the multiple forms of mushroom tyrosinase - the · pdf filethe journal of biological chemistry...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 238, No. 5, May1963
Printed in U.S.A.
The Multiple Forms of Mushroom Tyrosinase
PURIFICATION AND MOLECULAR PROPERTIES OF THE ENZYMES*
SIMONE BOUCHILLOUX,~ P. MCMAHILL, AND H. S. MASON
From the Department of Biochemistry, University of Oregon Medical School, Portland, Oregon
(Received for publication, November 26, 1962)
We have undertaken the purification of mushroom tyrosinase to obtain homogeneous enzyme for a study of its mechanism, which is unknown. Purified mushroom tyrosinases have already been reported (l-9). Some of these preparations were ho- mogeneous, or almost homogeneous, by either electrophoresis or ultracentrifugation; one was homogeneous by both criteria (3, 9). However, there are discrepancies among them. Ac- cording to one report, the molecular weight is 34,000 (3) ; accord- ing to others, it is close to 100,000 (2,4-6,9). The copper which the enzyme contains is either partially (6) or entirely cuprous (10). Moreover, if tyrosinase is a single protein able to catalyze a-hydroxylation of monophenols and dehydrogenation of o-diphe- nols, the ratio of these two kinds of activities, often referred to as cresolase and catecholase activities, depends on the method of purification (2).
In 1949, Mallette and Dawson (2) obtained mushroom tyro- sinases with different physical and enzymic properties which they ascribed to chemical or physicochemical changes in an original single enzyme during the purification process. Their enzymes contained different amounts of copper and different ratios of cresolase to catecholase activity, but they could not be separated from one another by ultracentrifugation or electro- phoresis. Recently, Smith and Krueger (8) showed that five tyrosinase activities can be separated from mushroom extracts by chromatography on hydroxylapatite columns. One of their active fractions moved as a single substance during starch gel electrophoresis, but the other fractions were heterogeneous.
In this study, we have confirmed the multiplicity of mushroom tyrosinase. Using classical procedures and an efficient prepara- tive electrophoresis followed by chromatography on hydroxylapa- tite, we obtained four proteins: the (Y-, fl-, y-, and d-tyrosinases, three in homogeneous form. In this article, we describe the preparation of the enzymes and some of their enzymic, chemical, and physical properties.
EXPERIMENTAL PROCEDURE
Materials-Commercial Agaricus bispora was used as starting material. This is probably identical with the commercial mush- rooms used as source material in other studies and identified as Psalliota campestris or Agaricus campestris. p-Cresol and catechol were purified from Eastman Kodak Company reagents.
* This research was supported by grants from the United States Public Health Service, the National Science Foundation, and the American Cancer Society, to whom we are indebted.
t On leave from Centre Nationale de Recherche Scientifique and the Faculty of Medicine and Pharmacy, University of Mar- seilles, France.
Sodium dodecyl sulfate was recrystallized from ethanol from Matheson, Coleman and Bell, Inc., reagent.’ Hydroxylapatite was prepared according to the method of Tiselius (11). All other materials were best commercial grade, and deionized, distilled water was used throughout the study.
Estimation of Enzymic Activities-Although the assay condi- tions and units of monophenol hydroxylase and o-diphenol dehydrogenase activities were defined some years ago (12, 13), modifications and novel assay procedures have been introduced. The units recommended by the Commission on Enzymes ought to be generally adopted, but we have in this paper retained the classical units for such comparisons as are possible. We deter- mined activity towards p-cresol and catechol according to the method of Gregg and Nelson (12), using, however, smaller War- burg flasks and smaller total volumes, i.e. 3.2 ml rather than 8.0 ml, and making the appropriate correction. The chrono- metric unit of Miller, Mallette, Roth, and Dawson (13) was also used where noted. Activity toward n-tyrosine and DL-3,4- dihydroxyphenylalanine was determined manometrically with 1 mg of substrate per 3.2 ml, total volume, at 25”, the unit being the amount of enzyme required to catalyze the consumption of 1 ~1 of 02 per minute (cf- Lerner (14)).
Protein Estimation-For first approximations, we used the spectrophotometric determination of Eiger and Dawson (15). However, it became obvious with our more purified fractions that a large discrepancy exists between this method and a biuret determination based on a commercial, stabilized globulin stand- ard, Lab-Trol (Dade Reagents, Inc.). The biuret method gave from 64 to 68% of the values obtained spectrophotometrically at this stage.
Total Copper Estimation-These were performed by the method of Stark and Dawson (16). To liberate the maximal amount of copper from the purified samples, 0.4 N rather than 0.2 N HCl was required. All determinations were made in duplicate, and calculations were based on a standard curve determined simul- taneously under identical conditions.
Cuprous Copper Determinations-We used several modifica- tions of cuprous copper determinations (10, 17, 18). Our experi- ence with these determinations is discussed at greater length under “Results.”
Cupric Copper Determination-Cupric copper was determined by electron spin resonance spectroscopy. We used a Varian V-4500 spectrometer with loo-kc field modulation. Samples and standard (2 mM CuS04 in 20 mM EDTA) were examined under exactly the same conditions, usually at -165”. The
1 We are indebted to Dr. D. Rigas for this compound.
1699
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Purification and Properties of il/lushroom Tyrosinase Vol. 238, No. 5
spectra were recorded as derivatives of a.bsorption with respect to field strength ; the double integral of the unknown and standard were determined by graphical methods and compared. This procedure applied to other copper proteins in our laboratory has been shown to be accurate to within 2% at 0.1 InM Cu++, and measures cupric copper only. The difference between this esti- mation and total copper gives an independent measure of cuprous copper.
dmino Acid Bn&sis-Exploratory analyses2 were made with the Beckman/Spinco model 120 automatic analyzer, after 20 hours of hydrolysis at 110” by 6 N HCl, with the reaction tube sealed under vacuum (19-20).3 Tryptophan was determined separately by the method of Goodwin and Morton (21).
