the journal of vol. 268, 31, of 5, pp. 23390-23398. 1993 0 ... · nucleotide sequence and predicted...
TRANSCRIPT

THE JOURNAL OF BIOLCGICAL CHEMSTRY 0 1993 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 268, No. 31, Issue of November 5, pp. 23390-23398. 1993 Printed in U.S.A.
Brn-5 Is a Divergent POU Domain Factor Highly Expressed in Layer IV of the Neocortex*
(Received for publication, May 28, 1993)
Bo@ Andersen8, Marcus D. Schonemann, Richard V. Pearse II, Keith Jenne, Jeff Sugarman, and Michael G. Rosenfeldsn From the Eukaryotic Regulatory Biology Program and the $Howard Hughes Medical Institute, School of Medicine, University of California at Sun Diego, La Jolla, California 92093-0648
We have identified rat cDNA clones that encode a POU domain protein, referred to as brain-5 (Brn-5). During embryogenesis in the rat, Brn-5 is widely ex- pressed with highest levels in the developing brain and spinal cord from embryonic day 12.5. In the adult, Bm-5 mRNA is most abundant in the brain, where it is diffusely expressed with the exception of an enrich- ment in layer IV of the neocortex. However, Brn-6 is also found in multiple adult tissues outside the central nervous system, including kidney, lung, heart, adrenal, skin, testis, and anterior pituitary. This expression pat- tern contrasts with that of most other POU domain genes that are expressed predominantly in the develop- ing nervous system and are progressively restricted to discrete regions of the brain. The predicted amino acid sequence of Brn-5 is considerably divergent from previ- ously described POU domains and thus defines a new POU domain subclass (class VI). Consistent with its di- vergent sequence, the DNA-binding characteristics of Brn-6 overlap with, but are clearly distinct from, that of Oct-2. Although Brn-5 can bind to non-octamer sites, a random site selection indicates that its preferred binding site contains a variant octamer core motif. Fi- nally, we show that the amino terminus of Brn-5 con- tains a transactivation domain.
Mammalian development is regulated by the temporal- and spatial-specific activity of factors that stimulate or suppress the transcription of developmentally important genes. Among the best defined developmental regulators are the homeotic genes which encode transcription factors characterized by a 60-amino acid long structural motif referred to as the homeobox (Scott and Carroll, 1987; Levine and Hoey, 1988). This domain resembles the helix-turn-helix motif of bacterial DNA-binding proteins and is responsible for the DNA-binding activity of these regulators (Gehring, 1987). The importance of homeodo- main-containing proteins in development was initially revealed by homeotic mutations in Drosophila (McGinnis et al., 1984; Scott and Weiner, 1984). Subsequently, homologous genes with
* This work was supported by National Institutes of Health Grant MH 47137 (to M. G. R.). Aportion of this work was presented as a poster and published as an abstract in the Meeting on Mouse Molecular Ge- netics, Cold Spring Harbor Laboratory, Cold Spring Harbor, N Y , Au- gust, 1992. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
to the GenBankTMIEMBL Data Bank with accession numberk) L23204. The nucleotide sequenceh) reported in this paper has been submitted
$: Bugher Fellow of the American Heart Association. 1 Investigator with the Howard Hughes Medical Institute. To whom
correspondence and reprint requests should be addressed. Tel.: 619-534- 5858; Fax: 619-534-8180.
important developmental functions were described in several other species, including mammals (for review see Holland and Hogan (1988) and Kessel and Gruss (1990)).
A specific subclass of homeodomain proteins was identified by the cloning of factors that activate specific transcription units; the octamer-binding proteins Oct-1 (Sturm et al., 1988) and Oct-2 (Clerc et al . , 1988; KO et al . , 1988; Muller et al . , 1988; Scheidereit et al., 1988) and the pituitary transcription factor Pit-1 (Bodner et al., 1988; Ingraham et al., 1988). Com- parison of these factors with the nematode developmental regulatory gene unc-86 (Finney et al., 1988) revealed a con- served 69- to 72-amino acid long domain on the NH2-terminal side of their homeodomain. This novel domain, referred to as the POU-specific domain, is required in conjunction with the POU homeodomain for high affinity interactions with most DNA-binding sites. The POU-specific domain is joined to the POU homeodomain by a 14- to %-amino acid variable linker sequence, and together they are referred to as the POU do- main (Herr et al., 1988). The four initially described members of the POU domain family are thought to exert critical roles in gene regulation and metazoan development (Fletcher et al . , 1987; Bodner et al., 1988; Ingraham et al., 1988; Muller et al., 1988; Scheidereit et al., 1988; Tanaka et al., 1988; Finney and Ruvkun, 1990; Li et al., 1990; Corcoran et al., 1993). In addi- tion, Oct-1 and several other POU domain factors are capable of regulating viral DNA replication in vitro (Verrijzer et al., 1990a, 1992a), and Pit-1 appears to regulate pituitary cell proliferation (Li et al . , 1990).
Since the molecular cloning of the original POU domain genes, several additional members of this family have been isolated (for review see Rosenfeld, 1991; Ruvkun and Finney, 1991; Scholer, 1991; Wegner et al., 1993). These additional POU domain proteins have permitted a classification of existing POU genes into five subclasses based on similarities in the sequence of the POU domain, including the linker region, and weaker homologies of the NH2-terminal domains (He et al., 1989; Hara et al . , 1992). Although no unifying rule for expres- sion pattern has emerged, examination of the developmental expression of POU domain genes in the mammalian nervous system has revealed that all known family members, except Oct-314 and S k n - l a l i are expressed during development ofthe nervous system (He et al., 1989; for review see Rosenfeld (1991) and Treacy and Rosenfeld (1992)). Thus, transcripts of the Oct-1, Oct-2, Pi t -I , Brn-1, Bm-2, Brn-3.0, Brn-4, and l3t -1 genes are detected in the developing neural tube of the rat at embryonic days 9 to 12. During early stages, expression is diffise, and, in contrast to most classic homeodomain genes, the region of expression includes the developing midbrain and forebrain. During later embryological stages, and in the adult, expression of most of these genes becomes completely restricted to subregions of the brain (He et al., 1989,1991; Hatzopoulos et al., 1990; Monuki et al . , 1990; Stoykova et al . , 1992; Le Moine
23390

Brn-5 Is Enriched in Neocortex 23391
S.... CK .Iw\ OCA AGG CTC TK CK CAC CCA CAG A T AGT Gcc GcG TCC CIT GGG GGA
CAG A 0 3 CAA A K CTC GGC 'KC CK ACT ACA CCT CCA (3" A l T ACC AAC ACC A l T CCC
AGC ATG CCC GGG ATC AGC AGT CCG ATC CTC ACC AAT GCT CAG CGA CAG G l T A l T GGA met pro gly ile ser ser pm ile leu thr asn ala gln gly gln Val ile gly 18
GCA CTT CCA Tuj GTA GTG AAC TCA GCC AGT GTG GCC da ku pm np val Val am m da ST Val rph
CTG CAG GTC CAA GCT GTG ACT CCC CAG CTC 'ITG ITG leu gln wl gln da v d thr pm gln ku leu leu
GCA ACA CTA GCC AGC AGC CCC CTG CCT CAA CCT GTG oh fhr leu ab ST m pm leu pm gln pm Val
CCG GAG TCC CCT GCA AAG AGT GAG GTG CAA CCC ATC pm glu ser pm ab l y s ser glu Val gln pm ile
