the journal of biological vol. 267, no .26, of … · 2001-06-18 · assay of cu,zn-sod...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1992 by The American Society for Biochemistry and Molecular Biology, he.
Vol. 267, No .26, Issue of September 15, PP. 18505-18510.1992 Printed in U. S.A.
Site-specific and Random Fragmentation of Cu,Zn-Superoxide Dismutase by Glycation Reaction IMPLICATION OF REACTIVE OXYGEN SPECIES*
(Received for publication, January 2, 1992)
Tomomi OokawaraS, Naohisa KawamuraS, Yasuyuki Kitagawas, and Naoyuki TaniguchiSll From the $Department of Biochemistry, Osaka University Medical School, 2-2 Yamadaoka, Suita, Osaka 565, Japan and the Slnstitute for Protein Research, Osaka University, Yamadaoka 3-2, Suita, Osaka 565, Japan
Site-specific and random fragmentation of human Cu,Zn-superoxide dismutase (Cu,Zn-SOD) was ob- served following the glycation reaction (the early stage of the Maillard reaction). The fragmentation pro- ceeded in two steps, In the first step, Cu,Zn-SOD was cleaved at a peptide bond between Pros2 and HissS, as judged by amino acid analysis and sequencing of frag- ment peptides, yielding a large (16 kDa) and a small (6 kDa) fragment. In the second step, random fragmen- tation occurred. The ESR spectrum of the glycated Cu,Zn-SOD suggested that reactive oxygen species was implicated in the both steps of fragmentation.
The same fragmentations were seen upon exposure of the enzyme to an H202 bolus. Catalase completely blocked both steps of the fragmentation process, whereas EDTA blocked only the second step.
Incubation with glucose resulted in a time-dependent release of Cu2+ from the Cu,Zn-SOD molecule. The released Cu2+ then likely participated in a Fenton’s type of reaction to produce hydroxyl radical, which may cause the nonspecific fragmentation. Evidence that EDTA abolished only the second step of fragmen- tation induced by an H202 bolus supports this mecha- nism. This is the first report that a site-specific frag- mentation of a protein is caused by reactive oxygen species formed by the Maillard reaction.
Cu,Zn-SOD’ is an anti-oxidant enzyme in tissues. Our previous studies indicated that human Cu,Zn-SOD undergoes glycation (Maillard reaction) at specific lysine residues and that the enzyme is inactivated by glycation in vitro (1) as well as in vivo (2). The level of glycated Cu,Zn-SOD is increased in the erythrocytes of patients with diabetes mellitus (2, 3), as well as patients with Werner’s syndrome, an age-acceler- ated disease (4). Involvement of the glycation reaction in the pathogenesis of diabetic complications has also been suggested (5,6).
Several groups recently reported that the Schiff base and/ or Amadori products generate ROS (7-9). Brownlee’s (7)
* This work was supported by grants-in-aid for Cancer Research and Scientific Research from the Ministry of Education, Science and Culture of Japan. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
7314. 7 To whom correspondence should be addressed. Fax: 81-6-875-
The abbreviations and trivial names used are: Cu,Zn-SOD, Cu,Zn- superoxide dismutase; ROS, reactive oxygen species; DMPO, 53- dimethyl-1-pyrroline-N-oxide; SDS, sodium dodecyl sulfate; PAGE, polyacrylamide gel electrophoresis.
group described that the production of superoxide anion by the glycation of protein is implicated in atherogenesis and diabetic complications. Implication of divalent metals for the production of ROS also has been suggested (8-9). On the other hand, Wolff et al. (10) reported that the autoxidation of reducing sugars produces ROS.
It is well known that Cu,Zn-SOD is sensitive to H202 (ll), and recently Sal0 et al. (12) reported that Cu,Zn-SOD under- goes random fragmentation after exposure to H2Oz. In the present study we found that glycation of Cu,Zn-SOD initially brought about a site-specific fragmentation of the molecule, and further incubation of the enzyme with glucose brought about complete fragmentation. The role of reactive oxygen species in the fragmentation reaction is discussed.
MATERIALS AND METHODS
Recombinant human Cu,Zn-SOD was a kind gift from Ube Indus- tries, Japan. The amino acid sequence of the recombinant enzyme is the same as that of the human erythrocyte enzyme except that the N terminus Ala is not acetylated. Outdated human blood was obtained from the Japan Red Cross (Osaka, Japan). Horse erythrocytes were a gift from Dr. Masuo Nakano, Obihiro Veterinary School. Human and horse blood Cu,Zn-SODS were purified according to the method described (13). Cow milk xanthine oxidase and bovine liver catalase were purchased from Boehringer (Mannheim, Germany) and P-L Biochemicals, Inc. (Milwaukee, WI), respectively. o-Glucose, D-fruc- tose, and D-sorbitol were purchased from Nacalai tesque, Inc. (Kyoto, Japan). DMPO was purchased from Labotec (Tokyo, Japan). Poly- vinylidine difluoride (Imobillon) membrane was a product of Milli- pore Ltd. The other reagents were of the highest grade available.
