de esta hipótesis proviene de la observación de que AKT también
promueve traducción de EDA mRNAs obligado por 9G8 fosforilados y SF2/ASF. Por lo tanto, la
activación de una sola vía de transducción de señales controla de manera integrada tanto empalme y la
traducción de los ARNm específicos y estimula la producción de proteínas específicas. El efecto final
es un aumento drástico tanto en la velocidad y la fuerza de la respuesta de señalización tal como se
mide por la producción de la proteína inducida.
Aunque estos ejemplos proporcionan ideas interesantes en los efectos de las vías de transducción de
señales en la regulación de empalme, la caracterización molecular sigue siendo escasa y todavía no se ha
proporcionado la aclaración de la vía completa. Por lo tanto, además de para ilustrar una función de
transducción de señales en el control de empalme, estos casos también sugieren la necesidad de estudios
adicionales para dilucidar este importante mecanismo de regulación de genes.
Un cuerpo creciente de datos implica corte y empalme alternativo como un mecanismo para el control
de la apoptosis o muerte celular programada, un proceso esencial en el desarrollo y en el mantenimiento
de la homeostasis celular en los organismos multicelulares. Bcl-X es un miembro de la familia Bcl II que
dirige desglose mitocondrial durante la apoptosis. El uso de 5 'sitios de empalme alternativos dentro de
exón 2 determina la producción de dos isoformas de proteínas: una forma larga antiapoptótica (Bcl-XL)
y una proteína corta la promoción de la apoptosis (Bcl-XS).Por lo tanto, un cambio en el patrón de
empalme de estas transcripciones puede tener efectos profundos sobre la actividad proliferativa de
las células cancerosas y de su respuesta a los tratamientos pro-apoptóticos. Varios factores de
empalme, incluyendo Sam68, SF2/ASF, hnRNP F / H y SAP155, contribuyen en el control de la elección
entre los dos sitios 5'-empalme alternativo.En particular, Sam68 sobre-expresión promueve la
producción de pro-apoptótica Bcl-XS y este efecto se revierte a Sam68 fosforilación. Por el contrario,
las proteínas hnRNP F / H, mediante la unión a un elemento tramo rico en G, promover el uso de la Bcl-
XS - 5 'del sitio de empalme. La relación de Bcl-X variantes de empalme contribuir a determinar la
sensibilidad de las células a una amplia variedad de agentes apoptóticos y puede tener importancia en la
resistencia a las drogas y la capacidad de respuesta quimioterapéutico. Por ejemplo, la ceramida lipídica,
un mediador / regulador de la apoptosis promueve la expresión de las variantes de empalme pro-
apoptóticas Bcl-XS, la elección entre los dos sitios 5'-alternativas de empalme está controlada por un
elemento sensible a la ceramida (CRCE 1) situado en el exón 2 y obligados por SAP155. Por otra parte,
la ceramida es capaz de modular el estado de fosforilación de las proteínas SR en una fosfatasa 1 (PP1)
forma-dependiente. Curiosamente, uno de los objetivos es el PP1 SR factor de SF2/ASF, otro
importante regulador de la Bcl-X procesamiento pre-mRNAs.

Hay varios ejemplos de eventos de empalme alternativos que controlan la actividad de las proteínas
implicadas en la motilidad celular y la invasión, un pre-requisito para la formación de metástasis de
cáncer. Este es el caso de las isoformas de empalme de la andrógenos y receptores de estrógenos que
están involucrados en carcinomas mamarios. Curiosamente, una isoforma de receptor de estrógeno alfa,
debido a la omisión del exón 3 (delta3ER), es un activador más potente del factor de crecimiento
endotelial vascular que el receptor de tipo salvaje. El Rac1b variante de empalme, que se genera por la
inclusión de un exón casete 57-nucleótidos, se ha demostrado que conducen a un crecimiento celular
independiente de anclaje. En particular, Rac1b es hasta reguladas en tumores colorrectales en las
diversas etapas de la progresión neoplásica, en comparación con los tejidos normales adyacentes. Otros
ejemplos, que muestran claramente el potencial efecto funcional de corte y empalme aberrante en la
tumorigénesis, son el receptor del factor de crecimiento de fibroblastos 2 ( FGFR2 ), la fibronectina y
la survivina .