CCA CCT GCT GTA ATC CTC ACC AGC CCA GCT CCA GCA pm pm aka VRI ile leu thr m pm oh pro ab
CCC A X CCA ATC ACC TGC TCA GAG ACA CCT ACA GTC pm ile pm ile thr cys m glu thr pm thr Val
ACA CCA GCA CCA GCA CAG AGC thr pro do pm da gln ser 37
AAT GCC CAG GGC CAG GKi ATC asn ala gln gly gln Val ile 56
GCT GTC CGG AAG CCA AGC ACA ab val arg lys pm ser fhr 75
CAG CCG ACA CAA GCC GTG-CCC gln pm fhr gln oh Val pm 94
CTC AAG CCA TCA GCC TCA GCT leu lys pm m do ser ab I 1 3
AGT CAG 1TG GTA TCA AAA CCG m gln leu Val ser lys pm 132
POU Swcitic Domain CAC ACT CCG AGT CTG GAT GAG GAC OGG ATC AAC lTA
leu asn glu ala g l u u mg am gln glu gly gln gln am leu mer glu phe Val 227 TTG AAC GAG GCAGAG TC Cffi AAC CAG GAA GGC CAG CAG AAT CTG ATG GAG 'ITT GTG
208 l y s l eu asp i l c thr pro l y s ser ala g ln l y s leu l y s pro val leu g l u l y s t rp A A G T T A G A C A T C A C A C C C A A G A G T G C C C A G A A G C T G A A G C C G G T T T T G G A A A A G T G G
189 ala leu thr ala thr g l u g I y pro ala tyr scr g ln scr ala i l c c y s arg phc glu G C T C T G A C C G C A A C A G A G G G G C C A G C C T A C A G C C A A T C A G C C A T C T G C A G G T T C G A G
170 l y s asn phc Iys i l e arg arg leu scr leu g l y leu thr g ln thr g l n Val g l y g ln AAG AAT TTT AAGATC CGG AGG CTC TCC CTG GGT CTG ACACAG ACC CAG GTG GGC CAG
151 g l u glu i l e arg g l u phe ala his thr pm ser leu arp glu arp ply ile am leu G A A G A G A T C C G G G A G T T T G C C
GGC GGC GAG CCC TCC AAG
arb arg g ln thr leu I y s asn thr ser lys leu am val phe gln ile pm **** 301 C G A C G C C A G A C A C T C A A G AA ACC AGC AAG CTG AAT GTC TlT CAG ATC CCC TAG ..... 3'
284 glu i l e ala Ips g l u leu asn ty r asp arg g l u Val Val arg Val trp phc cys asn G A G A T C G C T A A G G A G C T C AACTATGACCGCGAGGTGGTGAGGGTCTGGTTC TGT AAT
265 g l u ala leu asn ala tyr phe g l u l y r asn pro leu pro thr g I y g l n g l u i l e thr GAG GCT CTC AAT GCC TAC TTT GAGAAA AAC CCC CTG CCCACC GGC CAG GAG ATC ACC
246 l y s arg l y s arg arg thr scr phc thr pro g l n ala i l e gly gly glu pm ST lys AAA CGCAAG CGG CGT ACT TCC TTC ACA CCA CAG GCC ATA
FIG. 1. Nucleotide sequence and predicted amino acid sequence of Bm-6. The sequence shown includes in-frame stop codons 5' and 3' (designated ****) to the open reading frame. The POU-specific and the POU homeodomains are bored. The two direct repeats of 7 amino acids in the NH2 terminus are underlined. The numbers on the right refer to amino acids with the initial in-frame methionine assigned number 1.
POU Homeo Domain
and Young, 1992; Mathis et al., 1992; Gerrero et al., 1993). Because many POU domain genes were initially identified
using a polymerase chain reaction approach with a common set of oligonucleotides, the divergence of this family may have been underestimated. Using a modified PCR-basedl approach, we have cloned cDNAs encoding a new POU domain protein, Brn-5, which has a prominent neuronal expression and is char- acterized by a highly divergent POU domain.
EXPERIMENTAL PROCEDURES PCR Cloning of POU Domains-Poly(A+) RNA from rat anterior pi-
tuitary glands was used as a template for cDNA synthesis with random hexamer primers and reverse transcriptase (Superscript, Life Technolo- gies Inc.) according to instructions from the vendor. This cDNA was used as a template in PCR reactions with degenerate primers to con- served regions in the POU domain. A primer corresponding to the third helix of the POU-specific domain [5'-TAGAA'ITCA(AG)(AT)tCG)tGAT- C)AC(GATC)AT(ACT)TG(CT)(AC)G(GATC)'IT(TC)GA-3'] was used in combination with one of two primers corresponding to the third helix of the POU homeodomain [5'-TAGGATCCTG(GACT)(CG)o(GAT)(GATC)- (CT)(GT)(GA)'IT(AG)CAtAG)AACCA(GATC)AC-3' or 5"TAGGATC- CTG(GATC)(GC)(GAT)tGATC)(TC)(GT)(GA)'ITIT(GATC)(GC)(AT)(GA)A-
The abbreviations used are: PCR, polymerase chain reaction; e, embryonic day; CMV, cytomegalovirus; SAAB assay, Selected And Am- plified Binding site assay; bp, base paids); kb, kilobase(sl
ACCA(GATC)AC-3']. The conditions of the PCR were as follows: 30 cycles a t 94 "C for 1 min, 50 "C for 1 min, and 72 "C for 3 min. The PCR products were treated with restriction enzymes XhoI or BstXI and size- fractionated on a 2% agarose gel. Gel slices corresponding to full-length products were cut out, and the DNAwas isolated and used as a template for PCR with the same primers. The products from these reactions were cloned into a plasmid vector and sequenced.
Complementary DNA ClonineOne of the POU domains obtained in the screen described above was Bm-5. This fragment was 32P-labeled by random priming (Life Technologies Inc.) and used to screen a rat ante- rior pituitary cDNA library (Ingraham et d . , 1988). The cDNA clones obtained in this screen were sequenced on both strands as previously described (Andersen et al., 1993).
Messenger RNA Analyses-Northern blot analysis was done as pre- viously described (Yu et al., 1991). The probe was a random primed 32P-labeled DNA fragment corresponding to the coding region for amino acids 39 to 189. RNase protection assays were done as previously de- scribed (Yu et al., 1991). The probe was a 32P-labeled cRNA correspond- ing to the coding region for amino acids 186 to 281.
In Situ Hybridization--Rat embryos and adult brain were treated as previously described (Simmons et al., 1989; Andersen et al., 1993). The probe that was used in most experiments was a 36S-labeled sense or antisense RNA corresponding to the coding region for amino acids 39 to 186. In some experiments, we used a 282-bp antisense probe containing 167 bp of 5'-untranslated sequence in addition to the first 115 bp of the coding region. Similar results were obtained with both antisense probes.

23392 Bm-5 Is Enriched in Neocortex
I POU SPECIFIC DOMAIN I I- LINKER I
CLASS GENE (SP) 1 10 ao 40 YI
I
11
111
N
V VI
..
..
..
.. a. as
. . * * * * ** * *
I a1 1 - rn r a4 J
7 POU HOMEODOMAIN CLASS GENE (SP) 10 10 30 40 50
I
I1
III
IV
V VI
n i t - 1
act-1 oat-a skn-lali
brn-1
bra-4 brn-a
t.t-1
brn-3.0 brn-3.1
ocC-3I.
bra-5
........... ..?aI.aL#.. lI?#LO?DOL? .+ L..V.....
0. ......... Q .......... Q .......... Q .......... a .......... a .......... Y O A . . . . . . . .
aa.........
* * * * I HELIX 1 1 I HELIX2 I I HELIX 3
FIG. 2. Amino acid sequence comparison of mammalian POU domains. Known mammalian POU domains were assigned to classes I to V and compared as previously described (Rosenfeld, 1991). Bm-5 was placed in a new class (VI). The sequences are organized from left to right, and the lower halfis continuous from the top. The numbering systems for the POU-specific domain and the POU homeodomain are according to Assa-Munt et al. (1993) and Laughon (19911, respectively. Highly conserved residues are inside black and white boxes. Residues that are different in Bm-5 compared to most other POU domains are indicated with anasterisk. The four a helices in the POU-specific domain and the three predicted a helices in the POU homeodomain are indicated below. r indicates rat; h, human; and m, mouse. Other names that have been used for these mammalian POU domain factors are (for references see Wegner et al. (1993)): GHF-1 (for Pit-1); OTF-1, OBP100, NFIII, and NF-A1 (for Oct-1); OTF-2 and NF-A2 (for Oct-2); Oct-11 (for Skn-la); N-Oct-3 (for Bm-2); RHS2 (for Bm-4); Oct-6 and SCIP (for Tst-1); Bm-3 (for Bm-3.0); Oct-3 and Oct-4 (for Oct-3/4); Emb (for Bm-5).