Glycation of Cu,Zn-SOD in Vitro and the Effect of Reactive Oxygen Species on Fragmentation of Cu,Zn-SOD-The standard reaction mixture contained 50 mM potassium/sodium phosphate buffer, pH 7.4, 150 mM NaCI, 0.025% NaN3, 0.1 M reducing sugar, and 1-2 mg/ ml Cu,Zn-SOD (human recombinant or human or horse erythrocyte). After incubation at 37 “C for the indicated duration with gentle shaking in a water bath, the reaction was stopped by freezing the mixture. Each sample was stored at -35 “C until use.
In experiments on the effect of HZ02 on fragmentation of Cu,Zn- SOD, the H202 concentration was determined spectrophotometrically according to the extinction coefficient of 43.6 m”’. cm” at 240 nm (14). In experiments on the effect of superoxide anion on the frag- mentation, superoxide anion and HZ02 were generated by a hypoxan- thine/xanthine oxidase reaction. All solutions were sterilized by fil- tration through disposable cellulose nitrate filter DISMIC (Advantec, Tokyo, Japan).
Assay of Cu,Zn-SOD Activity-Cu,Zn-SOD activity was assayed using the xanthine-xanthine oxidase/nitro blue tetrazolium system as described previously (15).
Measurement of Ketoamine Adduct by Thiobarbituric Acid Reac- tion-Glycation of Cu,Zn-SOD was measured by the thiobarbituric acid reaction as described (16). In brief, 1 mg of glycated Cu,Zn-SOD dissolved in 500 11 of sodium phosphate buffer, pH 7.4, was hydrolyzed in 250 pl of 0.9 M oxalic acid for 4 h at 116 ”C. After cooling, 250 pl of 50% (w/v) trichloroacetic acid was added to the mixture and centrifuged and the supernatant was obtained. To the supernatant
18505

18506 Glycation and Fragmentation of Cu,Zn-Superoxide Dismutase (600 pl) 200 pl of 50 mM thiobarbituric acid was added. After incu- bation of the mixture for 60 min at 37 "C, the absorbance a t 443 nm was determined by a Hitachi U-2000 spectrophotometer.
Polyacrylamide Gel Electrophoresis in the Presence of S D S (SDS- PAGE) and Quantitation of Large and Small Fragments-SDS-PAGE was carried out as described (17) after the sample was reduced with 2.5% mercaptoethanol. Fifteen percent slab gels (90 X 55-mm) were used. The gels were stained with 0.15% Coomassie Brilliant Blue R- 250. The amount of fragmentation was quantitated by the staining intensity as judged by densitometric scanning with the flying spot scanner CS-9000 (Shimadzu, Kyoto, Japan).
Amino Acid Sequence Analysis-For the amino acid sequence analysis, recombinant Cu,Zn-SOD which had been incubated with 0.1 M glucose for 7 days, 5 mM HzOz for 30 min, or 1.25 mM hypoxanthine, 0.5 unit/ml xanthine oxidase for 4 h was subjected to SDS-PAGE. The proteins were then transferred onto polyvinylidine difluoride membrane using a Sartoblot semi-dry electroblotting sys- tem (Sartorius, Goettingen, Germany) a t 4 mA/cm2 for 15 min in a Tris/norleucine discontinuous buffer system according to the manu- facturer's instruction. The blotted spots were cut into slices and subjected to a gas-phase sequenator. Amino acid sequencing was carried out with an Applied Biosystems 477A Protein Sequencer equipped with an on-line analyzer for phenylthiohydantoin-deriva- tized amino acids.
Amino Acid Analysis of a Small Fragment-A 5-kDa fragment was obtained by the electroelution from SDS-PAGE slab gels (18). One- dimensional SDS-PAGE was carried out with 15% slab gels as de- scribed above and stained with Coomassie Brilliant Blue G-250. A band corresponding to 5 kDa was cut out from the gel and sliced into small pieces. The sliced gels were boiled in the same volume of 30 mM Tris-HCI, pH 6.5, containing 0.5% SDS for 5 min and then placed in a hand-made electroeluter as described (18). The fragments were eluted into a dialysis bag (Spectra/Por 3 membrane), concentrated, and then separated on a reversed phase high performance liquid chromatography column (Capcell Pak C8, Shiseido, Japan) using the trifluoroacetic acid/acetonitrile buffer system. The purified small fragment was hydrolyzed in 6 N HCI a t 100 "C for 24 h under in vacuo. Amino acid analysis was carried out with a Hitachi L-8500 automatic amino acid analyzer.