Free Boundary Electrophoresis-These determinations4 were made with the Spinco instrument, and the @- and y-enzymes in phosphate buffer, pH 6.8, ionic strength, 0.1.
Ultracentrifugal Analysis-Experiments to test the homogene- ity of our tyrosinases were performed, with the kind help of Dr. S. Inouye, with the Spinco model E centrifuge at 45,000 r.p.m., and solutions containing from 3 to 5.4 mg of protein per ml (biuret) in 5 mM NazHPOd.
A preliminary study of the behavior of an homogeneous tyro- sinase (p) in the presence of increasing amounts of sodium dodecyl sulfate was made by R. D. Wade, using a similar instrument at 59,780 r.p.m., and a double sector, capillary-type, synthetic boundary cell. The protein concentration was 2.68 mg per ml in 5 InM NasHPOh. The comparison solution contained the same amount of sodium dodecyl sulfate as the enzyme solution in each experiment.
Molecular Weight Determinations-These measurements were made3 with homogeneous /I- and y-tyrosinases with sedimentation equilibrium with the short column multichannel cell described by Yphantis (22). Values of the initial concentrations were ob- tained from synthetic boundary cell runs. A partial specific volume of 0.75 ml was assumed for the proteins.
Continuous Flow Electrophoresis-We used the refrigerated Beckman/Spinco model CP continuous flow electrophoresis apparatus. The enzyme extracts were always introduced at the cathodic upper side of the curtain. Preliminary experiments showed that most of the components in the extracts migrated towards the anode. Similar distributions were obtained with various buffers in the pH range 5.9 to 8.7, ionic strength, 0.02, and we adopted for routine use, Tris-HCl, pH 7.69, ionic strength, 0.02, with a current of 20 to 22 ma at 340 to 360 volts, the buffer flow and sample feed rates being dependent on protein concentra- tion (no more than 35 mg per ml for crude extracts). The en- zyme localization was determined by oxidation of tyrosine, p-cresol, or catechol. Protein distribution was determined spectrophotometrically at 280 rnp, after appropriate dilution and correction for the presence of pigment if necessary (15).
Chromatography on Hydroxylapatite-The gel was prepared according to Tiselius (11) in 0.001 M phosphate buffer, pH 6.8. A column 2.2 cm in diameter and 25 cm long was three-fourths filled with gel and washed with buffer. The sample, containing no more than 200 mg of protein in no more than 15 ml of buffer, was introduced, absorbed, and covered with a filter paper disk,
2 These were kindly made by Dr. R. Jones and Miss M. Duerst. a S. Moore and W. H. Stein, to appear in Methods in enzymology,
Vol. VI, published by Academic Press, Inc. 4 Kindly made by R. D. Wade, Department of Biochemistry,
University of Washington.
then washed twice with small amounts of buffer. An approxi- mately linear gradient of buffer concentration, from 1 rnM to 50 mM, was established by a two-vessel syphon, and the final con- centration was maintained until the third enzyme fraction (y- tyrosinase) was eluted. A new gradient from 0.05 M to 0.2 M
was then established. During the separation, buffer flow was controlled by a Milton Roy Minipump, usually at 4 ml/30 min- utes, this being the volume collected in each tube of an automatic constant interval collector. The optical transmittance at 280 rnp of the eflluent was recorded continuously; absorbancies of the effluent fractions of interest were determined later.
Concentration of Dilute Enzyme-It was often necessary to concentrate the dilute enzyme solutions obtained from continuous flow electrophoresis, chromatography, or after dialysis of am- monium sulfate fractionations of crude extracts. In the early stages, Carbowax 4000 was generally used around a dialysis bag containing the enzyme, but this was avoided in later steps. If the enzyme were not too dilute and already somewhat purified, it was brought to 0.7 saturation with ammonium sulfate, the precipitate was redissolved in a minimal volume, and the solution was dialyzed against any desired buffer. In case of higher dilu- tions of purified enzymes (for example, after chromatography), the dialysis bag containing enzyme was immersed in sucrose powder, in accordance with a suggestion of Dr. N. H. Horowitz, until a maximal decrease of volume had taken place. The solution was then dialyzed against buffer, and passed through a second sucrose cycle. The final dialyses against buffer were exhaustive in the case of the purified last stage enzyme.
Extraction of Tyrosinase-Fresh white mushrooms were frozen at -20” at least 1 day before extraction. We will describe now the first steps of enzyme preparation, modified from procedures reported by other investigators in the light of observations made in this laboratory over the last several years.