Protein-DNA-binding Assays-The proteins used for gel-shift analy- ses were in vitro translated or expressed in bacteria using a T, pro- moter, as previously described (Ingraham et al., 1990; Andersen et al., 1993). For the SAAB assay, we expressed Bm-5 holoprotein as a gluta- thione S-transferase fusion. The Bm-5 protein was purified by gluta- thione agarose affinity chromatography (Smith and Johnson, 1988) fol- lowed by cleavage with factor X. We observed no difference in binding specificity between proteins prepared by the three different methods. Gel mobility shift assays were done as previously described (Andersen et al., 1993). All binding sites were 32P-labeled to similar specific activ- ity with T4 polynucleotide kinase. The sequences and origin of the binding sites have been previously described (Mathis et al., 1992) except for En, 5'-AAGGGGATCCAAATGTCAATTAAATATCAA-3'; POMC CE2, 5'-TCCTCAITAGTGATA'I"Il'ACCTCCAAATGC-3'; and I12 Oct, 5'-TTTGAAAATATGTGTAATATGTAAAACATlTFG-3'. The SAAB as- say was performed as previously described (Blackwell and Weintraub, 1990). The sequence of the template was: 5"CGATGAA'ITCC- TAAGCGCATNNNNNNNNGAGCTCAGATC TC-3'. The sequences of the primers used to amplify the template were: 5"CGAT- GAAITCCTAAG-3'(sense) and 5'-ACGAGATCTGAGCTC-3' (anti- sense). The conditions of the PCR were as follows: 25 to 35 cycles of 94 "C for 45 s,48 "C for 2 min, and 72 "C for 30 s. After three rounds of selection, the template was sequenced using a 32P-labeled sense primer as previously described (Blackwell and Weintraub, 1990).
Cell Culture and Dansfections-CV-1 cells were grown and trans- fected with the calcium phosphate method as previously described (Andersen et al., 1993). CMV Lex A Bm-5 contains the coding region for amino acids 1 to 146 fused in-frame with the DNA-binding domain of
LexA and placed downstream of the CMV enhancer/promoter as previ- ously described (Ingraham et al., 1990). CMV LexA Pit-1, 2X LexA -36 luciferase and -36 luciferase plasmids have been described before (In- graham et al., 1990).
RESULTS
Cloning and Characterization of Brn-5 cDNh-Degenerate oligonucleotides representing all possible codons in two differ- ent 9-amino acid conserved regions in Oct-1, Oct-2, Pit-1, and Unc-86 were initially utilized as primers for PCR to identify novel POU domain genes (He et al., 1989). However, with the subsequent cloning of the Oct-3/4 gene (Okamoto et al., 1990; Rosner et al., 1990; Scholer et al., 1990a), it became evident that the 5' region on which one of the degenerate oligonucleo- tide primers was based, located in the POU-specific domain, was not fully conserved. This observation raised the possibility that additional POU domain genes might have escaped detec- tion by the initial PCR approach. Therefore, we used an oligo- nucleotide to another highly conserved region in the middle of the POU-specific domain (5'-oligonucleotide) in combination with an oligonucleotide to the highly conserved third helix of the POU homeodomain (3'-oligonucleotide). The cDNA was generated from rat anterior pituitary mRNA using random hexanucleotides to increase the chance of detecting POU

Brn-5 Is Enriched in Neocortex 23393 domain transcripts with long 3"untranslated sequences. Be- cause Pit-l is a highly abundant POU domain transcript in the pituitary, we excluded Pit-1 transcripts by treating the ampli- fied material with either restriction endonuclease XhoI or BstXI, both of which cleave in the Pit-1 POU domain. The amplified material was then size-fractionated by agarose gel electrophoresis, and DNA corresponding to a full-length PCR fragment was isolated and reamplified. These products were subsequently cloned into a plasmid vector and analyzed by dideoxynucleotide sequencing.
We used this method to isolate several copies of a novel POU domain, referred to as Bm-5, which was selected for fur- ther analysis. The Bm-5 PCR fragment was used to screen a rat pituitary cDNA library, yielding three overlapping clones of 4.6 kb, 4.7 kb, and 2.1 kb denoted A Z A P Bm-5A, -B, and -C, respectively. All three clones contained an identical open read- ing frame of 900 bp (Fig. 1). The translation start site was as- signed based on an in-frame stop codon 5' to the putative ini- tiating ATG, as well as homology to a consensus Kozak sequence (Kozak, 1984). All three clones contained poly(A) tracks at their 3' end. Clones A and B were similar in that both contained 2.9 kb of 3'-untranslated sequences, whereas clone C contained only 750 bp of 3"untranslated sequences, suggesting the use of an alternative poly(A) signal site. Al- though heterogeneity was detected in the 5'-untranslated re- gions, this did not alter the open reading frame. Brn-5 is a single copy gene (data not shown), and we have localized the gene to mouse chromosome 15 and human chromosome 12 (Xia et al., 1993).
The POU domain of Bm-5 is unusual because it has signifi- cant alterations in several predicted amino acids that are in- variant or exhibit only conservative changes in previously de- scribed mammalian POU domains (Fig. 2). In the POU-specific domain, 11 of the highly conserved amino acids are different, and 6 of these changes are nonconservative. In the POU ho- meodomain, seven near-invariant amino acids are altered in Bm-5. Five of the changes in the POU homeodomain are non- conservative. The 20-amino acid linker region of Bm-5 bears no homology to any of the previously described POU proteins. Because of this high degree of divergence, we assigned Bm-5 to a new subclass (class VI) of the POU domain gene family (Fig. 2). Indeed, two highly similar sequences from zebrafish and mouse, referred to as pou[cl (Johansen et al., 1993) and Emb (Okamoto et al., 19931, respectively, were recently reported. The predicted amino acid sequences of pou[c] and Emb are 70% and 97% homologous to that of rat Bm-5, respectively. The pou[cl coding region contains a 302-amino acid NHz-terminal extension that is not in Emb or Bm-5. Other structural fea- tures of Bm-5 include an unusually short COOH terminus of only 10 amino acids. The N H z terminus, which is 144 amino acids long, bears no obvious homology to previously described POU domain genes. However, it features an over-representa- tion of prolines (17%), a characteristic which has been associ- ated with trans-activation domains in several other transcrip- tion factors (Mitchell and Tjian, 19891, including Oct-314 (Imagawa et al., 1991). In addition, the NHz terminus contains a 7-amino acid sequence, NAQGQVI, that is repeated twice (Fig. 1).
Expression of Brn-5 mRNA during Development and in the Adult-RNA blot analysis using RNA from multiple tissues and cell lines was used to determine the size and distribution of Bm-5 transcripts (Fig. 3). A major transcript of 5.5 kb was observed in several somatomammotroph pituitary cell lines (GC, MMQ, 235-1) and in a thyrotroph tumor (TtT 97). Upon longer exposure, low level expression was found in embryonic cell lines F9 and P19 and in the corticotroph cell line AtT-20, but no expression was detected in HeLa, HL-60, or CV-1 cells
9.5 - 1.5 - 4.4 -
2.4 - 1.4 -
of poly(A+) RNA from the indicated cell lines were size-fractionated on FIG. 3. RNA blot analyses of Bra-6 expression. Four micrograms
a 0.8% formaldehyde-agarose gel and transferred to nitrocellulose. The filter was hybridized with a random-primed 32P-labeled probe from the
Numbers on the left represent sizes (in kb) of RNA markers (Life Tech- NHz terminus of Bm-5. Tt97 refers to the TtT 97 thyrotroph tumor.
nologies Inc.) run on the same gel. All the RNA samples were intact and approximately equal in quantity as assessed by ethidium bromide stain- ing and hybridization to an RXRbeta probe (data not shown).