ESR Spectra Measurements-ESR spectra were measured at room temperature using an E-12 EPR spectrometer system (Varian). The magnetic fields were calculated from the splitting of MnO. After a 3- day incubation of native or heat denatured Cu,Zn-SOD in the pres- ence or absence of 0.1 M glucose a t 37 "C, DMPO was added to the incubation mixture to a final concentration of 0.1 M as the hydroxyl radical trapping reagent and ESR signals were measured. Catalase (0.2 mg/ml) or 5 mM EDTA was added to the reaction mixture. Heat denatured Cu,Zn-SOD was obtained by boiling a t 100 "C for 60 min and released copper ion was separated on a PD-10 column which had been equilibrated with 50 mM sodium phosphate buffer, pH 7.5. In order to eliminate the autoxidation of glucose as a source of ROS, after incubation of the reaction mixture glucose was removed by a gel filtration column and the glycated Cu,Zn-SOD was further incubated at 37 "C for 12 h and then DMPO was added to measure the ESR signals.
Determination of Free Copper Ion Concentration-The concentra- tion of copper ion released from glycated Cu,Zn-SOD was determined using bathocupuroine disulfonic acid by the method described previ- ously (14). CuSOl solution for atomic absorption spectrometry (Na- calai tesque, lnc., Kyoto, Japan) was used as the standard.
RESULTS
Cleavage of Cu,Zn-SOD by Glycatwn Reaction-Incubation of human Cu,Zn-SOD with 0.1 M glucose for 0-14 days at 37 "C resulted in a time-dependent decrease in the amount of intact enzyme. SDS-PAGE showed a gradual decrease in the intensity of the original band with an approximate molecular mass of 20 kDa and the simultaneous appearance of a large molecular weight fragment with a distinct 15-kDa band and a small molecular mass fragment of about 5 kDa (Fig. LA). The kinetics of the decrease of intact Cu,Zn-SOD and ap- pearance of these fragments were assessed by densitometric scanning of staining intensity on SDS-PAGE (Fig. 1B). The figure represents percent contents and is based on a measure of weight amount. Therefore, assuming that staining intensity
A
1- Original Cu,Zn-SOD Large fragment - Small fragment
0 8 1 3 5 7 1014 hrs -days-
Incubation time
B
" 0 1 3 5 7 1 0 1 4 Incubation time (days)
FIG. 1. Fragmentation of Cu,Zn-SOD after incubation with glucose. After incubation with 100 mM glucose as described under "Materials and Methods," an aliquot of the incubation mixture con- taining 30 pg of Cu,Zn-SOD was directly loaded onto an SDS- polyacrylamide gel electrophoresis (A ) . Relative staining intensity of SDS-PAGE gels was on the basis of weight amount. ( B ) . W, a small fragment; 0, a large fragment; 0, original Cu,Zn-SOD.
L
FIG. 2. Effect of glucose concentration on the fragmenta- tion reaction of Cu,Zn-SOD was incubated with glucose for l, 3, and 7 days. The incubation mixtures were then analyzed as described in Fig. 1. Each lane contained 15 pg of Cu,Zn-SOD.
is equal per peptide mass, at equimolar amounts the staining of a large fragment would be more intense. The site-specific fragmentation increased promptly until 3 days and then in- creased gradually until 7 days. After 7 days, the fragments gradually decreased. These results indicate that the site- specific fragmentation occurred at first, and after 7 days, random fragmentation occurred predominantly. After 7 and 14 days incubation, approximately 80 and 50%, respectively, of the original Cu,Zn-SOD remained.
The effect of glucose concentration on the fragmentation of Cu,Zn-SOD was studied. At 0.1 mM glucose, essentially no fragmentation was observed (data not shown). At 10 mM glucose, however, fragmentation could be detected after in- cubation for 3 days, as shown in Fig. 2. At 500 mM glucose, pronounced fragmentation was observed after incubation for 1 day. The specific fragmentation reached a maximum at 3 days, and after 7 days the discrete fragments had been largely replaced by nonspecific degradation products. This suggests that the fragmentation proceeded in the two steps, with cleavage at a specific site as the initial step and random fragmentation as the second step.
Fructose has much more reducing capacity as compared to glucose in the Maillard reaction (19). The effect of fructose on the fragmentation reaction was studied. Incubation with 100 mM fructose for 3-7 days resulted in rapid and marked fragmentation of the Cu,Zn-SOD, whereas incubation with a nonreducing sugar such as sorbitol had no effect (Fig. 3). This
Glycation and Fragmentation of Cu,Zn-Superoxide Dismutase 18507
eb'J L"a";l ;: ;:; glucose fructose xxbitd Imimlim~imc
c 7d-
- la,- buyun, 14.4k
(Mr)
FIG. 3. Effects of various sugars on the fragmentation of Cu,Zn-SOD. The incubation mixture contained 100 mM sugar. Lanes a, 6, and c indicate the incubation times of 1, 3, and 7 days, respec- tively. The reaction mixture was directly loaded onto an SDS-poly- acrylamide gel electrophoresis. Each lane contained 15 pg of Cu,Zn- SOD.
L-I f
0 1 2 3 4 5 6 7 Incubation time (days)
FIG. 4. Inactivation of Cu,Zn-SOD during incubation with 0.1 M glucose for 1-7 days. After incubation for the appropriate time, an aliquot was taken and the Cu,Zn-SOD activity in the pres- ence (0) or absence (0) of EDTA and thiobarbituric acid reaction (W) was determined as described under "Materials and Methods."
suggests that the fragmentation is related to interaction of the Cu,Zn-SOD with reducing sugar and that the Maillard reaction (glycation reaction) may play a role in the fragmen- tation. Under anerobic conditions, the fragmentation was not observed (data not shown), indicating that the dissolved oxy- gen molecules are requisite for the fragmentation reaction.
