the application of synchrotron radiation induced x-ray emission in the measurement of zinc and lead...
TRANSCRIPT

The application of synchrotron radiation induced X-rayemission in the measurement of zinc and lead in Wistarrat ameloblasts
Manish Arora a,*, Brendan J. Kennedy b, Chris G. Ryan c, Ross A. Boadle d,D. Murray Walker a, Cathy L. Harland e, Barry Lai f, Zhonghou Cai f,Stefan Vogt f, Hans Zoellner a, Sheena W.Y. Chan a
aCellular and Molecular Pathology Research Unit, Department of Oral Pathology, Faculty of Dentistry, University of Sydney,
Westmead Centre for Oral Health, Westmead Hospital, Westmead, NSW 2145, Australiab School of Chemistry, University of Sydney, NSW 2006, AustraliacCSIRO Exploration and Mining, Geosciences, Building 28E, Monash University, Clayton, VIC. 3168, AustraliadElectron Microscope Unit, ICPMR, Westmead Hospital, Westmead, NSW 2145, AustraliaeAustralian Synchrotron Research Program, Australian Nuclear Science and Technology Organisation, Lucas Heights, NSW, AustraliafAdvanced Photon Source, Argonne National Laboratory, 9700 S. Cass Avenue, Argonne, IL 60439, USA
a r c h i v e s o f o r a l b i o l o g y 5 2 ( 2 0 0 7 ) 9 3 8 – 9 4 4
a r t i c l e i n f o
Article history:
Accepted 6 April 2007
Keywords:
Synchrotron radiation
X-ray emission
Ameloblasts
Wistar rats
Trace elements
Zinc
Lead
a b s t r a c t
The development of analytical techniques for the measurement of trace elements in cellular
compartments of developing teeth remains an important methodological issue in dental
research. Recent advances in third generation synchrotron facilities have provided high
brilliance X-ray sources that can be effectively used to study trace element distributions in
small spatial regions with low detection limits. The present study describes for the first time
the application of synchrotron radiation induced X-ray emission (SRIXE) in measuring the
distribution of zinc and lead in the ameloblasts of developing Wistar rat teeth. Wistar rats
were fed a standard rat diet, containing the normal dietary requirements of zinc, ad libitum
and exposed to 100 ppm of lead in drinking water. Resin embedded sections of first
mandibular molars were analysed using a 13.3 keV incident monochromatic X-ray beam
focussed to a 0.2 mm spot. Characteristic X-rays arising from the entire thickness of the
sample were measured using an energy dispersive detector for quantitative analysis of
elemental concentrations. The results showed that intranuclear concentrations of zinc were
greater than levels in the cytoplasm. Furthermore, nuclear and cytoplasmic concentrations
of zinc in the maturation stage (742 � 27 and 424 � 25 ppm, respectively) were significantly
higher than the zinc levels observed in the nucleus and cytoplasm of presecretory stage
ameloblasts (132 � 10 and 109 � 10 ppm, respectively) ( p < 0.05). A clear lead signal above
the background was not detected in the ameloblasts and lead concentrations could only be
reliably measured in the developing enamel. Overall, SRIXE was an effective method of
studying the spatial distribution of zinc in the cells of developing teeth and offered a unique
combination of sub-micron spatial resolution and parts-per-million detection limits (0.8–1
and 0.6–1 ppm for zinc and lead, respectively).
# 2007 Elsevier Ltd. All rights reserved.
avai lable at www.sc iencedi rec t .com
journa l homepage: www. int l .e lsev ierhea l th .com/ journals /arob
* Corresponding author. Tel.: +61 2 9845 7892.E-mail addresses: [email protected] (M. Arora), [email protected] (B.J. Kennedy), [email protected]
(C.G. Ryan), [email protected] (R.A. Boadle), [email protected] (C.L. Harland), [email protected] (B. Lai), [email protected](Z. Cai), [email protected] (S. Vogt), [email protected] (H. Zoellner).