Step 1: Frozen mushrooms, 3 kg, were homogenized in 4 liters of acetone at -20” in a large Waring Blendor, for no more than 1 minute. The homogenate was filtered through a 40-cm Buch- ner vacuum filter covered by a large nylon fabric cap on which was powdered Dry-Ice, placed in contact with the pulp as soon as most of the acetone phase had been sucked away. The whole process was repeated with the partially frozen, once-extracted
pulp. Step 2: The pulp obtained after the second dehydration was
homogenized with 309, acetone in water (volume for volume) at 0” for 2 to 3 minutes in the same Waring Blendor. We used 3 liters of liquid per 3 kg of starting mushrooms. The mixture was centrifuged at 6000 r.p.m. in six 390-ml cups of the refriger- ated Servall model RC-2 centrifuge. It is convenient to use two centrifuges when large quantities of mushrooms are being worked
UP. Step 3: To the supernatant aqueous acetone extract obtained
in the last step, 1.5 volumes of acetone at -20” were added. The mixture was allowed to settle in a freezer at -20”; most of the clear supernatant fluid was decanted and discarded, the remain- der was centrifuged as in Step 2, and the precipitate was dissolved quickly in ice-cold water. Approximately 450 ml were used for the precipitate from a 6-kg extraction. This extract was then made 1 y0 in calcium acetate by slowly adding a 10% calcium acetate solution. (Addition of calcium acetate was finally shown to give no increase in specific activity, at least as described here, although other investigators apparently derived some advantage from its use.) The turbid mixture was frozen at -20”. It was
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
May 1963 S. Bouchilloux, P. McMahill, and H. S. Mason 1701
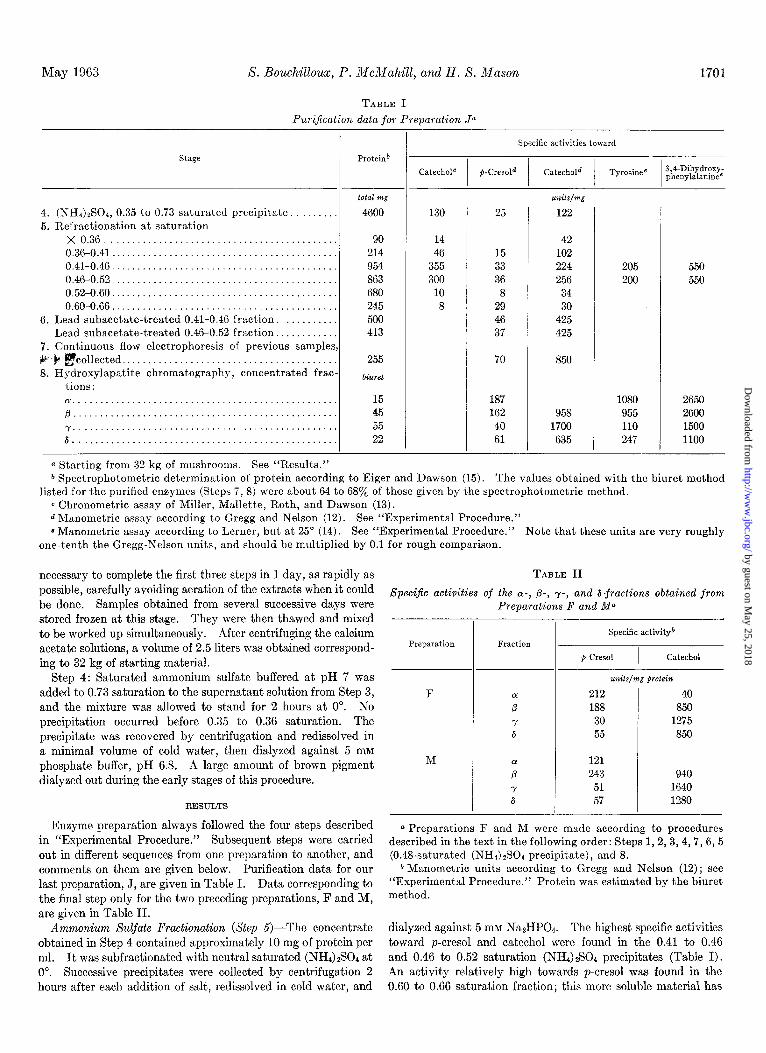
TABLE I Pur&ation data .for Preparation Ja
stage Proteinb
4. (NH4)zS0+ 0.35 to 0.73 saturated precipitate. . . 5. Refractionation at saturation
total mg
4600
X 0.36..........................................., 0.36-0.41.. . . . . . 0.41-0.46.. . 0.46-0.52.. . . 0.52-0.60.. . . . . . . 0.60-0.66.. .
6. Lead subacetate-treated 0.41-0.46 fraction. . . . . . Lead subacetate-treated 0.46-0.52 fraction..
7. Continuous flow electrophoresis of previous samples V .b gcollected.. . . . . 8. Hydroxylapatite chromatography, concentrated frac-
tions :
90 214 954 863 680 245 500 413
255
bilrrd
~.................................................. p . . . . . . . . . . . . . . y.,..,,............................................ 6..................................................
15 45 55 22
(1 Starting from 32 kg of mushrooms. See “Results.”
- Specific activities toward
130 25
14 46
355 300
10 8
15 33 36 8
29 46 37
70
187 162 40 61
42 102 224 256
34 30
425 425
850
958 1700 635
205 550 200 550
1080 2650 955 2600 110 1500 247 1100
b Spectrophotometric determination of protein according to Eiger and Dawson (15). The values obtained with the biuret method listed for the purified enzymes (Steps 7, 8) were about 64 to 68a/o of those given by the spectrophotometric method.
c Chronometric assay of Miller, Mallette, Roth, and Dawson (13). d Manometric assay according to Gregg and Nelson (12). See “Experimental Procedure.” 8 Manometric assay according to Lerner, but at 25” (14). See “Experimental Procedure.” Note that these units are very roughly
one-tenth the Gregg-Nelson units, and should be multiplied by 0.1 for rough comparison.
necessary to complete the first three steps in 1 day, as rapidly as TABLE II possible, carefully avoiding aeration of the extracts when it could 8 pec@c activities of the (Y-, p-, y-, and &fractions obtained from be done. Samples obtained from several successive days were Preparations F and Ma stored frozen at this stage. They were then thawed and mixed to be worked up simultaneously. After centrifuging the calcium acetate solutions, a volume of 2.5 liters was obtained correspond- ing to 32 kg of starting material.