The same fragmentation pattern was obtained using puri- fied Cu,Zn-SODS from human and horse erythrocytes (data not shown).
Loss of Cu,Zn-SOD Actiuity after Glycation-During incu- bation of Cu,Zn-SOD with glucose, the enzyme activity grad- ually decreased and simultaneously glycated products in- creased as reported previously (2). As reported earlier by other groups (8, 9), the ROS production by glycated proteins re- quires divalent metals. In the presence of EDTA, fragmenta- tion of Cu,Zn-SOD by glycation did not occur (data not shown). Therefore, the loss of activity in the presence of EDTA is considered to be due to the glycation of lysine residues (see open circle of Fig. 4). The reactivity of the thiobarbituric acid reaction which estimates the formation of ketoamine adducts in the glycation reaction was also parallel with the loss of enzyme activity. In the absence of EDTA (see closed circle of Fig. 4) the loss of activity was found to be marked, particularly after 2-7 days of incubation. These data will be well explained by the increased fragmentation of Cu,Zn-SOD as shown in Fig. 1B.
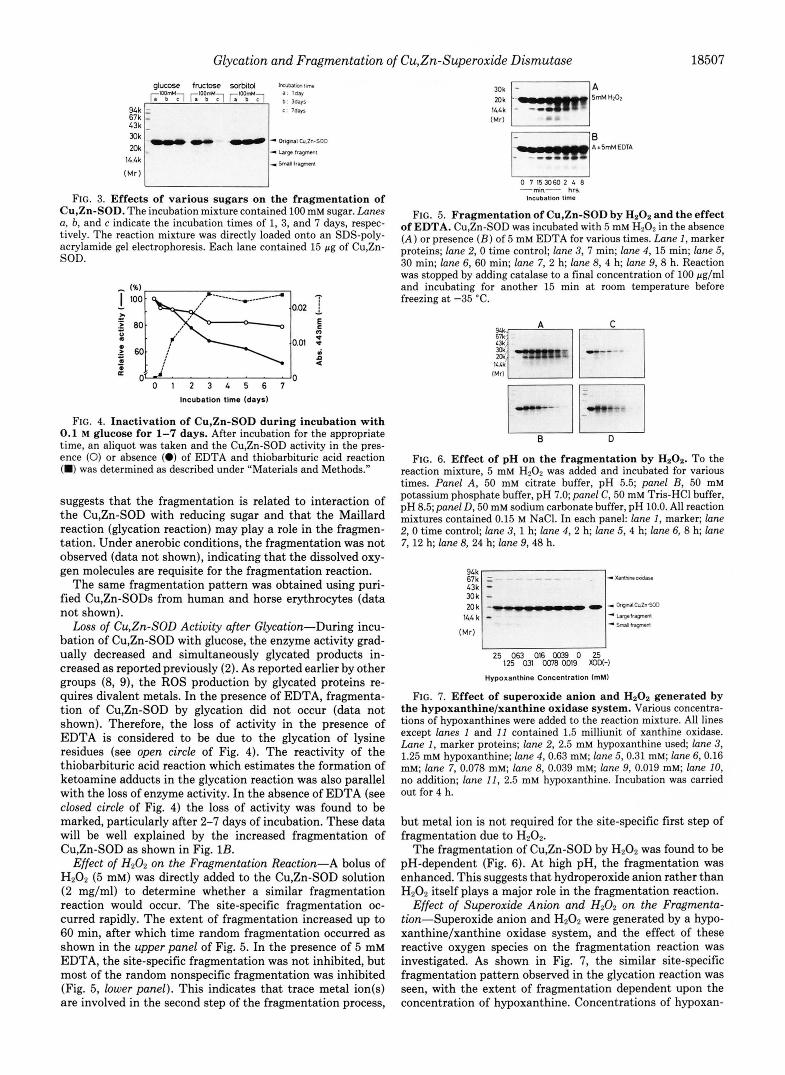
Effect of H202 on the Fragmentation Reaction-A bolus of H202 (5 mM) was directly added to the Cu,Zn-SOD solution (2 mg/ml) to determine whether a similar fragmentation reaction would occur. The site-specific fragmentation oc- curred rapidly. The extent of fragmentation increased up to 60 min, after which time random fragmentation occurred as shown in the upper panel of Fig. 5. In the presence of 5 mM EDTA, the site-specific fragmentation was not inhibited, but most of the random nonspecific fragmentation was inhibited (Fig. 5, lower panel). This indicates that trace metal ion(s) are involved in the second step of the fragmentation process,
Bnk - I A
E l B
" Ar5-EDTA
0 7 153060 2 4 8 -min- hrs
Incubation time
FIG. 5. Fragmentation of Cu,Zn-SOD by HzOz and the effect of EDTA. Cu,Zn-SOD was incubated with 5 mM H202 in the absence ( A ) or presence ( B ) of 5 mM EDTA for various times. Lune 1, marker proteins; lane 2, 0 time control; lane 3, 7 min; lane 4, 15 min; lane 5, 30 min; lane 6, 60 min; lane 7, 2 h; lane 8, 4 h; lane 9, 8 h. Reaction was stopped by adding catalase to a final concentration of 100 pg/ml and incubating for another 15 min a t room temperature before freezing a t -35 'C.