0003–9969/$ – see front matter # 2007 Elsevier Ltd. All rights reserved.doi:10.1016/j.archoralbio.2007.04.003

a r c h i v e s o f o r a l b i o l o g y 5 2 ( 2 0 0 7 ) 9 3 8 – 9 4 4 939
1. Introduction
Zinc is an essential dietary element and serves numerous
biological functions including protein synthesis, maintenance
of neuro-sensory functions, cell-mediated immunity, thyroid
function as well as bone formation and mineralisation.1–3
There is also an increasing body of evidence supporting the
role of zinc in the development and maintenance of dental
tissues. Data from animal models shows that restriction of
dietary zinc increases the incidence of dental caries,4 reduces
the lipid content of developing enamel, and causes morpho-
logical alterations in ameloblasts.5 In addition, a number of
zinc-finger transcription factors have recently been identified
in developing mouse tooth germs and current evidence
suggests that they serve an important role in cytodifferentia-
tion of enamel epithelium and amelogenesis.6–9 In contrast to
zinc, lead exerts an adverse effect on developing teeth, with
lead exposed rats showing an increased incidence of dental
caries, disruption of salivary gland function, as well as delays
in enamel mineralisation and tooth eruption.10–12 The positive
association of lead exposure to dental caries has also been
confirmed in epidemiological studies.13–14
In addition to the individual effects of lead and zinc on
oral and systemic health, a number of studies suggest an
interaction between these elements. Co-administration of
zinc is known to reduce tissue and organ lead concentrations
in animal models.15–17 Furthermore, lead may displace zinc
from a number of zinc-binding sites including d-aminolevu-
linic acid dehydratase (ALAD), metallothioneins and some
transcription factors.18–20
An important methodological issue in understanding the
oral health effects of zinc, lead and other metal toxins is
the development of suitable analytical techniques to measure
the spatiotemporal distribution of theseelements in the cellular
compartments of developing tooth germs. While bulk chemical
methods of analysis including inductively coupled plasma-
mass spectrometry (ICP-MS), inductively coupled plasma-
optical emission spectrometry (ICP-OES) and atomic absorption
spectrometry (AAS), are capable of providing detection limits in
the parts-per-million (ppm) or parts-per-billion (ppb) range,
these methods are not suitable for determining the spatial
distribution of trace elements. Techniques such as energy
dispersive X-ray analysis (EDAX), electron probe analysis (EP)
and nuclear microprobe analysis (NMP) have proven useful in
measuring the intracellular concentrations of trace elements in
various cell types.21–23 These methods are, however, limited by
either high detection limits or poor spatial resolution. Over the
past two decades, developments in third generation synchro-
tron sources have provided high intensity X-ray beams that can
be effectively used to study trace element distribution in small
spatial regions with low detection limits.
In synchrotron radiation induced X-ray emission (SRIXE)
analysis, the high-energy X-rays generated by an undulator
source are focussed into a sub-micron spot on the sample,
where they create vacancies in the inner shells of the target
atoms by photoelectric absorption. When these vacancies are
filled by electrons from outer shells, characteristic X-rays are
emitted that are measured in an energy dispersive detector for
quantitative analysis of elemental concentrations.24 For the
purposes of trace element analysis, SRIXE offers a unique
combination of sub-micron resolution and detection limits in
the ppm range or better. While this technology is rapidly
gaining acceptance in many areas of science, its application in
the measurement of trace element concentrations in dental
soft tissues remains largely unexplored. The primary aim of
this study was to measure the change in intracellular
distribution of lead and zinc during ameloblast differentiation
from presecretory to early maturation stage. To our knowledge
this paper describes for the first time the application of SRIXE
in measuring the intracellular distribution of zinc and lead in
ameloblasts of developing Wistar rat teeth. The advantages
and limitations of this technique are also discussed.
2. Materials and methods
2.1. Animal handling
Three female Wistar rats, aged 12–14 weeks, of normal weight
were mated with similarly aged male Wistar rats of normal
weight.25 The animals received 100 ppm of lead nitrate (Asia
Pacific Speciality Chemicals, NSW, Australia) in the drinking
water and were fed a standard rodent diet containing 40 mg/kg
of zinc (AIN93G, Glen Forrest Stockfeeders, Western Australia)
ad libitum. Lead exposure was commenced 5 days before the
expected birth of the pups. The pups were housed with their
mothers, fed only by lactation and had no access to the
drinking water.
Groups of three rat pups were sacrificed by decapitation
from each of the litters at birth and on the third and seventh
day after birth. These specific time points were chosen
because the presecretory and secretory stages of ameloblast
development can be observed in a rat first molar tooth germ at
birth and 3 days after birth, while the early maturation stage of
ameloblast development can be visualised on day-7.26 The
continuously erupting rat incisor has been widely used as a
model for tooth development. Rat molars were used in the
present analyses, however, to permit the measurement of
trace elements in radicular tissues, which was the aim of a
parallel-running project. All procedures in the present study
were approved by the Animal Ethics Committee, Westmead
Hospital, NSW, Australia (Ethics clearance no. 124.06).
2.2. Preparation of tissue sections
Immediately after culling, the mandibles were removed using
sterile surgical instruments. The samples were processed in a
manner similar to that reported by Dillon et al.27 for SRIXE
analysis of hamster lung cells. First mandibular molar tooth
germs were removed under a dissecting microscope and fixed
overnight at 4 8C in Karnovsky’s fixative, followed by washing
in MOPS buffer (3-(N-morpholino)-propane-sulphonic acid).
The samples were dehydrated with graded ethanols (50–100%)
(Selby Biolab, Australia) for 15 min each at room temperature.