Preparation
Step 4: Saturated ammonium sulfate buffered at pH 7 was added to 0.73 saturation to the supernatant solution from Step 3, F and the mixture was allowed to stand for 2 hours at 0”. No precipitation occurred before 0.35 to 0.36 saturation. The precipitate was recovered by centrifugation and redissolved in a minimal volume of cold water, then dialyzed against 5 mM phosphate buffer, pH 6.8. A large amount of brown pigment M
dialyzed out during the early stages of this procedure.
RESULTS
Enzyme preparation always followed the four steps described in “Experimental Procedure.” Subsequent steps were carried out in different sequences from one preparation to another, and comments on them are given below. Purification data for our last preparation, J, are given in Table I. Data corresponding to the final step only for the two preceding preparations, F and M, are given in Table II.
Q Preparations F and M were made according to procedures described in the text in the following order: Steps 1,2,3,4,7,6,5 (0.48~saturated (NH4) $304 precipitate), and 8.
b Manometric units according to Gregg and Nelson (12); see “Experimental Procedure.” Protein was estimated by the biuret method.
Ammonium Sulfate Fractionation (Step 5)-The concentrate dialyzed against 5 IRM Na2HP04. The highest specific activities obtained in Step 4 contained approximately 10 mg of protein per toward p-cresol and catechol were found in the 0.41 to 0.46 ml. It was subfractionated with neutral saturated (NHSGS04 at and 0.46 to 0.52 saturation (NH4)$04 precipitates (Table I). 0”. Successive precipitates were collected by centrifugation 2 An activity relatively high towards p-cresol was found in the hours after each addition of salt, redissolved in cold water, and 0.60 to 0.66 saturation fraction; this more soluble material has
-
Fraction Specific activity”
p-cm01 I
Catechol
units/mg protein
212 40 188 850 30 1275 55 850
121 243 51 57
940 1640 1280
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1702 PuriJication and Properties of Mushroom Tyrosinase Vol. 238, No. 5
not been examined closely, but it is probably analogous to the soluble high cresolase fraction observed by Mallette et al. (23).
Pigment Removal by Lead Subacetate (Step 6)-Most of the brown pigments (probably melanins) adsorbed to proteins were removed by stepwise additions of saturated aqueous lead sub- acetate at 0”. This opera.tion was delicate because excess lead subacetate removed enzyme as well as pigment; with care, 70% recovery could be achieved. To a solution containing approxi- mately 10 mg of proteins per ml, a volume of reagent equivalent to +T of the initial volume was added slowly with gentle stirring. The subsequent additions of reagent were made a few drops at a time, waiting 10 to 15 minutes after each. The pale brown super- natant (yellow if too much lead subacetate was added) was centrifuged at 16,000 r.p.m. and then exhaustively dialyzed first against water and then 5 mM Na2HP04. The specific activity of the supernatant either increased or remained the same, depending on the preparation. In some cases, we tried to increase recovery by retreating a solution of the pigmented precipitate, which, in general, had only weak activity, but this procedure should be avoided (see below).
Continuous Flow Electrophoresis (Step 7)-If the order here described (Steps 4,5,6, and 7) was followed, a solution containing 5 to 15 mg of proteins per ml gave a fairly narrow enzymic band separated from inactive proteins when subjected to continuous flow electrophoresis. Fig. 1 depicts the pattern obtained with a concentrate from 0.42 to 0.46 saturation ammonium sulfate precipitate from which most of the pigments had been removed by lead subacetate. The enzyme was well located, principally in tubes 15 to 19. Fig. 2 depicts the results obtained with cruder 0.35 to 0.73 saturation precipitate from which pigments had not been removed. In this case, the relative amount of inactive protein was much higher; brown pigments moved toward the anode faster than the enzyme, but only a partial separation was possible. The proteins within the enzyme fraction were probably
FIG. 1. Continuous flow electrophoresis of a 0.42 to 0.46 satu- rated (NH&SO, precipitate treated with lead subacetate. Initial concentration: 5 mg of protein per ml; Tris-HCI buffer, pH 7.65, ionic strength, 0.02; 21 ma, 360 volts. Absorbancy in tube 17, A 280 ,,,,, = 1.765; A330 m,, = 0.106; Ass5 mll = 0.088. Pigmentation increased slightly toward the anode. Specific activities in tube 17: toward catechol (12), 1280; 3,4-dihydroxyphenylalanine (14), 3400, the highest value observed (cf. Table I). The reason is un- known.
0 0 ENZYME (formation of
i dopochrome
I 5 IO 15 20 25 30 Tube Number
FIG. 2. Continuous flow electrophoresis of a crude 0.35 to 0.73 saturated (NH&Son precipitate containing 33.5 mg of protein per ml, specific activity toward catechol (12), 100. Tris-HCl buffer, pH 7.65, ionic strength, 0.02; 20 ma, 340 volts. O--O, absorbancy at 280 rnp, each tube diluted 1:5; A--A, pigment absorbancy (15); X---X, absorbancy at 475 rnp 20 minutes after addition of 20 ~1 of diluted eluate to 1 ml of 0.5 mM tyrosine, pH 6.8.
PREPARATION M
. D IO 20 30 40 50 60 70 SO 90 100
G k40 PREPARATION J N
p" P Y
2.0 -
- I 20 40 60 SO 100 120
Tube Number
FIG. 3. Chromatography of purified tyrosinase on hydroxylapa- tite. Upper graph, elution pattern after 120 mg of protein, Prep- aration M, was applied to a 2.2- X 16-cm column equilibrated with 1 mM phosphate buffer, pH 6.8 (see “Experimental Procedure”). The 0.05 to 0.2 M buffer gradient was begun at tube 62. Lower graph, elution pattern after 180 mg of protein, Preparation J, was applied to a 2.2- X 17.5-cm column. The 0.05 to 0.2 M buffer gradient was begun at tube 80. In both graphs, 4.2-ml fractions were collected from the column every 30 minutes.
heterogeneous; they could not be successfully resolved by this technique.