Recientemente, se ha utilizado el Ron (recepteur d'origine nantais) proto-oncogén como modelo para
investigar la relación entre el splicing alternativo y la progresión tumoral [40]. Ron, la tirosina quinasa
del receptor humano para la proteína macrophagestimulating (MSP), es una proteína heterodimérica
(p185-Ron) compuesta de subunidades α y β tanto derivadas de la transformación de un precursor
común. La unión a MSP estimula la actividad tirosina quinasa intrínseca de Ron y resultados en la
fosforilación de su sitio de acoplamiento para el transductor múltiple y adaptador de proteínas que
conducen a la activación de cascadas de señalización (Fig. 3A ). Junto con Met, el receptor del factor
de crecimiento de hepatocitos (HGF), Ron pertenece a una subfamilia de receptores de tirosina quinasas
(RTK) con los patrones de expresión únicos y actividades biológicas. Además de promover el crecimiento
celular y la protección de la apoptosis, estos receptores de célula de control de la disociación, la
motilidad y la invasión de matrices extracelulares, un proceso conocido como el crecimiento invasivo''''
o'''' dispersión de células [84]. Crecimiento invasivo es fisiológicamente relevante durante el desarrollo,
la organogénesis y la regeneración de tejidos, pero también es importante para mediar en la invasividad
y para promover la progresión maligna.Actualmente, se han identificado seis variantes incluyendo
RonΔ170, Δ165, Δ160, Δ155, Δ110, y Δ55 con diversas deleciones o truncamientos en las regiones
extracelular o intracelular. Todas estas variantes son constitutivamente activo, pero difieren en sus
propiedades bioquímicas y biológicas. Por otra parte, el perfil de corte y empalme del gen Ron se altera
con frecuencia en los cánceres epiteliales, tales como cánceres de colon y de mama, lo que sugiere que
la producción de múltiples isoformas Ron podría contribuir a la patogénesis de estos tumores. El exceso
de expresión de cualquiera de estas isoformas aumenta la motilidad celular (dispersión actividad
similar). Sin embargo, sólo RonΔ160 o RonΔ155 son capaces de inducir la formación de foco, sostenido
crecimiento independiente de anclaje y la capacidad para formar tumores metastásicos en ratones. Este
potencial oncogénico se canaliza a través de la vía de la PI3-quinasa / AKT. Transcripciones se reunió
también sufren splicing alternativo y una isoforma, llamado Met-SM, se origina en la omisión del exón
14, que codifica un segmento de 47 aminoácidos en el dominio yuxtamembrana. Esta isoforma se ha
demostrado recientemente que desempeñar un papel importante en el desarrollo y la progresión de los
cánceres humanos. Entre los mecanismos que controlan la expresión de las diferentes isoformas de Ron
en las células cancerosas, el interruptor de constitutiva de corte y empalme alternativo desempeña el
papel principal. Por lo tanto, la aclaración de las vías de regulación que controlan el perfil de empalme
de las transcripciones Ron arrojará nueva luz sobre la iniciación y progresión del cáncer. Hemos
estudiado en detalle el caso de corte y empalme alternativo que conduce a la producción de
Δ Ron ARNm. Esta transcripción carece de un exón de 147-pb (exón 11). La proteína codificada tiene
una 49-amino-ácido en la deleción en marco del dominio extracelular que afecta a la maduración
proteolítica y resulta en la acumulación de una sola cadena de pro-RonΔ165 en el citoplasma. Por otra
parte, la proteína eliminado se caracteriza por puentes disulfuro intracelulares aberrantes que facilitan
RonΔ165 oligomerización que conduce a la fosforilación y activación constitutiva. Hemos demostrado
que la elección entre la inclusión y la omisión del exón 11 es controlada por dos elementos reguladores

adyacentes, un silenciador y un potenciador, ambos ubicados en el exón constitutiva 12 (Fig. 3B ). La
unión de la proteína SR SF2/ASF al ESE estimula la omisión del exón 11 y la producción de ΔRon. Por lo
tanto, la sobreexpresión de SF2/ASF en las células que normalmente expresan Ron desencadena la
producción de la RonΔ165 con dramáticas consecuencias sobre las propiedades de la célula. En efecto,
de manera similar a lo observado después ΔRon sobre-expresión, una expresión aumentada de SF2/ASF
afecta profundamente la morfología celular y provoca la acumulación nuclear de β-catenina, la
reorganización del citoesqueleto de actina, y la baja regulación de la E-cadherina, un tumor y supresor
de la invasión en los carcinomas humanos. Todos estos cambios morfológicos y moleculares representan
características de la transición epitelial a mesenquimal (EMT), que está implicada en la diseminación
metastásica de los carcinomas humanos (Fig. 3B ). En particular, desmontables de SF2/ASF por
interferencia de ARN (ARNi) reduce los niveles de ΔRon y de forma concomitante disminuye la motilidad
celular. Del mismo modo, el ARNi dirigido específicamente contra Δ Ron reduce la motilidad celular y
revierte parcialmente los cambios morfológicos inducidos por SF2 / ASF sobre-expresión.