Following dehydration, tooth germs were placed in a 50:50
mixture of low viscosity Spurr’s resin (Polysciences Inc., USA)
and ethanol for 1 h at room temperature. The tooth germs
were then transferred to a 90:10 mixture of Spurr’s resin and
ethanol overnight. The final infiltration was in Spurr’s resin at
70 8C for 10 min, and this was repeated three times. For

Fig. 1 – Zinc concentrations in the nucleus and cytoplasm of
presecretory, secretory and early maturation stage
ameloblasts measured using synchrotron radiation
induced X-ray emission. From two tooth germs, 6–8 cells
were analysed per stage. Nuclear concentrations of zinc
were significantly different between all three
developmental stages ( p < 0.05). However, cytoplasmic
concentrations of zinc only showed a significant increase
in the early maturation stage ( p < 0.05).
a r c h i v e s o f o r a l b i o l o g y 5 2 ( 2 0 0 7 ) 9 3 8 – 9 4 4940
embedding, tooth germs were placed individually in large
embedding moulds (Leica Microsystems, Austria) and orien-
tated to ensure that the buccal surfaces of respective tooth
germs were parallel to the cutting faces of the resin blocks.
Spurr’s resin was then placed in the mould using a dropper
and care was taken to ensure that orientation of the tooth
germs remained unchanged. The moulds were placed in a
sealed container and the resin was polymerised at 70 8C
for 10 h.
Serial sections (1.0–1.5 mm) of the embedded tooth germ
samples were cut using an ultramicrotome (Reichert-Jung
Ultracut E, Austria). Adjacent sections were either stained with
methylene blue or mounted on an aluminium sample holder
prepared at the Advanced Photon Source, Argonne National
Laboratory, Illinois, USA. The aluminium mounts were 3.5 cm
long, 0.5 cm wide and two holes, 2 mm in diameter, were
present at one end of the mount. Gold finder grids (Prositech,
Australia) were placed on one hole to serve as reference points.
A thin coating of 0.3% (w/v) Formvar 15/95E (Monsanto
Chemicals, USA) was placed over the second hole to provide
support to the resin-embedded tissue section. Photomicro-
graphs of methylene blue-stained sections were used to
identify ameloblasts for SRIXE analysis. Using these photo-
micrographs as guides, tissue sections on aluminium mounts
were imaged under optical phase contrast microscopy and co-
ordinates for regions of interest were recorded. The finder
grids on each aluminium mount served as fixed reference
points. The co-ordinates recorded under phase contrast
microscopy were then transferred to the motorised SRIXE
sample stage, permitting accurate positioning of the samples.
To further confirm the location of the ameloblasts, coarse
SRIXE scans were conducted prior to high-resolution scans
(see following section on SRIXE Analysis). To avoid sample
contamination, tooth germ sections were mounted on the
aluminium holders only 5 days prior to the scheduled SRIXE
analysis. Mounted samples were stored in sterile tissue
culture dishes (Becton Dickinson, NJ, USA) and transported
to the Advanced Photon Source, Argonne National Laboratory,
Illinois, USA.
2.3. SRIXE analysis
SRIXE experiments were performed on the 2-ID-D beamline at
the Advanced Photon Source. All experiments were conducted
under a helium atmosphere to eliminate the argon K-shell
fluorescence signal and to reduce absorption of low-Z
fluorescence by air, and thus improve the signal-to-noise
ratio of the acquired data. Fluorescence lines were collected
using a Canberra germanium detector (Canberra, Meriden, CT,
USA). Full spectra were acquired at every scan position and the
K and L shell fluorescence signals were used for zinc and lead,
respectively.
A 13.3 keV incident monochromatic X-ray beam was
focussed to a 0.2 mm spot. The regions of interest were located
using the coordinates determined by light microscopy. Coarse
scans, with step sizes of 1–5 mm, were conducted to locate
individual cells in the X-ray microprobe. Detailed maps
were then collected using 0.2–0.5 mm steps and 2–10 s dwell
times per pixel. Mapped areas varied in dimensions from
15 mm � 20 mm to 80 mm � 80 mm. Because the tissue sections
analysed in the present study were 1.0–1.5 mm in thickness,
the emitted radiation was derived from the entire thickness of
the samples. The standard reference material (SRM) 1832
(National Institute of Standards and Technology, USA) was
used during the calibration process. SRM 1832 consists of a
silica-based glass film that has been deposited onto a
polycarbonate filter and is primarily used as a standard in
X-ray fluorescence spectrometry. Data analysis was con-
ducted using GeoPIXE II software (Commonwealth Scientific
and Industrial Research Organisation, Australia) and MS Excel
2000 (Microsoft Corporation, USA).
2.4. Statistical analysis
For each developmental stage the data from the two tooth
germs analysed were pooled and differences in element
concentrations between the different stages were analysed
using Kruskal–Wallis test at p < 0.05 using Stata (Intercooled
Stata 9.0; Stata Corporation, College Station, Texas, USA)
3. Results
Results of the SRIXE analysis of the developing rat tooth germs
are presented in relation to the stages of ameloblast devel-
opment in Figs. 1–3. The secretory and maturation stage
ameloblasts were analysed in detail with fine scans. However,
due to time restrictions, presecretory ameloblasts could only
be analysed using a coarse scan. A total of six tooth germ
sections from six different animals were analysed. The
method detection limits were 0.8–1 ppm for zinc and 0.6–
1 ppm for lead.