g - q F 1 F I I I - 1 IMrl
I l l k
0 D
FIG. 6. Effect of pH on the fragmentation by H202. To the reaction mixture, 5 mM H202 was added and incubated for various times. Panel A , 50 mM citrate buffer, pH 5.5; panel B, 50 mM potassium phosphate buffer, pH 7.0; panel C, 50 mM Tris-HCI buffer, pH 8.5;panel D, 50 mM sodium carbonate buffer, pH 10.0. All reaction mixtures contained 0.15 M NaCI. In each panel: lane 1, marker; lane 2,O time control; lane 3 , l h; lane 4 , 2 h; lane 5 , 4 h; lane 6, 8 h; lane 7, 12 h; lane 8, 24 h; lane 9,48 h.
Hypoxanthine Concentration (rnM)
FIG. 7 . Effect of superoxide anion and Hz02 generated by the hypoxanthine/xanthine oxidase system. Various concentra- tions of hypoxanthines were added to the reaction mixture. All lines except lanes 1 and 11 contained 1.5 milliunit of xanthine oxidase. Lane 1, marker proteins; lane 2, 2.5 mM hypoxanthine used; lane 3, 1.25 mM hypoxanthine; lane 4,0.63 mM; lane 5,0.31 mM; lane 6, 0.16 mM; lane 7, 0.078 mM; lane 8, 0.039 mM; lane 9, 0.019 mM; lane 10, no addition; lane 11, 2.5 mM hypoxanthine. Incubation was carried out for 4 h.
but metal ion is not required for the site-specific first step of fragmentation due to H202.
The fragmentation of Cu,Zn-SOD by HZ02 was found to be pH-dependent (Fig. 6). At high pH, the fragmentation was enhanced. This suggests that hydroperoxide anion rather than H202 itself plays a major role in the fragmentation reaction.
Effect of Superoxide Anion and H202 on the Fragmenta- tion-Superoxide anion and H202 were generated by a hypo- xanthinelxanthine oxidase system, and the effect of these reactive oxygen species on the fragmentation reaction was investigated. As shown in Fig. 7, the similar site-specific fragmentation pattern observed in the glycation reaction was seen, with the extent of fragmentation dependent upon the concentration of hypoxanthine. Concentrations of hypoxan-

18508 Glycation and Fragmentation of Cu,Zn-Superoxide Dismutase
thine above 1 pM resulted in fragmentation, and the cleavage was inhibited by the addition of catalase (data not shown). These results indicate that superoxide anion or HzOz or both are involved in the fragmentation process.
Identification of the Site of Cu,Zn-SOD Cleavage by Glyca- tion-In order to identify the site of the initial cleavage, the glycated human recombinant Cu,Zn-SOD was subjected to amino acid sequence analysis. As shown in Table I, the intact Cu,Zn-SOD and the 5-kDa fragment were found to have the same amino acid sequence from Ala at the N terminus. The 15-kDa band was found to contain residues from His63. Frag- mentations by a bolus HzOz and hypoxanthine/xanthine oxi- dase were also examined. The same cleavage site was found on amino acid sequence analysis when Cu,Zn-SOD was ex- posed to a bolus H20z. When exposed to hypoxanthine/ xanthine oxidase, His71, which is coordinated with Cu metal was identified as one of the fragment peptides (data not shown) but multiple cleavage sites may also exist.
Amino acid analysis of the 5-kDa fragment indicated that the fragment contained 3 prolines as shown in Table 11. The value was consistent with the reported amino acid residues (20), suggesting that the Pro6' was found to be intact. This suggests that the cleavage site was a peptide bond between Pro6' and His63 of the human Cu,Zn-SOD. His63 is one of 4 histidines coordinated with the Cu ion of Cu,Zn-SOD, and Hisfi3 is also coordinated with the Zn ion (21). Consequently,
TABLE I Amino acid sequence data for the 5- and 15-kDa fragments and
native Cu,Zn-SOD X , not identified.
Corresponding
region Fragment Sequence amino acid
5-kDa protein A-T-K-A-V-X-V Amino acids
15-dKa protein 1-7
Produced by H-F-N-P-L-X-R-K-H-G Amino acids
Produced by H-F-N-P-L-X-X-K Amino acids 5 mM H202 63-70
Native Cu, Zn- A-T-K-A-V-C-V-L-K-G Amino acids SOD 1-9
H-F-N-P-L-S-R-K-H-G Amino acids
0.1 M glucose 63-72
63-72
TABLE I1 Amino acid composition of a 5-kDa fragment
All integral numbers in parentheses are theoretical values deduced from the known complete amino acid sequence of human Cu,Zn-SOD (20). The amino acid compositions are based on isoleucine (see asterisk). Tryptophan was not determined. The values indicate the mean of the duplicate experiments.