(data not shown). In addition, minor bands of 2.8 kb and 2.1 kb were found in pituitary cell lines and embryonic cell lines, respectively. Although the nature of these minor transcripts is unclear, the 2.8-kb mRNA is consistent with the use of the alternative poly(A) signal site found in A Z A P Bm-5C.
To further determine the pattern of Brn-5 expression, we performed RNase protection assays on RNAs from several dif- ferent rat adult and embryonic tissues in addition to cell lines (Fig. 4, A and B). In the adult, Brn-5 is most highly expressed in the brain but transcripts are also readily detected in kidney, lung, heart, skin, adrenal, and placenta. Low level expression was observed in spleen, muscle, liver, anterior pituitary, testis, and ovary. During development, expression was readily de- tected in the head a t embryonic day 15, with levels decreasing gradually until postnatal day 10. On embryonic day 17, expres- sion was detected in brainstem, cortex, and hypothalamus, with highest levels in the cortex. The low level of expression in testis appears to be developmentally controlled, being highest during the prepubertal stage (Fig. 4B). Together, these data suggest that Bm-5 is expressed a t highest levels in the central nervous system, but at lower levels in many different organs.
We utilized in situ hybridization to localize the expression of Bm-5 more precisely both during development and in the adult brain. A series of rat embryos ranging from stage e9.5 through e15.5 were probed with sense and antisense Bm-5 RNAprobes. No expression of Bm-5 was detected before e12.5 (Fig. 5, left panel, and data not shown). On e13.5 and e15.5, expression of Bm-5 was detected diffisely throughout the developing brain and spinal cord (Fig. 5, middle and right panels). Because Bm-5 signal could be detected in whole adult brain by RNase protection, it was likely that Bm-5 was either expressed dif- fusely or, alternatively, restricted with very high levels in ex- pressing cells. To distinguish between these possibilities, we camed out in situ hybridization studies on regularly spaced coronal brain sections. Expression was diffise throughout the brain, and, in general, the signal correlated with cell density, including regions such as the hippocampus and cerebellum.

23394 Brn-5 Is Enriched in Neocortex
4-290 bp
4-160bp
4 - 1 0 0 bp
B
4- 240 bp
Actin
FIG. 4. Expression of B m d in development and in the adult rat analyzed by RNaee protection atmay& A, 20 pg of total RNA from the indicated tissues, cell lines, and yeast tRNA were hybridized to a 3ZP-labeled antisense riboprobe from part of the POU domain of Bm-5. Atter treatment with RNase, the products were size-fractionated on a 5% denaturing polyacrylamide gel and analyzed by autoradiography. The numbers on the right show the size of the protected fragment in rat tissues (290 bp) as judged by the migration of a labeled 1-kb DNA ladder (Life Technologies Inc.). Hybridization to RNA of mouse origin gives rise to two fragments, 160 and 100 bp long, apparently due to sequence divergence between mouse and rat in this region of the Brn-5 gene. B50 and BI09 are neuroblastoma cell lines. CA 77 and GC are thyroid C cell and somatotroph cell lines, respectively. TtT 97 is a mouse thyrotroph tumor. d l 0 Head refers to a brain of a 10-day-old rat. B, 20 pg of total RNA from the indicated sources were hybridized to a Bm-5 (top panel) and P-actin probes (lower panel) and analyzed as described above.
FIG. 5 s i t u hyl
e 11.5 e 13.5 e 15.5
Localization of Bm-6 mRNA during development by in ridization. Rat embryos from the embrvonic stages indicated
I.
b1 at the iop were hybridized with-a 36S-labeled cRNAprobe corresponding to the NH2 terminus of Bm-5. Basal telencephalic plate ( B ) , cerebellum angle ( C ) , diencephalon (D), lateral ventricle (L), mesencephalon (M), rhombencephalon (R), spinal cord (S), and telencephalon (TI are indi- cated.
The one clear exception was a higher expression level found in neocortex, and, within the cortex, the signal was most intense in the inner granular layer (layer W, Fig. W). Observation under high magnification light field showed that silver grains were localized over neurons (data not shown). No signal was observed when we used a sense Brn-5 riboprobe (data not shown). An RNase protection assay using RNA from various brain regions was used to independently confirm the expres- sion pattern showed by the in situ hybridization studies. This experiment revealed that Bm-5 was expressed in all brain regions, with highest expression in the neocortex (Fig. 6B).
Taken together, the in situ hybridization and RNase protection data indicate that Bm-5 mRNA has widespread distribution in adult brain, and that the expression is enriched in the neocor- tex.
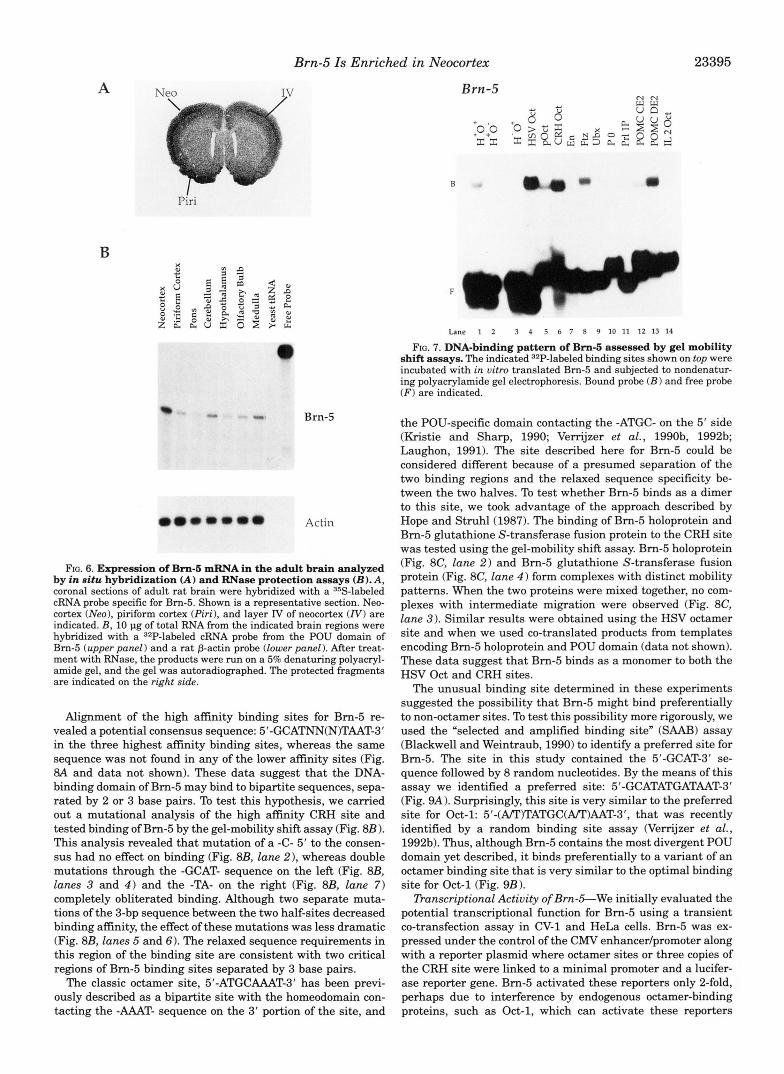
DNA-binding Properties of Brn-S-To test whether the diver- gent sequence of the Bm-5 POU domain led to different DNA- binding properties, we expressed Bm-5 both in bacteria and in uitro using rabbit reticulocyte lysate. Binding of the expressed protein was tested in the gel-mobility shiR assay using a series of AT-rich candidate binding sites for POU proteins or classic homeodomain proteins (Fig. 7). Bm-5 binding to the following elements was easily detected in order of decreasing affinity: HSV Oct, CRH Oct, POMC DE2, Ftz, pOct, and H+O+. Unex- pectedly, Brn-5 bound poorly to an element, H-O+, where the heptamer binding site was mutated. This suggests that, in contrast to classic octamer-binding proteins, the Bm-5 binding site overlaps with both the octamer and heptamer sequences. We could not detect binding to unrelated sequences, such as a thyroid hormone response element and a site that binds helix- loop-helix proteins (E box; data not shown). This binding pat- tern is different from that exhibited by Oct-2, which prefers all sites that contain a core octamer element: H’O’, H-O+, pOct, and I12 Oct (data not shown). Thus, consistent with the diver- gent sequence of the Brn-5 POU domain, its binding preference appears to be distinct from that of Oct-2.