Dada la informó sobre regulación de varias proteínas SR, incluyendo SF2/ASF, durante la progresión
del tumor, es tentador especular que el factor de empalme SF2/ASF podría promover la transformación
maligna induciendo una ΔRon mediada EMT. Esta hipótesis es consistente con un informe reciente que
muestra que SF2/ASF se comporta como un auténtico protooncogén.SF2/ASF es hasta reguladas en un
conjunto de tumores humanos y, el gen que codifica para SF2/ASF se amplifica específicamente en
algunos tumores de mama, pero no en el tejido de mama normal del mismo paciente. Limitado sobre-
expresión de SF2/ASF, comparable a la de las muestras de cáncer, dio lugar a la formación de tumores
en ratones desnudos. En particular, varios objetivos de corte y empalme endógenas de SF2/ASF, entre
ellos una nueva isoforma oncogénico del sustrato mTOR, S6K1, son esenciales para la transformación
SF2/ASFmediated. Además, ARN de interferencia (ARNi) de SF2/ASF o la oncogénico S6K1 isoforma,
dio lugar a la reversión del fenotipo transformado.
La identificación de los reguladores de empalme como una nueva clase de proto-oncogenes puede revelar
nuevos aspectos de la biología del cáncer y ofrecer nuevas oportunidades para el diagnóstico y la terapia.
Aunque todos los ejemplos presentados anteriormente, sin duda, demostrar que los cambios en la
actividad de corte y empalme de las células tumorales está vinculada la progresión del tumor, sin
embargo, los mecanismos moleculares responsables de estos cambios permanecen indefinidos. Varias
preguntas siguen merecen investigación. ¿Cuáles son los eventos que conducen a la desregulación de la
expresión y / o actividad de los miembros de SR y familias hnRNP? ¿Hay una reorganización generalizada
de la maquinaria de empalme en las células de cáncer o hace el resultado interruptor de corte y empalme
de una desregulación de un subconjunto de los reguladores de corte y empalme? ¿De qué manera las vías
de transducción de señales que cooperen para afectar a los perfiles de empalme para determinar la
conversión maligna?
Firmas splicing alternativo como indicadores de diagnóstico / pronóstico
potenciales y perspectivas de terapias
En las secciones anteriores hemos discutido varios ejemplos que ilustran un vínculo causal entre el
empalme y el desarrollo de neoplasia alternativa. Vale la pena notar que en muchos casos isoformas de
empalme parecen ser específica del cáncer. Por otra parte, existen evidencias de que en diferentes
tejidos de un conjunto diferente de genes se somete a cambios de los perfiles de corte y empalme
durante la tumorigénesis. Por lo tanto, en los tumores renales, Tropomyosin 1 , Actinin α 1 , la
integrina β 4 , catenina δ 1 y de crecimiento de fibroblastos receptor del factor 2 cambiar sus patrones
de splicing alternativo, mientras que N-glycanase1 no es así, a pesar de sus aparentes alteraciones en
los tumores de pulmón y útero. Estos descubrimientos argumentan en contra de la idea de que los
cambios en los perfiles de corte y empalme pueden ser resultado de una alteración generalizada de la
maquinaria de empalme en las células de cáncer y sugieren que sólo un conjunto específico de empalme

reguladores se ven afectados por la tumorigénesis de una manera tissuespecific. La identificación de
tissuespecific isoformas específicas de cáncer, empalmados alternativamente inmediatamente incitó a
su uso potencial como diagnóstico, pronóstico, o biomarcadores predictivos. Aunque los estudios a gran
escala aún no se han llevado a cabo, los resultados iniciales son prometedores. Una correlación
importante entre aberrante splicing alternativo y la progresión del tumor se ha demostrado
para CD44. En particular, las isoformas que contienen el exón v6 de la variante son upregulated
frecuentemente en carcinomas de cabeza y cuello o en cánceres avanzados gástrico en la etapa
y CD44v6 tumores positivos se asocian a mal pronóstico.También, aberrante corte y empalme alternativo
de MDM2 se encontraron transcripciones de asociarse con un factor de pronóstico pobre para la
supervivencia en pacientes con cáncer de mama, mientras que HDMX se ha relacionado con sarcoma de
tejidos blandos. Por otra parte, la expresión alterada de una variante empalmada del P73 , una proteína
relacionada con p53, parece ser un marcador de pronóstico negativo en pacientes con neuroblastoma.