Hydroxylapatite Chromatography (Step @-The pattern of elution of protein fractions from hydroxylapatite columns is depicted in Fig. 3, and properties of the fractions are given in Tables I and II. The tube contents corresponding to each peak were collected, excluding the very diluted portions, the overlap- ping portions, and portions of suspected heterogeneity, e.g. tubes 75 to 79 in the b-peak of Preparation M (Fig. 3). The a-peak of Preparation J was clearly heterogeneous, but the second section
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

May 1963 S. Bouchilloux,FP. McMahill, and H. S. Mason
(a’) possessed cnzymic specificities similar to those of the main a-peak itself. This feature, as well as the possible heterogeneity of the &peak in this preparation may have been connected with an attempt, in this case, to increase yield by subjecting the pig- mcntcd lead subacetate precipitate to a lead subacetate refrac- tionation. This procedure should, accordingly, be avoided.
SpeciJk Activities of CY-, fl-, y-, and &Tyrosinase-These activi- ties are listed in Table I (Preparation J) and Table II (Prepara- tions F and M). The results are comparable. The CC fraction, as observed by Smith and Krueger (S), is able to oxidize catechol but is rapidly inactivated during the reaction. It is, however, very active towards 3,4-dihydroxyphenylalanine and p-cresol. The /3-tyrosinase had high cresolase and high catecholase activi- ties. The y-tyrosinase has the highest catecholase and lowest cresolase activities. &Tyrosinase, like the P-tyrosinase, pos- sesses both activities although at lower levels of specific activity. Thus, a-tyrosinase can be considered “high-cresolase”; y-tyro- sinase, “high-catecholasc”; and p- and Ctyrosinases, mixed catecholase-cresolases. These ratios of phenol o-hvdroxylating and o-diphenol dehydrogrnating activities do not extend to other sets of substrates. The activity of cr-tyrosinase (“high-cresol- ase”) towards 3,4-dihydroxyphenylalanine is greater than that of y-tyrosinase (“high-catecholase”), and one cannot generalize concerning the ratio of activities.
Non<, of the four tyrosinases possessed lactase activity when hydroquinone was used as substrate.
Enzyme Stability-We did not systematically study enzyme stability, but decreased activity was always observed after pro- longed storage, particularly toward p-cresol. Purified enzymes were divided into small portions and frozen below -20”; as a rule, they were less stable than partially purified preparations.
Homogeneity of o(-, /3-, y-, and b-Tyrosinases-The p- and y- tyrosinases (Preparations F and J) moved as single components during ultracentrifugation (Fig. 4). Several determinations of s20,2u extrapolated to zero concentration gave a value of 7.25 for both. &Tyrosinase (Preparation M) was essentially homo- geneous (Fig. 4) ; it was not studied at several concentrations, but the once-determined sedimentation coefficient agreed with that of the others. Lu-Tyrosinase, although obtained from well defined rhromatographic peaks (Preparations F and XI), was heterogeneous by ultracentrifugation (Fig. 4). The heavier component would probably have an ~~0,~ slightly lower than the values found for the fi- and y-tyrosinases; a substantial amount of a light component was present in all three preparations of oc-tyrosinase, although in different proportions. A lighter com- ponent was also found in the Ltyrosinase of Preparation J.
/3- and y-Tyrosinases were essentially homogeneous by analyti- cal electrophoresis at pH 6.8 (Fig. 5), although there was an indication of some faster migrating material in y-tyrosinase. An average of two determinations of electrophoretic mobilities gave, for fi-tyrosinase, -6.07 X lo+ (descending) and -6.07 x 1 Oe5 (ascending) ; for y-tyrosinasc, the values were - 6.01 x 1 OW5 (descending) and -6.40 x lo+ (ascending).
d~olecular Weights-The molecular weights of fl- and y- tyrosinases (Preparation J) found by sedimentation equilibrium (see “Experimental Procedure”) were 118,600 and 119,500 for /3, and 118,800 for y. The precision was ~10% (22).
Dissociation oj /I-Tyrosinase in Presence of Sodium Dodecyl Sulfate-The molecular weight of /%tyrosinasc (Preparation J) was taken as 119,000. Sodium dodccyl sulfate was added in molecular ratios of 400, 266, and 133 to 1. Ultracentrifugal
analysis of the mixtures (see “Experimental Procedure”) showed the appearance of a new, light component (Fig. 6) which com- prised 33% of the total protein in the case of the highest ratio, but no enzymic activity was lost. The SZO.~ values for the en- zyme control, and the 400, 266, and 133 molecular ratios were, respectively, (heavier component) 6.61, 6.50, 6.39, and 6.51; and (lighter component) 2.57, 2.49, and 2.33, in the corresponding runs. This experiment indicates that P-tyrosinase dissociated into subunits in the presence of sodium dodecyl sulfate, although the possibility of unfolding rather than dissociation cannot be
c
.A”.--
Fro. 4. Sedimentation patterns of the tyrosinases. Solutions in 5 mM Na2HP04: rotor sneed. 45.000 r.o.m. LY fraction (Preaara- tion F) : 3.0 mg of’ protein-per ‘ml;’ average temperature‘ = 27.4”. Photographs were taken at 22, 54, and 86 minutes. SZO,~ of the heavier component = 6.41; of the lighter component, = 1.49. Pfraction (Preparation F) : 4.45 mg of protein per ml, average tem- perature = 28”. Photographs were taken at 21, 53, and 85 min- utes. spO,ur = 6.03. y fraction (Preparation F): 3.70 mg of pro- tein per ml, average temperature = 26.8”. Photographs were taken at 20, 52, and 84 minutes. .sP~,~ = 6.24. 6 fraction (Prep- aration M): 5.90 mg of protein per ml, average temperat.ure = 27”. Photographs were taken at 26, 58, and 74 minutes. sZO+ = 6.20.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1704 Purification and Properties 01 Mushroom Tyrosinase Vol. 238, No. 5
excluded. A value of 34,500 was once observed for the molecular weight of mushroom tyrosinase (3) ; the sB ,W values now observed for the “subunit” would be consistent with this value. A sub- unit containing one atom of copper per molecule (@-tyrosinase contains 0.19 $& copper) would have a minimal molecular weight of approximately 30,000.