Mean zinc concentrations in nuclei and the cytoplasm
immediately adjacent to the nuclei in ameloblasts during the
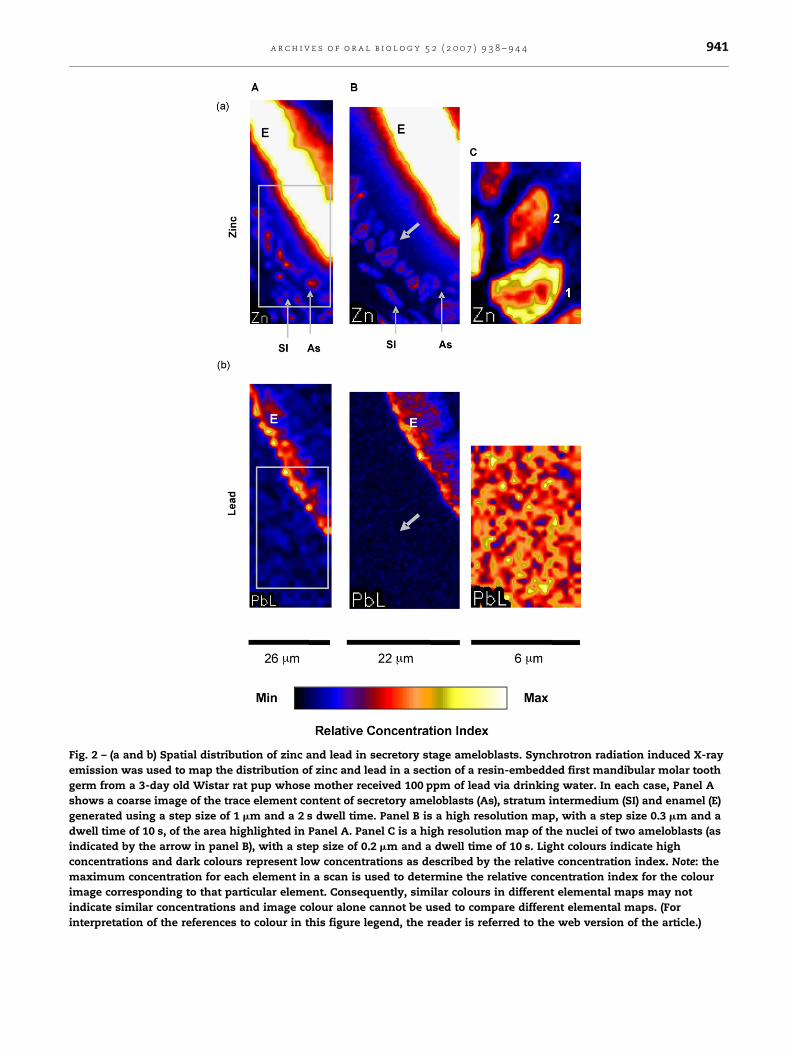
Fig. 2 – (a and b) Spatial distribution of zinc and lead in secretory stage ameloblasts. Synchrotron radiation induced X-ray
emission was used to map the distribution of zinc and lead in a section of a resin-embedded first mandibular molar tooth
germ from a 3-day old Wistar rat pup whose mother received 100 ppm of lead via drinking water. In each case, Panel A
shows a coarse image of the trace element content of secretory ameloblasts (As), stratum intermedium (SI) and enamel (E)
generated using a step size of 1 mm and a 2 s dwell time. Panel B is a high resolution map, with a step size 0.3 mm and a
dwell time of 10 s, of the area highlighted in Panel A. Panel C is a high resolution map of the nuclei of two ameloblasts (as
indicated by the arrow in panel B), with a step size of 0.2 mm and a dwell time of 10 s. Light colours indicate high
concentrations and dark colours represent low concentrations as described by the relative concentration index. Note: the
maximum concentration for each element in a scan is used to determine the relative concentration index for the colour
image corresponding to that particular element. Consequently, similar colours in different elemental maps may not
indicate similar concentrations and image colour alone cannot be used to compare different elemental maps. (For
interpretation of the references to colour in this figure legend, the reader is referred to the web version of the article.)