Durante la última década clásico perfil de la expresión genética realizado por microarray ha sido una
poderosa herramienta para el descubrimiento de nuevos biomarcadores de cáncer. Sin embargo, la
mayoría de las plataformas de gama desarrollado hasta la fecha no han sido diseñados para distinguir
isoformas de ARNm. Una dirección importante para el futuro será la aplicación de todo el
genoma pantallas diseñado para el análisis de firmas de corte y empalme alternativos asociados con
cáncer, pero prácticamente ausente en las células normales. Estas micromatrices de splicingsensitive
tienen el potencial de identificar mejor sub-clases de tumores. Por otra parte, no requieren que las
relaciones causa-efecto entre los perfiles de empalme y los cánceres se identifican (o incluso que
existían!), Sino sólo que esta asociación son suficientemente consistentes para ser
predictivo. Curiosamente, Li ha utilizado un enfoque para cuantificar simultáneamente los cambios en el
empalme ("splicing interruptor") y la abundancia de transcripción (de arriba a abajo-regulación). La
ventaja de esta estrategia de perfiles "bidimensional'' es que funciona con muestras biológicas
parcialmente degradadas, tales como ARN derivados de los bloques de tejido que han sido formalinfixed
y embebido en parafina. Estos autores identificaron un conjunto específico de biomarcadores de
isoformas de mRNA del cáncer de próstata mediante paneles independientes de muestras de tejido. En
particular, informaron de dos variantes de AMACR gen (que codifica una-metilacil-CoA racemasa)
resultantes de la utilización alternativa del último exón junto con ción poliadenilación alternativa: una
isoforma mostró un máximo de regulación cuantitativa en el cáncer de próstata en comparación con la
próstata normal, tejidos, el otro parecían ser expresado sólo en el cáncer de próstata. Más
recientemente, la novela de la tecnología de microarrays se ha utilizado para medir todo el genoma exón
expresión en 102 muestras normales y de cáncer de tejidos de diferentes etapas de colon, vejiga
urinaria, y la próstata. Se identificaron siete genes con variantes específicas de tumores de empalme
( alfa-actinina 1 , Caldesmon , Collagen, type VI alpha 3 , ricos en leucina repetir interacción proteína
2 , fosfatidilinositol-4-quinasa beta polipéptido catalítico , Tropomyosin 1 y Vinculina ). En particular,
los patrones de empalme tumorspecific de alfa-actinina 1 , caldesmón y Vinculina se encontraron, los
componentes clave del citoesqueleto, en los tres órganos que sugieren que pueden representar eventos
de splicing relacionadas con el cáncer en general. Por otra parte, en el análisis de proteínas in silico
predice que estas variantes de empalme del cáncer-específicos identificados codifican proteínas con
funciones potencialmente alteradas, lo que indica que pueden estar implicados en la patogénesis y por
lo tanto representan nuevos biomarcadores y dianas potenciales.
Variantes del cáncer-específicas de corte y empalme no sólo pueden servir como marcadores de
diagnóstico y pronóstico de tumores, sino que también proporcionan objetivos potenciales para el
desarrollo de nuevas estrategias terapéuticas (Fig. 4 ). Una vía prometedora para el desarrollo de
medicamentos contra el cáncer más selectivos consiste en la administración dirigida de compuestos
bioactivos para el tumor por medio de moléculas (por ejemplo, anticuerpos) que son específicos para los
marcadores asociados a tumores. La accesibilidad, así como el patrón de restricción de la expresión de

la antígeno es un criterio importante en la selección de un objetivo para la intervención bio-molecular. En
particular, muchos receptores que median las interacciones célula-célula y célula-matriz son regulados
por corte y empalme alternativo y empalmados alternativamente variantes específicas de estas
moléculas se asocian con muchos tumores malignos humanos. Curiosamente, casi el 90% de eventos de
empalme alternativos afecta a las regiones situadas en la superficie expuesta de la proteína y para
muchas moléculas de la membrana celular, los exones alternativos codifican nuevos epítopos que se
encuentran generalmente en el dominio extracelular de la proteína. Estos epítopos parecen ser ideal
para las estrategias de tumor de orientación. Una vez más, la proteína CD44, discutido en detalle en la
sección anterior, es un ejemplo notable ya que los anticuerpos marcados radiactivamente dirigidos
contra el CD44-v6 isoforma parecen ofrecer una herramienta terapéutica prometedora y están
actualmente en ensayos clínicos para el tratamiento del cáncer de cabeza y cuello. Sin embargo, a pesar