Amino Acid Analysis of p-, y-, and &Tyrosinases-Table III presents the amino acid composition of /3-, y-, and Ltyrosinases derived from a ZO-hour acid hydrolysis. Threonine, serine, proline, cystine, and tyrosine may have undergone some decom- position, and tryptophan is known to be lost under our conditions. More complete analyses will be reported in another paper. How- ever, Table III indicates that a striking likeness exists among the
three tyrosinases. The total recovery does not include trypto- phan (approximately 6% by the method of Goodwin and Morton (21) in fl- and y-tyrosinases, Preparation J) nor ammonia. Tyrosine estimated spectrophotometrically was found to be approximately 30% higher than the Stein and Moore analysis value. Several small, unknown components were also detected during the chromatographic analysis.
Absorption Spectrum-The four tyrosinases were pale yellow in solution and showed very similar absorption spectra, a maxi- mum at 280 to 281 rnp, a shoulder at 290 rnp, and a low, broad shoulder between 320 and 360 rnp (Fig. 7) with At80 = 2.45 to 2.65 (1 mg per ml, 1 cm) for the homogeneous fl-, y-, and 6- tyrosinases. The ratio of the absorbancies at 330 rnp and 280 rnp were variable even for purified enzymes. They ranged from 0.11 to 0.06, and tended to increase during prolonged storage with the concomitant development of a pale brown color. The mechanism of this process is unknown.
Total Copper Content-The copper contents of the purified tyrosinases are given in Table IV. The values were variable, even for the same type of tyrosinase obtained in different prepa- rations. If one assumes a molecular weight of 119,000, four atoms of copper per molecule corresponds to a copper content of 0.21%, the closest agreement, at least for /3- and y-tyrosinases. In spite of precautions to avoid contamination from water,
FIG. 5. Electrophoretic patterns of p- and y-tyrosinases: 4.45 mg of protein per ml (Preparation F) phosphate buffer, pH 6.8, ionic strength, 0.1,8.84 volts per cm. Photographs were taken at 108 minutes (P-tyrosinase, left) and 111 minutes (y-tyrosinase, right). Ascending (ax.) and descending (desc.) boundaries are indicated by arrows.
FIG. 6. Sedimentation pattern of p-tyrosinase in the presence of sodium dodecyl sulfate. Solutions in 5 mM NaQHPOd; rotor speed, 59,700; 2.68 mg of protein per ml (Preparation J), tempera- ture between 21.1” and 24.2”. Photographs were taken at zero and 24 minutes. A, no sodium dodecyl sulfate; B, 133 moles of sodium dodecyl sulfate per mole of enzyme; C, 266 moles of sodium dodecyl sulfate per mole of enzyme.
reagents, and glassware, the presence of extraneous copper cannot be ruled out.
Cuprous Copper Content-In determining cuprous copper, the copper is usually released from enzyme by acid. If it is initially in the cuprous state, it may either autoxidize or be bound to a cuprous-specific reagent such as biquinoline, forming a complex which is estimated calorimetrically. If initially in the cupric state, the copper may be reduced by groups on the protein. These circumstances make cuprous copper determination ambig- uous. Our results with y-tyrosinase (Preparation J) are shown in Fig. 8. Similar results were obtained with the cy, 0, and 6 fractions of the same preparation. In the absence of added re- ducing agent, the development of the cuprous-biquinoline color was slow and no well defined plateau was reached, in contrast to the results of Kertesz (10). Ascorbic acid added any time
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

May 1963 8. Bowhilloux, P. McMahill, and H. S. Mason 1705
after the start of the reaction immediately increased the cuprous- biquinoline concentration to roughly the same value, but p- chloromercurisulfonic acid had no effect on the course of the appearance of cuprous-biquinoline. It is clear that cuprous- biquinoline forms slowly after the decomposition of tyrosinase
TABLE III
Amino acid analysis of the p, y, and &fyrosinasesa
Amino acid Amino acid residue” Amino acidC
-_ g/100 g protein
Lysine. . . . . Histidine. . . . Arginine. . Aspartic acid. . . Threonine . . . Serine. . . . Glutamic acid . . Proline. . . . . Glycine . . . . Alanine. . . Half-cystine. . . . . Valine.. . Methionine. . . Isoleucine Leucine . . . . . Tyrosine . Phenylalanine . .
4.20 4.52 4.42 2.88 3.28 2.88 5.25 4.72 4.46
10.93 11.22 11.02 4.75 5.18 5.18 2.99 2.85 3.14
10.96 11.17 11.17 4.89 4.81 4.41 3.50 3.55 3.64 3.28 3.34 3.40
4.25 4.41 4.25 1.76 0.77 1.87 4.66 4.66 4.37 6.37 6.65 6.46 4.52 4.66 4.52 5.81 5.93 5.93
Total.. . 81.00 81.82 81.12 -
_-
1
-
39 25 40
113 56 41
101 60 73 55
Tracer 51 16 49 67 33 47
865
42 41 28 25 36 34
116 114 61 61 39 43
103 103 59 54 74 76 56 57
Traces I ‘. Traces 53 51 7 17
49 46 70 68 34 33 48 48
875 -
871
a p- and r-Tyrosinases from Preparation F, &tyrosinase from Preparation M.