Amino acid Composition
Asx 5.4 (5) Thr 3.4 (4) Ser 3.5 (3) Glx 6.5 (6) Pro 3.1 (3) Glu 11.7 (11) Ala 4.1 (4)
Val 5.9 (6) Ile* 3.0 (3) Leu 3.8 (3)
Phe 3.1 (3)
His 1.9 (3)
CYS 0.8 (2)
TY r 0.4 (0 )
LY s 4.6 (5)
Arg 0.3 (0)
cleavage at this position results in structural and functional damage to the enzyme.
Most mammalian Cu,Zn-SODS have the P r ~ " - H i s ~ ~ se- quence, while the enzyme from horse erythrocytes has Ala6'- His63 (22). In order to determine whether Pro6' is requisite for the cleavage site, horse Cu,Zn-SOD was incubated with glucose and SDS-PAGE was carried out. The same fragmen- tation pattern was seen, indicating that the Pro residue is not essential for this fragmentation. Addition of HzOz also re- sulted in fragmentation of the horse Cu,Zn-SOD.
Copper Ion Release from CyZn-SOD during Glycatwn Re- action-Hydroxyl radical can be formed from HzOz through a Fenton's-type reaction in the presence of metals. As described above, metal ion(s) are implicated in the second step, random fragmentation of Cu,Zn-SOD. As shown in Fig. 8, glycation lead to release of Cu ions from the Cu,Zn-SOD molecule. The released Cu ions could then enhance the Fenton's-type reac- tion to produce hydroxyl radicals from the HzOZ generated by the glycation reaction and play a major role in the random fragmentation.
Electron Spin Resonance Spectra of the DMPO-OH Adduct by Glycation and Fragmentation of Reaction Mixture-In order to identify radical species, ESR spectra were measured after incubation of Cu,Zn-SOD with 0.1 M glucose at 37 "c for 3 days using DMPO as a trapping agent. In the presence of DMPO, glycation of Cu,Zn-SOD generated 'OH which was detected by ESR as a DMPO-OH adduct. The adduct spec- trum consisted of a quartet (1:2:2:1) with hyperfine splitting constants of aN=aH= 15.0 G (Fig. 9). No signals were observed after addition of EDTA, which suggests involvement of metal ions in this reaction as suggested by several groups (8, 9). Addition of catalase also prevented the appearance of the DMPO-OH signals. This suggests that superoxide anion is
-* """" ~ I 0 1 2 3 4 5 6 7
Incubation time (Days)
FIG. 8. Copper ion release from the Cu,Zn-SOD after incu- bation with glucose. Cu,Zn-SOD (10 pg) was incubated with or without 0.1 M glucose at 37 "C under standard reaction conditions for various time intervals. An aliquot was taken, and free copper ion was determined according to the method described (12).
A: &+LW-.-',~++&~*~.-J"- (Control) 0.1M Glucose
0 ii +\ V I .I %JJ@ P*I',* 0.1M Glucose ' b,
4- Img/ml Cu,Zn-SOD
I : ! c: + , - + % ~ ~ & d ~ ~ ~ - y B with EDTA
D: , , d y ' &+...- B wlth Catalase
E: + " + + w b , ~ v + " - (Control)
-
lms/ml Cu,Zn-SOD
IOG
FIG. 9. Electron spin resonance spectrum of DMPO-OH ad- ducts formed after incubation of Cu,Zn-SOD with glucose at 37 OC for 3 days. A , 0.1 M glucose; B, Cu,Zn-SOD with 0.1 M glucose; C, Cu,Zn-SOD with 0.1 M glucose and 5 mM EDTA; D, Cu,Zn-SOD with 0.1 M glucose and 0.1 mg/ml catalase; E , Cu,Zn- SOD (1 mg/ml) alone. Spectra were recorded with a modulation amplitude of 5.0 G.

Glycation and Fragmentation of Cu, Zn-Superoxide Dismutase 18509
generated at first from the glycation reaction and then Hz02 is formed by a dismutation reaction of Cu,Zn-SOD. In order to inhibit the dismutation reaction by Cu,Zn-SOD in the reaction mixture, Cu,Zn-SOD was heat-denatured and then incubated with 0.1 M glucose, and then ESR spectra were measured. As shown in Fig. 1OA, the same DMPO-OH adduct was also observed, This adduct was eliminated by inclusion of EDTA (Fig. 1OB) but not by catalase (Fig. 1OC). These data again suggest that the dismutation reaction is highly involved in the fragmentation reaction, and possibly super- oxide anion as a substrate for Cu,Zn-SOD is supplied from the glycation reaction in the presence of a trace level of metal even though no direct evidence for the presence of DMPO- OOH adduct was observed.