a r c h i v e s o f o r a l b i o l o g y 5 2 ( 2 0 0 7 ) 9 3 8 – 9 4 4 941
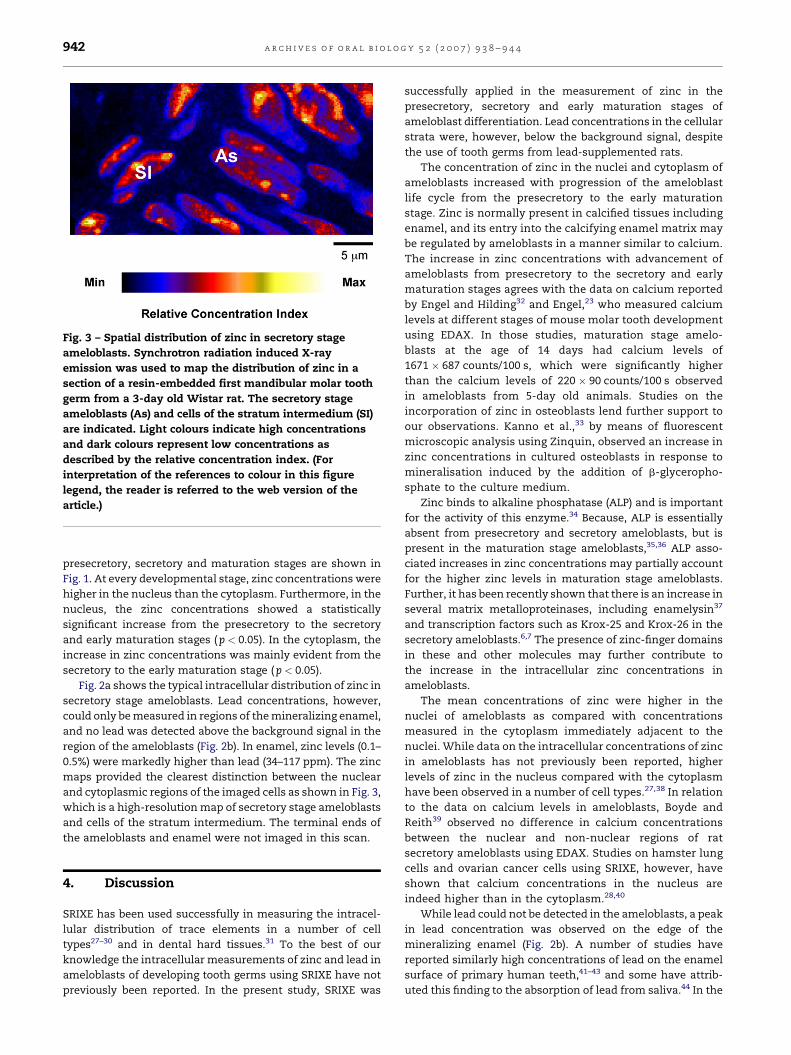
Fig. 3 – Spatial distribution of zinc in secretory stage
ameloblasts. Synchrotron radiation induced X-ray
emission was used to map the distribution of zinc in a
section of a resin-embedded first mandibular molar tooth
germ from a 3-day old Wistar rat. The secretory stage
ameloblasts (As) and cells of the stratum intermedium (SI)
are indicated. Light colours indicate high concentrations
and dark colours represent low concentrations as
described by the relative concentration index. (For
interpretation of the references to colour in this figure
legend, the reader is referred to the web version of the
article.)
a r c h i v e s o f o r a l b i o l o g y 5 2 ( 2 0 0 7 ) 9 3 8 – 9 4 4942
presecretory, secretory and maturation stages are shown in
Fig. 1. At every developmental stage, zinc concentrations were
higher in the nucleus than the cytoplasm. Furthermore, in the
nucleus, the zinc concentrations showed a statistically
significant increase from the presecretory to the secretory
and early maturation stages (p < 0.05). In the cytoplasm, the
increase in zinc concentrations was mainly evident from the
secretory to the early maturation stage (p < 0.05).
Fig. 2a shows the typical intracellular distribution of zinc in
secretory stage ameloblasts. Lead concentrations, however,
could only be measured in regions of the mineralizing enamel,
and no lead was detected above the background signal in the
region of the ameloblasts (Fig. 2b). In enamel, zinc levels (0.1–
0.5%) were markedly higher than lead (34–117 ppm). The zinc
maps provided the clearest distinction between the nuclear
and cytoplasmic regions of the imaged cells as shown in Fig. 3,
which is a high-resolution map of secretory stage ameloblasts
and cells of the stratum intermedium. The terminal ends of
the ameloblasts and enamel were not imaged in this scan.
4. Discussion
SRIXE has been used successfully in measuring the intracel-
lular distribution of trace elements in a number of cell
types27–30 and in dental hard tissues.31 To the best of our
knowledge the intracellular measurements of zinc and lead in
ameloblasts of developing tooth germs using SRIXE have not
previously been reported. In the present study, SRIXE was
successfully applied in the measurement of zinc in the
presecretory, secretory and early maturation stages of
ameloblast differentiation. Lead concentrations in the cellular
strata were, however, below the background signal, despite
the use of tooth germs from lead-supplemented rats.
The concentration of zinc in the nuclei and cytoplasm of
ameloblasts increased with progression of the ameloblast
life cycle from the presecretory to the early maturation
stage. Zinc is normally present in calcified tissues including
enamel, and its entry into the calcifying enamel matrix may
be regulated by ameloblasts in a manner similar to calcium.
The increase in zinc concentrations with advancement of
ameloblasts from presecretory to the secretory and early
maturation stages agrees with the data on calcium reported
by Engel and Hilding32 and Engel,23 who measured calcium
levels at different stages of mouse molar tooth development
using EDAX. In those studies, maturation stage amelo-
blasts at the age of 14 days had calcium levels of
1671 � 687 counts/100 s, which were significantly higher
than the calcium levels of 220 � 90 counts/100 s observed
in ameloblasts from 5-day old animals. Studies on the
incorporation of zinc in osteoblasts lend further support to
our observations. Kanno et al.,33 by means of fluorescent
microscopic analysis using Zinquin, observed an increase in
zinc concentrations in cultured osteoblasts in response to
mineralisation induced by the addition of b-glyceropho-
sphate to the culture medium.