de estos avances prometedores, los tumores sólidos son con frecuencia relativamente resistentes a las
terapias basadas en anticuerpos. Esto es debido, en parte, a la relativa inaccesibilidad de las células
tumorales y a la escasa penetración de los anticuerpos en el tejido tumoral. Dado que las células
tumorales se separan de la sangre por las células endoteliales y los componentes de la matriz
extracelular que rodea la vasculatura, la captación del tumor es muy limitado por la capacidad del
anticuerpo para cruzar esta capa. Es importante destacar que, los enfoques terapéuticos dirigidos
contra la vasculatura del tumor neo-han ganado significativa para un número de razones: (i) las células
endoteliales vasculares tumorales son fácilmente accesibles para los medicamentos a través de la
circulación de la sangre; (ii) existe una creciente evidencia de que la inhibición o regresión de los vasos
del tumor conduce a la muerte celular del cáncer ya que se basan en los vasos sanguíneos de nutrientes
y oxígeno para satisfacer sus necesidades metabólicas. Un ejemplo interesante es el EDB isoforma de
corte y empalme alternativo de fibronectina (FN). El EDB isoforma se genera a través de la inclusión
de un solo exón que codifica una región de 91 aminoácidos y está presente en los vasos sanguíneos de
tejidos neoplásicos, mientras que está virtualmente ausente en tejidos maduros / adulto. EDB está
implicada en la regulación de la proliferación celular endotelial y la morfogénesis vascular; un anticuerpo
radiomarcado EDB-específica se encuentra actualmente en ensayos clínicos de fase II para el
tratamiento del cáncer anti-angiogénica. Recientemente, se ha demostrado que el corte y empalme
alternativo del dominio extra-A (EDA) de la fibronectina puede representar a otro, igualmente
atractivos, objetivo para la entrega basada en anticuerpos de agentes bioactivos a la neovasculatura no
sólo de tumores sólidos, pero también de lesiones metastásicas . Del mismo modo, las variantes de corte
y empalme podrían representar nuevos antígenos específicos de leucemia con uso potencial en enfoques
inmunoterapéuticos. En leucemias Phpositive, la (9; 22) t translocación genera la Bcr / Abl proteínas de
fusión, cuya activación de la tirosina quinasa constitutiva es responsable de la aparición del fenotipo de
la leucemia. Además de la principal BCR / ABL transcritos de fusión, se ha informado de que BCR /
ABL transcripciones, que surge de corte y empalme alternativo, también se producen en un alto
porcentaje de la leucemia mielógena crónica (LMC) y leucemia linfoblástica aguda (LLA) de los
pacientes. En particular, BCR / ABL transcripciones alternativas que implican ABL exón 4 son muy
atractivos porque las proteínas de fusión resultantes contienen una novela epítopo inmunogénico que es
capaz de provocar una respuesta específica de antígeno de células T.
Enfoques terapéuticos innovadores también podrían ser diseñados para apuntar a la maquinaria de
empalme. Por ejemplo, modificaciones post-traduccionales de empalme de los reguladores pueden ser
relevantes para la progresión tumoral. SRPK1 es hasta reguladas en tumores de mama y de colon en
comparación con el epitelio normal adyacente, y los niveles de la quinasa aumentar junto con el grado
del tumor. Fuerte expresión de la proteína SRPK1 también fue evidente en la mayoría de los de mama y
las líneas celulares tumorales de colon. Curiosamente, la baja regulación de SRPK1 el uso de pequeños
ARN de interferencia (siRNA) afecta a la expresión de factores apoptóticos clave BAX y BCL2, aumenta
la proporción de las células tumorales (pero no de las células nontrasformed) sometidas a la apoptosis y
amplifica su sensibilidad a los dos quimioterapéuticos utilizados frecuentemente agentes tales como

gemcitabina y cisplatino. Estos resultados sugieren que la inhibición farmacológica de SRPK1 actividad
puede ser eficaz como agente independiente o en combinación con regímenes quimioterapéuticos
convencionales. Por otra parte, las pequeñas moléculas con SRPK-bloqueo de la actividad, tales como
inhibidores de Clk1/Sty o inhibidores potentes de la reacción de corte y empalme, tales como derivados
de indol, se pueden utilizar para alterar los patrones de empalme. Curiosamente, los datos recientes
muestran que las proteínas E (SME) Funciones spliceosomal como supresor del crecimiento de células
tumorales de manera independiente de p53. Por lo tanto, teniendo en cuenta que una gran parte de los
cánceres humanos son defectuosos en la actividad de p53, la orientación de las PYME en la terapia
génica contra el cáncer será de especial interés.