In order to eliminate autoxidation of glucose as a source of ROS, glycated Cu,Zn-SOD was separated from free glucose and ESR spectra was examined. Glycated Cu,Zn-SOD free from glucose also gave a DMPO-OH adduct (data not shown).
DISCUSSION
Our previous studies indicated that human Cu,Zn-SOD undergoes glycation reaction at specific lysine residues such as Lys"* and Lys"* (1) which comprise a positively charged channel track. We have proposed that the glycation of the lysine residues resulted in the relatively negative charge, which may interfere with the electrostatic guidance of the substrate superoxide anion to the active site.
In the present study, however, we found that Cu,Zn-SOD undergoes a site-specific cleavage between P r ~ ~ ' - H i s ~ ~ follow- ing the glycation reaction. Moreover, the lengthly incubation of the enzyme with glucose resulted in random fragmentation as judged by SDS-PAGE. In order to investigate the mecha- nism for the fragmentation reaction, we calculated the com- puter image of human Cu,Zn-SOD using the known tertiary structures of the spinach and bovine enzymes, which are very similar to the human enzyme in terms of tertiary structure (23). Data for the human enzyme is not available at present. According to the computer image, Pro6' is located on the surface of the molecule. Calculation of the extent of exposure suggested that Pro6' protruded from the surface.
The glycation reaction of Cu,Zn-SOD appears to be rather specific, the primary sites of glycation are L Y S ' ~ ~ and L Y S ' ~ ~ as reported previously by our group (1). Watkins et al. (24) and Acharya et al. (25) reported that the interatomic distance between His and Lys residues may play an important role in the glycation reaction because the Amadori formation in the Maillard reaction is an acid-base catalysis. The interatomic distances between Lys''' and His'" and between and His43 are small, 9.3 and 10.9 A, respectively. The principal glycated sites are not close to the cleavage site a t Pr~"-His~~, which suggests that the glycation reaction is not directly involved in the fragmentation. The generation of superoxide
B: A With EDTA
c A wth catalase - 1OG
FIG. 10. Electron spin resonance spectrum of DMPO-OH adducts formed after incubation of heat-denatured Cu,Zn- SOD with glucose. Experimental conditions were the same as described in the legend to Fig. 9. Cu,Zn-SOD was completely inacti- vated by heating at 100 "C for 1 h. Copper released from the molecule during the heat-inactivation process was separated from Cu,Zn-SOD on a column of gel filtration.
anion in the glycation reaction and the subsequent generation of H20z or hydroxyl radical, however, may play an important role in the site-specific fragmentation reaction.
In diabetic patients, the glycated form of Cu,Zn-SOD is increased approximately %fold (2,3). Therefore, the fragmen- tation of Cu,Zn-SOD likely also occurs in vivo. Sal0 et al. (12) reported that Cu,Zn-SOD undergoes fragmentation by oxi- dation reaction in red cells and the damaged enzyme is sus- ceptible to proteolytic attack by Macrooxyprotease. Presum- ably Cu,Zn-SOD molecules damaged by glycation in vivo rapidly undergo proteolytic degradation, and hence it might be difficult to confirm the occurrence of fragmentation in vivo. This remains for further study.
In the present study we found the cleavage site to be located at Pro6'-Hisa3. The mechanism for fragmentation of Cu,Zn- SOD remains unknown. Reactive oxygen species such as superoxide anion, H202, and hydroxyl radical are likely in- volved in both site-specific and random fragmentation. It has been reported that the superoxide anion is generated in the presence of trace metals from glycated products (&9), and the dismutation of superoxide anion results in further accu- mulation of H202. One possible mechanism is that the en- zyme-bound Cu2+ is reduced by H202 or superoxide anion to Cu+, followed by a Fenton's-type reaction of the Cu' with additional H202. This would generate Cu2+-OH. or its ionized equivalent, Cu2+-O', which could then attack the adjacent Pro-His residues as proposed by Hodgson and Fridovich (11). As described above, we found that horse Cu,Zn-SOD, which has Ala instead of Pro at the P r ~ ~ ' - H i s ~ ~ site of the human enzyme, underwent a similar fragmentation reaction, indicat- ing that Pro6' was not requisite for this fragmentation.
Stadtman's group extensively studied the effects of active oxygens on the modification of amino acids (26). They re- ported that one of the amino acid residues sensitive to oxi- dative damage is proline, which is converted to 5-hydroxypro- line or glutamyl semialdehyde and then converted to glutamic acid via pyroglutamic acid (27). In the present study we could not identify such a conversion because a 5-kDa fragment had 3 intact proline residues as described above. Instead cleavage of the peptide bond between Pro6' and H i P seemed to occur. On the other hand, we cannot deny the possibility that mod- ification of proline occurs, but that it is reversed under con- ditions of acid hydrolysis prior to amino acid analysis.
The second step of fragmentation may involve hydroxyl radicals generated from H202 through a Fenton's-type reac- tion with Cu released from the damaged enzyme molecules (see Fig. 8). Recently Yim et al. (28) reported that Cu,Zn- SOD catalyzes the production of hydroxyl radical from HzOz. The random fragmentation may, therefore, be augmented by the generation of free hydroxyl radicals from HzOz due to the residual intact Cu,Zn-SOD after the glycation reaction.