Zinc binds to alkaline phosphatase (ALP) and is important
for the activity of this enzyme.34 Because, ALP is essentially
absent from presecretory and secretory ameloblasts, but is
present in the maturation stage ameloblasts,35,36 ALP asso-
ciated increases in zinc concentrations may partially account
for the higher zinc levels in maturation stage ameloblasts.
Further, it has been recently shown that there is an increase in
several matrix metalloproteinases, including enamelysin37
and transcription factors such as Krox-25 and Krox-26 in the
secretory ameloblasts.6,7 The presence of zinc-finger domains
in these and other molecules may further contribute to
the increase in the intracellular zinc concentrations in
ameloblasts.
The mean concentrations of zinc were higher in the
nuclei of ameloblasts as compared with concentrations
measured in the cytoplasm immediately adjacent to the
nuclei. While data on the intracellular concentrations of zinc
in ameloblasts has not previously been reported, higher
levels of zinc in the nucleus compared with the cytoplasm
have been observed in a number of cell types.27,38 In relation
to the data on calcium levels in ameloblasts, Boyde and
Reith39 observed no difference in calcium concentrations
between the nuclear and non-nuclear regions of rat
secretory ameloblasts using EDAX. Studies on hamster lung
cells and ovarian cancer cells using SRIXE, however, have
shown that calcium concentrations in the nucleus are
indeed higher than in the cytoplasm.28,40
While lead could not be detected in the ameloblasts, a peak
in lead concentration was observed on the edge of the
mineralizing enamel (Fig. 2b). A number of studies have
reported similarly high concentrations of lead on the enamel
surface of primary human teeth,41–43 and some have attrib-
uted this finding to the absorption of lead from saliva.44 In the

a r c h i v e s o f o r a l b i o l o g y 5 2 ( 2 0 0 7 ) 9 3 8 – 9 4 4 943
present study, however, uptake of lead from saliva is unlikely
as the tooth germs had not erupted into the oral cavity. High
levels of lead in the superficial layers of enamel in unerupted
rat molars, measured using a nuclear microprobe, have been
previously reported.45 It is possible that the presence of lead in
the outer layers of enamel is due to ionic exchange with
calcium in the apatite lattice, similar to the exchange of
fluoride ions.46 Lead uptake in enamel may also be influenced
by considerable changes in the enamel organ during early
maturation, which is known to coincide with variations in the
uptake of calcium.47,48 However, the inability of the present
study to reliably detect the intracellular distribution of lead in
the ameloblasts precludes further analysis of the cause of high
concentrations of lead found at the mineralizing front of
enamel using this methodology. One limitation to the
approach used is that sample preparation requires fixation,
dehydration and resin infiltration. Each step may affect
concentrations of unbound trace elements, potentially con-
founding the data. However, zinc concentrations are unlikely
to be affected by this, as intracellular zinc is largely in bound
form.49
Despite these limitations, the present study demon-
strates the value of SRIXE as an effective and high-resolution
method for the measurement of intracellular trace elements
in developing rodent tooth germs. In comparison to EDAX,
EP and NMP, SRIXE offers a unique combination of low
detection limits and sub-micron resolution.50 It is also a non-
destructive technique and shows limited susceptibility to
matrix effects when analysing thin tissue sections as used in
the present study. The main limitations of SRIXE include the
long data collection times, limited access to synchrotron
facilities, higher detection limits when compared with
neutron activation analysis, and laser-ablation inductively
coupled plasma-mass spectrometry. SRIXE, however, offers
significantly better spatial resolution than these other
techniques.
In summary, the results presented here have shown that
SRIXE is a powerful technique for the determination of trace
elements in the cellular compartment of developing tooth
germs. Concentrations of zinc in the ameloblasts of the Wistar
rat tooth germs increased from the presecretory to early
maturation stage, and nuclear concentrations of this element
were higher than levels in the cytoplasm.
Acknowledgements
The authors thank Dr. Ross Mathews and the technical staff at
the Westmead Animal Holding, Westmead Hospital, Sydney,
for their help with animal handling, and the staff of the
Electron Microscope Unit, Westmead Hospital, Sydney, for
their assistance in sample preparation. Dr. Arora was
supported by the Early Career Development Award from the
College of Health Sciences, University of Sydney. This work
was supported by the Australian Synchrotron Research
Program, which is funded by the Commonwealth of Australia
under the Major National Research Facilities Program. Use of
the Advanced Photon Source was supported by the US
Department of Energy, Office of Science, Basic Energy
Sciences, under Contract no. W-31-109-Eng-38.
r e f e r e n c e s
1. Meunier N, O’Connor JM, Maiani G, Cashman KD, Secker DL,Ferry M, et al. Importance of zinc in the elderly: the ZENITHstudy. Eur J Clin Nutr 2005;59(Suppl. 2):S1–4.