Diferentes enfoques terapéuticos están estudiando actualmente para corregir aberrante de corte y
empalme alternativo a nivel del ARN. Estas estrategias pueden o bien alterar los patrones de splicing
alternativo de genes específicos o reconocer especies de ARNm particulares para provocar su
degradación (por ejemplo, ARNi, microARN y ribozimas). La corrección de defectos de empalme a través
de la manipulación de ARNm requiere el uso de estrategias de genetherapy, que a menudo están
relacionadas con problemas tales como la entrega, la toxicidad y la inmunogenicidad . terapia SiARN
basada ha demostrado una gran promesa para muchas enfermedades tales como el cáncer. Los
principales objetivos para la terapia siRNA incluyen oncogenes y genes que están involucrados en la
angiogénesis, la metástasis, la supervivencia celular, Antiapoptosis y la resistencia a la quimioterapia. Por
otra parte, las últimas generaciones de antisentido OLIGON ucleotides que contienen modificaciones
químicas aparecen más estable en comparación con cadenas principales de nucleótidos
convencionales. Algunos de estos oligos de evitar montajes ribosómico, y por lo tanto mRNA traducción,
y parece ser bien tolerada en los pacientes. Además, los oligonucleótidos modificados sintéticamente
dirigidos a través de la hibridación específica de secuencia de los sitios de empalme o secuencias
adyacentes pueden bloquear la selección inadecuada de exón mediante la inhibición de la unión
de trans factores de acción prolongada. Se han aplicado con éxito para corregir corte y empalme
aberrante de la β -globina , CFTR , distrofina , tau genes y para reprimir la generación de variantes de
corte y empalme relacionadas con el cáncer de FGFR1 , Bcl-X , y MDM2. Sola o en combinación con
agentes quimioterapéuticos, tales como cisplatino, irinotecan, paclitaxel, fluorouracilo se puede utilizar
con soltura en la terapia. Por último, hemos diseñado un oligómero de morfolino fosforodiamidato
dirigidas a la región de la Ron potenciador que contiene el sitio de unión SF2/ASF (Mo-SF2) para
bloquear estéricamente la unión de este factor de empalme y, por lo tanto, la omisión del exón 11. La
eficacia de Mo-SF2 se ensayó usando células de carcinoma gástrico KATOIII humanas caracterizadas
por altos niveles de SF2/ASF, ΔRon y fenotipo invasivo. Hemos encontrado que Mo-SF2 inhibe
Δ Ron empalme y corrige la morfología celular y las propiedades de migración de las células [datos no
publicados].
OBSERVACIONES FINALES
Después de la secuenciación de la totalidad de los genomas eucariotas, corte y empalme alternativo se
ha convertido en la estrategia dedicada a la interpretación de la información genética. Como
ampliamente discutido en esta revisión, de hecho, este mecanismo de procesamiento de cuentas pre-
mRNA de una gran parte de la diversidad proteómica en eucariotas superiores. Mientras regulación de
la transcripción determina la presencia y abundancia de transcripciones de genes primarios, corte y
empalme alternativo está diseñado para manipular el mensaje en respuesta a una serie de estímulos. De
esta manera se puede sintonizar las proteínas características, tales como la capacidad de participar a
hetero-complejos o la localización subcelular, con el fin de satisfacer las necesidades de las
células. Raramente son los programas de corte y empalme alternativos asociados a "sí o no" decisiones;
en la mayoría de los casos, los productos de corte y empalme co-existir en una sola célula y contribuir a
la capacidad celular para hacer frente a un número de diferentes estímulos internos y externos. Desde
este punto de vista, por lo tanto, este mecanismo parece estar íntimamente conectada a la complejidad

de los organismos metazoos que, una vez más, pone de relieve la necesidad de entender las reglas
combinatorias que subyacen opciones del sitio de empalme como un paso obligatorio para llegar a una
visión global de la lógica de la vida cuestión.
Descifrar las implicaciones biológicas de este proceso de regulación es una tarea difícil que requiere la
integración de una gran cantidad de datos en relación con los cis secuencias de acción ytrans -actuando
factores involucrados, los circuitos de regulación y vías de señalización que modulan las decisiones de
empalme sitio y finalmente la consecuencias fisiológicas y / o patológicas resultantes de la expresión
de las distintas isoformas de empalme. El logro de este resultado será sin duda acelerada por el
desarrollo reciente de nuevos y muy sofisticados bioinformática y enfoques de biología de sistemas.
Se espera que un avance importante para comprender completamente cómo splicing alternativo impregna
los programas de expresión para derivar a partir de la aplicación de metodologías de alto
rendimiento. Durante la última década, las ciencias biomédicas han sido fuertemente influenciados por
las "ómicas": la genómica, la proteómica, transcriptómica y metabolómica. La aplicación de los enfoques
de alto rendimiento para estudiar los perfiles de empalme es sólo en su comienzo. Este retraso es sin
duda debido a una serie de problemas técnicos que se derivan de la necesidad de evaluar contemporánea
para cada gen, el nivel absoluto de los transcritos totales y la abundancia relativa de las isoformas de
empalme. Por otra parte, la interpretación es complicada significativamente por la necesidad de
comprender las implicaciones fisiológicas que resultan de un cambio en los perfiles de corte y empalme.
Sin embargo, es fácil predecir que este tipo de análisis proporcionará información crucial para dar a
conocer las vías de regulación que subyacen a la co-regulación de los perfiles de empalme y para
comprender la relevancia de splicing alternativo en el contexto del desarrollo del organismo. La
esperanza es que las matrices splicingsensitive guiarán la identificación de los circuitos que, al igual que
la transducción de señales y las vías de transcripción, puede ser causalmente vinculados a los programas
de desarrollo, organogénesis, definición de esquema corporal y la identidad celular.Otra cuestión
pendiente se refiere a la identificación de la sub-genoma que no experimentan eventos de splicing
alternativo. ¿Estos genes identificar cualquier función celular crítico particular?Por lo tanto, el cambio
de perspectiva, desde la caracterización detallada de los mecanismos moleculares a enfoques globales,
que se espera que mejore nuestra comprensión de la importancia fisiológica de corte y empalme
alternativo y para aumentar drásticamente la comprensión de las condiciones fisiológicas y patológicas
importantes, tales como la plasticidad neuronal y la complejidad del cáncer. Estamos seguros de que los
enfoques de la biología de sistemas pueden ayudar a la identificación de los eventos de splicing
alternativo que pueden desempeñar un papel fundamental en la progresión tumoral. Esto ofrecerá la
oportunidad de desarrollar estrategias innovadoras de intervención terapéutica que se dirigen a
variantes específicas de splicing alternativo.