From these results, we suggest that the glycation reaction first produces superoxide anion. The superoxide anion is then converted to HzOz and hydroxyl radical by a combination of the Fenton's-type reaction, the reverse reaction of superoxide dismutase (ll), and the catalytic reaction of Cu,Zn-SOD (28). The fragmentation of Cu,Zn-SOD under hyperglycemic con- ditions in vivo is now under study in our laboratory.
Acknowledgments-We thank Satomi Kometani and Dr. Jun-ichi Yamauchi for performing amino acid sequence analysis.
REFERENCES 1. Arai, K Maguchi, S. Fujii S., Ishibashi, H., Oikawa, K., and Taniguchi,
2. Arai, K., Iizuka, S., Tada, Y. Oikawa, K., andTaniguchi, N. (1987) Biochirn.
3. Kawamura, N., Ookawara, T., Suzuki, K., Konishi, K., Mino, M., and
N. (1&37) J. Bbl. dhern. 262,16969-16972
Biaphys. Acta 924, 292-i96
Taniguchi, N. (1992) J. Clin. Endocrinol. Metab. 74, 1352-1354

18510 Glycation and Fragmentation of CyZn-Superoxide Dismutase 4. Taniguchi, N., Kinoshita, N., Arai, K., Iizuka, S., Usui, M., and Naito, T. 15. Beauchamp, C., and Fridovich, I. (1971) Anal. Biochem. 44,276-287
(1989) in The Maillurd Reaction in Aging, Dia6etes and Nutrition 16. Ney, K. A., Colley, K. J., and Pizzo S. B. (1981) Anal. Biochem. 118 , 294- (Baynes, J. W., and Monnier, V. M., eds) pp. 277-290, Alan R. Liss, Inc., 300
5. Kinoshita, N., Ookawara, T., Suzuki, K., Uchiyama, I., Kuwayama, Y., 18. LeGendre, N., and Mastudaira, P: T. (1989) in A Practical Guide to Protein New York 17. Laemmli, U. K. (1970) Nature 27,680-685
Manabe. R.. and Tanirmchi. N. (1990) in The Maillard Reaction in Food and Peptide Purificatzon for Mwrosequenclng (Mataudaira, P. T., ed) pp.
6.
7.
8. 9.
10. 11. 12.
13. 14.
Monnier, V. M. (1989) in The Maillard Reaction in Aging, Dio6etes and Verlag, Switzerland
Nutrition (Baynes, J. W., and Monnier, V. M., eds) pp. 1-22, Alan Liss, Inc., New York
Mullarkey, C. J., Edelstein, D., and Brownlee, M. (1990) Biochem. Biophys. Res. Commun. 173,932-939
Sakurai, T., and Tsuchiya, S. (1988) FEBS Lett. 236,406-410 Kawakishi, S., Tsunehiro, J., and Uchida, K. (1991) Carbohydr. Res. 2 1 1 ,
Wolff, S. P., and Dean, R. T. (1987) Biochem. J. 246,243-250 Hodgson, E. K., and Fridovich, I. (1975) Biochemistry 14,5294-5299 Salo, D. C., Pacifici, R. E., Lin, S. W., Giulivi, C., and Davies, K. J. A.
McCord, J. M., and Fridovich, I. (1969) J. Biol. Chem. 244,6049-6955 Jewett, S. L., Cushing, S., Gillespie, F., Smith, D., and Sparks, S. (1989)
167-171
(1990) J. Biol. Chem. 2 6 5 , 11919-11927
Eur. J . Biochem. 180,569-575
20.
21.
22. 23.
24.
25.
26.
28. 27.
Jabusch, J. R., Farb, D. L., Kershensteiner, D. A., and Deutsch, H. F.,
Tainer, J. A,, Getzoff, E. D., Richardson, J. S., and Richardson, D. C.
Lerch, K., and Anmer, D. (1981) J. Biol. Chem. 2 5 6 , 11545-11551 Kitagawa, Y., Tanaka, N., Hata. Y., Kusunoki, M., Lee, G-P., Katsube, Y.,
Watkins, N. G., Thorpe, S. R., and Baynes, J. W. (1985) J. Biol. Chem.
(1980) Biochemistry 19.2310-2316
(1983) Nature 306,284-286
Asada, K., and Aibara, S. (1991) J. Bwchem. 109,477-485
Acharya, A. S., Syssman, L. G., and Manning, J. M. (1983) J. Biol. Chem.
Amici, A., Levine, R. K. M., Tsai, L., and Stadtman, E. R. (1989) J . Biol.
Stadtman, E. R., and Oliver, C. N. (1991) J. Biol. Chem. 266,2005-2008 Yim,,M. B., Chock, P. B., and Stadtman, E. R. (1990) Proc. Natl. Acad.
2 6 0 , 10629-10636
258,2296-2302
Chem. 264,3341-3346
Scr. U. S. A. 87,5006-5010