2. Cuevas LE, Koyanagi A. Zinc and infection: a review. AnnTrop Paediatr 2005;25(3):149–60.
3. Frederickson CJ, Koh JY, Bush AI. The neurobiology of zinc inhealth and disease. Nat Rev Neurosci 2005;6(6):449–62.
4. Cerklewski FL. Effect of suboptimal zinc nutrition duringgestation and lactation on rat molar tooth composition anddental caries. J Nutr 1981;111(10):1780–3.
5. Goldberg M, Carreau JP, Driessens FC, Septier D. Zincdeficiency-induced changes in the lipid composition andultrastructure of rat incisor teeth. Arch Oral Biol1990;35(4):255–64.
6. Ganss B, Teo W, Chen H, Poon T. Krox-26 is a novel C2H2zinc finger transcription factor expressed in developingdental and osteogenic tissues. Connect Tissue Res 2002;43(2/3):161–6.
7. Lee SK, Kim YS, Lee SS, Lee YJ, Song IS, Park SC, et al.Molecular cloning, chromosomal mapping, andcharacteristic expression in tooth organ of rat and mouseKrox-25. Genomics 2004;83(2):243–53.
8. Jheon A, Chen J, Teo W, Ganss B, Sodek J, Cheifetz S.Temporal and spatial expression of a novel zinc fingertranscription factor, AJ18, in developing murine skeletaltissues. J Histochem Cytochem 2002;50(7):973–82.
9. Bouwman P, Gollner H, Elsasser HP, Eckhoff G, Karis A,Grosveld F, et al. Transcription factor Sp3 is essential forpost-natal survival and late tooth development. Embo J2000;19(4):655–61.
10. Watson GE, Davis BA, Raubertas RF, Pearson SK, Bowen WH.Influence of maternal lead ingestion on caries in rat pups.Nat Med 1997;3(9):1024–5.
11. Gerlach RF, Cury JA, Krug FJ, Line SR. Effect of lead on dentalenamel formation. Toxicology 2002;175(1–3):27–34.
12. Gerlach RF, Toledo DB, Novaes PD, Merzel J, Line SR. Theeffect of lead on the eruption rates of incisor teeth in rats.Arch Oral Biol 2000;45(11):951–5.
13. Gemmel A, Tavares M, Alperin S, Soncini J, Daniel D, Dunn J,et al. Blood lead level and dental caries in school-agechildren. Environ Health Perspect 2002;110(10):A625–30.
14. Moss ME, Lanphear BP, Auinger P. Association of dentalcaries and blood lead levels. JAMA 1999;281(24):2294–8.
15. Cerklewski FL. Influence of dietary zinc on lead toxicityduring gestation and lactation in the female rat. J Nutr1979;109(10):1703–9.
16. Bandhu HK, Dani V, Garg ML, Dhawan DK. Hepatoprotectiverole of zinc in lead-treated, protein-deficient rats. Drug ChemToxicol 2006;29(1):11–24.
17. Batra N, Nehru B, Bansal MP. The effect of zincsupplementation on the effects of lead on the rat testis.Reprod Toxicol 1998;12(5):535–40.
18. Jones KW, Gordon BM, Hanson AL, Kwiatek WM, Pounds JG.X-ray fluorescence with synchrotron radiation.Ultramicroscopy 1988;24:313–28.
19. Pass D, Freeth G. The rat. ANZCCART News 1993;6:1–4.20. Bernard A, Lauwerys R. Metal-induced alterations of delta-
aminolevulinic acid dehydratase. Ann NY Acad Sci1987;514:41–7.
21. Kagi JH, Kojima Y. Chemistry and biochemistry ofmetallothionein. Experientia 1987;52(Suppl.):25–61.
22. Zawia NH, Crumpton T, Brydie M, Reddy GR, RazmiafshariM. Disruption of the zinc finger domain: a common targetthat underlies many of the effects of lead. Neurotoxicology2000;21(6):1069–80.

a r c h i v e s o f o r a l b i o l o g y 5 2 ( 2 0 0 7 ) 9 3 8 – 9 4 4944
23. Engel MB. Microprobe analysis of calcifying matrices andformative cells in developing mouse molars. Histochemistry1981;72(3):443–52.
24. Takano Y, Yamamoto T, Domon T, Wakita M.Histochemical, ultrastructural, and electron microprobeanalytical studies on the localization of calcium in ratincisor ameloblasts at early stage amelogenesis. Anat Rec1990;228(2):123–31.
25. Bench G, Corzett MH, Kramer CE, Grant PG, Balhorn R. Zincis sufficiently abundant within mammalian sperm nuclei tobind stoichiometrically with protamine 2. Mol Reprod Dev2000;56(4):512–9.