Brn-5 Is Enriched in Neocortex 23395
A Neo
B
0. ”0.0
Brn-5
Actin
FIG. 6. Expression of Bm-6 mRNA in the adult brain analyzed by in situ hybridization (A) and RNase protection assays (B) . A, coronal sections of adult rat brain were hybridized with a 35S-labeled cRNA probe specific for Bm-5. Shown is a representative section. Neo- cortex (Neo), piriform cortex (Piri), and layer IV of neocortex (N) are
hybridized with a 32P-labeled cRNA probe from the POU domain of indicated. B , 10 pg of total RNA from the indicated brain regions were
Bm-5 (upper panel) and a rat p-actin probe (lower panel ). After treat- ment with RNase, the products were run on a 5% denaturing polyacryl- amide gel, and the gel was autoradiographed. The protected fragments are indicated on the right side.
Alignment of the high affinity binding sites for Bm-5 re- vealed a potential consensus sequence: 5’-GCATNN(N)TAAT-3’ in the three highest affinity binding sites, whereas the same sequence was not found in any of the lower affinity sites (Fig. 8A and data not shown). These data suggest that the DNA- binding domain of Bm-5 may bind to bipartite sequences, sepa- rated by 2 or 3 base pairs. To test this hypothesis, we carried out a mutational analysis of the high affinity CRH site and tested binding of Bm-5 by the gel-mobility shift assay (Fig. 8B 1. This analysis revealed that mutation of a -C- 5’ to the consen- sus had no effect on binding (Fig. 8B, lane 21, whereas double mutations through the -GCAT- sequence on the leR (Fig. 8B, lanes 3 and 4 ) and the -TA- on the right (Fig. 8B, lane 7) completely obliterated binding. Although two separate muta- tions of the 3-bp sequence between the two half-sites decreased binding affinity, the effect of these mutations was less dramatic (Fig. 8B, lanes 5 and 6). The relaxed sequence requirements in this region of the binding site are consistent with two critical regions of Bm-5 binding sites separated by 3 base pairs.
The classic octamer site, 5’-ATGCAAAT-3’ has been previ- ously described as a bipartite site with the homeodomain con- tacting the -AAAT- sequence on the 3’ portion of the site, and
B m - 5
B
k
(I
Lane 1 2 3 4 5 6 7 8 9 10 11 12 13 14
FIG. 7. DNA-binding pattern of Bm-6 assessed by gel mobility shift assays. The indicated 32P-labeled binding sites shown on top were incubated with in vitro translated Bm-5 and subjected to nondenatur- ing polyacrylamide gel electrophoresis. Bound probe ( B ) and free probe ( F ) are indicated.
the POU-specific domain contacting the -ATGC- on the 5’ side (Kristie and Sharp, 1990; Verrijzer et al., 1990b, 1992b; Laughon, 1991). The site described here for Bm-5 could be considered different because of a presumed separation of the two binding regions and the relaxed sequence specificity be- tween the two halves. To test whether Bm-5 binds as a dimer to this site, we took advantage of the approach described by Hope and Struhl(1987). The binding of Bm-5 holoprotein and Bm-5 glutathione S-transferase fusion protein to the CRH site was tested using the gel-mobility shift assay. Bm-5 holoprotein (Fig. 8C, lane 2) and Bm-5 glutathione S-transferase fusion protein (Fig. 8C, lane 4 ) form complexes with distinct mobility patterns. When the two proteins were mixed together, no com- plexes with intermediate migration were observed (Fig. 8C, lane 3) . Similar results were obtained using the HSV octamer site and when we used co-translated products from templates encoding Bm-5 holoprotein and POU domain (data not shown). These data suggest that Bm-5 binds as a monomer to both the HSV Oct and CRH sites.
The unusual binding site determined in these experiments suggested the possibility that Bm-5 might bind preferentially to non-octamer sites. To test this possibility more rigorously, we used the “selected and amplified binding site” (SAAB) assay (Blackwell and Weintraub, 1990) to identify a preferred site for Bm-5. The site in this study contained the 5’-GCAT-3’ se- quence followed by 8 random nucleotides. By the means of this assay we identified a preferred site: 5’-GCATATGATAAT-3’ (Fig. 9A). Surprisingly, this site is very similar to the preferred site for Oct-1: 5’-(AlT)TATGC(APT)AAT-3’, that was recently identified by a random binding site assay (Verrijzer et al., 1992b). Thus, although Bm-5 contains the most divergent POU domain yet described, it binds preferentially to a variant of an octamer binding site that is very similar to the optimal binding site for Oct-1 (Fig. 9B).
!lYanscriptional Activity of Brn-&We initially evaluated the potential transcriptional function for Bm-5 using a transient co-transfection assay in CV-1 and HeLa cells. Bm-5 was ex- pressed under the control of the CMV enhancer/promoter along with a reporter plasmid where octamer sites or three copies of the CRH site were linked to a minimal promoter and a lucifer- ase reporter gene. Bm-5 activated these reporters only 2-fold, perhaps due to interference by endogenous octamer-binding proteins, such as Oct-1, which can activate these reporters

Brn-5 Is Enriched in Neocortex
A SITE SEQUENCE BINDING
G G A T C C G C A T G C - T A A T G A +++ G C T C C T G C A T I A I T A A T A G +++ C C A A C I G C A T C C T T A A T T A +++
A T G ~ ~ ~ ~ ~ A T A T I T A A T C A ++ T I T C A T C C - 8 A A T A T ++
C I A : T G ~ C A I G - - T A A T C G ++ G T C A A G : A A T C G - T A A A ++ r c r r c ; a r A T c c - ~ A A T C A + T G ~ ~ ~ : G C A T A T - T C A T G R +
B 1 2 3 4 5 6 7
* ...................... ,, ......................... I c,\'I I 1 I . , I I ( ' . ( ; C ' : U " . \ ) 'I 'AATAcc
8 , ......................... ,, <; ................................ 4 . ................................. G C ......................... 5. ...................................... CCT .................. 6, ...................................... GCC .................. 'i, ............................................... C G ........... CONSENSUS: GCATNNNTAAT
1 2 3 4
FIG. 8. Identification of a consensus high affinity DNA-binding site for Bm-6. A, alignment of highest affinity binding sites for Bm-5 identified in Fig. 7. In some instances, two possible alignments are shown. Sense (SI and antisense (AS) strands are indicated. B, muta- tional analyses of the CRH site. The wild type CRH oligonucleotide or the indicated mutant sites were radioactively labeled, incubated with Bm-5 protein, and analyzed by the gel-mobility shift assay. Bound ( B ) and free ( F ) probes are indicated on the right. C , Bm-5 binds as a monomer. A radioactively labeled CRH site was incubated in the ab- sence of protein (lune 1 ), with Bm-5 holoprotein (lune 2) , with Bm-5 glutathione S-transferase fusion protein (lam 41, or with Bm-5 holo- protein and Bm-5 glutathione S-transferase fusion protein together (lane 3). Bound ( B l , Bm-5 glutathione S-transferase-DNA complex, and B2, Bm-5 holoprotein-DNA complex) and free ( F ) probes are indi- cated on the right.