AGRADECIMIENTOS
Este trabajo fue apoyado por becas de la Associazione Italiana per la Ricerca sul Cancro (AIRC), de la
Red de la Unión Europea (EURASNET) de Excelencia sobre splicing alternativo (EURASNET) y de la
Fondazione Cariplo a GB

Figura (1). cis secuencias de acción
necesarios para la reacción de empalme y
los diferentes tipos de eventos de splicing
alternativo. (A) de empalme secuencias
consenso de un gen eucariota típica (señales
exón / intrón empalme sitio, sitio de sucursal
y el tracto polypyrimidine). (B) mRNAs
empalmados alternativamente el resultado
de la omisión de exón, intrón retención, el uso
de la alternativa 3 '(aceptante) o 5'-
(donante) y los sitios de la selección de los
exones que se excluyen mutuamente. En el
nivel de proteína, corte y empalme
alternativo afecta drásticamente a la
secuencia de aminoácidos por deleción o
inserción de dominios, marco-turnos o
codones de parada. Splicing alternativo en
las regiones no codificantes del ARNm
maduro podría repercutir en la estabilidad y
la traducción del ARNm.
Figura (2). Cis-y elementos
reguladores que actúan en trans que
el control de corte y empalme
alternativo y modelos para la función
de empalme potenciadores y
silenciadores. (A) los exones
empalmados alternativamente por lo
general se caracterizan por los sitios
de empalme débiles. El reconocimiento
de estos sitios depende de elementos
reguladores de empalme: potenciadores
exonic empalme (ESE) y silenciadores
(ESS) y intronic potenciadores de
empalme (ISE) y silenciadores
(ISS). (B) los elementos ESE están
obligados por los factores de empalme
de la familia SR. través
de interacciones con las proteínas del
aparato de empalme, el dominio RS de
factores SR estimula el ensamblaje de
la splicesome en el intrón adyacente (a
la izquierda).Además, los factores SR pueden contrarrestar la actividad inhibidora de proteínas hnRNP
unidos a elementos ESS (derecha). (C) El mecanismo de acción de los elementos de silenciador depende
de la su posición a lo largo de la premRNA. En algunos casos, factores inhibidores (por ejemplo proteínas
hnRNP) se unen a secuencias que flanquean ISS un exón alternativo y causan un bucle y saltar fuera de
este exón (a la izquierda). Alternativamente, los factores inhibidores unidos a ESS pueden polimerizar
a lo largo del exón y desplazar las proteínas SR ESEbound (derecha).
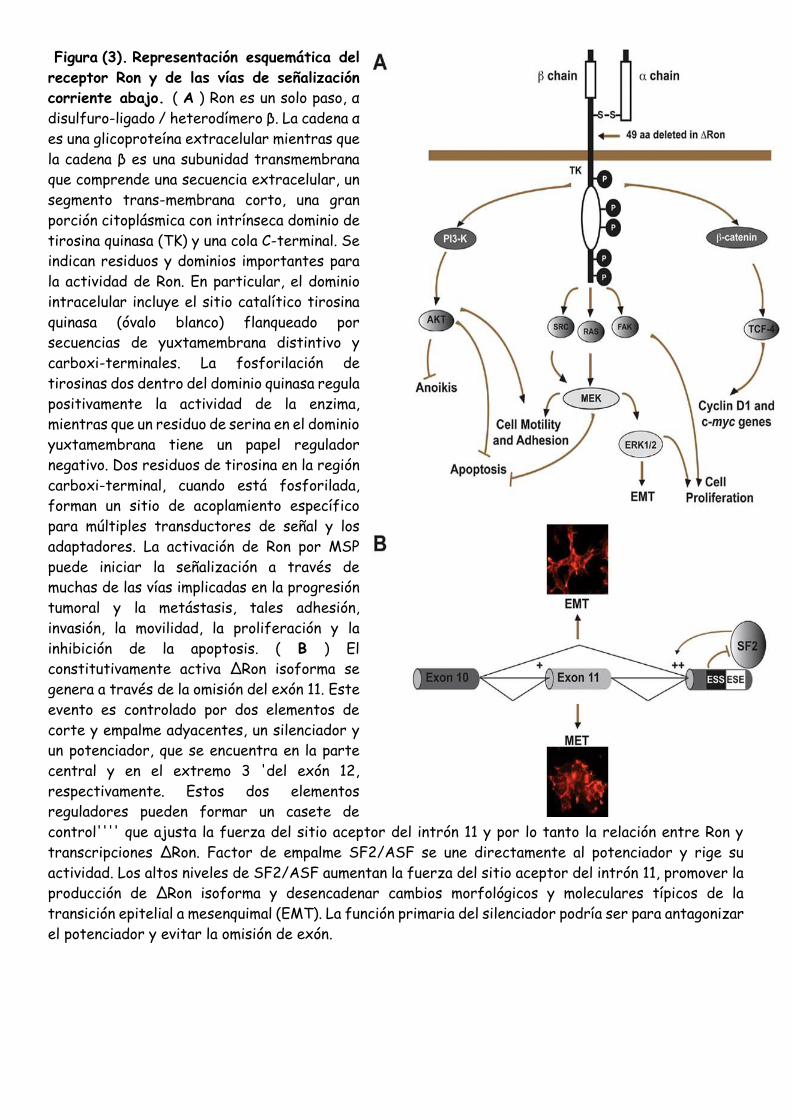
Figura (3). Representación esquemática del
receptor Ron y de las vías de señalización
corriente abajo. ( A ) Ron es un solo paso, α
disulfuro-ligado / heterodímero β. La cadena α
es una glicoproteína extracelular mientras que
la cadena β es una subunidad transmembrana
que comprende una secuencia extracelular, un
segmento trans-membrana corto, una gran
porción citoplásmica con intrínseca dominio de
tirosina quinasa (TK) y una cola C-terminal. Se
indican residuos y dominios importantes para
la actividad de Ron. En particular, el dominio