26. Reith EJ. The stages of amelogenesis as observed in molarteeth of young rats. J Ultrastruct Res 1970;30(1):111–51.
27. Dillon CT, Lay PA, Kennedy BJ, Stampfl AP, Cai Z, Ilinski P,et al. Hard X-ray microprobe studies of chromium(VI)-treated V79 Chinese hamster lung cells: intracellularmapping of the biotransformation products of a chromiumcarcinogen. J Biol Inorg Chem 2002;7(6):640–5.
28. Waern JB, Harris HH, Lai B, Cai Z, Harding MM, Dillon CT.Intracellular mapping of the distribution of metals derivedfrom the antitumor metallocenes. J Biol Inorg Chem2005;10(5):443–52.
29. Harris HH, Levina A, Dillon CT, Mulyani I, Lai B, Cai Z, et al.Time-dependent uptake, distribution andbiotransformation of chromium(VI) in individual and bulkhuman lung cells: application of synchrotron radiationtechniques. J Biol Inorg Chem 2005;10(2):105–18.
30. Ueki T, Takemoto K, Fayard B, Salome M, Yamamoto A,Kihara H, et al. Scanning X-ray microscopy of living andfreeze-dried blood cells in two vanadium-rich ascidianspecies Phallusia mammillata and Ascidia sydneiensis samea.Zool Sci 2002;19(1):27–35.
31. Martin RR, Naftel SJ, Nelson AJ, Feilen AB, Narvaez A.Synchrotron X-ray fluorescence and trace metals in thecementum rings of human teeth. J Environ Monit2004;6(10):783–6.
32. Engel MB, Hilding OH. Mineralization of developing teeth.Scan Electron Microsc 1984;Pt 4:1833–45.
33. Kanno S, Anuradha CD, Hirano S. Localization of zinc afterin vitro mineralization in osteoblastic cells. Biol Trace ElemRes 2001;83(1):39–47.
34. Genge BR, Sauer GR, Wu LN, McLean FM, Wuthier RE.Correlation between loss of alkaline phosphatase activityand accumulation of calcium during matrix vesicle-mediated mineralization. J Biol Chem 1988;263(34):18513–9.
35. Reith EJ. The ultrastructure of ameloblasts during matrixformation and the maturation of enamel. J Biophys BiochemCytol 1961;9:825–39.
36. Takano Y, Ozawa H, Crenshaw MA. Ca-ATPase and ALPaseactivities at the initial calcification sites of dentin andenamel in the rat incisor. Cell Tissue Res 1986;243(1):91–9.
37. Llano E, Pendas AM, Knauper V, Sorsa T, Salo T, Salido E,et al. Identification and structural and functionalcharacterization of human enamelysin (MMP-20).Biochemistry 1997;36(49):15101–8.
38. Ono H, Wada O, Ono T. Distribution of trace metals in nucleiand nucleoli of normal and regenerating rat liver withspecial reference to the different behavior of nickel andchromium. J Toxicol Environ Health 1981;8(5/6):947–57.
39. Boyde A, Reith EJ. Scanning electron microscopy of ratmaturation ameloblasts. Cell Tissue Res 1977;178(2):221–8.
40. Hall MD, Dillon CT, Zhang M, Beale P, Cai Z, Lai B, et al. Thecellular distribution and oxidation state of platinum(II) andplatinum(IV) antitumour complexes in cancer cells. J BiolInorg Chem 2003;8(7):726–32.
41. Petersson LG, Lodding A, Koch G. Elemental microanalysisof enamel and dentin by secondary ion mass spectrometry(SIMS) deciduous and permanent teeth from high and lowfluoride area. Swed Dent J 1978;2(2):41–54.
42. Purchase NG, Fergusson JE. Lead in teeth: the influence ofthe tooth type and the sample within a tooth on lead levels.Sci Total Environ 1986;52(3):239–50.
43. Budd P, Montgomery J, Cox A, Krause P, Barreiro B, ThomasRG. The distribution of lead within ancient and modernhuman teeth: implications for long-term and historicalexposure monitoring. Sci Total Environ 1998;220(2/3):121–36.
44. Malik SR, Fremlin JH. A study of lead distribution in humanteeth, using charged particle activation analysis. Caries Res1974;8(3):283–92.
45. Arora M, Chan SW, Ryan CG, Kennedy BJ, Walker DM.Spatial distribution of lead in enamel and coronal dentine ofwistar rats. Biol Trace Elem Res 2005;105(1–3):159–70.
46. Aoba T. The effect of fluoride on apatite structure andgrowth. Crit Rev Oral Biol Med 1997;8:136–53.
47. Hammarstrom l. Different localization of tetracycline andsimultaneously injected radiocalcium in developingenamel. Calc Tiss Res 1967;1:229–42.
48. Robinson C, Briggs HD, Atkinson PJ. Histology of enamelorgan and chemical composition of adjacent enamel in ratincisors. Calcif Tiss Int 1981;33:513–20.
49. Maret W. The function of zinc metallothionein: a linkbetween cellular zinc and redox state. J Nutr 2000;130(5SSuppl.):S1455–8.
50. Ortega R. Chemical elements distribution in cells. Nucl InstMeth Phys Res B 2005;231:218–23.