(data not shown). To independently test whether Brn-5 con- tains a domain capable of transcriptional activation, a CMV LexA Brn-5 fusion plasmid containing the NH2 terminus of Brn-5 linked to a LexA DNA-binding domain, was co-trans- fected with a reporter plasmid where 2 LexA binding sites were linked to a minimal promoter and a luciferase reporter. Under these conditions, we observed a consistent &fold activation imprinted by the NH2-terminal Brn-5 information (Fig. 10, black bars). This activation was similar to the activation by CMV LexA Pit-1 (Fig. 10) that contains the transcriptional activation domain of Pit-1, a POU protein known to be capable of activating both the growth hormone and prolactin genes, as well as to that of CMV LexA Brn-4 (Mathis et al., 1992). The activation was specific because a Lex fusion containing the NH2 terminus of Skn-li, which is incapable of DNA binding, did not activate in these same experiments (data not shown). The ac-
A CTAG
L *
3- e
m Ir
Selected
B
FIXED SELECTED SITE SITE
5 ' -&ATATDATAAT- 3 ' 3"CGTATACTATTA-5'
BINDING SITES PREFERRED
Bm-5 ( ~ I ~ S ~ V I ) 5'- ATATGATAAT -3'
oct-l 11) 5'- $TA&+T -3'
FIG. 9. Binding site selection for Brn-6. A, a template containing the sequence 5'4CAT-3' followed by random nucleotides in 8 consecu- tive positions was used in a gel-mobility shift assay with Bm-5 protein. The original template, labeled random (on the lef t ) , and the selected template (on the right) were sequenced after three rounds of selection. B, the selected sequence is shown on the top, and a comparison with the preferred site for Oct-1 (Verrijzer et al., 1992b) is shown below.
tivation was site-dependent because a reporter plasmid with- out Lex sites was not stimulated (Fig. 10, white bars). These data suggest that Brn-5 can function as a transcription factor.
DISCUSSION Defining additional POU domain proteins is of interest be-
cause at least three of these factors, Pit-l (Li et al., 1990),Oct-2 (Corcoran et al., 19931, and unc-86 (Finney and Ruvkun, 1990) have been genetically shown to determine cell fate or differen- tiated function, suggesting that there may be important roles for each of the diverse members of this family. We have iden- tified a cDNA encoding a novel POU domain factor referred to as Brn-5, whose predicted amino acid sequence is highly di- verged from other POU domain proteins, thus defining a new class of POU domain factors (He et al., 1989; Johansen et al., 1993; Okamoto et al., 1993).
The POU domain is a bipartite DNA-binding domain com- posed of a POU-specific domain joined by a short linker se- quence to the POU homeodomain. The POU-specific domain contains four a helices, homologous to the helix-turn-helix mo- tif in the h repressor (Assa-Munt et al., 1993; Dekker et al., 1993). The structure of the POU homeodomain is thought to be similar to that of the classic homeodomains with a cluster of basic amino acids at the N H 2 terminus and three a helices (Laughon, 1991; Rosenfeld, 1991; Scholer, 1991). In fact, POU domains are highly conserved, even across species including Drosophila (Johnson and Hirsh, 1990; Billin et al., 1991; Dick et al., 1991; Lloyd and Sakonju, 1991; Treacy et al., 1991, 1992; Prakash et al., 19921, Caenorhabditis elegans (Finney et al., 1988), Xenopus (Agarwal and Sato, 1991; Frank and Harland, 1992; HinMey et al., 1992; Whitfield et al., 1993), zebrafish (Matsuzaki et al., 1992; Johansen et al., 19931, chicken (Petry- niak et al., 19901, and mammals (Rosenfeld, 1991; Ruvkun and Finney, 1991; Scholer, 1991; Hara et al., 1992; Wegner et al., 1993). Although the predicted structure of the Brn-5 POU do- main probably conforms to these general rules, it has alter- ations of many amino acids that are conserved across all pre- viously described mammalian POU domains. For instance, the serine residue that replaces a lysine at position 22 in the basic region of helix 1 of the POU-specific domain of Brn-5 creates a potential phosphorylation site for protein kinase A (Kennelly

Brn-5 Is Enriched in Neocortex 23397
2 X L e x A -36
[EqiGq-"- + m-""- Fold Induction
2X Lex A 1 2 3 4 5
I I I I I
- +
- +
-36 prl luc - 0 2X Lex A -36 prl luc + I
FIG. 10. Brn-6 contains a transferable activation domain. The indicated expression plasmids (3 pg) were co-transfected with a reporter plasmid (3 pg) without (white bars) or with (black bars) two Led binding sites upstream of the minimal prolactin promoter and luciferase. The control luciferase activity was 2,500 light units per plate. Results represent the mean and standard error of the mean from a triplicate transfection.
and Krebs, 1991). Similarly, Oct-3/4 contains a threonine at the same position. This is reminiscent of the basic region in the NH2 terminus of the POU homeodomain in which a phospho- rylation site regulates the binding activity of l'it-1 (Kapiloff et al., 1991) and Oct-1 (Segil et al., 19911, raising the possibility that activity of the POU-specific domain of Bm-5 and Oct-3/4 might also be regulated by phosphorylation. Interestingly, Brn-5 has an alteration in the DNA recognition helix (alanine instead of threonine at position 46) of the POU-specific domain. However, this residue may not be directly involved in DNA recognition (Dekker et al., 1993). Two of the altered amino acids in the POU homeodomain, at position 25 (leucine instead of a basic residue) and 55 (threonine instead of a basic residue), are involved in phosphate backbone contacts in binding of classic homeodomains to their cognate site (Laughon, 1991). The NH2 terminus of Brn-5 contains a 7-amino acid sequence, NAQGQVI, that is repeated twice. Although its function is unknown, it is likely to be important because it is completely conserved in the mouse. Furthermore, in zebrafish, one of the repeats is unchanged while the other has two conservative changes. Because the NH2 terminus is a transactivation do- main, it is tempting to speculate that this domain might be involved in protein-protein interactions that are required for transcriptional activation.
Brn-5 exhibits a pattern of expression that is qualitatively distinct from most other POU domain factors. The most strik- ing feature of Brn-5 gene expression is the widespread distri- bution of Brn-5 transcripts, somewhat similar to that of Oct-1 (Sturm et al . , 1988; Kambe et al., 1993). Whereas, Oct-314 and Tst-1 are expressed early in embryogenesis and in embryonic cell lines (Meijer et al., 1990; Okamoto et al., 1990; Rosner et al., 1990; Scholer et al., 1990a, 1990b; Suzuki et al., 19901, Brn-5 is expressed at low levels in these cell lines and is not detected by in situ hybridization in the rat embryo until embryonic day 12.5. Therefore, high levels of Brn-5 expression appear to be associated with the appearance of more differentiated cell phe- notypes.
Similar to most POU domain genes, Bm-5 is initially dif- fusely expressed in the developing central nervous system. However, in contrast to neuronally expressed POU domain genes of class I11 and IV, Brn-5 mRNA expression does not subsequently become restricted to limited regions of the nerv-
ous system. Instead, Brn-5 is expressed diffusely throughout the adult brain, and signal strength correlates with cell density in structures such as the hippocampal formation (Okamoto et al . , 1993) and cerebellum. A clear exception to this pattern is focally enriched expression in the neocortex, especially layer IV. In this regard, Brn-5 expression is distinct from Brn-2 and Tst-1, which are expressed in isocortex layers 2-5 and 5-6, respectively. In layer IV, sensory inputs from the thalamus or adjacent cortical regions terminate on neurons that connect to adjacent cortical laminae (Schmitt et al., 1981). Interestingly, we also observed enriched expression in the piriform cortex, which receives olfactory input. Therefore, Bm-5 joins the grow- ing list of POU domain genes that are expressed in neurons involved in sensory pathways. In C. elegans, unc-86 specifies several sensory neuronal pathways (Finney and Ruvkun, 1990) and in mammals the Bm-3 gene is highly expressed in sensory ganglia (He et al., 1989; Gerrero et al., 1993). Furthermore, various products of the Oct-2 gene (Stoykova et al., 1992) and Brn-2 (He et al., 1989) are expressed in the olfactory pathway. Therefore, it appears that several POU domain genes may have important roles in the development of neurons involved in dif- ferent aspects of processing sensory information in the mam- malian nervous system. Brn-5 is also expressed in several or- gans outside the nervous system. Expression of Brn-5 in testis becomes lower after puberty, suggesting that Bm-5 is ex- pressed in non-germ cells or in immature spermatogonia.