intracelular incluye el sitio catalítico tirosina
quinasa (óvalo blanco) flanqueado por
secuencias de yuxtamembrana distintivo y
carboxi-terminales. La fosforilación de
tirosinas dos dentro del dominio quinasa regula
positivamente la actividad de la enzima,
mientras que un residuo de serina en el dominio
yuxtamembrana tiene un papel regulador
negativo. Dos residuos de tirosina en la región
carboxi-terminal, cuando está fosforilada,
forman un sitio de acoplamiento específico
para múltiples transductores de señal y los
adaptadores. La activación de Ron por MSP
puede iniciar la señalización a través de
muchas de las vías implicadas en la progresión
tumoral y la metástasis, tales adhesión,
invasión, la movilidad, la proliferación y la
inhibición de la apoptosis. ( B ) El
constitutivamente activa ΔRon isoforma se
genera a través de la omisión del exón 11. Este
evento es controlado por dos elementos de
corte y empalme adyacentes, un silenciador y
un potenciador, que se encuentra en la parte
central y en el extremo 3 'del exón 12,
respectivamente. Estos dos elementos
reguladores pueden formar un casete de
control'''' que ajusta la fuerza del sitio aceptor del intrón 11 y por lo tanto la relación entre Ron y
transcripciones ΔRon. Factor de empalme SF2/ASF se une directamente al potenciador y rige su
actividad. Los altos niveles de SF2/ASF aumentan la fuerza del sitio aceptor del intrón 11, promover la
producción de ΔRon isoforma y desencadenar cambios morfológicos y moleculares típicos de la
transición epitelial a mesenquimal (EMT). La función primaria del silenciador podría ser para antagonizar
el potenciador y evitar la omisión de exón.

Figura (4). Uso potencial de los enfoques terapéuticos que se dirigen a splicing alternativo. Varias
estrategias se están utilizando actualmente para explotar splicing alternativo para el tratamiento de
cáncer. (A) En muchos casos, las variantes de corte y empalme del cáncer-restringidas contienen
epítopos únicos que podrían ser utilizados como dianas para los anticuerpos específicos conjugados a
toxinas de células del tumor. (B) Se han encontrado muchos compuestos químicos que afectan empalme
de numerosos genes. Aunque los mecanismos por los cuales los patrones de empalme se alteran son aún
poco conocidos, varios compuestos son capaces de bloquear la actividad de las quinasas de proteínas SR-
(SRpKs) y, en consecuencia, reducir el estado de fosforilación de factores de empalme
SR. (C oligonucleótidos) modificados sintéticamente, tales como fosforotioato, morfolino
phosphorodiamide y 2'-O-metil, son capaces de bloquear la interacción de la maquinaria spliceosome con
un sitio específico. Por otra parte, son más estables y activas de cadenas principales de nucleótidos
regulares y presentan baja toxicidad in vivo. (D) transcritos de ARNm específico de cáncer que
contienen secuencias únicas pueden ser dirigidos utilizando la degradación mediada por ARNi.