* Biuret determination. c Assumed molecular weight, 119,000.
I I I I I I I
240 280 320 360 rnp FIG. 7. Absorption spectrum of P-tyrosinase, 0.417 mg per ml
in 5 mM Na2HP04.
TABLE IV
Total copper and copper valence distribution in purified tyrosinases
Preparation
a, J.............. c-x, M............. p, J 8, M. . . . 0, F. . . r,J.............. r,M............. ?, F . 6, J . 6, M .
Total Cua CuYESR)* Cu+’ (difference)
pg/100 nzg protei?zc
0.13 0.02 0.11 0.18 0.08 0.10 0.19 0.04 0.15 0.24 0.07 0.17 0.23 0.10 0.13 0.20 0.03 0.17 0.23 0.07 0.16 0.23 0.04 0.19 0.11 0.32
a Determination according to Stark and Dawson (16); see “Experimental Procedure.”
* ESR analysis; see “Experimental Procedure.” c Biuret determination.
r
Time in Minutes
FIG. 8. Time course of the reaction between tyrosinase copper and 2,2’-biquinoline. V--V; 0.1 ml containing 0.535 mg of -y-tyrosinase (Preparation J, 0.20% total Cu) was mixed with 0.2 ml of 0.050/, (weight per volume) p-chloromercuriphenylsulfonic acid, and finally with 0.7 ml of 0.1% (weight per volume) 2,2’- biquinoline in glacial acetic acid; after 202 minutes, excess as- corbic acid was added. O--O, the same reaction system with- out p-chloromercuriphenylsulfonic acid; ascorbic acid was added after 35 minutes. O---U, the same reaction system as the last, but ascorbic acid was added 2 minutes after zero time. The dashed line on the left margin (Total Copper) indicates the ab- sorbancy corresponding to the total enzymic copper.
and forms more rapidly in the presence of ascorbic acid, but the underlying reasons for this behavior are not known.
Cupric Copper Determination-Electron spin resonance spec- troscopy has been used for the determination of cupric copper successfully, both in this laboratory and in others (24). All samples of purified tyrosinases gave cupric copper signals, such as depicted in Fig. 9. The concentration of cupric copper is given in Table IV. We have been unable to observe tyrosinase copper entirely in the cuprous form (lo), but we substantiate Krueger’s preliminary note (6) stating that both forms of copper exist in the enzyme. On mixing of tyrosinase with glacial acetic acid, all copper was converted into cupric copper according to the ESR criterion. Hydrogen peroxide appeared to have no effect on the native enzyme, but in the presence of urea (4 M),
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1706 Purification and Properties of Mushroom Tyrosinase Vol. 238, No. 5
4---GfAg=ioo \ . r-
l+-$%+l V FIG. 9. Derivative curve depicting a tvpical electron spin
resonance spectrum of a mushroom- tyrosmase (-y-tyrosinase). Temperature = -165”; modulation amplitude, 32 gauss. The arrol*, indicates the position of free spin (g = 2.00).
the copper was again converted entirely to cupric copper. The functional significance of cupric copper in tyrosinase is now under investigation.
The electron spin resonance spectra of all tyrosinases were typical of amino acid chelates of cupric copper (24), with hyper- fine splitting of approximately 180 gauss and some indication of two bands in the g = 2.0 region, due perhaps either to more than one type of cupric copper binding, or to the presence of free radical in addition to cupric copper.
DISCUSSION
That mushroom tyrosinase exists in multiple forms, indicated by Smith and Krueger (8), has now been confirmed by the isola- tion of purified enzymes. Smith and Krueger chromatographed on hydroxylapatite a crude concentrate obtained by extraction with 0.1 M NaCl solution and precipitation with ammonium sulfate at 0.7 saturation. They found five active components, while we observed only four. The difference may arise from the presence of a very soluble tyrosinase in the 0.66 to 0.70 saturated fraction which we set aside and have not yet examined. Otherwise, the relative specificities of our four tyrosinases agree with those found by Smith and Krueger in their first three and fifth peaks. Our absolute activities correspond more closely to those of Mallette and Dawson (2) than those of Smith and Krueger for reasons as yet unknown. Perhaps identical assay techniques would yield more consistent specific activities in differ- ent laboratories.
Multiplicity of tyrosinases has now been demonstrated at several levels of the phylogenetic scale (25-33). The identical or almost identical sedimentation constants and electrophoretic mobilities of the mushroom tyrosinases may explain the failure of other investigators who did not use hydroxylapatite chroma- tography to demonstrate heterogeneity of their apparently pure preparations. In one case (Neurospora), the molecular weights of the multiple enzymes have been shown to vary from 34,000 to over 100,000 (31), the lightest enzyme presumably containing only one copper atom per molecule, an important feature from the point of view of mechanism. Our results with sodium dodecyl sulfate are consistent with the hypothesis that subunits are involved in tyrosinase structure.
If tyrosinases are built up of subunits, the recombination of identical subunits in slightly different ways could lead to multi- plicity. Alternatively, two or more kinds of subunits may exist, each under genetic control and possessing characteristic enzymic activities towards monophenols and o-diphenols. Their random recombination would lead to “hybrid” tyrosinases (isoenzymes) and account for the multiplicity in another manner. Finally, the possibility that tyrosinase multiplicity is an artifact of isola-
tion has not been strictly ruled out, although the work of Smith and Krueger (8) on this point makes the hypothesis unlikely.