Although POU homeodomain DNA recognition is thought to be similar to that of the classic homeodomain, high affinity DNA binding to cognate sites also requires the POU specific domain (Sturm and Herr, 1988; Ingraham et al., 1990; Kristie and Sharp, 1990; Verrijzer et al., 1990b; Imagawa et al., 1991), which makes direct DNA contacts (Aurora and Herr, 1992; Verrijzer et al., 1992b). Like the POU domain, the DNA recog- nition sequences for POU homeodomain proteins also appear to be bipartite. Oct-1 binds with highest affinity to sites contain- ing an octamer motif: 5'-ATGCAAAT-3' (for review see Laughon (1991)). The 3' half of this site ( - U T - , or a variant thereon is recognized by the homeodomain whereas the 5' half (-ATGC-) is recognized by the POU-specific domain (Kristie and Sharp, 1990; Verrijzer et al . , 1990b, 1992b). Oct-1 can also bind to a different type of sequence, the so-called TAATGARAT motif (Sturm et al., 1987); however, it binds to this site with lower

23398 Brn-5 Is Enriched in Neocortex
affinity and may not require the POU-specific domain for this interaction (Verrijzer et al., 1990b). Because of the divergent sequence of the Brn-5 POU domain, it was of interest to deter- mine whether Bm-5 preferred octamer DNA-binding sites. These experiments are especially relevant because the ze- brafish homolog of Bm-5 was reported to represent the initial example of a non-octamer-binding POU domain protein (Johan- sen et al., 1993). Although our results indicate that Bm-5 is capable of binding to non-octamer sites containing the consen- sus sequence: B'-GCATNN(N)TAAT-3', a random binding site selection demonstrates that Bm-5 binds preferentially to a site containing a variant octamer element, surprisingly similar to that preferred by both Oct-1 (Verrijzer et al., 1992b) and a new class V POU domain protein.2 Therefore, our results are con- sistent with previous studies demonstrating that, although dif- ferent POU homeodomain proteins may recognize distinct se- quences, they generally have the ability to bind to certain sites containing the octamer motif (Singh et al., 1986; Sturm et al., 1987; Baumruker et al., 1988; LeBowitz et al., 1989; Poellinger and Roeder, 1989; Scholer et al., 1989; Meijer et al., 1990; Oka- moto et al., 1990; suzuki et al., 1990; Hinkley et al., 1992; Mathis et al., 1992).
Acknowledgments-We thank Dr. P. Sawchenko for dissection of adult brain and advice on in situ hybridizations, Drs. C. Glass, G. Kennedy, and M. Wegner for critical reading of the manuscript, Drs. H. Ingraham and C. Lin for giR of reagents, A. Naar for advice on DNA- binding sites, and Dr. R. Kurakawa for advice on the SAAB assay.
REFERENCES
Agarwal, V. R., and Sato, S . M. (1991) Den Biol. 147, 363-373 Andersen, B., Schonemann, M. D., Flynn, S . E., Pearse 11, R. V., Singh, H., and
Assa-Munt, N., Mortishire-Smith, R. J., Aurora, R., Herr, W., and Wright, P. E.
Aurora, R., and Herr, W. (1992) Mol. Cell. Biol. 12.45~5467 Baumruker, T., Sturm, R., and Herr, W. (1988) Genes & Deu. 2,1400-1413 Billin, A. N., Cockerill, K. A., and Poole, S . J. (1991) Mech. Ageing Deu. 94 ,7544 Blackwell, T. K., and Weintraub, H. (1990) Science 250,1104-1110 Bodner, M., Caatrillo, J.-L., Theill, L. E., Deerinck, T., Ellisman, M., and Karin, M.
Clerc, R. G., Corcoran, L. M., LeBowitz, J. H., Baltimore, D., and Sharp, P. A. (1988)
Corcoran, L. M., Karvelas, M., Nossal, G. J. V., Ye, Z.-S., Jacks, T., and Baltimore,
Dekker, N., Cox, M., Boelens, R., Verrijzer, C. I?, van der Vliet, P. C., and Kaptein,
Dick, T., Yang, X., Yeo, S., and Chia, W. (1991) Proc. Natl. Acad. Sci. U. S. A. 88,
Finney, M., and Ruvkun, 0. (1990) Cell 63,895-905 Finney, M., Ruvkun, G., and Horvitz, H. R. (1988) Cell 66,757-769 Fletcher, C., Heintz, N., and M e r , R. G. (1987) Cell 61. 773-781 Frank, D., and Harland, R. M. (1992) Development 116,439-448 Gehring, W. J. (1987) Science 2% 1245-1252 Gerrero, R., McEvilly, R., Turner, E., O'Connell, S., Jenne, K, Hobbs, M., and
Rosenfeld, M. G. (1993) Proc. Natl. Acad. Sci. U. S. A, , in press Hara, Y., Rovescalli, A. C., Kim, Y., and Nirenberg, M. (1992) Proc. Natl. Acad. Sci.
U. S. A. 89,3280-3284 Hatzopoulos, A. K, Stoykova, A. S. , Erselius, J. R., Goulding, M., Neuman, T., and
GNSS, P. (1990) Deuelopment 109,34%362 He, X., Treacy, M. N., Simmons, I). M., Ingraham, H. A,, Swanson, L. W., and
Rosenfeld, M. G. (1989) Nature 340, 35-42 He, X., Gerrero, R., Simmons, D. M., Park, R. E., Lin, C. R., Swanson, L. W., and
Rosenfeld, M. G. (1991) Mol. Cell. Bwl. 11, 1739-1744 Herr, W., Sturm, R. A,, Clerc, R. G., Comran, L. M., Baltimore, D., Sharp, P. A.,
Ingraham, H. A., Rosenfeld, M. G., Finney, M., Ruvkun, G., and Horvitz, H. R.
Hinkley, C. S., Martin, J. F., Leibham, D., and Perry, M. (1992) Mol. Cell. Bid . 12, (1988) Genes & Deu. 2,1513-1516
Holland, P. W. H., and Hogan, B. L. M. (1988) Genes & Den 2,773-782 638-649
Imagawa, M., Miyamoto, A,, Shirakawa, M., Hamada, H., and Muramatsu, M.
Ingraham, H. A,, Chen, R., Mangalam, H. J., Elsholtz, H. P., Flynn, S . E., Lin, C. (1991) Nucleic Acids Res. 19,4503-4508
R., Simmons, D. M., Swanson, L. W., and Rosenfeld, M. G. (1988) Cell 65, 51S529
Ingraham, H. A,, Flynn, S . E., Voss, J. W., Albert, V. R., Kapiloff, M. S., Wilson, L., and Rosenfeld, M. G. (1990) Cell 61, 1021-1033
Johansen, T., Moene, U., Holm, T., Fjose, A., and Krauss, S . (1993) Nucleic Acids Res. 21,475483
Rosenfeld, M. G. (1993) Science 260,7t?-82
(1993) Cell 73, 193-205
(1988) Cell 55,505418
Genes & Deu. 2, 1570-1581
D. (1993) Genes & Deu. 7,570-582
R. (1993) Nature 382,852-855
7645-7649
Hope, I. A., and Struhl, K (1987) EMBO J. S, 2781-2784
B. Andersen and M. G. Rosenfeld, unpublished observations.
Johnson, W. A., and Hirsh, J. (1990) Nature 343, 467470 bbe, F., Tsukahara, S. , Kato, T., and Seo, H. (1993) Biochim. Biophys. Acta
Kapiloff, M. S., Farkash, Y., Wegner, M., and Rosenfeld, M. G. (1991) Science 26S.
Kennelly, P. J., and Krebs, E. G. (1991) J. Biol. Chem. 266, 15555-15558 Kessel, M., and Gruss, F? (1990) Science 249, 374379 KO, H.-S., Fast, P., McBride, W., and Staudt, L. M. (1988) Cell 66, 135-144 Kozak, M. (1984) Nucleic Acids Res. 12, 857-872
Laughon, A. (1991) Biochemistry SO, 11357-11367 Kristie, T. M., and Sharp, P. A. (1990) Genes & Deu. 4, 2383-2396
Le Moine, C., and Young 111, W. S . (1992) Proc. Natl. Acad. Sci. U. S. A. 89,
LeBowitz, J. H., Clerc, R. G., Brenowitz, M., and Sharp, P. A. (1989) Genes & Deu.
Levine, M., and Hoey, T. (1988) Cell 66, 537440 Li, S . , Crenshaw 111, E. B., Rawson, E. J., Simmons, D. M., Swanson, L. W., and
Rosenfeld, M. G. (1990) Nature 347, 526-533 Lloyd, A,, and Sakonju, S . (1991) Mech. Ageing Deu. 38,87-102 Mathis, J. M., Simmons, D. M., He, X., Swanson, L. W., and Rosenfeld, M. G. (1992)
Matsuzaki, T., Amanuma, H., and Takeda, H. (1992) Bioehem. Biophys. Res. Com-
McGinnis, W., Levine, M. S., Hafen, E., Kumiwa, A,, and Gehring, W. J. (1984)
Meijer, D., Graus, A., Kraay, R., Langeveld, A,, Mulder, M. P., and Grosveld, G.
Monuki, E. S . , Kuhn, R., Weinmaster, G., Trapp, B. D., and Lemke, G. (1990) Mitchell, P. J., and Tjian, R. (1989) Science 246, 371-378
Science 249, 1300-1303 Muller, M. M., Ruppert, S., SchaEner, W., and Matthias, P. (1988) Nature SS6,
544-551 Okamoto, K., Okazawa, H., Okuda, A,, Sakai, M., Muramatsu, M., and Hamada, H.
(1990) Cell 80,461472 Okamoto, K, Wakamiya, M., Noji, S. , Koyama, E., Taniguchi, S . , Takemura, R.,
Copeland, N. G., Gilbert, D. J., Jenkins, N. A., Muramatsu, M., and Hamada, H. (1993) J. Biol. Chem. 268,7449-7457
Petryniak, B., Staudt, L. M., Postema, C. E., McCormack, W. T., and Thompson, C.
Poellinger, L., and Roeder, R. G. (1989) Mol. Cell. Biol. 9, 747-756 B. (1990) Proc. Natl. Acad. Sci. U. S. A. 87, 109%1103
Rakash, K, Fang, X.-D., Engelberg, D., Behal, A,, and Parker, C. S . (1992) Pmc.
Rosenfeld, M. G. (1991) Genes & Dev. 6,897-907 Rosner, M. H., Wgano, M. A, Ozato, K., Timmons, P. M., Poirier, F., Rigby, P. W. J.,
Ruvkun, G., and Finney, M. (1991) Cell 84,475478 and Staudt, L. M. (1990) Nature S46, 686-692
Scheidereit, C., Cromlish, J. A,, Gerster, T., Kawakami, K, Balmaceda, C.-G., Cume, R. A,, and Roeder, R. G. (1988) Nature 338, 551457
Schmitt, F. O., Worden, F. G., Adelman, G., and Dennis, S . G. (1981) The Organi- zation of the Cerebral Cortex. Proceedings of a Neurosciences Research Program Colloquium, The MIT Press, Cambridge, MA
1171,307-310
786789
3285-3289
3,1625-1638
EMBO J. 11, 2551-2561
mun. 187, 1-1453
Nature 308,42&433
(1990) Nucleic Acids Res. 18, 7357-7365
Natl. Acad. Sci. U. S. A. 89, 7080-7084
Schijler, H. R. (1991) Bends Genet. 7,323-329 Schijler, H. R., Balling, R., Hatzowulos, A. K, Suzuki, N., and Gruss, P. (1989)
EMBO J. 8,2551-2557. Scholer, H. R., Ruppert, S. , Suzuki, N., Chowdhury, K, and Gruss, P. (199Oa)
Schijler, H. R., Dressler, G. R., Balling, R., Rohdewohld, H., and Gruss, P. (1990b)
Scott, M. P., and Carroll, S . B. (1987) Cell 61,689-698 Scott, M. P., and Weiner, A. J. (1984) P m c . Natl. Acad. Sei. U. S. A. 81,4115-4119 Segil, N., Roberta, S. B., and Heintz, N. (1991) Science 264,1814-1816 Simmons, D. M., Arriza, J. L., and Swanson, J. L. (1989) J. Histotechnol. 12,
Singh, H., Sen, D., Baltimore, D., and Sharp, P. (1986) Nature 319, 154-158 Smith, D. B., and Johnson, K S . (1988) Gene (Amst.) 67, 3 1 4 0 Stoykova, A. S. , Sterrer, S . , Erselius, J. R., Hatzopoulos, A. K, and Gruss, P. (1992)
Sturm, R. A,, and Herr, W. (1988) Nature SS8,601-604 Sturm, R., Baumruker, T., Franza, B. R., Jr., and Herr, W. (1987) Genes & Deu. 1,
Sturm, R. A,, Das, G., and Herr, W. (1988) Genes & Deu. 2,1582-1599 Suzuki, N., Rohdewohld, H., Neuman, T., Gruss, P., and Schijler, H. R. (1990)
Tanaka, M., Grossniklaus, U., Herr, W., and Hernandez, N. (1988) Genes & Deu. 2,
Treacy, M. N., and Rosenfeld, M. G. (1992)Annu. Rev. Neumsci. 16, 139-165
Treacy, M. N., Neilson, L. I., Turner, E. E., He, X., and Rosenfeld, M. G. (1992) Cell Treacy, M. N., He, X., and Rosenfeld, M. G. (1991) Nature S O , 577-584
Verrijzer, C. P., Kal, A. J., and Van der Wet , P. C. (1990a) EMBO J. 9,1883-1888 Verrijzer, C. P., Kal, A. J., and van der m e t , P. C. (199Ob) Genes & Deu. 4,
Verrijzer, C. P., Strating, M., Mul, Y. M., and van der Vliet, P. C. (1992a) Nucleic
Verrijzer, C. P., Alkema, M. J., van Weperen, W. W., Van Leeuwen, H. C., Strating,
Wegner, M., Drolet, D., and Rosenfeld, M. G. (1993) Curr Opin. Cell Biol. 6,
Whitfield, T., Heasman, J., and Wylie, C. (1993) Deu. Biol. 166,361-370 Xia, Y.-R., Andersen, B., Mehrabian, M., Diep, A. T., Warden, C. H., Mohandas, T.,
McKelvy, R., Rosenfeld, M. G., and Lusis, A. J. (1993) Genornies, in press Yu, V. C., Delsert, C., Andersen, B., Holloway, J. M., Devary, 0. V., Naar, A M., Kim,
S . Y., Boutin, J.-M., Glass, C. K, and Rosenfeld, M. G. (1991) Cell 67,1251-1266
Nature 344,435-439
EMBO J. 9,2185-2195
169-181
Neuron 8,541658
1147-1160
EMBO J. 9,37233732
1764-1778
88,491405
1964-1974
Acids Res. 20,6369-6375
M. J. J., and van der Vliet, P. C. (1992b) EMBO J. 11,49935003
48a498