SUMMARY
Multiple forms of mushroom tyrosinase have been obtained in homogeneous state by a process involving preparative elec- trophoresis and chromatography on hydroxylapatite. Four enzymes, OL-, /3-, y-, and &tyrosinase, were obtained, the last three essentially pure. Although these enzymes possessed par- tially different activities towards mono- and o-diphenols, the three homogeneous ones had very similar amino acid composition. Both cuprous and cupric copper were present in each enzyme. Apparent dissociation into subunits with retention of activity occurs in the presence of sodium dodecyl sulfate.
Acknowledgments-We wish to express our gratitude to R. D. Wade, of the Department of Biochemistry, University of Wash- ington, for help in characterizing our enzymes; to Professor H. Neurath for the generous hospitality of his Department; to Dr. S. Inouye for much assistance with analytical ultracentrifuga- tion; to Dr. Richard Jones and Miss Marie Duerst for their pains- taking amino acid analyses and helpful discussion; and to Dr. D. Rigas for stimulating comment.
1.
2.
3. 4.
5.
6. 7.
8.
9.
10. Il.
12.
13.
14.
15. 16. 17. 18.
19.
20.
21.
22. 23.
24.
25.
REFERENCES
KEILIN, D., AND MANN, T., Proc. Roy. Sot., Ser. B, 125, 187 (1938).
MALLETTE, M. F., AND DAWSOX, C. R., Arch. Bioch,em., 23, 29 (1949).
KERTESZ, D., AND ZITO, R., Nature, 179, 1017 (1957) YASUNOBU, K. T., THOMSON, L. C. G., AND MASON, H. S.,
Federation Proc., 16, 275 (1957). FRIEDEN, E., AND OTTESEN, M., Biochim. et Biophys. Acta, 34,
248 (1959). KRUEGER, R. C., Federation Proc., 18, 267 (1959). SMITH, J. L., AND KRUEGER, R. C., Federation Proc., 20, 48
(1961). SMITH. J. L., AND KRUEGER. R. C.. J. Biol. Chem.. 237, 1121
(196i). ’ KERTESZ, D., AND ZITO, R., in 0. HAYAISHI (Editor), Osy-
uenases. Academic Press, Inc., New York. 1962. I). 307. KERTEsz,‘D., Nature, MO,‘506 (1957). ’ ’ ^ TISELIUS, A., HJERT~N, A., AND LEVIN, ii., Arch. Biochem.
Biophys., 66, 132, (1956). GREGG, D. C., AND NELSON, J. M., J. Am. Chem. Sot., 62,
2500 (1940). MILLER, W. H., MALLETTE, M. F., ROTH, L. J., AND DAWSON,
C. R., J. Am. Chem. Sot., 66, 514 (1944). LERNER, A. B., in S. P. COLOWICK AND N. 0. KAPLAN (Edi-
tors), Methods in enzymology, Vol. II, Academic Press, Inc., New York, 1955, p. 827.
EIGER, I., AND DAWSON, C. R., Arch. Biochem., 21, 181 (1949). STARK, G. R., AND DAWSON, C. R., Anal. Chem., 30,191 (1958). FELSENFELD, G., Arch. Biochem. Biophys., 87, 247 (1960). GRIFFITHS, D. E., AND WHARTON, D. C., J. Biol. Chem., 236,
1857 (1961). HIRS, C. H. W., STEIN, W. H., AND MOORE, S., J. Biol. Chem.,
211, 941 (1954). SPACKMAN, D. H., STEIN, W. H., AND MOORE, S., Anal. Chem.,
30, 1190 (1958). GOODWIN, T. W., AND MORTON, R. A., Biochem. J., 40, 628
(1946). YPHANTIS, D. A., Ann. S. Y. Acad. Sci., 88, 586 (1960). MALLETTE, M. F., LEWIS, S., AIWES, S. R., NELSON, J. M., AND
DAWSON, C. R., Arch. Biochem., 16, 283 (1948). BROMAN, L., MALSTR~M, B. G., AASA, R., AND VHNNG~RD, T.,
J. Molecular Biol., 6, 301 (1962). BROWN, F. C., AND WARD, D. N., Proc. Sot. Exptl. Biol. Med.,
100, 701 (1959).
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

May 1963 S. Bouchillou,x, P. McMahill, and H. S. Mason 1707
26. SHIMAO, K., Biochim. et Biophys. Acta, 62, 205 (1962). 27. CLAYTON, H. A., Arch. Biochem. Biophys 27. CLAYTON, R. A., Arch. Biochem. Biophys., 81, 404 (1959).
Y., Cold Spring Harbor Symposia on quantitative biology,
28. BJORKMAN O., AND HOLMGREM, P., Phi 28. BJORKMAN, O., AND HOLMGREM, P., Physiol. Plantarum, 13, VoZ. XXVI, Long Island Biological Assoc., Cold Spring
. ..-.- ,.,2x 582 (1960). Harbor, New York, 1961, p. 233.
29. MAYlH, A. WI., rnyszor. rm’n‘w-I 29. MAYER, A. M., Physiol. Plantarum, 14, 322 (1961). 32. SUSSMAN, A. S., Arch. Biochem. Biophys., 96, 407 (1961).
30. HOROWITZ, N. H., AND FLING, hi., VITOG~ 30. HOROWITZ, N. H., AND FLING, M., Genetics, 38, 360 (1953). 33. Fox, A. S., AND BURNETT, J. B., Biochim. et Biophys. Acta, 61,
31. HOROWITZ, N. H., FLING, M., MACLEOD, 31. HOROWITZ, N. H., FLING, M., MACLEOD, H., AND WATANABE, 108 (1962).
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Simone Bouchilloux, P. McMahill and H. S. MasonMOLECULAR PROPERTIES OF THE ENZYMES
The Multiple Forms of Mushroom Tyrosinase: PURIFICATION AND
1963, 238:1699-1707.J. Biol. Chem.
http://www.jbc.org/content/238/5/1699.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/238/5/1699.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from